Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
69
Chapter 5
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica leaf and Embelin from Embelia ribes
fruit 5.1 INTRODUCTION
Major pharmaceutical companies are not interested in the evaluation of plant extracts.
The reasons are quite simple. Firstly, when an extract shows activity in a bioassay, the
active principle must be isolated and characterized. This is expensive and may take a
long time, depending on the availability of appropriate amount of extract or plant
material, the time for bioassay, and the ease of unambiguously determining the
structure. By this time, the synthetic “hits” will have moved to the next stage of
decision making and the natural product is left behind. Moreover, concern over the
availability of enough quantity of a chemical entity required for development and
market needs of natural products, has been the one most limiting factor for the
pharmaceutical industry’s interest in natural products. Market demand can reach a
scale of hundreds to thousands of kilograms per annum. Total synthesis will not
economically provide the complex natural product to meet this market demand. As
compared to the minor compounds in the plants, the active compounds present in high
concentrations increase their probability to be used for clinical purpose. In contrast,
the present study provides an alternative, simple, rapid and inexpensive approach for
isolation, identification and activity of active compounds from plant extracts.
5.2 MATERIALS AND METHODS
5.2.1 Plant material collection and authentication
Adhatoda zeylanica leaf and Embelia ribes fruit were collected from Gujrat, India.
Their authenticity was confirmed by the taxonomist Dr. Sheetal Anandijiwala of our
department in year 2005-2006 and voucher specimen was deposited at the Department
of Pharmacognosy and Phytochemistry, B. V. Patel Pharmaceutical Education &
Research Development (PERD) Centre, Ahmedabad, India. The material was stored
in air tight containers at room temperature until use.

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
70
5.2.2 Isolation of vasicine and vasicinone from methanolic extract of Adhatoda
zeylanica leaf and embelin from methanolic extract of Embelia ribes fruit
5.2.2.1 Preparation of methanolic extract
Dried leaves of A. zeylanica (500 g) were ground and extracted with methanol (1000
ml × 3) under reflux for 30 min at a maximum temperature of 50ºC. Dried fruits of E.
ribes (300 g) were ground and extracted with methanol (500 ml X 3) under reflux for
30 min at a maximum temperature of 50ºC. Both the extracts were filtered; the
filtrates were pooled and concentrated to dryness by removing the solvent under
reduced pressure at 50ºC. A mass of dark greenish sticky crude extract was obtained
from A. zeylanica leaf (93 g; 18.6 %) w/w and a mass of blackish sticky crude extract
of E. ribes (34 g; 11.3 % w/w). The extracts were stored in airtight glass bottles at 4ºC
in a refrigerator.
5.2.2.2 Fractionation of the methanolic extract of A. zeylanica and E. ribes
Methanolic extract of A. zeylanica (25 g) and E. ribes (8 g) were then suspended in
100 ml and 50 ml water respectively and partitioned successively with solvents in
increasing order of polarity viz, petroleum ether, chloroform, ethyl acetate, n-butanol
and the n-butanol insoluble fraction was also prepared. For A. zeylanica (100 ml X 3)
and for E. ribes 50 ml (X 3) solvent were used for liquid-liquid extraction. A.
zeylanica fractions were evaporated in vacuum to yield the residues of petroleum
ether (4 g; 16.0%), chloroform (0.8 g; 3.2 %), ethyl acetate (6.2 g; 25 %), n-butanol
(10 g; 40 %) fractions and n-butanol insoluble portion (0.9 g; 3.8 %) respectively. E.
ribes fractions were evaporated in vacuum to yield the residues of petroleum ether
(3.4 g; 42.5 %), chloroform (0.4 g; 5.3 %), ethyl acetate (1.6 g; 20 %), n-butanol (1 g;
12.4 %) fractions and n-butanol insoluble portion (0.7 g; 9.5 %) respectively. For
assay, fractions were dissolved in DMSO. Each fraction was bio-assayed for PfLDH.
Chloroform fraction of A. zeylanica and petroleum ether fraction of E. ribes
methanolic extract was found to have significant plasmodial lactate dehydrogenase
inhibitory activity as compared to other fractions of the respective plants. (Figure 5.1
& Figure 5.2)
5.2.2.3 Isolation of vasicine and vasicinone from methanolic extract of A. zeylanica leaf
TLC fingerprint profile

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
71
TLC fingerprint profile of all the fractions of A. zeylanica were developed in solvent
system of ethyl acetate: methanol: ammonia (8: 0.5: 0.2) (at 25 ± 2°C temperature and
40 % relative humidity) and Co-TLC of all the fractions was carried out along with
reference standard vasicine and vasicinone (purchased from SPIC, Chennai India) on
precoated silica gel 60F254 TLC plates (E. Merck). Since, chloroform fraction of A.
zeylanica leaf exhibited good activity as compared to other fractions and vasicine and
vasicinone are the major constituents present in the chloroform fraction of A.
zeylanica leaf. Hence we decided to isolate vasicine and vasicinone from A. zeylanica
leaf.
Isolation of vasicine from methanolic extract A. zeylanica leaf
Vasicine was isolated from A. zeylanica leaf by the reported method of acid–base
extraction (Chaitali D et al., 2005). 10 gm of methanolic extract of A. zeylanica leaf
was extracted with 25 ml (X 3) 2N HCl (pH 2-2.5) under reflux for 30 min at a
maximum temperature of 50ºC, consequently this acidic fraction was basified with
liquor ammonia to pH 8-8.5. This basic solution was extracted with 150 ml (X 3)
chloroform, with subsequent TLC monitoring of each step for the presence of
vasicine. Chloroform extract was pooled and evaporated to dryness by removing the
solvent under reduced pressure at 50°C to get 3.1 gm of alkaloid fraction. Vasicine
was identified as the major compound in the alkaloid fraction by TLC in solent
system ethyl acetae: methanol: ammonia (8: 0.5: 0.2). The alkaloid fraction was
dissolved in a minimum quantity of ethanol and vasicine was crystallized out from the
alkaloid fraction. It was purified by repeated crystallization with ethanol. White
needles of vasicine (150 mg) was obtained having melting point of 210°C. For
isolation of vasicinone remaining fraction was subjected to column chromatography.
Isolation of vasicine from methanolic extract A. zeylanica leaf
Stationary phase
Silica gel (for flash chromatography) 230-400 mesh (Spectrochem, India), 100 g.
Sample load
2 g of fraction was dissolved in minimum volume of methanol. The solution was
mixed with 3 g of silica gel and the solvent was evaporated. The fraction adsorbed on
silica was loaded on silicagel column.

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
72
Mobile phase
Remaining fraction was subjected to column chromatography over silica gel (Merck)
using mixtures of chloroform and methanol of increasing polarity. Vasicinone was
eluted from the column with chloroform: methanol (49:1) as a white solid and was
crystallised from chloroform: ethyl acetate.
Vasicine and vasicinone wee characterized by recording IR, MS and NMR spectra
and melting point and the identity of vasicine and vasicinone was confirmed by
comparison of their spectral data with those reported (Jain et al., 1980; Joshi et al.,
1994).
5.2.2.4 Isolation of embelin from methanolic extract of E. ribes fruit
TLC fingerprint profile
TLC profile of all the fractions of E. ribes was developed in solvent system of n-
propanol: n-butanol : ammonia (7: 1: 2) (at 25 ± 2°C temperature and 40 % relative
humidity) and Co-TLC of the fractions was carried out along with reference standard
embelin on precoated silica gel 60F254 TLC plates (E. Merck).
Since, petroleum ether fraction of E. ribes fruit exhibited good activity as compared to
other fractions and embelin was the major constituents present in the petroleum
fraction of E. ribes fruit. Hence we decided to isolate embelin from E. ribes fruit.
Isolation of embelin from methanolic extract E. ribes fruit
25 gm of E. ribes fruit was ground and extracted with 100 ml X 4 petroleum ether
under reflux for 30 min at a maximum temperature of 50ºC. The extract was filtered,
concentrated to 1/3 volume and kept overnight at room temperature. Embelin settled
at the bottom, and after decanting the supernatant, orange coloured mass was
obtained. It was purified by repeated crystallization with methanol, to obtain
glistening orange crystals of embelin (722 mg) having a melting point of 142-143°C.
It was characterized by recording UV, IR, MS and NMR spectra and melting point
and comparing with the reported data (Kumara Swamy HM et al., 2007; Haq et al.,
2007).

