
CSIRO PUBLISHING
www.publish.csiro.au/journals/ajz Australian Journal of Zoology, 2004, 52, 283–291
© CSIRO 2004 10.1071/ZO03051 0004-959X/04/030283
Helminth community structure in Rattus leucopus (Gray) (Muridae) fromAustralia, Papua New Guinea and Papua
L. R. SmalesA and D. M. SprattB
ASchool of Biological and Environmental Sciences, Central Queensland University, Rockhampton, Qld 4702, Australia.BCSIRO Sustainable Ecosystems, GPO Box 284, Canberra, ACT 2601, Australia.
Abstract
This study describes the helminth community in 28 Rattus leucopus, from Australia, Papua New Guineaand Papua (formerly Irian Jaya), and compares the community in Australian R. leucopus with those knownfrom two other Australian endemics, R. sordidus and R. fuscipes. In total, 28 parasite species were found:one acanthocephalan, five cestodes, one trematode and 21 nematodes as well as two lots of unidentifiedspirurid nematode larvae. The diversity of helminths in R. leucopus was high but equitable; the reciprocalof Simpson’s Index was 1.75. The distribution of prevalence of helminth species was not indicative of acore-satellite structure. No species occurred at a prevalence >30% and more than half the species occurredat prevalences of <10%. Only 10% of the helminth species identified were unique to R. leucopus. Helminthcommunity structure in R. leucopus differed from that observed in R. sordidus and the community in R.fuscipes was markedly different from those in R. leucopus and R. sordidus. Determinants of the diversity ofthe helminth community in R. leucopus include the time intervals between the three waves of migration ofRattus spp. to Australia, biogeographic factors related to immigration and adaptive radiation of the hostgenus, and associated with coevolution, speciation and host switching of the helminths.ZO03051Helmi nth par as it es of Rat tus l eucopusL. R. Sm al es and D. M . Spr att
Introduction
The genus Rattus is the only endemic genus of the cosmopolitan rodent subfamily Murinaerepresented in the Australasian region. Rattus leucopus (Gray) is one of only two species ofRattus that occur on both sides of the Torres Strait in Papua (formerly Irian Jaya) and PapuaNew Guinea, and in Australia. It is found in both continuous and fragmented rainforest ofCape York Peninsula (Leung 1999; Harrington et al. 2001) and on the southern slopes ofthe highlands, lowland rainforest blocks, gallery forest and garden areas of Papua andPapua New Guinea (Flannery 1995). R. leucopus forages in leaf litter for insects but willalso eat seeds, fruits and fungi (Leung 1999; Moore and Leung 1995).
There have been no surveys of the helminths of R. leucopus. Cribb and Pearson (1993)reported Neodiplostomum intermedium Pearson, 1959, a trematode, from Queensland andthis, together with two new species of nematode to be described elsewhere but associatedwith this study, are the only records of helminths to date.
This study was undertaken (1) to describe and compare the helminth communities ofR. leucopus from north-east Australia, New Guinea and Papua, (2) to compare them tohelminth communities known to occur in related Rattus spp. in other parts of Australia, and(3) to determine to what extent community differences are explained by current theories ofrat colonisation of the Australasian region.
Materials and Methods
Specimens were examined from a variety of sources and collected during 1917 (1 host), 1948 (1 host), 1960(3 hosts), 1968 (5 hosts), 1975 (1 host), 1981 (1 host), 1982 (5 hosts), 1984 (1 host), 1990 (7 hosts) and

284 Aust. J. Zoology L. R. Smales and D. M. Spratt
1992 (3 hosts). Some hosts had been examined previously for helminths and representative specimensdeposited in the Natural History Museum, London (NHM), the Queensland Museum, Brisbane (QM), theAustralian Museum, Sydney (AM), the CSIRO Wildlife Parasite Collection, Canberra (CSIRO) and theSouth Australian Museum, Adelaide (SAM). Material was also dissected from hosts that had been storedin 70% ethanol and held in either the Bernice Bishop Museum, Honolulu (BBM), or the AM. All thehelminths so examined provided data from 28 R. leucopus from the following localities: Cooktown,Queensland, 14°57′S, 145°21′E (1 host); East McIlwraith Range, Queensland, 13°45′S, 143°20′E (6 hosts);Danbulla State Forest, Queensland, 17°11′S, 153°01′E (7 hosts); a Dutch boat, Sydney Harbour, New SouthWales, 33°51′S, 151°10′E (1 host); Brown River, Papua New Guinea, 9°S, 147°E–148°E (6 hosts);Bensbach River, Papua New Guinea, 9°S, 141°E (2 hosts); Eramboe near Merauke, Papua, 7°30′S–8°40′S,140°E (3 hosts); Aru Island, Papua, 5°30′S–6°30′S, 134°E–135°E (2 hosts).
