Embed Size (px)
DESCRIPTION
In response to pathogen attack, plants have evolved sophisticated defense mechanisms to delay or arrest pathogen growth.Unlike animals, plants lack a circulating immune system recognizing microbial pathogens. Plant cells are more autonomous in their defense mechanisms and rely on the innate immune capacity of each cell and systemic signals that disseminate from infection sites (Jones and Dangl, 2006). Plant innate immunity consists of preformed physical and chemical barriers (such as leaf hairs, rigid cell walls, pre-existing antimicrobial compounds) and induced defenses. Should an invading microbe successfully breach the pre-formed barriers, it may be recognized by the plant, resulting in the activation of cellular defense responses that stop or restrict further development of the invader.
Citation preview

The molecular basis of plant resistance and defense responses to pathogens: Current status
Sruthi.N

Introduction
Many plant-associated microbes are pathogens that impair plant growth and reproduction
Pathogens may proliferate in intercellular spaces (the apoplast) after entering through stomata or hydathodes (bacteria), enter plant epidermal cells, or extend hyphae onto the plant cells (fungi)
Innate immune receptors in plants detect the presence of microbial pathogens and trigger defense responses to terminate or restrict pathogen growth

Elicitors of defence responses
Any substance that has the capability of activating defense responses in plants
Include components of the cell surface as well as excreted metabolites
Elicitors
General Race specific
a) Oligosaccharide elicitors a)avr gene products
b) Protein/peptide elicitors(Ebel et al.,1998)

Perception of elicitor signals
Binding proteins:
Oligosaccharide-binding sites: -Specific glucan-binding sites on soybean root plasma
membranes -high-affinity binding sites for chitin fragments in tomato, rice
Glycopeptide- and peptide-binding sites: -Binding sites for peptidoglycans have been identified in wheat
plasma membranes

Plant defense to pathogens
• Plants respond to infection using a two-branched innate immune system
-Recognition and response to molecules common to many classes of microbes (basal disease resistance)
-Response to pathogen virulence factors (Liu et al.,2008)

Basal defense
Triggered by trans-membrane receptors that recognize conserved molecules released by a variety of (unrelated) microbes
Include cell wall fragments, chitin or peptide motifs in bacterial flagella - PAMPs or MAMPs
PAMP- triggered immunity (PTI)
(Liu et al.,2008)

Secondary defense response
Against virulence effector proteins produced by pathogens
Effector –triggered immunity (ETI)
Mediated by resistance (R) proteins
(Liu et al., 2008)
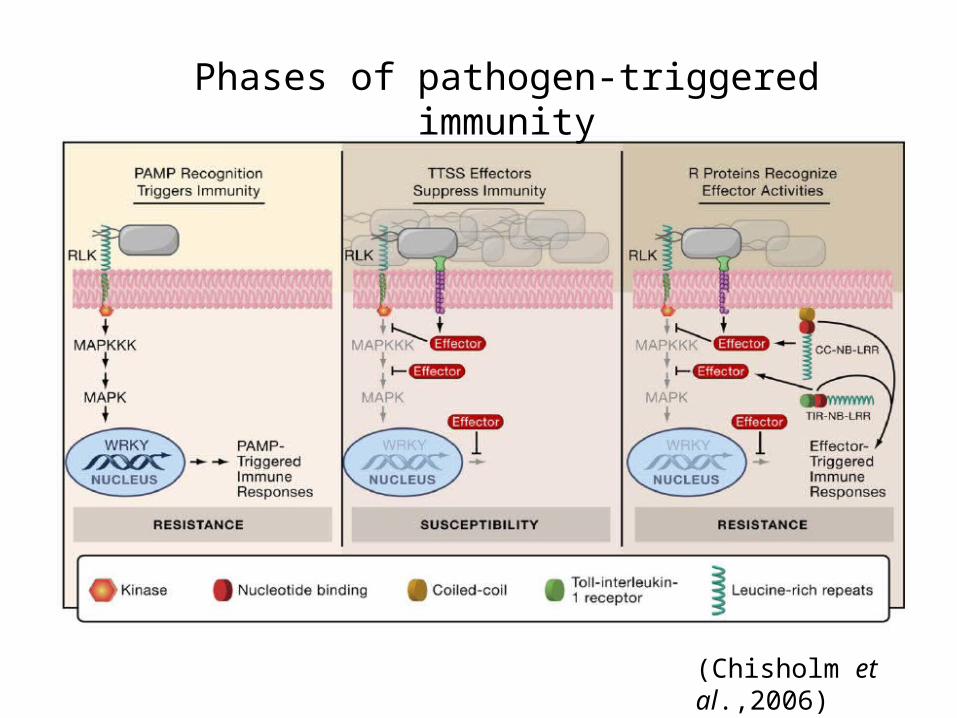
Phases of pathogen-triggered immunity
(Chisholm et al.,2006)

