Embed Size (px)
DESCRIPTION
These slides are from 3DSIG 2014, presented on July 11. I describe our investigation of internal symmetry in protein structures. This is quite common (24% of domains), and has many implications for function, folding, and evolution. I introduce the CE-Symm method, described in Myers-Turnbull, D., Bliven, S. E., Rose, P. W., Aziz, Z. K., Youkharibache, P., Bourne, P. E., & Prlić, A. (2014). Systematic Detection of Internal Symmetry in Proteins Using CE-Symm. Journal of Molecular Biology, 426(11), 2255–2268. doi:10.1016/j.jmb.2014.03.010 I discuss the results from running CE-Symm across the PDB, as well as some particularly compelling examples. See also my poster by the same title for more details.
Citation preview

Spencer Bliven July 11, 2014 3DSIG 2014
Myers-Turnbull, D., Bliven, S. E., Rose, P. W., Aziz, Z. K., Youkharibache, P., Bourne, P. E., & Prlić, A. (2014). Systematic Detection of Internal Symmetry in Proteins Using CE-Symm. Journal of Molecular Biology, 426(11), 2255–2268. PMID 24681267
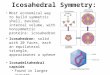
Hemoglobin [4HHB] C2
GTP Cyclohydrolase I [1A8R]
D5
Rhinovirus 2 [3DPR] Icosahedral
AmtB Ammonia Channel [1U7G]
C3
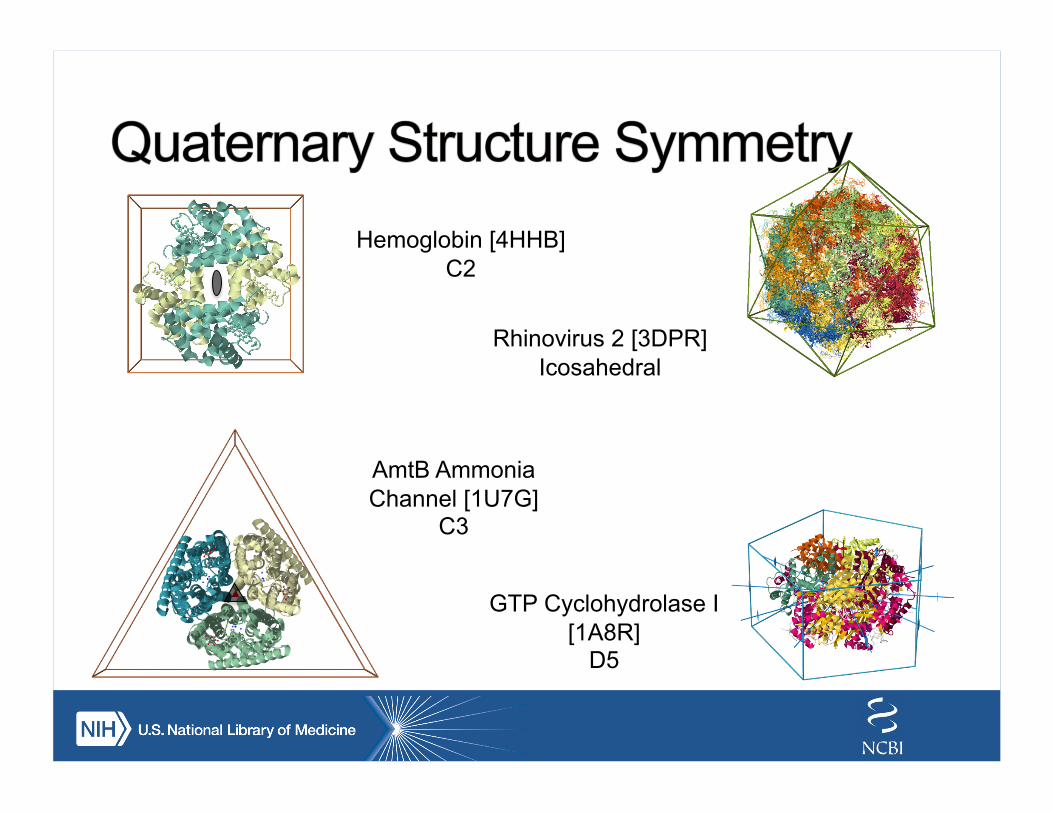
Ferredoxin-like [d2j5aa1]
C2
Beta-Propeller [d1u6dx_]
C6
Beta-trefoil [3JUT]
C3
TIM barrel [1TIM]
C8
Key: Crystallographic/NCS axis Pseudosymmetry axis

! Function ! Allosteric regulation/cooperativity ! Bind ligands symmetrically (e.g.
metals, palindromic DNA, channels) TATA Binding Protein
[1TGH]
Monod, J., Wyman, J., & Changeux, J.-P. (1965). J Mol Biol, 12, 88–118.

! Function ! Allosteric regulation/cooperativity ! Bind ligands symmetrically (e.g.
metals, palindromic DNA, channels)
! Folding ! Prevent infinite assembly ! Subunits fold quasi-
independently
TATA Binding Protein [1TGH]
Monod, J., Wyman, J., & Changeux, J.-P. (1965). J Mol Biol, 12, 88–118. Wolynes, P. G. (1996). PNAS, 93(25), 14249–14255.
