
REGULAR ARTICLE
Fine root turnover and litter production of Norway sprucein a long-term temperature and nutrient manipulationexperiment
Jaana Leppälammi-Kujansuu & Maija Salemaa &
Dan Berggren Kleja & Sune Linder &
Heljä-Sisko Helmisaari
Received: 29 May 2013 /Accepted: 16 July 2013 /Published online: 13 August 2013# Springer Science+Business Media Dordrecht 2013
AbstractBackground and aims Increased soil temperature andnutrient availability enhance soil biological activity.We studied how these affect fine root growth andsurvival, i.e. below-ground litter production, in relationto above-ground foliage litter production of Norwayspruce (Picea abies (L.) Karst.).Methods The treatments, irrigation (I), soil warming +irrigation (WI), fertilization + irrigation (FI) and soilwarming + fertilization + irrigation (WFI) were startedin 1987 (F, I) and in 1995 (W). The annual productionof fine root litter was estimated from minirhizotrons(survival) and soil-cores (biomass) and the annualabove-ground litter production from litter traps.
Results and conclusions The number and elongationof fine roots tended to be higher in WI and I comparedto the other treatments, which may indicate nutrientshortage. Fine roots in the WFI treatment had thelowest median longevity and from three to fourfoldhigher below-ground litter production compared toWI, FI or I - higher soil temperature increased the litterinput particularly into the mineral soil. Only fertiliza-tion increased the above-ground litter production. Aswarmer and more nutrient-rich soil significantly short-ened the fine root lifespan and increased the litter input,the storage of carbon in boreal forest soil may increasein the future.
Keywords Long-term soil warming . Long-termfertilization . Fine root longevity . Minirhizotron .
Litter production .Picea abies . Survival analysis
Introduction
Climatic scenarios suggest that climate change in-creases the mean annual temperature and increasesprecipitation in the boreal zone (IPCC 2007). Longergrowing season (Menzel and Fabian 1999) and elevat-ed temperature enhance nitrogen mineralization(Hyvönen et al. 2007) in the absence of moisturelimitations. As nitrogen is the most growth-limitingnutrient in the boreal zone (Kukkola and Saramäki1983; Linder and Flower-Ellis 1992) its increasedavailability gives trees the potential to grow fasterand accumulate more biomass, and thus carbon (C),
Plant Soil (2014) 374:73–88DOI 10.1007/s11104-013-1853-3
Responsible Editor: Peter J. Gregory.
J. Leppälammi-Kujansuu (*) :H.<S. HelmisaariDepartment of Forest Sciences, University of Helsinki,P.O. Box 27, 00014 Helsinki, Finlande-mail: [email protected]
M. SalemaaFinnish Forest Research Institute,P.O. Box 18, 01301 Vantaa, Finland
D. B. KlejaDepartment of Soil and Environment, Swedish University ofAgricultural Sciences,P.O. Box 7014, 75007 Uppsala, Sweden
S. LinderSouthern Swedish Forest Research Centre, SwedishUniversity of Agricultural Sciences,P.O. Box 49, 23053 Alnarp, Sweden

in both above- and below-ground compartments(Vanninen et al. 1996).
Soils are the major terrestrial storage of C (Lal 2005),and a large part of the C flux into the soil takes place viafine root and mycorrhizal turnover (Godbold et al. 2006;Helmisaari et al. 2002; Vogt et al. 1986). The responseof C flux into the forest soil is particularly interesting inrelation to climate change, but the environmental andbiological factors that control the fine root mortality rateand below-ground litter production are still poorlyknown. According to the C optimization theory(Eissenstat 1992), trees growing in nutrient-poor habi-tats invest large amounts of C in the construction of newfine roots, in order to acquire sufficient nutrients (Keyesand Grier 1981). As the cost of construction is high inrelation to the cost of maintenance and nutrient uptake innutrient-poor sites, root lifespan is expected to increase(Eissenstat et al. 2000; Schoettle and Fahey 1994).Furthermore, maintenance costs are dependent on theroot respiration activity which increases with soil tem-perature (Burton et al. 2002; Schindlbacher et al. 2009),leading to the lower lifespan expectancy of fine roots inwarmer soils (Eissenstat and Yanai 1997; Hendrick andPregitzer 1993).
In theory, root longevity can be predicted by ana-lyzing the costs and benefits of constructing andretaining roots (Eissenstat et al. 2000), but in practiceit is impossible to incorporate all the affecting compo-nents in the analysis, i.e. root diameter, root exudates,mycorrhizas, grazing, parasitism, and environmentalheterogeneity. The traits of potential growth rate of atree, such as fine root diameter and fine rootcarbon:nitrogen ratio, have recently been shown toexplain a large proportion of the variation in fine rootlongevity in a similar environment (McCormack et al.2012), but connections between root longevity andenvironmental factors still need further clarification.In addition to longevity, trees can adjust the biomassof the fine roots (Finér et al. 2011) or foraging strategy(Ostonen et al. 2011) to meet the altered environmentalconditions—or vary the allocation between the above-and below-ground parts of the tree (Keyes and Grier1981; Mäkelä et al. 2008; Ostonen et al. 2011).
Silviculture has long traditions in Scandinavia andthe growth responses of tree trunks to nutrient additionby fertilization have been widely studied. However,only by understanding the responses of both above-and below-ground compartments of the trees can theactual ecological effects of the environmental changes
be traced. Above the ground, the initial response tofertilization has been found to be expressed as amarked increase in the leaf area index through in-creased size and number of needles and concurrentlydecrease in the needle litterfall due to the loweredmortality of needles (Derome et al. 2009; Linder andFlower-Ellis 1992). However, when the fertilizationcontinued, the litter production increased compared tothe control (Derome et al. 2009; Linder and Flower-Ellis 1992). A similar trend of increasing amount offoliage litterfall can also be observed along a fertilitygradient from poor to fertile coniferous stands(Saarsalmi et al. 2007; Ukonmaanaho et al. 2008).
Helmisaari et al. (2007) found that the needle:fineroot biomass ratio of coniferous trees increased withthe increasing fertility level of the sites or with increas-ing nitrogen concentration of the roots. Likewise, Callocation models, based on empirical data fromFinnish Norway spruce and Scots pine stands, indicat-ed that allocation to root production decreases withincreasing nitrogen availability and the wood produc-tion of trunks uses the excess of C without remarkablechanges in the allocation to foliage (Mäkelä et al. 2008;Valentine and Mäkelä 2012). Thus, trees appear todirect the allocation of C substantially more to theabove-ground than below-ground compartments innutrient-rich conditions. However, biomass allocationrelates poorly to C flux and should not be used to infereither (Litton et al. 2007). The lifespan of differentabove- and below-ground litter compartments also af-fects markedly the amount of litter produced annually.
Despite the substantial coverage area of Norwayspruce forests in Eurasian boreal and temperate foreststhe major part of the publications on fine root turnoverof forest trees has focused on temperate forests and ondeciduous trees (Idol et al. 2000; López et al. 2001;Lukac et al. 2003), and less on Norway spruce (Gaulet al. 2009; van Praag et al. 1988; Withington et al.2006). The responses of Norway spruce fine roots toshort-term experimental manipulations in the borealzone have been studied (Clemensson-Lindell andPersson 1995; Majdi 2001; Majdi and Öhrvik 2004),but because the monitoring period lasted only a shorttime it is hard to distinguish the immediate or transientresponse e.g. from the permanent change in the alloca-tion patterns. Moreover, only a few studies have cov-ered both the above- and below-ground litter produc-tion on the same site (Hansson et al. 2013a; Kleja et al.2008; Röderstein et al. 2005). Therefore, the lack of
74 Plant Soil (2014) 374:73–88

long-term, interactive experiments is a fundamentalflaw as the climate-related environmental changes takeplace simultaneously and gradually, over decades.
Our main aim was to study whether long-term ele-vated temperature and/or better nutrient availabilityincrease the fine root mortality rate and litter input tothe soil. At the same site the short-term responses havebeen studied by Majdi and Öhrvik (2004). The annualproduction of fine root litter and the above-groundfoliage litter were compared after more than two de-cades of optimal fertilization and/or more than onedecade of soil warming. We recently reported that thesetreatments increased the needle and fine root biomasstogether with basal area (Leppälammi-Kujansuu et al.2013). As the productivity of boreal forest is restrictedby nutrient deficiency and low temperatures, we hy-pothesized that 1) both higher soil temperature andnutrient availability will shorten fine root longevity,and that the combined treatment would have an addi-tive effect. Further, we expected that 2) higher resourceavailability and warmer soil will increase the below-ground litter production, but 3) in relation to the above-ground fraction of the total litter production the below-ground fraction will decrease.
Material and methods
Site description
The study was conducted in a long-term nutrient opti-mization experiment at Flakaliden (64°07′ N, 19°27′ E,310m a.s.l.) in northern Sweden. At the site, the forest iseven-aged, the stand was planted in 1963 with 4-yearold Norway spruce seedlings of local provenance afterclear-felling. At the time of establishment, stand densitywas ca. 2,400 trees ha−1 with no subsequent thinnings.Understory vegetation consists of Vaccinium vitis-idaea,Vaccinium myrtillus, Deschampsia flexuosa andEmpetrum spp., and the ground is covered by feathermosses.
