
GINKGOALES
The order Ginkgoales is today represented by only one living member, i.e. Ginkgo biloba.
Ginkgoales was, however, very abundantly represented in the world by several species of
about 16 genera during the Triassic period of Mesozoic age, i.e. about 200,000,000 years ago.
Today, all the genera, except Ginkgo biloba, are extinct.
G. biloba is represented by five varieties viz. Ginkgo biloba var. aurea (Nelson) Beisson,
G.biloba var. fastigata Henry, G.biloba var. paciniata Carriere, G. biloba var. pendula Carnere
and G. biloba var. variegata Carriere. Due to the presence of a number of primitive
characters, as well as because of its long geological records, Ginkgo is called a “living
fossil”.
Details of the geological history of Ginkgoales indicate that its members started appearing on
the earth during Permian, achieved luxuriance and worldwide distribution during Triassic and
Jurassic periods of Mesozoic age, started fading out of existence during Cretaceous and now
represented only in some parts of Southern and Eastern China by only one living member i.e.
Ginkgo biloba.
Ramanujam (1976), while tracing the geological history of Ginkgoales, reported that a few
records of this order are known from Late Palaeozoic of India. These include Ginkgophyton,
Psygmophyllum and Rhipidopsis.
Seward (1938) considered Ginkgo “as one of the wonders of the world” and stated that has
persisted with little change until the present through a long succession of ages when the earth
was inhabited by animals and plants”. Arnold (1947) mentioned that “Ginkgo biloba is one of
the oldest living plants and may indeed be the oldest living genus of the seed plants”

Distribution of Ginkgo:
Ginkgo biloba is a tall slender and beautiful tree. It is commonly called Maiden-hair Tree
because its new leaves resemble very much like those of Adiantum (called maiden hair fern)
both in form and venation. Ginkgo still exists in the wild state in South-eastern China, along
“the north western border of Chekiang and south eastern Anhwei”. In China and Japan it is
grown as a sacred tree in temple gardens. It is cultivated in the United States as a shade tree.
It is also successfully cultivated in some gardens of Europe, America and India.
1. Tall, well-branched trees with short and long shoots. However, some earliest fossil
members were without short and long shoots.
2. Wood is pycnoxylic.
3. Leaves are large, leathery and fan-shaped or strap-shaped. They are often deeply divided.
4. Dichotomous venation is usually present in the leaves.
5. Un-branched, catkin-like male organs are axillary in position.
6. Male organs bear micro-sporangiophores.
7. Each micro-sporangiophore possesses 2-12 pendulous microsporangia.
8. Spermatozoids are motile and contain spiral bands of flagella.
9. Ovules are terminal in position on branched or un-branched axillary axes. They are 2-10 in
number.
10. Seeds are large-sized.
11. Each seed contains a fleshy outer layer and a middle stony layer.
Morphological Features of Ginkgo:
The plant body of Ginkgo biloba is sporophytic, and the sporophyte resembles several
conifers in general habit. The trees have a pronounced ex-current habit of growth and attain a
height up to 30 metres. A very irregular pattern of branching is shown by Ginkgo trees.
The branches are dimorphic i.e. bear long shoots which are of unlimited growth with
scattered leaves and dwarf shoots which are short branches of limited growth.
Long shoots elongate rapidly, sometimes as much as 50 cm in a year. Dwarf shoots grow
rather slowly. A dwarf shoot of 2-3 cm length may be several years old. Sometimes there is
no clear-cut distinction between two types of shoots, and the dwarf shoot may convert into a
long shoot while the latter may sometimes convert into a dwarf shoot for a year or two and
then revert back into a long shoot.

The foliage leaves, present on the long shoots, are deeply lobed while those on the dwarf
shoots are not so deeply lobed and sometimes more or less entire (Figs. 10.4, 10.5). Bierhorst
(1971) opined that differences in the growth pattern of long and dwarf shoots may be due to
the quantities of auxin produced in the apical meristems.
Ginkgo biloba possesses a long tap root system. The roots are extensively branched and
penetrate deep into the soil. The foliage leaves are simple,large, petiolate and wedge-shaped
or fan-shaped with expanded apex and narrow base.
They resemble Maiden-hair fern (Adiantum) and by this they can be distinguished among
gymnosperms In general, the leaves are bilobed, and hence the name ‘biloba’ was suggested
to the species by Linnaeus (1771).
However, the leaves with many lobes (Fig 10.6) are also present invariably on the tree.The
leaves of the long shoot and seedlings are deeply bilobed while that of the dwarf shoot are not
so deeply lobed and may be sinuate or even entire. They may be pale yellow, golden yellow
or dark green in colour. A typical dichotomous type of venation (Fig. 10.7) is present in the
leaves.

Foliar epidermis also exhibits some distinguishable characters in Ginkgo biloba. The
epidermal cells are rectangular over the veins while polygonal in outline in between the
veins. The leaves are hypostomatic (i.e. bear stomata only on the lower surface of the leaf).
Kanis and Karstens (1963), however, observed a few stomata on the upper surface of the
leaves on long shoot, thus making them amphistomatic. Stomata remain surrounded by 4-6
subsidiary cells with finger-like outgrowths overarching the guard cells.
5. Anatomy of Ginkgo:
(i) Root:
In transverse section (Fig. 10.8) the roots are somewhat circular in outline. Mature roots are
surrounded by phellogen or suberized cells of cortex. A large portion of the young root is
occupied by multilayered, thin-walled cortex which contains several tannin- filled cells and
calcium oxalate crystals. Mucilage canals are also prominently visible (Fig. 10.8)
In young roots, a layer of endodermis and uni-layered pericycle are clear. Mature
roots,however, lack such a distinction. Diarch condition is clearly visible in the young roots.

Xylem is exarch. It remains separated by the phloem strands. Sometimes the roots also show
triarch condition.
(ii) Stem:
The young stem (long shoot) is more or less circular in outline and remains surrounded by a
single-layered, thickly circularized epidermis (Fig. 10.9) made of brick-shaped cells.
Epidermis is replaced by periderm in the older stems.
Inner to the epidermal layer is present a well-marked region of parenchymatous cortex. It
contains mucilaginous canals, sphaeraphides and many tannin-filled cells. Cortex is
comparatively less extensive in long shoots than dwarf shoots. Endodermis and pericycle are
not well-marked in long shoot.
Several conjoint, collateral, open and endarch vascular bundles are arranged in a ring in very
young stem. Two leaf traces, one for each leaf, are given out. After the onset of the secondary
growth, the vascular cylinder of the stem becomes an endarch siphonostele with no
parenchyma in the wood except that of uniseriate medullary rays.
Protoxylem has spiral thickenings while bordered pits are present on the radial walls of the
metaxylem tracheids. Sieve tubes and phloem parenchyma constitute the phloem. A narrow
pith, containing mucilage canals and sphaeraphides, is present in the centre of long shoot,
while in dwarf shoot the pith is comparatively more extensive.