Crystal of Squalene synthase [3WCG]
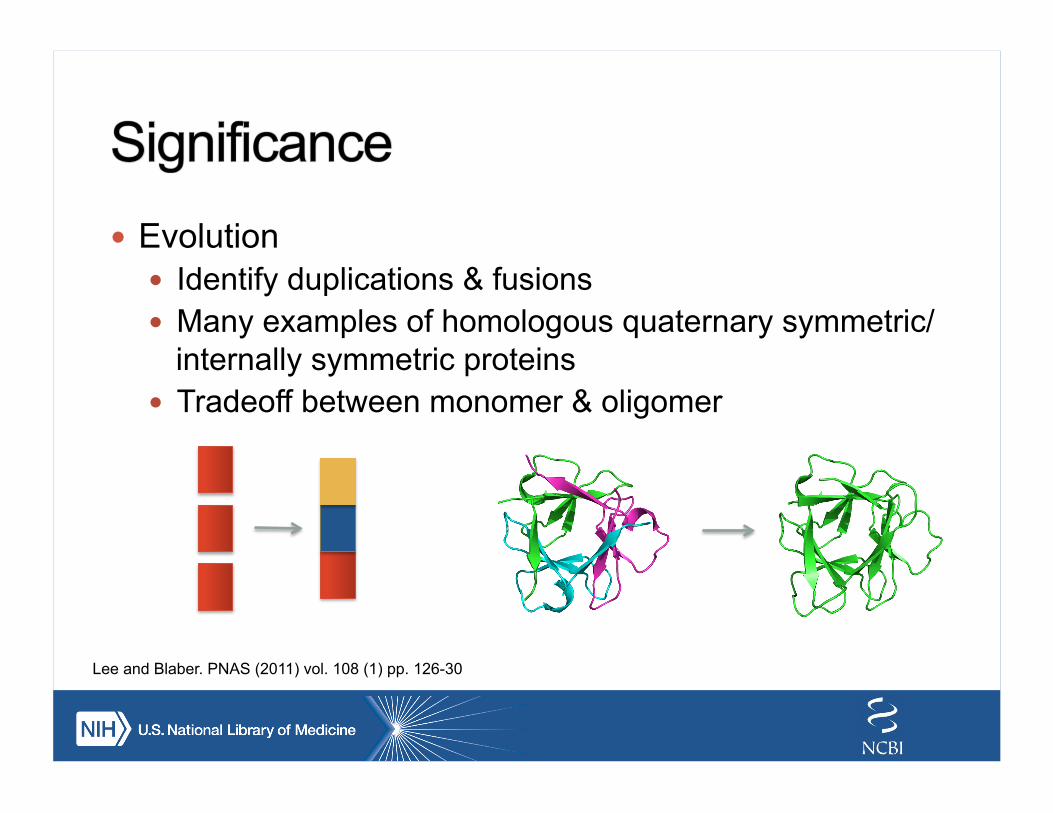
! Evolution ! Identify duplications & fusions ! Many examples of homologous quaternary symmetric/
internally symmetric proteins ! Tradeoff between monomer & oligomer
Lee and Blaber. PNAS (2011) vol. 108 (1) pp. 126-30
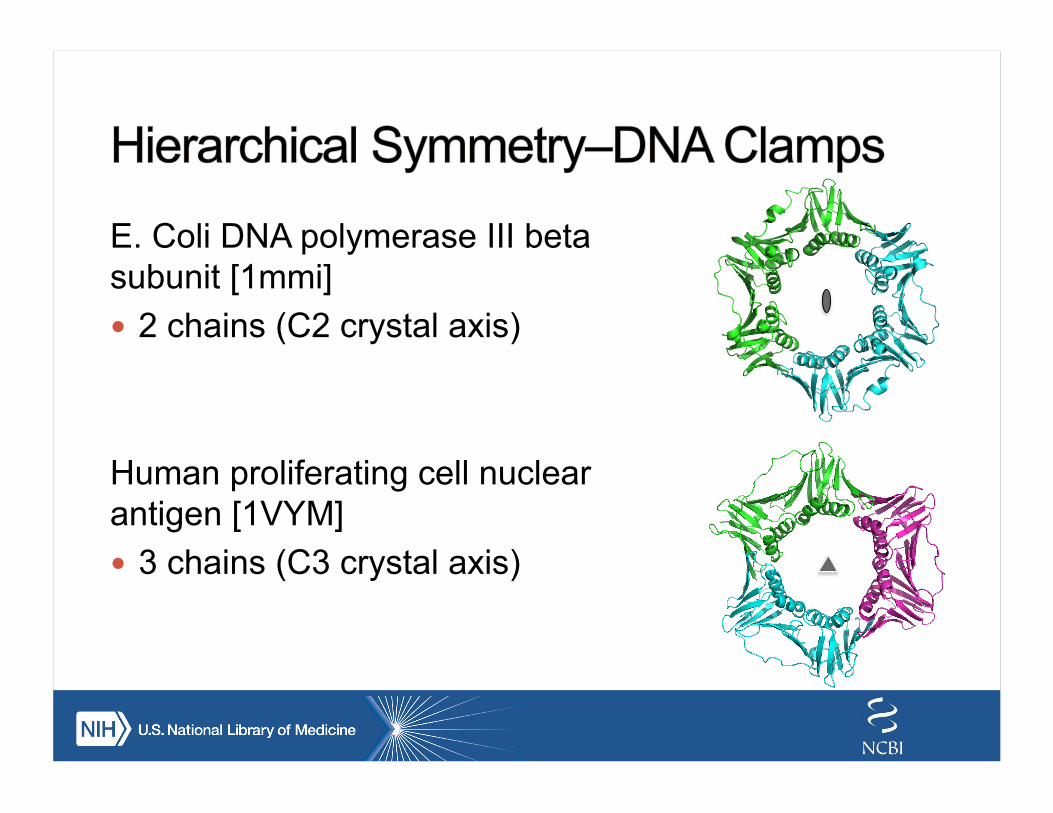
E. Coli DNA polymerase III beta subunit [1mmi] ! 2 chains (C2 crystal axis)
Human proliferating cell nuclear antigen [1VYM] ! 3 chains (C3 crystal axis)
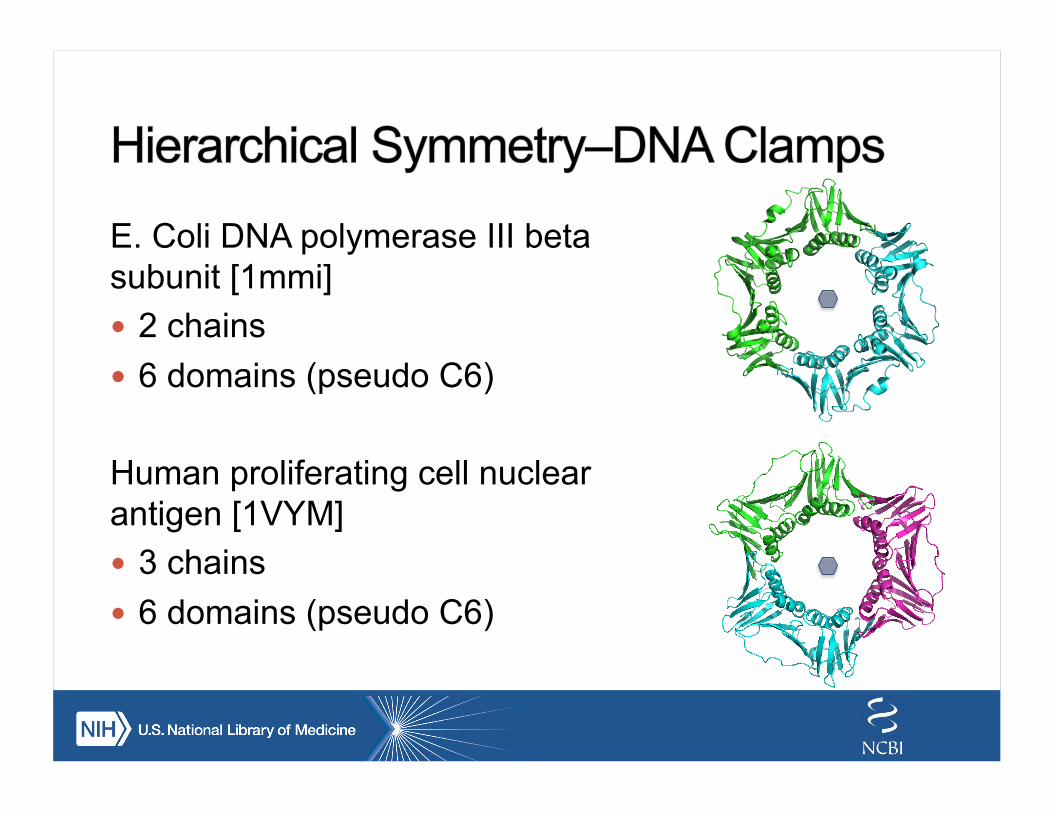