Soil at the site is a thin podzolic, sandy, post-glacialtill with mean depth of about 120 cm, classified asSpodosol according to USDA Soil Survey Staff(1999), with soil water content normally not limitingfor tree growth (Bergh et al. 1999). The site fertility isrelatively low (H100=17–19 m; Hägglund andLundmark 1977). The site belongs to the middle borealsub-zone (Sjörs 1999) with harsh boreal climate with
long cool days in the summer and short cold days in thewinter. The mean annual air temperature is 2.5 °C andthe mean monthly temperature varies from −7.5 °C inFebruary to 14.6 °C in July (mean for 1990–2009).Mean annual rainfall is ~600 mm with approximatelyone-third falling as snow, which usually covers thefrozen ground from mid-October to early May. Thelength of the growing season (the period with a dailymean air temperature ≥+5 °C) averages ~150 days with atemperature sum of ~900° days (°C), but with largebetween-year variations. The annual depositions of totalnitrogen and sulfur in the region are low (both ≤3 kg ha−1)(Berggren et al. 2004).
Fertilization and warming treatments
The fertilization experiment was established in 1987,with the aim of optimizing the nutritional status of thestand, and at the same time avoiding the leaching ofnutrients to groundwater. All essential macro- andmicro-nutrients were supplied every second day duringthe period of active growth (early June–mid-August).The amount and composition of the nutrient mix wasdetermined each year on the basis of foliar analysis,monitoring of nutrient concentrations in the soil water,and from the predicted growth response. The amountof irrigation was set to maintain soil water potentialabove −100 kPa. Net nitrogen mineralization is 4 and18 kg N ha−1 a−1 in control and fertilized stands,respectively (Andersson et al. 2002). For further detailson experimental design and treatments, see Linder(1995) and Bergh et al. (1999).
For the present study, of the 50×50 m treatmentplots, only 10×10 m sub-plots (Table 1) were used withfollowing treatments: soil warming-fertilization-irrigation (WFI), soil warming-irrigation (WI),fertilization-irrigation (FI) and irrigation (I) whichwas used as the reference level and represented acontrol, although since 2007 it has received a smallamount of fertilizers with irrigation water (Table 2).WFI and WI treatments consisted of two sub-plots andtheir data was pooled together.
In the summer of 1994, six 85-m long heating cables(DEVI, Elektrovärme AB, Vällingby, Sweden) wereburied in the soil of the buffer zone of the fertilizationand/or irrigation plot. The cables were installed betweenthe organic and mineral soil layer (spacing ~20 cm). Thefunctioning of the system was tested in the fall and soilwarming started in April 1995, 5 weeks before the
Plant Soil (2014) 374:73–88 75
snowmelt, with an increase of 1 °C per week, until a5 °C difference between warmed and non-warmed plotswas attained (Table 3). In falls, the soil temperature wascorrespondingly decreased by 1 °C per week after thesoil temperature in the control plots approached 0 °C.The aim of soil warming was to lengthen the growingseason by 2 months. Irrigation was included in the soilwarming treatment in order to avoid unwanted dryingeffects. For further information of the warming treat-ment, see Bergh and Linder (1999).
Soil temperature profiles were measured from theorganic layer and 5, 10, 20, 30 and 40 cm depth ofmineral soil. The depth of the frozen soil was measuredby a frost-depth gauge developed by Gandahl (1957),for English description, see Rydén and Kostov (1980),inserted to 50 cm depth and measured from one to threetimes per month (Table 3). After 10 years of optimizedfertilization, at the time of installing the heating cables,
in the fertilized plots the annual stem volume productionhad more than quadrupled (from 3 to 14 m3 ha−1 year−1)and the trees were higher compared to those in the non-fertilized treatment (Bergh et al. 1999 and Table 1).
Minirhizotrons
In September 1994, five acrylic minirhizotron tubes(length 45 cm, internal D 5 cm, external D 6 cm) wereinstalled at an angle of 45° to the soil surface of eachtreatment plot. Using the same tubes, the short-termresponse of root growth to soil warming was reportedby Majdi and Öhrvik (2004). The images were collect-ed using a minirhizotron camera (BTC-2; BartzTechnology, Santa Barbara, USA) three times in 2008(11.6; 19.8 and 25.9), once in 2009 (3.6) and four timesin 2010 (2.6; 15.7; 23.8 and 8.10). The camera waslowered to ten tubes at WFI, eight at WI, four at FI and
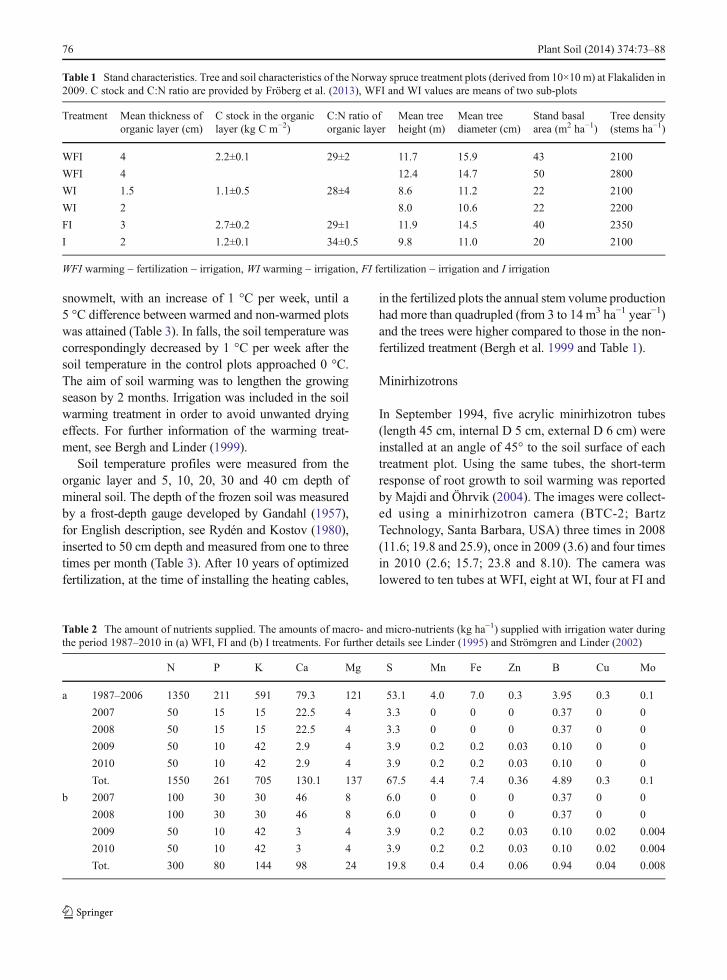
Table 1 Stand characteristics. Tree and soil characteristics of the Norway spruce treatment plots (derived from 10×10 m) at Flakaliden in2009. C stock and C:N ratio are provided by Fröberg et al. (2013), WFI and WI values are means of two sub-plots
Treatment Mean thickness oforganic layer (cm)
C stock in the organiclayer (kg C m−2)
C:N ratio oforganic layer
Mean treeheight (m)
Mean treediameter (cm)
Stand basalarea (m2 ha−1)
Tree density(stems ha−1)
WFI 4 2.2±0.1 29±2 11.7 15.9 43 2100
WFI 4 12.4 14.7 50 2800
WI 1.5 1.1±0.5 28±4 8.6 11.2 22 2100
WI 2 8.0 10.6 22 2200
FI 3 2.7±0.2 29±1 11.9 14.5 40 2350
I 2 1.2±0.1 34±0.5 9.8 11.0 20 2100
WFI warming – fertilization – irrigation, WI warming – irrigation, FI fertilization – irrigation and I irrigation
76 Plant Soil (2014) 374:73–88
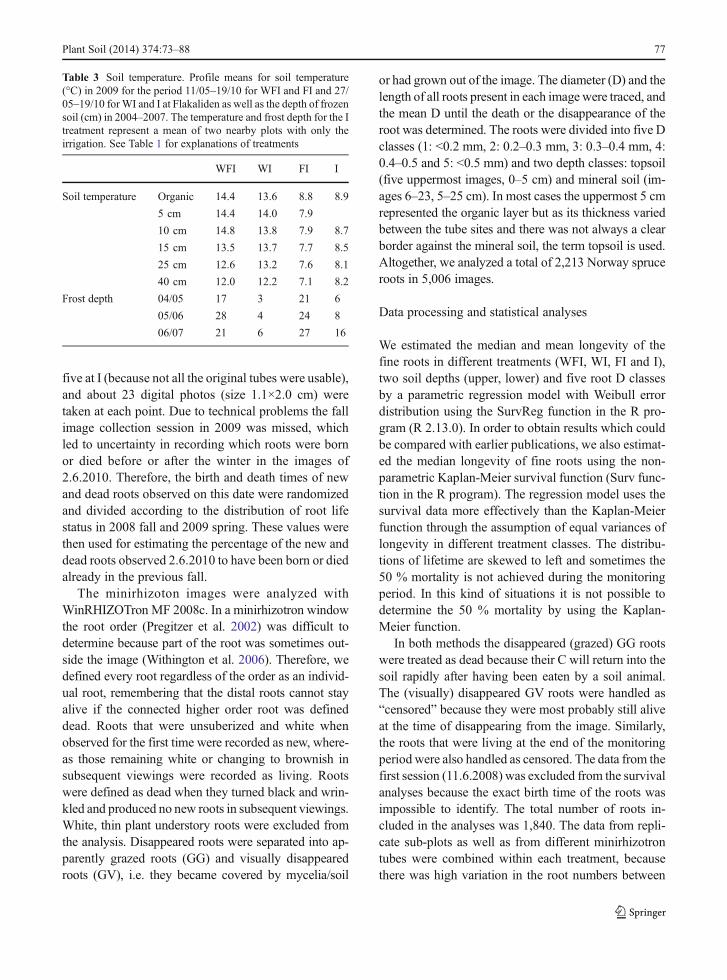
Table 2 The amount of nutrients supplied. The amounts of macro- and micro-nutrients (kg ha−1) supplied with irrigation water duringthe period 1987–2010 in (a) WFI, FI and (b) I treatments. For further details see Linder (1995) and Strömgren and Linder (2002)
N P K Ca Mg S Mn Fe Zn B Cu Mo
a 1987–2006 1350 211 591 79.3 121 53.1 4.0 7.0 0.3 3.95 0.3 0.1
2007 50 15 15 22.5 4 3.3 0 0 0 0.37 0 0
2008 50 15 15 22.5 4 3.3 0 0 0 0.37 0 0
2009 50 10 42 2.9 4 3.9 0.2 0.2 0.03 0.10 0 0
2010 50 10 42 2.9 4 3.9 0.2 0.2 0.03 0.10 0 0
Tot. 1550 261 705 130.1 137 67.5 4.4 7.4 0.36 4.89 0.3 0.1
b 2007 100 30 30 46 8 6.0 0 0 0 0.37 0 0
2008 100 30 30 46 8 6.0 0 0 0 0.37 0 0
2009 50 10 42 3 4 3.9 0.2 0.2 0.03 0.10 0.02 0.004
2010 50 10 42 3 4 3.9 0.2 0.2 0.03 0.10 0.02 0.004
Tot. 300 80 144 98 24 19.8 0.4 0.4 0.06 0.94 0.04 0.008
five at I (because not all the original tubes were usable),and about 23 digital photos (size 1.1×2.0 cm) weretaken at each point. Due to technical problems the fallimage collection session in 2009 was missed, whichled to uncertainty in recording which roots were bornor died before or after the winter in the images of2.6.2010. Therefore, the birth and death times of newand dead roots observed on this date were randomizedand divided according to the distribution of root lifestatus in 2008 fall and 2009 spring. These values werethen used for estimating the percentage of the new anddead roots observed 2.6.2010 to have been born or diedalready in the previous fall.
