Embed Size (px)
Citation preview

e at SciVerse ScienceDirect
Toxicon 60 (2012) 821–829
Contents lists availabl
Toxicon
journal homepage: www.elsevier .com/locate/ toxicon
Short communication
Potamotrygon cf. henlei stingray mucus: Biochemical features of a novelantimicrobial protein
Katia Conceição a,*, Juliane Monteiro-dos-Santos c, Carla Simone Seibert c,Pedro Ismael Silva Jr. b, Elineide Eugênio Marques c, Michael Richardson d,Mônica Lopes-Ferreira a
a Immunoregulation Unit, Special Laboratory of Applied Toxinology (CEPID/FAPESP), Butantan Institute, Avenida Vital Brazil 1500, Sao Paulo – SP 05503-900, Brazilb Functional Proteomic Units, Special Laboratory of Applied Toxinology (CEPID/FAPESP), Butantan Institute, Sao Paulo, Brazilc Federal University of Tocantins, Brazild FUNED (Fundação Ezequiel Dias – Belo Horizonte, MG), Brazil
a r t i c l e i n f o
Article history:Received 9 April 2012Received in revised form 4 May 2012Accepted 24 May 2012Available online 6 June 2012
Keywords:StingrayPotamotrygonMucusHemoglobinAntimicrobial
* Corresponding author. Tel.: þ55 11 3726 1024.E-mail address: [email protected]
0041-0101/$ – see front matter � 2012 Elsevier Ltdhttp://dx.doi.org/10.1016/j.toxicon.2012.05.025
a b s t r a c t
Antimicrobial molecules are important components of the innate immune system invertebrates. They have been studied widely in several fishes, but little is known aboutthese defence factors in stingrays, which are thought to have less sophisticated adaptiveimmune systems when compared to other teleosts. Stingrays from the specie Potamo-trygon cf. henlei are distributed throughout the rivers of central-west Brazil, being thecause of numerous envenomations occurring in the dry seasons.In a previous study, we reported that the mucus of the stingray P. cf. henlei showsantimicrobial effects. Here, to analyze the antimicrobial compounds from the mucus of P. cf.henlei, we employed solid-phase extraction, chromatographic separation followed by ESI-MS, and Edman degradation. A protein similar to the b-chain of hemoglobin was identi-fied, isolated and partially sequenced by Edman degradation. This protein has a molecularweight of 16072.8 Da, and was shown to be active against bacteria (Micrococcus luteus andEscherichiacoli) and yeast (Candida tropicalis) without hemolytic activity. Effects of this newprotein in the microcirculation environment were also evaluated. The results obtainedprovide fundamental information for future basic research, clinical diagnosis and develop-ment of new therapies to accident treatment. To the best of our knowledge, this is the firstdescription of a bioactive polypeptide from the mucus of a stingray.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The skin of fish constitutes a pivotal immunologicalprotection against the external environment. The layer ofmucus on the fish surface, considered the first line ofdefence, participates in a number of functions includingdisease resistance, respiration, ionic and osmotic regula-tion, locomotion, reproduction, communication, feedingand nest building (Negus, 1963; Ingram, 1980; Shephard,
(K. Conceição).
. All rights reserved.
1994; Zhao et al., 2008). The mucus, such as thatproduced by the skin of the stingrays, has a complex set ofcomponents, which may include amino acid residues,peptides, complex carbohydrates, glycopeptides, glyco-lipids and other chemicals (Klesius et al., 2008; Alexanderand Ingram, 1992; Birkemo et al., 2003).
Fish epidermal mucus was found to display antimicro-bial activity against broad range of infectious pathogens(Mozumder, 2005; Hellio et al., 2002). We recentlydescribed the antimicrobial activity of catfish Cathoropsspixii mucus (Ramos et al., 2012). Moreover, histone H2Band two ribosomal proteins are examples of proteins withantimicrobial activity that have been isolated from

K. Conceição et al. / Toxicon 60 (2012) 821–829822
epidermal mucus of Atlantic cod (Bergsson et al., 2005).Members of some families of antimicrobial peptides (AMPs)were also found to be important innate defence componentsin the epidermal mucosal layer of Moses sole fish (Parda-chirus marmoratus) (Oren and Shai, 1996), winter flounder(Pleuronectes americanus) (Cole et al., 2000), Atlantic halibut(Hippoglossus hippoglossus) (Birkemo et al., 2003), Hagfish(Myxine glutinosa L.) (Subramanian et al., 2009) and catfish(Pelteobagrus fulvidraco) (Su, 2011). These observationssuggest that mucus is a good source of novel molecules forfish and human health-related applications.
We also recently reported that themucus of the stingrayPotamotrygon. cf. henlei shows antimicrobial effects anda pro-inflammatory response (Monteiro-Dos-Santos et al.,2011). The aim of the present study was therefore toidentify and characterize the major component(s) withantimicrobial activity in the mucus of P. cf. henlei, which isa very common stingray found in northern and central-western rivers from Brazil (Carvalho et al., 2003). Theirspines are hard, sharp, bilaterally retroserrated and coveredby an integumentary sheath with a ventrolateral glandulargroove containing venom glands along both edges(Halstead, 1970) and the mucus of biological importancethat covers the entire body of these animals. This studyemployed a screening approach onmucus components thatwere purified by RP-HPLC, and characterized by ESI-MS andEdman degradation. By this approach, several compoundsincluding peptides were obtained and a protein similar toHemoglobin b-chain was identified, isolated and charac-terized. Following antimicrobial and hemolytic assays,intravital microscopy was used to image the effects of theprotein on the microcirculation.