The fixation method for some of this material is unknown. Specimens from the Danbulla State Forestand the East McIlwraith Range were fixed in 10% formalin and stored in 70% ethanol, prior to examination.All other specimens were stored in 70% ethanol. Acanthocephalans and nematodes were cleared inBeechwood creosote or lactophenol prior to microscopic examination. Cestodes were stained in celestinblue and trematodes in Mayer’s haematoxylin, dehydrated in a graded series of ethanol, cleared in methylsalicylate and mounted in Canada balsam prior to microscopic examination. All helminths were identifiedto species level where possible.
Helminth community diversity was determined using the reciprocal of Simpson’s Index (1/SI), asemployed by Holmes and Podesta (1968). Prevalence of infection was used to characterise communitystructure according to the core and satellite species concept of Hanski (1982) and as interpreted by Bushand Holmes (1986) and Stock and Holmes (1987). Ecological terminology follows Bush et al. (1997). Forthe purposes of analyses, data from R. leucopus from Papua and Papua New Guinea were pooled becausethere are not marked differences in the habitat, climate and geography and because R. leucopus is typicallya rainforest and gallery forest species (O’Connor et al. 2002).
Results
Data on the prevalence of helminth infection in 28 R. leucopus collected from Australia andthe island of New Guinea, comprising one acanthocephalan, five cestode, one trematodeand 21 nematode species plus two lots of unidentified spirurid nematode larvae are givenin Table 1.
The acanthocephalan, from a single individual from the East McIlwraith Range, was inpoor condition and the fragments could not be identified. Only the cosmopolitan cestodeHymenolepis diminuta was found in both regions and was also recovered from theR. leucopus taken from the Dutch boat. Oligorchis sp. was found only in the rats fromDanbulla State Forest on the Atherton Tablelands and may represent a new species. Thistaxon has not been reported previously from Australia. All other cestodes from R. leucopushave been reported previously from Rattus fuscipes in Australia (see Smales 1997).
The trematode Neodiplostomum intermedium was found only in Australian R. leucopus.The nematodes Toxocara mackerrasae, Protospirura kaindiensis, spirurid juveniles,
Hepatojarakus malayae and Trichuris muris were found only in R. leucopus from NewGuinea. Strongyloides sp., Ascarops sp. larvae, Ascaropsinae gen. nov., sp. nov.,Mastophorus muris, Physaloptera sp., Rictularia mackerrasae, Abbreviata sp. larvae,encapsulated spirurid larvae, Hepatojarakus pycnofasciatus, Nippostrongylus braziliensis,Odilia emanuelae, and Eucoleus gastricus were found only in rats from Australia.Heterakis spumosa, Ophidascaris robertsi, Subulura andersoni, Syphacia sp. nov.,Nippostrongylus magnus, and Eucoleus sp. were found in R. leucopus from both Australiaand New Guinea.
The diversity of helminths occurring in R. leucopus, as measured by the reciprocal ofSimpson’s Index, was 1.75. Prevalence distribution of helminth species is given in Fig. 1;no species reached a prevalence >30%. and the prevalence of many species was <10%.

Helminth parasites of Rattus leucopus Aust. J. Zoology 285
Discussion
This study presents detailed information on helminth community structure in R. leucopus.It is, however, limited by the number of hosts collected at each locality and that data on theage classes of the host sample were not available. Given that the detection of Strongyloidesspp. is best done by scraping the mucosa of the intestine and examining this under high powerusing a dissecting microscope, that Eucoleus spp. are difficult to detect during dissectionand that only the gut was examined from hosts that had been stored in ethanol, these datarepresent a conservative estimate of the presence and prevalence of species comprising thehelminth community of R. leucopus. The overall number of parasite species recorded, 28species and two lots of unidentified spirurid nematode larvae from 28 host individuals, issimilar to the report of 50 parasite species from 82 Rattus fuscipes but markedly differentfrom the report of 11 species from 94 R. sordidus (Gould) (see Smales 1997).
