
This article was downloaded by:[Instytut Biologii Doswiadczaln][Instytut Biologii Doswiadczaln]
On: 20 April 2007Access Details: [subscription number 731999660]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Molecular Membrane BiologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713693962
Lysenin-His, a sphingomyelin-recognizing toxin,requires tryptophan 20 for cation-selective channelassembly but not for membrane binding
To cite this Article: , 'Lysenin-His, a sphingomyelin-recognizing toxin, requirestryptophan 20 for cation-selective channel assembly but not for membrane binding',Molecular Membrane Biology, 24:2, 121 - 134To link to this article: DOI: 10.1080/09687860600995540URL: http://dx.doi.org/10.1080/09687860600995540
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.
© Taylor and Francis 2007

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
Lysenin-His, a sphingomyelin-recognizing toxin, requires tryptophan20 for cation-selective channel assembly but not for membrane binding
KATARZYNA KWIATKOWSKA1, RENATA HORDEJUK2, PIOTR SZYMCZYK1,
MAGDALENA KULMA1, ABO-BAKR ABDEL-SHAKOR1, ANDRZEJ PL UCIENNICZAK3,
KRZYSZTOF DOL OWY2, ADAM SZEWCZYK1, & ANDRZEJ SOBOTA1
1The Nencki Institute of Experimental Biology, Warsaw and 2Department of Biophysics, Agriculture University SGGW,
Warsaw, and 3The Institute of Biotechnology and Antibiotics, Warsaw, Poland
(Received 7 April 2006; and in revised form 2 August 2006)
AbstractLysenin is 297 amino acid long toxin derived from the earthworm Eisenia foetida which specifically recognizessphingomyelin and induces cell lysis. We synthesized lysenin gene supplemented with a polyhistidine tag, subcloned itinto the pT7RS plasmid and the recombinant protein was produced in Escherichia coli . In order to obtain lysenin devoid ofits lytic activity, the protein was mutated by substitution of tryptophan 20 by alanine. The recombinant mutant lysenin-Hisdid not evoke cell lysis, although it retained the ability to specifically interact with sphingomyelin, as demonstrated byimmunofluorescence microscopy and by dot blot lipid overlay and liposome binding assays. We found that the lytic activityof wild-type lysenin-His was correlated with the protein oligomerization during interaction with sphingomyelin-containingmembranes and the amount of oligomers was increased with an elevation of sphingomyelin/lysenin ratio. Blue native gelelectrophoresis indicated that trimers can be functional units of the protein, however, lysenin hexamers and nanomers werestabilized by chemical cross-linking of the protein and by sodium dodecyl sulfate. When incorporated into planar lipidbilayers, wild type lysenin-His formed cation-selective channels in a sphingomyelin-dependent manner. We characterizedthe channel activity by establishing its various open/closed states. In contrast, the mutant lysenin-His did not form channelsand its correct oligomerization was strongly impaired. Based on these results we suggest that lysenin oligomerizes uponinteraction with sphingomyelin in the plasma membrane, forming cation-selective channels. Their activity disturbs the ionbalance of the cell, leading eventually to cell lysis.
Keywords: Channel assembly, hemolysis, lysenin, oligomerization, plasma membrane, toxin
Abbreviations: BSA, bovine serum albumin; DSS, disuccinimidyl suberate; IPTG, isopropyl-1-thio-b-D-
galactopyranoside; lysenin-His, lysenin fused with a polyhistidine tag at the N terminus; PBS, phosphate-buffered
saline; PCR, polymerase chain reaction; TBST, Tris-buffered saline containing Tween 20; wt, wild type.
Introduction
Several toxins of prokaryotic and eukaryotic origin
exert their cytolytic activity by binding to the cell
surface via specific interaction with sphingomyelin.
As more than half of the total amount of sphingo-
myelin in the cell resides in the plasma membrane,
mostly in its outer leaflet [1,2], the use of sphingo-
myelin as a target ensures efficient binding of the
toxins to the plasma membrane. After binding, toxin
molecules self-assemble into transmembrane pores
and subsequently kill the cell via osmotic shock, as
has been described for equinatoxin II of the sea
anemone Actinia equina [3,4]. The binding of toxins
to sphingomyelin and their cytolytic activity often
either require or are augmented by cholesterol.
Thus, pleurotolysin and ostreolysin, toxins of the
mushroom Pleurotus ostreatus, and VacA of Helico-
bacter pylori, bind to cholesterol- and sphingomye-
lin-containing membranes that probably acquire
liquid-ordered phase [5�7]. Similarly, the Vibrio
cholerae cytolysin displays dual specificity for cho-
lesterol and sphingolipids [8]. In the plasma mem-
brane, sphingolipids and cholesterol are believed to
form distinct microdomains of liquid-ordered state,
so-called rafts, which serve as signaling platforms for
Correspondence: Andrzej Sobota, Dept. Cell Biology, Nencki Institute of Experimental Biology, 3 Pasteur Street, 02-093 Warsaw, Poland.
Tel: (48-22) 5892 234. E-mail: [email protected]
Molecular Membrane Biology, March�April 2007; 24(2): 121�134
ISSN 0968-7688 print/ISSN 1464-5203 online # 2007 Informa UK Ltd
DOI: 10.1080/09687860600995540

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
a subset of plasma membrane receptors [9�11].
Owing to the sphingomyelin- and cholesterol-de-
pendent activity of a variety of pore-forming toxins,
the lipid composition of rafts is likely to render them
susceptible to the action of the proteins. Accord-
ingly, it has been shown that perfringolysin O from
Clostridium perfringens targets in vivo cholesterol
accumulated in plasma membrane rafts of intact
cells [12].
Recently, several proteins isolated from the coe-
lomic fluid of the earthworm Eisenia foetida have
been added to the list of sphingomyelin-dependent
cytolytic toxins [13]. Among them lysenin, a unique
297 amino acid long protein recognizes sphingo-
myelin exclusively. The conserved tryptophan resi-
dues of the protein are crucial for this interaction
[14,15]. The binding of lysenin to sphingomyelin is
accompanied by oligomerization of the protein and
both events are facilitated upon incorporation of
cholesterol. The cholesterol effect can be attributed
to the changes of sphingomyelin topology in the
membrane yielding local concentration of sphingo-
myelin into discrete microdomains [14,16]. Accord-
ingly, lysenin was applied for identification of
sphingomyelin-rich domains in the plasma mem-
brane [17].
The aforementioned oligomerization of lysenin is
believed to be a prerequisite for the formation of
pores in the membrane by the toxin [15,18]. As a
result of pore formation, lysenin causes leakage of
liposomes and lysis of erythrocytes in a sphingomye-
lin-dependent manner [14]. The cytolytic activity of
lysenin has also been claimed to cause death of
cultured mammalian cells and vertebrate spermato-
zoa [19,20]. It is of interest, however, that the first
bioactivity of lysenin to be discovered was contrac-
tion of strips of rat aorta. Lysenin at 10�9 M evoked
contraction of aorta muscles intensity of which
reached 60% of the maximum contraction induced
by 60 mM KCl [21]. This phenomenon can be
ascribed to lysenin-induced changes of sarcolemma
permeability leading to ion influx and muscle con-
traction. These data prompted us to study whether
lysenin can form transmembrane pores having ion
channel activity. For these studies we prepared
recombinant wild type lysenin with a polyhistidine
tag (wt lysenin-His). Wt lysenin-His recognized
sphingomyelin exclusively, evoked hemolysis and
formed oligomers, thus resembling the activity of
the native protein. When incorporated into planar
lipid bilayers containing sphingomyelin, lysenin-His
formed large-conductance cation-selective channels.
Channel activity was characterized by various open/
closed states. The protein mutated at tryptophan 20,
although still able to bind sphingomyelin, was
defective in oligomer and channel formation. We
suggest that lysenin, upon binding to sphingomyelin-
containing membranes, self-assembles into cation-
permeable channels and the resulting intracellular
ion imbalance can cause the death of cells.