Concepts regarding pathogen recognition and defense
Gene-for-gene resistance (Flor , 1947)
Guard hypothesis (Beizen et al.,1998)

Gene-for-gene resistance
For resistance to occur, complementary pairs of dominant genes, one in the host and the other in the pathogen, are required (incompatibility)
A loss or alteration to either the plant resistance (R) gene or the pathogen avirulence (Avr) gene leads to disease (compatibility)
(Hammond-Kosack et al., 1997)
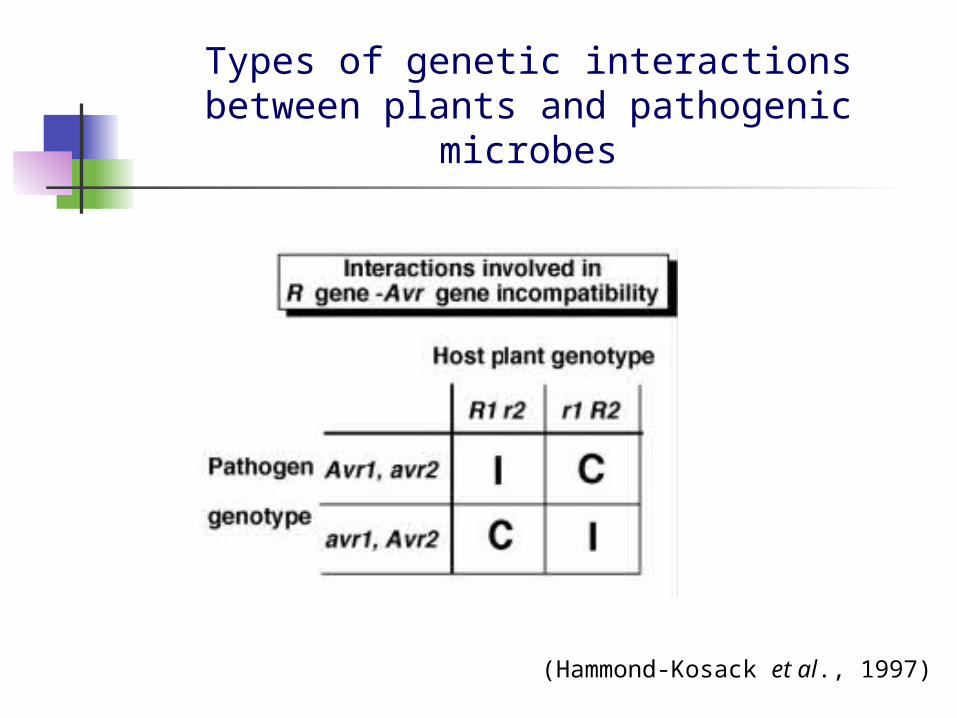
Types of genetic interactions between plants and pathogenic microbes
(Hammond-Kosack et al., 1997)

Plant disease resistance genes
Encode proteins that recognize Avr-gene-dependent ligands
Activate signaling cascade(s) that coordinate the initial plant defense responses to impair pathogen ingress
Capacity for rapid evolution of specificity
Common feature of resistance proteins is a leucine-rich repeat
(Hammond-Kosack et al., 1997)
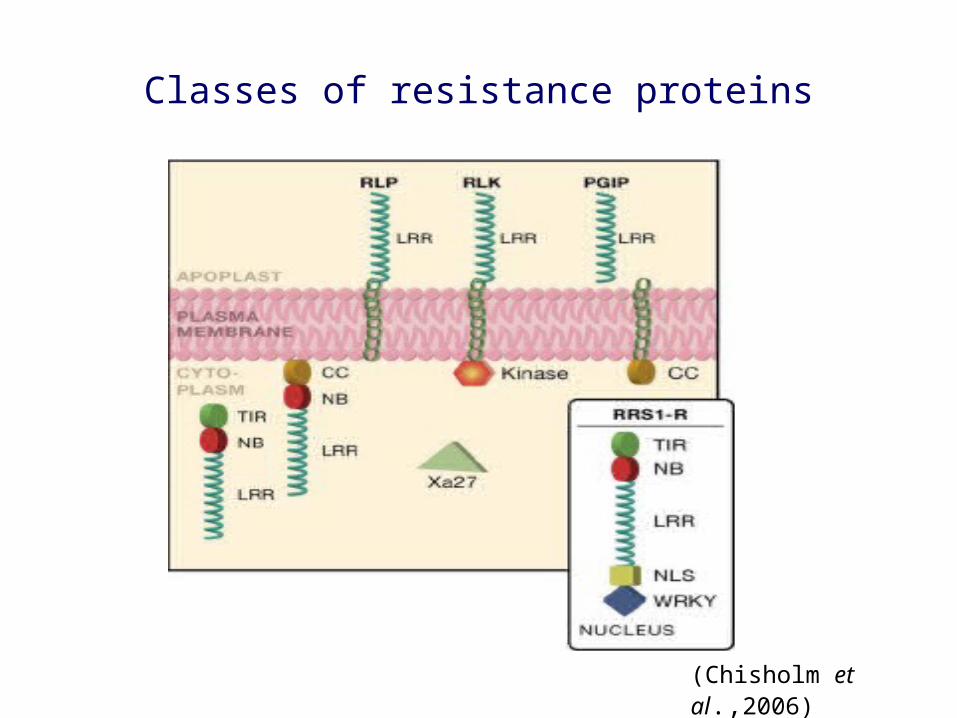
Classes of resistance proteins
(Chisholm et al.,2006)

