
ANNALS O F CLINICAL AND LABORATORY SCIENCE, Vol. 8, No. 4Copyright © 1978, Institute for Clinical Science
Renal Handling of Proteins and Peptides
F. A. CARONE, M.D.
Morrison Professor o f Pathology, Northwestern University School o f Medicine,
Chicago, IL 60611
ABSTRACT
The kidney plays an im portant role in the metabolism of proteins and peptides. C urrent evidence indicates that only the proximal tubule possesses the mechanisms for absorption, transport and/or degradation of these substances. Large proteins and polypeptide m olecules filtered by the glomerulus, are absorbed from proximal tubular fluid by luminal en- docytosis into apical vacuoles which fuse with primary lysosomes where hydrolysis occurs followed by diffusion of metabolites out of the cells and into the blood. Recent evidence indicates that small peptides are handled by a different mechanism. It appears that small peptides are degraded at the luminal surface of the brush-border of proximal tubules, which contains many hydrolytic enzymes, by the process of membrane or contact digestion with reabsorption of the breakdown products. Proximal tubular m echanisms for handling of proteins and peptides are probably important biologically to conserve amino acids, inactive toxic substances and help regulate the circulating level of protein and peptide hormones.
It is clearly established that the kidney plays an important role in the metabolism of a num ber of protein, polypeptide and sm all p e p tid e m o lecu les , in c lu d in g p lasm a p ro te in s , g row th horm one, L-chains of immunoglobulins, /^-micro- globulin, lysozyme, insulin and small vasoactive peptides. Absorption, transport and/or degradation of proteins or peptides are properties of the proximal tubules. T here is little ev idence that other segments of the nephron have the mechanisms for uptake or transport of these substances. In d irec t and d irect studies indicate that a variety of proteins
and po lypep tides filte red by the g lomerulus are absorbed by the proximal tubule by luminal endocytosis and hydrolyzed by lysosomal enzymes.
Recent studies suggest that small linear peptides, consisting of eight to ten amino acids, are handled by the proximal tubule by a different mechanism. It has been postu lated that small pep tides m icroinfused into proximal tubules are hydrolyzed at the luminal surface of the brush border, which is rich in a variety of enzymes, by the process of membrane or contact digestion with reabsorption of most of the breakdown products.
2870091-7370/78/0700-0287 $01.20 © Institute for Clinical Science, Inc.
2 8 8 CARONE
Proximal Tubular H andling of Proteins and Large Peptides
Qualitative morphologic and indirect functional stud ies have dem onstrated that proximal tubular cells absorb proteins in the. tubular lum inal fluid and have suggested a pathway by which protein undergoes intracellular digestion. These processes have been quantitated directly. Some findings suggest that certain proteins are transported intact across the proximal epithelial cells; however, other studies do not support this conclusion. Direct studies on isolated tubular segments indicate that album in and insulin are not transported into tubular cells across the peritubular cell m em brane, presumably owing to the absence of an endocytic mechanism on this side of the tubular epithelium.
L u m in a l U p t a k e
M any m orphologic, m icropuncture , m icro in fu sion and m icroperfu sion studies have dem onstrated that a wide variety of proteins are absorbed along the lum inal a sp ec t o f proxim al tu b u la r
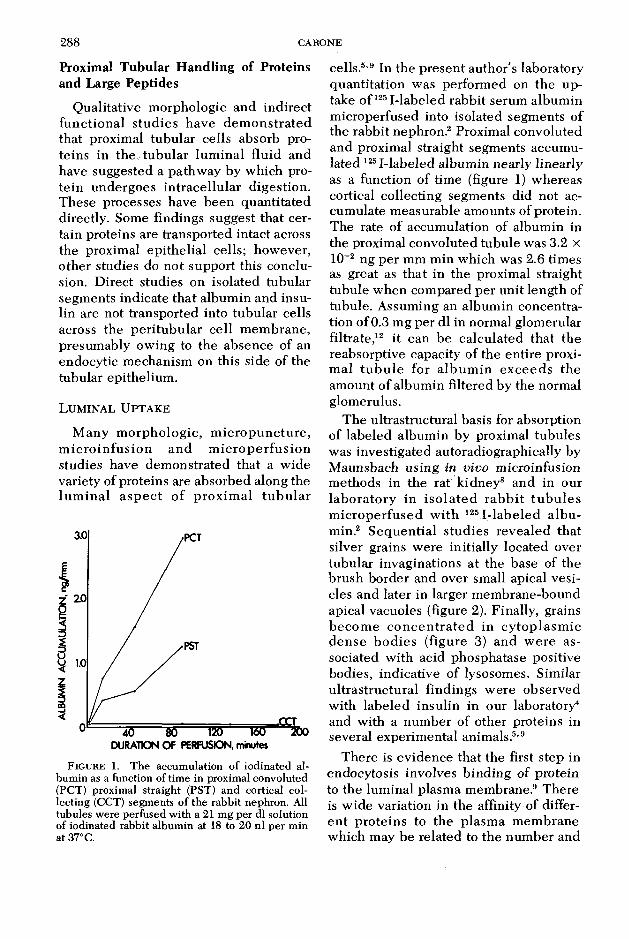
F ig u r e 1. The accumulation of iodinated albumin as a function of time in proximal convoluted (PCT) proximal straight (PST) and cortical collecting (CCT) segments of the rabbit nephron. All tubules were perfused with a 21 mg per dl solution of iodinated rabbit albumin at 18 to 20 nl per min at 37°C.