Materials and methods
Preparation of synthetic lysenin gene
Synthetic lysenin gene was obtained from DNA
oligos by PCR, as shown on-line in supplementary
Figure 1, using the nucleotide sequence of lysenin
cDNA cloned by Sekizawa et al. [22]. The oligos
were divided into four groups consisting of: group I
� oligos from U1 to U6 and from J1_2 to J5_J6;
group II � from U6 to U11 and from J6_7 to J10_11;
group III � from U10 to U15 and from J10_11 to
J14_15; and group IV � from U15 to U20 and from
J15_16 to J19_20. Mixtures of Ux oligos (250 pM of
each oligo) of each group were phosphorylated with
T4 polynucleotide kinase and mixed with equimolar
joining oligonucleotides Jx in a solution containing
ligation buffer [23]. The mixtures were heated to
608C for 5 min and left for 20 min at room
temperature. After that, T4 DNA ligase was added
to each mixture and ligation was carried out at 168Cfor at least 12 h. After ligation, 1 ml of the mixtures
was taken for PCR amplification with pairs of 50 pM
primers (U1, J5_6), (U6, J10_11), (U10, J14_15)
and (U15, P2) for groups I, II, III and IV,
respectively. Commonly, 18 cycles of amplification
were required to obtain a sufficient amount of the
necessary DNA fragment. The amplified DNA
fragments obtained from group I and II after
electrophoresis and isolation from polyacrylamide
gel [24] were mixed together, the oligos U1 and
J10_11 were added and PCR was carried out to join
the two DNA fragments and to amplify their sum.
Similarly, fragments obtained from groups III and IV
of oligos were treated in the same manner. After
that, both (I�/II) and (III�/IV) DNA fragments
were mixed in one tube and amplified by PCR using
the U1 and P2 oligos as primers. The final PCR
product containing cDNA of lysenin was isolated
from polyacrylamide gel, digested with EcoRI and
HindIII restriction nucleases and cloned into pBlue-
script plasmid (Stratagene, La Jolla, CA).
The nucleotide sequence of lysenin cDNA was
modified by introducing six histidine codons at the
5? end of the coding sequence. For this we used the
sense primer 5?GGGGAATTCATATGCATCATC
ACCATCATCACTCTGCAAAAGCTGCAGAAG
G3? designed to contain EcoRI and NdeI restriction
sites, codons for the 6�/His tag, and the 5? end of
the open reading frame, and the antisense primer
5?GCAGCGCGGGTGAAGCTTAACCAACCAC
122 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
CTCG3? designed to contain a HindIII site and 3?end of the open reading frame with a stop codon
(restriction sites and insert sequence are underlined,
stop and start codons are double underlined). The
product was initially cloned into pBluescript plasmid
using EcoRI and HindIII sites. Subsequently, the
900bp NdeI/HindIII fragment was cloned into the
expression plasmid pT7RS (the GenBank accession
number for the sequence of pT7RS expression
vector is AY923866). This construct of full length
lysenin-His was used to create the W20A point
mutant of recombinant lysenin. W20A lysenin-His
was amplified by PCR and the Quick Change
Mutagenesis kit (Stratagene) with the sense primer
5? GATGTGGTGGCAGTAGCGAAAGAAGGCT
ATGTATACG3?, and the antisense primer 5? CGT
ATACATAGCCTTCTTTCGCTACTGCCACCA
CATC3? designed to contain the W20A mutation
(underlined).
Purification of recombinant proteins
For bacterial expression of the His-tagged wt and
W20A lysenin, the coding sequences were subcloned
into the bacterial expression vector pT7RS using
EcoRI/HindIII restriction sites. The plasmids were
used to transform the BL-21-DE3 strain of Escher-
ichia coli . Transfectants were grown in LB medium
containing 100 mg/ml ampicillin at 378C to A600 0.6
when 1 mM isopropyl-1-thio-b-D-galactopyranoside
(IPTG) was added. After 20 h at 258C, bacteria were
harvested, sonicated and lysed with 50 mg/ml lyso-
zyme (20 min, 308C) followed by 1% Triton X-100
and 10 mg/ml DNAse (20 min, 308C). The recom-
binant proteins were purified on a HIS-Select Nickel
HC Affinity Gel column (Sigma-Aldrich, St Louis,
MO). They were eluted from the column with 100
mM imidazole, 300 mM NaCl, 5 mM b-mercap-
toethanol, 5 mM phosphate buffer, pH 8.0, and
dialyzed against phosphate-buffered saline (PBS)
containing 5 mM b-mercaptoethanol. The concen-
tration of wt and W20A lysenin-His was established
after 10% SDS-PAGE by quantifying the Coomas-
sie-Blue stained bands using bovine serum albumin
(BSA) as a standard.
Protein-lipid overlay assay
For analysis following lipids were used: dioleoyl
phosphatidylcholine, sphingosine, C6�/ and C16�/
ceramides, cholesterol, bovine brain or semisyn-
thetic bovine brain sphingomyelin, and galactocer-
ebrosides (all from Sigma-Aldrich). The molecular
weight of the latter lipid was tentatively estimated as
1200 Da. Lipid samples of 1 ml containing various
amounts of lipids (5-100 pmol) were spotted onto a
nitrocellulose membrane (0.45 mm, Santa Cruz
Biotechnology, Santa Cruz, CA) and allowed to
dry for 30 min [25]. To enhance binding of the
lipids to the membrane, the method of Taki and
Ishikawa [26] was used. The membrane was blocked
for 1 h at 208C with 1% gelatin and 1% polyvinyl-
pyrrolidone in Tris-buffered saline containing 0.03%
Tween-20 (TBST), exposed for 2 h at 208C to 3 mg/
ml wt or W20A lysenin-His in 1% gelatin/TBST,
and incubated with goat anti-His IgG-peroxidase
(Sigma-Aldrich) in 1% gelatin/TBST (1 h, 208C).
Immunoreactive spots were visualized with the
SuperSignal West Pico chemiluminescent substrate
(Pierce, Rockford, IL).
Hemolytic assay
Serial dilutions of recombinant wt and W20A
lysenin-His up to 200 ng/ml (6 nM) were incubated
with 7�/107 sheep erythrocytes in 1 ml of TBS for
1 h either at 48C or at 208C. After pelleting of non-
lysed cells (200 g , 48C, 5 min), the amount of
released hemoglobin was estimated at 405 nm. The
hemolysis caused by lysenin-His was expressed as
the percentage of maximal hemolysis (100%) found
after osmotic lysis of erythrocytes in distilled water.
Immunofluorescence
Sheep red blood cells (3�/107/sample) were fixed
with 1% glutaraldehyde/TBS (20 min, 48C) and
after quenching with 50 mM NH4Cl/TBS they were
pelleted onto coverlips coated with poly-L-lysine
(200 g , 5 min, 208C). After blocking with 3% BSA/
TBS, the cells were incubated with wt or W20A
lysenin-His (3 mg/ml in TBS containing 5 mM
imidazole; 30 min, 208C). The presence of imidazole
during incubation of lysenin-His with erythrocytes
abrogated nonspecific interactions of the His tag
with the negatively charged cell surface. Subse-
quently, cells were exposed to rabbit anti-His IgG
(Santa Cruz Biotechnology; 30 min, 208C) followed
by goat anti-rabbit IgG-FITC (Jackson Immuno
Research Labotratories, West Grove, PA). The
samples were examined under a Nikon microscope
[27]. Fluorescent cell images were collected with a
digital Nikon DS-5M camera.
In controls, cell incubation with lysenin-His was
omitted or lysenin-His denatured by 10 min heating
at 608C was applied. In both cases, no staining of the
erythrocyte membrane was detected. In a series of
experiments cells were treated with 70 mU/ml of
bacterial sphingomyelinase (1 h, 378C; Sigma-
Aldrich) prior to fixation and incubation with the
recombinant proteins.
