
ANALYTICAL BIOCHEMISTRY 73, 486-492 (1976)
Isoelectric Focusing of Brain Adenylate Cyclase
DOUGLAS J. FRANKS' AND DANIEL MALAMUD~
Shriners Burns Institute, Massachusetts General Hospital, Boston. Massachusetts 02114. and Department of Surgery, Harvard Medical School, Boston, Massachusetts 02115
Received August 15. 1975; accepted March 18. 1976
Mouse brain adenylate cyclase has been solubilized with Lubrol PX and separated by isoelectric focusing on polyacrylamide gels. The enzyme activity has been measured with a sensitive assay isolating cyclic AMP from Dowex and alumina columns. The technique allows a one-step analysis of this membrane enzyme from a heterogeneous sample within 6 hr.
A method is described for the separation of adenylate cyclase (EC 4.6.1.1.) from a solubilized mouse brain membrane preparation utilizing isoelectric focusing in polyacrylamide gels. Purification of adenylate cyclase from mammalian cell membranes has been difficult, as the enzyme rapidly loses activity (1,2). Since the adenylate cyclase from mammalian brain has a higher specific activity than other tissues, it has been used as the source of enzyme in a number of purification studies (l-3). After solubilization with Lubrol PX (4) the enzyme retains basal activity but may lose the ability to be stimulated by sodium fluoride (2). Estimations of the molecular weight of solubilized adenylate cyclase have been made using gel-filtration techniques (3,5), and recent work by Neer indicates that the enzyme could consist of more than one molecular species (6).
We report here the application of a highly sensitive adenylate cyclase assay, modified from the technique of Salomon et al. (7), to locate brain adenylate cyclase activity on polyacrylamide gels after isoelectric focusing. The assay can be performed on fragments of gel containing the enzyme within 6 hr of obtaining tissue.
MATERIALS
[cK-~~P]ATP, 3H-labeled cyclic AMP([3H]cAMP), and Omnifluor were obtained from New England Nuclear, and Dowex AG SOW-X4 (200-400 mesh) was from Bio-Rad. Neutral alumina (WN-3). ATP, CAMP, phos- phoenol-pyruvate, and pyruvate kinase were obtained from Sigma.
1 Present address: Clinical Research Institute of Montreal. 110 Pine Avenue West, Montreal. Quebec H2W IR7.
* All correspondence to Dr. Malamud at the Shriners Bums Institute.
486 Copyright 0 1976 by Academic Press. Inc. All rights of reproduction in any form reserved.

ISOFOCUSING OF ADENYLATE CYCLASE 487
METHODS
Whole brains obtained from male Charles River CD-l mice were homogenized in 0.25‘~ sucrose 10 mM Tris-HCl (pH 7.5) in a Teflon/ glass homogenizer (three strokes, 3000 rpm). The homogenate was centri- fuged at 20,OOOg for 10 min, and the pellet, containing little adenylate cyclase activity, was discarded. The supernatant was centrifuged at 200,OOOg for 30 min, and the resulting pellet was solubilized by stirring for 10 min at 0°C in a small volume of homogenizing medium containing 1% Lubrol PX (a gift of ICI America, Inc.). Particulate material was removed by centrifugation at 200,OOOg for 30 min.
Enzyme Assay
The adenylate cyclase assay medium contained 100 mM Tris- HCl (pH 7.4), 10 mM MgC&, 1 mM CAMP, 2.5 mM mercaptoethanol, 0.1 mM ATP, 0.5 @i of [(r-32P]ATP, 0.0075% bovine serum albumin, an ATP-regenerating system, consisting of 5.4 mM phospho-enolpyruvate and 4 pg of pyruvate kinase, and 50- 100 pg of enzyme protein in a total volume of 0.1 ml. All incubations were carried out in a nonshaking water bath for 10 min at 37°C. Enzyme activity was stopped by adding 0.1 ml of a solution containing 10 mM ATP and 1 mM [3H]cAMP (50,000 dpm). Protein was then denatured by boiling for 1 min; samples were di- luted to 0.6 ml with HzO, and CAMP was isolated. The assay conditions were optimized for the brain enzyme. An ATP concentration of 0.1 mM re- sulted in the maximum conversion of [32P]ATP to [32P]cAMP. Care was taken to ensure that the reaction was linear with protein concentra- tion and time of incubation under these conditions.