Table 1. Helminth parasites of 28 Rattus leucopus (Gray) from Australia and New GuineaLocation: A = Australia, NG = island of New Guinea
Taxon Prevalence (%) Site in host Location
AcanthocephalaUnidentifiable fragments 3.6 Small intestine A
CestodaRaillietina celebensis (Janicki, 1902) 25 Small intestine AMonopylidium ratticola (Sanders, 1957) 10.7 Small intestine AHymenolepis diminuta (Rudolphi, 1819) 28.5 Small intestine A, NGOligorchis sp. 3.6 Small intestine ASpirometra sp. spargana 7.2 Subcutaneous tissue A
TrematodaNeodiplostomum intermedium Pearson, 1959 3.6 Small intestine A
NematodaAscaridida
Heterakis spumosa (Schneider, 1866) 28.5 Caecum, colon A, NGOphidascaris robertsi larvae Sprent, 1963 25 Subcutaneous tissue, stomach A, NGToxocara mackerrasae (Sprent, 1957) 10.7 Stomach NGSubulura andersoni (Cobbold, 1876) 21.4 Caecum, colon A, NG
OxyuridaSyphacia n.sp. 17.9 Caecum, colon A, NG
RhabditidaStrongyloides sp. 3.6 Small intestine A,
SpiruridaAscarops sp. larvae 3.6 Stomach A,Ascaropsinae gen. nov., sp. nov. 10.7 Stomach AMastophorus muris (Gmelin, 1790) 3.6 Stomach APhysaloptera sp. 3.6 Stomach AProtospirura kaindiensis Smales, 2001 10.7 Stomach NGRictularia mackerrasae Mawson, 1971 10.7 Stomach A,Abbreviata sp. larvae 3.6 Mesentery, tissues ASpirurid larvae (enscapulated) 3.6 Small intestine, stomach A,Spirurid juveniles 3.6 – NG
StrongylidaHepatojarakus pycnofasciatus Mawson, 1961 10.7 Small intestine AH. malayae Yeh, 1955 3.6 Small intestine NGNippostrongylus brasiliensis (Travassos, 1919) 3.6 Small intestine AN. magnus (Mawson, 1961) 7.2 Small intestine A, NGOdilia emanuelae (Mawson, 1961) 17.8 Small intestine A
EnoplidaEucoleus gastricus (Bancroft, 1893) 3.6 Stomach A,Eucoleus sp. 28.5 Stomach, small intestine A, NGTrichurus muris (Schrank, 1788) 3.6 Caecum NG
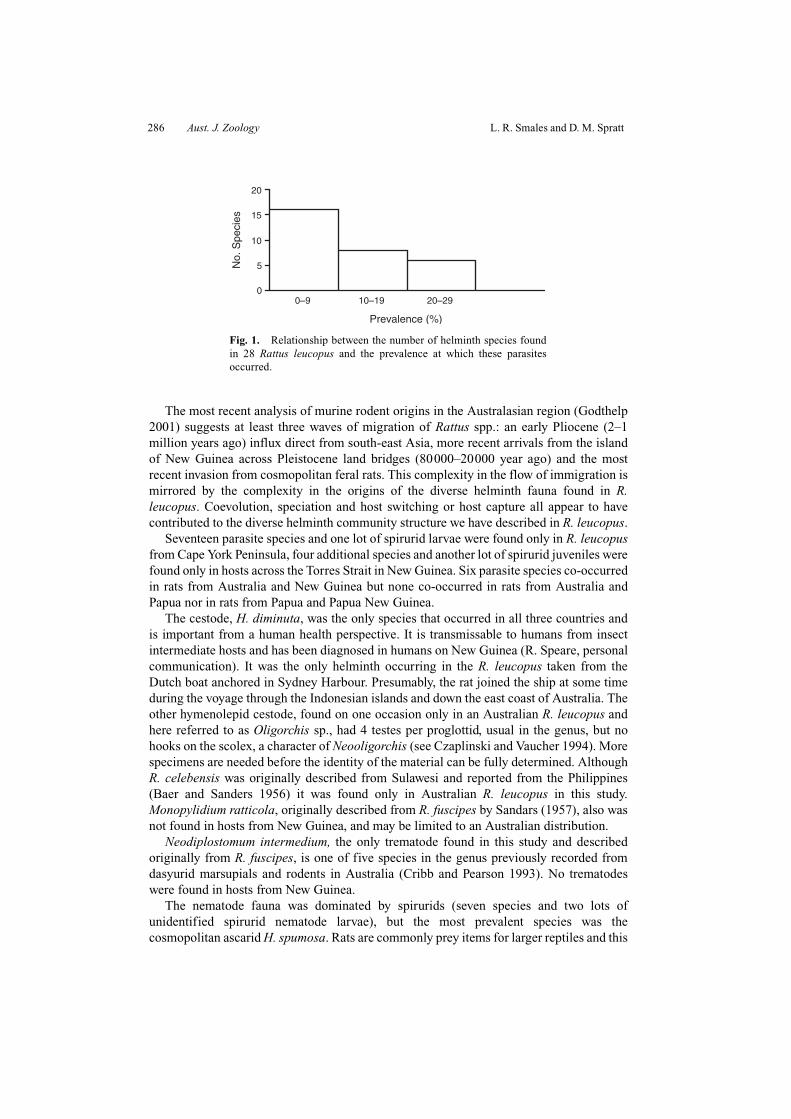
286 Aust. J. Zoology L. R. Smales and D. M. Spratt
The most recent analysis of murine rodent origins in the Australasian region (Godthelp2001) suggests at least three waves of migration of Rattus spp.: an early Pliocene (2–1million years ago) influx direct from south-east Asia, more recent arrivals from the islandof New Guinea across Pleistocene land bridges (80000–20000 year ago) and the mostrecent invasion from cosmopolitan feral rats. This complexity in the flow of immigration ismirrored by the complexity in the origins of the diverse helminth fauna found in R.leucopus. Coevolution, speciation and host switching or host capture all appear to havecontributed to the diverse helminth community structure we have described in R. leucopus.
