MOLECULAR BIOLOGY
Dicer uses distinct modules forrecognizing dsRNA terminiNiladri K. Sinha, Janet Iwasa, Peter S. Shen,* Brenda L. Bass*
Invertebrates rely on Dicer to cleave viral double-stranded RNA (dsRNA), and DrosophilaDicer-2 distinguishes dsRNA substrates by their termini. Blunt termini promote processivecleavage, while 3′ overhanging termini are cleaved distributively. To understand thisdiscrimination, we used cryo–electron microscopy to solve structures of Drosophila Dicer-2alone and in complex with blunt dsRNA. Whereas the Platform-PAZ domains have beenconsidered the only Dicer domains that bind dsRNA termini, unexpectedly, we found thatthe helicase domain is required for binding blunt, but not 3′ overhanging, termini. We furthershowed that blunt dsRNA is locally unwound and threaded through the helicase domain inan adenosine triphosphate–dependent manner. Our studies reveal a previously unrecognizedmechanism for optimizing antiviral defense and set the stage for the discovery of helicase-dependent functions in other Dicers.
Dicer ribonucleases cleave double-strandedRNA (dsRNA) precursors to generatemicro-RNAs (miRNAs) and small interferingRNAs(siRNAs) (1, 2). In concert with Argonautes,these small RNAs bind complementary
mRNAs todown-regulate their expression.miRNAsare processed by Dicer from small hairpins, where-as siRNAs are typically processed from longerdsRNA, from endogenous sources (3), or fromexogenous sources such as viral replication in-termediates (4–6). Someorganisms, such asHomosapiens and Caenorhabditis elegans, encode oneDicer that generates miRNAs and siRNAs, butother organisms have multiple Dicers with spe-cialized functions.Dicers exist throughout eukaryotes, and a sub-
set have an N-terminal helicase domain of theRIG-I-like receptor (RLR) subgroup (7) (Fig. 1Aand fig. S1A). RLRs often function in innate im-munity (8), and Dicer helicase domains some-times show differences in activity that correlatewith roles in immunity. For example,Drosophilamelanogaster expresses two Dicers, one special-ized for processing miRNAs (dmDcr-1) and asecond for siRNAs (dmDcr-2) (9). dmDcr-1 hasa degenerate helicase domain and is an adeno-sine triphosphate (ATP)–independent enzyme (10),whereas dmDcr-2, with dedicated antiviral roles(11–13), has a conserved helicase domain thathydrolyzes ATP (14–17). Under certain conditions,Homo sapiens Dicer-1 (hsDcr-1) also generatesviral siRNAs (18, 19). However, despite conser-vation of its helicase domain, hsDcr-1 does nothydrolyze ATP in vitro (20), and its helicase do-main is not implicated in viral siRNA biogenesisin vivo (19). Differences in activities of the heli-case domain of vertebrate and invertebrate Dicersmay reflect distinct roles in antiviral defense.dmDcr-2 activity depends on termini of its
dsRNA substrates (15, 16). Blunt (BLT) termini
promote a processive reaction whereby mul-tiple siRNAs are produced before dmDcr-2 dis-sociates, and this reaction requires a functionalhelicase domain and ATP (15, 17). In contrast,dsRNAs with 3′ overhanging (3′ovr) termini pro-mote an ATP-independent, distributive cleav-age, whereby dmDcr-2 dissociates after eachcleavage. hsDcr-1 does not require ATP for pro-cessing BLT or 3′ovr termini (fig. S1B, C and D,lanes 5, 7, 10, and 12), suggesting that, at leastin vitro, cleavage of BLT dsRNA is not dependenton its helicase domain.To understand themechanism of terminus dis-
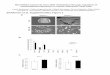
crimination by dmDcr-2, we used cryo–electronmicroscopy (cryo-EM) to determine structures ofdmDcr-2 alone and in complexwith a BLT52basepair (bp) dsRNA (52 dsRNA) and ATP-gS (Fig. 1,figs. S2 to S10, and table S1). We used full-lengthdmDcr-2 with a point mutation in each ribonu-clease III (RNase III) domain to preclude dsRNAcleavage (dmDcr-2RIII) (Fig. 1A). ATP hydrolysisis required for processive cleavage of BLT dsRNA,and dmDcr-2 cannot hydrolyze ATP-gS, whichstabilizes a helicase-dependent conformation ofdmDcr-2 (16).The structure of apo-dmDcr-2RIII (Fig. 1, B and
