Embed Size (px)
Citation preview

wel come

Non-host Resistance in Plants:
New insights
Jayappa PALB4255
JayappaID.No: 4255Sr. M.Sc (Agri)

OUTLINE OF SEMINAR
Introduction
Components
Types
Applications
Case studies
Conclusion

INTRODUCTION
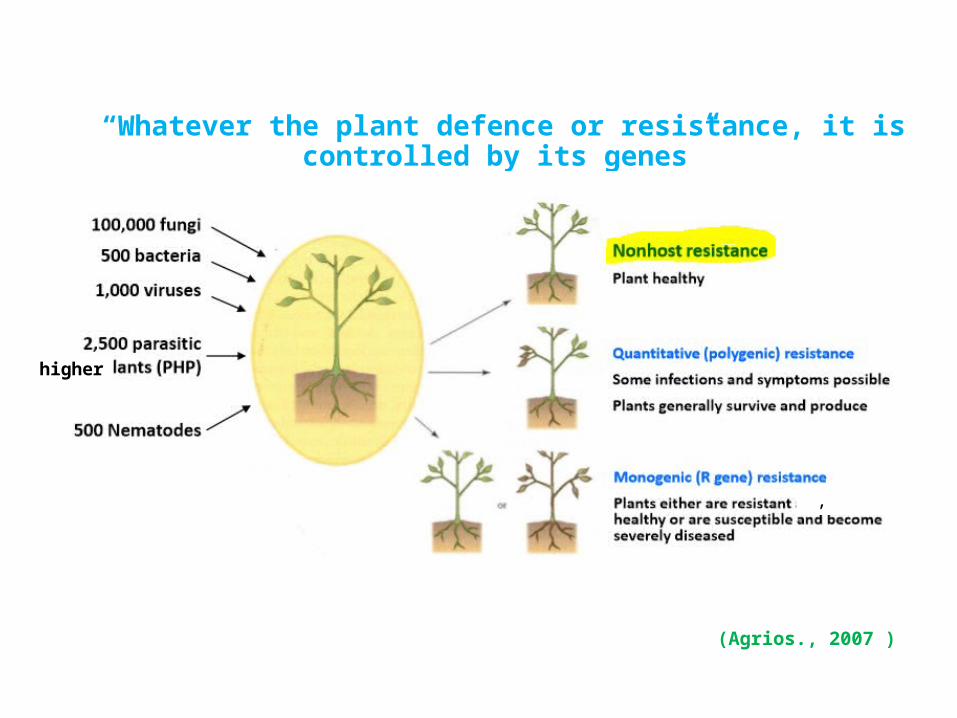
“Whatever the plant defence or resistance, it is controlled by its genes”
(Agrios., 2007 )
higher
,

Non Host response of the cell/ nucleus
against the pathogen . Ultimately
no disease to the plants
Host response of the cell/ nucleus against
the pathogen and cause disease
Non Host response of the cell/ nucleus
against the pathogen . Ultimately no disease
to the plants
Host response of the cell/ nucleus against the pathogen as R
Gene mediated resistance develops
and cause no disease
Nucleus response
NH pathogen signals
NH
pathogen
Avr
path
ogen
Vir
pathog
en Infection/
diseaseIndu
ced
defe
nce
&
PTI
Prefo
rmed
defen
c &
phys
ical
barr
iers
R- gene m
ediated im
munity
( Shamim et al., 2013)
No
dise
ase
No disease
Interaction and disease development phenomenon in plant

Is a broad spectrum resistance Resistance of an entire plant species to all isolates of a microbial species Preformed barriers such as cell wall, cuticle, phytoanticipins Induced defense responses such as lignin accumulation, production of
antimicrobials like phytoalexins, HR response, induction of pathogenesis-related (PR) proteins
Eg., Barley is typically susceptible to P. hordei, to Which wheat is a nonhost. The reverse is true for P. triticina
Non host resistance Plant Innate Immunity, Species Resistance, Durable resistance,
Basal defense and Nonspecific resistance

Components of Non host resistance
Preformed or passive defence mechanisms
Inducible plant defence mechanisms
Plant defense signalling
Broad-spectrum disease resistance genes

Preformed or passive defense mechanisms
Sheath around a hypha
Cork layer Phenolic compounds
T, Tyloses in xylem vessels
Abscission layer

Necrotic defense reaction in a cell
N, nucleus; PS, protoplasmic strands; Z, zoospore; H, hypha; G, granular material; NC, necrotic cell
( Tomiyama et al., 1999 )
Resistant potato variety infected by Phytophthora infestans.