The minirhizoton images were analyzed withWinRHIZOTron MF 2008c. In a minirhizotron windowthe root order (Pregitzer et al. 2002) was difficult todetermine because part of the root was sometimes out-side the image (Withington et al. 2006). Therefore, wedefined every root regardless of the order as an individ-ual root, remembering that the distal roots cannot stayalive if the connected higher order root was defineddead. Roots that were unsuberized and white whenobserved for the first time were recorded as new, where-as those remaining white or changing to brownish insubsequent viewings were recorded as living. Rootswere defined as dead when they turned black and wrin-kled and produced no new roots in subsequent viewings.White, thin plant understory roots were excluded fromthe analysis. Disappeared roots were separated into ap-parently grazed roots (GG) and visually disappearedroots (GV), i.e. they became covered by mycelia/soil
or had grown out of the image. The diameter (D) and thelength of all roots present in each image were traced, andthe mean D until the death or the disappearance of theroot was determined. The roots were divided into five Dclasses (1: <0.2 mm, 2: 0.2–0.3 mm, 3: 0.3–0.4 mm, 4:0.4–0.5 and 5: <0.5 mm) and two depth classes: topsoil(five uppermost images, 0–5 cm) and mineral soil (im-ages 6–23, 5–25 cm). In most cases the uppermost 5 cmrepresented the organic layer but as its thickness variedbetween the tube sites and there was not always a clearborder against the mineral soil, the term topsoil is used.Altogether, we analyzed a total of 2,213 Norway spruceroots in 5,006 images.
Data processing and statistical analyses
We estimated the median and mean longevity of thefine roots in different treatments (WFI, WI, FI and I),two soil depths (upper, lower) and five root D classesby a parametric regression model with Weibull errordistribution using the SurvReg function in the R pro-gram (R 2.13.0). In order to obtain results which couldbe compared with earlier publications, we also estimat-ed the median longevity of fine roots using the non-parametric Kaplan-Meier survival function (Surv func-tion in the R program). The regression model uses thesurvival data more effectively than the Kaplan-Meierfunction through the assumption of equal variances oflongevity in different treatment classes. The distribu-tions of lifetime are skewed to left and sometimes the50 % mortality is not achieved during the monitoringperiod. In this kind of situations it is not possible todetermine the 50 % mortality by using the Kaplan-Meier function.
In both methods the disappeared (grazed) GG rootswere treated as dead because their C will return into thesoil rapidly after having been eaten by a soil animal.The (visually) disappeared GV roots were handled as“censored” because they were most probably still aliveat the time of disappearing from the image. Similarly,the roots that were living at the end of the monitoringperiod were also handled as censored. The data from thefirst session (11.6.2008) was excluded from the survivalanalyses because the exact birth time of the roots wasimpossible to identify. The total number of roots in-cluded in the analyses was 1,840. The data from repli-cate sub-plots as well as from different minirhizotrontubes were combined within each treatment, becausethere was high variation in the root numbers between
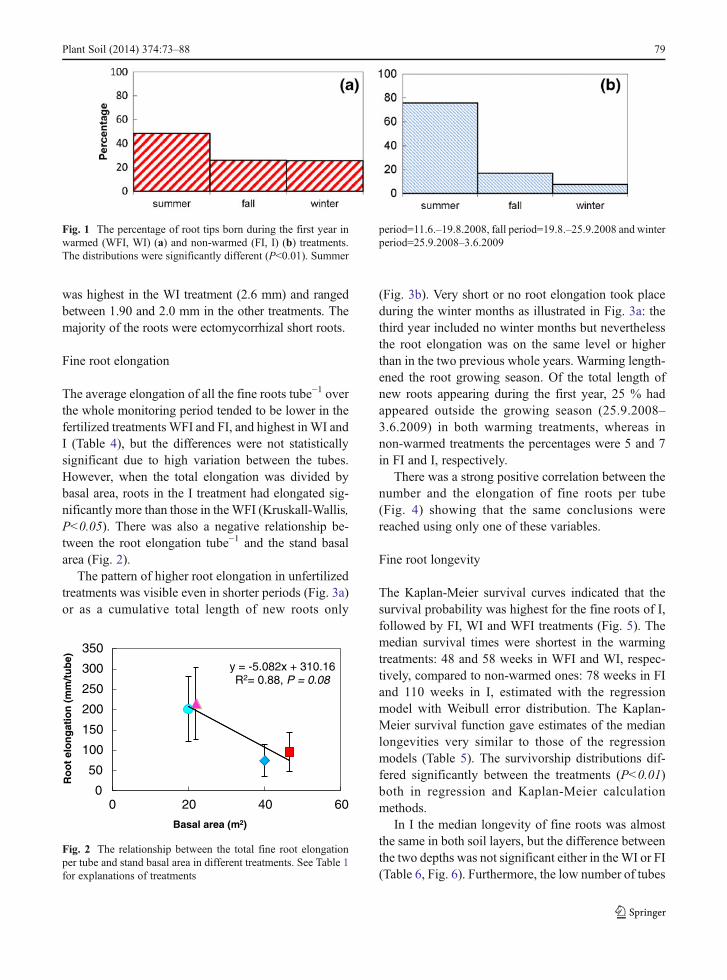
Table 3 Soil temperature. Profile means for soil temperature(°C) in 2009 for the period 11/05–19/10 for WFI and FI and 27/05–19/10 forWI and I at Flakaliden as well as the depth of frozensoil (cm) in 2004–2007. The temperature and frost depth for the Itreatment represent a mean of two nearby plots with only theirrigation. See Table 1 for explanations of treatments
WFI WI FI I
Soil temperature Organic 14.4 13.6 8.8 8.9
5 cm 14.4 14.0 7.9
10 cm 14.8 13.8 7.9 8.7
15 cm 13.5 13.7 7.7 8.5
25 cm 12.6 13.2 7.6 8.1
40 cm 12.0 12.2 7.1 8.2
Frost depth 04/05 17 3 21 6
05/06 28 4 24 8
06/07 21 6 27 16
Plant Soil (2014) 374:73–88 77

the tubes. In the regression method, the statistical dif-ferences between the treatments were tested by com-paring each treatment to the basic level (intercept)according to z values, so that all pair-wise comparisonswere carried out. In the Kaplan-Meier method the sur-vival curves for each treatment were tested using log-rank tests.
The differences between the fine roots birth timedistributions of warmed (WFI and WI combined) andnon-warmed (I and FI) treatments during the first mon-itoring year (from 11.6.2008 to 3.6.2009) were testedusing the χ2 analysis. The average number of fine rootsper tube and the summed elongation of fine roots indifferent treatments were tested using Kruskall-Wallistests.
Below- and above-ground litter production
For estimating below-ground litter production in dif-ferent treatments, we used the median fine root agefrom the survival analyses and fine root biomass data(Leppälammi-Kujansuu et al. 2013) from the soil coresof the same treatments in WFI, WI and FI. The biomassvalues for the I treatment represent a two nearby sub-plots with only the irrigation, without any fertilizers.Only the fine roots with D less than 1 mm were includ-ed to the biomass data as only 0.3 % of the traced rootsin minirhizotron tubes were thicker than 1 mm. Annualfine root production (kg ha−1) was estimated separatelyfor topsoil and for mineral soil by dividing the fine rootbiomass (kg ha−1) by fine root turnover time (yr). Thisis the inverse of turnover rate (year−1).
For calculating the annual above-ground litter pro-duction, eight conical traps (0.8 m2) had been placed atrandom on each plot. The litterfall for the I treatmentrepresent a two nearby sub-plots with only the irriga-tion, without any fertilizers. The traps were at a heightof 0.9 m, and therefore the litter falling into them wasonly from trees. The collection took place twice a yearfrom October 2008 to September 2011, but data wassummed to annual production, and the mean produc-tion of these 3 years was used. The samples were driedat 65 °C for 24 h, and sorted into two components:needles and other (including twigs, cones and all othermaterial) and weighed.
