
Indian Journal of Ex perimental Biology Vol. 4 1, August 2003 , pp. 850-856
Regional variations in fibre growth dynamics of myotomal and caudal fin muscles in relation to body size of a freshwater teleost, Barbus sarana (Cuv. & Val.)
Swati Pandya, Kamakshi Arora, Sudipta Mi sra & Rah ul Kundu *
Department of Biosc iences , Saurashtra Uni versity, Rajkot 360005. Indi a.
Received I Allgllsl 2002: revised 9 April 2003
The growth of red fibres in anterior and middle myotomal regions of B. samlla was mainly by hyperplasia in small er size classes. In higher size classes, growth by hyperplasia was greater in posteri or myotomal reg ion compared to the other two myolOmal regions. The growth of pin k fibres in anteri or myotomal regions was mainl y by hypertroph y. The middl e and posteri or myotomal regions showed fibre growth by hyperpl as ia. The growth dynamics of white fibres revealed morc or less similar pattern in all three myotomal regions aga inst the somatic development. White fibres grew by hyperplasia up to 8 cm F.L. size classes and thereafter by hypertrophy. However, in > 12 cm F.L. size classes, the mean diameter of white fib res did not increase significantl y. Similar pattern of growth was fou nd in the white fibres of caudal fin muscle. It is interesting 10
note that the hyperplasia was mostl y compl eted in the white fibres of the small es t fi sh stud ies, whereas. it continued to quite larger fish size in red and pi nk fibrc s. Thus. hyperp lasia and hypert rophy may be responsibl e fo r growt h in all fibre types in all myotomal regions in relat ion to somati c development in th is small and medium grow ing spec ies.
Keywords : Barbll .\" samlla , Fibre growth dynamics, Fish-Myotomal regions, Muscle fibre growth
In the ax ial musculature of many spec ies of fi h, white fi bres are more numerous than red I. In muscle tissue postembryonic hyperpl as ia and hypertrophy are normall y derived from myosatellite cells2, which di vides to form new small musc le fibres that mature by hypertroph/ -6. However, the somatic growth in fish can be very much plastic, affec ted by temperature, food, hormone etc.~ . The fast growing spec ies grow possibly by hyperplas ia rather than hypertrophy and by both processes in slow and medi um growing spec ies7
. The myotomal muscle of fi sh compri ses much of the larges t fraction of the ti ssues and its relative growth rate is close to that of other ti ssues or of the fi sh body as whole8
. Thus, it is possible that the muscle growth dynamics are related to the somatic growth rate and to the final body size7
.9
-t2.
The larva l stages of many teleost have a single superfici al layer of small diameter muscle fibres that express fast muscle myosin light chains t3 and have a myosin heavy-chain composition di stinct from that of adult red and white muscle fibres l4
. Postembryonic muscle growth of fish myotomes involves both new fibre recruitment (hyperplasia) and hypertrophy of ex isting fibres5. Fibre recruitment also varies with seasont 5, between males and females t6 . The growth of
*For correspondence : Phone and Fax : 09 1-0281-25864 19 E-mail: rskundu_ad J @sancharnet.in
the muscle fibres by hyperpl as ia declines stead il y from juvenile to adult stages and totally stops at around 40-50% of the maximum body length , as the recruitment of new small fibre ceases, and the fi sh reach to a comparatively large size by hypertrophi ·7 .
The present communication reports the growth dynami cs of red, pink and white fibres of a trop ica l freshwater small and medium growing teleost Barblls sarona (Cuv. & VaL) . This species is a small sized edible fi sh, reported to attain a maximum length of about a foot l7 and has a medium somatic growth rate. However, authors never came across fi sh more than 17 cm fork length during entire study period in the Saurashtra region of the Gujarat State.