2. Materials and methods
2.1. Collection of stingray mucus
Specimens of adult female and male (n ¼ 15) P. cf. henleifish were collected from the Manoel Alves River in the stateof Tocantins, Brazil. Mucus dispersed all over the body wascollected by scraping the skin with a glass slide, andimmediately stored on ice, then diluted in 0.15 Mphosphate-buffered sterile saline, pH 7.4, homogenized,and centrifuged (5000 � g for 20 min at 4 �C) for collectionof the supernatant. The supernatant was collected andstored at �20 �C. Protein content was determined by themethod of Bradford (1976) using bovine serum albumin(Sigma Chemical Co., St Louis, MO) as standard protein.
2.2. Purification of mucus
The supernatants were loaded onto solid phase extrac-tion cartridges Sep-Pak, C18 (Waters Corporation, Taunton,MA, USA) equilibrated in acidified water (0.1% trifluoro-acetic acid (TFA)). A single aliquot of 3 mg diluted in 3mL of0.1% TFA was loaded and the elution was performedsequentially with 40 and 80% acetonitrile. These fractionswere further concentrated by a vacuum centrifugation.Aliquots of 1 mg of the samples were dissolved in 1 mL ofdeionized water in 0.1% TFA and centrifuged at 5000� g for20 min (10 �C). The supernatants were applied to a system
of RP – HPLC (Äkta basic, Amersham Biosciences – Sweden)for the sample separation. The sample was loaded ina Jupiter C18 column (4.6mm� 150mm, 5 mm, Phenomenex,USA) in a two-solvent system: (A) TFA/H2O (1:1000) and (B)TFA/Acetonitrile (ACN)/H2O (1:900:100). The column waseluted at a flow rate of 1.0mL/minwith a 10–80% gradient ofsolvent B over 40 min. For protein purification, furtheradditional steps of chromatography were necessary, usinga Bio Basic C8 column (4.6 mm � 250 mm, 5 mm, Thermo,USA) with optimized gradients. The HPLC column eluateswere monitored by their absorbance at 214 nm.
2.3. Sodium dodecyl sulphate-polyacrylamide gelelectrophoresis (SDS-PAGE)
SDS-PAGE was carried out according to Laemmli (1970).Proteins (10 mg) from the mucus of P. cf. henlei wereanalyzed by SDS-PAGE 4–20% acrylamide gradient underreducing conditions. Prior to electrophoresis, the sampleswere mixed 1:1 (v/v) with sample buffer. The gels werestained with the Silver method.
2.4. Mass spectrometry (MS) and peptide mass fingerprint(PMF)
The fractions were analyzed by electrospray, with directinjection in an LC–MS Surveyor MSQ Plus (Thermo Elec-tron, USA) under positive ionization mode. The needle andcone potential were set to 3.1 kV and 40 V, respectively. Theaqueous sample solutions (10 mL) were directly injected ata 50 mL/min constant flow rate of acetonitrile H2O/0.1%formic acid (1:1). External calibration was performed withNaI (Sigma) over m/z 100–2000.
Protein band was excised and in-gel trypsin digestionwas performed according to Hellman et al. (1995). Nano-spray MS/MS analysis was performed on tryptic digests ofSDS-Page band of purified PcfHb using Q-Tof mass spec-trometry (Q-TOF Ultima API Waters/Micromass, Man-chester, United Kingdom). An aliquot (5 mL) of the resultingpeptide mixture were injected into Symmetry C18 trappingcolumn (5 mm particles, 180 mm i.d.� 20 mm, Waters, USA)to desalt the peptide mixture. The nano UPLC (Waters)conditions were 0.1% formic acid in water (solvent A) andacetonitrile with 0.1% formic acid as solvent B. The sepa-rations were performed at a flow rate of 0.6 ml/min usinga 0–80% gradient of solvent B over 45 min. The LC systemwas coupled to a nano ESI source of the Q-ToF instrumentusing a BEH130C18 column (75 mm � 100 mm, 1.7 mmparticles; Waters, MA, USA). Typical conditions werea capillary voltage of 3.1 kV, a cone voltage of 50 V, andsource temperature of 70 �C. Data dependent acquisition(parent ions with 2, 3 and 4 charges) were automaticallyrecognized by the charge state recognition software Mas-sLynx 4.1 (Waters, USA). The peptide ions were detected byscanning fromm/z 200 tom/z 2000 at a rate of 1 scan/s, andwere subjected to collision-induced dissociation withargon in the 13–55 eV collision energy range. Product ionsfrom MS/MS experiments were detected by scanning fromm/z 50 tom/z 2000 at a rate of 1 scan/s. External calibrationwas performed using phosphoric acid (Merck, Darmstadt,Germany).

K. Conceição et al. / Toxicon 60 (2012) 821–829 823
All MS/MS spectra were acquired using MassLynx 4.0software (Waters), deisotoped and converted (by WatersProteinLynx Global server 1.0 software) and searched usinga licensed copy of the Mascot Server 2.2 program (MatrixScience, London, UK). In each instance, the search wascarried out against the non-redundant protein database atthe National Center for Biotechnology Information bank(NCBI www.ncbi.nlm.nih.gov) assuming the parameters ofIodoacetamide derivative of cysteine as fixed modificationand methionine oxidation as variable modification, mon-oisotopic mass values,� 0.5 Da peptidemass tolerance, and�0.5 Da fragment mass tolerance. Mascot identificationsrequired that at least the ion scores must be greater thanthe associated identity scores, and 20, 30, 40 and 50 forsingle, double, triple and quadruple charged peptides.Furthermore, Mascot searches were followed by manualinterpretation of MS/MS spectra to eliminate false positiveswith the help of the PepSeq tool (MassLynx 4.1 software,Waters, USA).