C, and figs. S2 to S5) reiterated the “L shape” oflower resolution (~15 to 30 Å) EM reconstruc-tions of hsDcr-1 (21–23). Our 7.1-Å EM densitymap (Fig. 1B and fig. S2C) enabled fitting (fig. S4,B to F) and homology modeling (Fig. 1B) ofPlatform-PAZ domains at the cap and tandemRNase III domains in the core. An additionalround of three-dimensional (3D) classification(fig. S3) revealed an 8.7-Å map (fig. S2C), allow-ing assignment of the helicase domain at thebase (Fig. 1C and fig. S4A). Fitting of related apo-helicases into the EM density is consistent withthe helicase domain adopting an open confor-mation (fig. S5, A to D).The 2D class averages of the dmDcr-2 complex
revealed protein with well-resolved secondarystructure features bound to the BLT dsRNA ter-minus (fig. S6, B and C). Some protein densitywas missing, and because control experiments
indicated that protein on the grid was intact (fig.S7A), this was likely due to inherent flexibility(fig. S7B). Measurements of the dsRNA, guided bymajor grooves, showed that visible protein foot-printed ~8 to 9 bps (fig. S6B). The crystal struc-ture of RIG-I’s helicase domain bound to dsRNAhas a similar footprint (24, 25), suggesting thatdmDcr-2’s helicase domain bound to BLT dsRNAtermini. Indeed, our 6.8-Å reconstruction of thecomplex (figs. S6E and S8) resembled RIG-I in aclosed conformation (Fig. 1D and fig. S9). TheHel1 and Hel2 subdomains of RIG-I’s helicase,along with the pincer helices (fig. S1A), could befit as a single rigid body (fig. S9A). The reconstruc-tion also revealed ahelical bundle characteristic ofthe Hel2i subdomain that could be fitted sepa-rately as a rigid body (fig. S9B). These fittingsenabled a homologymodel of dmDcr-2’s helicasedomain bound to BLT dsRNA (Fig. 1D and fig.S9, D and E).Ourmodels of dmDcr-2’s helicase in open (apo)
and closed (substrate-bound) conformations im-plied clamping of the helicase on BLT dsRNAtermini (Fig. 1E and movies S1 and S2). In theopen conformation, Hel2 and Hel2i extend awayfrom Hel1, creating a C-shaped opening for sub-strate engagement (Fig. 1C). In the BLT dsRNA-bound state, Hel2 and Hel2i swivel toward Hel1to clamp on the terminus (Fig. 1, D and E, andmovies S1 and S2).Within the helicase domain, density was ob-
served for only one RNA strand, indicative oflocal unwinding (Fig. 1F and fig. S9C). Unwindingwould likely require ATP hydrolysis and possiblywas enabled by contaminating ATP in commer-cial preparations of ATP-gS (16). Using dsRNAswith a nick in sense or antisense strands (fig.S11A), we performed in vitro unwinding assays(fig. S11B). With ATP, dmDcr-2RIII, but not theadenosine triphosphatase (ATPase)–defectiveWalker A mutant dmDcr-2RIII,K34A (16), unwoundBLT dsRNA termini (fig. S11B; compare lanes 3and 7, top and bottom panels).The unwound single strand maintained an
A-form conformation (Fig. 1F), likely tominimizeentropic costs of reannealing before cleavagein RNase III sites. Whether the RIG-I helicaseunwinds dsRNA is controversial (26, 27), butrelated helicases exhibit unwinding activity (28).Local unwinding may facilitate dmDcr-2’s heli-case domain in binding and translocating alongdsRNA.The Platform-PAZ domains have been consi-
dered the only Dicer domains that bind dsRNAtermini (29–32), but our structures suggested thatthe helicase domain also binds termini. To inves-tigate this idea, we purified dmDcr-2DHel,RIII
(Fig. 2A),which lacked thehelicasedomain (Fig. 1A).Consistent with previous studies (16), in gel-shiftassays full-length dmDcr-2RIII bound both BLTand 3′ovr dsRNA (Fig. 2B, dsRNA design; andFig. 2C, top); ATP increased affinity for BLTdsRNA and decreased affinity for 3′ovr dsRNA(Fig. 2, C and D and table S2). However, whereasdmDcr-2DHel,RIII bound 3′ovr dsRNA with an af-finity similar to dmDcr-2RIII, its binding to BLTdsRNA was not detected (Fig. 2C, bottom panel,
RESEARCH
Sinha et al., Science 359, 329–334 (2018) 19 January 2018 1 of 6
Department of Biochemistry, University of Utah, Salt LakeCity, UT 84112, USA.*Corresponding author. Email: [email protected](B.L.B.); [email protected] (P.S.S.)