Extracellular LRR class of R genes
Have classic receptor-kinase formats - an extracelluar LRR, a membrane spanning region and an intracellular protein kinase domain
Against pathogens that have an extracellular lifestyle
Examples: rice Xa21 against Xanthomonas, cf genes of tomato against Cladosporium fulvum.

NB-LRR R proteins
Consists of four domains connected by linkers
A leucine-rich repeat domain (LRR) fused to a nucleotide binding (NB) domain
NB-LRR core equipped with variable amino- and carboxy-terminal domains
(Tameling et al., 2008)

NB-LRR subfamilies
TIR NB-LRRs (Toll/interleukin-1 receptor like NB-LRRs)
CC-NB-LRRs (Coiled Coil NB-LRRs) Solanaceous domain (SD) BED zinc finger DNA-binding domain
(Tameling et al.,2008)
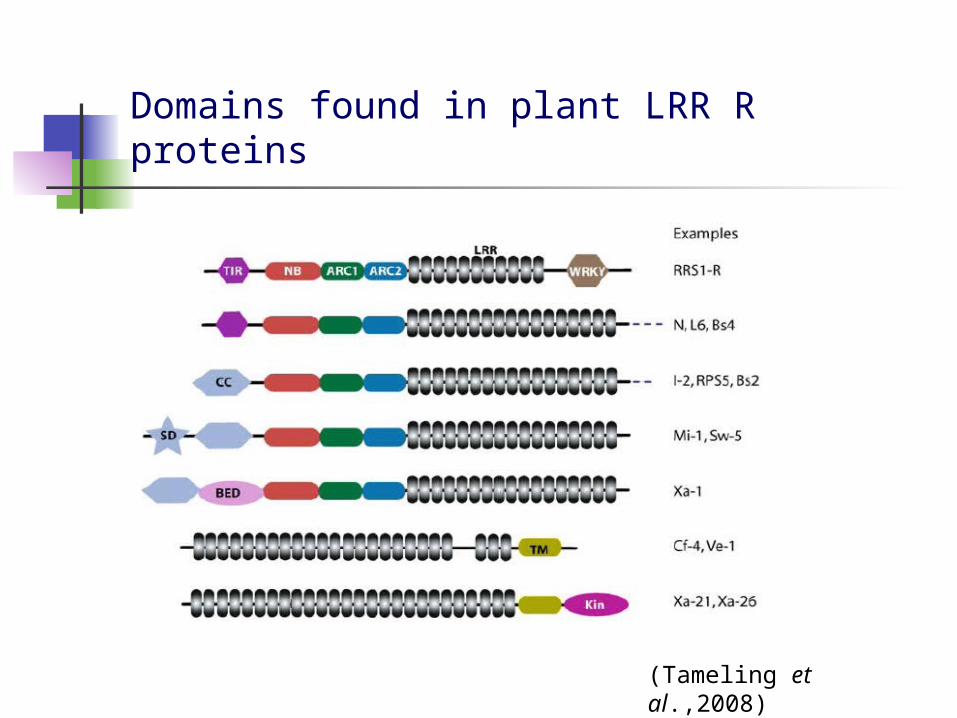
Domains found in plant LRR R proteins
(Tameling et al.,2008)

Deviations from gene-for-gene concept
One R gene may confer specificity to more than one ligand- RPM1 in Arabidopsis confers resistance against P.syringae expressing
either avrRpm1 and avrB
More than one R gene may exist for a given Avr gene- Pto and Prf genes encode biochemically distinct components of the
same pathway- Two genes at the Cf-2 locus furnish identical functions (Bent, 1996)