Materials and Methods Live specimens of B. sarana were co llected
locally . A total of 35 fishes fallin g under seven size classes, viz. 5 cm (n = 3), 8 cm (n = 5). 12 cm (n = 6), 14 cm (n=6), IS cm (n=8), 16 cm (n:= 3) and 17 cm (n = 4) fork length were examined. Since the fi shes were collected from the local fish vendors, fish size classes were selected according to the availability of various sizes in the catch. Looking into the function al role in the body, muscle samples were taken from the three lateral myotomal regions of the fish . Samples of myotomal muscles were obtained from (a) anterior myotomal region , next to the operculum; (b) middle myotomal region, on the lateral line above the anal

PANDYA 1'1 ClI .: M USCLE FIBRE GROWTH DYNAMICS OF FISH /3A /?B US 851
aperture; and (c) posterior myotomal reg ion, prior to the caudal region of the f ish body. Apart from these three myotomal regions, caudal fi n muscle, immediately before commencement of the caudal fin rays was also taken. Di ssected muscle samples were immediately frozen w ith dry ice and transferred onto the stage of a cryosta t microtome at - 20°e. Transverse secti ons of about 14 11m were cut and histochemica l demonstrati on of lipids, SD H and LDH~·' x. were carried out in order to iden tify vari ous musc le fibre types . Fibre diameter was measured by using ocular micrometer fitted in an eyepiece')·' '!. Since large pink and white fibres were nor always circul ar in shape, diameter of each pink and white fibres was measured at least thri ce from three different angles and the mean va lue is taken as standard diameter of that fi bre. Diameter of 100 ri bres of a single type from each myotomal reg ion was measured. T he data were subjected to stati sti ca l analyses l ike regress ion and correlati on coefficient as per Soka l & Rohlf20.
Results
Red fibres showed high f ibre frequency va lues in intermediate diameter modes. In all three myotoma l regions. lower diameter mode ( 1- 10 11m ). indica ting hyperplas ia, was present in sma ller size classes. T he frequency value of thi s mode declined in the higher size classes, replaced by the sh i fti ng of model frequency values to the nex t higher d il.llneter mode (hypertrophy ). In anteri or myotomal reg ion , the growth of red fibres occurred mainl y by hyperpl as ia in smaller sized classes (Fig. I ). However, in higher size classes, growth was by hyperplasia and hypertroph y. In the midd le myotomal reg ion the main mode of growth was by hyperplasia up to 8 cm (F.L.) f ish. thereafter the musc le growth occurred mainl y by hypert rophy. In the posteri or myotomal region the amou nt of red fibres was greater than other two myotomal regions (Fig. I ). On the other hand, the red fibres of the caudal fin muscle showed substanti al recruitment of small new fibres in all size classes studied, however, the hyperpl as ia was less in higher size classes (Fig. 4). In the anteri or and middle myotoma l regions, pink fibres showed even lower recruitment ra te than posteri or myotomal reg ion where very lillie add ition of small new pink fibres was observed. In higher size classes, in vo lvement of hypertrophy was observed as recruitment o f small new fibres was ceased in > 14 cm F.L. size class
(Fig. 2) . However, hypertroph y was ev ident in caudal fi n muscl e as we l l (Fig. 4). I n the present invest igation , white fibres showed very little recruitment o f small new f ibres in few lower sizc classes. White fibres showed high frequency values in the higher diameter modes hav ing diameter of about 2 1-60 11m . In anteri or myotomal region, the frequency va lues were high in moderate diameter modes (21-60 11m). However, the shirting or model freq uency values towards higher diameter modes was observed in middle and posterior body reg ions (Fig. 3). Very large f ibres hav ing diameter of about 90 ,um was observed in the anteri or and middle myoto l11al reg ions. The white fibres of th e caudal fi n muscle showed abso lutely no hyperpl as ia (Fig. 4) .
Discussion In the present study, growth dYl1<.lIl li cs of reel fi bres
in vo lved a certain degree of hyperp lasia in myotomal musc le. The results suggest more or less no regional variat ions among the three myotomal region:--. However, growth by hyperplasia was Illuch greater in posteri or myotomal region than ot her two myotomal
80 ~ 5cm 70 • Scm 60 o 12cm 50 o 14cm 40 ~ 15cm 30 ~ 16cm 20 ~ 17cm 10
0 80
~ 70
>- 60 u c: 50 (1) :;;J
40 r::r ~ 30 (1) 20 .0 Ii: 10
0 80
70
60
50
40
30
20
10
0 1-10 11 -20 21.30
Fibre diameter mode
Fig. I - Mean percell! I"requLncics of red fihn;s in Ihrcc lllyolOmal regions against fi bre diamelcr modes (/lIl1 ) in various fi sh size c lasses.

