
Primate Report: Research and Observation on the Common Squirrel Monkey (Saimiri sciureus)
Anthropology 67: Monkeys, Apes and Humans
Ishan Dutt
12/19/2012
Abstract: This paper consists of two components. The first is an overview of academic research conducted on squirrel monkeys in the wild. It presents their distribution, physical characteristics, social behavior, parenting strategies, food preference, vocalizations and conservation issues. The second component presents the findings of 6 hours of observations of Jack, a sub-adult male squirrel monkey at the Los Angeles Zoo. A comparison of observations on his behavior with squirrel monkey behavior in the wild and from other research on individuals in captivity presents insights into how zoos and wildlife parks can better provide desirable living conditions for squirrel monkeys.

2
RESEARCH COMPONENT
Distribution and Physical Characteristics The common squirrel monkey (Saimiri sciureus) is a small species of New World primate and is the
most populous of the Cebidae family, which includes the squirrel monkey and capuchin species (Boubli et al.
2008). The common squirrel monkey is distributed throughout much of the tropical rainforests of South
America, with populations present in Bolivia, Brazil, Colombia, Ecuador, French Guiana, Guyana, Peru,
Suriname, Paraguay and Venezuela (Hershkovitz 1984).
Fig 1: Distribution of common squirrel monkey (Saimiri sciureus). Image from: http://www.theanimalfiles.com/mammals/primates/monkey_common_squirrel.html
The common squirrel monkey is on the smaller scale of the New World monkeys, weighing
between 1.1 and 1.7lbs on average, and a total length (including the tail, which is about 1.5 times
the length of their bodies) of about 19 inches (Hershkovitz 1984). Both males and females are
marked by grey or olive coloration, with yellowish orange limbs and a white face. In the wild, the
common squirrel monkey has a lifespan of about 20 years (Hartwig 1995).

3
Fig 2: Common Squirrel Monkey at the Lakeland Wildlife Oasis, Milnthorpe, Cumbria, UK. © Lakeland Wildlife Oasis
Social Behavior
Squirrel monkey social groups are multi-male and multi-female and exhibit a high size
variance, ranging anywhere from 10-75 individuals (Mendoza et al. 1991; Boinski 1999). Within
these large groups, squirrel monkeys tend to exhibit closest association with members that match
their age-sex profile, with this segregation generally being attributed to the behavior of females
(Mendoza et al. 1991). Females often form small social circles within the context of the larger
group. These ‘cliques’ can be exclusionary, and allow certain females to gain certain benefits, such
as access to males or food resources. An individual’s association with a particular social circle is
impermanent, such that the makeup of smaller groups changes often, allowing different individuals
to enjoy advantages at different times.
Contrastingly, individual male-male relationships tend to be weaker, but also less prone to
change (Coe & Rosenblum, 1974). Within-sex relationships for both male and females are primarily
characterized by linear hierarchies, although such patterns of dominance are generally more
apparent in interactions between adult males (Mendoza et al. 1978). Female behavior also
influences social structure through dispersal behaviour – if females decide to stay within the group
they were born to, males may choose to form alliances and leave the larger group to avoid the
problem of inbreeding (Boinski 1999).

4
Another major factor that influences intra-group social behavior is the availability and
defendability of food resources (Boinski 1999). Scarcity of food resources leads to intense
competition, a condition under which the structure of small groups of strong female-female bonds
is advantageous to both acquiring food and defending it. In turn, the success of these female
groups influences the availability and defendability of females, which drives the distribution of
males within the larger group context (Boinski 1999). Monopolization and defence of food
resources is also a major outlet through which social dominance plays out. Subordinates of either
sex avoid any competition with dominants over food resources (Boinski 1999).
Empirical research has also shown significant variations in social behavior based on geography
(Boinski 1999). Groups in Peru have exhibited female dominance over males (Mitchell 1990)
whereas in some Surinamese groups, all males are dominant over any female, and female-female
bonds tend to be relatively weak. Throughout Costa Rica, squirrel monkey interactions between
both males and females are perhaps the least aggressive of any large primate social groups in the
world (Boinski 1987). Thus, while female behavior and food availability are general factors that
influence group dynamics, specific groups of common squirrel monkeys exhibit nuances in social
interactions.
Parental Strategies
Compared to the majority of nonhuman primates, which tend to exhibit careful and active
maternal care for infants (De Vore, 1965), squirrel monkeys generally seem to take on a more
passive role in raising their young (Rumbaugh 1965). Infants are fully dependent on their mothers
for the first 4-6 weeks of their lives and usually undergo weaning at sometime between 2-6 months
(Mendoza et al. 1991). During this time however, mothers display relatively little interaction
towards their young than most primates. For example, beyond cleaning them immediately after