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
73
5.2.3 Structure elucidation of vasicine, vasicinone and embelin
The melting point, Infra-red (IR), Mass, 1H-NMR and 13C-NMR spectra were
recorded for vasicine and embelin. The melting point was measured using a melting
point apparatus (Toshniwal, India). IR spectrum was recorded using KBr disk method
on IR spectrophotometer (Model 500, BUCK SCIENTIFIC). Mass spectra were
recorded in the positive ion mode on mass spectrophotometer (API 165, PERKIN-
ELMER). NMR spectra were recorded on FT-NMR spectrophotometer (DRX 300,
Bruker) at 300 MHz, using tetramethylsilane as internal standard.
5.2.4 In vitro testing of the antiplasmodial activity of vasicine, vasicinone,
embelin
The three isolated compounds were evaluated for antimalarial activity in schizont
maturation inhibition assay and Plasmodium falciparum lactate dehydrogenase
inhibition assay (PfLDH).
5.2.4.1 Schizont maturation inhibition assay
Experimental protocol
The test procedure for antimalarial screening described in section 2.2.5.1 was used.
Sample preparation
One mg of each compound was dissolved in 1 μl DMSO and 999 μl RPMI-1640 to
obtain a stock of 1 mg/ml (stock solution). A series of eight concentrations were
prepared from the stock solutions by 2-fold dilutions (0.0625–2 mg: ml) to give
concentrations around the range of 50% inhibition (IC50).
5.2.4.2 Plasmodium falciparum lactate dehydrogenase inhibition assay
Experimental protocol
The test procedure for antimalarial screening described in section 2.2.5.2 was used.
Sample preparation
The compounds were dissolved in DMSO separately to produce a stock solution of 1
mg/ml each. These stock solutions were subsequently diluted with culture medium
containing 10% human serum before being transferred in triplicate of 10 µl each at
concentrations of 1,10,25,50 and 100 μg/ml into two 96-well microtiter plates.
5.2.5 In vivo testing of the antiplasmodial activity of vasicine, vasicinone, embelin

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
74
The three isolated compounds were evaluated for antimalarial activity in Plasmodium
berghei model.
5.2.5.1 Introduction
Plasmodium berghei belongs to a group of four Plasmodium species that infect
murine rodents. These species are P. vinckei, P. chabaudi, P. yoelii and P. berghei.
After the original discovery of P. berghei was made by Vincke and Lips in 1948,
since then a number of isolates (strains) have been collected from the wild. P. berghei
infects laboratory hamsters, rats and mice. A susceptible mosquito vector for P.
berghei, which is widely used in the laboratory, is Anopheles stephensi (Sinden,
1996).
These rodent parasites exhibited similarity to human malaria parasites with respect to
both proteomic and genomic level. For example, a number of (surface) proteins
demonstrate conservation both in structure and function between rodent and human
malaria parasites (such as CS, TRAP, P45/48, CTRP, P25, P28, AMA-1, MSP-1)
(Sinden, 1978; Aikawa and Seed, 1980; Lin et al., 2000). Also studies demonstrated a
high level of conservation of genome organisation between rodent and human
parasites. The genome of both P. falciparum and the four rodent parasites are
organised into 14 linear chromosomes, ranging in size from 0.5-3.8 Mb. They possess
identical telomeric repeats [CCCT (A/G) AA], organised into tandem arrays.
Comparative mapping of genes located in the central regions of the chromosomes has
shown that both linkage and gene order appear to be well conserved between human
and rodent parasites, resulting in significant level of synteny between these species
(Carlton et al., 1998; Lin et al., 2001; Rich and Ayala; 2003). Moreover, conservation
of specific gene domains, regulatory control elements (Janse et al., 1994),
organisation of complex genomic loci and the presence of conserved multi-gene
families in rodent and human parasites (Homewood and Neame, 1980) all emphasize
the high similarity of genome organisation, gene content and gene-regulation.
These facts suggest that murine malaria models are particularly valuable in studies of
the erythrocytic stage of malaria infection, because the morphology and parasite
development stages are similar to those in human malaria infections. An important
advantage with murine malaria models is that all parasite stages can be observed in
the peripheral circulation (Gibbons et al., 2007; Mackenstedt et al., 1989). Also, it is

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
75
easy and safe to handle and manipulate any stage of the life cycle of murine malaria
parasites in the laboratory, and such models often represent the only practical means
towards in vivo experimentation (Janse and Waters, 1995). Thus, species of malaria
parasites that infect rodents provide models for the study of the biology of malaria
parasites that infect humans (Sinden, 1996).
5.2.5.2 Experimental
Parasite density determination and inoculum preparation
A standard inoculum was prepared from a donor mouse with P. berghei parasitized
erythrocytes. Infected blood from the donor mouse was obtained by cardiac puncture
after anesthesia with chloroform Parasitaemia was established by microscopic
examination of a thin blood film under oil immersion at x 100 magnification and
measured as a percentage of infected erythrocytes in fields of 500 erythrocytes. Each
mouse was infected with a standard inoculum of the 106 parasitized erythrocyte
suspension in phosphate buffered saline (0.2 ml) from a donor mouse that was
prepared based on percentage parasitaemia and number of erythrocytes counted per
microlitre of blood using an improved Neubear Haematocytometer.
Treatment
A 4-day suppressive test according to Peter et al (1975) was used. The animals were
divided into 5 groups of 6 mice each. Animals in group 1 were given the vehicle only
(i.e. 0.2 ml PBS) and served as the uninfected control. Animals in group 2 were
administered 20 mg/kg b wt. suppressive dose of chloroquine and those in groups 3,4
and 5 were administered vasicine (40 and 100 mg/kg b wt.), vasicinone (500 mg/kg b
wt.) and embelin (500 mg/kg b wt.) respectively. Treatments commenced
immediately after inoculation (Day 0) and continued on days 1, 2 and 3 via an oral
route using a canula. Tail blood films of the infected animals were prepared, fixed in
methanol, stained with 4% Giemsa stain for 20 min and examined microscopically
under oil immersion on day 4 post inoculations. Parasite density and percentage
reduction in parasitaemia relative to chloroquine control on group by-group basis
were calculated.
5.3 RESULTS AND DISCUSSION

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
76
5.3.1 Isolation of vasicine and vasicinone from methanolic extract of Adhatoda
zeylanica leaf and embelin from methanolic extract of Embelia ribes fruit
Methanolic extract of A. zeylanica leaf (IC50 5.80 µg/ml) and E. ribes fruit (IC50 13.10
µg/ml) showed good antiplasmodial activity in inhibiting PfLDH
5.3.1.1 Fractionation of the methanolic extract
Methanolic extracts of A. zeylanica and E. ribes were fractionated in different polarity
solvents viz, hexane, chloroform, ethyl acetate, n-butanol fractions and n-butanol
insoluble fraction. IC50 values for A. zeylanica fractions were as follows (Figure 5.1):
petroleum ether fraction 243.70 µg/ml, chloroform fraction 6.76 µg/ml, ethyl acetate
fraction 971.16 µg/ml, n-butanol fraction 99.84 µg/ml and n-butanol insoluble
fraction 1283.73 μg/ml. IC50 values for E. ribes fractions were as follows: petroleum
ether fraction 5.26 µg/ml, chloroform fraction 100.33 µg/ml, ethyl acetate fraction
130.00 µg/ml, n-butanol fraction 3883.87 µg/ml and n-butanol insoluble fraction
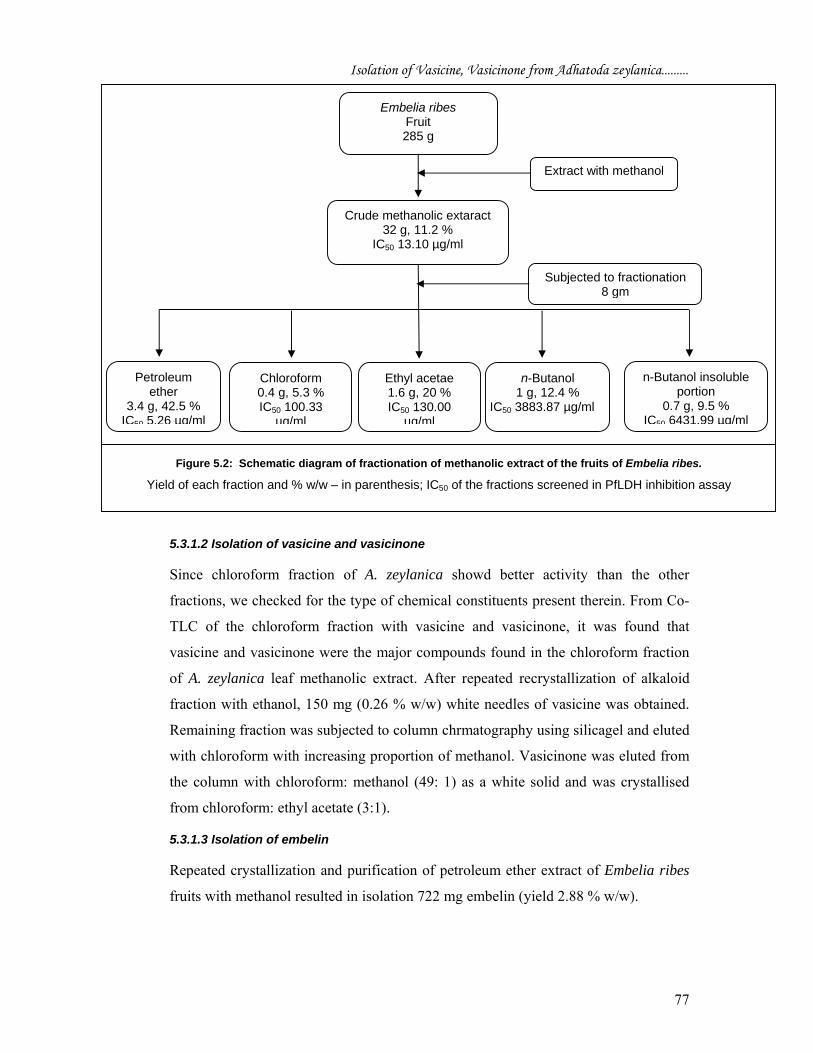
6431.99 μg/ml (Figure 5.2).