Guard hypothesis
Key points
a) An effector acting as a virulence factor has a target(s) in the host
b) By manipulating or altering this target(s) the effector contributes to pathogen success in susceptible host genotypes
c) Effector perturbation of a host target generates a “pathogen induced modified self” molecular pattern, which activates the corresponding NB-LRR protein, leading to ETI
(Jones et al.,2006)
a) 9
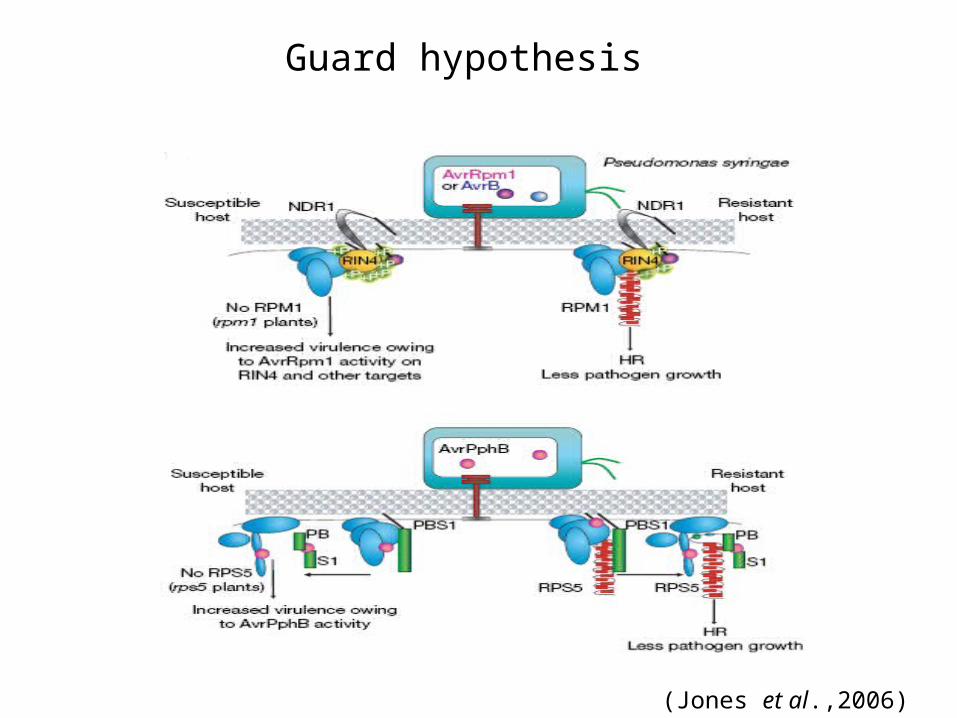
Guard hypothesis
(Jones et al.,2006)

Plant defense responses
Hypersensitive response
Production of reactive oxygen species
Cell wall fortification
Production of antimicrobial metabolites (phytoalexins)
Defense signal transduction
Synthesis of enzymes harmful to pathogen (eg. chitinases, glucanases)
(Nurnberger et al.,2006)

Types of activated defense responses
(A) Hypersensitive response in single lettuce mesophyll cells penetrated by haustoria of an incompatible isolate of the biotrophic fungus Bremia lactucae B) H2O2 generation in lettuce cell walls, in the vicinity of the incompatible bacterium P. syringae pv phaseolicola(C) Papillae formation. Papillae develop beneath the penetration peg (PP) and germinating spore (S) of an avirulent isolate of the biotrophic fungusErysiphe graminis f sp hordei on barley leaves expressing the Mlg gene.
(Hammond-Kosack et al., 1997)

Defense signaling pathways
SA-dependant signaling
Effective against biotrophic pathogens
Activated by the initiation of HR in plants
Rise in SA levels dissociation of NPR1 oligomers to monomers interaction with TGA-type transcription factors
activation of PR gene expression
TGAs 2,5, and 6 and WRKY70 required for full expression of PR-1 (Glazebrook, 2005)

Defense signaling pathways
JA- and ET- dependent signaling
Effective against necrotrophic pathogens
Increase in JA levels and induction of effector genes (PDF1.2, VSP1)
Induction of transcription factors ERF1, RAP2.6 and JIN1 which activates many defense related genes
Some JA regulated genes also regulated by ET (PDF1.2 , ERF1) (Glazebrook, 2005)

Defense signaling pathways
JA- and ET- dependent signaling
JA levels regulated by cellulose synthases
JAR1 involved in conversion of JA to active form by conjugation with amino acids like isoleucine
In Arabidopsis, all activities of JA requires the function of CO11.
Some responses to JA require the function of an MAP kinase encoded by MPK4
(Glazebrook, 2005)

Defense signaling pathways
Cross-talk between SA and JA/ET signaling
Helps the plant to minimize energy costs and create a flexible signaling network that allows the plant to finely tune its defense response to the invaders encountered
Most reports indicate a mutually antagonistic interaction between SA- and JA dependent signaling.
(Koornneef et al., 2008)