The method of White and Zenser (8) was originally used for separation of CAMP. Unfortunately, with [a-32P]ATP from New England Nuclear, blank values ranged from 200 to 700 cpm of 32P. The Dowex/alumina method described by Salomon et al. (7) reduced the blank value to levels that approached scintillation counter background. In our hands, the method as described only gave recoveries of applied CAMP of about 50%. We have instead used a “piggy back” method based on the Note Added in Proof in the paper by Salomon et al. (7). In our modifica- tion, the sample was added to a 5.0 x 0.5 cm column of Dowex (AG SOW-X4) resin, the column was washed with 2 ml of HzO, and the washings were discarded. Over 99% of the applied 32P was removed in this wash. The column was then placed over a 3.0 x 0.5-cm column of neutral alumina and washed with a further 4 ml of H,O. Because the eluate from the Dowex has a low pH (pH 3-4), CAMP is bound to the alumina and does not elute. The Dowex column was then removed, and CAMP was eluted from the alumina with 4 ml of Tris-HCl (50 mM, pH 7.5). The eluate was collected directly into a scintillation vial containing
488 FRANKS AND MALAMUD
10 ml of a scintillation fluid composed of 8 g of Omnifluor, 200 ml of ethanol, 500 ml ofTriton X-100, and 500 ml of xylene. Using this procedure the assay blank was 15-20 cpm. and 80-90% of the applied CAMP was recovered.
Isoelectric Focusing
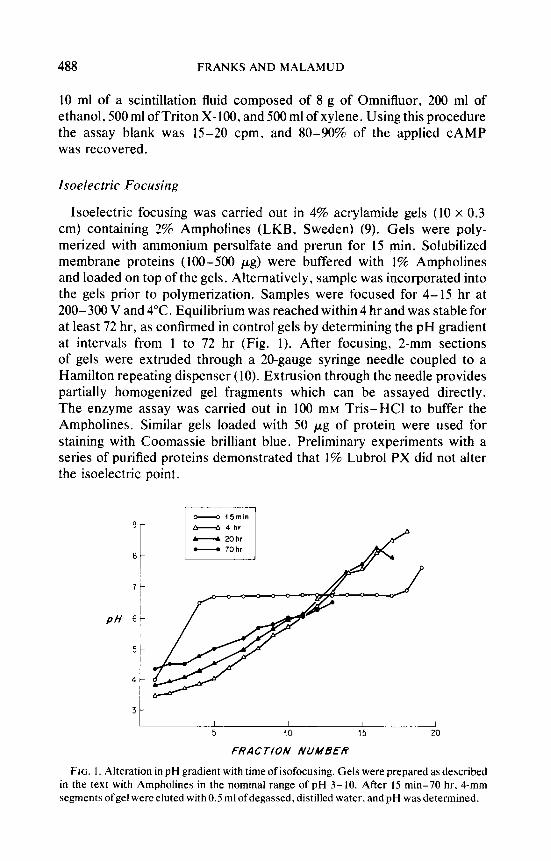
Isoelectric focusing was carried out in 4% acryiamide gels (10 x 0.3 cm) containing 2% Ampholines (LKB, Sweden) (9). Gels were poly- merized with ammonium persulfate and prerun for 15 min. Solubilized membrane proteins (100-500 pg) were buffered with 1% Ampholines and loaded on top of the gels. Alternatively. sample was incorporated into the gels prior to polymerization. Samples were focused for 4-15 hr at 200-300 V and 4°C. Equilibrium was reached within 4 hr and was stable for at least 72 hr, as confirmed in control gels by determining the pH gradient at intervals from 1 to 72 hr (Fig. 1). After focusing, 2-mm sections of gels were extruded through a 20-gauge syringe needle coupled to a Hamilton repeating dispenser (10). Extrusion through the needle provides partially homogenized gel fragments which can be assayed directly. The enzyme assay was carried out in 100 mM Tris-HCl to buffer the Ampholines. Similar gels loaded with 50 pg of protein were used for staining with Coomassie brilliant blue. Preliminary experiments with a series of purified proteins demonstrated that 1% Lubrol PX did not alter the isoelectric point.
PH
Y-
8-
6-
o--o (5min
I I I I 1 5 10 15 20
FRAC T/ON NUMBER
FIG. I. Alteration in pH gradient with time of isofocusing. Gels were prepared as described in the text with Ampholines in the nominal range of pH 3-10. After 15 min-70 hr, 4-mm segments ofgel were eluted with 0.5 ml ofdegassed. distilled water. and pH was determined.

ISOFOCUSING OF ADENYLATE CYCLASE 489
RESULTS AND DISCUSSION
Brain adenylate cyclase could be solubilized by either stirring or homog- enizing of the 200,OOOg pellet in 0.25~ sucrose, 10 mM Tris-HC1 (pH 7.5) containing 1% Lubrol PX. It was possible to solubilize 40% of the protein by this method. However, as shown in Table 1 the specific activity of adenylate cyclase in the soluble fraction was fourfold greater than that in the starting material, partly due to activation of enzyme activity by the detergent. Substantial adenylate cyclase activity remained “particu- late,” but the use of more vigorous methods of dispersion or higher Lubrol concentrations resulted in a reduction in the specific activity of the solubilized enzyme.