The total annual litter input was calculated by com-bining the above-ground and below-ground litter pro-duction dry weights converted to C input by assuminga C content of 50 %.
Results
The number of fine roots
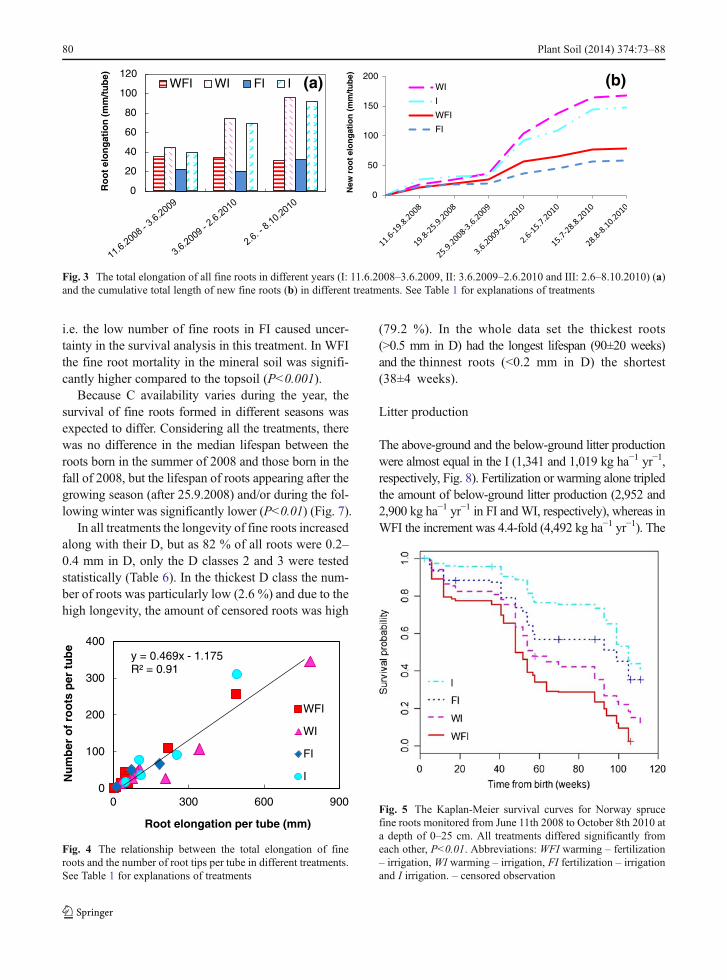
In the whole data the proportion of fine roots was 23 %in the five centimeters thick topsoil (mainly organic)layer and 77 % in the mineral soil. The average numberof fine roots tube−1 tended to be higher in I and WI thanin FI and WFI, but the differences were not statisticallysignificant due to high variation between the tubes(Table 4). Also, the I treatment tended to have moreroots per stand basal area than theWFI (Kruskall-Wallis,P<0.10). Monitoring of the roots during the first yearshowed that in the treatments including warming theappearance of the new roots was seasonally more evenlydistributed and continued later in the fall compared tothe non-warmed treatments (P<0.001, Fig. 1).
At the end of the monitoring period over 40 % of thefine roots were still alive in all the other treatments exceptin WFI, in which only 18 % of the fine roots were alive(Table 4). In the whole data, there was a low number ofgrazed fine roots (GG, 5%) compared to the high numberof visually disappeared fine roots (GV, 14–20 %).
All traced root were less than 1.3 mm in D. In all theother treatments expect in I the majority of fine rootswere 0.3–0.4 mm in D. In the I treatment roots wereslightly thinner, most being 0.2–0.3 mm in D. These twoD classes together comprised of 82 % of all the tracedfine roots. The mean length of an individual fine root
Table 4 The number and elongation of fine roots. The averagenumber of fine roots (a) and the fine root sum elongation, mm (b)per tube in different treatments and life status categories in theend of the experiment
WFI % WI % FI % I % Tot. %
a) Alive 9 18 35 42 19 51 67 63 129 47
Dead 31 62 29 35 10 26 18 17 88 32
GG 3 6 3 3 1 3 6 5 13 5
GV 7 14 16 19 7 20 15 15 45 16
Tot. 51 82 37 106
b) Alive 28 28 116 54 47 62 143 71 334 56
Dead 55 55 59 28 16 22 24 12 155 26
GG 5 5 3 2 3 4 8 4 19 3
GV 12 12 36 17 9 12 27 13 84 14
Tot. 101 215 75 201
GG grazed roots, GV visually disappeared roots, See Table 1 forexplanations of treatments
78 Plant Soil (2014) 374:73–88
was highest in the WI treatment (2.6 mm) and rangedbetween 1.90 and 2.0 mm in the other treatments. Themajority of the roots were ectomycorrhizal short roots.
Fine root elongation
The average elongation of all the fine roots tube−1 overthe whole monitoring period tended to be lower in thefertilized treatments WFI and FI, and highest in WI andI (Table 4), but the differences were not statisticallysignificant due to high variation between the tubes.However, when the total elongation was divided bybasal area, roots in the I treatment had elongated sig-nificantly more than those in theWFI (Kruskall-Wallis,P<0.05). There was also a negative relationship be-tween the root elongation tube−1 and the stand basalarea (Fig. 2).
The pattern of higher root elongation in unfertilizedtreatments was visible even in shorter periods (Fig. 3a)or as a cumulative total length of new roots only
(Fig. 3b). Very short or no root elongation took placeduring the winter months as illustrated in Fig. 3a: thethird year included no winter months but neverthelessthe root elongation was on the same level or higherthan in the two previous whole years. Warming length-ened the root growing season. Of the total length ofnew roots appearing during the first year, 25 % hadappeared outside the growing season (25.9.2008–3.6.2009) in both warming treatments, whereas innon-warmed treatments the percentages were 5 and 7in FI and I, respectively.
There was a strong positive correlation between thenumber and the elongation of fine roots per tube(Fig. 4) showing that the same conclusions werereached using only one of these variables.
Fine root longevity
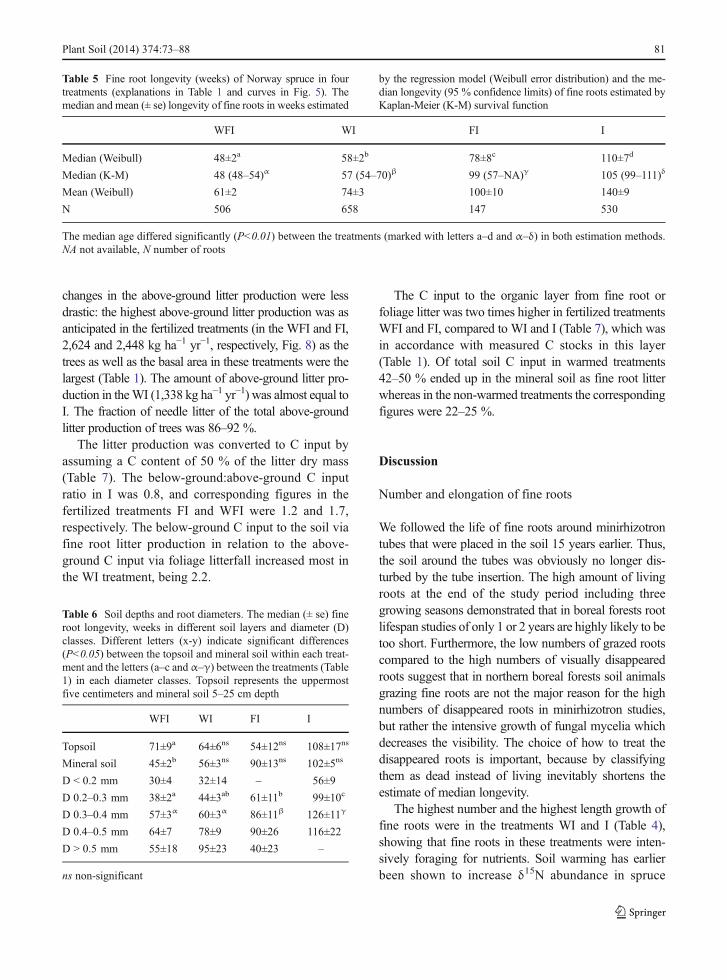
The Kaplan-Meier survival curves indicated that thesurvival probability was highest for the fine roots of I,followed by FI, WI and WFI treatments (Fig. 5). Themedian survival times were shortest in the warmingtreatments: 48 and 58 weeks in WFI and WI, respec-tively, compared to non-warmed ones: 78 weeks in FIand 110 weeks in I, estimated with the regressionmodel with Weibull error distribution. The Kaplan-Meier survival function gave estimates of the medianlongevities very similar to those of the regressionmodels (Table 5). The survivorship distributions dif-fered significantly between the treatments (P<0.01)both in regression and Kaplan-Meier calculationmethods.
In I the median longevity of fine roots was almostthe same in both soil layers, but the difference betweenthe two depths was not significant either in theWI or FI(Table 6, Fig. 6). Furthermore, the low number of tubes
Fig. 1 The percentage of root tips born during the first year inwarmed (WFI, WI) (a) and non-warmed (FI, I) (b) treatments.The distributions were significantly different (P<0.01). Summer
period=11.6.–19.8.2008, fall period=19.8.–25.9.2008 and winterperiod=25.9.2008–3.6.2009
y = -5.082x + 310.16R2= 0.88, P = 0.08
0
50
100
150
200
250
300
350
0 20 40 60
Ro
ot
elo
ng
atio
n (
mm
/tu
be)
Basal area (m2)
Fig. 2 The relationship between the total fine root elongationper tube and stand basal area in different treatments. See Table 1for explanations of treatments
Plant Soil (2014) 374:73–88 79
i.e. the low number of fine roots in FI caused uncer-tainty in the survival analysis in this treatment. In WFIthe fine root mortality in the mineral soil was signifi-cantly higher compared to the topsoil (P<0.001).