Seventeen parasite species and one lot of spirurid larvae were found only in R. leucopusfrom Cape York Peninsula, four additional species and another lot of spirurid juveniles werefound only in hosts across the Torres Strait in New Guinea. Six parasite species co-occurredin rats from Australia and New Guinea but none co-occurred in rats from Australia andPapua nor in rats from Papua and Papua New Guinea.
The cestode, H. diminuta, was the only species that occurred in all three countries andis important from a human health perspective. It is transmissable to humans from insectintermediate hosts and has been diagnosed in humans on New Guinea (R. Speare, personalcommunication). It was the only helminth occurring in the R. leucopus taken from theDutch boat anchored in Sydney Harbour. Presumably, the rat joined the ship at some timeduring the voyage through the Indonesian islands and down the east coast of Australia. Theother hymenolepid cestode, found on one occasion only in an Australian R. leucopus andhere referred to as Oligorchis sp., had 4 testes per proglottid, usual in the genus, but nohooks on the scolex, a character of Neooligorchis (see Czaplinski and Vaucher 1994). Morespecimens are needed before the identity of the material can be fully determined. AlthoughR. celebensis was originally described from Sulawesi and reported from the Philippines(Baer and Sanders 1956) it was found only in Australian R. leucopus in this study.Monopylidium ratticola, originally described from R. fuscipes by Sandars (1957), also wasnot found in hosts from New Guinea, and may be limited to an Australian distribution.
Neodiplostomum intermedium, the only trematode found in this study and describedoriginally from R. fuscipes, is one of five species in the genus previously recorded fromdasyurid marsupials and rodents in Australia (Cribb and Pearson 1993). No trematodeswere found in hosts from New Guinea.
The nematode fauna was dominated by spirurids (seven species and two lots ofunidentified spirurid nematode larvae), but the most prevalent species was thecosmopolitan ascarid H. spumosa. Rats are commonly prey items for larger reptiles and this
0
5
10
15
20
0–9 10–19 20–29
Prevalence (%)
No.
Spe
cies
Fig. 1. Relationship between the number of helminth species foundin 28 Rattus leucopus and the prevalence at which these parasitesoccurred.

Helminth parasites of Rattus leucopus Aust. J. Zoology 287
is reflected in the presence of larval stages of O. robertsi, Abbreviata sp., and encapsulatedspirurid larvae. O. robertsi was previously thought to be endemic to Australian rodents butis here recorded from R. leucopus in both Australia and New Guinea. The other spiruridlarva found in this survey was the spirocercid, Ascarops sp. Dung beetles and othercoleopterans serve as intermediate hosts for species of Ascarops, and larvae have beenfound in a range of insectivorous vertebrates, including rodents, as accidental or paratenichosts (Anderson 2000). The definitive hosts usually are wild or domesticated Suidae butlagomorphs, guinea pigs and cattle have also been infected (Anderson 2000). Ascaropsstrongylina (Rudolphi, 1819) has been reported from domestic pigs in Queensland(Mackerras 1958).
An adult spirocercid was dissected from lesions in the stomach wall of one R. leucopusfrom Queensland. These nematodes were most similar to species of Leiuris, a neotropicalgenus occurring in sloths (Chabaud 1975), but sufficiently different to be considered a newgenus, to be described elsewhere. There is no simple explanation for the origins of the twospirocercids.
Rictularia mackerrasae was found only in Australian R. leucopus and was knownpreviously only from R. fuscipes near Innisfail in north Queensland. A second speciesoccurs in R. fuscipes on Pearson Island in South Australia and a third and possibly fourthspecies are known from Rattus villosissimus (Waite) in the Northern Territory (Smales1997).