~52 INDIAN J EXP BIOL. AUGUST 2003
regions. Since the posterior myotomal region to a greater ex tent provides force for the movement, the amount of red fibres was much higher than the other two myotomal regions, showing hyperplasia as predominant mode of growth . An increase in diameter of red fibres occurred only in middle myotomal region indicating that the small new red fibres were recruited up to certain extent and then ceased, suggesting rest of the growth was by hypertrophy (Tab le I ). Thu s, a mixed type of growth dynamics was ev ident in red fibres in almost all myotomal regions. The experimenta l animal , B. sarona is not a cons tant sw immer but ex hibit occas ional fast swimming activities l 7
. Poss ibly due to thi s reason, the amount of red f ibres is less than white fibres in all myotomal regions. It is possib le that the hyperp lasia was restricted to the posterior myotomal reg ion that provides maximum force for slow and susta ined
80
70 Anterior myo tomal
60
50
40
30
20
10
0 80
70 Middle myotomal
;g !!..- 60 >- 60 u c: CI> 40 ::l C" CI> 30 ... -CI> 20 ... .0
10 u:: 8S
70 Poste rior myotomal
60
60
40
30
20
10
0
1·10 11·20 21-30
movement of caudal fin in thi s fish (Figs. j , 4 and Table 2). The caudal region of the fi sh along wi th th e posterior myoromal reg ion is the most active portion of the fi sh body during locomotion . Since the present experimental fish B. sarona is a moderately ac ti ve fish, the caudal portion is rich in red and pink fibres that are associated with slow and sLlstai ned movement of the fin .
The growth dynamics of the pink fibres showed certain similarity with that of red fibres. The max imum number of f ibres in both anteri or and midd le myotomal reg ions was in the range of 11-30 p.m. However, the recruitment of small new pink fibres was present in all myotomal regions (Fig. 2), which strongly resembles that of red fibres. The regression and correlation coeffIcient analyses showed hypertrophy in the pink f ibres of an terior myotomal reg ions (Tab les I and 2). rn the midd le
~ 5em.8em 0 12em
0 14 em 111 15 em
e:ll 16em 1§ll 17em
31-40 41-60 61~0
Fibre diameter mode
Fig. 2 - Mea n perce nt frequencies of pink fib res in three myotomal reg ions against fibre d iameter modes (lim) in various fi sh size c lasses.
..,
. .-t I
PANDY A el al.: MUSCLE FIBRE GROWTH DYN AMI CS OF FISH BARBUS 853