on October 18, 2020
http://science.sciencem
ag.org/D
ownloaded from

Fig. 2D, and table S2). The inability of dmDcr-2DHel,RIII to bind BLT dsRNAwas not due to theabsence of ATP hydrolysis because dmDcr-2RIII,K34A bound BLT dsRNA efficiently (fig. S12,A and B, and table S2). Thus, the helicase domainis required for binding BLT, but not 3′ovr, dsRNA.Single-turnover cleavage assays showed that
neither dmDcr-2WT nor dmDcr-2DHel (fig. S12C)cleaved BLT dsRNAwithout ATP (Fig. 2, E and F,lanes 5 and 6). With ATP, cleavage of BLT dsRNAby dmDcr-2WT gave heterogeneous cleavage pro-ducts characteristic ofDicer enzymeswithATPaseactivity (16, 33), but strikingly, dmDcr-2DHel wasincapable of cleaving BLT dsRNA (Fig. 2, E and F;compare lanes 9 and 10). As expected (15, 16),cleavage of 3′ovr dsRNAwas independent of ATPand with both dmDcr-2WT and dmDcr-2DHel pro-duced a single siRNA-sized 22-nucleotide (nt)band (Fig. 2E; compare lanes 7, 8, 11, and 12).dmDcr-2DHel cleaved 3′ovr dsRNAmore efficientlythan dmDcr-2WT (Fig. 2, E and F, compare lanes7, 8, 11, and 12), suggesting that the helicase do-main hinders cleavage of 3′ovr dsRNA. Thisobservation is reminiscent of autoinhibition byhsDcr-1’s helicase domain in processing 3′ovrdsRNAs (34).Our biochemical and structural studies in-
dicated that dmDcr-2 has twomodes of substraterecognitionand cleavage: onemediatedbyPlatform-PAZdomains and resulting in precise cleavage of 3′ovr dsRNAs into base-paired 22-nt siRNAs (22mersiRNAs) and a second mediated by the helicasedomain and resulting in heterogeneous cleavageof BLT dsRNAs. We searched for amino acidsthat might separately affect cleavage of a 3′ovror BLT dsRNA.We created one variant of dmDcr-2(Fig. 1A, PP) containing five point mutations inthe Platform and PAZ domains (35). Multiplecrystal structures show that a phenylalanine inthe C-terminal domain (CTD) of RIG-I recognizesBLT dsRNA by stacking on the terminal basepair (24). Dicer enzymes do not have a CTD,but for the second variant we searched for re-gions in dmDcr-2 with sequence similarity to theCTD.Within the region identified (fig. S13, A to C),we mutated a single phenylalanine to a glycine(dmDcr-2F225G).We compared activities of purified dmDcr-2PP
and dmDcr-2F225G (fig. S13D) to dmDcr-2WT usingsingle-turnover cleavage assays (Fig. 3, A and B).As expected, cleavage of BLT dsRNA was notobserved without ATP (Fig. 3, A and B, lanes 5to 7). However, with ATP, cleavage of BLT dsRNAby dmDcr-2WT or dmDcr-2PP appeared nearly iden-tical (Fig. 3, A and B, lanes 11 and 13), whereascleavage was completely disrupted by the heli-case point mutation in dmDcr-2F225G (Fig. 3, Aand B, lane 12). [At least part of this effect is dueto weakened BLT dsRNA binding (fig. S13, E andF, and table S2)]. By contrast, cleavage of 3′ovrdsRNA was independent of ATP and minimallyaffected by the F225G helicase mutation (Fig. 3,A and B, lanes 8, 9, 14, and 15). However, cleavagewas eliminated bymutations in the Platform-PAZdomains (Fig. 3, A and B, lanes 10 and 16). Thesedata reiterate that cleavage of 3′ovr dsRNA ismediated by Platform-PAZ domains, whereas the
helicase domain coordinates recognition and cleav-age of BLT dsRNA.While dmDcr-2PP cleaved BLT dsRNA to yield
a pattern nearly identical to dmDcr-2WT, levels of22 nt siRNA decreased (Fig. 3, A and B, lanes 11and 13). Because 22-nt siRNA was not observedwith dmDcr-2F225G (Fig. 3A, lane 12), we hypothe-sized that this species derived from dsRNA thatwas threaded through the helicase domain untilthe BLT terminus encountered the Platform-PAZdomains. To confirm that smaller products (<22 nt)did not result from degradation of 22-nt siRNA,we monitored cleavage of chimeric dsRNAs con-taining deoxynucleotides at positions 21 to 23from the 5′ terminus (Fig. 3C). Cleavage of 3′ovr
dsRNA was eliminated with chimeric molecules(Fig. 3, D and E; compare lanes 8, 9, 15, and 16),as expected for Platform-PAZ–mediated cleavage.However, for BLT dsRNA, while 22-nt siRNAwasabsent, all other fragments were visible (Fig. 3, Dand E; compare lanes 6 and 13), consistent with ahelicase-mediated threading mechanism.Studies of Dicer fromother organisms indicate
Platform-PAZ domains bind termini of 3′ovrdsRNA to allow measuring to RNase III activesites and production of an siRNA length (29, 30).Our cryo-EM structure of the dmDcr-2 complexand subsequent biochemical studies suggestedthat BLT dsRNA is cleaved differently, and in anATP-dependent manner, threaded through the
Sinha et al., Science 359, 329–334 (2018) 19 January 2018 2 of 6
Hel2