cells.5,9 In the present author’s laboratory quantitation was performed on the uptake o f1251-labeled rabbit serum album in m icroperfused into isolated segments of the rabbit nephron.2 Proximal convoluted and proximal straight segments accumulated 125I-labeled albumin nearly linearly as a function of time (figure 1) whereas cortical collecting segments did not accumulate measurable amounts of protein. The rate of accumulation of albumin in the proximal convoluted tubule was 3.2 x 10~2 ng per mm min which was 2.6 times as great as that in the proximal straight tubule when compared per unit length of tubule. Assuming an album in concentration of 0.3 mg per dl in normal glomerular filtrate,12 it can be calculated that the reabsorptive capacity of the entire proxim al tu b u le for a lbum in exceeds the amount of albumin filtered by the normal glomerulus.
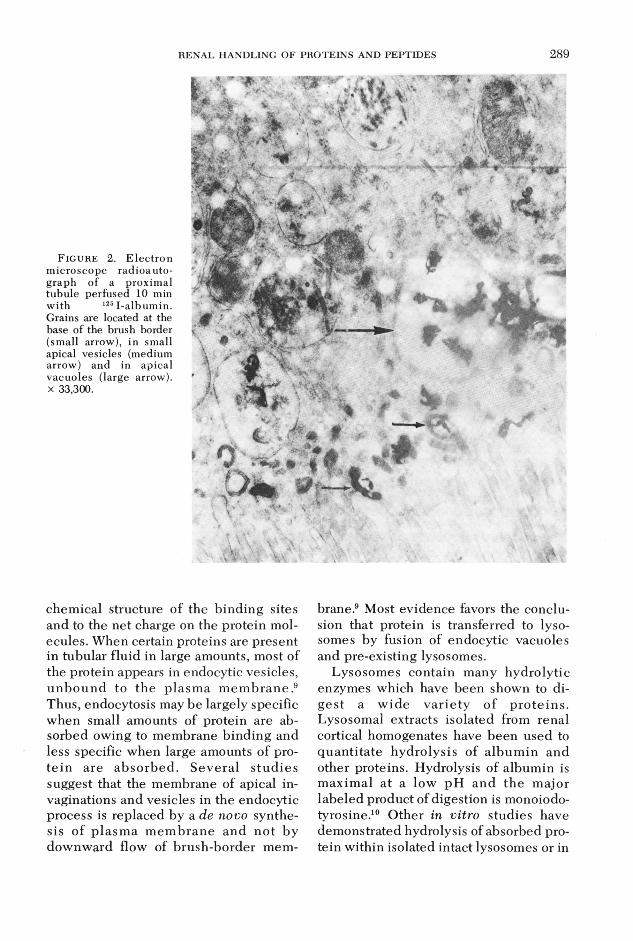
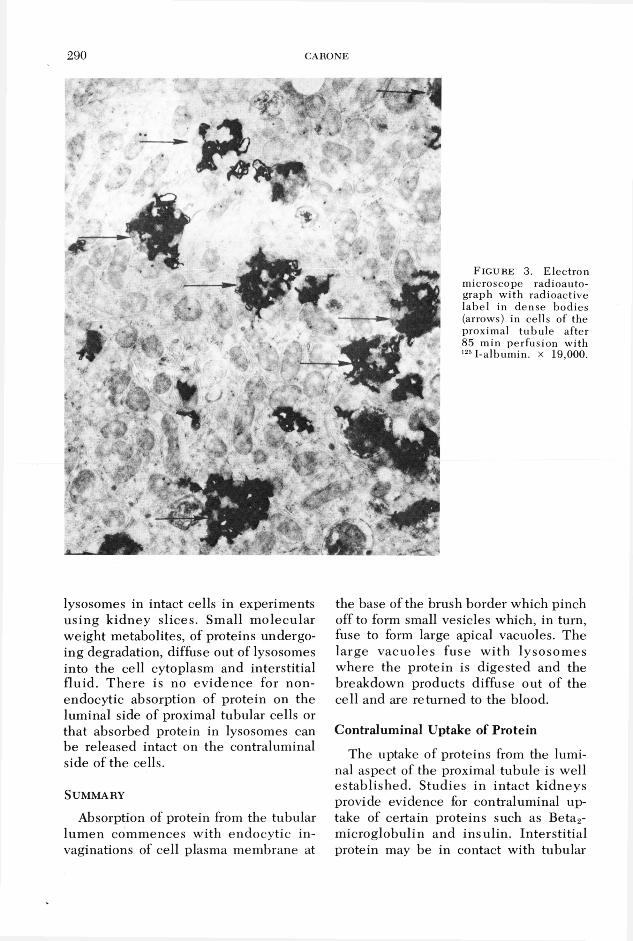
The ultrastructural basis for absorption of labeled album in by proximal tubules was investigated autoradiographically by M aunsbach using in vivo microinfusion methods in the rat kidney8 and in our laborato ry in iso la te d rab b it tu b u le s m icroperfused w ith l25I-labeled a lb u min.2 Sequential studies revealed that silver grains were initially located over tubular invaginations at the base of the brush border and over small apical vesicles and later in larger m em brane-bound apical vacuoles (figure 2). Finally, grains becom e c o n cen tra ted in cy top lasm ic dense bod ies (figure 3) and w ere associated with acid phosphatase positive bodies, indicative of lysosomes. Similar u ltrastructural findings w ere observed with labeled insulin in our laboratory4 and with a num ber of other proteins in several experimental animals.5,9
There is evidence that the first step in endocytosis involves b inding of protein to the luminal plasma membrane.9 There is w ide variation in the affinity of differen t proteins to the plasm a m em brane which may be related to the num ber and
RENAL HA N D LIN G O F PRO TEIN S AND PEPT ID E S 289
I ’’’- -xS % • 4
' ^ is