Molecular players in SA/JA crosstalk
NPR1, required for transduction of SA signaling is a suppressor of JA response
Expression of GRX480 is SA inducible and dependent on NPR1
Overexpression of GRX480 completely abolished MeJA-induced PDF1.2 expression, but does not affect the induction of the JA-responsive genes LOX2 and VSP2
(Koornneef et al., 2008)

Molecular players in SA/JA crosstalk
Overexpression of WRKY70 caused enhanced expression of SA-responsive PR genes and suppressed methyl jasmonate (MeJA)-induced expression of PDF1.2
MPK4 is a negative regulator of SA signaling and a positive regulator of JA signaling in Arabidopsis (Koornneef et al., 2008)

PR proteins
Coded by host plants as a response to pathological or related situations
Accumulate not only locally in the place of infection, formed systemically following infection by pathogens.
Wide array of functions: hydrolases, transcription factors, protease inhibitors etc.
(Scherer et al.,2005)
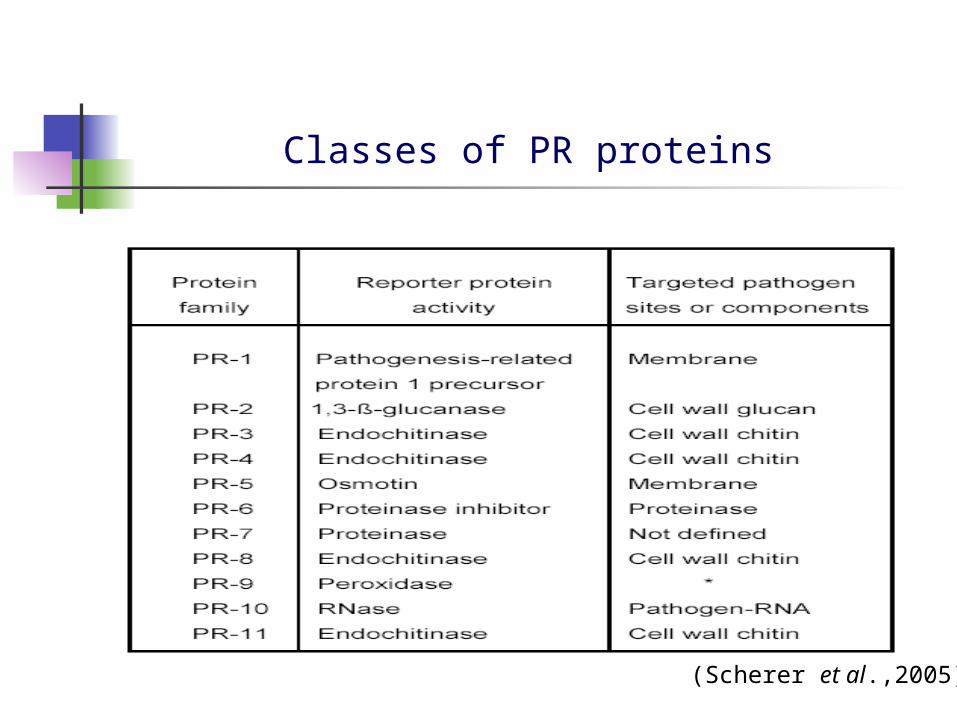
Classes of PR proteins
(Scherer et al.,2005)

Non-Host Resistance
Two mechanisms In case of a potentially new host, pathogen’s effectors could be ineffective, resulting in
little or no supression of PTI, and failure of pathogen growth
One or more of the effector complement of the would-be pathogen could be recognized by the NB-LRR proteins of plants other than it’s coadapted host , resulting in ETI
Arabidopsis resistance to non-adapted powdery mildew Blumeria graminis f. sp. hordei (Jones et al., 2006)
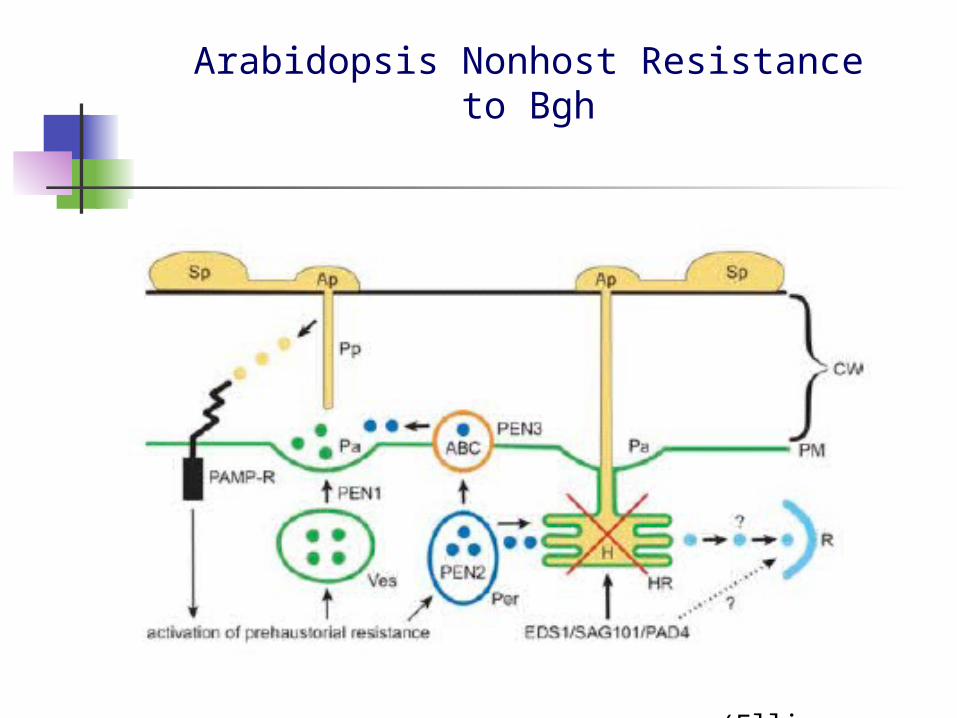
Arabidopsis Nonhost Resistanceto Bgh
(Ellis, 2006)