5
birth, squirrel monkeys rarely groom their infants (Rumbaugh 1965). Mothers do not actively carry
their young; instead they are simply clung onto for the majority of the first month of the infant’s
life. After this time, mother and infant tend to remain physically separated except if the infant is
frightened or in danger (Rumbaugh 1965).
Although maternal care towards an infant is predominately carried out by its mother (Boinski
1987), the common squirrel monkey does occasionally display allo-parenting behavior (Soltis et al.
2004). This is almost exclusively carried out by females within the small social group, or ‘cliques’,
discussed above. These females will sometimes protect, carry, retrieve and nurse infants belonging
to others. On rare occasions, sub-adult males have been known to carry or protect young (Soltis et
al. 2004). Despite the existence of allo-parenting, mother squirrel monkeys have been observed to
aggressively restrict the interactions that other individuals in their social group may have with their
infant by physically stopping them from touching or carrying their young (Baldwin 1985).
Food Preference and Feeding Strategies
Although omnivorous, squirrel monkeys tend to show a preference for small fruits and
invertebrates, which make up the majority of their diet (Boinski 1999). Insects make up the
majority of their diet, and they spend about 75-80% of their waking day foraging for them (Boinski
1987). The preference for insects has a physiological reason as due to their short intestine tracts,
they find it easier to digest invertebrate protein than fruit or vegetation (Petrucci 1993). Insects are
supplemented with a variety of small, soft fruits, particularly berries although under conditions of
seasonality and shortages, squirrel monkeys can depend almost entirely on animal prey (Boinski
1999). Squirrel monkeys tend to forage in sizable groups, leading to food competition particularly
over less abundant sources such as fruits or vertebrate prey (e.g eggs) (Boinski 1999). Food
abundance usually leads to a reduction in overt exhibitions of dominance, and when plenty of food

6
is available subordinate individuals will be seen eating near dominants (DuMond 1968). Alcock
(1984) found that, in their foraging habits, squirrel monkeys tend to show a preference for diet
variety rather than purely pursuing food items with high caloric content. While foraging and
feeding, squirrel monkeys display unique positional behavior, generally preferring to sit on top of
branches rather than hang from them, which often leaves them vulnerable to predators (Boinski
1999).
Vocalizations
Squirrel monkeys have a large variety of calls and vocalizations, each of which pertains to a
particular social situation (Hammerschmidt et al. 2001). The most frequent vocalizations made in
agonistic social interactions are ‘caws’, ‘bawls’ and ‘shrieks’. These three are used when individuals
are claiming or defending a territory, before and during fights, during displays of dominance or
while engaging with predators (Hammerschmidt et al. 2001). An individual squirrel monkey may
also utter a ‘yap’ to warn of predators and to seek reinforcements to attack the predator as a mob.
During less aggressive social interactions, such as grooming, huddling or caring for an infant,
squirrel monkey make a low-pitched ‘chuck’ sound. Finally, ‘twitters’ and ‘cackles’ tend to be
produced in larger group situations, such as when a food source has been located or a lost
individual returns to the group and wants to make his announcement (Hammerschmidt et al. 2001).
Interestingly, the researchers who identified these various calls also concluded that the squirrel
monkey’s ability to produce these vocalizations is genetic rather than learned, as even infants
removed from groups and deaf individuals were able to produce all the different calls.

7
Conservation Issues and Species Survival The International Union for Conservation of Nature (IUCN) lists the common squirrel monkey
under the conservation status of Least Concern (Boubli et al. 2008). The reasons it provides for this
classification are a relatively wide range, adaptability to disturbed forest and lack of threat from
hunting due to its small size. Nevertheless, the squirrel monkey faces population decline from
habitat loss due to increased deforestation in the Amazon forest. According to IUCN, the squirrel
monkey is a protected species in every country that is in its distribution.
Two specific human activities that have placed some subspecies of the common squirrel
monkey under further threat or are pertinent to the discussion of species survival are medical
testing and exotic wildlife trade. Squirrel monkeys are popular test animals for numerous aspects
of biomedical research such as atherosclerosis, neuroscience, pharmacology, psychiatry, toxicology,
and vision research (Williams & Glasgow 2000). This is due to the physiological similarities between
humans and nonhuman primates that allow research done on the squirrel monkey to be pertinent
to human treatments. Due to their small size and relative ease of looking after, squirrel monkeys
are also popular as exotic pets. Nevertheless, habitat destruction poses a far more significant
threat to squirrel monkey populations than either primate medical research or the exotic wildlife
trade.