2.5. Antimicrobial assay
The antimicrobial activities were determined usinga modified microtiter broth dilution method. The antimi-crobial activity wasmonitored by a liquid growth inhibitionassay against gram positive bacteria Micrococcus luteusA270, gram negative Escherichia coli SBS 363 and yeastCandida tropicalisMDM8, as described by Bulet et al. (1993)and Ehret-Sabatier et al. (1996). Pre inocula of the strainswere prepared in Poor Broth (1.0 g peptone in 100 mL ofH2O containing 86 mM NaCl at pH 7.4; 217 mOsM for M.luteus and E.coli and 1.2 g potato dextrose in 100 mL of H2Oat pH 5.0; 79 mOsM for C. albicans) and incubated at 37 �Cwith shaking. The absorbance at 595 nm was determinedand one aliquot of this solution was taken to obtain cells inlogarithmic growth (A595nm w 0.6), and diluted 600 times(A595 nm ¼ 0.0001). The venom, mucus and fractions weredissolved in sterileMilli-Qwater, at a final volume of 100 mL(10 mL of the sample and 90 mL of the inoculum in PB broth).After incubation for 18 h at 30 �C the inhibition of bacterialgrowth was determined by measuring absorbance at595 nm.
2.6. Hemolytic studies
For hemolytic studies human red blood cells froma healthy donor (type A) were collected in 0.15 M citratebuffer, pH7.4, and washed 3 times by centrifugation with0.15 M phosphate-buffered saline, pH 7.4. To determine thehemolytic activity, protein samples were assayed in tripli-cate and tested up to 100 mM: 1.563, 3.125, 6.250, 12.5, 25,50 and 100 mM in a 3% suspension of erythrocytes incu-bated for 3 h at room temperature. Hemolysis was deter-mined by reading the absorbance at 595 nm of each well ina plate reader. A suspension of erythrocytes incubated withwater was used as a positive control (100% hemolysis).
2.7. Mice and intravital microscopy
Male Swiss mice (5–6 weeks old) were obtained froma colony at the Butantan Institute, São Paulo, Brazil. Animals
were housed in a laminar flow holding unit (GelmanSciences, Sydney, Australia) on autoclaved bedding, inautoclaved cages, in an air-conditioned room under a 12 hlight/dark cycle. Irradiated food and acidified water wereprovided ad libitum. All procedures involving animals werein accordance with the guidelines provided by the BrazilianCollege of Animal Experimentation.
The dynamics of alterations in the microcirculatorynetwork were determined using intravital microscopy bytransillumination of mice cremaster muscle after subcuta-neous application 10 ml of protein dissolved in sterile saline.Administration of the same amount of sterile saline wasused as control. In three independent experiments (n ¼ 4),mice were injected with 0.4% Xilazine (Coopazine�,Schering-Plough) and then anaesthetized with 0.2 g/kgchloral hydrate, and the cremaster muscle was exposed formicroscopic examination in situ as described by Conceiçãoet al. (2009), Baez (1973) and Lomonte et al. (1994). Theanimals were maintained on a board thermostaticallycontrolled at 37 �C, which included a transparent platformon which the tissue to be transilluminated was placed.After the stabilization of the microcirculator, the number ofroller cells and adherent leukocytes in the postcapillaryvenules were counted 10 min after venom injection. Thestudy of the microvascular system of the transilluminatedtissue was accomplished with an optical microscope (AxioImager A.1, Carl-Zeiss, Germany) coupled to a camera (IcC 1,Carl-Zeiss, Germany) using a 10/025 longitudinal distanceobjective/numeric aperture and 1.6 optovar.
2.8. N-terminal sequence analysis
To determine the amino acid sequence, HPLC purifiedsamples of the native proteins were subjected to Edmandegradation using a Shimadzu PPSQ-21 automated proteinsequencer, following manufacturer’s instructions.
2.9. Statistical analysis
All results were presented as means � SEM of at leastfour animals in each group. Differences among data weredetermined by ne way analysis of variance (ANOVA) fol-lowed by Dunnett’s test. Differences between two meanswere determined using unpaired Student’s t-test. Datawere considered significant at p < 0.05.