Adhatoda zeylanica
Leaf 480 g
Petroleum ether 4.0 g, 16 %
IC50 243.70 µg/ml
Chloroform 0.8 g, 3.2 %
IC50 6.76 µg/ml
Ethyl acetae 6.25 g, 25 % IC50 971.16
µg/ml
n-Butanol 10.0 g, 40 %
IC50 99.84 µg/ml
Subjected to fractionation 25 g
Extract with methanol
n-Butanol insoluble portion
0.9 g, 3.8 % IC50 1283.73 µg/ml
Crude methanolic extaract 89 g, 18.6 %
IC50 5.80 µg/ml
Figure 5.1: Schematic diagram of fractionation of methanolic extract of the leaf of Adhatoda zeylanica.
Yield of each fraction and % w/w – in parenthesis; IC50 of the fractions screened in PfLDH inhibition assay
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
77
5.3.1.2 Isolation of vasicine and vasicinone
Since chloroform fraction of A. zeylanica showd better activity than the other
fractions, we checked for the type of chemical constituents present therein. From Co-
TLC of the chloroform fraction with vasicine and vasicinone, it was found that
vasicine and vasicinone were the major compounds found in the chloroform fraction
of A. zeylanica leaf methanolic extract. After repeated recrystallization of alkaloid
fraction with ethanol, 150 mg (0.26 % w/w) white needles of vasicine was obtained.
Remaining fraction was subjected to column chrmatography using silicagel and eluted
with chloroform with increasing proportion of methanol. Vasicinone was eluted from
the column with chloroform: methanol (49: 1) as a white solid and was crystallised
from chloroform: ethyl acetate (3:1).
5.3.1.3 Isolation of embelin
Repeated crystallization and purification of petroleum ether extract of Embelia ribes
fruits with methanol resulted in isolation 722 mg embelin (yield 2.88 % w/w).
Chloroform 0.4 g, 5.3 % IC50 100.33
µg/ml
Embelia ribes Fruit 285 g
Petroleum ether
3.4 g, 42.5 % IC50 5.26 µg/ml
Ethyl acetae 1.6 g, 20 % IC50 130.00
µg/ml
n-Butanol 1 g, 12.4 %
IC50 3883.87 µg/ml
Subjected to fractionation 8 gm
Extract with methanol
n-Butanol insoluble portion
0.7 g, 9.5 % IC50 6431.99 µg/ml
Crude methanolic extaract 32 g, 11.2 %
IC50 13.10 µg/ml
Figure 5.2: Schematic diagram of fractionation of methanolic extract of the fruits of Embelia ribes.
Yield of each fraction and % w/w – in parenthesis; IC50 of the fractions screened in PfLDH inhibition assay
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
78
5.3.1.4 Spectral details of vasicine
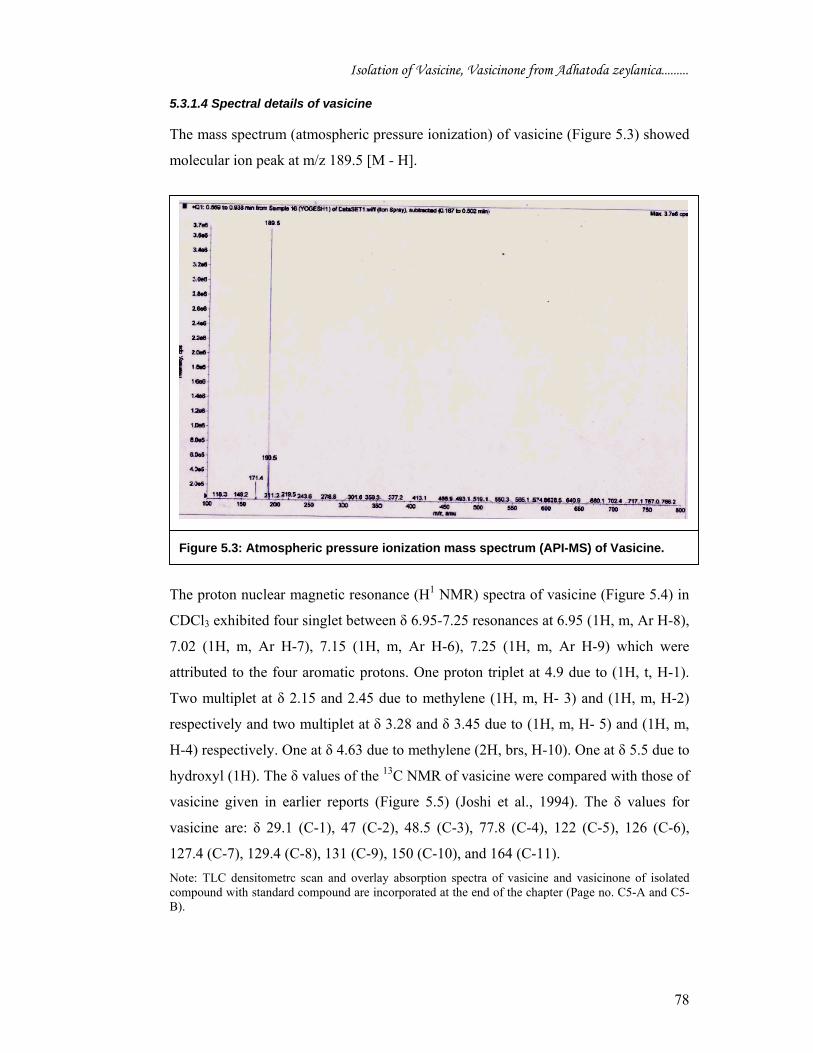
The mass spectrum (atmospheric pressure ionization) of vasicine (Figure 5.3) showed
molecular ion peak at m/z 189.5 [M - H].