myotomal region and posteri or myotomal reg ions the rate of hyperpl as ia gradually increases which was supported by the results obtained from the regress ion as well as correlati on coefficient analyses. It was ev ident that the growth pattern of pink fibres in middle myotomal reg ions was possi ble by both hyperpl as ia and hypertrophy. A mi xed type of growth dynamics of pink fibres predominantl y by hyperpl as ia and partiall y by hypertrophy was observed in posteri or myotomal regions. The growth dy namics of
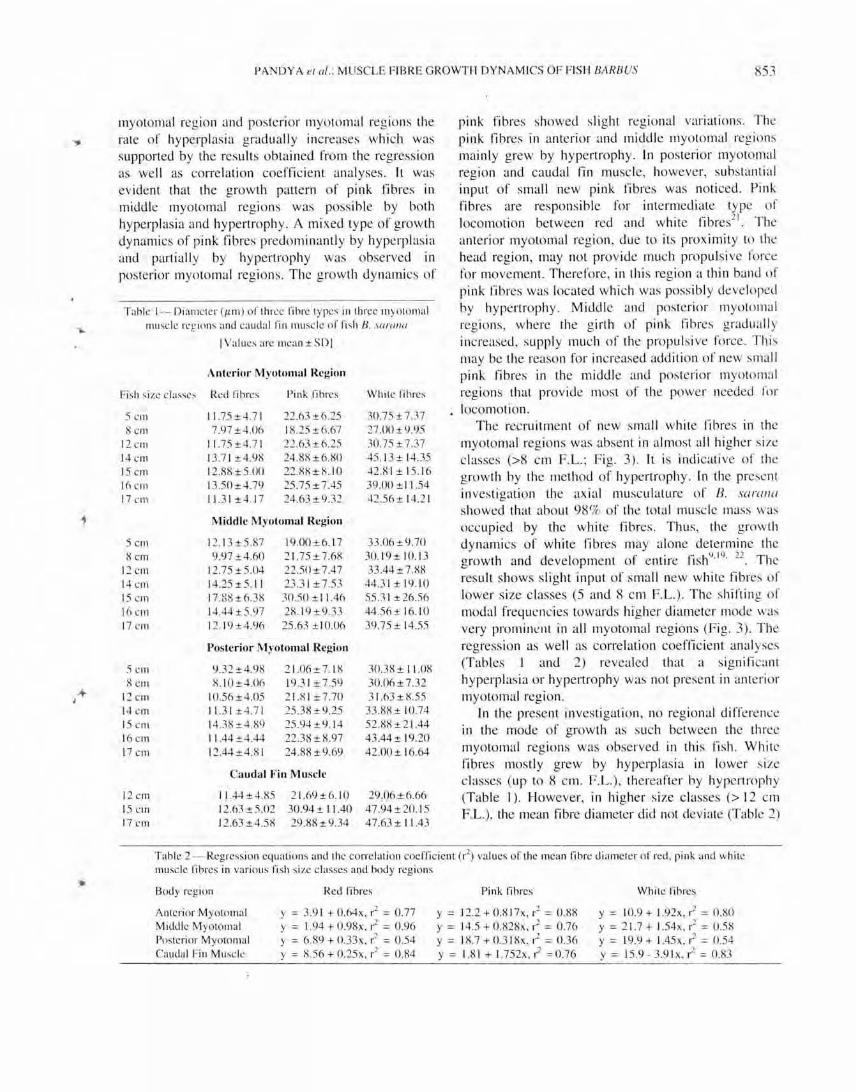
Table 1- Diameler (/1m ) o r lhree fibre lypes in lhree myolo mal ll1usc le regions and ca udal rin mu sc le or fi sh 8. sO/'{/Il{/
I Values are mean ± SDI
Ante rior Myotomal Region
Fi sh size classe, Red fibres Pink fibres While fibres
S cm 11.75 ±4.7 1 22.63±6.25 30.75 ±7.37 8elll 7.97±4.06 18.25±6.67 27.00 ±9.95
12 CIll 11.75±4.7 1 22 .63 ±6.25 30.75 ± 7.37 14 cm 13.7 1±4.98 24.88 ±6.80 45. 13 ± 14.35 15 Cill 12.88 ± 5.00 22.88 ± 8. 10 42.8 1 ± 15. 16 16 crn 13.50±4.79 25.75 ± 7.45 39.00 ± 11 .54 17 Cill 11.31 ±4.1 7 24 .63 ±9.32 42.56± 14.2 1
Middle Myolomal Region
SCIll 12.13±5.87 19.00 ± 6.1 7 33.06 ±9.70 8cIll 9.97 ±4.60 2 1.7S±7.68 30. 19± 10. 13
12 cm 12.75 ±5.04 22.50±7.47 33.44 ± 7.88 14 CI11 14 .2S±5. 11 23.3 1 ± 7.53 44.3 1 ± 19. 10 15 Cill 17 .88±6.38 30.50 ± I 1.46 55.3 1 ± 26.56 16 CIll 14.44±5.97 28. 19 ± 9.33 44.S6± 16.1 0 17 CIll 12. 19±4.96 25 .63 ± I 0.06 39.75 ± 14.55
Posterior M yotomal Region
5cIll 9.32±4.98 2 1.06± 7. 18 30.38 ± 11.08 8cIll 8. 10±4.06 19.3 1 ±7.S9 30.06± 7.32
12 Cill 10.56±4.05 2 1.8 1 ±7.70 3 1.63 ±8.55 14 Cill 11.3 1±4.7 1 25.38 ±9.25 33 .88 ± 10 .74 15 CIll 14.38±4.89 25.94±9. 14 52.88±2 1.44 16 Cill 11 .44 ±4.44 22 .38 ± 8.97 43.44± 19.20 17 Cill 12.44±4.8 1 24.88±9.69 42 .00 ± 16.64
Cauda l Fin Muscle
12 Cill 11 .44±4.85 2 1.69±6. 10 29 .06±6.66 15 cm 12.63±5.02 30.94 ± 11 .40 47 .94±20.I S 17 c rn 12.63 ±4.58 29.88 ±9.34 47.63 ± 11 .43
pink fibres showed sli ght regional variati ons. The pink fibres in anterior and middle myotomal reg ions mainl y grew by hypertrophy. In posterior myotomal region and caudal fi n muscle, however, substanti al input of small new pink fibres was noticed. Pink fibres are responsible for intermediate type of locomotion between red and white fibres21
. The anterior myotomal region, due to its proximity to the head region , may not prov ide much propulsive fo rce for movement. Therefore, in thi s region a thin band of pink fibres was located which was poss ibl y developed by hypertrophy . Middle and posterior myotomal reg ions, where the girth of pink fibres grad uall y increased, suppl y much of the propulsive force . This may be the reason for increased add ition of new sma ll pink fibres in the middle and posterior myolomal reg ions th at prov ide most of the power needed for locomotion .