3. Results
3.1. PcfHb structural characterization
PcfHb mucus was partially purified by solid-phaseextraction to identify the mucus component(s) respon-sible for the antimicrobial activity (Monteiro-Dos-Santoset al., 2011). Three fraction eluates containing 0, 40 and80% of acetonitrile were obtained. The eluate sample con-taining 80% acetonitrile reported an enhanced antimicro-bial activity against M. luteus, E. coli and C. Tropicalis. Whenthe 80% acetonitrile eluate active factor was purified,a fraction with antimicrobial activity against the microor-ganisms tested was detected (Fig. 1A). The antimicrobialfraction 8 was subjected to further purification by the C8
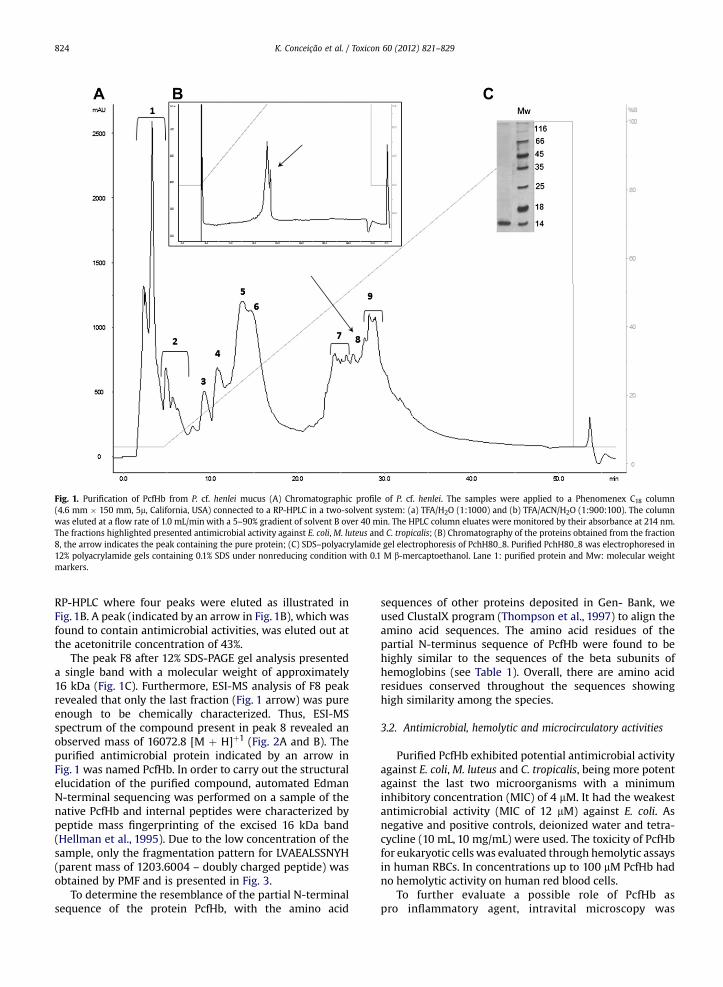
Fig. 1. Purification of PcfHb from P. cf. henlei mucus (A) Chromatographic profile of P. cf. henlei. The samples were applied to a Phenomenex C18 column(4.6 mm � 150 mm, 5m, California, USA) connected to a RP-HPLC in a two-solvent system: (a) TFA/H2O (1:1000) and (b) TFA/ACN/H2O (1:900:100). The columnwas eluted at a flow rate of 1.0 mL/min with a 5–90% gradient of solvent B over 40 min. The HPLC column eluates were monitored by their absorbance at 214 nm.The fractions highlighted presented antimicrobial activity against E. coli, M. luteus and C. tropicalis; (B) Chromatography of the proteins obtained from the fraction8, the arrow indicates the peak containing the pure protein; (C) SDS–polyacrylamide gel electrophoresis of PchH80_8. Purified PchH80_8 was electrophoresed in12% polyacrylamide gels containing 0.1% SDS under nonreducing condition with 0.1 M b-mercaptoethanol. Lane 1: purified protein and Mw: molecular weightmarkers.
K. Conceição et al. / Toxicon 60 (2012) 821–829824
RP-HPLC where four peaks were eluted as illustrated inFig. 1B. A peak (indicated by an arrow in Fig. 1B), which wasfound to contain antimicrobial activities, was eluted out atthe acetonitrile concentration of 43%.
The peak F8 after 12% SDS-PAGE gel analysis presenteda single band with a molecular weight of approximately16 kDa (Fig. 1C). Furthermore, ESI-MS analysis of F8 peakrevealed that only the last fraction (Fig. 1 arrow) was pureenough to be chemically characterized. Thus, ESI-MSspectrum of the compound present in peak 8 revealed anobserved mass of 16072.8 [M þ H]þ1 (Fig. 2A and B). Thepurified antimicrobial protein indicated by an arrow inFig. 1 was named PcfHb. In order to carry out the structuralelucidation of the purified compound, automated EdmanN-terminal sequencing was performed on a sample of thenative PcfHb and internal peptides were characterized bypeptide mass fingerprinting of the excised 16 kDa band(Hellman et al., 1995). Due to the low concentration of thesample, only the fragmentation pattern for LVAEALSSNYH(parent mass of 1203.6004 – doubly charged peptide) wasobtained by PMF and is presented in Fig. 3.
To determine the resemblance of the partial N-terminalsequence of the protein PcfHb, with the amino acid
sequences of other proteins deposited in Gen- Bank, weused ClustalX program (Thompson et al., 1997) to align theamino acid sequences. The amino acid residues of thepartial N-terminus sequence of PcfHb were found to behighly similar to the sequences of the beta subunits ofhemoglobins (see Table 1). Overall, there are amino acidresidues conserved throughout the sequences showinghigh similarity among the species.
3.2. Antimicrobial, hemolytic and microcirculatory activities
Purified PcfHb exhibited potential antimicrobial activityagainst E. coli,M. luteus and C. tropicalis, being more potentagainst the last two microorganisms with a minimuminhibitory concentration (MIC) of 4 mM. It had the weakestantimicrobial activity (MIC of 12 mM) against E. coli. Asnegative and positive controls, deionized water and tetra-cycline (10 mL, 10 mg/mL) were used. The toxicity of PcfHbfor eukaryotic cells was evaluated through hemolytic assaysin human RBCs. In concentrations up to 100 mM PcfHb hadno hemolytic activity on human red blood cells.