E. Coli DNA polymerase III beta subunit [1mmi] ! 2 chains ! 6 domains (pseudo C6)
Human proliferating cell nuclear antigen [1VYM] ! 3 chains ! 6 domains (pseudo C6)
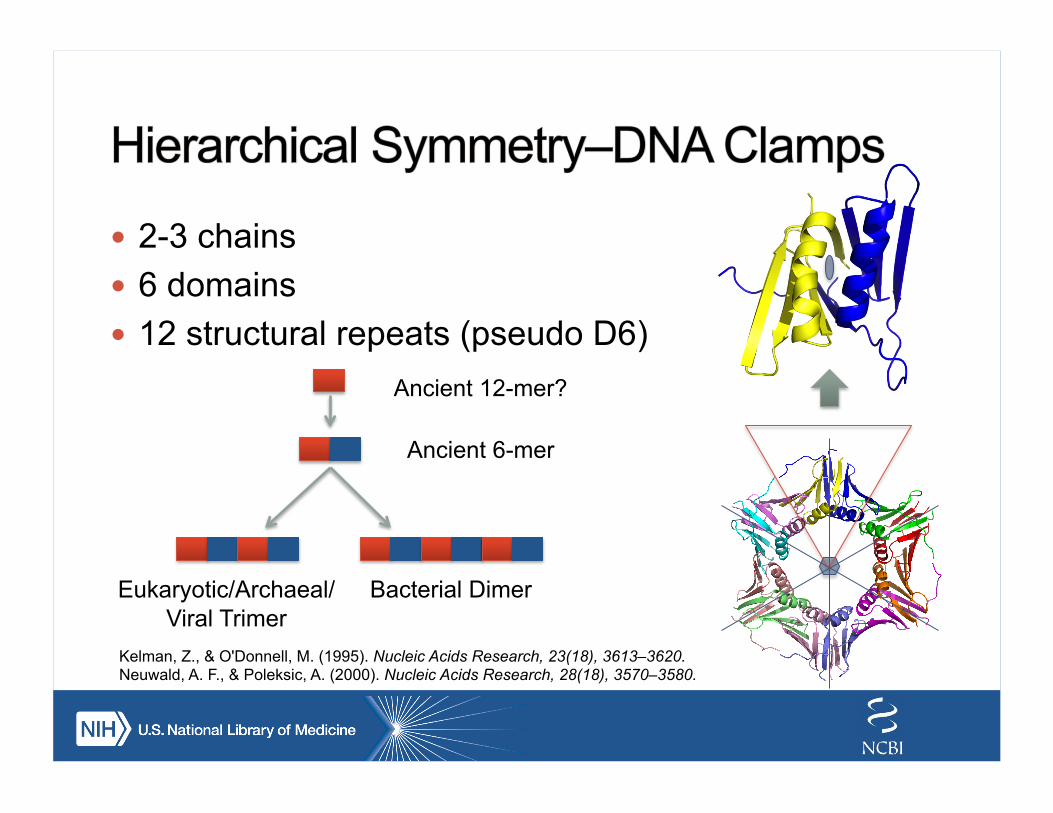
! 2-3 chains ! 6 domains ! 12 structural repeats (pseudo D6)
Ancient 12-mer?
Ancient 6-mer
Bacterial Dimer Eukaryotic/Archaeal/Viral Trimer
Kelman, Z., & O'Donnell, M. (1995). Nucleic Acids Research, 23(18), 3613–3620. Neuwald, A. F., & Poleksic, A. (2000). Nucleic Acids Research, 28(18), 3570–3580.