How R genes initiate defense signaling pathways R proteins recognize pathogen effectors in the cytoplasm
Effector perception may result in altered intra and inter-molecular R protein interactions, including oligomerization.
Activated R proteins cycle into the nucleus and directly bind transcriptional repressors of innate immunity, resulting in transcriptional reprogramming of the plant cell
Studies conducted for N, MLA,RPS4, and RX (Liu et al., 2008)
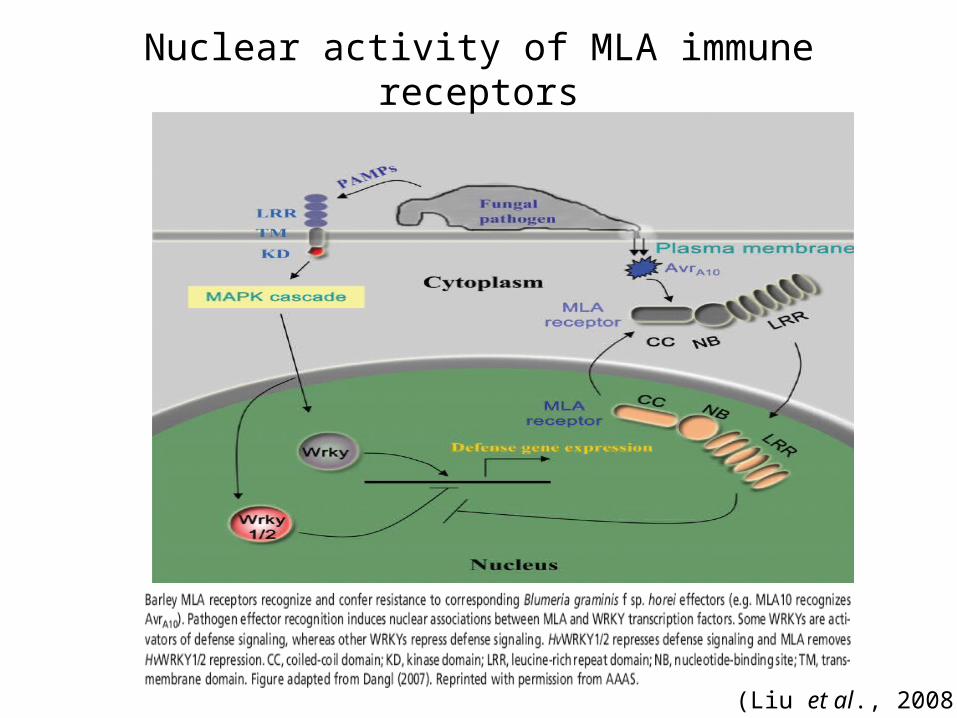
Nuclear activity of MLA immune receptors
(Liu et al., 2008)