To further evaluate a possible role of PcfHb aspro inflammatory agent, intravital microscopy was

Fig. 2. ESI-MS spectrum of charge distribution from the fraction PcfHb. Molecular weight of the PcfHb fraction that was performed by mass spectrum decon-volution using the program Mag Tran 1.0 beta 8.
K. Conceição et al. / Toxicon 60 (2012) 821–829 825
employed to assess the effects of the protein in cellularrecruitment on microcirculation, as well to detecteventually a toxic effect of the protein. The topicalapplication of 4 mM of the protein induced an increase ofcellular recruitment characterized by an increase in the
Fig. 3. MS/MS spectrum. The fragmentation pattern for LVAEALSSNYH with a parea fragment found in chain b form of hemoglobin from Dasyatis akajei (UniProt ID:
number of leukocyte rolling (Fig. 4). The cellularrecruitment was more pronounced 5 min after topicalapplication and decreased continuously, returning tonormal after 30 min of experiment. No change innumbers of adhered cells was observed.
nt mass of 1203.6004 (a doubly charged peptide) is shown. The peptide isP56692).

Table 1Sequence alignment between PcfHb and three stingray hemoglobin b-chain performed by Clustal (Thompson et al., 1997). To optimize maximal alignment,gaps were introduced in the sequences indicated as dots. Proteins: HBB_DASAK (P56692) – hemoglobin subunit beta (beta-globin), [Dasyatis akajei (redstingray) (Akaei)]; HBB2_TORMA (P20247): Hemoglobin subunit beta-2 (beta-2-globin) (hemoglobin beta-2 chain) [HBB2] [Torpedo marmorata (Marbledelectric ray)]; Q4JDG3_BATEA – beta 1 globin OS ¼ Bathyraja eatonii; HBB1_TORMA (P20246) – hemoglobin subunit beta-1 (beta-1-globin) (hemoglobinbeta-1chain) [HBB1] [T. marmorata (Marbled electric ray)]; HBB_BATEA hemoglobin subunit beta OS ¼ Bathyraja eatonii. Amino acid sequences that areconserved are highlighted with asterisk.
K. Conceição et al. / Toxicon 60 (2012) 821–829826
4. Discussion
Although the basic features of the adaptive immunesystem have been described in fish, the limited classes ofimmunoglobulin present and functional variants of fishimmunoglobulins show the lack of a well-developedadaptive immune system in fish (Rajanbabu and Chen,2011).
To overcome this limitation, epithelial surfaces of fish,like mucus and skin have a wide variety of host defences,characterizing an efficient innate immune system whichacts as the first line of defense against the broad spectrumof pathogens encountered in the aquatic environment(Subramanian et al., 2009; Robinette et al., 1998).
Antimicrobial activity is most commonly found inspecialized peptides with a number of unusual structures(Rajanbabu and Chen, 2011). However, an increasingnumber of antimicrobials are found to be larger proteins orfragments obtained thereof. Among the most common of
these are histone-like proteins: 13.5–21 kDa proteins iso-lated from the skin, gill, liver and spleen of various fish.Highly homologous to histones, they have potent, broad-spectrum activity against Gram-negative bacteria, watermolds and parasites (Richards et al., 2001; Fernandes et al.,2002). Another example is the antimicrobial peptide hip-posin from the skin mucus of Atlantic halibut (Hippoglossushippoglossus L.) derived from the histone H2A (Birkemoet al., 2003). Other antimicrobial proteins isolated fromfish and having other primary functions include apolipo-proteins A-I and A-II, present in skin or serum of carp(Cyprinus carpio) and active against some fish bacterialpathogens (Concha et al., 2004). These proteins with otherwell established functions appear to be recruited toa second antimicrobial role in nature.
In the present work we purified and identified thefraction of the P. cf henlei mucus responsible for antimi-crobial activity against E. coli,M. luteus and C. tropicalis. Thepurified PcfHb exhibited a lower MIC against gram-
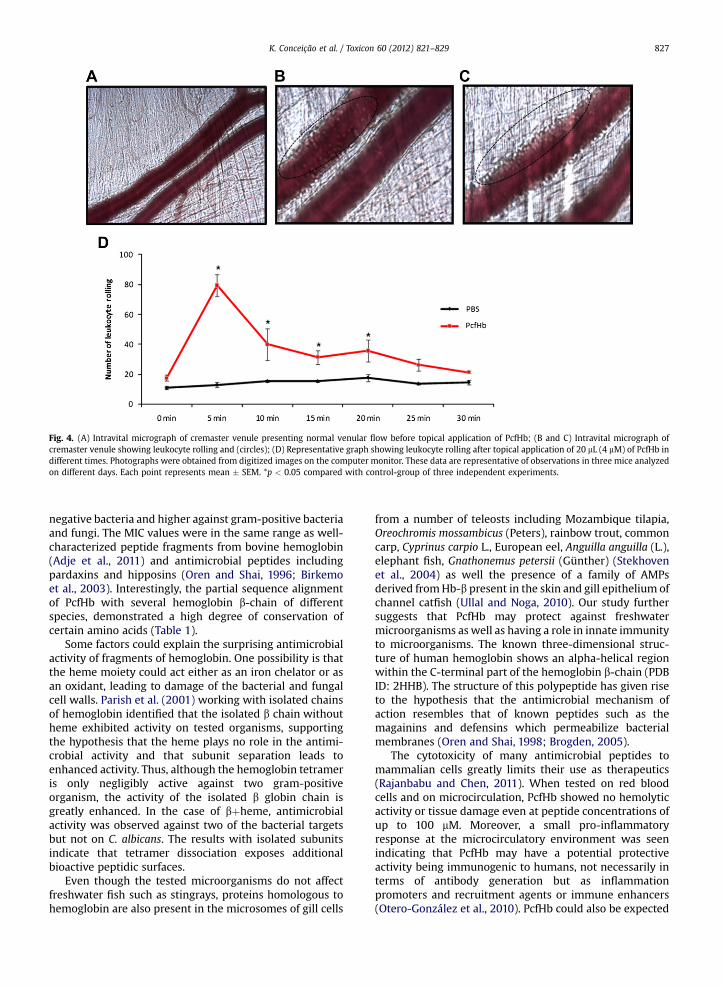
Fig. 4. (A) Intravital micrograph of cremaster venule presenting normal venular flow before topical application of PcfHb; (B and C) Intravital micrograph ofcremaster venule showing leukocyte rolling and (circles); (D) Representative graph showing leukocyte rolling after topical application of 20 mL (4 mM) of PcfHb indifferent times. Photographs were obtained from digitized images on the computer monitor. These data are representative of observations in three mice analyzedon different days. Each point represents mean � SEM. *p < 0.05 compared with control-group of three independent experiments.