! Extends Combinatorial Extension (CE) algorithm for structural alignment
! Web server: source.rcsb.org/jfatcatserver/symmetry.jsp
! Download & Source code: github.com/rcsb/symmetry (LGPL)
Shindyalov, I. N., & Bourne, P. E. (1998). Protein Engineering, 11(9), 739–747.
Jia, Y., Dewey, T. G., Shindyalov, I. N., & Bourne, P. E. (2004). J Comput Biol, 11(5), 787–799.

Fibroblast Growth Factor [3JUT]
120° 120°
Myers-Turnbull, D., Bliven, S. E., Rose, P. W., Aziz, Z. K., Youkharibache, P., Bourne, P. E., & Prlić, A. (2014). Journal of Molecular Biology, 426(11), 2255–2268.
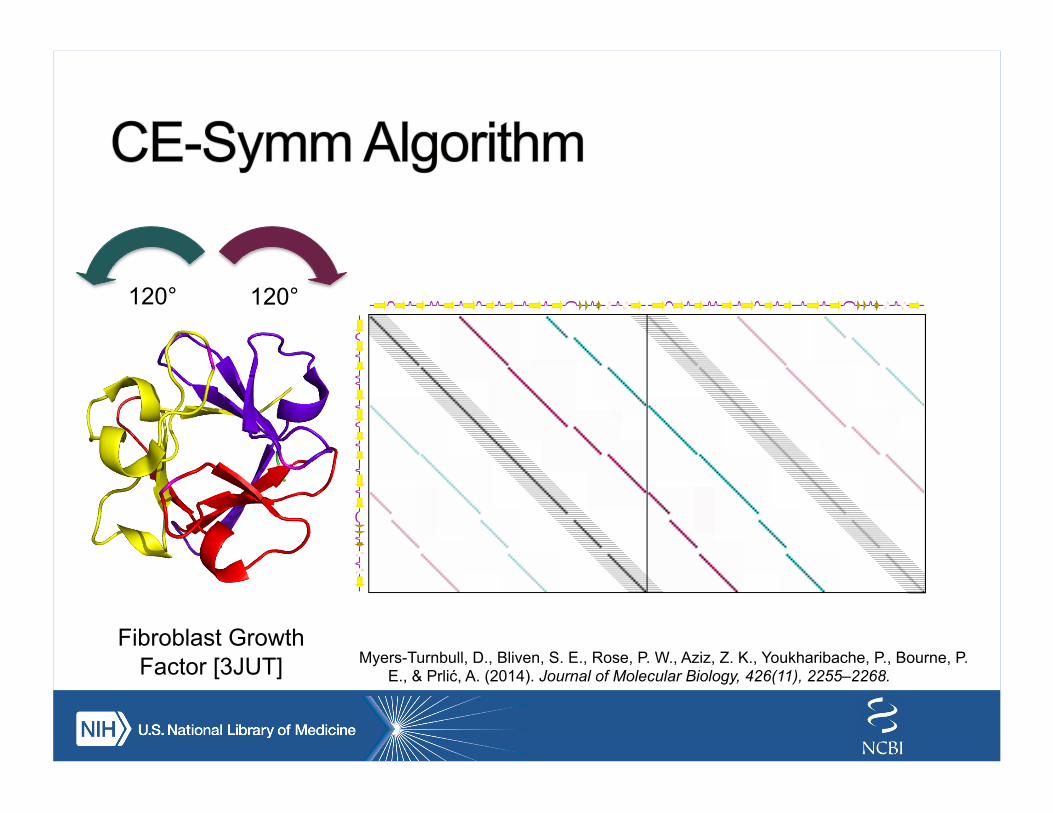
Fibroblast Growth Factor [3JUT]
120° 120°