The recruitment of new small white fibres in the myotomal regions was absent in almost all hi gher size classes (>8 cm F.L.; Fig. 3). It is indicative of the growth by the method of hypertrophy . In the present in vesti gati on the axia l muscul ature of B. soml/o showed that about 98% of the total muscle mass was occupied by the white fibres. Thus, the growth dynamics of white fibres may alone determine the growth and development of entire fish9
.19
. 22 . The result shows slight input of small new white fibre of lower size classes (5 and 8 cm F.L.) . The shifting of modal frequencies towards higher di ameter mode was very promill~ l1t in all myotomal regions (Fi g. 3). The regress ion as well as correlati on coeffici ent analyses (Tables J and 2) revealed that a significant hyperpl as ia or hypert rophy was not present in anterior myolomal region.
In the present in vestigation, no regional difference in the mode of growth as such between the three myotomal regions was observed in thi s fish. While fibres mostly grew by hyperpl as ia in lower size classes (up to 8 cm. F.L.) , therea fter by hypert rophy (Table J ). However, in higher size classes (> 12 cm F.L.), lhe mean fibre diameter did not deviate (Table 2)
Table 2 - Regression eq ual ions and lhe corre lali on coeffic ienl (r2) va lues of lhe Illean fibre diameler of red , pink and while Illuscle fibres in va ri ous fi sh size classes and body regions
Body region
Aillcrior Myolomal Middle MyolOmal Poslerior Myololllal Caudal Fi n Musele
Red fibres
y = 3.91 + O.li4x , r2 = 0.77 Y = 1.94 + 0 .98x , r2 = 0.96 y = 6.89 + 0. 33x, r2 = 0.54 Y = 8.56 + 0.25x, r2 = 0 .84
Pink fibres
y = 12.2 + O.8 17x, 1'2 = 0 .88 Y = 14.5 + 0 .828x, 1'2 = 0.76 Y = 18.7 +0.3 18x, r2 = 0.36 y = 1.8 1 + 1.752x , r2 =0.76
While fibres
y = 10.9 + l.92x , r2 = 0.80 y = 2 1.7 + 1.54x , 1'2 = 0 .58 Y = 19.9 + 1.45x,r2 = 0.54 Y = 15.9 - 3.9 Ix, r2 = 0 .83

t-l54 INDI AN J EXP 13 10L, AUGUST 2003
70
60 Apterior myotomal
60
~ 5 em .8 em 0 12 em 014 em 1111 15 em e!:I16 em I!:ro 17 em
40
30
20
10
78 Middle myotomal
;g- 60 ~
>- 60 u c: Q) 40 ::J 0-Q) 30 ... -Q) 20 ... .0 u::: 10
0 70
Posterior myotomal 60
60
40
30
20
10
0 1·10 11·:10 21..:Jl 31~ 41-50 51-60 61·70 71-60 81-00 91·100 1()1·110 111·1:10
Fibre diameter mode
Fig. :I _ M ean percent I reqllcneies of white fibrcs in three myotomal reg ions against fihre diamete r mocies (/1111 ) in vari oLis f ish size
classes.
even though there was more or less no input of small new white fibres . The growth or white fibres o f the cauda l rin muscle also showed si milar trend (Fig. 4) . The locomotion of thi s fish is mostl y short but high ~; peed burst type, which is indicative of the involvement of mostl y wh ite fibres and, up to certain ex tent, pink fibres. In B. sarallO, wh ite fibres showed greater recruitment of small new fibres in lower size classes. The recruitment of small new fibres ceased at certain stage in thi s f ish and further musc le growth occurred mostly by hypertroph y. However, in this species, the mean diameter of white fibres did not increase significantl y in higher size classes.