Hel1 Hel2iPincer
90°
Hel1 Hel2
Hel2i
BLTdsRNA
90°
Hel1 Hel2Hel2i
BLTdsRNA
5’3’
~ 90 Å ~ 75 Å
~ 50
Å~
90 Å
dmDcr-2
dsRBMRNase IIIa RNase IIIbHel2Hel2i DUF283 platform PAZ rulerHel1
976
16531722
219 231 380
381
508 571 6737 729 855
856 977
1194 1391 1448 1649
D1217A D1476AF225G H743AR752A R759A
R943A R956A
PP
RIIIK34A
548
pincer
CapC
ore
Base (Helicase)
PAZ Platform
RIIIaRIIIa
120°
RIIIb
Hel1
Hel2
Hel2i
90°
5’3’
5’
3’
5’
3’5’
dsR
NA
ssR
NA
~ 14
0 Å
Fig. 1. Cryo-EM reconstructions of apo-dmDcr-2 and dmDcr-2•BLTdsRNA•ATP-gS. (A)dmDcr-2 domains numbered at boundaries. Mutations/deletions are in bold and are designatedin text as superscripts. All mutations were in the context of the full-length protein, unlessspecified by D. Two full-length variants had multiple mutations: PP (H743A, R752A, R759A, R943A,and R956A) and RIII (D1217A and D1476A). (B) Cryo-EM density map of apo-dmDcr-2 (7.1 Å) fittedwith homology models of subdomains. RIIIa, RNase IIIa; RIIIb, RNase IIIb. (C) Homology modelof apo-dmDcr-2 in open conformation, based on apo-RIG-I, and fitted as a rigid body into an8.7-Å cryo-EM density map (see also figs. S4A and S5, B to D). (D) Cryo-EM reconstruction ofdmDcr-2RIII•BLT dsRNA•ATP-gS showing helicase in closed, ligand-bound conformation.(E) Superimposition of open (light) and closed (dark) helicase conformations showing clampingof Hel2 and Hel2i on BLT dsRNA. Arrow, direction of clamping. (F) EM density and modeling of BLTsingle- and double-stranded RNA.
RESEARCH | REPORTon O
ctober 18, 2020
http://science.sciencemag.org/
Dow
nloaded from

helicase domain to encounter the RNase IIIactive sites. We tested the threading model bydesigning dsRNAwith blocks at specific positions(Fig. 4A). Measurements using our EM densitymaps predicted that BLT dsRNAs are threadedthrough the helicase domain ~20 bp before en-countering RNase III domains. To trap threadingintermediates, we put biotin-dT analogs on bothstrands of 52 BLT dsRNA, at positions 28 or 37,counting from the 5′ end of the sense strand (Fig.4A); there was no significant difference in cleav-age of these modified dsRNAs (figs. S14, A andB). However, we hypothesized that the additionof streptavidin to biotin-dT–substituted dsRNAs
(Block dsRNAs) would arrest threading of dsRNAsthrough the helicase. When dsRNA was incubatedwith streptavidin before initiating cleavagewithdmDcr-2WT and ATP, we trapped early (<11 nt,Block-28) and intermediate (11 to 20 nt, Block-37)threading products without observing 22 ntsiRNAs (Fig. 4, B and C; see fig. S14C for sche-matic). By contrast, cleavage by hsDcr-1 was un-affected by blocks, indicating that, at least underthese conditions, hsDcr-1 cannot thread dsRNAthrough its helicase domain (Fig. 4D; comparelanes 12 to 14 with lanes 15 to 17).We anticipated that short threading intermedi-
ates (<22 nts) might be unique to the initial
cleavage event. However, threading intermediateswere observed with dmDcr-2WT under multiple-turnover conditions using internally 32P-labeleddsRNAs, increasing proportionally with 22mersiRNAs through the reaction time course (fig.S14, D and E). Thus, at least in vitro, threadingintermediates are recurring by-products of pro-cessive cleavage andnot specific to the initial cleav-age. dmDcr-2’s highly efficient, helicase-dependent,processive cleavage is likely advantageous in anti-viral defense. The generation of heterogeneouscleavage products during processive cleavage ispredicted to dampen the phasing signal of viralsiRNAs and is consistent with the overlapping
Sinha et al., Science 359, 329–334 (2018) 19 January 2018 3 of 6
Fig. 2. dmDcr-2’s helicase domain isrequired to recognize and cleaveBLTdsRNA. (A) Gel-filtration andSDS–polyacrylamide gel electrophore-sis analyses of dmDcr-2DHel,RIII.(B) Cartoon of dsRNA used in (C) to (F),showing modifications that blockbinding at one end. (C) Gel mobility shiftassays of dmDcr-2RIII (top) anddmDcr-2DHel,RIII (bottom) with52 BLT or 3′ovr dsRNA, ±5 mM ATP(n ≥ 3). (D) Binding curves usingdata as in (C). Data points, mean ± SD(n = 3). (E) Single-turnover cleavageassays of 52 BLT or 3′ovr dsRNA(1 nM) with dmDcr-2WTor dmDcr-2DHel
(30 nM), ± 5 mM ATP (n = 3).Only initial cleavage is monitored,because this removes 5′ 32P. Arrow,22-nt siRNA product. AH, alkalinehydrolysis. Left, nt lengths.(F) Quantification of cleavageassays as in (E). Percent dsRNAcleaved (all dsRNA except uncleaved)and percent siRNAs (21 to 23 ntproducts) resulting from firstcleavage were quantified. Datapoints, mean ± SD (n = 3). *P < 0.05;**P < 0.01; ***P < 0.001.