Oligomers of lysenin form channels 123

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
Preparation of liposomes, erythrocyte ghosts and
lysenin-His binding
To prepare small unilamellar vesicles, semisynthetic
sphingomyelin and dioleoyl phosphatidylcholine
with or without cholesterol were mixed in different
molecular ratios and dried from a chloroform/
methanol (1:1, by vol.) under nitrogen. The lipids
were resuspended in 100 ml of PBS, vortexed and
sonicated on ice for 30 min. Vesicles were pelleted
(200,000 g , 2 h, 48C) and resuspended in 16.5 ml of
PBS. For experiments, 4 ml of the liposome suspen-
sion was mixed with 8 ml of a solution containing 1
mg (2.5 mM) of lysenin-His, 50 mM NaCl, 20 mM
imidazole and 50 mM phosphate buffers, pH 8.0. In
the mixture, the total concentration of sphingomye-
lin and phosphatidylcholine was 1 mM. After 45 min
incubation at 208C, the liposomes were diluted with
300 ml of PBS and pelleted as above. In a series of
experiments, prior to the centrifugation, the mixture
was supplemented with 2 mM disuccinimidyl sube-
rate (DSS, Pierce) and 2 h later, 20 mM Tris, pH
7.4, was added to stop cross-linking of the protein.
For preparation of sheep red blood cell ghosts,
erythrocytes (7�/109/sample) were lysed in distilled
water and washed five times with the water by
centrifugation (15,000 g , 6 min, 48C). The ghosts
were suspended in 50 ml of solution containing 0.4
mg lysenin-His (0.24 mM), 40 mM imidazole and 50
mM phosphate buffer, pH 8.0, and 0.1�0.8 M
NaCl. After incubation (30 min, 208C), ghosts
were washed with PBS, suspended in 20 ml SDS
sample buffer and analysed by SDS-PAGE.
SDS-PAGE, blue native gel electrophoresis and
immunoblotting
For SDS-PAGE, pellets of liposomes obtained after
incubation with lysenin-His were dissolved in 45 ml
of 2% SDS/1% b-mercaptoethanol sample buffer,
boiled for 5 min and loaded onto 7% gel (14 ml).
After electrophoresis and transfer to nitrocellulose,
the proteins were visualized by immunoblotting with
anti-His IgG-peroxidase and chemiluminescence.
Monomers and oligomers of lysenin-His were quan-
tified densitometrically in Fluor-S MultiImager
equipped with Quantity One software (Bio-Rad).
For normalization, the densitometric data were
expressed in relation to the monomer level detected
in liposomes at sphingomyelin/lysenin ratio 8:1
without cholesterol, and arbitrary equalized to 1.
For blue native gel electrophoresis, liposomes
pelleted after incubation with lysenin-His were
suspended in 12 ml of solubilization buffer contain-
ing 1% dodecyl-b-maltoside, 20% glycerol, 25 mM
BisTris-HCl, 50 mM 6-amino-caproic acid, pH 7.0,
and 0.5% Coomassie Blue Serva G. The samples
were loaded onto a 7% polyacrylamide gel and
developed for 5 h (160 V, 48C) according to Cline’s
protocols [http://www.hos.ufl.edu/clineweb/Proto-
cols/BNgel.htm]. Ferritin (880 kDa and 440 kDa)
and BSA (132 kDa and 66 kDa) were applied as
molecular weight standards. Subsequently, lanes of
interest were dissected from the gel, incubated in
SDS electrophoresis buffer for 10 min, mounted on
the top of 9% SDS-PAGE gels and run for the
second dimension analysis at 120 V. After transfer to
nitrocellulose, the samples were immunoblotted
with anti-His as above.
Electron microscopy
For ultrastructural studies, multilamellar liposomes
were prepared by mixing semisynthetic sphingomye-
lin, dioleoyl phosphatidylcholine and cholesterol
(1:2:2 molar ratio) or dioleoyl phosphatidylcholine
and cholesterol (3:2 molar ratio). The lipids were
dried under nitrogen, resuspended in 100 ml PBS,
vortexed, sonicated for 2 min on ice and collected
by centrifugation (15,000 g , 15 min, 48C). The
liposomes (6 mM total phospholipid content) were
incubated with 7 mM wt or W20A lysenin-His for
45 min at 208C. After incubation, the vesicles
were treated with 1% glutaraldehyde in PBS (20
min, 208C) and applied onto poly-L-lysine-coated
and formvar/carbon-treated grids. After 20 min, the
samples were washed twice with PBS and once with
water, and counterstained with 2% uranyl acetate.
They were examined under a JEM-1200EX (JEOL)
microscope.
Planar lipid bilayer technique and lysenin reconstitution
The planar lipid bilayer technique was applied as
described previously [28]. In brief, planar phospho-
lipid bilayers were formed in a 250 mm diameter hole
which separated two chambers (cis 2 ml and trans 3
ml internal volume). The chambers contained 450/
150 mM KCl, 5 mM Hepes, pH 7.0 (adjusted with
KOH). The outline of the aperture was coated with
a lipid suspension and dried with N2 prior to bilayer
formation to improve membrane stability. Planar
phospholipid bilayers were painted using azolectin
(L-a-phosphatidylcholine, Sigma-Aldrich) or azolec-
tin enriched with 3% sphingomyelin and 1% cho-
lesterol � all in n-decane at a final concentration of
25 mg of lipid/ml. Formation and thinning of the
bilayer were monitored by capacitance measure-
ments. Final capacitance values ranged from 110�200 pF. Electrical connections were made using
Ag/AgCl electrodes and agar salt bridges (3 M KCl)
to minimize liquid junction potentials. Voltage was
applied to the cis compartment of the chamber and
124 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
the trans compartment was grounded. Solutions of
wt lysenin-His or W20A lysenin-His in 50 mM KCl,
20 mM Hepes, pH 7.2 (adjusted with KOH) were
added to the trans compartment. In a series of
experiments native lysenin isolated from Eisenia
foetida (Peptides International, Louisville, KY) was
used. All measurements were carried out at room
temperature.
Data recording and analysis
The current was measured using a Bilayer Mem-
brane Admittance Meter (model ID 562, IDB,
Gwynadd, UK). Signal was filtered at 0.2 kHz
(Low Pass Bessel Filter 4 Pole, Warner Instrument
Corp.), digitized (A/D converter 1401, Cambridge
Electronic Design, UK) and transferred to a PC for
off-line analysis by the CED Electrophysiology
Package V6.41 and pClamp6 software (Axon
Instruments, Union City, CA) and plotted in
Microcal Origin. All measurements were conducted
in asymmetric ionic conditions (450/150 mM KCl,
pH 7.0 cis/trans). Recordings were low�pass filtered
at 200 Hz. Channel conductances were expressed as
the mean9/SD.
Results
Expression of wt lysenin-His
The synthetic lysenin gene was constructed from
partially overlapping DNA fragments obtained by
PCR as shown on-line in supplementary Figure 1.
The prepared cDNA of recombinant lysenin was
cloned into pT7RS expression plasmid and con-
firmed by sequencing. The predicted amino acid
sequence of the recombinant lysenin was identical to
the sequence of native lysenin cloned from Eisenia
foetida by Sekizawa et al. [22]. A tag consisting of six
histidine residues was added without any linker at the
N terminus of the protein to facilitate its purification
and detection in further analysis. The lysenin-His
cDNA was expressed in Escherichia coli that were
induced with IPTG and cultured at 258C. Under
these conditions, most of the protein remained
soluble and was purified under non-denaturing
conditions in one step affinity chromatography on
Ni2�-agarose. The purified protein migrated as a
41 kDa band on SDS-PAGE and was recognized by
anti-His antibody (see Figure 3B).
Wild type and W20A lysenin-His bind sphingomyelin
selectively
The tryptophan residues of lysenin were found to be
important for the protein activity [29]. Therefore, in
an attempt to interfere with the protein properties,
we introduced a point mutation in lysenin-His
cDNA replacing the N-terminal-most tryptophan
residue 20 with alanine (on-line supplementary
Figure 1, shadowed). To analyse the ability of
W20A lysenin-His to recognize and bind sphingo-
myelin, the protein-lipid overlay assay was applied.