Enzyme activity was stimulated by 10e2 M NaF, but the degree of stimulation decreased as the enzyme was purified. Storage of the solu- bilized enzyme for 15 hr at 0°C resulted in a loss of 50% of basal activity and almost complete disappearance of NaF stimulation.
The effect of the presence of gel constituents on the activity of the solubilized enzyme was evaluated. As shown in Table 2 there was little effect of nonpolymerized acrylamide on adenylate cyclase activity. Ampholines at the concentrations present in enzyme preparations (1%) or gels (2%) had minima1 effects on adenylate cyclase activity. For determina- tion of recovery, enzyme activity on gels was compared to unfractionated enzyme preparations containing 1% Ampholines. The polymerizing agent ammonium persulfate had only a slight effect on enzyme activity at concentrations used to polymerize gels (0.04%). In top-loading experi- ments, gels were preisofocused for 15 min to remove ammonium per- sulfate. A sample of solubilized membranes containing 100-500 pg of protein was either top-loaded or incorporated into a gel and isofocused to equilibrium. Gel fragments were incubated in the reaction mixture and, after termination of the reaction, passed through an additional 5 x 0.5-
TABLE 1
SOLUBILIZATION OF MOUSE BRAIN ADENYLATE CYCLASE”
Basal Fluoride (10-2~)
Preparation (pmole CAMP producedimgi IO min)
Membrane preparation 187 t 4 844 k 66 Lubrol supematant 815 2 23 1646 k 51 Lubrol pellet 1852 It 202 3228 + 60 Lubrol supematant after storage for 15 hr at 0°C 366 + 12 458 2 59
LI Specific activity of adenylate cyclase in subcellular and solubilized fractions of mouse brain. Assay conditions are as described in Methods. Each value is the mean k SE of at least four determinations.
490 FRANKS AND MALAMUD
TABLE 2
EFFECT OF GEL CONSTITUENTS ON ADENYLATE CYCLASE ACTIVITY”
Enzyme assayed in presence of
Adenylate cyclase activity
(pmol/mgllO min) Percent
inhibition
No additions 848 ? 17 -
Acrylamide, 4% 837 -c I5 0
Ampholines. 1% 975 k 21 - 2% 604 k 36 30
Ammonium persulfate 0.04% 685 k I I I9 0.1% 527 k 8 38 0.4% 0 100
n Adenylate cyclase activity was assayed as described in Methods. Each value is the mean 2 SE of at least four determinations.
cm column containing only a glass wool plug. This step prevented clogging of the Dowex columns by the gel fragments.
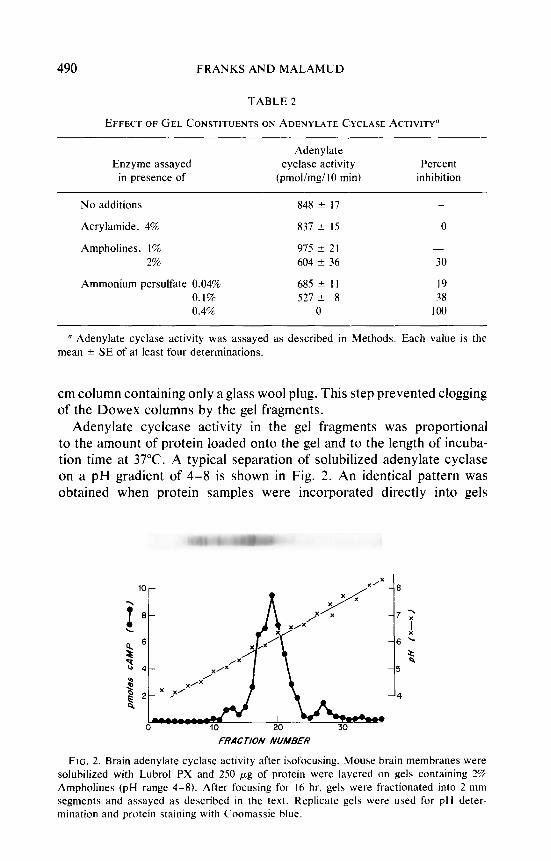
Adenylate cyclcase activity in the gel fragments was proportional to the amount of protein loaded onto the gel and to the length of incuba- tion time at 37°C. A typical separation of solubilized adenylate cyclase on a pH gradient of 4-8 is shown in Fig. 2. An identical pattern was obtained when protein samples were incorporated directly into gels
lo- / -6
FRACTION NUMBER
FIG. 2. Brain adenylate cyclase activity after isofocusing. Mouse brain membranes were solubilized with Lubrol PX and 250 pg of protein were layered on gels containing 2% Ampholines (pH range 4-8). After focusing for I6 hr. gels were fractionated into 2-mm segments and assayed as described in the text. Replicate gels were used for pH deter- mination and protein staining with Coomassie blue.