Plant Defense Signalling
(Yang et al., 1999)
Non host pathogen

Gene/protein Function in non-host resistance (NHR) References
PAMPsPep-13 Induces defense responses in non-host plants
like PotatoNurnberger et al. (1994)
Harpin (Hrp Z) Elicits HR-like cell death and defense responses in various plants
He et al. (1993)
Genes PEN1(Penetration1)/ROR2
This gene is involved in timely deposition of papillae during non-host interactions.
Collins et al. (2003)
NHO1 Required for NHR of Arabidopsis against Pseudomonas syringae pv. phaseolicola
Kang et al. (2003)
R-gene mediated genes SGT1 Silencing of SGT1 in N. benthamiana
compromises NHR against P. syringae pv. maculicola and Xanthomonas axonopodis pv. Vesicator
Peart et al.(2002)
Heat-shock proteins (Hsps)
Silencing of Hsp90 and Hsp70 in N. benthamiana individually compromised NHR against P. cichorii.
Kanzaki et al. (2003)
Genes involved in NHR

Types of Non host resistance
( Mysore et al.,2004)

Examples of type I
( Mysore et al.,2004)
Pathogen Strain NH plants Symptoms
P. s. pv. syringe B76 Arabidopsis None
P.s. pv. phaseolicola S2 N. tabacum None
P.s.pv.delphini PDDCC529 Arabidopsis None
P. infestans 88069 N. clevalandi None
P. infestans 88069 N. tabacum cv. xanthi None

Cont..
( Mysore et al.,2004)
Type II non host resistance
Pathogen Strain NH plants Symptoms
P. s.pv. tomato DC3000 N. tabacum HR
P. s. pv. glycinea PG4180 N. tabacum HR
P. s. pv. pisi ATCC 11055 N. tabacum HR
P. s. pv. syringae 61 N. tabacum HR
P. cichorii 83-1 Arabidopsis HR

Application of NHR in Agriculture
Application References
1. Successful use of a PRR gene, EFR, from Arabidopsis in N. benthamiana and tomato to reduce the growth of their respective host pathogens
(Lacombe et al., 2010)
2. R gene, Bs2, from pepper has been shown to impart resistance to X. campestris pv. vesicatoria in tomato (Tai et al., 1999)
3. Resistance to Erwinia carotovora, the causal agent of potato (Solanum tuberosum) tuber soft rot, was achieved by replacing chromosome 8 of potato with the corresponding chromosome from its wild relative Solanum brevidens
(Tek et al.,2004)

CASE STUDIES

Objective : To know the function of MPKs in NHR to M.oryzae in A.thaliana

Quantitative analysis of NHR to M. oryzae in A. thaliana
Mean frequency of M. oryzae penetration into A. thaliana mutants @ 48hpi (Expressed as a total no. of infection sites.)
Mean length of infection hyphae measured @48hpi
(Okawa and Ishikawa, 2013)

Quantitative analysis of Post penetration resistance to M.oryzae in A.thaliana
I. Cell wall penetrationII. Establishment of infection
hyphaeIII. Elongation of infection
hyphaeIV. Branch formation on
infection hyphae
Microscopic views of infection sites in Arabidopsis mutants
Light microscopic view @ 48hpi
Fluorescence microscopic
view @ 48hpi
pen2
Pen2 mpk6
48hpi
(Okawa and Ishikawa, 2013)