Myers-Turnbull, D., Bliven, S. E., Rose, P. W., Aziz, Z. K., Youkharibache, P., Bourne, P. E., & Prlić, A. (2014). Journal of Molecular Biology, 426(11), 2255–2268.
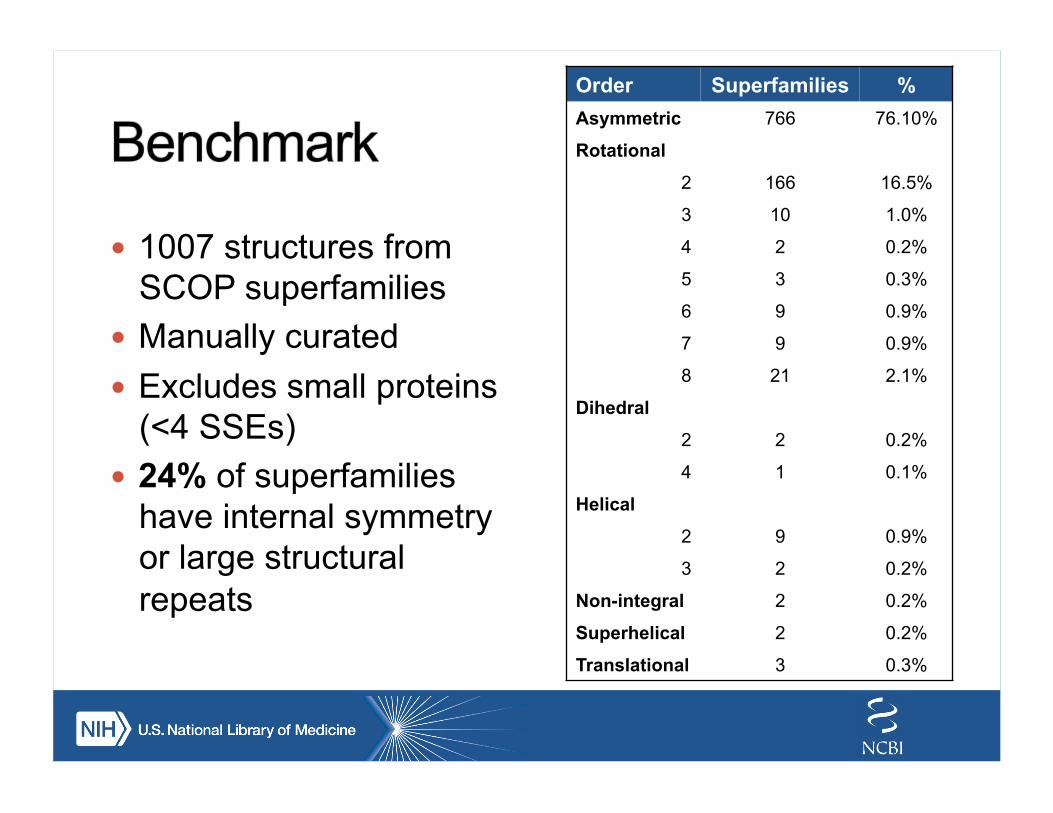
! 1007 structures from SCOP superfamilies
! Manually curated ! Excludes small proteins
(<4 SSEs) ! 24% of superfamilies
have internal symmetry or large structural repeats
Order Superfamilies % Asymmetric 766 76.10%
Rotational
2 166 16.5%
3 10 1.0%
4 2 0.2%
5 3 0.3%
6 9 0.9%
7 9 0.9%
8 21 2.1%
Dihedral
2 2 0.2%
4 1 0.1%
Helical
2 9 0.9%
3 2 0.2%
Non-integral 2 0.2%
Superhelical 2 0.2%
Translational 3 0.3%
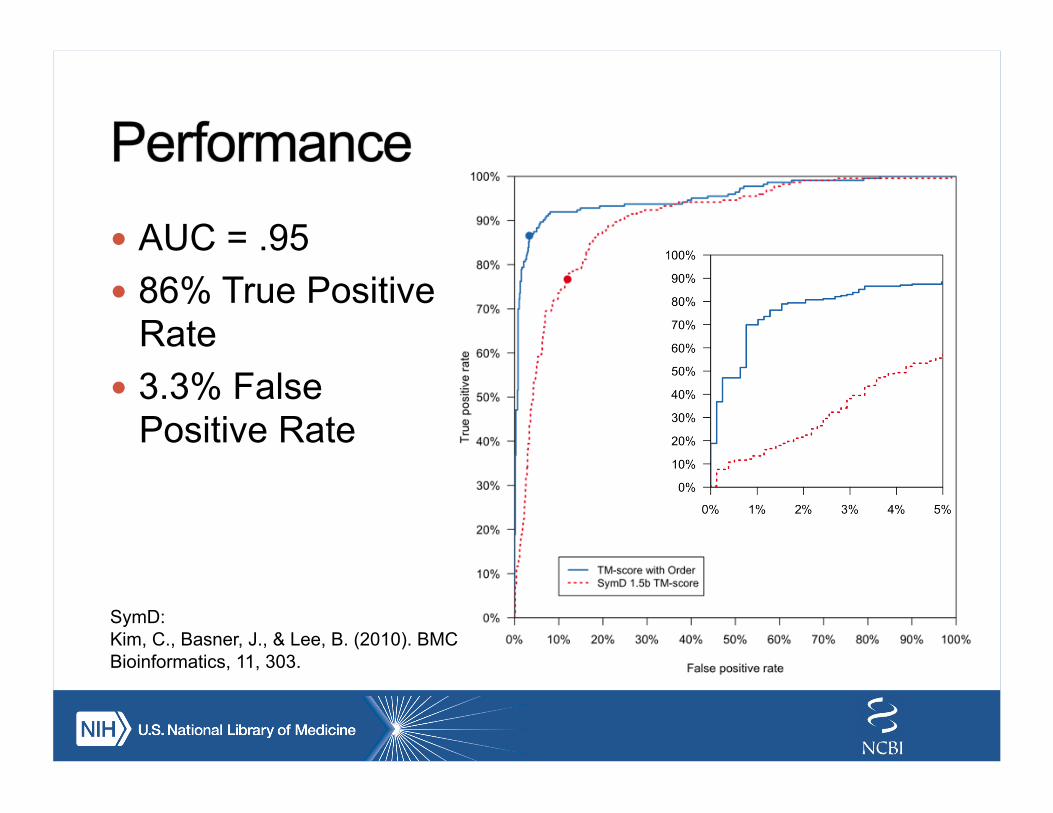
! AUC = .95 ! 86% True Positive
Rate ! 3.3% False
Positive Rate
SymD: Kim, C., Basner, J., & Lee, B. (2010). BMC Bioinformatics, 11, 303.