K. Conceição et al. / Toxicon 60 (2012) 821–829 827
negative bacteria and higher against gram-positive bacteriaand fungi. The MIC values were in the same range as well-characterized peptide fragments from bovine hemoglobin(Adje et al., 2011) and antimicrobial peptides includingpardaxins and hipposins (Oren and Shai, 1996; Birkemoet al., 2003). Interestingly, the partial sequence alignmentof PcfHb with several hemoglobin b-chain of differentspecies, demonstrated a high degree of conservation ofcertain amino acids (Table 1).
Some factors could explain the surprising antimicrobialactivity of fragments of hemoglobin. One possibility is thatthe heme moiety could act either as an iron chelator or asan oxidant, leading to damage of the bacterial and fungalcell walls. Parish et al. (2001) working with isolated chainsof hemoglobin identified that the isolated b chain withoutheme exhibited activity on tested organisms, supportingthe hypothesis that the heme plays no role in the antimi-crobial activity and that subunit separation leads toenhanced activity. Thus, although the hemoglobin tetrameris only negligibly active against two gram-positiveorganism, the activity of the isolated b globin chain isgreatly enhanced. In the case of bþheme, antimicrobialactivity was observed against two of the bacterial targetsbut not on C. albicans. The results with isolated subunitsindicate that tetramer dissociation exposes additionalbioactive peptidic surfaces.
Even though the tested microorganisms do not affectfreshwater fish such as stingrays, proteins homologous tohemoglobin are also present in the microsomes of gill cells
from a number of teleosts including Mozambique tilapia,Oreochromis mossambicus (Peters), rainbow trout, commoncarp, Cyprinus carpio L., European eel, Anguilla anguilla (L.),elephant fish, Gnathonemus petersii (Günther) (Stekhovenet al., 2004) as well the presence of a family of AMPsderived fromHb-b present in the skin and gill epithelium ofchannel catfish (Ullal and Noga, 2010). Our study furthersuggests that PcfHb may protect against freshwatermicroorganisms as well as having a role in innate immunityto microorganisms. The known three-dimensional struc-ture of human hemoglobin shows an alpha-helical regionwithin the C-terminal part of the hemoglobin b-chain (PDBID: 2HHB). The structure of this polypeptide has given riseto the hypothesis that the antimicrobial mechanism ofaction resembles that of known peptides such as themagainins and defensins which permeabilize bacterialmembranes (Oren and Shai, 1998; Brogden, 2005).
The cytotoxicity of many antimicrobial peptides tomammalian cells greatly limits their use as therapeutics(Rajanbabu and Chen, 2011). When tested on red bloodcells and on microcirculation, PcfHb showed no hemolyticactivity or tissue damage even at peptide concentrations ofup to 100 mM. Moreover, a small pro-inflammatoryresponse at the microcirculatory environment was seenindicating that PcfHb may have a potential protectiveactivity being immunogenic to humans, not necessarily interms of antibody generation but as inflammationpromoters and recruitment agents or immune enhancers(Otero-González et al., 2010). PcfHb could also be expected

K. Conceição et al. / Toxicon 60 (2012) 821–829828
to function in conjunction with the histone-like proteinsthat were found in the same epithelial mucus in thisstingray (data not shown), providing a strong line of innatehost defence against eukaryotic as well as prokaryoticpathogens.