Both wt and W20A lysenin-His at 3 mg/ml detected
as little as 5 pmol of sphingomyelin (Figure 1). The
mutant lysenin-His retained selectivity for sphingo-
myelin binding found for wt lysenin-His. Aside from
sphingomyelin, neither of the recombinant proteins
recognized any other sphingolipid nor phosphatidyl-
choline or cholesterol (Figure 1).
To assess whether the recombinant proteins were
able to bind sphingomyelin of the plasma mem-
brane, the proteins were incubated with sheep
erythrocytes, whose membrane contains 51% sphin-
gomyelin of total phospholipids [30]. Distinct stain-
ing of the erythrocyte surface was found when the
cells were incubated with wt lysenin-His or with
W20A lysenin-His (Figure 2). Both recombinant
proteins labeled the cells with similar intensity.
Pretreatment of erythrocytes with bacterial shingo-
myelinase markedly reduced the staining, as shown
in Figure 2 (inset) for wt lysenin-His.
Wild type, but not W20A lysenin-His, forms oligomers
Recombinant wt lysenin-His evoked hemolysis of
sheep erythrocytes in a dose dependent manner,
Figure 1. Selective recognition of sphingomyelin by wt and W20A
lysenin-His. Interaction of the proteins with various lipids was
examined using a protein-lipid overlay assay. Indicated amounts of
lipids were spotted onto nitrocellulose and exposed to 3 mg/ml of
wt or W20A lysenin-His and anti-His IgG-peroxidase. Immuno-
reactive spots were revealed by chemiluminescence. DOPC,
dioleoyl phosphatidylcholine; SM, sphingomyelin, Sph, sphingo-
sine; Gal-Cer, galactocerebrosides; Cer, ceramide; Chol, choles-
terol. A representative of five experiments is shown.
Oligomers of lysenin form channels 125

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
resembling the activity of the native protein [15]. To
cause 50% hemolysis at 48C, 28 ng/ml of wt lysenin-
His was required (Figure 3A, open squares). In
contrast, W20A lysenin-His did not cause hemolysis
even at 200 ng/ml both at 48C and at 208C (Figure
3A, closed circles, and results not shown).
This difference prompted us to compare the
ability of wt and W20A lysenin-His to oligomerize.
For this purpose, proteins were mixed with sphin-
gomyelin/phosphatidylcholine liposomes, and after
pelleting the samples were subjected to SDS-PAGE
analysis. In the liposomes, the sphingomyelin con-
tent varied from 0.02 mM to 0.7 mM yielding
sphingomyelin/lysenin ratio in the range from 8:1
to 280:1. When the ratio exceeded 40:1, a SDS-
resistant oligomer of wt lysenin-His of about 280
kDa was assembled at the expense of the 41 kDa
monomer (Figure 3B). Oligomerization of wt lyse-
nin-His was facilitated by cholesterol and was
observed already at sphingomyelin/protein ratio�/
20:1 when liposomes were supplemented with cho-
lesterol in an equimolar ratio to sphingomyelin. At
the cholesterol presence, the oligomer of wt lysenin-
His prevailed with minute amounts of the monomer
left when the sphingomyelin/lysenin ratio reached
280:1 (Figure 3B). However, an increasing sphingo-
myelin/lysenin ratio facilitated the protein oligomer-
ization regardless of the cholesterol presence in
liposomes (Figure 3B). Studies performed with
W20A lysenin-His confirmed that the protein bound
to sphingomyelin-containing liposomes in amounts
comparable to wt lysenin-His (Figure 3B). In con-
trast to wt lysenin-His, the vast majority of W20A
lysenin-His bound to the liposomes existed as the
monomer. The mutant lysenin formed only minute
amounts of the SDS-resistant 280 kDa oligomer at
sphingomyelin/protein ratio�/40:1 and the protein
oligomerization was slightly promoted by cholesterol
(Figure 3B). Similarly, the monomer of W20A
lysenin-His prevailed after incubation of the protein
with sheep erythrocyte ghosts instead of sphingo-
myelin-containing liposomes (Figure 3C). Oligo-
merization of W20A lysenin-His was moderately
improved by increasing salt concentration up to 0.8
M during erythrocyte ghost binding. However, high
salt concentrations, 0.1�0.8 M NaCl, impaired
binding and oligomerization of wt lysenin-His dur-
ing incubation with erythrocyte ghosts (Figure 3C).
The assembly of lysenin oligomers was further
analysed using blue native gel electrophoresis which
allowed us to isolate native protein complexes.
Dissected lanes of the native gel were subsequently
subjected to SDS-PAGE at denaturing conditions
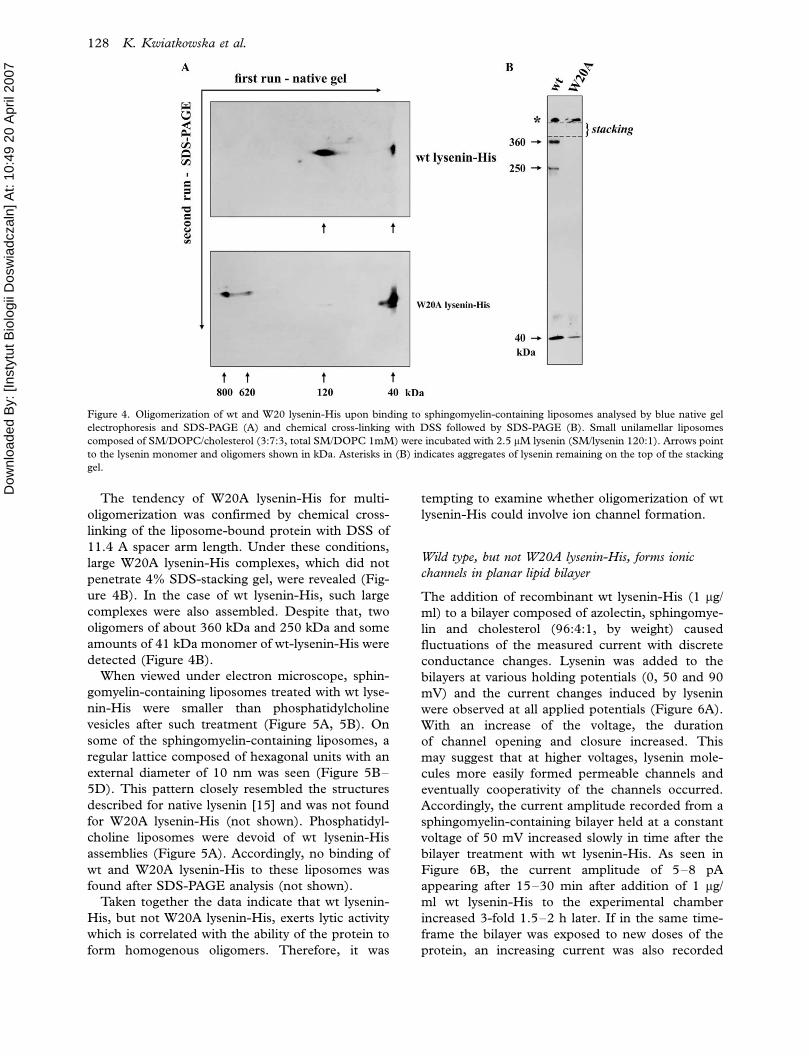
for analysis in the second dimension (Figure 4A).
This approach confirmed that wt lysenin-His oligo-
merized efficiently upon binding to sphingomyelin-
containing lipososmes (sphingomyelin/protein ratio
of 120:1), leaving small amounts of the monomer.
However, the apparent molecular weight of the wt
lysenin-His oligomer found in the native gel electro-
phoresis was the half of the value of the SDS-
resistant oligomer and reached about 120 kDa
(Figure 4A, compare with Figure 3B). In contrast,
W20A lysenin-His bound to the liposomes mainly as
the monomer (Figure 4A). Aside from the mono-
mer, the protein formed very large oligomers of
about 800 kDa and 620 kDa with traces of 120 kDa
complex (Figure 4A). It is of note, that in the second
run during SDS-PAGE, lysenin complexes and the
monomer migrated at the same velocity. This
indicates that after isolation at native conditions,
including treatment of liposomes with a non-ionic
detergent dodecyl-b-maltoside, the oligomers were
decomposed by SDS.