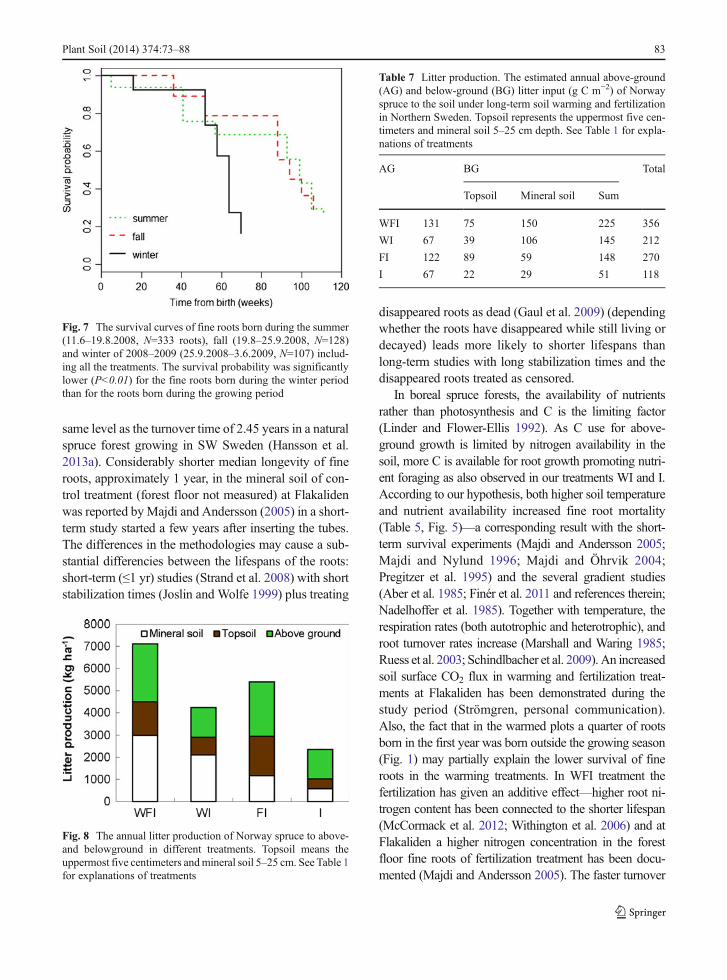
Because C availability varies during the year, thesurvival of fine roots formed in different seasons wasexpected to differ. Considering all the treatments, therewas no difference in the median lifespan between theroots born in the summer of 2008 and those born in thefall of 2008, but the lifespan of roots appearing after thegrowing season (after 25.9.2008) and/or during the fol-lowing winter was significantly lower (P<0.01) (Fig. 7).
In all treatments the longevity of fine roots increasedalong with their D, but as 82 % of all roots were 0.2–0.4 mm in D, only the D classes 2 and 3 were testedstatistically (Table 6). In the thickest D class the num-ber of roots was particularly low (2.6 %) and due to thehigh longevity, the amount of censored roots was high
(79.2 %). In the whole data set the thickest roots(>0.5 mm in D) had the longest lifespan (90±20 weeks)and the thinnest roots (<0.2 mm in D) the shortest(38±4 weeks).
Litter production
The above-ground and the below-ground litter productionwere almost equal in the I (1,341 and 1,019 kg ha−1 yr−1,respectively, Fig. 8). Fertilization or warming alone tripledthe amount of below-ground litter production (2,952 and2,900 kg ha−1 yr−1 in FI andWI, respectively), whereas inWFI the increment was 4.4-fold (4,492 kg ha−1 yr−1). The
0
20
40
60
80
100
120
Ro
ot
elo
ng
atio
n (m
m/t
ub
e) (a)WFI WI FI I
0
50
100
150
200
New
ro
ot
elo
ng
atio
n (
mm
/tu
be) (b)WI
I
WFI
FI
Fig. 3 The total elongation of all fine roots in different years (I: 11.6.2008–3.6.2009, II: 3.6.2009–2.6.2010 and III: 2.6–8.10.2010) (a)and the cumulative total length of new fine roots (b) in different treatments. See Table 1 for explanations of treatments
0
100
200
300
400
0 300 600 900
Nu
mb
er o
f ro
ots
per
tu
be
Root elongation per tube (mm)
WFI
WI
FI
I
y = 0.469x - 1.175R² = 0.91
Fig. 4 The relationship between the total elongation of fineroots and the number of root tips per tube in different treatments.See Table 1 for explanations of treatments
Fig. 5 The Kaplan-Meier survival curves for Norway sprucefine roots monitored from June 11th 2008 to October 8th 2010 ata depth of 0–25 cm. All treatments differed significantly fromeach other, P<0.01. Abbreviations: WFI warming – fertilization– irrigation, WI warming – irrigation, FI fertilization – irrigationand I irrigation. – censored observation
80 Plant Soil (2014) 374:73–88
changes in the above-ground litter production were lessdrastic: the highest above-ground litter production was asanticipated in the fertilized treatments (in the WFI and FI,2,624 and 2,448 kg ha−1 yr−1, respectively, Fig. 8) as thetrees as well as the basal area in these treatments were thelargest (Table 1). The amount of above-ground litter pro-duction in theWI (1,338 kg ha−1 yr−1) was almost equal toI. The fraction of needle litter of the total above-groundlitter production of trees was 86–92 %.
The litter production was converted to C input byassuming a C content of 50 % of the litter dry mass(Table 7). The below-ground:above-ground C inputratio in I was 0.8, and corresponding figures in thefertilized treatments FI and WFI were 1.2 and 1.7,respectively. The below-ground C input to the soil viafine root litter production in relation to the above-ground C input via foliage litterfall increased most inthe WI treatment, being 2.2.
The C input to the organic layer from fine root orfoliage litter was two times higher in fertilized treatmentsWFI and FI, compared to WI and I (Table 7), which wasin accordance with measured C stocks in this layer(Table 1). Of total soil C input in warmed treatments42–50 % ended up in the mineral soil as fine root litterwhereas in the non-warmed treatments the correspondingfigures were 22–25 %.
Discussion
Number and elongation of fine roots
We followed the life of fine roots around minirhizotrontubes that were placed in the soil 15 years earlier. Thus,the soil around the tubes was obviously no longer dis-turbed by the tube insertion. The high amount of livingroots at the end of the study period including threegrowing seasons demonstrated that in boreal forests rootlifespan studies of only 1 or 2 years are highly likely to betoo short. Furthermore, the low numbers of grazed rootscompared to the high numbers of visually disappearedroots suggest that in northern boreal forests soil animalsgrazing fine roots are not the major reason for the highnumbers of disappeared roots in minirhizotron studies,but rather the intensive growth of fungal mycelia whichdecreases the visibility. The choice of how to treat thedisappeared roots is important, because by classifyingthem as dead instead of living inevitably shortens theestimate of median longevity.
The highest number and the highest length growth offine roots were in the treatments WI and I (Table 4),showing that fine roots in these treatments were inten-sively foraging for nutrients. Soil warming has earlierbeen shown to increase δ15N abundance in spruce
Table 5 Fine root longevity (weeks) of Norway spruce in fourtreatments (explanations in Table 1 and curves in Fig. 5). Themedian and mean (± se) longevity of fine roots in weeks estimated
by the regression model (Weibull error distribution) and the me-dian longevity (95 % confidence limits) of fine roots estimated byKaplan-Meier (K-M) survival function
WFI WI FI I
Median (Weibull) 48±2a 58±2b 78±8c 110±7d
Median (K-M) 48 (48–54)α 57 (54–70)β 99 (57–NA)γ 105 (99–111)δ
Mean (Weibull) 61±2 74±3 100±10 140±9
N 506 658 147 530
The median age differed significantly (P<0.01) between the treatments (marked with letters a–d and α–δ) in both estimation methods.NA not available, N number of roots
Table 6 Soil depths and root diameters. The median (± se) fineroot longevity, weeks in different soil layers and diameter (D)classes. Different letters (x-y) indicate significant differences(P<0.05) between the topsoil and mineral soil within each treat-ment and the letters (a–c and α–γ) between the treatments (Table1) in each diameter classes. Topsoil represents the uppermostfive centimeters and mineral soil 5–25 cm depth
WFI WI FI I
Topsoil 71±9a 64±6ns 54±12ns 108±17ns
Mineral soil 45±2b 56±3ns 90±13ns 102±5ns
D < 0.2 mm 30±4 32±14 – 56±9
D 0.2–0.3 mm 38±2a 44±3ab 61±11b 99±10c
D 0.3–0.4 mm 57±3α 60±3α 86±11β 126±11γ
D 0.4–0.5 mm 64±7 78±9 90±26 116±22
D > 0.5 mm 55±18 95±23 40±23 –
ns non-significant
Plant Soil (2014) 374:73–88 81
needles (Strömgren 2001) and tree volume growth(Jarvis and Linder 2000) at the same site. However, thehigher nitrogen availability (Peterjohn et al. 1994;Rustad et al. 2001) or the higher uptake of nitrogen bythe fine roots due to higher soil temperature seem to havebeen more pronounced in the first years of soil warming.In addition the morphological changes in theectomycorrhizal root tips suggest the insufficient avail-ability of nutrients in the WI treatment (Leppälammi-Kujansuu et al. 2013).