Secondary Growth in Stem:
Cork cambium cuts periderm towards outer side and secondary cortex towards inner side
(Fig. 10.10) Periderm replaces the epidermis. Mucilage canals are absent.
A single ring of cambium remains active throughout and cuts secondary phloem towards
outer side and secondary xylem towards inner side. Crushed patches of primary phloem
towards outer side and primary xylem towards inner side are present. Secondary phloem
consists of sieve tubes and phloem parenchyma. Secondary xylem consists of tracheids. Ill-
defined annual rings are also seen.
Study of the tangential longitudinal sections of long and dwarf shoots shows that uniseriate
medullary’ rays are 1 -3 cells in height in long shoot (Fig. 10.11) while 1-15 cells in height in
dwarf shoot (Fig. 10.12).
One or two rows of bordered pits are present on the radial walls of the tracheids (Fig. 10.13).
Pits are circular in outline and have a clear torus. Bars of Sanio are also present. Bars of

Sanio do not occur in primary wood. Trabeculae of Sanio, which cross the lumen of the
tracheids, are also present.
(iii) Leaf:
A layer of epidermis is present on upper as well as lower sides of leaf. The epidermis is
thickly cuticularised and consists of rectangular to polygonal cells. Haplocheilic type of
stomata, restricted only to the lower epidermis, are present. Kanis and Karstens (1963),
however, reported some stomata on the upper epidermis of the leaves on long shoots of male
plants only.
Mesophyll is present in between the two epidermal layers. It is not well-differentiated into
palisade and spongy parenchyma (Fig. 10.14). The leaves of long shoot, however, show a
distinct palisade region. Many mucilage canals or secretory canals and a few tannin-filled
cells are also present in the mesophyll region.

Vascular bundles of the vein have an indistinct mesarch structure (Fig. 10.14). Each vascular
bundle may be surrounded by a sclerenchymatous bundle sheath in mature leaves.
(iv) Petiole:
The petiole remains covered by a layer of thickly cuticulanzed epidermis, the continuity of
which is broken by stomata. Inner to the epidermis are present a few hypodermal layers. Few
mucilage canals, tannin-filled cells and sphaeraphides are irregularly distributed in the cortex.
The petiole is supplied by a pair of endarch vascular bundles (Fig. 10.15).
The detailed structure of a vascular bundle is shown in Fig. 10.16. It remains surrounded by a
sclerenchymatous bundle sheath. Protoxylem has spiral thickenings. Cambium is clearly
visible Uniseriate rays are present in the xylem. These are continuous with those of the
phloem.
6. Reproductive Structures of Ginkgo:
Ginkgo biloba is dioecious. Male and female plants are,however, difficult to be differentiated,
when young. According to Lee (1954) sex in Ginkgo is determined by sex chromosomes (XY
in male and XX in female). Reproductive bodies of Ginkgo are most primitive among living
seed plants except some Cycadales.
(i) Male Strobilus:
Male or microsporangiate fructifications develop in catkin-like clusters (Fig. 10.17 A) on the
dwarf shoots of male frees.Each male strobilus contains several microsporophyll’s arranged
loosely on a central axis (Fig. 10.17B). Each microsporophyll has a long stalk terminating
into a hump or knob.
It contains two pendant microsporangia (Fig. 10.17C). According to some workers this
terminal knob represents an abortive sporangium. A mucilage duct is present in the knob.
Rarely more than two sporangia are present in a microsporophyll.

Each sporangium is a tubular structure surrounded by many layers (Fig. 10.17D). The
outermost layer of sporangial tissue differentiates into a tapetum. The sporogenous cells of
the sporangium undergo reduction division and form many haploid microspores Wolniak
(1976) has studied the ultrastructure of the microspore mother cell.
(ii) Development of Microsporangium:
It is of eusporangiate type, i.e., single archesporial cell divides by a periclinal wall forming
primary wall cell and primary’ sporogenous cell (Fig. 10.18A,B). The former develops into
wall of microsporangium while the latter develops into sporogenous tissue. Sporangium
dehisces by means of a longitudinal slit.