dmDcr-2ΔHel,RIII
MWobs : 160 kDaMWcalc : 135 kDaMWobs / MWcalc : 1.2
RIII, BLT, - ATPRIII, 3’ovr, - ATP
RIII, 3’ovr, + ATPRIII, BLT, + ATP
ΔHel,RIII, BLT, - ATPΔHel,RIII, 3’ovr, - ATP
ΔHel,RIII, 3’ovr, + ATPΔHel,RIII, BLT, + ATP
3.2 12.5 50 400 3.2 12.5 50 400 3.2 12.5 50 400 3.2 12.5 50 4000 0
dmD
cr-2
RIII
nM
– ATP + ATP
3.2 12.5 50 400 3.2 12.5 50 400 3.2 12.5 50 400 3.2 12.5 50 4000 0 nM
Free
ds
RN
AC
ompl
exFr
ee
dsR
NA
Com
plex
52 BLTdsRNA
52 3’ovrdsRNA
52 BLTdsRNA
52 3’ovrdsRNA
52 B
LT52
3’o
vr
dmD
cr-2
ΔHel
,RIII
100
6050
30
20
10
40
80
1 2 3 4 5 6 7 8 9 10 11 12Lanes
52 B
LT d
sRN
A
52 3
’ovr
dsR
NA + ATP– ATP
WT
ΔHel
WT
ΔHel
WT
ΔHel
WT
ΔHel
52 BLT dsRNA
52 3’ovrdsRNA
52 BLTdsRNA
52 3’ovrdsRNA
0
20
40
60
80
0
20
40
60
80
+ ATP– ATP52 BLT dsRNA
52 BLTdsRNA
52 3’ovr dsRNA
52 3’ovrdsRNA
52 B
LT d
sRN
A
52 3
’ovr
dsR
NA
52 B
LT d
sRN
A
52 3
’ovr
dsR
NA
+ ATP– ATP52 BLT dsRNA
52 BLT dsRNA
52 3’ovr dsRNA
52 3’ovr dsRNA
3 4 5 6 7 8 9 10 11 12Lanes
***
* *****
*
*
***
***
Unc
leav
edC
leave
d
52 3’ovr dsRNA
AH Dec
ade
dmDcr-2
dmDcr-2WT dmDcr-2ΔHel
dsR
NA
cle
aved
(%
) si
RN
As
from
1st
cle
avag
e (%
)
32Psense strand
antisense strand
deoxynucleotides
biotin
sense strand
antisense strand
deoxynucleotides
biotinblocked end
3’
5’
32P
52 BLT dsRNAopen end
3’ 5’
3’
blocked endopen end
3’
250150100
50
25
RESEARCH | REPORTon O
ctober 18, 2020
http://science.sciencemag.org/
Dow
nloaded from

and discontinuous viral siRNAs observed in in-vertebrate cells (6, 13, 36).The dsRNAbinding protein (dsRBP) Loquacious-
PD (Loqs-PD) allows dmDcr-2 to cleave inde-pendent of termini (16, 37) and is required forprocessing endogenous siRNAs (38), but not foran antiviral response (13). This suggests thatdmDcr-2’s intrinsic termini preferences functionin viral defense, whereas Loqs-PD allows process-ing of endogenous dsRNA with diverse termini.
By monitoring cleavage of dsRNAs with 5′ovrtermini, or overhangs on both strands (fig. S15),we determined that dsRNA with an accessible3′ terminus is preferentially recognized by thePlatform-PAZ domain and, without this feature,is processed by threading through the helicasedomain.RIG-I distinguishes capped termini of cellular
transcripts from tri- and diphosphorylated ter-mini of viral transcripts, and this is inferred to
allow self versus nonself discrimination (8). Wefound that, like RIG-I, dmDcr-2 cannot efficientlyprocess dsRNAs capped at the 5′ terminus, al-though the phosphorylation state does not affectcleavage (fig. S16, A and B, and table S3). Theseresults may reflect dmDcr-2’s ability to processprecursors of both endogenous and viral siRNAs.We show that dmDcr-2 has twomodes of cleav-
age (Fig. 4E and movies S3 and S4). dmDcr-2 iscapable of using its Platform-PAZ domain to
Sinha et al., Science 359, 329–334 (2018) 19 January 2018 4 of 6
Fig. 3. Helicase and Platform-PAZdomains differentially contributeto cleavage of BLT and 3′ovrdsRNA. (A) Single-turnovercleavage assays of 52 BLT or 3′ovrdsRNA (1 nM) with dmDcr-2WT,dmDcr-2F225G, and dmDcr-2PP
(30 nM), ±5 mM ATP (n = 3).Substrates were as described inFig. 2, B and E. AH, arrow, as inFig. 2E. (B) Quantification ofcleavage assays, as in (A). Datapoints, mean ± SD (n = 3). *P <0.05; **P < 0.01; ***P < 0.001;****P < 0.0001; P > 0.05, n.s.(nonsignificant). (C) Substratesused in (D) and (E), with additionaldetails in Fig. 2, B and E. In chime-ras, nts 21 to 23 from the 5′ endof the sense strand, and nts 32 to34 from the 5′ end of the antisensestrand were deoxyribonucleotides(dashed circles). (D) Single-turnovercleavage assays of regular orchimeric 52 BLT or 3′ovr dsRNA(1 nM) with dmDcr-2WT (30 nM),±5 mM ATP (n = 4). (E) Quantifica-tion of cleavage assays as in (D).Data points, mean ± SD (n = 4).