Objective of the study
Study of the interaction of tomato plants with tomato powdery mildew fungus, Oidium neolycopersici
The monogenic genes Ol-1, ol-2 and Ol-4 confer resistance to tomato powdery mildew Oidium neolycopersici via different mechanisms
Study of the molecular and biochemical mechanisms involved
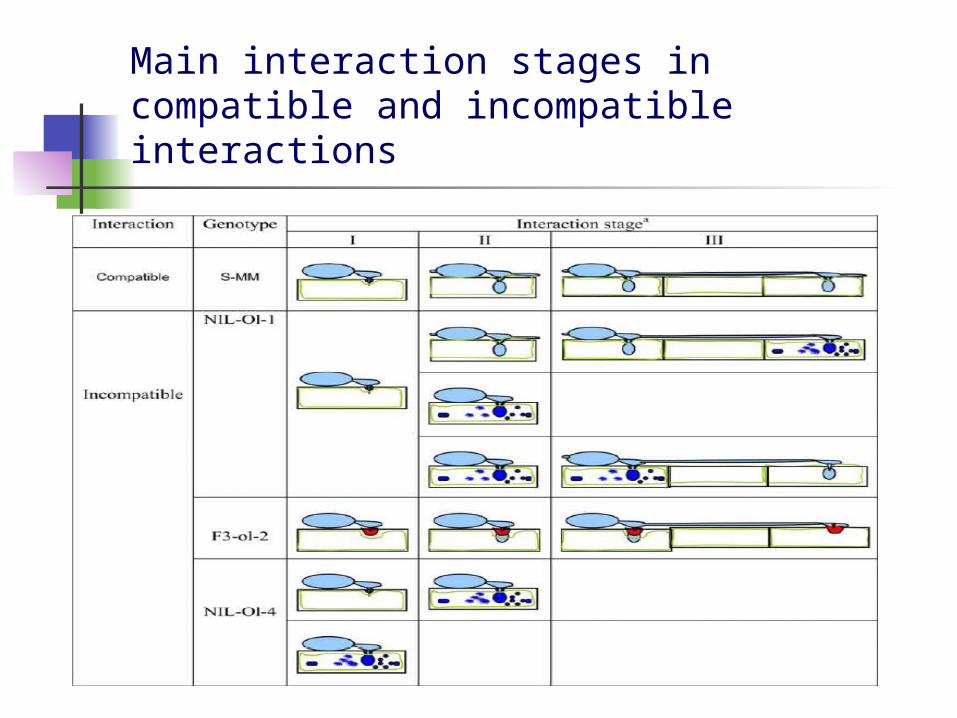
Main interaction stages in compatible and incompatible interactions
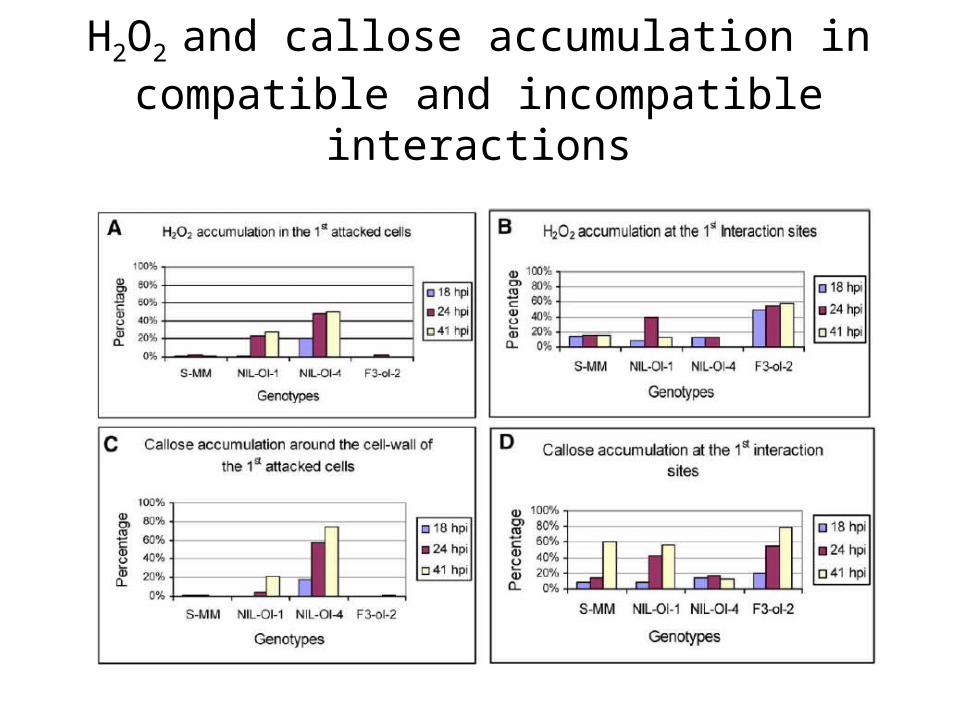
H2O2 and callose accumulation in compatible and incompatible interactions