(iii) Female Strobilus:
The female strobili develop in groups in the axil of leaves or scaly leaves present on the
dwarf shoots (Fig. 10.19). These are much reduced structures, each having a long stalk or
peduncle, which bifurcates apically. Each bifurcation usually bears a single ovule. Out of the
two sessile ovules one generally aborts earlier.
Rarely, three, four or more ovules on a peduncle are also observed. Sprecher (1907) reported
an axis with seven ovules, each borne singly on its independent peduncle, and not in pairs. At
the base of each ovule is present a fleshy cup-like structure called ring or collar. The leaves
surrounding the ovules do not show their bilobed character.
The development of ovule, mega-sporogenesis and structure of the mature ovule is similar to
that of Cycas. There is a thick integument consisting of three layers, i.e., outer fleshy, middle
stony and inner fleshy layers (Fig. 10.20). A major difference between Ginkgo and Cycas is
that the integuments of Ginkgo do not have any noticeable vascular supply.
Each ovule has a large and prominent nucellus. Free apex of the nucellus breaks down into a
pollen chamber (Fig. 10.21) forming the nucellar beak. Quite deep in the nucellus tissue, a
functional spore mother cell becomes prominent. The spore mother cell develops into a
tetrad, of which only the innermost megaspore remains functional and develops into female
gametophyte.
The vascular supply of the ovule is ill-developed. Two vascular strands enter the inner fleshy
layer and reach up to the free part of the nucellus without branching. Outer fleshy layer lacks
any prominent vascular supply.
7. Gametophytes in Ginkgo:
(i) Male Gametophyte:
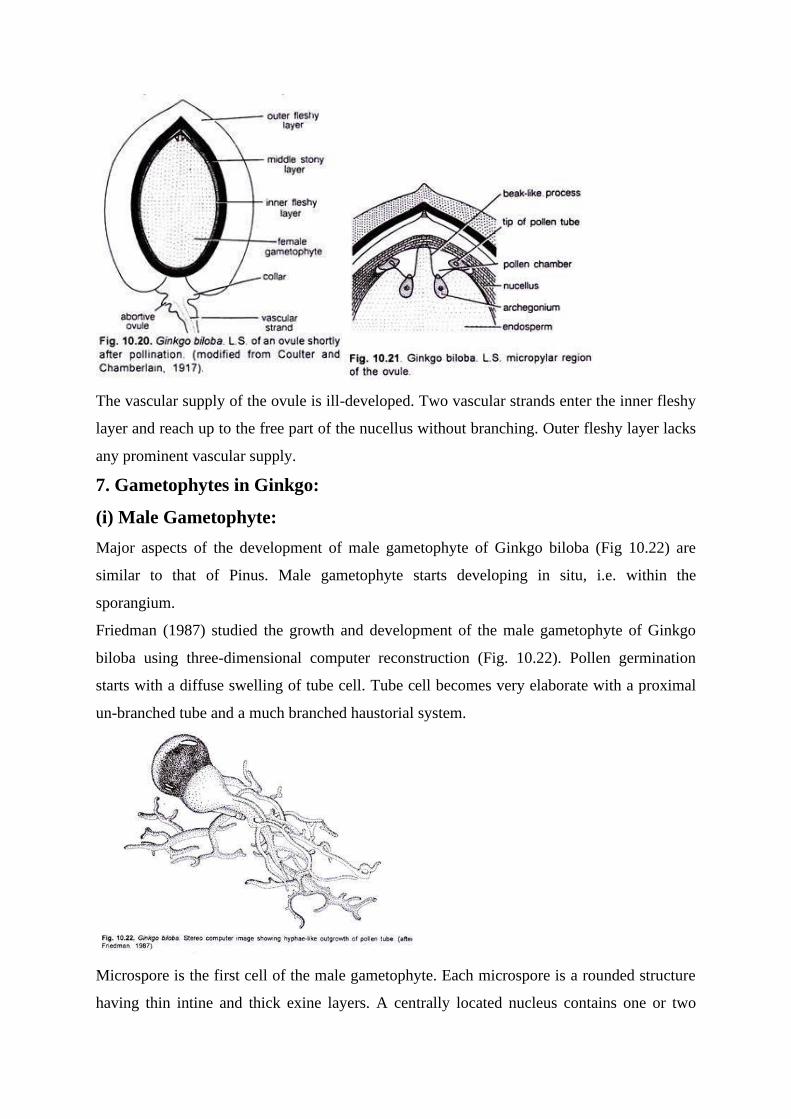
Major aspects of the development of male gametophyte of Ginkgo biloba (Fig 10.22) are
similar to that of Pinus. Male gametophyte starts developing in situ, i.e. within the
sporangium.
Friedman (1987) studied the growth and development of the male gametophyte of Ginkgo
biloba using three-dimensional computer reconstruction (Fig. 10.22). Pollen germination
starts with a diffuse swelling of tube cell. Tube cell becomes very elaborate with a proximal
un-branched tube and a much branched haustorial system.
Microspore is the first cell of the male gametophyte. Each microspore is a rounded structure
having thin intine and thick exine layers. A centrally located nucleus contains one or two
nucleoli and remains surrounded by dense cytoplasm. An un-thickened portion, called pore,
is also present in each microspore. It is a region where exine is not covering the intine.
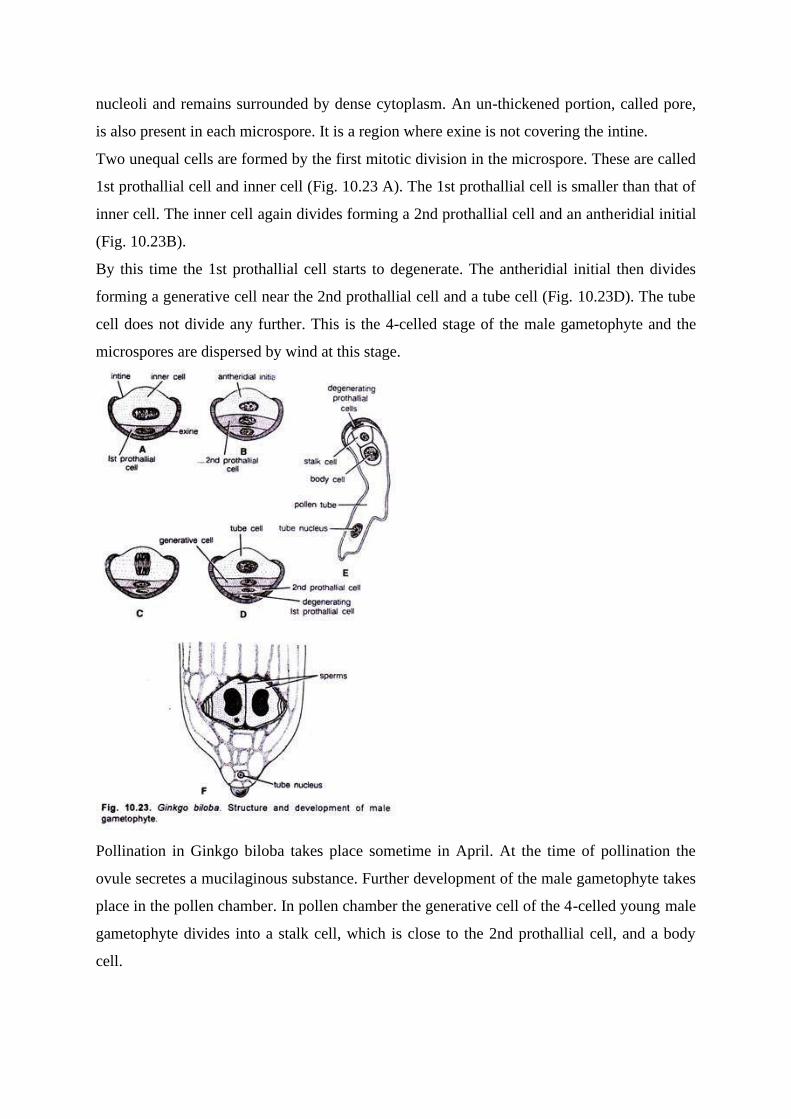
Two unequal cells are formed by the first mitotic division in the microspore. These are called
1st prothallial cell and inner cell (Fig. 10.23 A). The 1st prothallial cell is smaller than that of
inner cell. The inner cell again divides forming a 2nd prothallial cell and an antheridial initial
(Fig. 10.23B).
By this time the 1st prothallial cell starts to degenerate. The antheridial initial then divides
forming a generative cell near the 2nd prothallial cell and a tube cell (Fig. 10.23D). The tube
cell does not divide any further. This is the 4-celled stage of the male gametophyte and the
microspores are dispersed by wind at this stage.
Pollination in Ginkgo biloba takes place sometime in April. At the time of pollination the
ovule secretes a mucilaginous substance. Further development of the male gametophyte takes
place in the pollen chamber. In pollen chamber the generative cell of the 4-celled young male
gametophyte divides into a stalk cell, which is close to the 2nd prothallial cell, and a body
cell.

Now the intine protrudes out near the pore and functions as a pollen tube (Fig. 10.23E). The
pollen rube swells up and advances towards the archegonia by protruding into the nucellus.
Just before fertilization the body cells divides into two, and the wall formation is at right
angle to the base of pollen grains.
The blepharoplasts appear on the body of these so-formed two cells which function as sperms
or male gametes (Fig. 10.23F). The structure of these multi-flagellated sperms, as well as
their behaviour in the tube at the time of fertilization has been investigated by
Shimamura(1937), Lee (1955) and Favre-Duchartre (1956).
Motile sperms in gymnosperms were observed for the first time on September 9, 1896 by
Hirase, and his this discovery is considered as one of the most remarkable events in plant
morphology. The ultrastructure of male gamete of Ginkgo has been studied by Gifford and
Lin (1975).
(ii) Female Gametophyte:
The development of the ovule and female gametophyte in Ginkgo biloba has been studied by
Carothers (1907). Out of the four megaspores formed from the megaspore mother cell, only
the lowermost remains functional and the remaining three degenerate (Fig. 10.24) A).
The nucleus of the functional megaspore migrates towards the micropylar end and divides
into two followed by a number of free-nuclear divisions forming hundreds of free nuclei.
A large vacuole is present in the megaspore at this stage (Fig 10.24B). Now the wall
formation starts (Fig. 10.24B,C) and progresses from the periphery towards the centre. Dunng
wall formation first the anticlinal walls are formed followed by the vertical walls.
Each cell generally contains one nucleus but in some cells 2-3 nuclei are also seen. Due to
more rapid cell divisions on the micropylar end a pole-like structure develops. This is called
tent-pole (Fig. 10.24D). The female gametophyte possesses abundant chlorophyll.