Figure 2. Binding of wt and W20A lysenin to the surface of sheep erythrocytes. Cells were fixed with 1% glutaraldehyde and exposed to the
proteins in the presence of 5 mM imidazole, followed by rabbit anti-His IgG and anti-rabbit IgG-FITC. Inset shows cells treated with
bacterial sphingomyelinase (70 mU/ml, 1 h, 378C) before fixation and staining with lysenin. Bar, 10 mm.
126 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
Figure 3. W20A lysenin-His is devoid of hemolytic activity and inefficiently forms oligomers which are characteristic for wt lysenin-His. (A)
Hemolytic activity of wt lysenin-His (open squares) and its lack in W20A lysenin-His (closed circles). Sheep erythrocytes (7�/107/ml) were
incubated with the recombinant proteins at indicated concentrations for 1 h at 48C. In controls, cells were suspended in H2O to estimate
100% hemolysis. The results are mean9/SE from five experiments. (B, C) Wt lysenin-His forms a distinct SDS-resistant oligomer upon
binding to sphingomyelin-containing liposomes (B) and sheep red blood cell ghosts (C). (B) Small unilamellar liposomes composed of SM/
DOPC with or without cholesterol (1 mM total phospholipids) were incubated with 2.5 mM wt or W20A lysenin-His at the SM/lysenin ratio
from 8:1 to 280:1. Pelleted liposomes were subjected to 7% SDS-PAGE under denaturing condition and analyzed for the presence of
lysenin monomers and oligomers by immunoblotting with anti-His (upper panel). In lane ‘‘no liposomes’’ 0.2 mg of lysenin-His was applied.
On the left, molecular weight standards are shown. Lower panel: Quantification of lysenin monomers (closed symbols) and oligomers (open
symbols) based on a densitometric analysis of blots shown in the upper panel. Triangles, liposomes with cholesterol; circles, liposomes
without cholesterol. Data are mean9/SE from three experiments. (C) Erythrocyte ghosts were incubated with 0.24 mM recombinant
proteins in the presence of 0.1�0.8 M NaCl and analysed by SDS-PAGE and immunoblotting with anti-His. Data shown are representative
of three experiments.
Oligomers of lysenin form channels 127
Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
The tendency of W20A lysenin-His for multi-
oligomerization was confirmed by chemical cross-
linking of the liposome-bound protein with DSS of
11.4 A spacer arm length. Under these conditions,
large W20A lysenin-His complexes, which did not
penetrate 4% SDS-stacking gel, were revealed (Fig-
ure 4B). In the case of wt lysenin-His, such large
complexes were also assembled. Despite that, two
oligomers of about 360 kDa and 250 kDa and some
amounts of 41 kDa monomer of wt-lysenin-His were
detected (Figure 4B).
When viewed under electron microscope, sphin-
gomyelin-containing liposomes treated with wt lyse-
nin-His were smaller than phosphatidylcholine
vesicles after such treatment (Figure 5A, 5B). On
some of the sphingomyelin-containing liposomes, a
regular lattice composed of hexagonal units with an
external diameter of 10 nm was seen (Figure 5B�5D). This pattern closely resembled the structures
described for native lysenin [15] and was not found
for W20A lysenin-His (not shown). Phosphatidyl-
choline liposomes were devoid of wt lysenin-His
assemblies (Figure 5A). Accordingly, no binding of
wt and W20A lysenin-His to these liposomes was
found after SDS-PAGE analysis (not shown).
Taken together the data indicate that wt lysenin-
His, but not W20A lysenin-His, exerts lytic activity
which is correlated with the ability of the protein to
form homogenous oligomers. Therefore, it was
tempting to examine whether oligomerization of wt
lysenin-His could involve ion channel formation.
Wild type, but not W20A lysenin-His, forms ionic
channels in planar lipid bilayer
The addition of recombinant wt lysenin-His (1 mg/
ml) to a bilayer composed of azolectin, sphingomye-
lin and cholesterol (96:4:1, by weight) caused
fluctuations of the measured current with discrete
conductance changes. Lysenin was added to the
bilayers at various holding potentials (0, 50 and 90
mV) and the current changes induced by lysenin
were observed at all applied potentials (Figure 6A).
With an increase of the voltage, the duration
of channel opening and closure increased. This
may suggest that at higher voltages, lysenin mole-
cules more easily formed permeable channels and
eventually cooperativity of the channels occurred.
Accordingly, the current amplitude recorded from a
sphingomyelin-containing bilayer held at a constant
voltage of 50 mV increased slowly in time after the
bilayer treatment with wt lysenin-His. As seen in
Figure 6B, the current amplitude of 5�8 pA
appearing after 15�30 min after addition of 1 mg/
ml wt lysenin-His to the experimental chamber
increased 3-fold 1.5�2 h later. If in the same time-
frame the bilayer was exposed to new doses of the
protein, an increasing current was also recorded
Figure 4. Oligomerization of wt and W20 lysenin-His upon binding to sphingomyelin-containing liposomes analysed by blue native gel
electrophoresis and SDS-PAGE (A) and chemical cross-linking with DSS followed by SDS-PAGE (B). Small unilamellar liposomes
composed of SM/DOPC/cholesterol (3:7:3, total SM/DOPC 1mM) were incubated with 2.5 mM lysenin (SM/lysenin 120:1). Arrows point
to the lysenin monomer and oligomers shown in kDa. Asterisks in (B) indicates aggregates of lysenin remaining on the top of the stacking
gel.
128 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
(Figure 6C). Channel formation induced by wt
lysenin-His appeared to be a function of the protein
concentration in the range of 1�5 mg/ml. At con-
centrations exceeding 6 mg/ml the current increased
rapidly until the membrane broke.
Wt lysenin-His formed channels only in mem-
branes containing sphingomyelin, since no channel-
like activity was recorded from bilayers composed of
azolectin alone in the presence of 2�6 mg/ml of the
protein (Figure 6D).
Substitution of tryptophan 20 by alanine in
lysenin-His abolished the ability of the protein to
form channels in sphingomyelin-containing mem-
branes (Figure 6E). Addition of 2�6 mg/ml of W20A
lysenin-His to an azolectin/sphingomyelin/choles-
terol bilayer had no effect on current measurements
(Figure 6E).
Lysenin forms cation-permeable channels
To characterize the current-voltage relationship for
single wt lysenin-dependent channel openings, cur-
rent-time traces were recorded at holding potentials
ranging from 70 mV to �/150 mV (450/150 mM
KCl, cis/trans) (Figure 7A). At positive voltage,
frequent short-lasting channel closures were ob-
served, while under negative voltage, up to �/110
mV, the channels displayed longer openings of lower
amplitude. However, the potential of �/150 mV
induced prolonged closure of the channels. All the
traces shown were recorded when the lysenin
channels exhibited activities devoid of initial flicker-
ing observed immediately after incorporation of
lysenin into the membrane.
The current-voltage plot derived from the re-
cordings presented in Figure 7A was a straight
line with a reversal potential Urev�/�/399/2 mV
(Figure 7B), indicating that the channel is selective
for cations. The most frequent single channel
conductance was 1009/9 pS, as determined from
30 measurements for 1 mg/ml wt lysenin-His. In
experiments with higher concentrations of the
protein (2�5 mg/ml) we observed cation-selective
channels with conductances ranging from 350 pS
to 890 pS and complex kinetics. Occasionally, we
also found a similar behavior of the channels
when 1 mg/ml of wt lysenin-His was applied (not
shown).
Since wt lysenin-His is a recombinant protein, the
channel-forming properties of native lysenin isolated
from Eisenia foetida were also examined. The native
protein formed cation-selective channels of similar
properties (not shown).