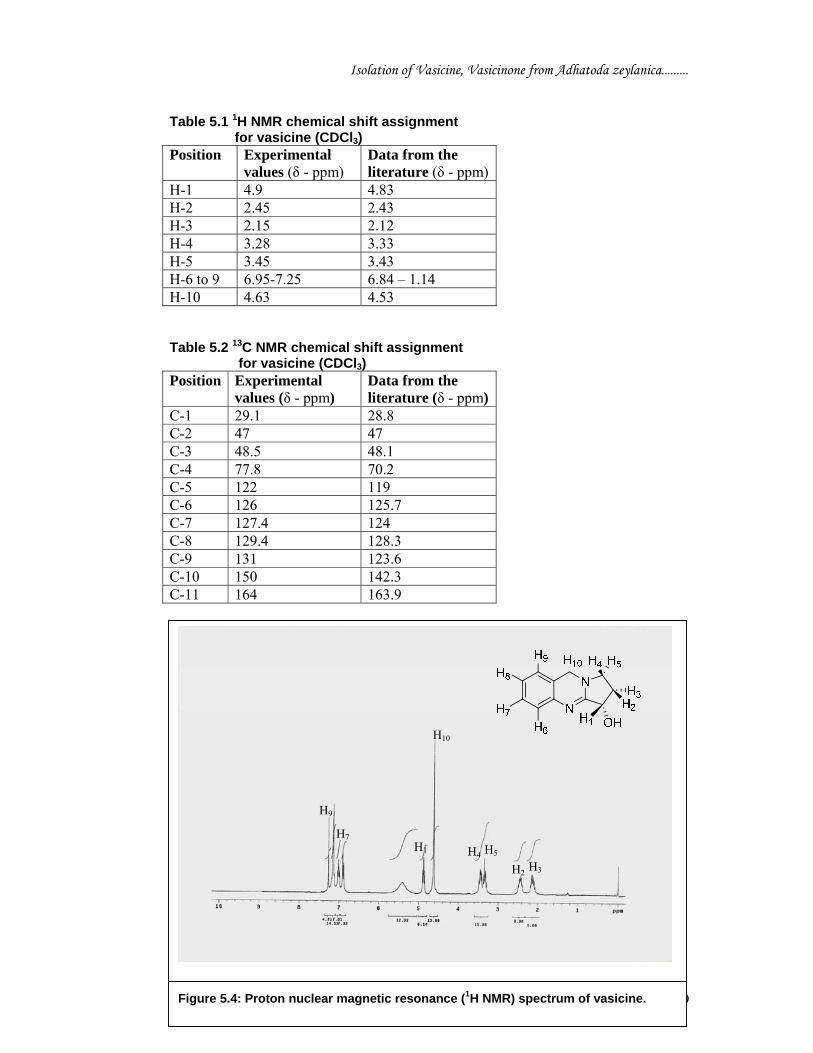
The proton nuclear magnetic resonance (H1 NMR) spectra of vasicine (Figure 5.4) in
CDCl3 exhibited four singlet between δ 6.95-7.25 resonances at 6.95 (1H, m, Ar H-8),
7.02 (1H, m, Ar H-7), 7.15 (1H, m, Ar H-6), 7.25 (1H, m, Ar H-9) which were
attributed to the four aromatic protons. One proton triplet at 4.9 due to (1H, t, H-1).
Two multiplet at δ 2.15 and 2.45 due to methylene (1H, m, H- 3) and (1H, m, H-2)
respectively and two multiplet at δ 3.28 and δ 3.45 due to (1H, m, H- 5) and (1H, m,
H-4) respectively. One at δ 4.63 due to methylene (2H, brs, H-10). One at δ 5.5 due to
hydroxyl (1H). The δ values of the 13C NMR of vasicine were compared with those of
vasicine given in earlier reports (Figure 5.5) (Joshi et al., 1994). The δ values for
vasicine are: δ 29.1 (C-1), 47 (C-2), 48.5 (C-3), 77.8 (C-4), 122 (C-5), 126 (C-6),
127.4 (C-7), 129.4 (C-8), 131 (C-9), 150 (C-10), and 164 (C-11). Note: TLC densitometrc scan and overlay absorption spectra of vasicine and vasicinone of isolated compound with standard compound are incorporated at the end of the chapter (Page no. C5-A and C5-B).
Figure 5.3: Atmospheric pressure ionization mass spectrum (API-MS) of Vasicine.
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
79
Table 5.1 1H NMR chemical shift assignment for vasicine (CDCl3) Position Experimental
values (δ - ppm) Data from the literature (δ - ppm)
H-1 4.9 4.83 H-2 2.45 2.43 H-3 2.15 2.12 H-4 3.28 3.33 H-5 3.45 3.43 H-6 to 9 6.95-7.25 6.84 – 1.14 H-10 4.63 4.53 Table 5.2 13C NMR chemical shift assignment for vasicine (CDCl3) Position Experimental
values (δ - ppm) Data from the literature (δ - ppm)