T h growth of red, pink and whi te fibres is by hyperplasia as well as hypertrophy. However, the frequencies of some intermediate diameter mode were
high in all size classes . The results clearl y show that hyperplasia was mostl y completed in the white fibres of the smallest fish studi es whereas, in red and pink fibres it continued up to q!Jite large body size. Postembryon ic musc le growth of fish myotome involves both hyperplasia and hypertrophy of exist ing fibres at different phases of their li vess.t4
. It is believed that new muscle fibres are initially formed in distinct germi nal zones at the lateral apices of the myotomes and only later in the early juvenile stage, when these zones are exhausted, fibre hyperplasia observed throughout the myotome. T he addi ti on of persistence myoblasts or myosatell ile cellsH24 also attributed towards the overall growth and development of muscle fibres in all myotomal regions25
.26
The contribution of hyperplas ia to muscle growth and

70 Red Fibres
60
50
40 o 12 em Bl 15 em ~ 17 em
30
20
10
0
60 Pink Fibres
~ 50 ~
>- 40 to) c: Q) 30 ::J CT
~ 20
Q) 10 .0
u: 0
60 White Fibres
50
40
30
20
10
1-10 11-20 21 -30 31-40 41 -50 51 -60 61-70 71 -311 81-90 91-100
Fibre d iameter mode
Fig. 4 - Mean percent frcqucncics of red, pink and wh i te fibres in the cauda l fin musc le against fibre diameter modes (/Lm) in vari ous fi sh size classcs.
deve lopment decl ines throughout the juvenile and the adu lt stages and in some spec ies ceases at around 40~7o
of the maxi mum body length)·7. Accordin g to the existing know ledge of f ish growth and deve lopment. a fast growing ri sh spec ies ex hibits fibre growth dynamics mainl y by hyperplasia where myostallite ce ll s produce small new whi te f ibres17.28_ The fi sh of the present study B. sarall a is a sma ll and medium growing spec ies where growth or musc le fibres
. 1 b I I' I <; 7 1<; OCCUITeu y l yperp aSia as we I as hypertrophy" .-. .
Acknowledgement Thi s work was funded by DS A and COSIST
Programmes of UGC, New Delhi sanctioned to Department of Biosc iences, Saurashtra Uni vers ity. Partia l support through a minor research proj ec t by Saurashtra Uni versit y is also acknowledged.
References I Bone Q. Locomo(or Illusc le . in Fish phvsiologv. Vol. 7.
edited by W S Horc & D J Randall (Academic Press. London ) 1978. 36 1.
2 Johnston I A . Musc le development and growth : Potential implication for Oesh quality in fi sh. Aqllaclllfllre. 177 (1999) 99.
3 Johnston I A. M c Lay H A _ Abercromby M & Robbi n, D. Phenotypic plasticity o f earl y myogenes is and satellite cell numbers in At lalll ic sa lmon spawning in an upland and a lowland tributary of a river system. J Ex!, Bioi. 203 (2000) 2539.
4 Johnston I A. Alderson R. Sandham C, Mitchell D. Sdkirk C. Dingwell A . icke ll D. Baker Remi , Robertson B. Whytc D & Springatc J. Pallerns of musc lc growth in ea rl y and late maturing populations of At lantic salmon (Saill /(l sa lor L. ). Aqllaclllllire. 189 (2000) 307.
5 Wea ther!ey A H. Approaches to understa nding fi sh growth. TraIlS Alii Fish Soc. I 19 ( 1(90) 662.
6 Ennion S. Gauvry L. BUllerworth P & Goldspi nk G. Small diameter white myotomal muscle fibrcs assoc iatcd wi th growth hyperplasia in the carp (Cv!, rillli s m rpio) exprcss a di stinct myosin heavy chain gene . .I Ex!, iJiol. 19, (1995) 1603.
7 Kundu R. Pate l D & M ansuri A P, Growth dynamics of myolOmal mu,cle fibres and their relationship with somatic dcvelopmcnt in somc mari ne teleosts. III dill II .I Ert' thol. 3~ ( 1994) 261.
X Weathcrl ey A H & Gill H S. Growth increasl:s produced hy hovinc growth hormone in grass pickeral. Es()x allleric{((IIiS l'enll iCir/aflls (Le Sucur), and thc underl y ing dynamics of musc le fibre growth . Aqllaclllfllre. 65 ( 1987) 55.
9 Kundu R & Mansuri A P. Growth dynamics of myotom31 muscle fibres in a carangid, CaJ"((lIx lI/alab((riclIs (Cuv .:t Val). J Fish Bioi , 35 ( 1990) 57.
10 Kundu R. Lakshmi R & Mansuri A P. Growth dynamics or the red. pink and whitc fibres in the caudal fin musclc in relation to the somati c growth in few marine fi sh, Mar /Jeh((l' Phv.liol, 19 ( 199 1) 113.