**• é , M•A
F ig u r e 2. E lectron m icroscope radioautograph of a proximal tubule perfused 10 min w ith 1251-albumin.Grains are located at the base of the brush border (small arrow), in small apical vesicles (medium arrow) and in apical vacuoles (large arrow), x 33,300.
«a*»**'. Afnr. ■
. %• ' ’ 4 ? ;
» '• £ mu
jf . w «
I r .%
«
% ; f
*
chemical structure of the b ind ing sites and to the net charge on the protein molecules. W hen certain proteins are present in tubular fluid in large amounts, most of the protein appears in endocytic vesicles, u n b o u n d to th e p la sm a m e m b r a n e .9 Thus, endocytosis may be largely specific w hen small amounts of protein are absorbed owing to m em brane b ind ing and less specific w hen large amounts of prote in are a b s o rb e d . S ev era l s tu d ie s suggest that the m em brane of apical invaginations and vesicles in the endocytic process is replaced by a de novo synthesis o f p la s m a m e m b ra n e an d n o t by dow nw ard flow of b rush-border m em
brane.9 Most ev idence favors the conclusion that protein is transferred to lyso- somes by fusion of endocytic vacuoles and pre-existing lysosomes.
Lysosom es contain many hydro ly tic enzymes which have been shown to dig e s t a w id e v a r ie ty o f p ro te in s . Lysosomal extracts isolated from renal cortical homogenates have b een used to q u an t i ta te hydro lys is of a lb u m in and other proteins. Hydrolysis of a lbum in is m axim al at a low p H an d th e m ajor labeled product of digestion is monoiodo- tyrosine.10 O ther in vitro s tud ies have dem onstrated hydrolysis of absorbed protein within isolated intact lysosomes or in
290 CARONE
F ig u re 3. Electron microscope radioautograph w ith radioactive label in dense bodies (arrows) in cells of the proximal tubu le after 85 min perfusion with 1251-albumin. x 19,000.
lysosomes in intact cells in experiments using k idney slices. Small m olecular weight metabolites, of proteins undergoing degradation, diffuse out of lysosomes into the cell cytoplasm and interstitial fluid. T here is no ev idence for non- endocytic absorption of protein on the luminal side of proximal tubular cells or that absorbed protein in lysosomes can be released intact on the contraluminal side of the cells.