Objective: Detection of DNAse and their activity in Non host resistance
Non-host to Fusarium solani f.sp. phaseoli (Fsph) Host to Fusarium solani f.sp. pisi (Fspi )
Pea
,2013
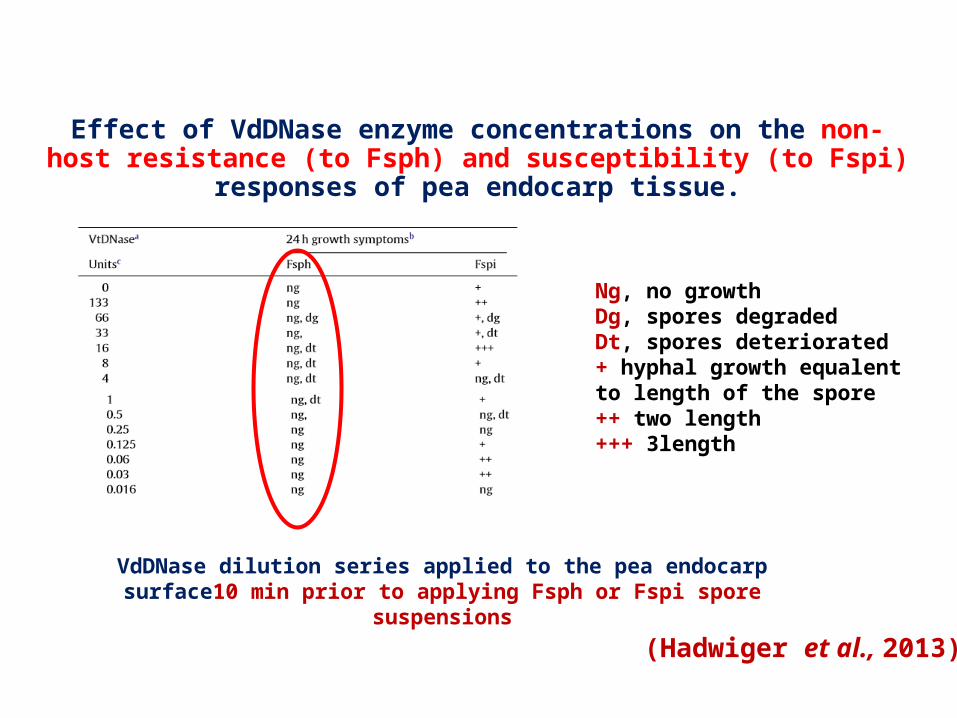
Effect of VdDNase enzyme concentrations on the non-host resistance (to Fsph) and susceptibility (to Fspi) responses of pea endocarp tissue.
Ng, no growthDg, spores degradedDt, spores deteriorated+ hyphal growth equalent to length of the spore++ two length+++ 3length
VdDNase dilution series applied to the pea endocarp surface10 min prior to applying Fsph or Fspi spore suspensions
(Hadwiger et al., 2013)

VdDNase induces pisatin accumulation
Vd DNase assay and pisatin eliciting activity:(A) Pod halves (0.5 g fr. wt.) were treated
with 2 ml of superdex 75 fractions. (B) Superdex 75 fractions (2.0 ml) were incubated in the standard DNase buffer for 10 min wit 0.2micro gram of plasmid DNA.
DNase activity
24 h
(Hadwiger et al., 2013)

A northern blot analysis of the expression of PR gene, DRR49
Treatments: 1 = 02 = 5000 Units VdDNase, 3 = 2500 units VdDNase, 4 = 1250 units VdDNase 5 = 06 = 5000 units VdDNase7 = 2500 units VdDNase 8 = 1250 units VdDNase
4Hr
7Hr
mRNA levels
(Hadwiger et al., 2013)

Objective : To confirm that PRR activity is retained after its transfer between two plant families
EFR(Elongation factor receptor, PRR) Elf18( Elongation factor 18aa)
EF-TU(Elongation factor thermo unstable)EF-Tu is highly conserved in all Phytopathogenic bacteria
2010