C-1 29.1 28.8 C-2 47 47 C-3 48.5 48.1 C-4 77.8 70.2 C-5 122 119 C-6 126 125.7 C-7 127.4 124 C-8 129.4 128.3 C-9 131 123.6 C-10 150 142.3 C-11 164 163.9
H8
H6
Figure 5.4: Proton nuclear magnetic resonance (1H NMR) spectrum of vasicine.
H1
H10
H4 H5 H2 H3
H7
H9
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
80
Thus, identity of vasicine was confirmed by recording IR, MS and NMR spectra and
by comparison of the spectral data with those reported (Jain et al., 1980; Joshi et al.,
1994). 5.3.1.5 Spectral details of vasicinone
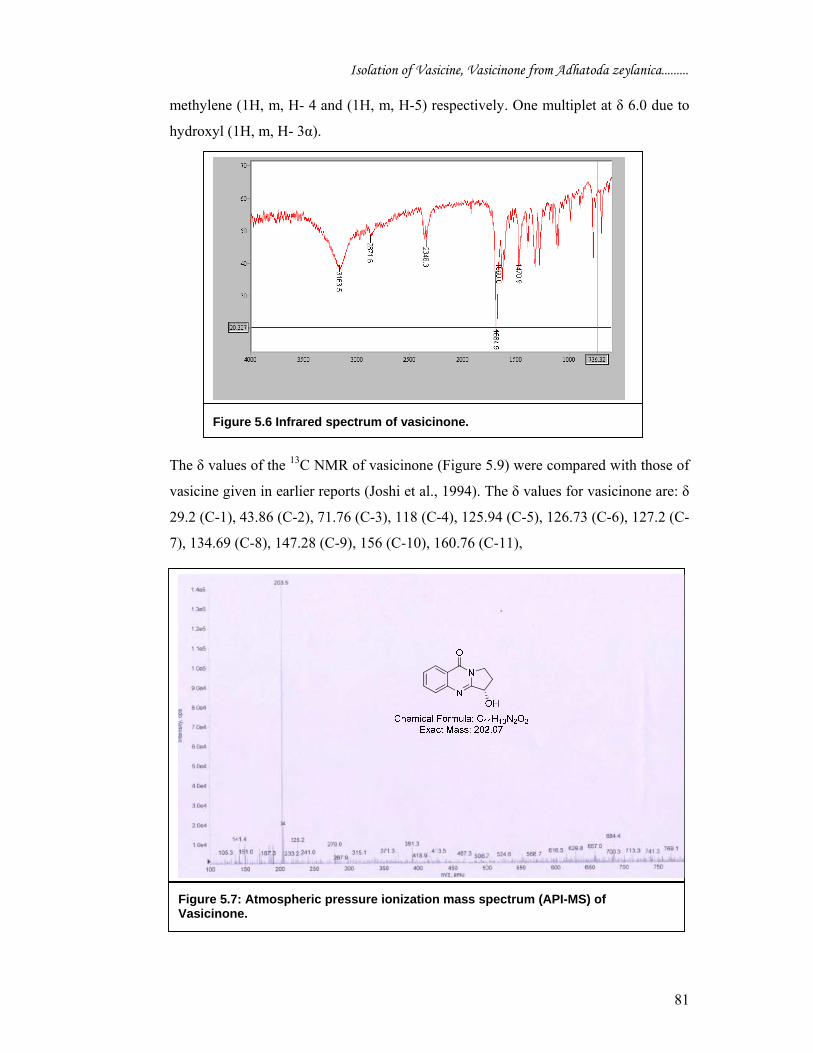
The IR (KBr) spectrum of vasicinone (Figure 5.6) showed the characteristic
absorption at 1470 cm-1 (due to CN bond), 1660, 1684 cm-1 (due to stretching
vibration of C = O), 2871, 3163 cm-1 (due to stretching vibration of OH group). The
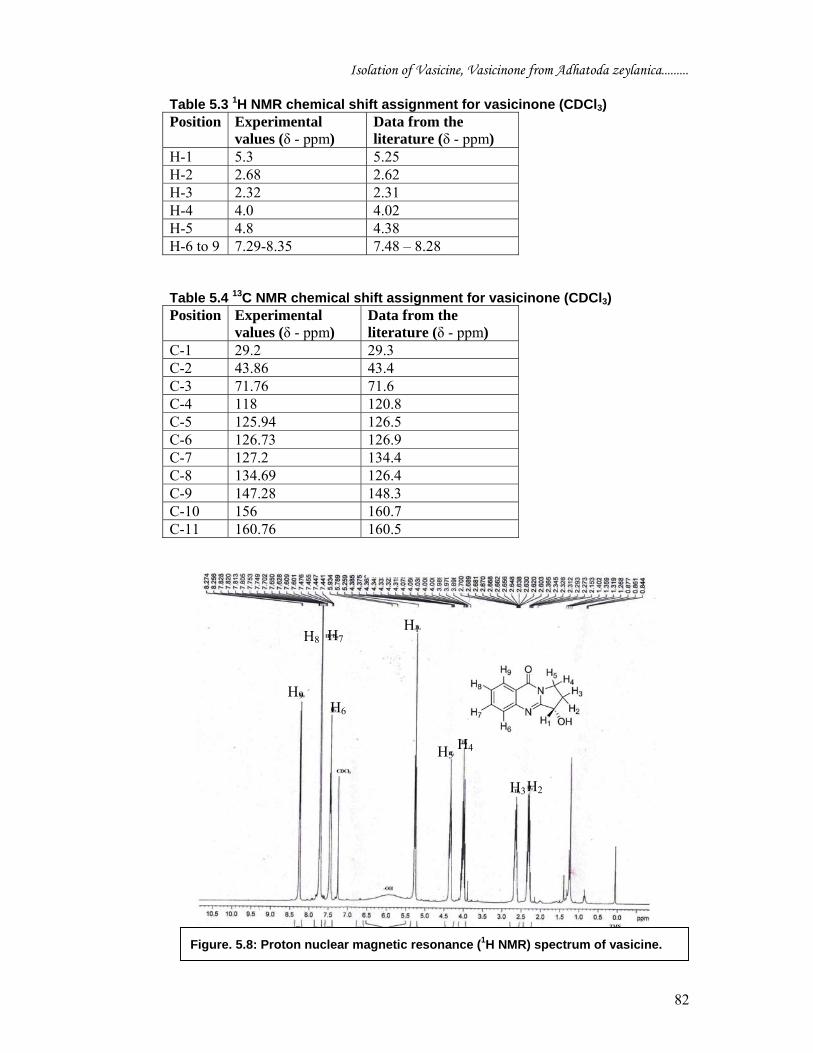
mass spectrum (atmospheric pressure ionization) of vasicinone [Figure 5.7] showed
molecular ion peak at m/z 203.5 [M - H]-.
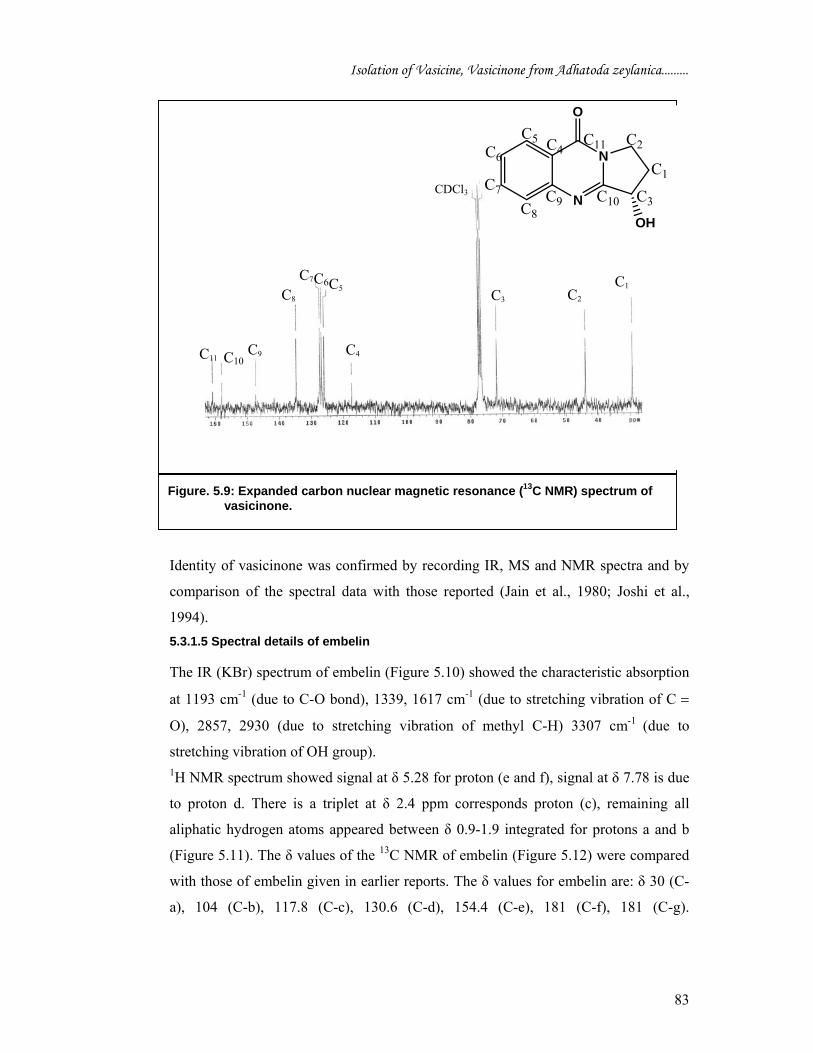
The proton nuclear magnetic resonance (H1 NMR) spectra of vasicinone (Figure 5.8)
in CDCl3 exhibited four singlet at between δ 7.29-8.35 resonances at δ 7.29 (1H, m,
Ar H-6), δ 7.8 (1H, m, Ar H-7), δ 7.53 (1H, m, Ar H-8), δ 8.35 (1H, m, Ar H-9)
which were attributed to the four aromatic protons. One proton triplet at 5.3 due to
methane (CH) (1H, t, H1). Two multiplet at δ 2.32 and 2.68 due to methylene (1H, m,
H- 3) and (1H, m, H-2) respectively and two multiplet at δ 4.0 and 4.8 due to
C1 C3 C4 C2
C6 C7
C9
C10 C11
CDCl3
C8
C5
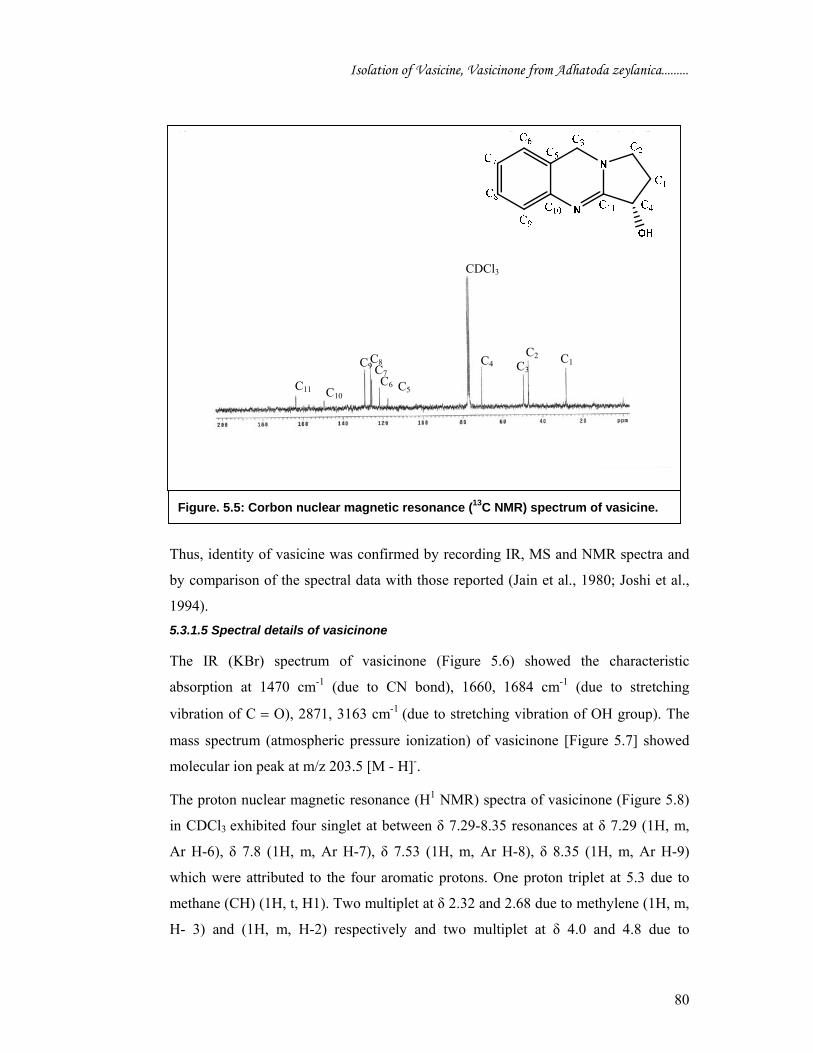
Figure. 5.5: Corbon nuclear magnetic resonance (13C NMR) spectrum of vasicine.