Majdi and Öhrvik (2004) reported that half of theannual root production in the warmed plots takes placeduring Oct 5th–June 4th. In this study the root elongationoutside the most active growing season (Sept 25th–June3th) was lower, only a quarter of the whole first year’selongation. Nevertheless, the number of new roots
formed during this period was significantly higher thanin non-warmed treatments (Fig. 1). Obviously thehigher soil temperature lengthens the growing seasonand the active period in soil, but does not correspond-ingly lengthen the period of active photosynthesis(Bergh and Linder 1999). If C allocated to fine rootsmostly originated from current photosynthesis, the fineroots born outside the growing season may have hadless C available for their construction. This was reflectedby their significantly lower survival compared to thecohorts born during the previous summer (Fig. 7).
Fine root longevity
In the I treatment the Norway spruce fine root turnoverwas almost 2 years (Table 5, Fig. 5), which is on the
Fig. 6 The survival curves for the Norway spruce fine roots in different depths in different treatments. Topsoil represents the uppermostfive centimeters and mineral soil 5–25 cm depth. See Table 1 for explanations of treatments
82 Plant Soil (2014) 374:73–88
same level as the turnover time of 2.45 years in a naturalspruce forest growing in SW Sweden (Hansson et al.2013a). Considerably shorter median longevity of fineroots, approximately 1 year, in the mineral soil of con-trol treatment (forest floor not measured) at Flakalidenwas reported byMajdi and Andersson (2005) in a short-term study started a few years after inserting the tubes.The differences in the methodologies may cause a sub-stantial differencies between the lifespans of the roots:short-term (≤1 yr) studies (Strand et al. 2008) with shortstabilization times (Joslin and Wolfe 1999) plus treating
disappeared roots as dead (Gaul et al. 2009) (dependingwhether the roots have disappeared while still living ordecayed) leads more likely to shorter lifespans thanlong-term studies with long stabilization times and thedisappeared roots treated as censored.
In boreal spruce forests, the availability of nutrientsrather than photosynthesis and C is the limiting factor(Linder and Flower-Ellis 1992). As C use for above-ground growth is limited by nitrogen availability in thesoil, more C is available for root growth promoting nutri-ent foraging as also observed in our treatments WI and I.According to our hypothesis, both higher soil temperatureand nutrient availability increased fine root mortality(Table 5, Fig. 5)—a corresponding result with the short-term survival experiments (Majdi and Andersson 2005;Majdi and Nylund 1996; Majdi and Öhrvik 2004;Pregitzer et al. 1995) and the several gradient studies(Aber et al. 1985; Finér et al. 2011 and references therein;Nadelhoffer et al. 1985). Together with temperature, therespiration rates (both autotrophic and heterotrophic), androot turnover rates increase (Marshall and Waring 1985;Ruess et al. 2003; Schindlbacher et al. 2009). An increasedsoil surface CO2 flux in warming and fertilization treat-ments at Flakaliden has been demonstrated during thestudy period (Strömgren, personal communication).Also, the fact that in the warmed plots a quarter of rootsborn in the first year was born outside the growing season(Fig. 1) may partially explain the lower survival of fineroots in the warming treatments. In WFI treatment thefertilization has given an additive effect—higher root ni-trogen content has been connected to the shorter lifespan(McCormack et al. 2012; Withington et al. 2006) and atFlakaliden a higher nitrogen concentration in the forestfloor fine roots of fertilization treatment has been docu-mented (Majdi and Andersson 2005). The faster turnover
Fig. 7 The survival curves of fine roots born during the summer(11.6–19.8.2008, N=333 roots), fall (19.8–25.9.2008, N=128)and winter of 2008–2009 (25.9.2008–3.6.2009, N=107) includ-ing all the treatments. The survival probability was significantlylower (P<0.01) for the fine roots born during the winter periodthan for the roots born during the growing period
Fig. 8 The annual litter production of Norway spruce to above-and belowground in different treatments. Topsoil means theuppermost five centimeters andmineral soil 5–25 cm. See Table 1for explanations of treatments
Table 7 Litter production. The estimated annual above-ground(AG) and below-ground (BG) litter input (g C m−2) of Norwayspruce to the soil under long-term soil warming and fertilizationin Northern Sweden. Topsoil represents the uppermost five cen-timeters and mineral soil 5–25 cm depth. See Table 1 for expla-nations of treatments
AG BG Total
Topsoil Mineral soil Sum
WFI 131 75 150 225 356
WI 67 39 106 145 212
FI 122 89 59 148 270
I 67 22 29 51 118
Plant Soil (2014) 374:73–88 83

in the mineral soil than in the topsoil in both warmingtreatments (Table 6, Fig. 6) was surprising, because theshortest lifespan has generally been recorded in the soilsurface (Baddeley and Watson 2005; Chen and Brassard2013), with a few exceptions (López et al. 2001;McCormack et al. 2012).
The definition of the fine roots according to the arbi-trary diameter borderline, less than 2 mm in D, is acommon practice, despite of the knowledge that thereare noticeable functional and morphological differencesbetween the fine roots of different orders. However, asminirhizotron tracing is a less suitable method for thetracing of root orders (Withington et al. 2006), thenarrower D classes have been proposed to divide hetero-geneous root populations (Guo et al. 2008). Our findingsupports the general rule of thumb that root lifespancorrelates with diameter (Eissenstat and Yanai 2002), butalso that the root survival may vary markedly among fineroots differing in D by only a few tenths of a millimeter(Wells and Eissenstat 2001): the median longevity ofspruce fine roots at least doubled within a root D class ofonly a few millimeters (Table 6). By including all fineroots into one fraction root longevity may beoverestimated, especially in the case of fine roots >1 mmin D (Finér and Laine 1998; Hansson et al. 2013a; Kinget al. 2002).
Litter production
In this study the above-ground litter productionconsisted of needle, branch, twig and cone litter pro-duction of trees; the litter production from understoryvegetation and tree trunks was not included. Likewise,the below-ground litter production consisted only offine root litter, whereas the tree coarse root and under-story species root litter production were excluded.
In the I treatment the above- and below-ground litterproduction were nearly equal (Table 7), both being sur-prisingly close to litter productions published earlier fromFlakaliden (1,200 and 1,140 kg ha−1 yr−1, in above- andbelow-ground, respectively) (Kleja et al. 2008) and threeother sites from Sweden (Hansson et al. 2011, 2013a;Kleja et al. 2008). In all of these sites the ratio betweenthe above- and below-ground litter was 1±0.3.
As the nutrient availability improves, trees need toinvest less in nutrient foraging and more resources arereleased for allocation to the above-ground biomass(Axelsson and Axelsson 1986; Iivonen et al. 2006;Vanninen and Mäkelä 1999; Vogel et al. 2008). The
positive response of fertilization to tree stemwood yieldis well known and documented for silvicultural pur-poses (Bergh et al. 1999; Linder 1987; Tamm 1968),and there is a positive correlation between stand basalarea and above-ground litter production (Hansson et al.2011; Saarsalmi et al. 2007; Starr et al. 2005). InFlakaliden the trees in the WFI and FI were taller, thebasal area larger and the above-ground litter productiondouble compared to the WI and I. Higher soil tempera-ture alone had no effect on the above-ground biomass orthe above-ground litter production (Table 7.).
We hypothesized that both the increased temperatureand the nutrient additions would increase the amount offine root litter produced annually, which was confirmedand was in accordance with several studied (e.g.Brassard et al. 2009 and references therein). The highfine root biomass (Leppälammi-Kujansuu et al. 2013)and high mortality in the mineral soil of both warmingtreatments and in the organic layer of fertilization treat-ment led to increased fine root production: the below-ground litter production was almost threefold in the WIand FI treatments compared to the I, and in the WFI thisdifference was more than fourfold. Mainly due to thesesubstantial increments of the below-ground litter pro-duction, the total annual litter production was 1.8–3.0times higher in the other treatments than in the I. In agradient study in Sweden, the total tree litter productionalso increased towards the south, i.e. towards higherfertility (Kleja et al. 2008).
Our hypothesis, the expectation of the lower below-ground C input to the soil compared to the above-ground,could not be confirmed as the below-ground:above-ground litter production ratio was higher in all othertreatments compared to I. Our result contradicts the litterproduction studies by Gower et al. (1992), Keyes andGrier (1981), Ruess et al. (1996) and Röderstein et al.(2005). The ratio of below-ground:above-ground litterproduction of Norway spruce in different sites inSweden varied without any clear pattern (Hansson et al.2013b; Kleja et al. 2008).
Conclusions
Litter production is affected both by the amount andthe longevity of biomass components. C allocationstudies are often based on differences in above- andbelow-ground biomasses only, ignoring the longevitiesof the components. This should be kept in mind when
84 Plant Soil (2014) 374:73–88

comparing the results of biomass studies with those onC flux (Litton et al. 2007) or biomass production.Furthermore, the amount of C allocated to the treetrunks is substantial but as the trunks in managedforests are harvested, their input to litter production isminimal.
Tree fine roots and their turnover rate have a funda-mental role in determining the response of C input tothe soil via fine root litter in a changing climate. Herewe showed that increased nutrient availability and/orhigher soil temperature significantly shortened fineroot longevity, which led (together with the increasedfine root biomass in these treatments) to three to fourtimes higher below-ground litter production comparedto the reference treatment. In the above-ground, onlyfertilization increased the amount of litter. Thus, thehigher total litter yield in warmed and/or fertilizedtreatments mostly resulted from the substantial incre-ments of production in the below-ground. However, itmust be borne in mind that although substantial, thereported C input to the soil via fine root productionforms only a part of the total below-ground C flux.Root respiration, root exudates, mycorrhizal mycelia,coarse tree roots and understory roots and rhizomeswere not studied in our study but have all been shownto contribute markedly to the below-ground C flux(Högberg et al. 2002; Wallander et al. 2004; Godboldet al. 2006; Hobbie 2006; Kleja et al. 2008), and furtherresearch should also be directed to their quantification.