Toxocara mackerrasae, described from R. fuscipes and also reported from Hydromyschrysogaster Geoffroy in Australia (Warren 1970), was found only in R. leucopus fromNew Guinea. The nematode may be present in Australian R. leucopus, but was simplymissed in this survey; alternatively, T. mackerrasae may be present in other host species inNew Guinea and only recently has host switching occurred and the parasite been ‘captured’(sensu Chabaud 1965) by R. leucopus.
The specimens of Physaloptera sp. were all females. They appeared most similar toP. troughtoni, described by Johnston and Mawson (1941) from R. fuscipes from KangarooIsland, South Australia. The same authors also described P. banfieldi from M. cervinipesfrom Dunk Island, North Queensland. Male specimens are needed before this identificationcan be confirmed.
The spirurid nematode Protospirura kaindiensis, previously known from New Guineabut not Australia, was not found in Australian R. leucopus The cosmopolitan spiruidMastophorus muris occupied the same niche in the stomach of R. leucopus from Australia.This suggests a more recent arrival for M. muris, travelling to the region with recentintroductions of Rattus spp. and evolving with endemic rodent species, as appears to be thecase with P. kaindiensis. The ascarid nematode Subulura andersoni has previously beenreported from India, Sri Lanka [Ceylon] (Thwaite 1927), Malaysia (Betterton 1979),Indonesia (Wiroreno 1978) and now both Papua New Guinea and Australia. Thisdistribution is congruent with the putative migration patterns of murid rodents fromsouth-east Asia (Godthelp 2001). It is not consistent with the presence of Subulura ortleppi,an African species from the four-striped grass mouse, Rhabdomys pumilio (Sparrman), andthe African soft-furred rat, Aethomys namaquensis (Smith), (as Praomys), in R. fuscipesfrom Pearson Island off the coast of South Australia (Mawson 1971). That author reportedthat her identification had been confirmed by Inglis, who had described this species. In hisobservations on the comparative morphology of the genus Subulura, Inglis (1960) indicatedthat prior to knowledge of the genus Subulura, species of the family Subuluridae Yorke &Maplestone, 1926, had been highly host specific. Since then, S. andersoni has been

288 Aust. J. Zoology L. R. Smales and D. M. Spratt
reported from a variety of rodent hosts (Ow Yang 1971; Singh and Chee-Hock 1971; Fonget al. 1977; Singh et al. 1987). Further examination of material from hosts across thedistribution of these species leading to a full revision of the genus may be timely.
The oxyurid nematode Syphacia sp. nov. appears to be specific to R. leucopus across thehost’s geographic range. The cosmopolitan S. muris, the Australian S. darwini and thePapua New Guinean S. longaecauda were not found in this study. The cosmopolitanStrongyloides ratti has been reported from R. sordidus from Ingham, Queensland (seeSmales 1992); however, there was insufficient material available to confirm the specificidentification of the Strongyloides sp. from R. leucopus. Similarly, Mackerras (1958) wasunable to identify to species material from a northern Queensland population of R. fuscipes.
The Heligmonellidae, usually dominant in the helminth community of Rattus spp. (seeObendorf 1979 for R. fuscipes; Smales 1992 for R. sordidus) were present but in a smallnumber of hosts. Odilia emanuelae, described from R. fuscipes and R. sordidus (seeMawson 1961), was the most prevalent species but occurred in only five host individuals.The genus Odilia was previously designated an Australian endemic genus (Durette-Desset1985), coevolving with the old endemic hosts, the Hydromyinae, and subsequentlyswitching hosts to the new endemic Rattus spp. (Gibbons and Spratt 1995). Species ofOdilia have since been described from Papua and Sulawesi, Indonesia (Hasegawa andSyafruddin 1994; Hasegawa et al. 1999) and recorded from Papua New Guinea (Smales2001). It is therefore now unclear where the ancestral Odilia species originated but therehave been foci of speciation in Indonesia, Papua New Guinea and Australia.
The genus Nippostrongylus was proposed by Durette-Desset et al. (1994) to be Asiaticin origin, reaching Australia with migrating Rattus spp. Two species, N. magnus andN. typicus, are known from Australian hosts (Smales 1997). Of these, N. magnus,previously thought to be endemic in Australian rodents, was found in R. leucopus fromAustralia and Papua New Guinea. The cosmopolitan species N. brasiliensis occurred at lowprevalence in Australian R. leucopus and was not present in R. leucopus from New Guinea.These host relationships and geographic distributions are supportive of host migrationacross Torres Strait from Australia to New Guinea being responsible for the presence of N.magnus in the latter country. On the other hand, N. brasiliensis may have arrived inAustralia during recent migrations of cosmopolitan Rattus spp., been captured byR. leucopus in Australian localities but not reached hosts in Papua New Guinea or Papua.