The development of archegonium starts from the cells towards the micropylar end of the
female gametophyte. According to Favre-Duchartre (1958) the archegonial initials start
appearing sometime in June.
Each archegonium possesses a short neck made up of only four cells and a small venter
having a central cell. The central cell later on forms a ventral canal cell and an egg cell (Fig.
10.25). According to Bierhorst (1971) only two cells are present in the archegonial neck.
Ultra structural study of the female gametophyte of Ginkgo biloba has been made by
Dexheimer (1973). He observed four zones based on food reserves.
These are:
(1) Lipid zone,
(2) Starch proteolipid zone with large vacuolated cells,
(3) Starch zone made of large vacuolated cells with peripheral cytoplasm, and
(4) Deep or central zone with very little reserve contents.
Fertilization:
The pollen tube reaches up to the neck of the archegonium just after its (archegonium)
differentiation. The tube ruptures releasing the sperms and the other contents in the
archegonial chamber. A sperm passes through the neck of the archegonium, comes in contact
and fuses with the egg nucleus exactly in the same way as in Cycas.
8. Embryogeny in Ginkgo:
The zygote becomes enlarged and starts dividing by many free-nuclear divisions just after
fertilization. The nuclei are distributed irregularly (Fig. 10.26A). The cell wall formation
starts just after a few divisions of the nucleus and soon a pro-embryo is differentiated (Fig.
10.26B). Three types of cells are clearly differentiated in the pro-embryo (Fig. 10.26C).

The cells towards the chalazal end are smaller in size with dense cytoplasm, followed by
another zone of comparatively larger cells, which do not possess such a dense cytoplasm.
Elongated cells are present in the uppermost zone. There is no differentiation of suspensor in
the embryo.
The development of the cotyledon starts from the lowermost zone soon after the
differentiation of these zones (Fig 10.26D,E). Roots, stem and cotyledons are soon
differentiated (Fig. 10.26 E,F). Generally, five leaves are present in the mature embryo. Out
of these five leaves the first two are decussate with cotyledons while the remaining leaves are
irregular in arrangement in embryo and seedling.
The germination of the seed is of hypogeal type and quite similar to that of Cycas. A strong
tap root develops soon and the seedling bears many bilobed leaves, which is characteristic
feature of Ginkgo biloba. Soon, a mature plant, with many more leaves, develops.
In a recent study, Dogra (1992) opined that development of embryo in Ginkgo biloba is a
continuous process from the time of fertilization until the seed germination. Actually, the
major part of the growth of embryo takes place when the seed is detached from the tree and is
lying on the ground.
9. Economic Importance of Ginkgo:
Ginkgo biloba tree is treated as a sacred plant in China and Japan and worshipped by the
people. It is regarded as “Holy Tree” by Buddhist monks and nuns because of its tall, strong
and majestic appearance. It is grown as an ornamental plant in gardens in several countries.
In autumn, the green leaves turn yellow, showing a magnificent autumn scenery. The
endosperm of its roasted seeds is edible, but seeds may prove fatal if taken in large quantity.
After the outer skin of its apricot-like seeds is peeled off or decayed, the kernel left is called ”
White Fruit”. Ginkgo is, therefore, also named “White fruit tree”.
The kernel is also used as food and Chinese medicine as well. If it is taken raw, it functions
as detoxicating and asthma-curing medicine. Although the plants are highly resistant to
pathogenic infections, specially of bacteria and fungi, their wood is not of much commercial
importance. It also possesses a strong capacity of resistance to air pollution by smoke and
poisonous gases.

Since an unpleasant odour is emitted from the ripe seeds of female trees, male trees are
largely preferred and grown as shade trees in China, Japan and United States. Ginkgo trees
grow and develop very slowly.
It is said that the tree planted by grandfather will bear “fruit” at the time when his grandson
grows up. Therefore, it is also known as “grandfather grandson tree”.An extract of leaves of
G. biloba is useful in the treatment of cerebral insufficiency and vertigo.
Phylogeny of Ginkgoales:
Some of the striking peculianties of Ginkgo biloba, the only living representative of
Ginkgoales, include:
(i) Bilobed, fan-shaped leaves with dichotomous open venation,
(ii) Ovule bearing a collar at the base,
(iii) Micro-sporangiophore bearing a hump-like outgrowth at its apex,
(iv) Presence of tent pole at the tip of the female gametophyte, and
(v) Absence of suspensor in its embryo.
In-spite of these characteristics, Ginkgoales resemble in some or other aspects with
Cordaitales, Pteridospermales, Filicales, Cycadales and even Coniferales.
Resemblances with Cordaitales
(i) Presence of double leaf trace,
(ii) Endospermic beak in the mature ovule, and
(iii) The probable motility of the spermatozoids of the members of both Ginkgoales and
Cordaitales are some of the resemblances between these two groups.
Resemblances with Pteridospermales:
(i) Presence of leaf gap in the stem,
(ii) Dichotomous open venation in the wedge-shaped leaves of Ginkgo and pinnules of some
seed ferns (Ptendospermales),
(iii) A distinct pollen chamber,
(iv) A massive fleshy layer in the ovules, and
(v) Collar at the base of the ovules in Ginkgo and cupule in ovules of some Pteridospermales,
are some of the characteristics showing resemblance between Ginkgoales and
Pteridospermales.
Resemblances with Filicales:
(i) The similarities between the primary xylem structure, secondary xylem structure and
periderm of Ginkgo and some ferns (Ophioglossum and Botrychium),

(ii) Dichotomous open venation in their leaves,
(iii) Multi-flagellated and motile spermatozoids, and
(iv) Presence of ventral canal cell in the archegonia in some of their members.
Resemblances with Cycadales:
(i) Presence of multi-flagellated spermatozoids,
(ii) Well-developed nucellar beak and pollen chamber,
(iii) Haustorial nature of pollen tube,
(iv) Presence of large egg, massive female gametophyte and well-developed venter,
(v) Endoscopic embryo with two cotyledons,
(vi) Seed with thick and well-developed integument, and
(vii) Hypogeal type of seed germination.
In-spite of these above-mentioned similarities, there are several major differences between
Ginkgoales and Cycadales which put them quite apart from each other.
They differ from each other in the:
(i) General structure of their male and female reproductive organs,
(ii) Vascular supply of their ovules,
(iii) Development of their male gametophytes,
(iv) Branched (Ginkgoales) and generally un-branched (Cycadales) nature of their stems,
(v) Simple (Ginkgoales) and compound (Cycadales) nature of their leaves, and
(vi) Presence (Cycadales) or absence (Ginkgoales) of circinate venation in their leaves, etc.
Resemblances with Coniferales:
Ginkgo has several characteristics in common with many members of Coniferales which
compel one to think Ginkgoales as well-defined Coniferophytes. Florin opined that all
Ginkgoales, Coniferales, Cordaitales and Taxales belong to the same natural group named as
Coniferopsida.
All these four orders of Coniferopsida constitute parallel lines of evolution and separated
from each other in the Upper Devonian or Carboniferous times according to Florin.
Some major characteristics common to both Ginkgoales and Coniferales
are listed below:
(i) General appearance of tree is cone like,
(ii) Extensively branched stem with two types of branches, i.e. long shoots and dwarf shoots,
(iii) Leaves are simple with sunken stomata,
(iv) Cortex and pith are narrow and wood is pycnoxylic,