8
OBSERVATION COMPONENT Review of Observational Studies
Observations of squirrel monkey groups in captivity have provided a wealth of information
on social and individual behavior that complement our understanding of this species gained from
research in the wild. They also provide an opportunity for zoos and wildlife parks to improve the
living standards of their captive squirrel monkeys. Petrucci (1993) observed a group of 8 squirrel
monkeys at Tulsa Zoological Park in Oklahoma and found that changes in food distribution have an
impact on the relative time spent on foraging, feeding and social interactions such as grooming and
playing. When food is in abundance, as in captivity, displays of social dominance tend to be
reduced and monkeys of different hierarchy levels will be seen eating near each other comfortably,
although food sharing was not observed (Petrucci 1993). Food stealing, and associated agonistic
behavior such as fighting, is occasionally observed but these tend to be quickly resolved. Another
key conclusion from observation of squirrel monkeys is that the quality, rather than the quantity, of
space is the most important determinant in preventing boredom (Heideger 1967).
Squirrel monkeys, which spend the majority of their time in the wild foraging for food, can
easily become bored in a captive setting where food is abundant and easily obtained (DeVore
1965). Alcock (1984) observed that under certain conditions of food provision leading to boredom,
abnormal behavior such as regurgitation and food re-ingestion can occur. To prevent such
behaviors, zoos and wildlife parks must ensure that squirrel monkeys have some forms of
stimulation to allow for a greater variety of activities other than just eating or sleeping. Resource
distribution is one mechanism through which this may be achieved. Petrucci (1993) showed that
when no food bowls were provided and seeds were simply scattered and hidden throughout the
exhibit, squirrel monkeys spent almost all of their time foraging and the remainder of the time