The molineid genus Hepatojarakus is an Australasian genus, H. pycnofasciatusoccurring in rodents in northern Australia (Smales 1997) and H. malayae in rodents inPapua, Java (Wiroreno 1978) and Malaya (Betterton 1979).
The cosmopolitan whip-worm, Trichuris muris, was found in a single R. leucopus fromPapua New Guinea and was known previously in Australia only from Hydromyschrysogaster (Smales 1997).
Enoplid nematodes of the genus Eucoleus (=Capillaria sensu lato) were present in 32%of hosts examined and overall were the most prevalent parasite group. They are small,difficult to find in their habitat in epithelial tissues, and may be overlooked duringparasitological examination, particularly of fixed tissues. Thus, their prevalence may beunderestimated in this study and may be closer to the 64% prevalence reported by Obendorf(1979) for E. gastricus in R. fuscipes in south-eastern Australia.
The diversity of the helminth community was high (28 species plus two lots ofunidentified spirurid nematode larvae found) but relatively equitable, as indicated by thediversity index (1/SI = 1.75). No single species, or group of species, dominated. This wassupported by an analysis of community structure: examination of prevalence classes did not

Helminth parasites of Rattus leucopus Aust. J. Zoology 289
reveal a core, dominant species or group of species. No species occurred at prevalence>30% and more than half the species occurred at prevalences of <10%. This communitystructure is neither isolationist, with a species-poor assemblage, nor interactive, withregular co-occurrences of large populations of many species, as defined by Stock andHolmes (1987). Further, it does not form a typical bimodal community, with a core andsatellite structure, core species having prevalences >60% (Holmes and Podesta 1968;Hanski 1982). Only four of the helminth species identified, namely Ascarops sp. larvae, thenew ascaropsine taxon, Oligorchis sp. and Syphacia sp. nov., are unique to R. leucopus, buteach of these should be classed as rare species occurring at low prevalences, rather than assatellite species occurring as occasional infections of the host species being studied (Stockand Holmes 1987). The remaining 20 species with low prevalences (<11%) could either bedefined as similarly rare in other host species, e.g. M. muris or as occasional infections ofspecies that occur in the satellite component of the helminth community of other hostspecies, e.g. N. magnus or O. emanuelae (see Obendorf 1979; Smales 1992).
In contrast, Rattus sordidus has a less diverse community of 8 species (1/SI = 2.2) and aunique species, Cyclodontostomum purvisi Adams, 1933 (see Smales 1992; Hasegawa andSyafruddin 1994). There were no core species, but a bimodal distribution, indicative of theheligmonellids N. typicus and O. emanuelae (42 and 44% prevalence respectively)representing the secondary species of Bush and Holmes (1986).
The Victorian population of R. fuscipes studied by Obendorf (1979), the only other studyfor which prevalence data are available, also had a diverse but equitable community(1/SI = 0.5), with only two unique species, Paraustrostrongylus ratti Obendorf, 1979 andCapillaria praeputialis Obendorf, 1979. In contrast to the helminth communities inR. leucopus and R. sordidus, the community structure in R. fuscipes was represented by twocore species (prevalence >60%), seven secondary species (prevalence 20–50%) and 11satellite species (prevalence <20%). This appears congruent with the description of aninteractive community by Stock and Holmes (1987).
Two determinants of the helminth community of R. leucopus may relate to the relativeuniformity and stable food supply of a rainforest habitat (Leung 1999) and the diversity ofdiet items available to the rats. The nematode parasites, for example, include monoxenousspecies (heligmonellids and heterakids), autoinfective species (oxyurids) and heteroxenousspecies (subulurids and ascarids). These two factors, together with biogeographic featuresrelated to immigration and adaptive radiation of the host genus, will have played a role inthe evolution of the diversity of the helminth community in R. leucopus.
Acknowledgements
Thanks are due to Mick Burt, Ian Mason and Peter Catling for collection of rats in northQueensland and Peter Haycock and Elizabeth Walter for assistance in post mortemexamination of these animals. The cooperation of Eileen Harris from the Natural HistoryMuseum, London, Carla Kisinami from the Bishop Museum, Honolulu, Ian Whittingtonfrom the South Australian Museum, Adelaide, and Tim Flannery from the AustralianMuseum, Sydney, in lending specimens is also gratefully acknowledged. Ken Aplin kindlyoffered advice concerning the suitability of pooling the data from samples from Papua andPapua New Guinea.
References
Anderson, R. C. (2000). ‘Nematode Parasites of Vertebrates. Their Development and Transmission.’ (CABIPublishing: Wallingford, UK.)