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
81
methylene (1H, m, H- 4 and (1H, m, H-5) respectively. One multiplet at δ 6.0 due to
hydroxyl (1H, m, H- 3α).
The δ values of the 13C NMR of vasicinone (Figure 5.9) were compared with those of
vasicine given in earlier reports (Joshi et al., 1994). The δ values for vasicinone are: δ
29.2 (C-1), 43.86 (C-2), 71.76 (C-3), 118 (C-4), 125.94 (C-5), 126.73 (C-6), 127.2 (C-
7), 134.69 (C-8), 147.28 (C-9), 156 (C-10), 160.76 (C-11),
Figure 5.6 Infrared spectrum of vasicinone.
Figure 5.7: Atmospheric pressure ionization mass spectrum (API-MS) of Vasicinone.
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
82
Table 5.3 1H NMR chemical shift assignment for vasicinone (CDCl3) Position Experimental
values (δ - ppm) Data from the literature (δ - ppm)
H-1 5.3 5.25 H-2 2.68 2.62 H-3 2.32 2.31 H-4 4.0 4.02 H-5 4.8 4.38 H-6 to 9 7.29-8.35 7.48 – 8.28 Table 5.4 13C NMR chemical shift assignment for vasicinone (CDCl3) Position Experimental
values (δ - ppm) Data from the literature (δ - ppm)
C-1 29.2 29.3 C-2 43.86 43.4 C-3 71.76 71.6 C-4 118 120.8 C-5 125.94 126.5 C-6 126.73 126.9 C-7 127.2 134.4 C-8 134.69 126.4 C-9 147.28 148.3 C-10 156 160.7 C-11 160.76 160.5
Figure. 5.8: Proton nuclear magnetic resonance (1H NMR) spectrum of vasicine.
H1
H2 H3
H4H5
H6
H7 H8
H9
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
83
Identity of vasicinone was confirmed by recording IR, MS and NMR spectra and by
comparison of the spectral data with those reported (Jain et al., 1980; Joshi et al.,
1994). 5.3.1.5 Spectral details of embelin
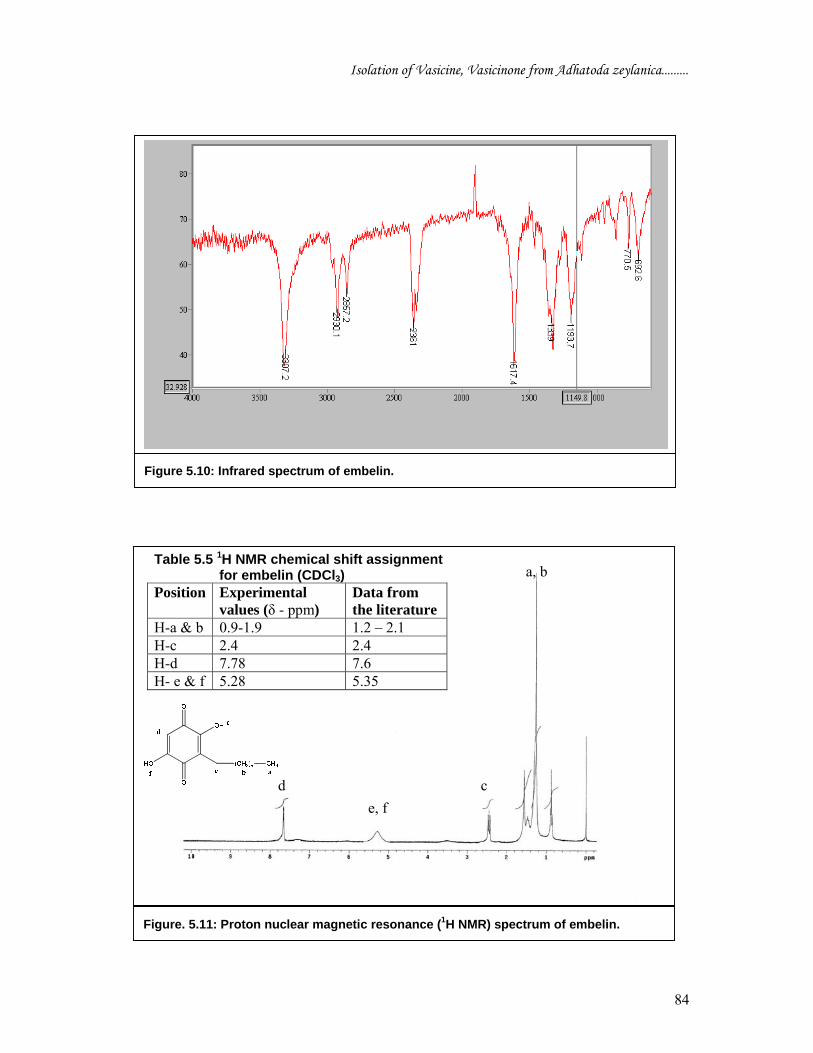
The IR (KBr) spectrum of embelin (Figure 5.10) showed the characteristic absorption
at 1193 cm-1 (due to C-O bond), 1339, 1617 cm-1 (due to stretching vibration of C =
O), 2857, 2930 (due to stretching vibration of methyl C-H) 3307 cm-1 (due to
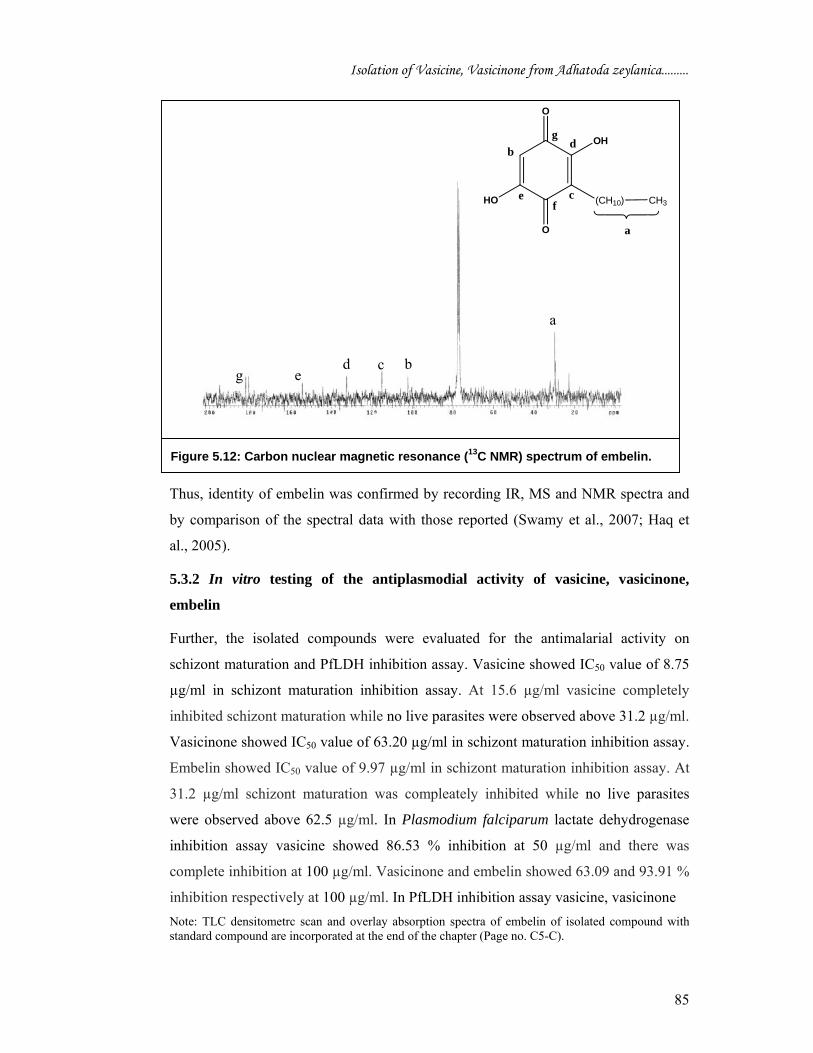
stretching vibration of OH group). 1H NMR spectrum showed signal at δ 5.28 for proton (e and f), signal at δ 7.78 is due
to proton d. There is a triplet at δ 2.4 ppm corresponds proton (c), remaining all
aliphatic hydrogen atoms appeared between δ 0.9-1.9 integrated for protons a and b
(Figure 5.11). The δ values of the 13C NMR of embelin (Figure 5.12) were compared
with those of embelin given in earlier reports. The δ values for embelin are: δ 30 (C-
a), 104 (C-b), 117.8 (C-c), 130.6 (C-d), 154.4 (C-e), 181 (C-f), 181 (C-g).