Different expression classes of the DE-TDF in cDNA AFLP analysis
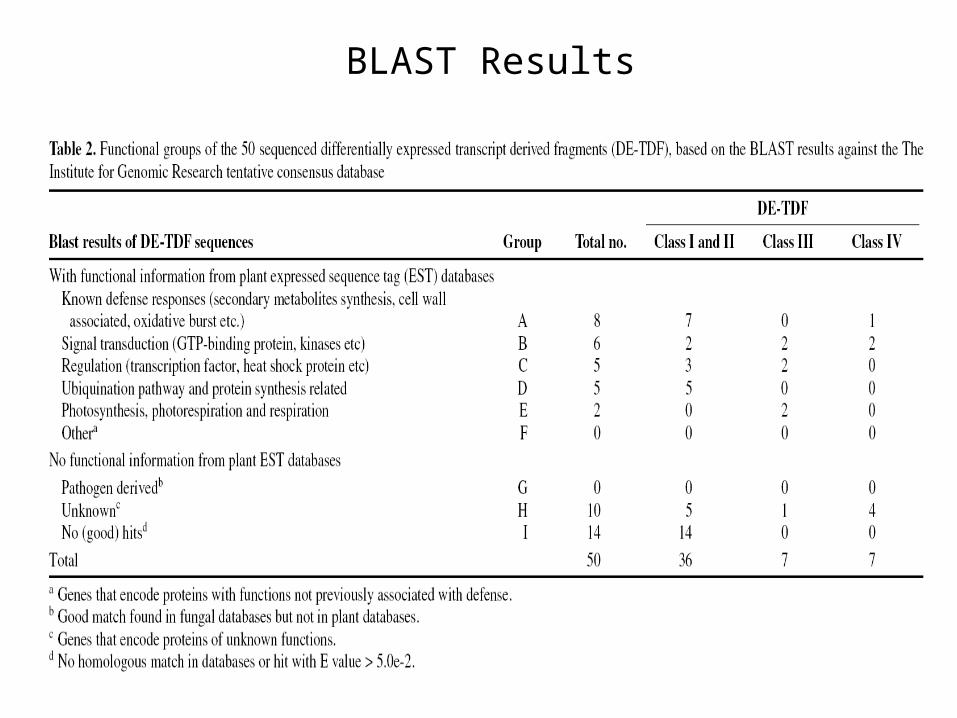
BLAST Results
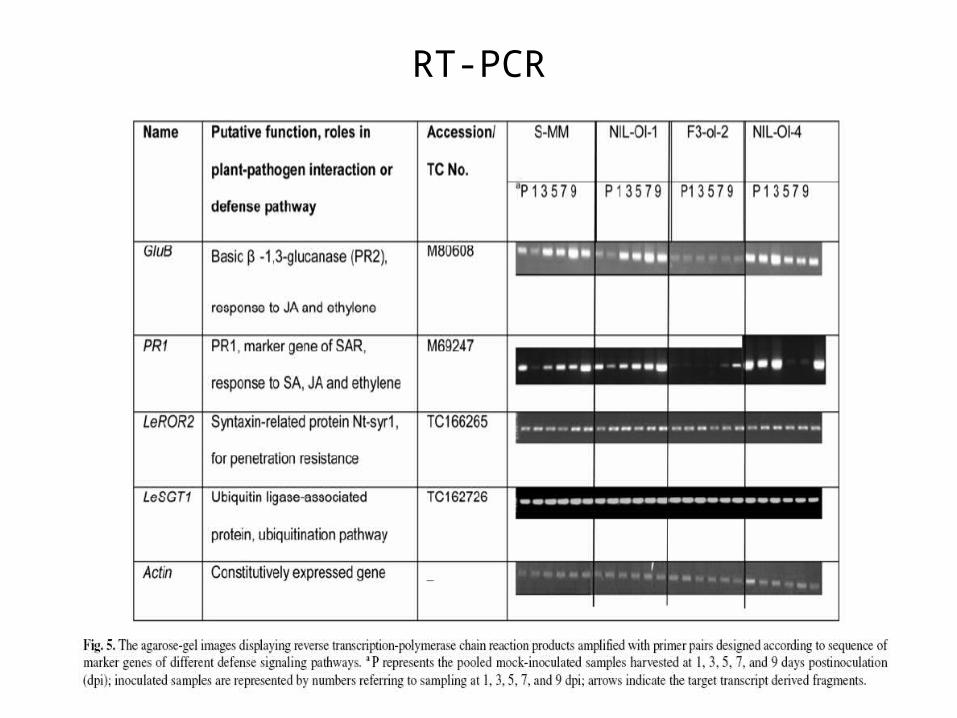
RT-PCR

Conclusion of the study
ROS, callose accumulation, and upregulation of DE-TDF associated with resistances conferred by dominant and recessive Ol genes
cDNA – AFLP profiling clarified that 81% of upregulated DE-TDF are common for both compatible and incompatible interactions
Class III DE-TDF were up regulated only in incompatible interactions and are specific for specific resistance genes
DE-TDF profiles of NIL-OL-1 and NIL-OL-4 deviated much but similarities were observed between NIL-OL-1 and F3-ol-2

Conclusion
An evolutionary arms race..

Discussion

THANK U…





