3 4 5 6 7 8 9 10111213 151614
+ ATP– ATP52 BLTdsRNA
52 BLTdsRNA
52 3’ovrdsRNA
52 3’ovr dsRNA
52 B
LT d
sRN
A52
3’o
vr d
sRN
A
WT
+ ATP– ATP52 BLTdsRNA
52 BLTdsRNA
52 3’ovr dsRNA
52 3’ovrdsRNA
52 B
LT d
sRN
A52
3’o
vr d
sRN
A
n.s.n.s.
n.s.n.s.
****
******
****n.s.
******
***
***
0
20
40
60
80
0
20
40
60
80
siR
NA
s fr
om1s
t cle
avag
e (%
)
Lanes
52 BLT dsRNA 52 3’ovr dsRNA 52 BLT chimeradsRNA
52 3’ovr chimeradsRNA
52 BLTchimeradsRNA
100
6050
30
20
10
40
52 BLTdsRNA
++
++ ++ –
–––−–
52 3’ovrdsRNA
dmDcr-2WT
ATP
1 2 3 4 5 6 7 8 9 10 11 12Lanes 13 14 15 16
*
Unc
leav
edU
Cle
ave
d
++
++ ++ –
–––−–
52 3’ovrchimeradsRNA
0
20
40
60
80
n.s.
n.s.
***
n.s.
******
**
0
20
40
60
80
*** *
n.s. n.s.
*****
**
++
++ ++ –
–––−–
52 BLTdsRNA
52 3’ovrdsRNA
++
++ ++ –
–––−–
dmDcr-2WT
ATP
52 BLTchimeradsRNA
52 3’ovrchimeradsRNA
++
++ ++ –
–––−–
52 BLTdsRNA
52 3’ovrdsRNA
++
++ ++ –
–––−–
dmDcr-2WT
ATP52 BLTchimeradsRNA
52 3’ovrchimeradsRNA
100
6050
30
20
10
40
80
52 B
LT d
sRN
A52
3’o
vr d
sRN
A
WT
WT
F22
5G
PP
F22
5G
PP
52 BLT dsRNA
52 3’ovr dsRNA
+ ATP
1 2 3 4 5 6 7 8 9 10 11 12Lanes 13 15 16
AH
14
WT
WT
F22
5G
PP
F22
5G
PP
52 BLTdsRNA
52 3’ovr dsRNA
– ATP
Dec
ade
Unc
leav
edU
lC
leave
d
dmDcr-2
Decade
(AH)
F225G PP
dsR
NA
cle
aved
(%
)
(AH)
siR
NA
s fr
om1s
t cle
avag
e (%
)ds
RN
A c
leav
ed (
%)
32P 32P 32P 32P
dmDcr-2
deoxynucleotides deoxynucleotides
4 5Lanes 6 7 8 9 11 12 13 14 15 16
4 5Lanes 6 7 8 9 11 12 13 14 15 16
RESEARCH | REPORTon O
ctober 18, 2020
http://science.sciencemag.org/
Dow
nloaded from

recognize 3′ovr dsRNAs in vitro, but it is un-knownwhether dmDcr-2 processes such dsRNAsin vivo. dmDcr-2’s cognate dsRBP, R2D2, mayinhibit recognition and processing of substrateswith 3′ovr termini (17). As such, the Platform-PAZ domains of dmDcr-2 may function solely ondsRNAs that are threaded through the helicasedomain. At least in vitro, hsDcr-1 does not dis-tinguish termini and does not exhibit helicase-dependent threading. Unlike dmDcr-2, hsDcr-1may rely on the Platform-PAZ domain for gen-erating viral siRNAs. Indeed, mutations to thePlatform-PAZ domains of hsDcr-1 disrupt viralsiRNA biogenesis (19). However, given the con-servation of hsDcr-1’s helicase domain, it is intrigu-ing to consider that, under certain conditions,perhaps with additional factors, hsDcr-1 mightmediate processive cleavage by threading ofdsRNA through the helicase domain.
REFERENCES AND NOTES
1. M. Ha, V. N. Kim, Nat. Rev. Mol. Cell Biol. 15, 509–524(2014).
2. R. C. Wilson, J. A. Doudna, Annu. Rev. Biophys. 42, 217–239(2013).
3. K. Okamura, E. C. Lai, Nat. Rev. Mol. Cell Biol. 9, 673–678(2008).
4. R. Aliyari et al., Cell Host Microbe 4, 387–397 (2008).5. A. Flynt, N. Liu, R. Martin, E. C. Lai, Proc. Natl. Acad. Sci. U.S.A.