ISOFOCUSING OF ADENYLATE CYCLASE 491
before polymerization. Some experiments were also run with gels contain- ing 1% Lubrol. These gels were most difficult to fractionate, and there was no apparent requirement for addition of Lubrol to the gel. In a total of 16 experiments the enzyme had a pl of 5.9-6.1. As can be seen from the stained gel, adenylate cyclase is separated from most of the major protein bands. Attempts to determine the amount of protein associated with the enzyme peak were unsuccessful due to interference of the Ampholines with Lowry and biuret assays. In several experiments, Coomassie-stained gels were scanned at 600 nm to estimate the relative amount of protein associated with the enzyme peak. Such estimates suggest that 5- 15% of the applied protein migrates with adenylate cyclase activity.
An aliquot of the enzyme that was applied to the gel was stored at 0°C and assayed at the same time as the gel fractions. The activity of this prep- aration indicated that the enzyme in the gel could produce 104.4 pmol of CAMP. The actual recovered activity was 47.9, a recovery of 46%. In the many experiments performed, the recovery of adenylate cyclase activity was always less than 50% (mean 23 + 3%, rz = 16). The recovery was not improved by isoelectric focusing in the presence of dithiothreitol (1 mM), GTP (lO-4 M), MgCl, ( 10e2 M), and CaCI, (lO-5 M) or by assaying fractionated gels in the presence of the above compounds.
To assess the physical factors associated with assaying adenylate cyclase within a polymerized gel, samples were isofocused for 30 min. sufficient time for protein to enter the gel but not to separate into discrete components. Gels assayed at this time also showed recoveries of 20-30%. This suggests that the observed loss in enzyme activity may result from an underestimate of activity assayed in a polymerized gel. An alternative explanation, that a protein activator (11.12) is separated by isofocusing, would not explain the above results.
The techniques described here have allowed a rapid separation of adenylate cyclase from a heterogeneous preparation of solubilized mem- brane proteins. Since isoelectric focusing is an equilibrium method, large amounts of material may be loaded onto analytical sized gels. While it has not proven necessary to remove the gel or the detergent prior to the enzyme assay for analysis of adenylate cyclase activity on a preparative scale, it may be desirable to electrophorese the protein out of the gel to increase the recovery of enzyme activity.
ACKNOWLEDGMENTS
This study was supported by USPHS Grant GM 21700. The second author holds a National Institutes of Health Career Development Award,
REFERENCES
1. Sutherland, E. W., Rail. R. W., and Menon. T. (1962) J. Bid. Chem. 237, 1220- 1227. 2. Johnson, R. A., and Sutherland. E. W. (1973) J. Bid. Chew]. 248, 5114-5121.

492 FRANKS AND MALAMUD
3. Swislocki, N. I., and Tiemey, J. (1973) Biochemistry 12, 1862-1866. 4. Levey, G. S. (1970) Biochem. Biophys. Res. Commun. 38, 86-92. 5. Neer, E. J. (1973) J. Biol. Chem. 248, 3742-3744. 6. Neer, E. J. (1974) J. Biol. Chem. 249, 6527-6531. 7. Salomon, T., Londos, C., and Rodbell, M. (1974)Anal. Biochem. 58, 541-548. 8. White, A., and Zenser, T. (1971) Anal. Biochem. 41, 372-396. 9. Righetti, P., and Drysdale, J. W. (1971) Biochim. Biophys. Acta 236, 17-28.
10. Bagshaw, J. C., Drysdale, J. W., and Malt, R. A. (1973) Ann. N.Y. Acad. Sci. 209, 363-371.
11. Brostrom, C. O., Huang. Y. C.. Breckenridge, B. McL.. and Wolff, D. J. (1975) Proc. Nat. Acad. Sci. USA 72, 64-68.
12. Cheung, W. Y., Bradham, L. S., Lynch, T. J., Lin. Y. M., and Tallant, E. A. (1975) Biochem. Biophys. Res. Commun. 66, 1055-1062.
Recommended

![Development of Glucagon Sensitivity Neonatal Rat Liver · activity of ['3P]ATP. Protein was determined fluorometri-cally (18) and adenylate cyclase activity was expressed as picomoles](https://img.pdfslide.us/doc/110x75/5cc4ce6288c993474e8c3ac5/development-of-glucagon-sensitivity-neonatal-rat-liver-activity-of-3patp.jpg)