9
grooming and playing. When seed was scattered and food bowls were provided, they spent 19% of
the time foraging, 29% eating and 26% playing or grooming. Thus, changing the manner in which
food is provided can stimulate different amounts of a variety of behaviors. Petrucci also noted that
squirrel monkeys rarely drank water. When they did, they drank from a small water fountain in the
exhibit .
Squirrel Monkey Observation at the Los Angeles Zoo
Table 1: 6 hours of observation of sub-adult male squirrel monkey (Jack) at the Los Angeles Zoo. Table shows exhibit areas frequented and activities observed during 24 15-minute intervals
Period Time Area 1 Area 2 Area 3 Notes
1 1000-1015 Rested in A alone
2 1015-1030 Retrieved corn from feeding area and returned to A
Feeding Area
Water
Fig 3: Layout of the Los Angeles Zoo Squirrel Monkey exhibit. For the purpose of observation, the exhibit was divided into three sections. Areas A,B,C and D are small shelters built on trees in the exhibit. Food and water areas are marked.
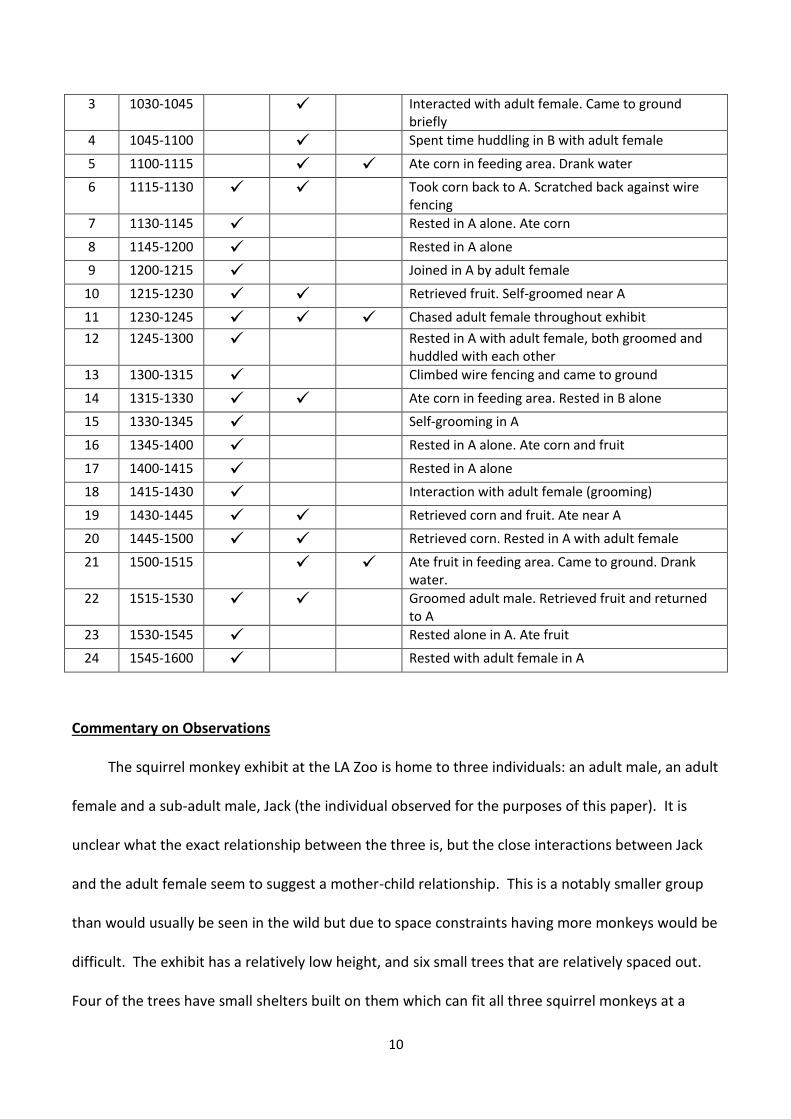
10
3 1030-1045 Interacted with adult female. Came to ground briefly
4 1045-1100 Spent time huddling in B with adult female
5 1100-1115 Ate corn in feeding area. Drank water
6 1115-1130 Took corn back to A. Scratched back against wire fencing
7 1130-1145 Rested in A alone. Ate corn
8 1145-1200 Rested in A alone
9 1200-1215 Joined in A by adult female
10 1215-1230 Retrieved fruit. Self-groomed near A
11 1230-1245 Chased adult female throughout exhibit
12 1245-1300 Rested in A with adult female, both groomed and huddled with each other
13 1300-1315 Climbed wire fencing and came to ground
14 1315-1330 Ate corn in feeding area. Rested in B alone
15 1330-1345 Self-grooming in A
16 1345-1400 Rested in A alone. Ate corn and fruit
17 1400-1415 Rested in A alone
18 1415-1430 Interaction with adult female (grooming)
19 1430-1445 Retrieved corn and fruit. Ate near A
20 1445-1500 Retrieved corn. Rested in A with adult female
21 1500-1515 Ate fruit in feeding area. Came to ground. Drank water.
22 1515-1530 Groomed adult male. Retrieved fruit and returned to A
23 1530-1545 Rested alone in A. Ate fruit
24 1545-1600 Rested with adult female in A
Commentary on Observations The squirrel monkey exhibit at the LA Zoo is home to three individuals: an adult male, an adult
female and a sub-adult male, Jack (the individual observed for the purposes of this paper). It is
unclear what the exact relationship between the three is, but the close interactions between Jack
and the adult female seem to suggest a mother-child relationship. This is a notably smaller group
than would usually be seen in the wild but due to space constraints having more monkeys would be
difficult. The exhibit has a relatively low height, and six small trees that are relatively spaced out.
Four of the trees have small shelters built on them which can fit all three squirrel monkeys at a

11
time. At the back of the exhibit there is a feeding area, where small fruits and corn are placed. One
end of the exhibit (in section 3 of Fig 3) has a small pool of water for drinking.
Over the course of 6 observation hours, Jack showed a preference for areas 1 and 2, only
venturing into area 3 on three occasions, twice to drink water and once during a play session with
the adult female. The primary reason for this seems to be that zoo visitors approached the squirrel
monkey exhibit from the side closest to area 1 and Jack (as well as the other two squirrel monkeys)
seemed extremely curious, often jumping onto the wire fence when visitors came closer to the
exhibit. When in any of the three given areas, Jack showed a preference for either sitting on top of
branches or resting inside one of the shelters. He spent a significant amount of time throughout
the day in shelter A, usually feeding alone or grooming and huddling with the adult female.
Compared to the Petrucci’s observations in the Tulsa Zoological Park, the squirrel monkeys in
the LA Zoo seemed to display greater signs of boredom. The easy access to food from the feeding
area meant that they had almost no foraging to do, and thus spent a lot of their time stationary on
branches or inside shelters. Feeding was observed during 12 of the 24 time periods, there were no
agonistic behaviors associated with feeding between any of the individuals and there were also no
examples of food sharing, which parallels Petrucci’s observations. Jack also only drank from the
pool in section 3 twice, confirming the observation that squirrel monkeys rarely drink water. There
were five observations of Jack coming to ground: twice to drink water, twice to retrieve food he
had dropped earlier and once during the play session with the adult female. This play session,
which lasted for slightly less than a minute during the 11th time period, was the most notable
interaction between Jack and the other squirrel monkeys. It was followed by a long grooming and
huddling session between the two, suggesting that they enjoyed a close relationship. Interactions
with the adult male were rare and Jack was only observed grooming him once during the entire
observation period.