290 Aust. J. Zoology L. R. Smales and D. M. Spratt
Baer, J. G., and Sanders, D. F. (1956). The first record of Raillietina (Raillietina) celebensis (Janicki, 1902),(Cestoda) in man from Australia, with a critical survey of previous cases. Journal of Helminthology 30,179–182.
Betterton, C. (1979). The intestinal helminths of small mammals in the Malaysian tropical rainforest,patterns of parasitism with respect to lost ecology. International Journal for Parasitology 9, 313–320.doi:10.1016/0020-7519(79)90080-8
Bush, A. O., and Holmes, J. L. (1986). Intestinal helminths of lesser scaup ducks: patterns of association.Canadian Journal of Zoology 64, 132–141.
Bush, A. O., Lafferty, K. D., Lotz, J. M., and Shostak, A. W. (1997). Parasitology meets ecology on its ownterms: Margolis et al. revisited. Journal of Parasitology 83, 575–583.
Chabaud, A. G. (1965).Spécificité parasitaire. I. Chez les Nématodes Parasites des Vertébrés. In ‘Traité deZoologie. Anatomy, Sytématique, Biologie. Vol 4. Part 2. Némathelminthes (Nématodes)’. (Ed. P. P.Grassé.) pp. 548–564. (Masson: Paris.)
Chabaud, A. G. (1975). Keys to genera of the Order Spirurida. Part 2. Spiruroidea, Habronematoidea andAcuarioidea. In ‘CIH Keys to the Nematode Parasites of Vertebrates.’ (Eds R. C. Anderson, A. G.Chabaud and S. Willmott.) pp. 29–58. (CAB: Farnham Royal, UK.)
Cribb, T. H., and Pearson, J. C. (1993). Neodiplostomum spratti n. sp. (Digenea: Diplostomidae) fromAntechinus spp. (Marsupialia: Dasyuridae) in Australia, with notes on other diplostomids fromAustralian mammals. Systematic Parasitology 25, 25–35.
Czaplinski, B., and Vaucher, C. (1994). Family Hymenolepididae. In ‘Keys to the Cestode Parasites ofVertebrates’. (Eds L. F. Khalil, A. Jones and R. A. Bray.) pp. 595–664. (CAB International: Wallingford,UK.)
Durette-Desset, M.-Cl. (1985). Trichostrongyloid nematodes and their vertebrate hosts: reconstruction ofthe phylogeny of a parasitic group. Advances in Parasitology 24, 239–306.
Durette-Desset, M. C., Beveridge, I., and Spratt, D. M. (1994). The origins and evolutionary expansion ofthe Strongylida (Nematoda). International Journal for Parasitology 24, 1139–1165. doi:10.1016/0020-7519(94)90188-0
Flannery, T. (1995). ‘The Mammals of New Guinea.’ (Reed Books: Sydney.)Fong, Y. P., Liat, L. B., de Witt, G. F., Krishnasamy, M., Sivanandam, S., and Foong, P. Y. K. (1977).
Protozoan and helminth parasites of small wild mammals in a new FELDA settlement Jenderak Utara,Central Pahang, peninsular Malaysia. Southeast Asian Journal of Tropical Medicine and Public Health8, 345–353.
Gibbons, L. M., and Spratt, D. M. (1995). Two new species of Odilia (Nematoda: Heligmonellidae) fromAustralian rodents, with comments on O. bainae Beveridge & Durette-Desset 1992. SystematicParasitology 31, 67–79.
Godthelp, H. (2001). The Australian rodent fauna, flotilla’s flotsam or just fleet footed? In ‘Faunal andFloral Migrations and Evolution in S.E. Asia–Australasia’. (Eds I. Metcalfe, J. M. B. Smith, M.Morwood, I. Davidson.) pp. 319–322. (A.A.Balkema: Lisse.)
Hanski, J. (1982). Dynamics of regional distribution: the core and satellite species hypothesis. Oikos 38,210–221.
Harrington, G. N., Freeman, N. D., and Crome, F. H. J. (2001). The effects of fragmentation of an Australiantropical rainforest on populations and assemblages of small mammals. Journal of Tropical Ecology 17,225–240. doi:10.1017/S0266467401001158
Hasegawa, H., and Syafruddin. (1994) Odilia mallomys sp. n. (Nematoda: Heligmonellidae) fromMallomys rothschildi weylandi (Rodentia: Muridae) of Irian Jaya, Indonesia. Journal of theHelminthological Society of Washington 61, 208–214.
Hasegawa, H., Miyata, A., and Syafruddin. (1999). Six new nematodes of the Heligmonellidae(Trichostrongylina) collected from endemic murines of Sulawesi, Indonesia. Journal of Parasitology 85,513–524. doi:10.1007/S004360050589
Holmes, J. C., and Podesta, R. (1968). The helminths of wolves and coyotes from the forested regions ofAlberta. Canadian Journal of Zoology 46, 1193–1203.