Although innate immunity to microbial infection isa property common to almost all forms of life, it was quiteunexpected that hemoglobin, one of the most well-char-acterized proteins due to its function in oxygen transport,should contribute to innate immunity. However, recentstudies have identified Hb-derived AMPs from humans andother animals; some of which were more inhibitory toeukaryotes than bacteria (Ullal et al., 2008; Ullal and Noga,2010). In view of the fact that different hemoglobin-derivedpeptide fragments exhibit diverse antibiotic activities, it isconceivable that, in addition to its role in oxygen transporthemoglobin functions as an important multi-defense agentagainst a wide range of microorganisms. In conclusion, wehave shown for the first time that a protein with highsequence similarity to the hemoglobin b chain is anantimicrobial polypeptide naturally occurring in the mucusof stingrays. This finding is in accordance with the data ofParish et al. (2001) which identified the region containingthe antimicrobial fragments at the amino acid sequences offree b -hemoglobin chain with a greater activity on gram-positive bacteria. Due to the broad antimicrobial action ofPcfHb against Gram-positive and Gram-negative bacteriaand yeast and its pro-inflammatory action, it may be sug-gested that this antimicrobial polypeptide could playa significant role in the innate immune response of this andother fishes. Beyond that, it might ultimately lead to newtherapies with novel engineered antimicrobial moleculesthat support an integrative and complementary actionagainst pathogens related to infectious diseases invertebrates.
Acknowledgement
This work has been supported by a FAPESP (2007/55148-9), CNPq and FAPEMIG. Alessandra Cardoso isacknowledged for technical assistance, Marta Maria BatistaRibeiro and Vera Luisa Neves for helpful discussions.
Conflict of interest statement
None to declare.
References
Adje, E.Y., Balti, R., Kouach, M., Dhulster, P., Guillochon, D., Nedjar-Arroume, N., 2011. Obtaining antimicrobial peptides by controlledpeptic hydrolysis of bovine hemoglobin. Int. J. Biol. Macromol. 49,143–153.
Alexander, J.B., Ingram, G.A., 1992. Noncellular nonspecific defencemechanism of fish. Annu. Rev. Fish. Dis. 2, 249–279.
Baez, S., 1973. An open cremaster muscle preparation for the study ofblood vessels by in vivo microscopy. Microvasc. Res. 5, 384–396.
Bergsson, G., Agerberth, B., Jornvall, H., Gudmundsson, G.H., 2005. Isola-tion and identification of antimicrobial components from theepidermal mucus of Atlantic cod (Gadus morhua). FEBS J. 272, 4960–4969.
Birkemo, G.A., Lüders, T., Andersen, Ø, Nes, I.F., Nissen-Meyer, J., 2003.Hipposin, a histone-derived antimicrobial peptide in Atlantic halibut(Hippoglossus hippoglossus L.). Biochim. Biophys. Acta 1646, 207–215.
Bradford, M.M., 1976. A rapid and sensitive method for quantitation ofmicrogram quantities of protein utilizing the principle of protein dyebinding. Anal. Biochem. 72, 248–254.
Brogden, K.A., 2005. Antimicrobial peptides: pore formers or metabolicinhibitors in bacteria? Nat. Rev. Microbiol. 3, 238–250.
Bulet, P., Dimarcq, J.L., Hetru, C., Lagueux, M., Charlet, M., Hegy, G., VanDorsselaer, A., Hoffmann, J.A., 1993. A novel inducible antimicrobialpeptide of Drosophila carries an O-glycosylated substitution. J. Biol.Chem. 268, 14893–14897.
Carvalho, M.R., Lovejoy, N.R., Rosa, R.S., 2003. Family Potamotrygonidae.In: Reis, R.E., Kullander, S.O., Ferraris, C.J. (Eds.), Checklist of theFreshwater Fishes of South and Central America (CLOFFSCA). Edi-pucrs, Porto Alegre, pp. 22–29.
Cole, A.M., Darouiche, R.O., Legarda, D., Connell, N., Diamond, G., 2000.Antimicrob. Agents Chemother. 44, 2039–2045.
Conceição, K., Santos, J.M., Bruni, F.M., Klitzke, C.F., Marques, E.E.,Borges, M.H., Melo, R.L., Fernandez, J.H., Lopes-Ferreira, M., 2009.Characterization of a new bioactive peptide from Potamotrygon gr.orbignyi freshwater stingray venom. Peptides 30, 2191–2199.
Concha, M.I., Smith, V.J., Castro, K., Bastias, A., Romero, A., Amthauer, R.J.,2004. Apolipoproteins A-I and A-II are potentially important effectorsof innate immunity of the teleost fish Cyprinus carpio. Eur. J. Biochem.271, 2984–2990.
Ehret-Sabatier, L., Loew, D., Goyffon, M., Fehlbaum, P., Hoffmann, J.A., VanDorsselaer, A.V., Bulet, P., 1996. Characterization of novel cysteine-richantimicrobial peptides from scorpion blood. J. Biol. Chem. 271,29537–29544.
Fernandes, J.M.O., Kemp, G.D., Molle, M.G., Smith, V.J., 2002. Anti-microbial properties of histone H2A from skin secretions of rainbowtrout, Oncorhynchus mykiss. Biochem. J. 368, 611–620.
Halstead, B.W., 1970. Poisonous and Venomous Marine Animals of theWorld. The Darwin Press, Princeton, New Jersey, NJ. 1988:1168.
Hellio, C., Pons, A.M., Beaupoil, C., Bourgougnon, N., Gal, Y.L., 2002.Antimicrobial, antifungal and cytotoxic activities of extracts fromfish epidermis and epidermal mucus. Int. J. Antimicrob. Agents 20,214–219.
Hellman, U., Wernstedt, C., Góñez, J., Heldin, C.H., 1995. Improvement ofan “In-Gel” digestion procedure for the micropreparation of internalprotein fragments for amino acid sequencing. Anal. Biochem. 224,451–455.
Ingram, G.A., 1980. Substances involved in the natural resistance of fish toinfection – a review. J. Fish Biol. 16, 23–60.