S u m m a r y
Absorption of protein from the tubular lum en com m ences with endocytic invaginations of cell plasma membrane at
the base of the brush border which pinch off to form small vesicles which, in turn, fuse to form large apical vacuoles. The large vacuoles fuse w ith lysosom es where the protein is digested and the breakdown products diffuse out of the cell and are returned to the blood.
Contraluminal Uptake of Protein
The uptake of proteins from the luminal aspect of the proximal tubule is well established. Studies in intact k idneys provide evidence for contraluminal uptake of certain proteins such as Beta2- m icroglobulin and insulin . Interstitial protein may be in contact with tubular
RENAL HA N D LIN G O F PR O TEIN S AND PEPT ID E S 2 9 1
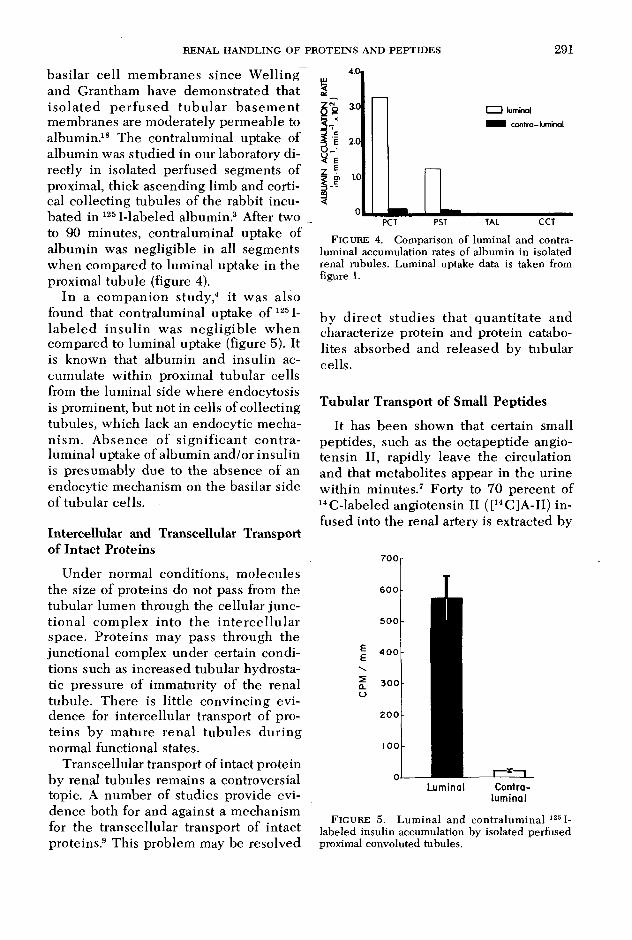
basilar cell m em branes since W elling and Grantham have dem onstrated that iso la ted p e rfu sed tu b u la r b asem en t membranes are moderately perm eable to albumin.18 The contraluminal uptake of album in was studied in our laboratory directly in isolated perfused segments of proximal, thick ascending limb and cortical collecting tubules of the rabbit incubated in 125I-labeled albumin.3 After two to 90 m inutes, contraluminal uptake of albumin was negligible in all segments when compared to luminal uptake in the proximal tubule (figure 4).
In a com panion study,4 it was also found that contralum inal uptake of 1251- labe led in su lin was neg lig ib le w hen compared to luminal uptake (figure 5). It is known that album in and insulin accumulate w ithin proximal tubular cells from the luminal side where endocytosis is prominent, but not in cells of collecting tubules, which lack an endocytic mechanism . A bsence of s ign ifican t co n tra luminal uptake of album in and/or insulin is presumably due to the absence of an endocytic mechanism on the basilar side of tubular cells.
Intercellular and Transcellular Transport o f Intact Proteins
Under normal conditions, m olecules the size of proteins do not pass from the tubular lumen through the cellular junctional com plex into th e in te rce llu la r space. Proteins may pass through the junctional complex under certain conditions such as increased tubular hydrostatic pressure of immaturity of the renal tubule. There is little convincing evidence for intercellular transport of proteins by m ature renal tubu les during normal functional states.