Inglis, W. G. (1960). Further observations on the comparative anatomy of the head of the nematode familySubuluridae with the description of a new species. Proceedings of the Royal Society, London 135,125–136.
Johnston, T. H. J., and Mawson, P. M. (1941). Some parasitic nematodes in the collection of the AustralianMuseum. Records of the Australian Museum 21, 9–16.

Helminth parasites of Rattus leucopus Aust. J. Zoology 291
http://www.publish.csiro.au/journals/ajz
Leung, L. K. (1999). Ecology of Australian tropical rainforest mammals. III. The Cape York rat Rattusleucopus (Muridae: Rodentia). Wildlife Research 26, 317–328.
Mackerras, M. J. (1958). Catalogue of Australian mammals and their recorded internal parasites.Proceedings of the Linnean Society of New South Wales 83, 101–160.
Mawson, P. M. (1961). Trichostrongyles from rodents in Queensland, with comments on the genusLongistriata (Nematoda: Heligmosomatidae). Australian Journal of Zoology 9, 791–826.
Mawson, P. M. (1971). Pearson Island Expedition 1969. 8. Helminths. Transactions of the Royal Society ofSouth Australia 95, 169–183.
Moore, L. A., and Leung, L. K. (1995). Cape York rat. Rattus leucopus. In ‘The Mammals of Australia’.(Ed. R. Strahan.) pp. 653–654. (Reed Books: Sydney.)
Obendorf, D. L. (1979). The helminth parasites of Rattus fuscipes (Waterhouse) from Victoria, includingdescription of two new nematode species. Australian Journal of Zoology 27, 867–879.
O’Connor, S., Aplin, K. P., Spriggs, M., Veth, P., and Ayliffe, L. K. (2002). From savanna to rainforest:changing environments and human occupation at Liang Lemdubu, Aru Islands, Maluku (Indonesia). In‘Bridging Wallace’s Line: The Environmental and Cultural History and Dynamics of the SEAsian–Australian Region’. (Eds P. Kershaw, B. David, N. Tapper, D. Penny and J. Brown.). pp. 279–306.Advances in Geoecology Series, No. 34. (Catena Verlag: Reiskirchen.)
Ow-Yang, C. K. (1971). Studies on the nematode parasites of Malaysian rodents. 1. The Rhabdiasidea,Trichuridea and Oxyuridea. Journal of Helminthology 45, 93–109.
Sandars, D. F. (1957). Cestoda from Rattus assimilis (Gould, 1858) from Australia. Journal ofHelminthology 31, 65–78.
Singh, M., and Chee-Hock, C. (1971). On a collection of nematode parasites from Malayan rats. SoutheastAsian Journal of Tropical Medicine and Public Health 2, 516–522.
Singh, K. I., Krishnasamy, M., and Ming, H. T. (1987). Collection of small vertebrates and theirendoparasites from Ulu Endau, Johore, Malaysia. Malaysian Nature Journal 41, 349–356.
Smales, L. R. (1992). A survey of the helminths of Rattus sordidus, the canefield rat, together with adescription of Ancistronema coronatum n.g., n. sp. (Nematoda: Chabertiidae). Systematic Parasitology22, 73–80.
Smales, L. R. (1997). A review of the helminth parasites of Australian rodents. Australian Journal ofZoology 45, 505–521.
Smales, L. R. (2001). Protospirura kainiensis n. sp. (Spirura: Spiruridae) and other helminths fromPseudohydromys (Muridae: Hydromyinae) from Papua New Guinea. Journal of Parasitology 87,169–172.
Smales, L. R., and Cribb, T. H. (1997). Helminth parasite communities of the water-rat Hydromyschrysogaster. Wildlife Research 24, 445–457.
Stock, T. M., and Holmes, J. C. (1987). Host specificity and exchange of intestinal helminths among fourspecies of grebes (Podicipididae). Canadian Journal of Zoology 65, 669–676.
Thwaite, J. W. (1927). On a collection of nematodes from Ceylon. Annals of Tropical Medicine andParasitology 21, 225–244.
Warren, E. G. (1970). Observations on the life-cycle of Toxocara mackerrasae. Parasitology 60, 239–253.Wiroreno, W. (1978). Nematode parasites in West Java, Indonesia. Southeast Asian Journal of Tropical
Medicine and Public Health 9, 520–525.
Manuscript received 22 September 2003; accepted 20 February 2004
Recommended

