Figure 5. Wild type lysenin-His assembles into a honeycomb-like structure when bound to sphingomyelin-containing liposomes.
Recombinant wt lysenin-His (7 mM) was incubated with (A) DOPC/cholesterol (3:2) or (B�D) SM/DOPC/cholesterol mulilamellar
liposomes (1:2:2, 6 mM total phospholipids, SM/lysenin 300:1) after which samples were negatively stained and examined under an
electron microscope. Bars, 200 nm in (A, B), 50 nm in (C, D). Arrow in (B) indicates lattice formed by wt-lysenin-His.
Oligomers of lysenin form channels 129

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
Discussion
The data presented in this report indicated that wt
lysenin-His forms oligomers the size of which varied
between 120 kDa and 280 kDa, depending on
conditions of their isolation. On the other hand,
lysenin-His mutated on tryptophan 20 failed to
form correct oligomers and existed mainly as the
monomer when bound to sphingomyelin-containing
membranes. The ability of wt lysenin-His for oligo-
merization was correlated with the ability of the
protein to assembly ion-permeable, cation-selective
channels in a sphingomyelin-dependent manner.
The formation of channels by wt lysenin-His
required an interaction of the protein with sphingo-
myelin, as no channel activity was detected from
azolectin membranes even at relatively high wt
lysenin-His concentrations. However, sphingomye-
lin binding was not sufficient for the protein activity.
Mutant W20A lysenin-His specifically recognized
and bound sphingomyelin as determined by lipid-
overlay assay, erythrocyte staining and liposome
binding. Despite the sphingomyelin binding, the
mutant lysenin was not able to assemble channels
and lacked hemolytic activity. These data point to
tryptophan 20 residue located in the N-terminus of
lysenin-His as important to the lytic activity of the
recombinant protein, but not required for specific
sphingomyelin recognition by the protein. The
crucial role of the conserved tryptophan residues of
lysenin for both sphingomyelin binding and hemo-
lytic activity of the protein was already shown by
Kobayashi’s group [29]. Accordingly, intrinsic tryp-
tophan fluorescence of lysenin exhibited blue shift in
the presence of sphingomyelin in membranes, sug-
gesting that at these conditions the aromatic residues
are exposed to a less polar environment [15,16].
However, contrary to those data, in our hands
W20A lysenin-His retained the ability to bind
sphingomyelin with a comparable sensitivity as wt
lysenin-His, although it lost the hemolytic activity.
When trying to find a reason for this discrepancy,
one can consider that the recombinant W20A
lysenin used by Kobayashi and co-workers that had
lost the sphingomyelin-binding ability was fused at
the N-terminus to a maltose-binding protein tag
[29]. In comparison to only six histidine residues
fused to the N-terminus of lysenin in our construct,
maltose-binding protein adds more than 300 amino
Figure 6. Incorporation of wt lysenin-His into planar lipid bilayers containing sphingomyelin leads to channel formation. All measurements
were conducted in asymmetric ionic conditions (450/150 mM KCl, pH 7.0 cis/trans ). (A) Wt lysenin-His (1 mg/ml) induces single channel
activity recorded in a planar lipid bilayer composed of azolectin/sphingomyelin/cholesterol (96:4:1, by weight) at indicated holding
potentials. (B) Time-dependent changes of current induced by 1 mg/ml wt lysenin-His in the azolectin/sphingomyelin/cholesterol bilayer.
The measurements were performed continuously for 2 h at 50 mV. (C) Currents recorded in the azolectin/sphingomyelin/cholesterol bilayer
in response to a series of wt lysenin-His doses (1�6 mg/ml) within 2 h at the holding potential of 50 mV. (D) Lack of channel activity of wt
lysenin-His (2�6 mg/ml) in a planar membrane devoid of sphingomyelin at 50 mV holding potential. (E) W20A lysenin-His (2�6 mg/ml)
does not form channels in bilayers composed of azolectin/sphingomyelin/cholesterol at 50 mV. The proteins were added to the trans
chamber of experimental cuvettes.
130 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
acids to the 297 amino acid long lysenin. Such an
extended tag could have markedly affected the
structure and properties of the N-terminal fragment
of W20A lysenin. Hence, the significantly dimin-
ished binding of sphingomyelin by W20A lysenin
fused to maltose-binding protein could ensue from
the plausible constraints exerted on the mutant
lysenin by the huge tag. In agreement with this
assumption it was recently reported that deletion
mutants of lysenin lacking up to 160 N-terminal
amino acids, including tryptophan 20, bound sphin-
gomyelin without causing hemolysis [31]. Altogether
the data of Kobayashi’s group and the results
presented here confirm that the N-terminus of
lysenin including tryptophan 20 is of importance
for the cytotoxic activity of the protein, being
dispensable for sphingomyelin binding. On the basis
of our results we suggest that the N-terminus of the
protein is important for efficient lysenin oligomer-
ization and/or membrane penetration.
Oligomerization is a typical stage during pore and
channel formation by cytotoxins [32�34]. When
studied by SDS-PAGE, an oligomer of wt lysenin-
His of about 280 kDa was found (Figure 3B, 3C) in
line with earlier reports [15,31]. The shift from 41
kDa monomer to 280 kDa oligomer of wt lysenin-
His started at 40:1 sphingomyelin/lysenin threshold
and was enhanced by increasing sphingomyelin/
protein ratio and by cholesterol, although cholesterol
was not required for the binding of the protein to
sphingomyelin, as shown by SDS-PAGE and an
overlay assay. A similar oligomer of wt lysenin-His of
about 250 kDa, accompanied by 360 kDa one, was
found after chemical cross-linking of the protein
bound to liposomes. However, during studies em-
ploying blue native gel electrophoresis, a predomi-
nant 120 kDa oligomer of wt lysenin-His was
detected. These data suggest that trimer of 120
kDa can be a functional unit of wt lysenin-His
responsible for lytic activity of the protein. Accord-
ingly, lytic-inactive W20A lysenin-His failed to
assembly the 120 kDa trimer forming instead
multi-oligomers of about 620 kDa and 800 kDa.
Further studies are required to determine whether
haxamer (250�280 kDa) or nanomer (360 kDa) of
wt lysenin-His detected after chemical cross-linking
and SDS-PAGE analysis represent higher form of
organization of the basic trimeric unit of the protein
or result from an action of the cross-linker as well as
SDS. The latter suggestion is in agreement with
reports that SDS led to non-specific oligomerization
of the pore-forming Pseudomonas aeruginosa cyto-
toxin [35].
To explain the lytic/channel activity of lysenin
three mechanisms can be considered. The presence
of positively charged amino acids spread along the
polypeptide chain of lysenin suggests that the protein
can cover the surface of negatively charged mem-
branes in a carpet-like manner, leading to membrane
Figure 7. Wild type lysenin-His forms cation-selective channels.
(A) Single channel activity induced by wt lysenin-His (1 mg/ml) in
an azolectin/sphingomyelin/cholesterol bilayer (96:4:1, by weight)
and recorded at indicated holding potentials. Lipid bilayers were
exposed to asymmetric ionic conditions (450/150 mM KCl, pH 7.0
cis/trans ). All recordings were low�pass filtered at 200 Hz. The
closed channel state is indicated by c . (B) Current/voltage relation-
ship of the wt lysenin-His-induced single channel currents from the
experiment shown in (A). The current/voltage relationship was
linear for holding potentials in the range from �/150 mV to 70 mV.
Oligomers of lysenin form channels 131

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
disintegration in an analogy to detergent action [36].
Accordingly, wt lysenin-His at 6 mg/ml no longer
formed channels but caused membrane rupture.