(v) Secondary wood is well-developed,
(vi) Medullary rays are uniseriate,
(vii) Circular bordered pits arranged uniseriately along the radial walls of the tracheids,
(viii) Microsporangia show longitudinal dehiscence, and
(ix) Sessile nature of ovule.
EPHEDRALES
Distribution of Ephedra:
Ephedra (commonly known as joint pine, joint fir, Mormon tea or Brigham tea) is the only
genus in family Ephedraceae and order Ephedrales. It is represented by 50 species.
These species grow in dry climate over wide areas of the Northern hemisphere including
North America,Europe, North Africa, and South west and central Asia. Eight species of
Ephedra are known from India. Some of the common Indian species are E. intermedia, E.
gerardiana, E. sexatilis, E.foliata etc. These species are distributed in dry parts of Punjab,
Haryana, Rajasthan and parts of Kashmir to Sikkim.
Morphological Features of Ephedra:
The plant body is sporophytic and shows xerophytic characters. Mostly the plants are woody
shrubs (Fig. 1 A), a very few species are lianas and some species grow into a small tree. E.
compacta reaches 30 cm in height E. triandra is a tree. Its height is several meters. Plant body
can be differentiated into three parts – root, stem and leaves.
1. Root:
There is a prominent underground tap root system. Later on the adventitious roots develop.
Many root hairs are present but there is no mycorrhiza.
2. Stem:
Like Equisetum, the stem is green, ribbed, branched, fluted and differentiated into nodes and
internodes (Fig.1B). It is distinctly jointed fir) (therefore, commonly known as jointed fir). It

performs the function of photosynthesis and may be called as phylloclade. The branches arise
from the axillary buds and are, therefore, in pairs of threes or fours according to the number
of the scaly leaves at the nodes in different species.
The branches are also green and differentiated into nodes and internodes. The branching
starts early at the cotyledonary stage. The apical meristem is having well marked tunica layer
but the growth of internode is independent due to the presence of the meristemetic zone at its
base. This zone dries up at the end of each growing season. It results in the brittleness and
shedding of the branches. These branches are again replaced in next season by new axillary
branches.
3. Leaves:
Leaves are small scaly, present in pairs at the nodes and are arranged in opposite decussate
manner. (Fig. 1 C, D). These leaves unite at the base to form a basal sheath. Each leaf
contains two unbranched, parallel veins. They are so minute that they are of no use i. e.,
unable to perform photosynthesis. The function of photosynthesis is carried by green stem. In
the axil of each leaf is present a bud for the branch. True foliage leaves are absent.
Internal Structures of Ephedra:
1. Stem:
The stem is ribbed; so, in tranverse section stem shows ridges and grooves (Fig. 2A).

A T.S. of stem at node shows the following structures (Fig. 2A, B):
a. Epidermis:
It is the outermost layer of thick walled cells, covered with a thick layer of cuticle. Sunken
stomata are present on the slopes of the ridges in the circular pits.
b. Hypodermis:
It is present just below the ridges. (Fig 2B). It is made up of sclerenchymatous cells and
provides mechanical strength to the plant.
c. Cortex:
In is present between the thick walled sclerenchyma and vascular cylinder. It can be
differentiated into outer and inner cortex. The outer cortex contains 2-3 layers of radially
elongated palisade tissue and inner cortex consists of 2-3 layers of spongy parenchyma.
The cells of outer and inner cortex are loosely arranged with large intercellular spaces and are
provided with chlorophyll to perform the function of photosynthesis. A few patches of
scleranchymatous cells may also occur in the cortex to provide mechanical support to the
young axis.
d. Endodermis:
It is the innermost layer of cortex. It is not easily distinguishable from the cortical cells.
e. Pericycle:
It is present below the endodermis. It is single layered and ill defined.
f. Vascular Cylinder:
It is endarch, siphonostele and consists of many vascular bundles arranged in a ring. Vascular
bundles are conjointed, collateral, open and endarch. The number of primary vascular strands
is generally eight, out of which four small represent the foliar traces while the other large four
are stem bundles.
Foliar traces run upto the node. Xylem consists of tracheids, vessels and xylem parenchyma.
Due to the presence of the vessels the Ephedra resembles angiosperms. The phloem consists
of sieve cells, phloem parenchyma and albuminous cells. Phloem and xylem are separated by
a narrow strip of cambium.
g. Medullary rays:
Broad, parenchymatous medullary rays are present in between the vascular bundles.
Medullary rays connect the pith with cortex.
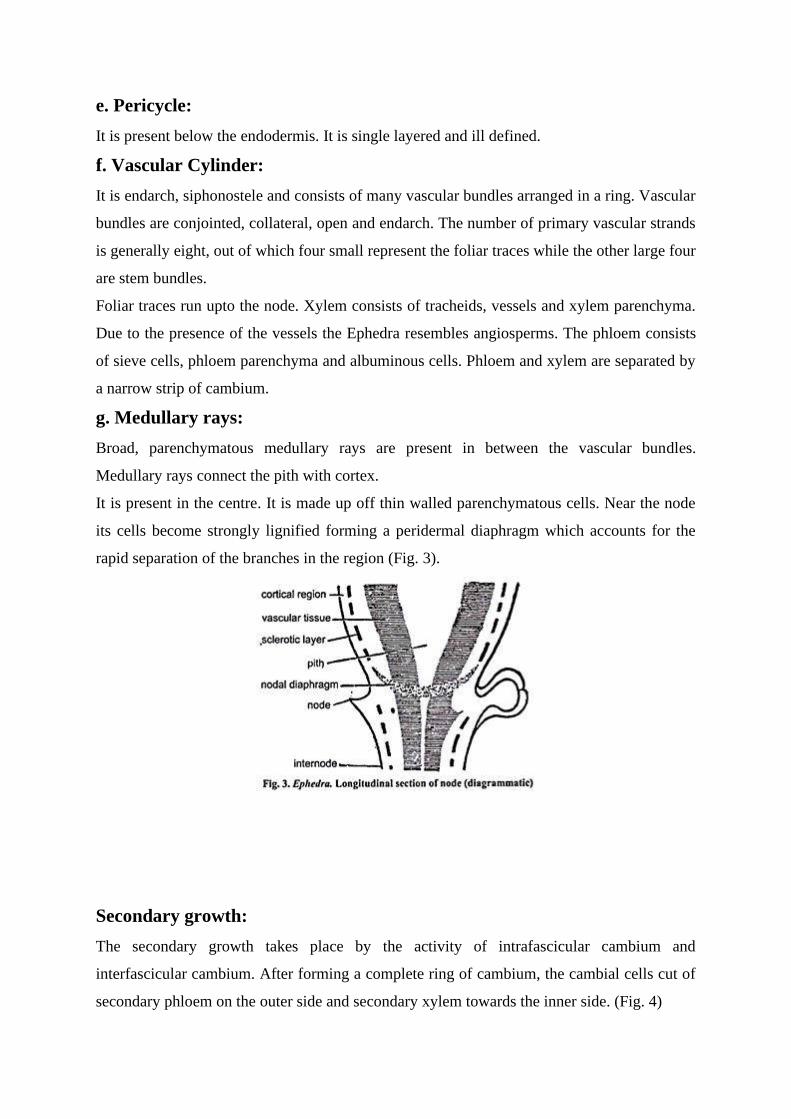
It is present in the centre. It is made up off thin walled parenchymatous cells. Near the node
its cells become strongly lignified forming a peridermal diaphragm which accounts for the
rapid separation of the branches in the region (Fig. 3).
Secondary growth:
The secondary growth takes place by the activity of intrafascicular cambium and
interfascicular cambium. After forming a complete ring of cambium, the cambial cells cut of
secondary phloem on the outer side and secondary xylem towards the inner side. (Fig. 4)