Two broader conclusions can be drawn from ourstudy of fine root turnover and litter production. First,estimating and modeling the C cycle in boreal forestsin a present and future climate require more attention tothe below-ground compartments. Although long-termtime series are available for above-ground litter pro-duction from several sites available (e.g. Bille-Hansenand Hansen 2001; Starr et al. 2005; Saarsalmi et al.2007), without information about below-ground litterproduction our understanding of the annual C flux intothe soil remains limited. Second, our results show agreat importance of fine root turnover time on rootlitter production and below-ground C flux, and indicatethat models solely based on tree biomass allocationstudies or above-ground parts most likely underesti-mate the below-ground C input to the soil. As there areonly a few reports on combined above- and below-ground litter production, especially from long-termenvironmental manipulation experiments, more effortshould be put into surveying these interactions.
Finally, in addition to determining how much Cflows into the soil from different compartments invarying environmental conditions, future researchshould concentrate on which proportion of the C fluxvia above- and below-ground litter production is actu-ally sequestered into the soil, i.e. transformed intorecalcitrant forms of C, and how this process willchange with changing climate.
Acknowledgements This study was funded by the SwedishResearch Council Formas, the Maj and Tor Nessling Foundationand the Academy of Finland. The authors are grateful to allinvolved in the establishment and maintenance of the Flakalidenresearch site over many years. The minirhizotron tubes used inthis study were originally set by prof. Hooshang Majdi, deceasedin 2007. At the Finnish Forest Research Institute we thank JuhaHeikkinen for statistical assistance and Tauno Suomilammi,Jarmo Mäkinen and Juha Kemppainen for technical assistance.The language of the manuscript was revised by Michael Bailey.
References
Aber JD, Melillo JM, Nadelhoffer KJ, McClaugherty CA, PastorJ (1985) Fine root turnover in forest ecosystems in relationto quantity and form of nitrogen availability: a comparisonof two methods. Oecologia 66:317–321
Andersson P, Berggren D, Nilsson I (2002) Indices for nitrogenstatus and nitrate leaching from Norway spruce (Picea abies(L.) Karst.) stands in Sweden. For Ecol Manag 157:39–53
Axelsson E, Axelsson B (1986) Changes in carbon allocationpatterns in spruce and pine trees following irrigation andfertilization. Tree Physiol 2:189–204
Baddeley JA, Watson CA (2005) Influences of root diameter,tree age, soil depth and season on fine root survivorship inPrunus avium. Plant Soil 276:15–22
Berggren D, Bergkvist B, Johansson M-B, Langvall O, Majdi H,Melkerud P-A, Nilsson Å, Weslien P et al (2004) A de-scription of LUSTRA’s common field sites. Reports inforest ecology and forest soils. Report 87. SwedishUniversity of Agricultural Sciences, Uppsala
Bergh J, Linder S (1999) Effects of soil warming during springon photosynthetic recovery in boreal Norway spruce stands.Glob Change Biol 5:245–253
Bergh J, Linder S, Lundmark T, Elfving B (1999) The effect ofwater and nutrient availability on the productivity of Norwayspruce in northern and southern Sweden. For Ecol Manag119:51–62
Bille-Hansen J, Hansen K (2001) Relation between defoliationand litterfall in some Danish Picea abies and Fagussylvatica stands. Scand J For Res 16:127–137
Brassard BW, Chen HYH, Bergeron Y (2009) Influence ofenvironmental variability on root dynamics in northernforests. Crit Rev Plant Sci 28:179
Burton AJ, Pregitzer KS, Ruess RW, Hendrick RL, Allen MF(2002) Root respiration in North American forests: effects
Plant Soil (2014) 374:73–88 85

of nitrogen concentration and temperature across biomes.Oecologia 131:559–568
Chen HYH, Brassard BW (2013) Intrinsic and extrinsic controlsof fine root life span. Crit Rev Plant Sci 32:151–161
Clemensson-Lindell A, Persson H (1995) Fine-root vitality in aNorway spruce stand subjected to various nutrient supplies.Plant Soil 168–169:167–172
Derome J, Saarsalmi A, Kukkola M (2009) Effects of nitrogen andsulphur “stress” treatment on soil acidity and growth re-sponse of a Scots pine stand. Boreal Environ Res 14:861–874
Eissenstat DM (1992) Costs and benefits of constructing roots ofsmall diameter. J Plant Nutr 15:763–782
Eissenstat DM, Yanai RD (1997) The ecology of root lifespan.In: Begon M, Fitter AH (eds) Advances in ecological re-search, vol 27. Academic, London, pp 1–60
Eissenstat DM, Yanai RD (2002) Root life span, efficiency, andturnover. In: Waisel Y, Eshel A, Kafkafi U (eds) Plantroots—the hidden half. Marcel Dekker, NewYork, pp 221–238
Eissenstat DM,Wells CE, Yanai RD, Whitbeck JL (2000) Buildingroots in a changing environment: implications for root longev-ity. New Phytol 147:33–42
Finér L, Laine J (1998) Root dynamics at drained peatland sites ofdifferent fertility in southern Finland. Plant Soil 201:27–36
Finér L, Ohashi M, Noguchi K, Hirano Y (2011) Fine rootproduction and turnover in forest ecosystems in relation tostand and environmental characteristics. For Ecol Manag262:2008–2023
FröbergM,GripH, Tippings E, SvenssonM, StrömgrenM,KlejaDB(2013) Long-term effects of experimental fertilization and soilwarming on dissolved organic matter leaching from a spruceforest in Northern Sweden. Geoderma 200–201:172–179
Gandahl R (1957) Bestämning av tjälgräns i mark med enkel typav tjälgränsmätare. Grundförbättring 10:7–19 (in Swedish)
Gaul D, Hertel D, Leuschner C (2009) Estimating fine rootlongevity in a temperate Norway spruce forest using threeindependent methods. Funct Plant Biol 36:11–19
Godbold DL, Hoosbeek MR, Lukac M et al (2006) Mycorrhizalhyphal turnover as a dominant process for carbon input intosoil organic matter. Plant Soil 281:15–24
Gower ST, Vogt KA, Grier CC (1992) Carbon dynamics ofRocky Mountain Douglas-fir: influence of water and nutri-ent availability. Ecol Monogr 62:43–65
Guo DL, Mitchell RJ, Withington JM, Fan P-P, Hendricks JJ(2008) Endogenous and exogenous controls of root lifespan, mortality and nitrogen flux in a longleaf pine forest:root branch order predominates. J Ecol 96:737–745
Hägglund B, Lundmark J-E (1977) Site index estimation bymeans of site properties of Scots pine and Norway sprucein Sweden. Stud For Suec 138:1–33
Hansson K, Olsson BA, Olsson M, Johansson U, Kleja DB(2011) Differences in soil properties in adjacent stands ofScots pine, Norway spruce and silver birch in SW Sweden.For Ecol Manag 262:522–530
Hansson K, Helmisaari H-S, Sah SP, Lange H (2013a) Fine rootproduction and turnover of tree and understorey vegetationin Scots pine, silver birch and Norway spruce stands in SWSweden. For Ecol Manag, In press
Hansson K, Fröberg M, Helmisaari H-S, Kleja DB, Olsson BA,Olsson M, Persson T (2013b) Carbon and nitrogen pools andfluxes above and below ground in spruce, pine and birchstands in southern Sweden. For Ecol Manag, In press
Helmisaari H-S, Makkonen K, Kellomäki S, Valtonen E,Mälkönen E (2002) Below- and above-ground biomass,production and nitrogen use in Scots pine stands in easternFinland. For Ecol Manag 165:317–326
Helmisaari H-S, Derome J, Nöjd P, Kukkola M (2007) Fine rootbiomass in relation to site and stand characteristics in Norwayspruce and Scots pine stands. Tree Physiol 27:1493–1504
Hendrick RL, Pregitzer KS (1993) Patterns of fine root mortalityin two sugar maple forests. Nature 361:59–61
Hobbie EA (2006) Carbon allocation to ectomycorrhizal fungicorrelates with below-ground allocation in culture studies.Ecology 87:563–569
Högberg P, Nordgren A, Ågren GI (2002) Carbon allocationbetween tree root growth and root respiration in boreal pineforest. Oecologia 132:579–581
Hyvönen R, Ågren GI, Linder S et al (2007) The likely impact ofelevated [CO2], nitrogen deposition, increased temperatureand management on carbon sequestration in temperate andboreal forest ecosystems: a literature review. New Phytol173:463–480
Idol TW, Pope PE, Ponder F Jr (2000) Fine root dynamics acrossa chronosequence of upland temperate deciduous forests.For Ecol Manag 127:153–167
Iivonen S, Kaakinen S, Jolkkonen A, Vapaavuori E, Linder S(2006) Influence of long-term nutrient optimization on bio-mass, carbon, and nitrogen acquisition and allocation inNorway spruce. Can J For Res 36:1563–1571
IPCC (2007) The physical science basis. In: Solomon S, Qin D,Manning M, Chen Z, Marquis M, Averyt KB, Tignor M,Miller HL (eds) Contribution of working group I to the fourthassessment report of the intergovernmental panel on climatechange. Cambridge University Press, Cambridge and NewYork