106, 5270–5275 (2009).6. L. R. Sabin et al., PLOS ONE 8, e55458 (2013).7. M. E. Fairman-Williams, U.-P. Guenther, E. Jankowsky,
Curr. Opin. Struct. Biol. 20, 313–324 (2010).8. S. Ahmad, S. Hur, Trends Biochem. Sci. 40, 576–585
(2015).9. Y. S. Lee et al., Cell 117, 69–81 (2004).10. A. Tsutsumi, T. Kawamata, N. Izumi, H. Seitz, Y. Tomari,
Nat. Struct. Mol. Biol. 18, 1153–1158 (2011).11. X.-H. Wang et al., Science 312, 452–454 (2006).12. D. Galiana-Arnoux, C. Dostert, A. Schneemann, J. A. Hoffmann,
J.-L. Imler, Nat. Immunol. 7, 590–597 (2006).13. J. T. Marques et al., PLOS Pathog. 9, e1003579 (2013).14. Q. Liu et al., Science 301, 1921–1925 (2003).15. N. C. Welker et al., Mol. Cell 41, 589–599 (2011).16. N. K. Sinha, K. D. Trettin, P. J. Aruscavage, B. L. Bass, Mol. Cell
58, 406–417 (2015).17. E. S. Cenik et al., Mol. Cell 42, 172–184 (2011).18. Y. Qiu et al., Immunity 46, 992–1004.e5 (2017).19. Y. Li et al., Nat. Microbiol. 2, 16250 (2016).20. H. Zhang, F. A. Kolb, V. Brondani, E. Billy, W. Filipowicz,
EMBO J. 21, 5875–5885 (2002).21. P.-W. Lau, C. S. Potter, B. Carragher, I. J. MacRae, Structure 17,
1326–1332 (2009).22. P.-W. Lau et al., Nat. Struct. Mol. Biol. 19, 436–440
(2012).23. D. W. Taylor et al., Nat. Struct. Mol. Biol. 20, 662–670
(2013).24. D. Luo et al., Cell 147, 409–422 (2011).25. E. Kowalinski et al., Cell 147, 423–435 (2011).26. K. Takahasi et al., Mol. Cell 29, 428–440 (2008).27. S. Myong et al., Science 323, 1070–1074 (2009).28. P. Linder, E. Jankowsky, Nat. Rev. Mol. Cell Biol. 12, 505–516
(2011).29. H. Zhang, F. A. Kolb, L. Jaskiewicz, E. Westhof, W. Filipowicz,
Cell 118, 57–68 (2004).30. I. J. Macrae et al., Science 311, 195–198 (2006).31. J.-E. Park et al., Nature 475, 201–205 (2011).32. Y. Tian et al., Mol. Cell 53, 606–616 (2014).33. S. U. Colmenares, S. M. Buker, M. Buhler, M. Dlakić, D. Moazed,
Mol. Cell 27, 449–461 (2007).34. E. Ma, I. J. MacRae, J. F. Kirsch, J. A. Doudna, J. Mol. Biol. 380,
237–243 (2008).35. S. K. Kandasamy, R. Fukunaga, Proc. Natl. Acad. Sci. U.S.A. 113,
14031–14036 (2016).36. Q. Wu et al., Proc. Natl. Acad. Sci. U.S.A. 107, 1606–1611
(2010).
Sinha et al., Science 359, 329–334 (2018) 19 January 2018 5 of 6
100
6050
30
20
10
40
AH Dec
ade
0 5 15 75 0 5 15 75 0 5 15 75 min
Unc
leav
edC
leave
d
1 2 3 4 5 6 7 8 9 10 11 12Lanes 13
Block-28
dmDcr-2WT, ATP, Streptavidin
Block-37 No Block
0
20
40
60
80
100
dsR
NA
cle
aved
(%
)
***********
********n.s.
********
***********
***n.s.***
n.s.