Fragmentation of liposomes by wt lysenin-His was
also detected in ultrastructural studies when high
doses of wt lysenin-His were used. These data
indicate that the detergent-like action of lysenin
can prevail at high concentrations of the protein
while channels are functioning at lower protein
content. This suggestion is in line with the sugges-
tion that formation of ion channels, transmembrane
pores and membrane damage can represent various
stages of lytic toxin action [37,38]. The channel
assembly requires formation of well-organized trans-
membrane pores. To this end, an analysis of the
secondary structure of lysenin has indicated that the
protein has no helices capable of forming transmem-
brane domain [39]. Nevertheless, lysenin shares
several properties with equinatoxin II, a sphingo-
myelin-binding toxin known to form cation-selective
channels of a toroidal pore structure. It was found
that for equinatoxin II to bind to sphingomyelin-
containing membranes, clusters of aromatic residues
of the protein, including two tryptophan residues,
are required [40]. After binding to the membrane,
the protein aggregates. Upon oligomerization, a
short N-terminal fragment of equinatoxin II inserts
into the membrane which is concomitant with the
formation of a toroidal lipid pore in the bilayer
[41,42]. On the basis of ultrastructural studies,
formation of toroidal pores by equinatoxin II was
connected with reorganization of lipids into hexago-
nal phase HII [41]. We did not detect corresponding
lipid structures in liposomes treated with wt lysenin-
His (Figure 5 and data not shown). This, however,
does not preclude insertion of wt lysenin-His into
the membrane to form toroidal pores since melittin,
another toxin forming such pores, also did not favor
HII lipid phase formation and cleaved the liposomes
into smaller vesicles [41], resembling action of wt
lysenin-His. The lack of pore-forming capacity of
W20A lysenin-His can ensue from its incorrect
oligomerization and possible failure in membrane
lipid distortion. Recent studies on a-hemolysin
demonstrated that insertion of the toxin into the
membrane and membrane lysis can be two un-
coupled phenomena [43]. In an analogy, one can
consider that W20A lysenin-His can also penetrate
the membrane but fails to change the lipid archi-
tecture to induce membrane permeation.
As demonstrated in this paper, lysenin forms
cation-selective ion channels that can be detected
by electrophysiological means. Their single channel
conductance is 1009/9 pS. The cation selectivity of
the lysenin channels is probably a result of the
presence of clusters of negatively charged amino
acids promoting passage of cations rather than
anions. Further studies are needed to establish
which parts of the protein may contribute to this
interesting feature. The formation of cation-selective
channels by lysenin can account for the observed
long-lasting contraction of strips of aorta exposed to
the toxin [21]. Presumably, this effect was related to
the influx of calcium ions through the lysenin-
formed channels. Formation of channels by lysenin
in the plasma membrane of erythrocytes and other
cells can lead to ion imbalance followed by osmotic
lysis of the cells. It is worthy of note that several
other cytolytic toxins also form ion channels in
membranes, indicating that the channel activity of
the toxins is relevant to their ability to kill cells
[3,44�46]. It is a matter of further investigation how
lysenin oligomerization influences channel kinetic
properties, and its conductance and selectivity.
While the manuscript was completed, another
report analysing hemolytic and microbicidal activity
of recombinant lysenin was announced [47]. It was
shown that lysenin oligomerization occurred on
erythrocytes but not on bacterial membranes. Aim-
ing to examine the interaction of lysenin with
sphingomyelin-containing membranes, the authors
found that in lipid planar bilayers lysenin formed ion
channels, in agreement with our data on wt lysenin-
His.
Acknowledgements
We thank Dr Andrzej Kubalski for critical reading of
the manuscript and valuable discussion, Dr Mariusz
Wieckowski for the introduction to the blue native
gel electrophoresis and Kazimiera Mrozinska for
excellent technical assistance. This work was sup-
ported by Grant no. 2PO4C 141 29 from the Polish
Ministry of Education and Science and by Innova-
tion Grant from the Nencki Institute of Experimen-
tal Biology.
References
[1] Lange Y, Swaisgood MH, Ramos BV, Steck TL 1989.
Plasma membranes contain half the phospholipid and 90%
of the cholesterol and sphingomyelin in cultured human
fibroblasts. J Biol Chem;264:3786�3793.
[2] Koval M, Pagano RE 1991. Intracellular transport and
metabolism of sphingomyelin. Biochim Biophys Acta;1082:
113�125.
[3] Zorec R, Tester M, Macek P, Mason WT 1990. Cytotoxicity
of equinatoxin II from the sea anemone Actinia equina
involves ion channel formation and an increase in intracel-
lular calcium activity. J Membr Biol;118:243�249.
[4] Bonev BB, Lam YH, Anderluh G, Watts A, Norton RS,
Separovic F 2003. Effects of the eukaryotic pore-forming
cytolysin equinatoxin II on lipid membranes and the role of
sphingomyelin. Biophys J;84:2382�2392.
132 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
[5] Geisse NA, Cover TL, Henderson RM, Edwardson JM
2004. Targeting of Helicobacter pylori vacuolating toxin to
lipid raft membrane domains analyzed by atomic force
microscopy. Biochem J;381:911�917.
[6] Sepcic K, Berne S, Rebolj K, Batista U, Plemenitas A,
Sentjurc M, Macek P 2004. Ostreolysin, a pore-forming
protein from the oyster mushroom, interacts specifically with
membrane cholesterol-rich lipid domains. FEBS Lett;575:
81�85.
[7] Tomita T, Noguchi K, Mimuro H, Ukaji F, Ito K, Sugawara-
Tomita N, Hashimoto Y 2004. Ostreolysin, a pore-forming
protein from the oyster mushroom, interacts specifically with
membrane cholesterol-rich lipid domains. J Biol
Chem;279:26975�26982.
[8] Zitzer A, Zitzer O, Bhakdi S, Palmer M 1999. Oligomeriza-
tion of Vibrio cholerae cytolysin yields a pentameric pore and
has a dual specificity for cholesterol and sphingolipids in the
target membrane. J Biol Chem;274:1375�1380.
[9] Brown DA, London E 1998. Functions of lipid rafts in
biological membranes. Annu Rev Cell Dev Biol;14:111�136.
[10] Horejsi V 2003. The roles of membrane microdomains
(rafts) in T cell activation. Immunol Rev;191:148�164.
[11] Rajendran L, Simons K 2005. Lipid rafts and membrane
dynamics. J Cell Sci;118:1099�1102.
[12] Waheed AA, Shimada Y, Heijnen H F, Nakamura M,
Inomata M, Hayashi M, Iwashita S, Slot JW, Ohno-Iwashita
Y 2001. Selective binding of perfringolysin O derivative to
cholesterol-rich membrane microdomains (rafts). Proc Natl
Acad Sci USA;98:4926�4931.
[13] Cooper EL, Kauschke E, Cossarizza A 2002. Digging for
innate immunity since Darwin and Metchnikoff. BioEssays;
24:319�333.
[14] Yamaji A, Sekizawa Y, Emoto K, Sakuraba H, Inoue K,
Kobayashi H, Umeda M 1998. Lysenin, a novel sphingo-
myelin-specific binding protein. J Biol Chem;73:5300�5306.
[15] Yamaji-Hasegawa A, Makino A, Baba T, Senoh Y, Kimura-
Suda H, Sato SB, Terada N, Ohno S, Kiyokawa E, Umeda
M, Kobayashi T 2003. Oligomerization and pore formation
of a sphingomyelin-specific toxin, lysenin. J Biol Chem;
278:22762�22770.
[16] Ishitsuka R, Yamaji-Hasegawa A, Makino A, Hirabayashi Y,
Kobayashi T 2004. A lipid-specific toxin reveals heteroge-
neity of sphingomyelin-containing membranes. Biophys J;
86:296�307.
[17] Abdel-Shakor AB, Kwiatkowska K, Sobota A 2004. Cell
surface ceramide generation precedes and controls FcgRII
clustering and phosphorylation in rafts. J Biol
Chem;279:36778�36787.
[18] Gouaux E 1997. Channel-forming toxins: tales of transfor-
mation. Curr Opin Struct Biol;7:566�573.
[19] Hanada K, Hara T, Fukasawa M, Yamaji A, Umeda M,
Nishijima M 1998. Mammalian cell mutants resistant to a
sphingomyelin-directed cytolysin. Genetic and biochemical
evidence for complex formation of the LCB1 protein with
the LCB2 protein for serine palmitoyltransferase. J Biol
Chem;273:33787�33794.