Transcellular transport of intact protein by renal tubules remains a controversial topic. A num ber of studies provide evidence both for and against a mechanism for the transcellular transport of intact proteins.9 This problem may be resolved
F igure 4. Comparison of luminal and contraluminal accumulation rates of albumin in isolated renal rubules. Luminal uptake data is taken from figure 1.
by d irec t stud ies that q u an tita te and characterize protein and protein catabo- lites absorbed and released by tubular cells.
Tubular Transport of Small Peptides
It has been shown that certain small peptides, such as the octapeptide angiotensin II, rapidly leave the circulation and that metabolites appear in the urine w ithin m inutes.7 Forty to 70 percent of 14C-labeled angiotensin II ([14C]A-II) infused into the renal artery is extracted by
Luminal Contraluminal
F igure 5. Luminal and contralum inal 1251- labeled insulin accumulation by isolated perfused proximal convoluted tubules.

292 CARONE
the dog kidney, indicating that renal extraction of angiotensin II involves more than glom erular filtration and urinary excretion.1 Furthermore, similar studies show th a t 75 pe rcen t of [14C] A-II is m etabolized in a single passage through the kidney and that 98.7 percent of injected radioactive material is recovered in renal venous blood, leading to the conclu sion tha t tissu e se q u es tra tio n o f labeled material does not occur.13 Support for this is the autoradiographic evidence that tritium -labeled angiotensin-II localizes in mesangial cells of the rat kidney but not in tubular cells.11
Recently, the role of individual nephrons or isolated tubu lar segm ents has been assessed by us in the reabsorption and/or metabolism of radioactive-labeled angiotensin II, bradykinin or oxytocin. The techniques of in vivo, tracer microinjection of surface tubules in rats and in vitro microperfusion of isolated segments of rabbit kidneys were employed. Recovery of radioactivity in urine or collection fluid was used to calculate the extent of tubular reabsorption of labeled peptide and two-dimensional peptide m apping was employed to identify and quantitate intact peptide and its metabolites in urine or collection fluid. To identify possible pep tide degradation products, labeled pep tid es w ere d ig ested w ith several pu rified enzym es, the p roducts w ere spotted on Whatman 3MM paper, subjec ted to descending chrom atography, d rie d and th e n su b je c ted to e le c trophoresis in a second dim ension.6,15 Approximately 95 percent of radioactivity applied to the map appeared in identifiable spots.
When [14C] A-II was infused in vivo, into individual surface proximal or distal tubules in rats,15 recovery of 14 C in the u rine was 11 pe rcen t w hen proxim al tubu les w ere infused and 95 percen t when distal tubules were infused. Follow ing proxim al tu b u la r in fusions of [14C] A-II, of the 11 percent of 14C recov
ered in the urine, most (95 percent) appeared as peptide fragments, the principal metabolic product was the Ile5-Phe8 fragment of angio tensin II. Follow ing distal tubular infusion of [14C] A-II, essentially a l l 14 C activity in the urine was intact angiotensin II. Similar data were obtained when the renal tubular handling o f3H-labeled bradykinin ([3H] BKN) was studied by the same techniques.6 After microinfusion of [3H] BKN, urinary recovery of 3H labeled was 24 percent when proximal tubules were infused and 98 percent when distal tubules were in fused. For proximal tubules, 85 percent of 3H activity in urine consisted of metabolites (81 percent Pro and 4 percent A rg^P he5) and 15 percen t was intact BKN. With distal tubules, all 3H activity appeared as intact BKN.
In another set of experiments [14C] A-II was microperfused, in vitro, in isolated segments o f rabbit tubules.14 Proximal straight tubules reabsorbed about 30 percent per mm tubular length of perfused 14 C label and most of the isotope was transported across the tubular cells, since no label was sequestered by the tubules during the perfusion period . E lectro phoresis of collected perfusate demonstrated that [14C] A-II was hydrolyzed to [14C] isoleucine.
C ollectively , these stud ies dem onstrate that the proximal tubules of rat and rabb it k idneys can enzym atically hydrolyze small peptides presented to the luminal aspect of the tubular cells. Furthermore, distal segments of the nephron do not appear to possess this mechanism. The concentrations of angiotensin II and bradykinin in the infusates of these experim ents were several thousand times greater than that of normal plasma indica ting an enorm ous capacity of the mammalian proximal tubule to hydrolyze and reabsorb small peptide molecules.