C6
C10
Figure. 5.9: Expanded carbon nuclear magnetic resonance (13C NMR) spectrum of vasicinone.
C1 C2 C3
C4
C5 C7
C8
C9 C11
CDCl3 N
N
O
OH
C1
C2
C3
C4C5
C6
C7
C8C9 C10
C11
C6
C10
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
84
Figure 5.16: Carbon nuclear magnetic resonance (13C NMR) spectrum of embelin.
Figure 5.10: Infrared spectrum of embelin.
a, b
Figure. 5.11: Proton nuclear magnetic resonance (1H NMR) spectrum of embelin.
c e, f
d
Table 5.5 1H NMR chemical shift assignment for embelin (CDCl3) Position Experimental
values (δ - ppm) Data from the literature
H-a & b 0.9-1.9 1.2 – 2.1 H-c 2.4 2.4 H-d 7.78 7.6 H- e & f 5.28 5.35
a, b
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
85
Thus, identity of embelin was confirmed by recording IR, MS and NMR spectra and
by comparison of the spectral data with those reported (Swamy et al., 2007; Haq et
al., 2005).
5.3.2 In vitro testing of the antiplasmodial activity of vasicine, vasicinone,
embelin
Further, the isolated compounds were evaluated for the antimalarial activity on
schizont maturation and PfLDH inhibition assay. Vasicine showed IC50 value of 8.75
µg/ml in schizont maturation inhibition assay. At 15.6 µg/ml vasicine completely
inhibited schizont maturation while no live parasites were observed above 31.2 µg/ml.
Vasicinone showed IC50 value of 63.20 µg/ml in schizont maturation inhibition assay.
Embelin showed IC50 value of 9.97 µg/ml in schizont maturation inhibition assay. At
31.2 µg/ml schizont maturation was compleately inhibited while no live parasites
were observed above 62.5 µg/ml. In Plasmodium falciparum lactate dehydrogenase
inhibition assay vasicine showed 86.53 % inhibition at 50 µg/ml and there was
complete inhibition at 100 µg/ml. Vasicinone and embelin showed 63.09 and 93.91 %
inhibition respectively at 100 µg/ml. In PfLDH inhibition assay vasicine, vasicinone Note: TLC densitometrc scan and overlay absorption spectra of embelin of isolated compound with standard compound are incorporated at the end of the chapter (Page no. C5-C).
f
Figure 5.12: Carbon nuclear magnetic resonance (13C NMR) spectrum of embelin.
O
O
OH
HO
b
(CH10) CH3
a
c
d
ef
g
a
b c d e g

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
86
and embelin showed an IC50 of 2.05, 57.20 and 3.33 µg/ml respectively. The results
indicate that vasicine and embelin showed dose dependent schizonticidal activity. At
higher concentrations embelin was found to kill all the parasites. In vasicine treated
samples a few dead parasites were seen.
5.3.3 In vivo testing of the antiplasmodial activity of vasicine, vasicinone, embelin
The results of the previous in vitro experiments provided the rationale for determining
whether vasicine, vasicinone and embelin could prevent or suppress in vivo growth of
the rodent malarial parasite P. berghei. This In vivo evaluation of antimalarial
compounds typically begins with the use of rodent malaria parasites. Of these, P.
berghei, P. yoelii, P. chabaudi and P. vinckei have been used extensively in drug
discovery and early development (Childs et al., 1984). These rodent models have been
validated through the identification of several antimalarials-for example, mefloquine,
halofantrine and artemisinin derivatives (Peters et al. 1977; Peters et al., 1987;
Vennerstrom et al., 2000; Posner et al., 2003). In view of their proven use in the
prediction of treatment outcomes for human infections, these models remain a
standard part of the drug discovery and development pathway.
Of these above mentioned models, Plasmodium berghei model is the most extensively
used model. Swiss mice are tolerant of P. berghei infection at a low to moderate level
of parasitaemia (<5%), with organ failure and death occurring only at high
parasitaemia (Cox, 1988; Landau and Gautret, 1998; Peters and Robinson, 1999).
Hence, the features of P. berghei infection model are beneficial for pharmacodynamic
studies because tolerance of the infection allows the mice to reach a parasitaemia of
up to 5% before drug treatment strategies are commenced and the parasitaemia can be
monitored down to a level of approximately 0.005%.
Five groups of mice were infected with 106 parasitized erythrocytes from the blood of
a single mouse parasitized with P. berghei. After treatment with compounds and drug,
the suppression of parasitemia (chemo-suppression) and mean survival rate (time)
were used as measures of efficacy. The effect of the drug was considered ineffective
if: before and after treatment no significant difference in degrees of parasitaemia or a
continuous increase of parasitaemia is noted. In the present experiment animals
treated with compounds showed decrease in parasitaemia. Samples were categorized
as highly active when chemo-suppression was above 60% or moderately active
Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
87
between 30 and 60%, but lowly active below 30%. Mean survival time of the
experimental groups of test animals correlated to treatment.
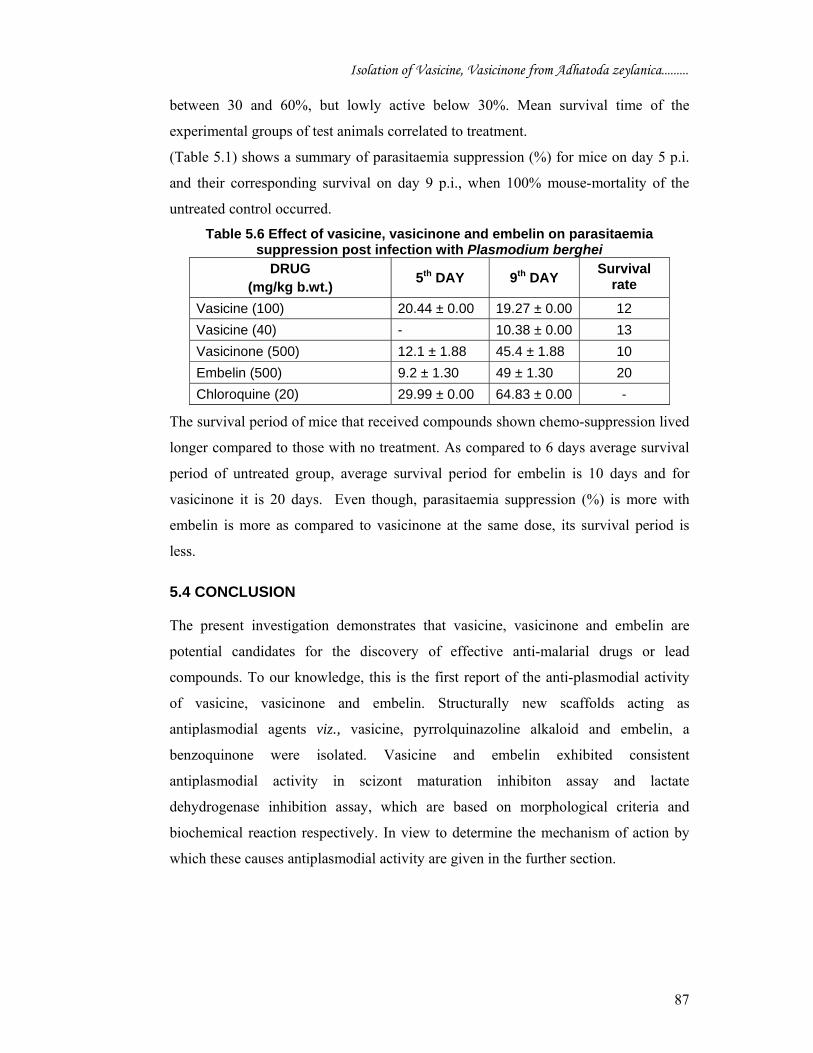
(Table 5.1) shows a summary of parasitaemia suppression (%) for mice on day 5 p.i.
and their corresponding survival on day 9 p.i., when 100% mouse-mortality of the
untreated control occurred.