12
The small group size and lack of stimulation in the LA Zoo seems to have given rise to
boredom amongst the squirrel monkeys. Compared to individuals in the wild, or in Tulsa Zoological
Park, Jack and the other two monkeys spent very little time foraging or interacting with each other
compared to time spent feeding and resting. This seems to confirm Heideger’s conclusion that
quality of space is more important than quantity. Despite the relatively small exhibit, Jack spent
almost all his time in either section 1 or 2, suggesting that he does not require a large space, but
simply a more stimulating environment. Encouraging foraging by hiding food, as was done in Tulsa
Zoological Park, would help stimulate both individual and social behavior to more accurately
represent how squirrel monkeys spend their time in the wild.

13
References
Alcock, J. Animal behavior: an evolutionary approach. Sunderland, MA: Sinauer As-sociates, Inc., Publishers; 1984.
Boinski S. (1987) Mating patterns in squirrel monkeys: implications for seasonal sexual dimorphism. Behavioral Ecology and Sociobiology 21:13–21.
Boinski, S. (1999), The social organizations of squirrel monkeys: Implications for ecological models of social evolution. Evolutionary Anthropology, 8: 101–112
Boubli, J.-P., Rylands, A.B., de la Torre, S. & Stevenson, P. (2008). Saimiri sciureus. In: IUCN 2012. IUCN Red List of Threatened Species. Version 2012.2. <www.iucnredlist.org>.
Coe, C. L., and Rosenblum, L.A (1974) Sexual segregation and its ontogeny in squirrel monkey social structure. Journal of Human Evolution 3: 551-561, 1974
DeVore, I., editor. Primate behavior. New York: Holt, Rinehart and Winston, Inc.; 1965
DuMond, F. V. The squirrel monkev in a seminatural environment. In: Rosenblum, L. A.; Coe, C. L., eds. The squirrel monkey. New York: Academic Press; 1968
Hammerschmidt, K., Freudenstein, T., & Jürgens, U. (2001). Vocal development in squirrel monkeys. Behaviour, 1179-1204.
Hartwig, W. C. (1995), Effect of life history on the squirrel monkey (Platyrrhini, Saimiri) cranium. American Journal of Physical Anthropology, 97: 435–449
Hediger, H (1967). Experiences of a zoo director in teaching animal behavior. The use of zoos and aquariums in teaching animal behavior. A symposium sponsored by The Amer-ican Association of Zoological Parks and Aquariums. Tampa, FL; 1967.
Hershkovitz, P. (1984), Taxonomy of squirrel monkeys genus Saimiri (Cebidae, platyrrhini): A preliminary report with description of a hitherto unnamed form. American Journal of Primatology., 7: 155–210.
Mendoza, S. P., Lyons, D. M. and Saltzman, W. (1991), Sociophysiology of squirrel monkeys. American Journal of Primatology, 23: 37–54
Mitchell CL. (1990). The ecological basis for female social dominance: a behavioral study of the squirrel monkey (Saimiri sciureus) in the wild. Ph.D. thesis, Princeton University: Princeton, NJ
Petrucci, S. T. (1993). Observations of the Foraging Behavior of Squirrel Monkeys (Saimiri
sciureus boliviensis) at the Tulsa Zoological Park. Bios, 64-71.

14
Rumbaugh, D (1965) Maternal care in relation to infant behavior in the squirrel monkey. Psychological Reports: Volume 16, Issue , pp. 171-176.
Soltis, J., Wegner, F. H., & Newman, J. D. (2005). Urinary prolactin is correlated with mothering and allo-mothering in squirrel monkeys. Physiology & behavior, 84(2), 295-301.
Williams L, Glasgow M. (2000) Squirrel Monkey Behavior in Research. ILAR Journal V41(1) 2000
Recommended





![Seminal quality and global proteomic analysis of ... · 3 43 Introduction 44 The squirrel monkey (Saimiri collinsi), a Neotropical primate endemic to the Amazon in 45 Brazil [1],](https://img.pdfslide.us/doc/110x75/60d88ab056669c0ded0c1857/seminal-quality-and-global-proteomic-analysis-of-3-43-introduction-44-the-squirrel.jpg)