Due to formation of the secondary tissues, primary phloem is crushed and the primary xylem
is pushed towards the inner side at the base of the secondary xylem. In addition to vascular
tissue cambium also forms medullary rays (secondary). These rays are long, broad
(multiseriate) and traverse between secondary xylem and secondary phloem.
Radial Longitudinal Section (R.L.S.):
In R.L.S. xylem tracheids, vessels and medullary rays are clearly visible. Medullary rays are
cut lengthwise and their length and height are revealed (Fig. 5A). Each medullary ray is
composed of irregularly dispersed ray cells and ray tracheids. Tracheids possess bordered pits

on their radial and tangential walls. In vessels, the bordered pits are also arranged in the same
way as tracheids (Fig. 5 B, C).
Tangential Longitudinal Section (T.L.S.) of wood:
Like R.L.S. in T.L.S. also, the xylem, tracheids, vessels and medullary rays are clearly visible
but they are cut transversely here. (Fig. 6). Bordered pits and simple pits are seen on the
radial and tangential walls. The medullary rays are elongated and on their tangential walls are
present simple pits.
2. Leaf:

The transverse section of scaly leaf is oval in shape and can be differentiated into epidermis,
mesophyll tissue and vascular tissue.
a. Epidermis:
It is outer most single layer of thick walled elongated cells. The cells are covered with thick
cuticle. Sunken stomata are present (Fig. 7).
b. Mesophyll tissue:
Two or three layers of palisade tissue are present inner to epidermis. The cells are filled with
chloroplast and large intercellular spaces are present between them. In the centre of the leaf
parenchymatous tissue is present.
c. Vascular tissue:
Two vascular bundles are embedded in the parenchymatous tissue. The vascular bundles are
collateral and closed. Xylem is present towards the upper side.
3. Root:
The transverse section of root shows single layer epiblema, outer cortex (composed of
collenchymatous cells), inner cortex (composed of parenchymatous cells) endodermis and
pericycle. Vascular bundles are radial and exarch. The root may be diarch or triarch.
Reproduction in Ephedra:
Ephedra is heterosporous (produces two types of spores: microspores and macrospores) and
dioecious (both these types of spores are produced on two different plants of the same
species. E. fuliata is monoecious. Microspores are formed in male flowers while megaspores
are formed in female flowers.

These flowers are present in the form of cone like compound strobili. Male flowers are
present in the form of male strobilus while female flowers are present in the form of female
strobilus. Both male and female strobili are compound i. e.,the cone axis bears pairs of bracts
which subtend either microsproangiate or ovulate shoots.
Male Strobilus (Staminate Strobilus):
Male strobili arise in clusters from the nodes of the branches. Each strobilus is rounded,
ovoid or spherical in shape and arises in the axis of a scale leaf. Their number at the node
depends upon the number of scale leaves.
Each strobilus has a central axis which bears 2-12 pairs decussately arranged simple, broad
and cupped bracts. Lower most 1-2 pairs of bracts are sterile. In the axil of each fertile bract
arises a male flower or staminate flower (Fig. 8 A-C). A male strobilus with several male
flowers can be compared with an inflorescence.
Male flowers:

Each male flower has two lipped thin bractioles (perianth) which encloses a stamen.
Bracteoles are united at the base. The flower has a short stalk known as microsporangiophore
and two, eight to twelve microsporangia at its tip (Fig. 8 D).
Microsporangia are sessile and dehisce terminally. Male flower is also called simple
strobilus. A compound male strobilus, therefore, consists of many such strobili.
Structure of microsporangium:
Each microsporangium has 2-3 loculi and is often called as synangium. Its wall is two layered
followed by a prominent tapetal layer enclosing a sporangial sac having many pollen grains
or microspores (Fig. 8E).
Development of microsporangium:
The development of microsporangium is eusporangiate. Microspangia arise at the tip of
microsporangiophore. The microsporangiophore arises as small protuberance in the axil of
the fertile bract of male strobilus. The apex of microsporangiophore becomes lobed after
growing for some time.
Each lobe represents a sporangium. Few hypodermal cells in each lobe enlarge in size. These
cells consist large nuclei, denser cytoplasts and are known as archesporial cells. These cells
divide periclinally into outer primary wall cells or primary parietal cells and primary
sporogenous cells (Fig. 9A).
Primary sporogenous cells further divide by two periclinal divisions to differentiate middle
wall layer, inner tapetal layer and sporogenous cells. The primary wall cells function directly
as the outer wall of the sporangium.
However, according to some workers, the primary wall cells divide periclinally to form three
layered thick wall. The sporogenous cells divide further to form large number of microspore
mother cells. Each microspore mother cell divides by meiosis to form four haploid
microspores arranged in a linear tetrad.