Table 5.6 Effect of vasicine, vasicinone and embelin on parasitaemia suppression post infection with Plasmodium berghei
DRUG (mg/kg b.wt.) 5th DAY 9th DAY Survival
rate
Vasicine (100) 20.44 ± 0.00 19.27 ± 0.00 12 Vasicine (40) - 10.38 ± 0.00 13 Vasicinone (500) 12.1 ± 1.88 45.4 ± 1.88 10 Embelin (500) 9.2 ± 1.30 49 ± 1.30 20 Chloroquine (20) 29.99 ± 0.00 64.83 ± 0.00 -
The survival period of mice that received compounds shown chemo-suppression lived
longer compared to those with no treatment. As compared to 6 days average survival
period of untreated group, average survival period for embelin is 10 days and for
vasicinone it is 20 days. Even though, parasitaemia suppression (%) is more with
embelin is more as compared to vasicinone at the same dose, its survival period is
less.
5.4 CONCLUSION
The present investigation demonstrates that vasicine, vasicinone and embelin are
potential candidates for the discovery of effective anti-malarial drugs or lead
compounds. To our knowledge, this is the first report of the anti-plasmodial activity
of vasicine, vasicinone and embelin. Structurally new scaffolds acting as
antiplasmodial agents viz., vasicine, pyrrolquinazoline alkaloid and embelin, a
benzoquinone were isolated. Vasicine and embelin exhibited consistent
antiplasmodial activity in scizont maturation inhibiton assay and lactate
dehydrogenase inhibition assay, which are based on morphological criteria and
biochemical reaction respectively. In view to determine the mechanism of action by
which these causes antiplasmodial activity are given in the further section.

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
88
References
Aikawa, M. and Seed, T.M. (1980). Morphology of Plasmodia. In: Malaria, vol. 1 (Kreier, J.P. ed.) pp. 285-345. Academic Press, New York.,
Carlton JMR, Vinkenoog R, Waters AP, Walliker D (1998) Gene synteny in species of Plasmodium. Molecular and Biochemical Parasitology 93, 285-94
Chaitali Das, Rajlakshmi Poi, Ashim Chowdhury (2005) HPTLC determination of vasicine and vasicinone in Adhatoda vasica. Phytochemical Analysis 16, 90–92
Childs GE, Boudreau EF, Milhous WK, Wimonwattratee T, Pooyindee N, Pang L, Davidson DE (1984) Comparison of in vitro and in vivo antimalarial activities of 9-phenanthrenecarbinols. Annals of Tropical Medicine and Parasitology 78, 13–20
Cox F (1988) Major animal models in malaria research: rodent. In: Wernsdorfer, W.H., McGregor, I. (Eds.), Malaria Principles and Practice of Malariology, vol. 2. Churchill Livingstone, New York, pp. 1503–1543
Gibbons PL, Batty KT, Barrett PHR, Davis TME, Ilett KF (2007) Development of a pharmacodynamic model of murine malaria and antimalarial treatment with dihydroartemisinin. International Journal for Parasitology 37, 1569–1576
Haq, K, Ali M, Siddiqui AW. New compounds from the seeds of Embelia ribes Burm. Pharmazie 60: 69-71 (2005)
Homewood CA and Neame KD (1980) Biochemistry of malaria parasites. In: Malaria, vol. 1 (Kreier, J.P. ed.) Academic Press, New York.pp. 346-406. Academic Press, New York.
Jain, M.P., Koul, S.K., Dhar, K.L., Atal, C.K., 1980. Novel nor-harmal alkaloid from Adhatoda vasica. Phytochemistry 19, 1880–1882.
Janse CJ and Waters AP (1995) Plasmodium berghei: The Application of Cultivation and Purification Techniques to Molecular Studies of Malaria Parasites. Parasitiology Today 11 138 - 143
Janse CJ, Carlton JMR, Walliker D, Waters AP (1994) Conserved location of genes on polymorphic chromosomes of four species of malaria parasites Molecular Biochemical Parasitology 68, 285-96
Joshi BS, Bai Y, Puar MS, Dubose KK, Pelletier SW (1994) 'H- and 13C-NMR assignments for some pyrrolo[2,1b] quinazoline alkaloids of Adhatoda vasica. Journal of Natural Products 57, 953-962
Landau I and Gautret P (1998) Animal models: rodents. In: Sherman, I.W. (Ed.), Malaria: Parasite Biology, Pathogenesis and Protection. American Society for Microbiology, Washington, DC, pp. 401–417
Lin, L.H.M. et al. (2000) Int. J. Parasitol. 30, 357-370.) Lin LHM, Janse CJ, Waters AP (2000) The conserved genome organisation of non-falciparum malaria species: the need to know more. International Journal of Parasitology 30, 357-370
Lin, L.H.M. et al. (2001) Nuc. Acids Res. 15, 2059-68., Lin LHM, Pace T, Janse CJ, Birago C, Ramesar J, Picci L, Ponzi M, Waters AP (2001) Interspecies conservation of gene order and intron–exon structure in a genomic locus of high gene density and complexity in Plasmodium. Nucleic Acids Research 29, 2059-2068
Mackenstedt U, Brockelman CR, Mehlhorn H, Raether W (1989) Comparative morphology of human and animal malaria parasites. I. Host-parasite interface. Parasitology Research 75, 528–535
Peters W and Robinson BL (1999) Malaria. In: Zak, O., Sande, M. (Eds.), Handbook of Animal Models of Infection: Experimental Models in Antimicrobial Chemotherapy. Academic Press, London, pp. 757–773

Isolation of Vasicine, Vasicinone from Adhatoda zeylanica.........
89
Peters W, Portus JH, Robinson BL (1975) The chemotherapy of rodent malaria, XXII. The value of drug-resistant strains of Plasmodium berghei in screening for blood schizontocidal activity. Annals of Tropical Medicine and Parasitology 69, 155–171
Peters W, Robinson BL, Ellis DS (1987) The chemotherapy of rodent malaria. XLII. Halofantrine and halofantrine resistance. Annals of Tropical Medicine and Parasitology 81, 639–646
Peters W, Howells RE, Portus J, Robinson BL, Thomas S, Warhurst DC (1977) The chemotherapy of rodent malaria. XXVII. Studies on mefloquine (WR 142490). Annals of Tropical Medicine and Parasitology 71, 407-418
Posner GH, Paik Ik-Hyeon, Sur S, McRiner AJ, Borstnik Kristina, Xie S, Shapiro TA (2003) Orally active, antimalarial, anticancer, artemisinin-derived trioxane dimers with high stability and efficacy. Journal of Medicinal Chemistry 46, 1060–1065
Rich SM and Ayala FJ (2003) Progress in malaria research: the case for phylogenetics. Advances in Parasitology 54, 255-80
Sinden RE (1996) Infection of mosquitoes with rodent malaria. In: Molecular Biology of Insect Disease Vectors: A methods manual. Crampton, J.M, Beard, C.B. and Louis, C. eds. (London: Chapman and Hall), pp. 67-91
Sinden, R.E. (1978) Cell Biology. In: Rodent Malaria (R. Killick-Kendrick and W. Peters, eds.) Academic Press, London. pp 85-168.
Swamy HMK, Krishna V, Shankarmurthy K, Abdul Rahiman B, Mankani KL, Mahadevan KM, Harish BG, Raja Naika H (2007) Wound healing activity of embelin isolated from the ethanol extract of leaves of Embelia ribes Burm. Journal of Ethnopharmacology 109, 529–534
Vennerstrom JL, Dong Y, Andersen SL, Ager, Jr AL, Fu Hong-ning, Miller RE, Wesche DL (2000) Synthesis and antimalarial activity of sixteen dispiro-1,2,4, 5-tetraoxanes: alkyl-substituted 7,8,15,16-tetraoxadispiro[5.2.5.2]hexadecanes. Journal of Medicinal Chemistry 43, 2753–2758
Recommended