II Kundu R. Lakshmi R & Mansuri A P. Proporti on of difTc rcllI musc le fibres and their orientation in the myotomal and rin Illusc lcs of fi sh in rclati on to sw irnmin)! acti vit y . .I Cllrr /Jios(". 8 ( 1991 ) 89.
12 Kundu R & Man, uri A P. Growt h of pec toral muscle fibres in relati on to somati c growth in some marine fi shes. Nelfla J Zool, 42 ( 1992) 595 .
l :l Johnston I A & Horne Z_ Inllnunoc)'tnchl:lll ical invcsti gations of musc le dilTercnciation in the ,\t\antic Ilerri ng (C/ II[!e(( harellg lls: Tcleostci) . .1 M ar !:Jilll : \ .\\"(1(" UK. 74 ( 1994)79
14 Mascerello F, Rowclerson /\. Radae lli G_ Sea polo P/\ & Veg,e.elli A _ Differenciation and growth of musc le in fi sh SPlI /"{{S allra/({ (L. ) I. M yos in expression and organi LJti on of ribre types in lateral musc le from hatching to adull. J MIII"('le Res Cell MOIilily. 16 ( 1995) 2 13.
IS Carpene E & Veggelli A. Increase in musc lc ribres in the laterali s musc le (white portion ) of Mugilidae (Pieces. Teleostei ), E.rperielll io. 37 ( 198 1) 191.
16 Ca l vo J, Sexual differences in the increase of white musc lc fih res in A rge llline hake. M erlllccills IlIlbhis fro m the San Mata is Guld (Argell tina) . .1 Fisl; iJiol , 35 ( 1989) 207.
17 Day F. nil' Fishes of 11Ii1((( (Today and Tomorrows Bonk Agency. Ncw Del hi ) 1986.559.

856 INDIAN J EXP BIOL. AUGUST 2003
18 Lojda Z. Gossrau R & Schiebler T II. EII ; rlll l'
histlichelllistrv: A laborator v IIwllllol (Springer-Verl ag. New York ) 1979, 127.
1<) Kundu R. Lakshmi R & Mansuri AP, Growth dynamics of caudal and pectora l Illusc le fibres in a carangid. Carmlx
IIwlabariclIs (Cuv. & VaL ) and their poss ible relation w ith somati c growth. J Fish Bioi , :'7 ( 1990) 845.
20 Soka l R R & Rohlf F J. lJ iOlll l' IIY (W H Freeman & Co, San Francisco) 1969. 34:' .
21 Coughlin D J. Zhang G & Rome L C. Contracti on dynamics and power production of pink muscle of the scup (StI'1I0tOIllIlS chrysops) , J E.1'f} Bioi. 199. ( 1996) 270:..
22 Kundu R. Musc le fibre diameter and its relationship w ith body shape and size in some marine fi sh. J Cllrr Biosc. 8 ( 199 1) 3.
D Koumans J T M . Akster H A . Booms G H R. Lem/llens C J J & Osse J W M. Numbers of myosatellite cell s in wh ite ax ial muscle of growing fi sh Cvprilllls carpio L. (Teleostei). Alii J Allat. 192 ( 1991 ) 4 18.
24 I-liggi ns P J & Thrope J E, Hyperplasia and hypertrophy in the grow th of skeletal muscle in ju venile Atlantic Salmon. Sallllo salar L. J Fish Biol. ] 7 ( 1990) 505 .
25 Galloway T F, Kj orsvik E & Kryv i H. Musc le growth and development in At lantic cod larvae (Gadlls II lO rhlla L. ) related to diffe rent somalie growth rate. J EXI } Bioi. 202 ( I 999) 2 I I I.
26 Row lerson A . M ascarello F, Radae ll i G & Veggctti A . D i Ilcrenciation and growth of musc le in the fi sh Sparus (l lIrata (L. ): II. Hyperpl as ti c and hypert roph ic growth of lateral muscle from hatching to adult . J MlIscl l' Res Cell Motility. 16 ( 19<)5 ) 223.
27 Mau ro A . Sat ellite cel i oI' skeletal Illusc le fihres. J Biophv Bioc/,ell' Cw ol , <) ( 196 1) 493 .
28 Nag A C & arsall J R, Histogenesis of while and red musc le fibres of trunk Illusc le of fish Sallllo gai rrill er i . CytoiJios. 6 ( 1972) 227.
Recommended

