Proximal tubu lar hand ling o f small peptides may be accounted for by (1) hydrolysis at the luminal cell surface with
RENAL H A N D LIN G O F PR O TEIN S AND PE PT ID E S 2 9 3
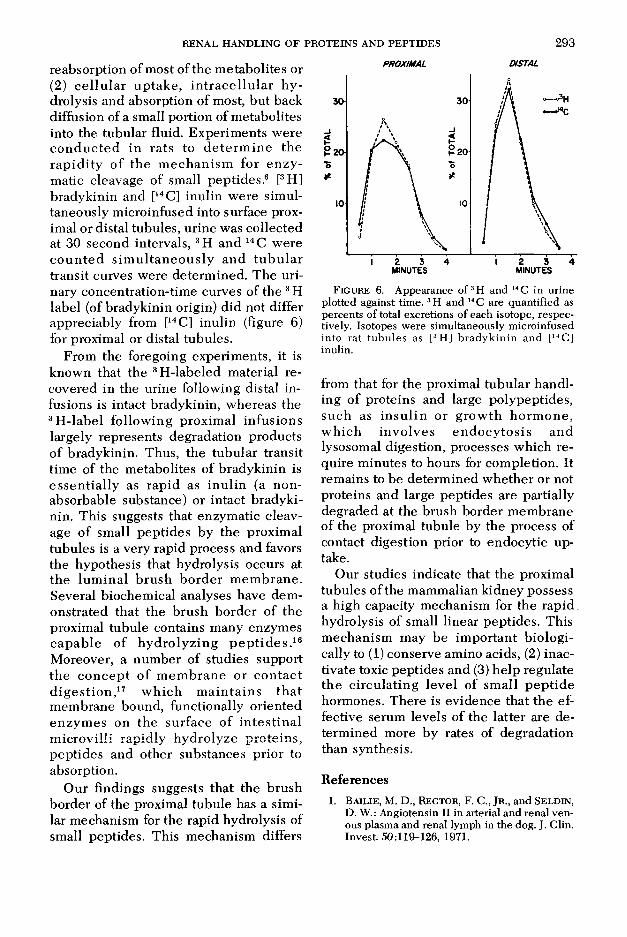
reabsorption of most of the m etabolites or (2) c e llu la r up take , in tra ce llu la r h y drolysis and absorption of most, but back diffusion of a small portion of metabolites into the tubular fluid. Experim ents were co n d u c ted in rats to d e te rm in e the rap id ity o f the m echan ism for en zy matic cleavage of small peptides.6 [3H] bradykinin and [14C] inulin were simultaneously microinfused into surface proximal or distal tubules, urine was collected at 30 second intervals, 3H and 14 C were c o u n te d s im u ltan eo u sly and tu b u la r transit curves were determ ined. The urinary concentration-time curves of the 3 H label (of bradykinin origin) did not differ appreciably from [14C] inulin (figure 6) for proximal or distal tubules.
From the foregoing experiments, it is known that the 3H-labeled material recovered in the urine following distal infusions is intact bradykinin, whereas the 3H -label follow ing proxim al infusions largely represents degradation products of bradykinin. Thus, the tubular transit time of the metabolites of bradykinin is essen tia lly as rapid as inu lin (a nonabsorbable substance) or intact bradykinin. This suggests that enzymatic cleavage of small peptides by the proximal tubules is a very rapid process and favors the hypothesis that hydrolysis occurs at the lum inal b rush border m em brane. Several biochemical analyses have dem onstrated that the brush border of the proximal tubule contains many enzymes capab le of hydro lyz ing p e p tid e s .16 Moreover, a num ber of studies support the co n cep t o f m em brane or con tac t d ig es tio n ,17 w h ich m ain ta in s th a t m embrane bound, functionally oriented enzym es on the surface of in tes tina l m icrovilli rapidly hydrolyze pro teins, peptides and other substances prior to absorption.
Our findings suggests that the brush border of the proximal tubule has a similar mechanism for the rapid hydrolysis of small peptides. This mechanism differs
PROXIMAL DISTAL
Figure 6. Appearance o f3H and 14 C in urine plotted against time. 3H and 14 C are quantified as percents of total excretions of each isotope, respectively. Isotopes were simultaneously microinfused into rat tubules as [3H] bradykinin and [14C] inulin.
from that for the proximal tubular handling of proteins and large polypeptides, such as in su lin or grow th horm one, w h ich invo lves endocy to sis and lysosomal digestion, processes which require m inutes to hours for completion. It remains to be determ ined w hether or not proteins and large peptides are partially degraded at the brush border membrane of the proximal tubule by the process of contact digestion prior to endocytic uptake.