Structure of pollen grain:
Pollen grain is the first cell of the male gametophyte. Each pollen grain is elliptical,
uninucleate and has two wall layers. The outer wall layer is thick and is called exine while the
inner male layer is then and is called intine (Fig. 10A, B).
Female Strobilus (Ovulate Strobilus) or Female Cone:
They usually arise in pairs at each node in the axil of scale leaves. A female strobilus appears
to be an elliptical structure with a pointed apex (Fig. 11 A, B). It retains the same compound

structure as the male strobilus. It consists of a short axis to which are attached three or four
pairs of decussate bracts.
In E. Americana these bracts are swollen and juicy (Fig. 11E). All the pairs of bracts are
sterile except the uppermost one which bears a pair of ovules in its axil (Fig. 11C, D) and
may be variously coloured. Out of the pairs of the ovules only one survives and it takes up a
false terminal position.
Female flower:
The female flower has short stalk and an ovule (megasporangium)
Structure of ovule (megasporangium):
Longitudinal section of an ovule shows that it consists of a mass of parenchymatous cells in
the centre. It is called nucellus. The nucellus is surrounded by a two-layered envelope. These
are usually designated as outer and inner integuments. The outer envelope is formed by four
segments and receives four bundles while the inner one is formed of two segments and
receives two bundles.
The lower half of the inner envelope is fused to the nucellus but upper half is free and
prolongs into a long micropyle tube. By the time of pollination just below the micropyle

pollen chamber develops. Pollen chamber in Ephedra is the deepest known among the
Gymnosperms. The floor of the pollen chamber is formed by female gametophytic tissue and
not by the nucellus as in other gymnosperms. (Fig. 12).
Development of Ovule:
Development of the ovule takes place in the form of a small cellular protuberance. This
protuberance increases in size and becomes the nucellus. Soon neighbouring cells of the base
forms inner and outer integuments. Inner integument surrounds the nucellus except the top
where it form a small opening called micropyle.
A hypodermal archesporial cell differentiates in the nucellus. It divides periclinally into outer
parietal cell and inner megaspore mother cell. The latter is pushed quite deep into the nucellar
tissue.
The megaspore mother cell divides meiotically to form four hapliod megaspores. Generally
the lowermost megaspore (towards the chalazal end) remains functional. It enlarges and gives
rise to female gametophyte (first cell of the female gametophyte) and the remaining upper
three megaspores degenerate.

Gametophytic Phase:
The sporogenesis results in the formation of micro- and megaspores representing the
gametophytic stage. They undergo gametogenesis to form the male and female gametophytes
respectively.
Development of male gametophyte before pollination:
It takes place in microsporangium. After the reduction division spores tetrads are formed. The
four cells of the tetrad separate and develop into microspores. The microspore divides by a
transverse wall to form a small prothallial cell and a large outer cell is (Fig. 13 A). The outer
cell again divides by a transverse wall and forms a second prothallial cell and an antheridial
cell. (Fig 13 B).
The antheridial cell divides to form a small generative cell and a large tube cell (Fig. 13 C,
D). The generative cell soon divides into the nuclei of stalk cell and body cell. The nuclei of
stalk cell and body cell remain surrounded by a common mass of cytoplasm (Fig. 13 E, F).
Pollens are shed at this five celled stage.
Development of female gametophyte:

As mentioned earlier, the functional megaspore is the first cell of the female gametophyte. It
enlarges and its nucleus divides into two. These nuclei move towards the opposite pole and
are separated by a large central vacuole.
Later these two nuclei divide by free nuclear division to form as many as 256 nuclei. These
nuclei are arranged in a peripheral layer around the central vacuole. Later the central vacuole
disappears and free nuclei are evenly distributed throughout.
Centripetal wall formation (from periphery towards the centre) starts and thus a mass of
cellular tissue is formed. It is called female gametophyte or endosperm. Gradually the female
gametophyte is differentiated into two regions.
Micropylar region and antipodal region. Micropylar region consists of loosely arranged thin
walled cells, which later on give rise to archegonia. Antipodal region is further differentiated
into lower storage zone and basal haustorial zone. Storage zone comprises of bulk of
endosperm. This zone consists of compactly arranged cells which are full of starch and other
food. The cells of the haustorial zone absorb the food material from the nucellus.
Structure and development of archegonium:
Archegonia arise in the micropylar region. The number of archegonia in Ephedra varies from
1-3 but they are generally two in number. Any superficial cell of female gametophyte
towards micropylar region acts as archegonial initial (Fig. 14A). It divides by a transverse
division to form outer primary neck cell or neck initial and inner central cell (Fig. 14B). The
neck cell undergoes a number of divisions to form a long neck of 8 or more tiers (minimum
of 32 cells). It encloses no neck canal.

The neck of archegonium of Ephedra is the longest in the gymnosperms. The central cell
enlarges in size. Its nucleus divides into a ventral canal nucleus and an egg nucleus but no
wall is laid down between the two.
As the archegonium reaches towards maturity, the cells of neck usually merge with
surrounding gametophytic cells and become undistinguishable from the surrounding cells of
female gametophyte. The cells adjacent to the central cell divide transversely to form a
distinct jacket layer, which may be two or three layer thick.
A mature archegonium consists of a long neck and a central cell having a ventral canal
nucleus and egg nucleus (Fig. 13, 14).
Pollination:
The pollination is anemophilous i.e. it takes place by wind. Pollen grains are carried by the
wind on the female strobilus. The cells of the nucellus secrete pollination drop which comes
out through the micropylar canal. Pollen grains to adhere to the pollination drop. Pollen
grains are sucked inside and come to lie in a deep pollen chamber.
Development of male gametophyte after pollination:
Pollen grains germinate in the pollen chamber. The exine ruptures and intine comes out in the
form of pollen tube. The nucleus of the body cell divides to form two male gametes (Fig. 16)
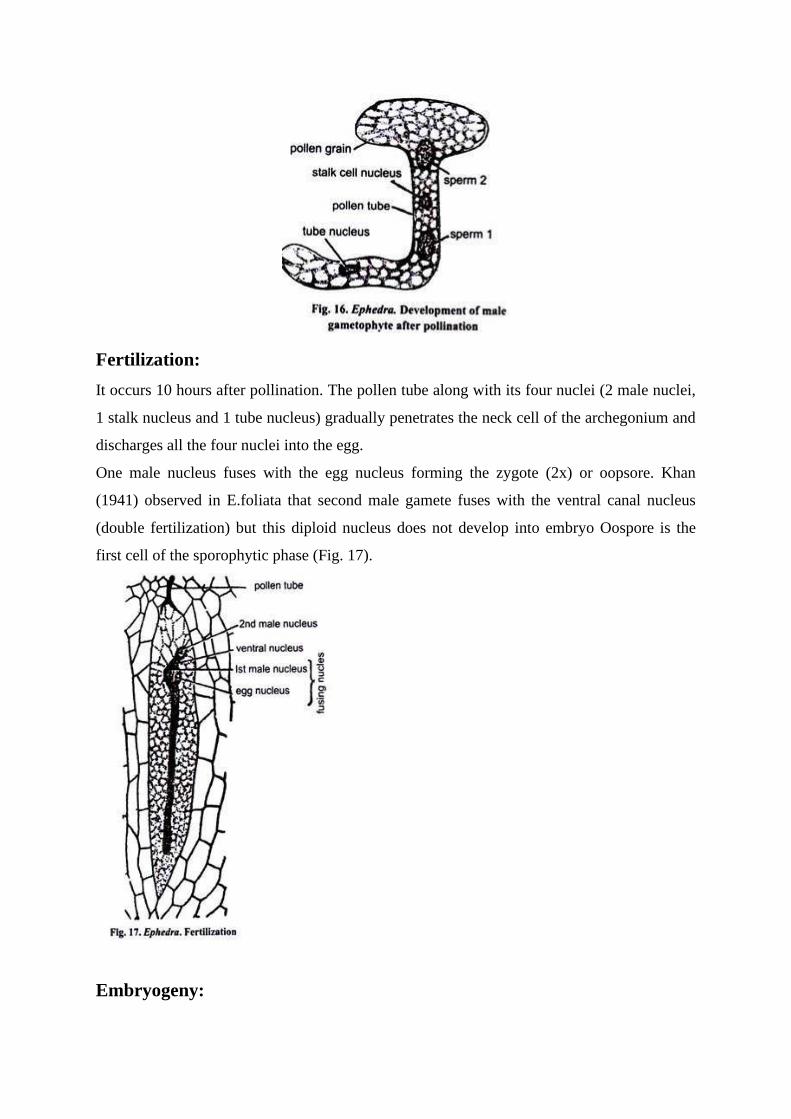
Fertilization:
It occurs 10 hours after pollination. The pollen tube along with its four nuclei (2 male nuclei,
1 stalk nucleus and 1 tube nucleus) gradually penetrates the neck cell of the archegonium and
discharges all the four nuclei into the egg.
One male nucleus fuses with the egg nucleus forming the zygote (2x) or oopsore. Khan
(1941) observed in E.foliata that second male gamete fuses with the ventral canal nucleus
(double fertilization) but this diploid nucleus does not develop into embryo Oospore is the
first cell of the sporophytic phase (Fig. 17).
Embryogeny:

More than one archegonium may be fertilized in an ovule, but only one oospore develops into
embryo. The zygote nucleus divides by three free nuclear divisions to form eight nuclei.
These nuclei are irregularly distributed in the cytoplasm of the archegonium.
Later wall-formation takes place and this structure is known as proembryo. Each cell of
inproembryo is capable to develop into an independent embryo. Three to five of these nuclei
individually enclose in somewhat irregular walls and become globular.
These are known as pro-embryonal cells, each of which produces an independent embryo. In
Ephedra, this type of polyembrony without any cleavage, it unique among gymnosperms.
Because the polyembryony occurs without any cleavage, it is known as embryo sac
polyembryony. Each proembryo grows into tubular structure called the suspensor (Fig. 18A-
C).
Tube nucleus of the proembryo divides into two. Both these nuclei move into the tube. A wall
separates these two daughter nuclei and forms two cell(Fig. 18D). The cell towards the
micropylar and disintegrates while the cell formed towards the chalazal end of the tube
survives and is called embryonal initial.
The tube grows more and carries the embryonal initial deep into theprothallus tissue. This
embryonal initial divides into a proximal suspensor cell and a distalembryo cell. The embryo
cell divides and develops into the embryo proper which contains two cotyledons (Fig. 18E-
G).Although several embryos may develop in a single ovule but only one survives and
reaches at maturity as seed.
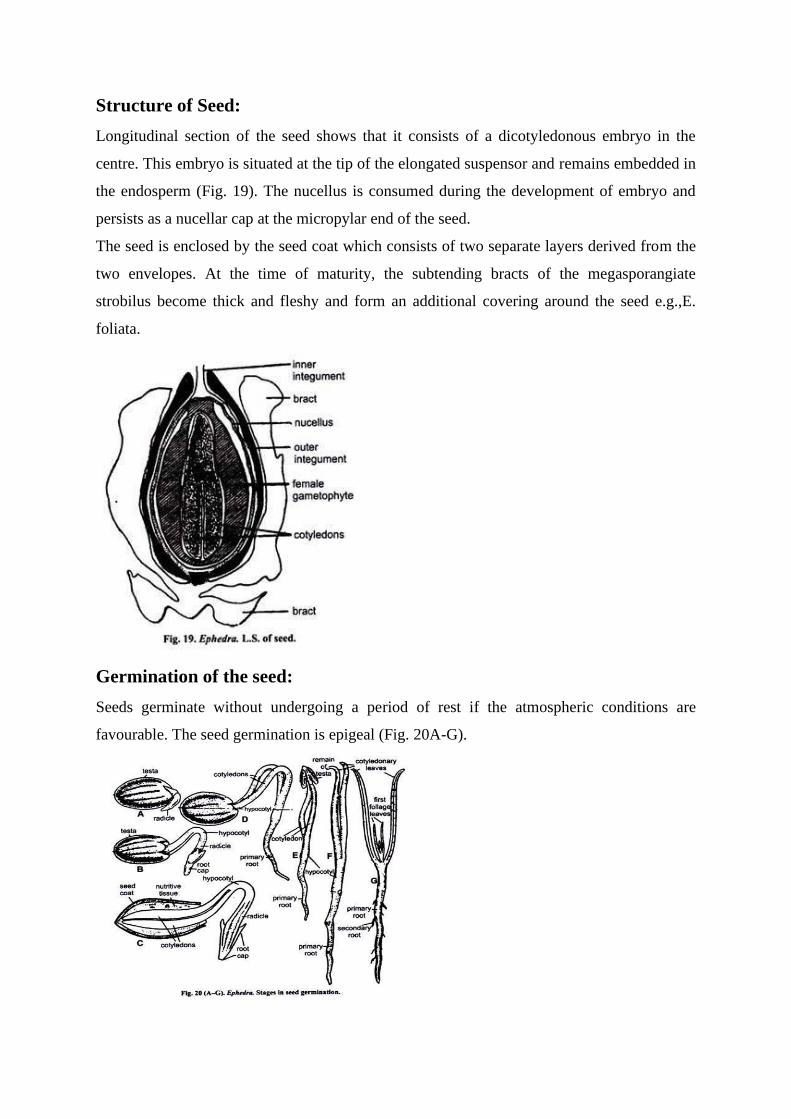
Structure of Seed:
Longitudinal section of the seed shows that it consists of a dicotyledonous embryo in the
centre. This embryo is situated at the tip of the elongated suspensor and remains embedded in
the endosperm (Fig. 19). The nucellus is consumed during the development of embryo and
persists as a nucellar cap at the micropylar end of the seed.
The seed is enclosed by the seed coat which consists of two separate layers derived from the
two envelopes. At the time of maturity, the subtending bracts of the megasporangiate
strobilus become thick and fleshy and form an additional covering around the seed e.g.,E.
foliata.
Germination of the seed:
Seeds germinate without undergoing a period of rest if the atmospheric conditions are
favourable. The seed germination is epigeal (Fig. 20A-G).


Economic Importance of Ephedra:
1. An alkaloid ephedrine is obtained from E. gerardiana, E. intermedia, E.
nebrodensis etc. It is used in preparation of medicines that cure cough,
bronchitis, asthma and hay fever.
2. Tincture of E. gerardiana is also used as a cardiac and circulatory stimulant.
3. Decoction of the stem and roots is used to cure rheumatism and syphilis
e.g.,E. antisyphilitica.
4. The juice of berry is used to cure respiratory disorders.
5. Mormon tea is brewed from the species of Ephedra in south western United
States.
6. Some species are grown as ornamentals.
www.google.com
All the above study materials were downloaded from Google search engine and edited by me.
Recommended