Jarvis P, Linder S (2000) Constraints to growth of boreal forests.Nature 405:904–905
Joslin JD, Wolfe MH (1999) Disturbances during minirhizotroninstallation can affect root observation data. Soil Sci SocAm J 63:218–221
Keyes MR, Grier CC (1981) Above- and below-ground netproduction in 40-year-old Douglas-fir stands on low andhigh productivity sites. Can J For Res 11:599–605
King JS, Albaugh TJ, Allen HL, Buford M, Strain BR,Dougherty P (2002) Below-ground carbon input to soil iscontrolled by nutrient availability and fine root dynamics inloblolly pine. New Phytol 154:389–398
Kleja DB, Svensson M, Majdi H et al (2008) Pools and fluxes ofcarbon in three Norway spruce ecosystems along a climaticgradient in Sweden. Biogeochemistry 89:7–25
Kukkola M, Saramäki J (1983) Growth response in repeatedlyfertilized pine and spruce stands on mineral soils. CommunInst For Fenn 144:1–55
Lal R (2005) Forest soils and carbon sequestration. For EcolManag 220:242–258
Leppälammi-Kujansuu J, Ostonen I, Strömgren M, Nilsson LO,Kleja DB, Sah SP, Helmisaari H-S (2013) Effects of long-term temperature and nutrient manipulation on Norwayspruce fine roots and mycelia production. Plant Soil366:287–303
Linder S (1987) Responses to water and nutrients in coniferousecosystems. In: Schulze E, Zwölfer H (eds) Ecologicalstudies, vol 61. Springer, Berlin Heidelberg, pp 180–202
86 Plant Soil (2014) 374:73–88

Linder S (1995) Foliar analysis for detecting and correcting nutri-ent imbalances in Norway spruce. Ecol Bull (Copenhagen)44:178–190
Linder S, Flower-Ellis JGK (1992) Environmental and physio-logical constraints to forest yield. In: Teller A, Mathy P,Jeffers JNR (eds) Responses of forest ecosystems to envi-ronmental changes. Elsevier Applied Science, London, pp149–164
Litton CM, Raich JW, Ryan MG (2007) Carbon allocation inforest ecosystems. Glob Change Biol 13:2089–2109
López B, Sabaté S, Gracia CA (2001) Fine-root longevity ofQuercus ilex. New Phytol 151:437–441
Lukac M, Calfapietra C, Godbold DL (2003) Production, turn-over and mycorrhizal colonization of root systems of threePopulus species grown under elevated CO2 (POPFACE).Glob Change Biol 9:838–848
Majdi H (2001) Changes in fine root production and longevity inrelation to water and nutrient availability in a Norwayspruce stand in northern Sweden. Tree Physiol 21:1057–1061
Majdi H, Andersson P (2005) Fine root production and turnoverin a Norway spruce stand in Northern Sweden: effects ofnitrogen and water manipulation. Ecosystems 8:191–199
Majdi H, Nylund J-E (1996) Does liquid fertilization affect fineroot dynamics and lifespan of mycorrhizal short roots?Plant Soil 185:305–309
Majdi H, Öhrvik J (2004) Interactive effects of soil warming andfertilization on root production, mortality, and longevity ina Norway spruce stand in Northern Sweden. Glob ChangeBiol 10:182–188
Mäkelä A, Valentine HT, Helmisaari H-S (2008) Optimal co-allocation of carbon and nitrogen in a forest stand at steadystate. New Phytol 180:114–123
Marshall JD, Waring RH (1985) Predicting fine root productionand turnover by monitoring root starch and soil tempera-ture. Can J For Res 15:791–800
McCormack LM, Adams TS, Smithwick EAH, Eissenstat DM(2012) Predicting fine root lifespan from plant functionaltraits in temperate trees. New Phytol 195:823–831
Menzel A, Fabian P (1999) Growing season extended in Europe.Nature 397:659
Nadelhoffer KJ, Aber JD, Melillo JM (1985) Fine roots, netprimary production, and soil nitrogen availability: a newhypothesis. Ecology 66:1377–1390
Ostonen I, Helmisaari H-S, Borken W et al (2011) Fine rootforaging strategies in Norway spruce forests across aEuropean climate gradient. Glob Change Biol 17:3620–3632
Peterjohn WT, Melillo JM, Steudler PA, Newkirk KM, BowlesFP, Aber JD (1994) Responses of trace gas fluxes and Navailability to experimentally elevated soil temperatures.Ecol Appl 4:617–625
Pregitzer KS, Zak DR, Curtis PS, Kubiske ME, Teeri JA, VogelCS (1995) Atmospheric CO2, soil nitrogen and turnover offine roots. New Phytol 129:579–585
Pregitzer KS, DeForest JL, Burton AJ, Allen MF, Ruess RW,Hendrick RL (2002) Fine root architecture of nine NorthAmerican trees. Ecol Monogr 72:293–309
Röderstein M, Hertel D, Leuschner C (2005) Above- and below-ground litter production in three tropical montane forests insouthern Ecuador. J Trop Ecol 21:483–492
Ruess RW, Cleve KV, Yarie J, Viereck LA (1996) Contributionsof fine root production and turnover to the carbon andnitrogen cycling in taiga forests of the Alaskan interior.Can J For Res 26:1326–1336
Ruess RW,Hendrick RL, BurtonAJ, Pregitzer KS, SveinbjornssönB,Allen MF, Maurer GE (2003) Coupling root dynamics withecosystem carbon cycling in black spruce forests of interiorAlaska. Ecol Monogr 73:643–662
Rustad LE, Campbell JL, Marion GM et al (2001) A meta-analysis of the response of soil respiration, net nitro-gen mineralization, and above-ground plant growth toexperimental ecosystem warming. Oecologia 126:543–562
Rydén BE, Kostov L (1980) Thawing and freezing in tundrasoils. Ecol Bull (Stockholm) 30:251–280
Saarsalmi A, Starr M, Hokkanen T, Ukonmaanaho L, KukkolaM, Nöjd P, Sievänen R (2007) Predicting annual canopylitterfall production for Norway spruce (Picea abies (L.)Karst.) stands. For Ecol Manag 242:578–586
Schindlbacher A, Zechmeister-Boltenstern S, Jandl R (2009)Carbon losses due to soil warming: do autotrophic and het-erotrophic soil respiration respond equally? Glob ChangeBiol 15:901–913
Schoettle AW, Fahey TJ (1994) Foliage and fine root longevityof pines. Ecol Bull (Stockholm) 43:136–153
Sjörs H (1999) The background: geology, climate and zonation.Acta Phytogeogr Suec 84:5–14
Soil Survey Staff (1999) Soil taxonomy. A basic system of soilclassification for making and interpreting soil surveys.USDA, Washington CD
Starr M, Saarsalmi A, Hokkanen T, Merilä P, Helmisaari H-S(2005) Models of litterfall production for Scots pine (Pinussylvestris L.) in Finland using stand, site and climate fac-tors. For Ecol Manag 205:215–225
Strand AE, Pritchard SG, McCormack ML, Davis MA, Oren R(2008) Irreconcilable differences: fine-root life spans andsoil carbon persistence. Science 319:456–458
Strömgren M (2001) Soil-surface CO2 flux and growth in a borealNorway spruce stand—effects of soil warming and nutrition.Acta Universitatis Agriculturae Sueciae, Silvestria 220,Doctoral thesis. Swedish University of Agricultural Sciences,Uppsala
Strömgren M, Linder S (2002) Effects of nutrition and soilwarming on stemwood production in a boreal Norwayspruce stand. Glob Change Biol 8:1194–1204
Tamm CO (1968) An attempt to assess the optimum nitrogenlevel in Norway spruce under field conditions. Stud ForSuec 61:1–67
Ukonmaanaho L, Merilä P, Nöjd P, Nieminen TM (2008)Litterfall production and nutrient return to the forest floorin Scots pine and Norway spruce stands in Finland. BorealEnviron Res 13:67–91
Valentine HT, Mäkelä A (2012) Modeling forest stand dynamicsfrom optimal balances of carbon and nitrogen. New Phytol194:961–971
van Praag HJ, Sougnez-Remy S, Weissen F, Carletti G (1988)Root turnover in a beech and a spruce stand of the BelgianArdennes. Plant Soil 105:87–103
Vanninen P, Mäkelä A (1999) Fine root biomass of Scots pinestands differing in age and soil fertility in southern Finland.Tree Physiol 19:823–830
Plant Soil (2014) 374:73–88 87

Vanninen P, Ylitalo H, Sievänen R, Mäkelä A (1996) Effects ofage and site quality on the distribution of biomass in Scotspine (Pinus sylvestris L.). Trees - Struct Funct 10:231–238
Vogel JG, Bond-Lamberty BP, Schuur EAG, Gower ST, MackMC, O'Connell KEB, Valentine DW, Ruess RW (2008)Carbon allocation in boreal black spruce forests acrossregions varying in soil temperature and precipitation. GlobChange Biol 14:1503–1516
Vogt KA, Grier CC, Vogt DJ (1986) Production, turnover, andnutrient dynamics of above- and below-ground detritus ofworld forests. Adv Ecol Res 15:303–377
Wallander H, Göransson H, Rosengren U (2004) Production,standing biomass and natural abundance of 15N and 13C inectomycorrhizal mycelia collected at different soil depths intwo forest types. Oecologia 139:89–97
Wells CE, Eissenstat DM (2001) Marked differences in survi-vorship among apple roots of different diameters. Ecology82:882–892
Withington JM, Reich PB, Oleksyn J, Eissenstat DM(2006) Comparisons of structure and life span in rootsand leaves among temperate trees. Ecol Monogr76:381–397
88 Plant Soil (2014) 374:73–88
Recommended

