! All domains from SCOPe 2.03 ! Interactive results:
source.rcsb.org/jfatcatserver/scopResults.jsp ! Underestimate based on conservative thresholds
SCOP Class Superfamilies % Symmetric α 507 18.5% β 354 24.6% α/β 244 16.8% α+β 551 14.3% Multi-domain 66 4.5% Membrane 109 23.8% Overall 1831 18.0%
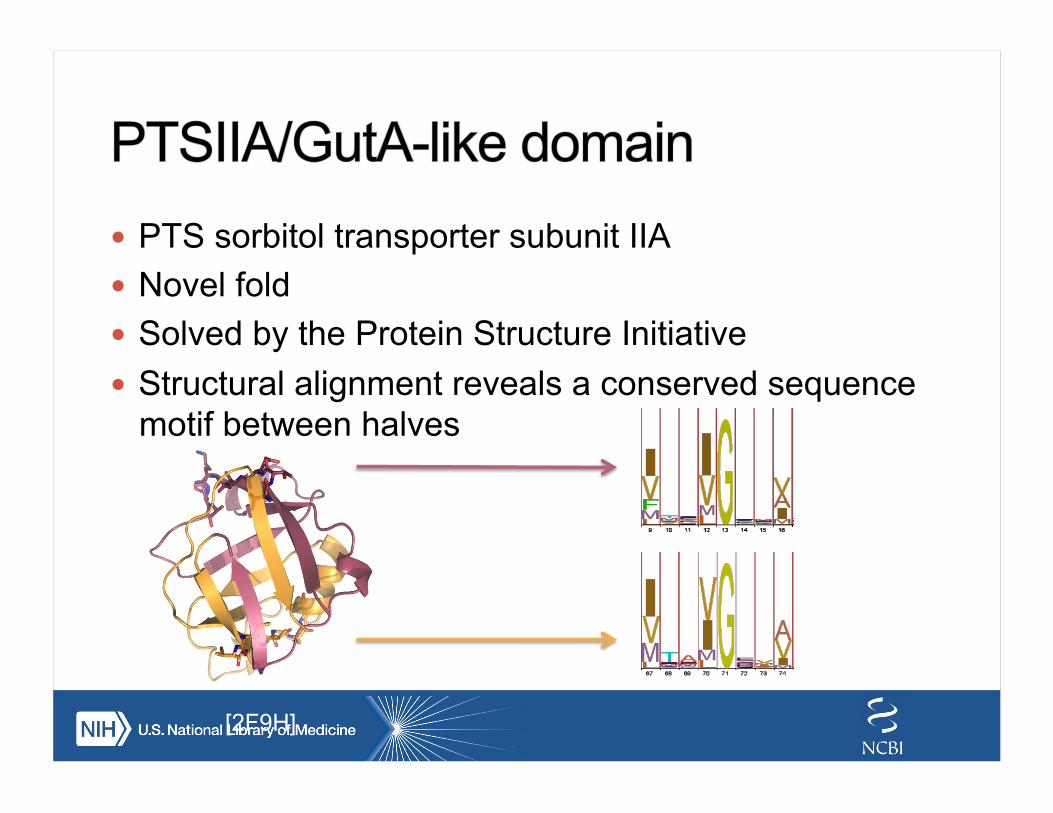
! PTS sorbitol transporter subunit IIA ! Novel fold ! Solved by the Protein Structure Initiative ! Structural alignment reveals a conserved sequence
motif between halves
[2F9H]
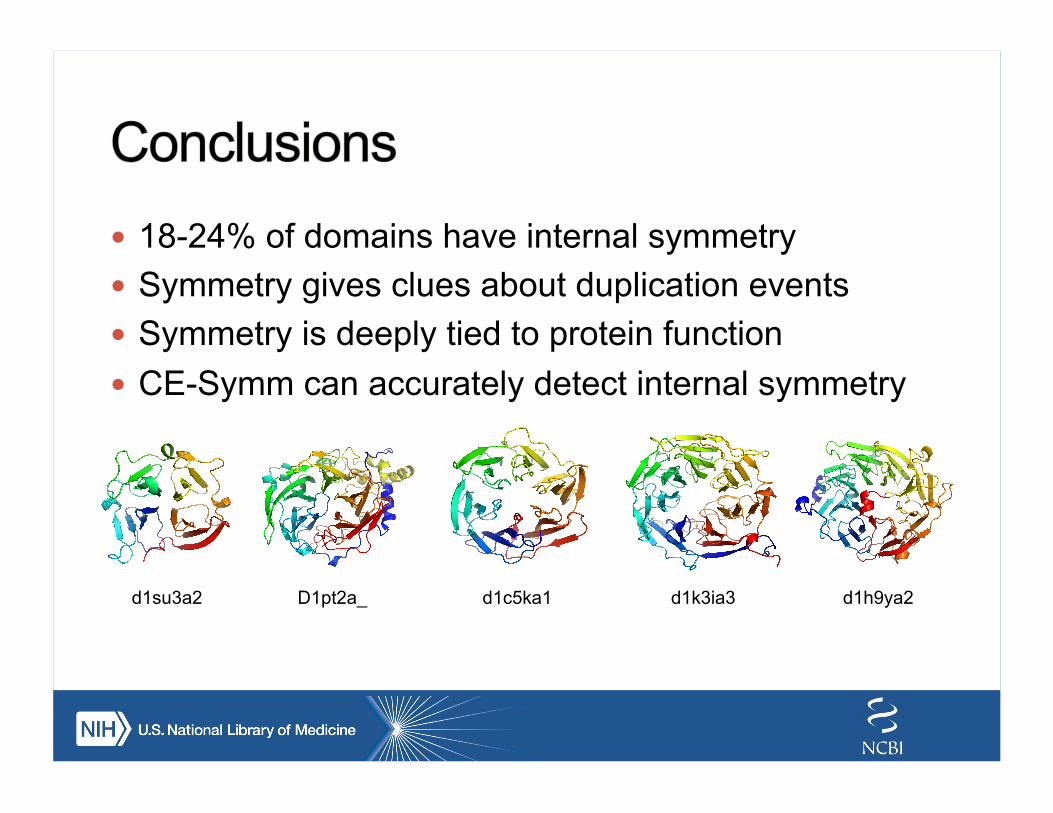
! 18-24% of domains have internal symmetry ! Symmetry gives clues about duplication events ! Symmetry is deeply tied to protein function ! CE-Symm can accurately detect internal symmetry