[20] Kobayashi H, Sekizawa Y, Aizu M, Umeda M 2000. Lethal
and non-lethal responses of spermatozoa from a wide variety
of vertebrates and invertebrates to lysenin, a protein from the
coelomic fluid of the earthworm Eisenia foetida . J Exp
Zool;286:538�549.
[21] Sekizawa Y, Hagiwara K, Nakajima T, Kobayashi H 1996. A
novel protein, lysenin, that causes contraction of the isolated
rat aorta: its purification from the coelomic fluid of the
earthworm, Eisenia foetida . Biomed Res;17:197�203.
[22] Sekizawa Y, Kubo T, Kobayashi H, Nakajima T, Natori S
1997. Molecular cloning of cDNA for lysenin, a novel
protein in the earthworm Eisenia foetida that causes con-
traction of rat vascular smooth muscle. Gene;191:97�102.
[23] Maniatis T, Fritsch EF, Sambrook J 1982. Preparation of
reagents and buffers used in molecular cloning. In: Mole-
cular cloning: a laboratory manual. Cold Spring Harbor,
NY: Cold Spring Harbor Laboratory Press. p. B27.
[24] Dybczynski I, Pl ucienniczak A 1988. A protocol for DNA
fragment extraction from polyacrylamide gels. Biotech-
niques;6:924�926.
[25] Thomas C L, Steel J, Prestwich GD, Schiavo G 1999.
Generation of phosphatidylinositol-specific antibodies and
their characterization. Biochem Soc Trans;4:648�652.
[26] Taki T, Ishikawa D 1997. TLC blotting: application to
microscale analysis of lipids and as a new approach to lipid-
protein interaction. Anal Biochem;251:135�143.
[27] Kwiatkowska K, Frey J, Sobota A 2003. Phosphorylation of
FcgRIIA is required for the receptor-induced actin rearran-
gement and capping: the role of membrane rafts. J Cell
Sci;116:537�550.
[28] Hordejuk R, Lobanov NA, Kicinska A, Szewczyk A, Dol owy
K 2004. pH modulation of large conductance potassium
channel from adrenal chromaffin granules Mol Membr
Biol;21:307�313.
[29] Kiyokawa E, Makino A, Ishii K, Otsuka N, Yamaji-Hase-
gawa A, Kobayashi T 2004. Recognition of sphingomyelin
by lysenin and lysenin-related proteins. Biochemistry;43:
9766�9773.
[30] Barenholz Y, Gatt S. Sphingomyelin: metabolism, chemical
synthesis, chemical and physical properties. In: Hawthorne
JN Ansell GB, editors. Phospholipids. Amsterdam: Elsevier/
North-Holland Biomedical Press; 1982. pp 129�177.
[31] Kiyokawa E, Baba T, Otsuka N, Makino A, Ohno S,
Kobayashi T 2005. Spatial and functional heterogeneity of
sphingolipid-rich membrane domains. J Biol Chem;280:
24072�24084.
[32] Valeva A, Weisser A, Walker B, Kehoe M, Bayley H, Bhakdi
S, Palmer M 1996. Molecular architecture of a toxin pore: a
15-residue sequence lines the transmembrane channel of
staphylococcal alpha-toxin. EMBO J;15:1857�1864.
[33] Hotze EM, Heuck AP, Czajkowsky DM, Shao Z, Johnson
AE, Tweten RK 2002. Monomer-monomer interactions
drive the prepore to pore conversion of a b-barrel-forming
cholesterol-dependent cytolysin. J Biol Chem;277:11597�11605.
[34] Montoya M, Gouaux E 2003. b-Barrel membrane protein
folding and structure viewed through the lens of a-hemoly-
sin. Biochim Biophys Acta;1609:19�27.
[35] Sliwinska-Korell A, Engelhardt H, Kampka M, Lutz F 1999.
Oligomerization and structural changes of the pore-forming
Pseudomonas aeruginosa cytotoxin. Eur J Biochem;265:
221�230.
[36] Shai Y, Oren Z 2001. From ‘‘carpet’’ mechanism to de-novo
designed diastereomeric cell-selective antimicrobial pep-
tides. Peptides;22:629�1641.
[37] Brogden KA 2005. Antimicrobial peptides: pore formers or
metabolic inhibitors in bacteria? Nat Rev Microbiol;
3:238�250.
[38] Dathe M, Wieprecht T 1999. Structural features of helical
antimicrobial peptides: their potential to modulate activity
on model membranes and biological cells. Biochim Biophys
Acta;1462:71�87.
Oligomers of lysenin form channels 133

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
[39] Abel-Shakor AB, Czuryl o EA, Sobota A 2003. Lysenin, a
unique sphingomyelin-binding protein. FEBS Lett;542:1�6.
[40] Hong Q, Gutierrez-Aguirre I, Barlic A, Malovrh P, Kristan
K, Podlesek Z, Macek P, Turk D, Gonzalez-Manas JM,
Lakey JH, Anderluh G 2002. Two-step membrane binding
by equinatoxin II, a pore-forming toxin from the sea
anemone, involves an exposed aromatic cluster and a flexible
helix. J Biol Chem;277:41916�41924.
[41] Anderluh G, Dalla Serra M, Viero G, Guella G, Macek P,
Menestrina G 2003. Pore formation by equinatoxin II, a
eukaryotic protein toxin, occurs by induction of nonlamellar
lipid structures. J Biol Chem;278:45216�45223.
[42] Kristan K, Podlesek Z, Hojnik V, Gutierrez-Aguirre I,
Guncar G, Turk D, Gonzalez-Manas JM, Lakey JH, Macek
P, Anderluh G 2004. Pore formation by equinatoxin, a
eukaryotic pore-forming toxin, requires a flexible N-terminal
region and a stable b-sandwich. J Biol Chem;279:46509�46517.
[43] Sanchez-Magraner L, Cortajarena AL, Goni FM, Ostolaza
H 2006. Membrane insertion of Escherichia coli a-hemolysin
is independent from membrane lysis. J Biol Chem;281:
5461�5467.
[44] Kagan BL, Selsted ME, Ganz T, Lehrer RI 1990. Anti-
microbial defensin peptides form voltage-dependent ion-
permeable channels in planar lipid bilayer membranes. Proc
Natl Acad Sci USA;87:210�214.
[45] Falla TJ, Karunaratne DN, Hancock RE 1996. Mode of
action of the antimicrobial peptide indolicidin. J Biol
Chem;271:19298�19303.
[46] Laohachai KN, Bahadi R, Hardo MB, Hardo PG, Kourie JI
2003. The role of bacterial and non-bacterial toxins in the
induction of changes in membrane transport: implications
for diarrhea. Toxiconology;42:687�707.
[47] Bruhn H, Winkelmann J, Anderson C, Andra J, Leippe M
2006. Dissection of the mechanism of cytolytic and anti-
bacterial activity of lysenin, a defense protein of the annelid
Eisenia fetida. Dev Comp Immunol;30:597�606.
This paper was first published online on iFirst on 23 December 2006.
134 K. Kwiatkowska et al.

Dow
nloa
ded
By: [
Inst
ytut
Bio
logi
i Dos
wia
dcza
ln] A
t: 10
:49
20 A
pril
2007
Supplementary Figure 1. Nucleotide sequence of synthetic lysenin gene. The coding strand synthetic oligonucleotides are in capital and
lower-case letters, alternately, and are marked by symbols ‘‘Ux’’ on the right margin of the same row; oligonucleotides joining the upper
ones during ligation are written in bold capital letters and marked by symbols ‘‘Jx_x�/1’’ on the right margin. The amino acid sequence of
lysenin is given above the coding upper strand. Start and stop codons are in bold italics. The nucleotide sequence of oligonucleotide P2
which allowed amplification of the final PCR product is double underlined. Tryptophan 20 that was substituted with alanine in W20A
lysenin-His is indicated by black background.
Oligomers of lysenin form channels 135
Recommended

