Transgenic expression of EFR in N. benthamiana and tomato
N. benthamiana plants S. lycopersicum plants
Oxidative Burst triggered by 100 nM elf18 or flg22
Four-week-old Four-week-old
Oxidative Burst triggered by 100 nM elf18 or flg22
(Lacombe et al., 2010)RLU= measures cleanliness of leaf surface
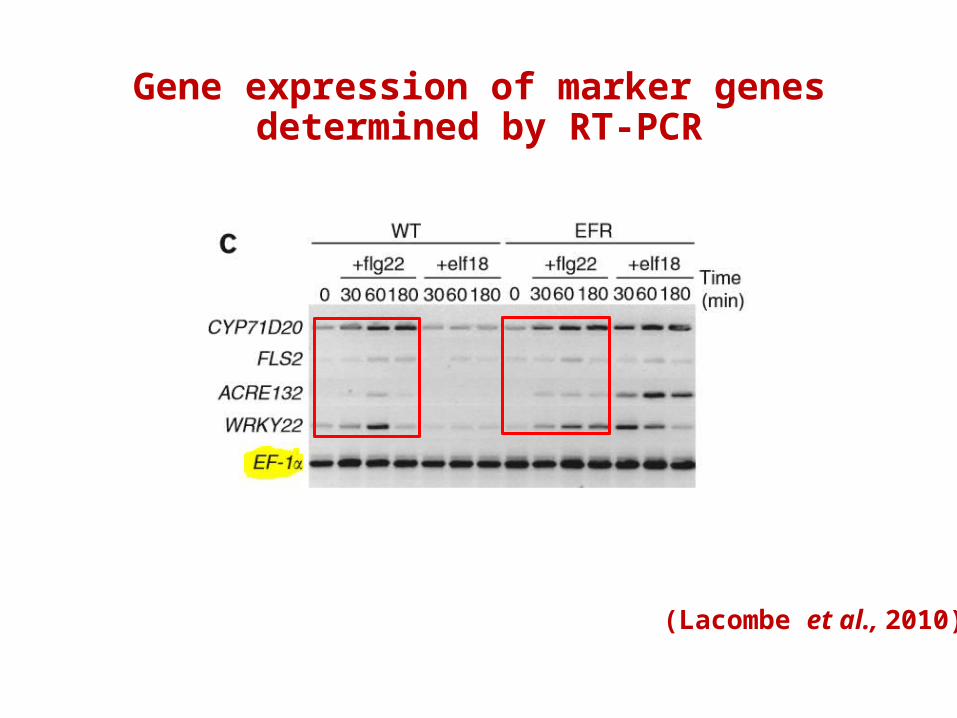
Gene expression of marker genes determined by RT-PCR
(Lacombe et al., 2010)

Transgenic expression of EFR in N. benthamiana
a. Infection with P. syringae pv. syringae (Pss) B728ab. Infection with P. syringae pv. tabaci (Pta) 11528
(Lacombe et al., 2010)

N. benthamiana infection with A. tumefaciens A281
(Lacombe et al., 2010)

Transgenic expression of EFR in tomato
(Lacombe et al., 2010)

Objective: 1. Identification of non-adapted P. cucumerina isolates 2. To find out the role of different trp-derived compounds.

Plectosphaerella cucumerina isolates Pc1187 and Pc2127 are non-adapted necrotrophic fungi on Arabidopsis wild-
type plants
Lactophenol Trypan Blue staining of
inoculated leaves at 12 and 20 h post
inoculation
Relative quantification of fungal DNA
Average disease rating
(Sanchez- Vallet et al., 2010)

Quantitative real-time PCR quantification of fungal DNA (Pcb-tubulin)
Average disease rating
Lactophenol Trypan Blue staining of inoculated leaves
(Sanchez- Vallet et al., 2010)

Objective: 1. Inoculation with a Xoo results in induction of HR and
nonhost resistance2. Exogenous supply of accelerated Xoo induced HR
,2015

Effect of the Xoo inoculum concentration on HR induction.
Xoo at 1×, cfuconcentrations was inoculated in fully expanded N. benthamianaleaves. The inoculated plants were grown at 28 °C.
24h
(Li et al., 2015)

is essential for HR induction during the interaction between Xoo and N. benthamiana
Effect of exogenous on Xoo-induced HR.
Effect of exogenous catalase on Xoo-induced HR
DAB staining for detection
O
O
(Li et al., 2015)

Ability of three Xoo-derived T3SS gene mutants to induce HR and accumulation in N. benthamiana
DAB staining for detection
(Li et al., 2015)

Bacterial numbers were counted in catalase treated leaves inoculated with Xoo and the Δhpa1 mutant
(Li et al., 2015)

CONCLUSION






