d1su3a2 D1pt2a_ d1c5ka1 d1k3ia3 d1h9ya2

! UC San Diego/RCSB ! Douglas Myers-Turnbull ! Andreas Prlić ! Peter Rose ! Zaid Aziz ! Milton Saier ! RCSB & Bourne Lab
members ! NIH
! Philip Bourne ! Philippe Youkharibache ! David Landsman
! Paul Scherrer Institute ! Guido Capitani & Lab
members
Resources: ! source.rcsb.org/jfatcatserver/
symmetry.jsp ! github.com/rcsb/symmetry ! Poster 25 ! www.slideshare.net/sbliven ! Funding: NSF, NIH, DOE,
Open Science Grid

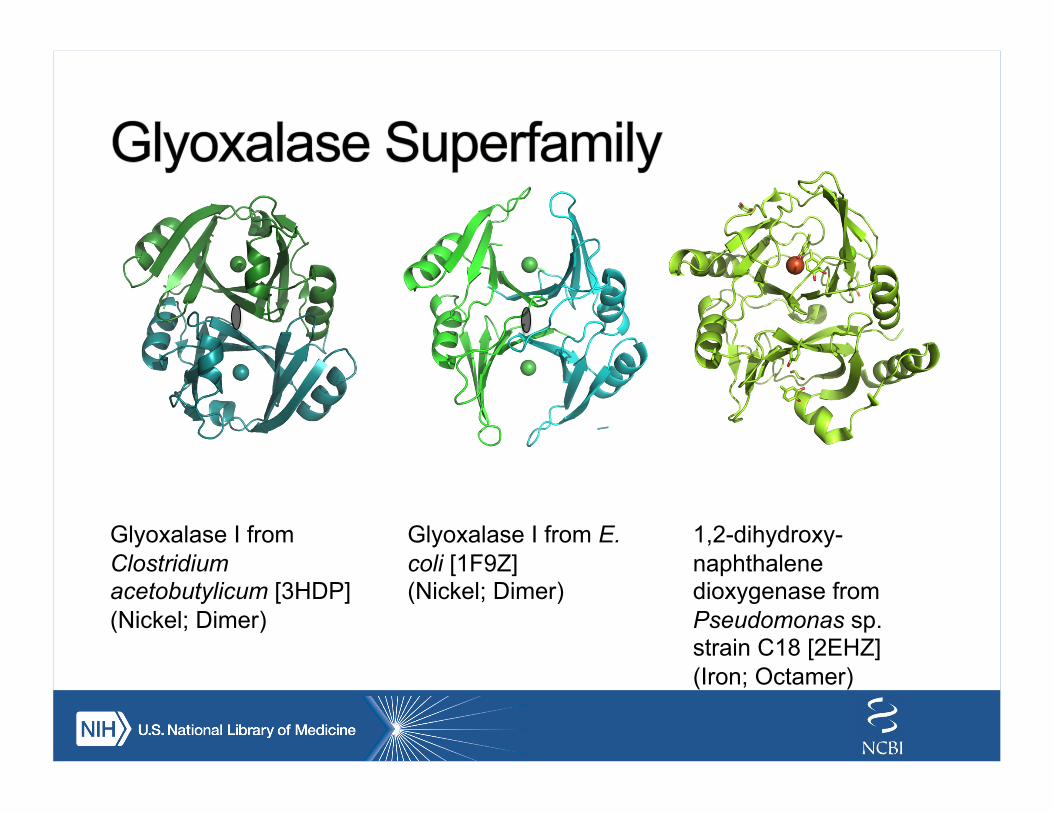
Glyoxalase I from Clostridium acetobutylicum [3HDP] (Nickel; Dimer)