Our studies indicate that the proximal tubules of the mammalian kidney possess a high capacity mechanism for the ra p id . hydrolysis of small linear peptides. This m echanism may be im portant biologically to (1) conserve amino acids, (2) inactivate toxic peptides and (3) help regulate the c ircu lating level of small pep tide hormones. There is evidence that the effective serum levels of the latter are determ ined more by rates of degradation than synthesis.
References1. Ba ilie , M. D., Rector , F. C., Jr ., and Seld in ,
D. W.: Angiotensin II in arterial and renal venous plasma and renal lymph in the dog. J. Clin. Invest. 50:119-126, 1971.

294 CARONE
2. Bourdeau, J. E., Carone, F. A., and Ganote , C. E.: Serum albumin uptake in isolated perfused renal tubules. J. Cell. Biol. 54:382-398,1972.
3. Bourdeau , J. E. and Carone , F. A.: Contra- luminal serum in albumin uptake in isolated perfused renal tubules. Amer. J. Physiol. 224:399-404, 1973.
4. Bourdeau , J. E., Chen , E. R. Y., and Carone , F. A.: Insulin uptake in the renal proximal tubule. Amer. J. Physiol. 225:1399-1404, 1973.
5. Bourdeau, J. E. and Carone, F. A.: Protein handling by the renal tubule. Nephron 13:22-34, 1974.
6. Carone , F. A., Pullman , T. N., Oparil , S., and NAKAMURA, S.: Micropuncture evidence of rapid hydrolysis of bradykinin by rat proximal tubule. Amer. J. Physiol. 230:1420-1424,1976.
7. H odge , R. L., N g, K. K. F., and Vane, J. R.: Disappearance of angiotensin from the circulation of the dog. Nature 215:138-141, 1967.
8. M AUN SBA C H , A. B.: Absorption of I 125- labeled homologous albumin by rat kidney proximal tubu le cells. A study of microperfused single proximal tubules by electron microscopic autoradiography and histochemistry. J. Ultrastruct. Res. 15:197-241,1966.
9. MAUNSBACH, A. B.: Cellular mechanisms of tubular protein transport. Kidney and Urinary Tract Physiology, II, Vol. 11, Thurau, K., ed. Baltimore, University Park Press, 1976, pp. 145-167.
10. MAUNSBACH, A. B.: Ultrastructure and digestive activity of lysosomes from proximal tubu
lar cells. Proceedings of the 4th Congress on Nephrology, Vol. 1. Stockholm, Basel, Karger,1971, p. 102.
11. Meyer , P., O sborne , M. J., Broz , B., and M orel , F.: Intrarenal localization of angiotensin II by autoradiograph. Abstract of 5th International Congress on Nephrology. Mexico City, 1972, p. 117.
12. O aken, D. E., Cotes, S. C., and Men de , C. W.: Micropuncture study of tubular transport of album in in rats w ith am inonucleoside nephrosis. Kidney Intern. 2:3-11, 1972.
13. O paril , S. and Bailie , M. D.: Mechanism of renal handling of angiotensin II in the dog. Circulation Res. 33:500-507, 1973.
14. Peterson, D. R., O paril , S., Flouret , G., and CARONE, F. A.: Handling of angiotensin II and oxytocin by renal tubular segments perfused in vitro. Amer. J. Physiol. 232:F319- F324, 1977.
15. Pullman , T. N., Oparil , S., and Carone , F.A.: Fate of labeled angiotensin II microinfused into individual nephrons in the rat. Amer. J. Physiol. 228:747-751, 1975.
16. Q uirk , S. J. and Robinson , G. B.: Isolation and characterization of rabbit kidney brush borders. Biochem. J. 128:1319-1328, 1972.
17. Ugolev , A. M. and de Laey, P.: Membrane digestion. A concept of enzymic hydrolysis on cell m em branes. Biochim. Biophys. Acta 300:105-128, 1973.
18. Well in g , L. W. and G rantham , J. J.: Physical properties of isolated perfused renal tubules and tubular basement membranes. J. Clin. Invest. 51:1063-1075, 1972.
Recommended

