Klesius, P.H., Shoemaker, C.A., Evans, J.J., 2008. Flavobacterium columnarechemotaxis to channel catfishmucus. FEMSMicrobiol. Lett. 288, 216–220.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assemblyof the head of bacteriophage T4. Nature 227, 680–685.
Lomonte, B., Lundgren, J., Johansson, B., Bagge, U., 1994. The dynamics oflocal tissue damage induced by Bothrops asper snake venom andmyotoxin II on the mouse cremaster muscle. An intravital and elec-tron microscopic study. Toxicon 32, 41–55.
Monteiro-Dos-Santos, J., Conceição, K., Seibert, C.S., Marques, E.E., IsmaelSilva Jr., P., Soares, A.B., Lima, C., Lopes-Ferreira, M., 2011. Studies onpharmacological properties of mucus and sting venom of Potamo-trygon cf. henlei. Int. Immunopharmacol. 11, 1368–1377.
Mozumder, M.M.H., 2005. Antimicrobial Activity in Fish Mucus fromFarmed Fish. Norwegian College of Fishery Science, University ofTromsø, Norway.
Negus, V.E., 1963. The functions of mucus. Acta Otolaryngol. 56, 204–214.Oren, Z., Shai, Y., 1996. A class of highly potent antimicrobial peptides
derived from pardaxin, a pore-forming peptide isolated from Mosessole fish Pardachirus marmoratus. Eur. J. Biochem. 237, 303–310.
Oren, Z., Shai, Y., 1998. Mode of action of linear amphipathic alpha-helicalantimicrobial peptides. Biopolymers 47, 451–463.
Otero-González, A.J., Magalhães, B.S., Garcia-Villarino, M., López-Abarrategui, C., Sousa, D.A., Dias, S.C., Franco, O.L., 2010. Antimicrobialpeptides from marine invertebrates as a new frontier for microbialinfection control. FASEB J. 24, 1320–1334.
Parish, C.A., Jiang, H., Tokiwa, Y., Berova, N., Nakanishi, K., McCabe, D.,Zuckerman, W., Xia, M.M., Gabay, J.E., 2001. Broad-spectrum antimi-crobial activity of hemoglobin. Bioorg. Med. Chem. 9, 377–382.
Rajanbabu, V., Chen, J.Y., 2011. Applications of antimicrobial peptides fromfish and perspectives for the future. Peptides 32, 415–420.
Ramos, A.D., Conceição, K., Silva Jr., P.I., Richardson, M., Lima, C., Lopes-Ferreira, M., 2012 Feb 21. Specialization of the sting venom and skinmucus of Cathorops spixii reveals functional diversification of thetoxins. Toxicon.
Richards, R.C., O’Neil, D.B., Thibault, P., Ewart, K.V., 2001. Histone H1. Anantimicrobial protein of Atlantic salmon (Salmo salar). Biochem.Biophys. Res. Commun. 284, 549–555.

K. Conceição et al. / Toxicon 60 (2012) 821–829 829
Robinette, D., Wada, S., Arroll, T., Levy, M.G., Miller, W.L., Noga, E.J., 1998.Antimicrobial activity in the skin of channel catfish Ictalurus punc-tatus: characterization of broad-spectrum histone-like antimicrobialproteins. Cell. Mol. Life Sci. 54, 467–475.
Shephard, K.L., 1994. Functions for fish mucus. Rev. Fish Biol. Fish 4,401–429.
Stekhoven, F., Bonga, S., Flik, G., 2004. Extranuclear histones in teleostgills: an evolutionary study. Fish Physiol. Biochem. 30, 201–211.
Su, Y., 2011. Isolation and identification of pelteobagrin, a novel antimi-crobial peptide from the skin mucus of yellow catfish (Pelteobagrusfulvidraco). Comp.Biochem.Physiol. B Biochem.Mol. Biol.158,149–154.
Subramanian, S., Ross, N.W., MacKinnon, S.L., 2009. Myxinidin, a novelantimicrobial peptide from the epidermal mucus of hagfish, Myxineglutinosa L. Mar. Biotechnol. (NY) 11, 748–757.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G.,1997. The CLUSTAL_X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. NucleicAcids Res. 25, 4876–4882.
Ullal, A.J., Litaker, R.W., Noga, E.J., 2008. Antimicrobial peptides derivedfrom hemoglobin are expressed in epithelium of channel catfish(Ictalurus punctatus, Rafinesque). Dev. Comp. Immunol. 32, 1301–1312.
Ullal, A.J., Noga, E.J., 2010. Antiparasitic activity of the antimicrobialpeptide HbbetaP-1, a member of the beta-haemoglobin peptidefamily. J. Fish. Dis. 33, 657–664.
Zhao, X., Findly, R.C., Dickerson, H.W., 2008. Cutaneous antibody-secreting cells and B cells in a teleost fish. Dev. Comp. Immunol. 32,500–508.









![EFFICACY OF ANESTHETICS FOR REDUCING STRESS IN … · Potamotrygon cf. histrix (Cururu stingray) fish [49]. Transportation may be considered a strong stressor to catfish [50]. Due](https://img.pdfslide.us/doc/110x75/5c4ac22893f3c34c50655dc3/efficacy-of-anesthetics-for-reducing-stress-in-potamotrygon-cf-histrix-cururu.jpg)



