Glyoxalase I from E. coli [1F9Z] (Nickel; Dimer)
1,2-dihydroxy-naphthalene dioxygenase from Pseudomonas sp. strain C18 [2EHZ] (Iron; Octamer)
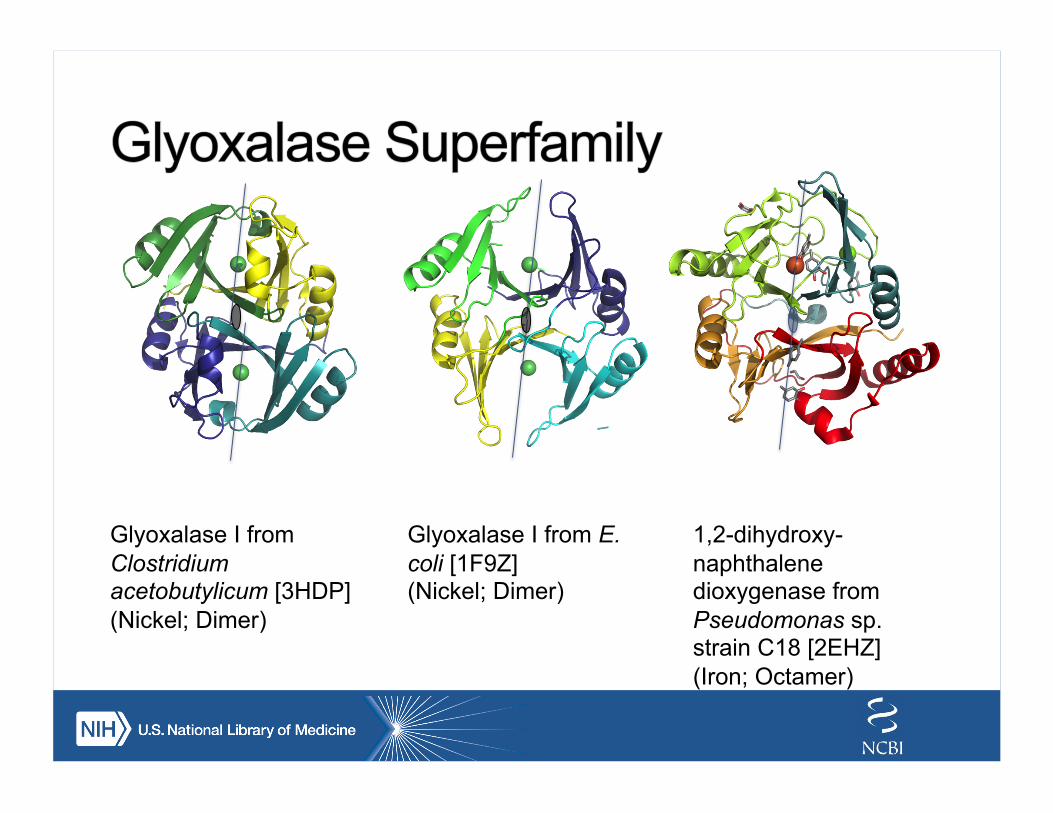
Glyoxalase I from Clostridium acetobutylicum [3HDP] (Nickel; Dimer)
Glyoxalase I from E. coli [1F9Z] (Nickel; Dimer)
1,2-dihydroxy-naphthalene dioxygenase from Pseudomonas sp. strain C18 [2EHZ] (Iron; Octamer)
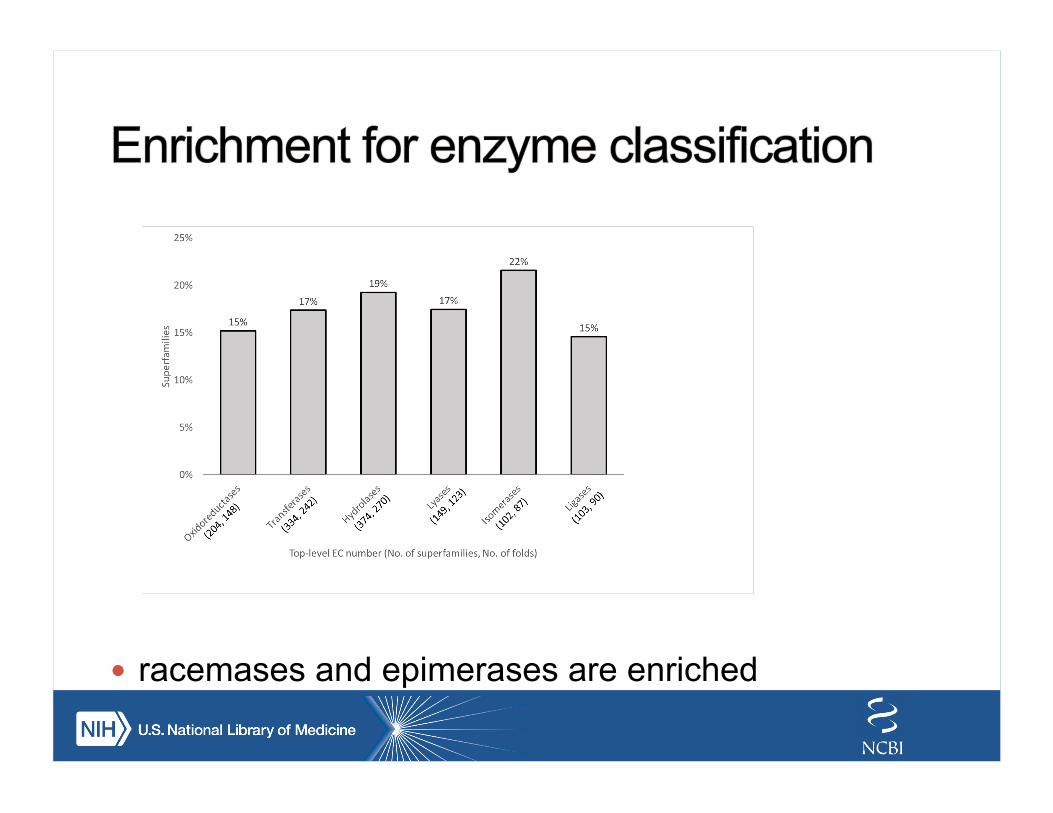
! racemases and epimerases are enriched
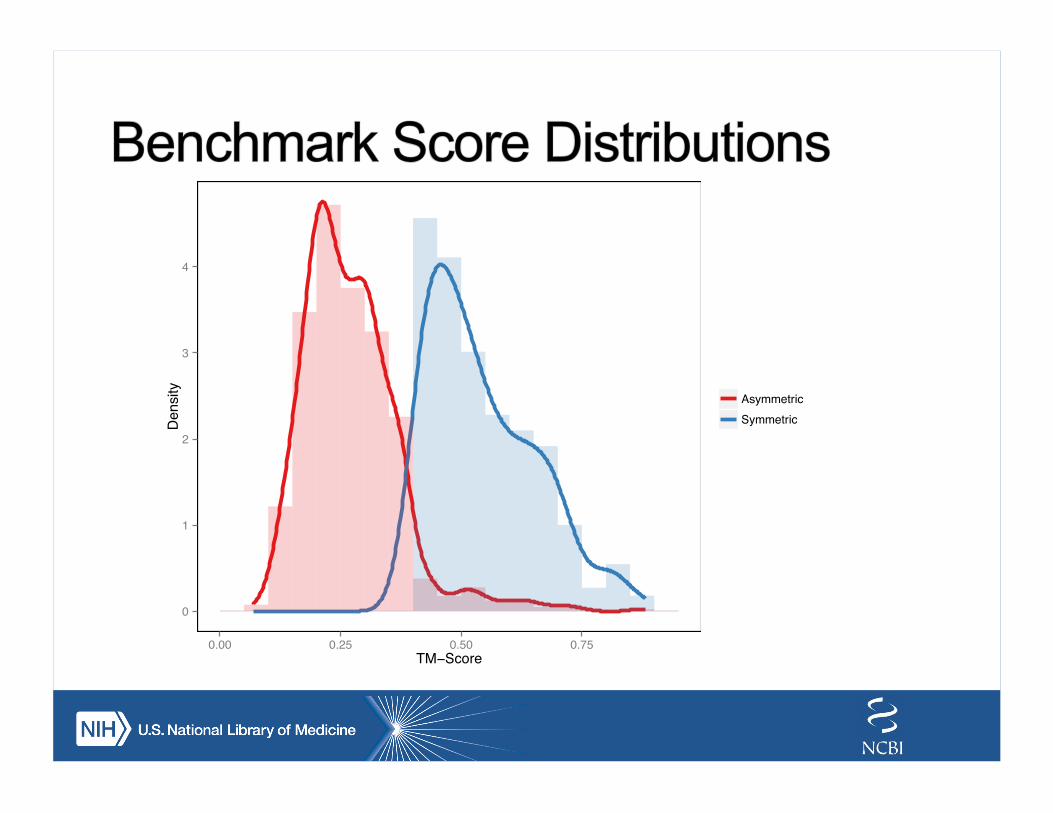
0
1
2
3
4
0.00 0.25 0.50 0.75TM−Score
Density
AsymmetricSymmetric

This work is licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License.