*******
Block-28 Block-37 No Block
dmDcr-2WT, ATP, Streptavidin
52 BLT dsRNA
% cleaved(total)
% cleaved> 23 nt
% cleaved21-23 nt
% cleaved11-20 nt
% cleaved< 11 nt
Time : 75 min
52 BLT Block-28 dsRNA
B
B
32P
52 BLT Block-37 dsRNA
B
B
32P
52 BLT No Block dsRNA
32P
100
6050
30
20
10
40
Unc
leav
edC
leave
d
52 BLT dsRNA, ATP
RNA only dmDcr-2WT dmDcr-2WThsDcr-1WT hsDcr-1WT
+ Streptavidin– Streptavidin
1 2 3 4 5 6 7 8 9 10 1112Lanes 13 14 15 16 17
AH Dec
ade
28 37 No
28 37 No
28 37 No
28 37 No
28 37 No Block
BLT dsRNA 3’ovr dsRNA
apo dmDcr-2Hel1
Hel2Hel2i
PAZPlatformRIIIa
RIIIbPincer
ATP dependent
ATP independent
Binding
Threading
Unwinding
ATPADP Pi
ATPADP Pi
22 nt siRNA cleavage
Pro
cess
ive
clea
vag
e
ATPADP Pi
substrate release
22 nt siRNA cleavage
52 BLT dsRNA
Fig. 4. BLTdsRNA threads through helicase domain. (A) Substrates for (B) to (D); somefeatures described in Fig. 2, B and E. Blocked dsRNAs contained biotin-dT (red B) onboth strands, with 28 and 37 indicating the position from the 5′ end of the sense strand.(B) Single-turnover cleavage assays of blocked or unblocked 52 BLT dsRNA (1 nM) withdmDcr-2WT (30 nM), 5 mM ATP, and 80 nM streptavidin (n = 3). dsRNA was preincubated withstreptavidin before adding dmDcr-2WT. Arrow, AH, as in Fig. 2E. (C) Quantification of cleavage at75 min, as in (B). dsRNA cleaved (%) is plotted based on all products (total), those >23 nt,siRNAs (21 to 23 nt products), those 11 to 20 nts, and those <11 nts. Data points, mean ± SD(n = 3). *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001; P > 0.05, n.s. (nonsignificant).(D) Single-turnover cleavage assays as in (B), with dmDcr-2WT or hsDcr-1WT, ±80 nM streptavidin(n = 3). Black arrow, siRNA product (22 nt) with dmDcr-2WT; green arrow, siRNA product(26 nt) with hsDcr-1WT. (E) Model for recognition and cleavage of BLT and 3′ovr dsRNA bydmDcr-2. Dotted orange arrow, clamping of helicase on BLT dsRNA; dotted white arrow,unwinding; dotted gray box, dmDcr-2RIII•BLT dsRNA•ATP-gS complex shown in Fig. 1D (see alsofig. S10); red arrow, cleavage; gray arrow, threading intermediates (see text and movie S3 fordetails). Model for 3′ovr recognition from data reported here and elsewhere (see also movie S4) (23, 34).
RESEARCH | REPORTon O
ctober 18, 2020
http://science.sciencemag.org/
Dow
nloaded from

37. K. D. Trettin, N. K. Sinha, D. M. Eckert, S. E. Apple,B. L. Bass, Proc. Natl. Acad. Sci. U.S.A. 114, E7939–E7948(2017).
38. R. Zhou et al., RNA 15, 1886–1895 (2009).
ACKNOWLEDGMENTS
We thank D. Cazalla and E. Cao for critique of the manuscriptand P. J. Aruscavage for technical assistance. EM wasperformed at University of Utah EM Core Laboratory, withcomputational support from Utah Center for High PerformanceComputing. RNA was synthesized by the DNA/Peptide
facility (Health Sciences Center Cores at Universityof Utah). This work was supported by funding from theNational Institute of General Medical Sciences(R01GM121706) and the H. A. and Edna BenningPresidential Endowed Chair (to B.L.B.). The authorsdeclare no competing financial interests. The modelsand cryo-EM maps are available via the followingaccession numbers: Protein Data Bank (PDB) 6BUA,Electron Microscopy Data Bank (EMDB) EMD-7291,EMD-7292 (apo-dmDcr-2RIII); PDB 6BU9, EMD-7290(dmDcr-2RIII•52 BLT dsRNA•ATP-gS complex).
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/359/6373/329/suppl/DC1Materials and MethodsSupplementary TextFigs. S1 to S16Tables S1 to S3References (39–66)
29 September 2017; accepted 11 December 2017Published online 21 December 201710.1126/science.aaq0921
Sinha et al., Science 359, 329–334 (2018) 19 January 2018 6 of 6
RESEARCH | REPORTon O
ctober 18, 2020
http://science.sciencemag.org/
Dow
nloaded from

Dicer uses distinct modules for recognizing dsRNA terminiNiladri K. Sinha, Janet Iwasa, Peter S. Shen and Brenda L. Bass
originally published online December 21, 2017DOI: 10.1126/science.aaq0921 (6373), 329-334.359Science
, this issue p. 329Scienceprovide invertebrates with the optimization capabilities needed for antiviral defense.
-overhang dsRNAs and ATP-dependent, processive threading of blunt-end dsRNAs. This flexibility might′cleavage of 3distinct mechanisms for recognizing and cleaving substrates: adenosine triphosphate (ATP)-independent, distributive
Dicer protein that shed light on its twoDrosophila report structures of a et al.interference and antiviral defense. Sinha The Dicer protein generates short RNAs from double-stranded RNA (dsRNA) substrates and is critical for RNA
Substrate recognition by Dicer elucidated
ARTICLE TOOLS http://science.sciencemag.org/content/359/6373/329
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2017/12/20/science.aaq0921.DC1
REFERENCES
http://science.sciencemag.org/content/359/6373/329#BIBLThis article cites 66 articles, 11 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Science. No claim to original U.S. Government WorksCopyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of
on October 18, 2020
http://science.sciencem
ag.org/D
ownloaded from
Recommended