
Enzymes and enzyme kinetics

• enzyme "in yeast"
• proteins and catalysts
• 1860: L. Pasteur "fermentation catalyzed by enzymes"
• 1897: E. Buchner demonstrated alcohol production independent of cells
• 1926: J.B. Sumner isolated pure crystalline urease
milestones

urease
NH2
NH2
C O + H2O 2NH3 + CO2
• accelerates reaction 1014 fold• average for enzymes is 108 – 1020 fold

Don't you hate definitions
• enzyme (catalyst) is substance that increases the rate of chemical reaction without undergoing permanent change itself
• influences rate only, not equilibrium
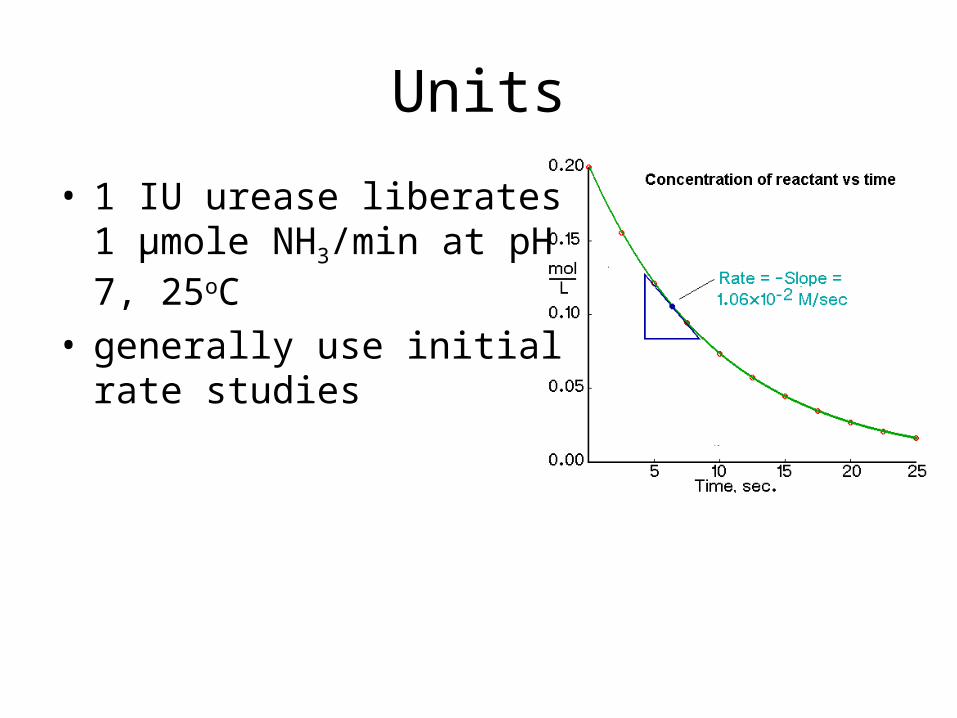
Units
• 1 IU urease liberates 1 μmole NH3/min at pH 7, 25oC
• generally use initial rate studies

Free energy of activation
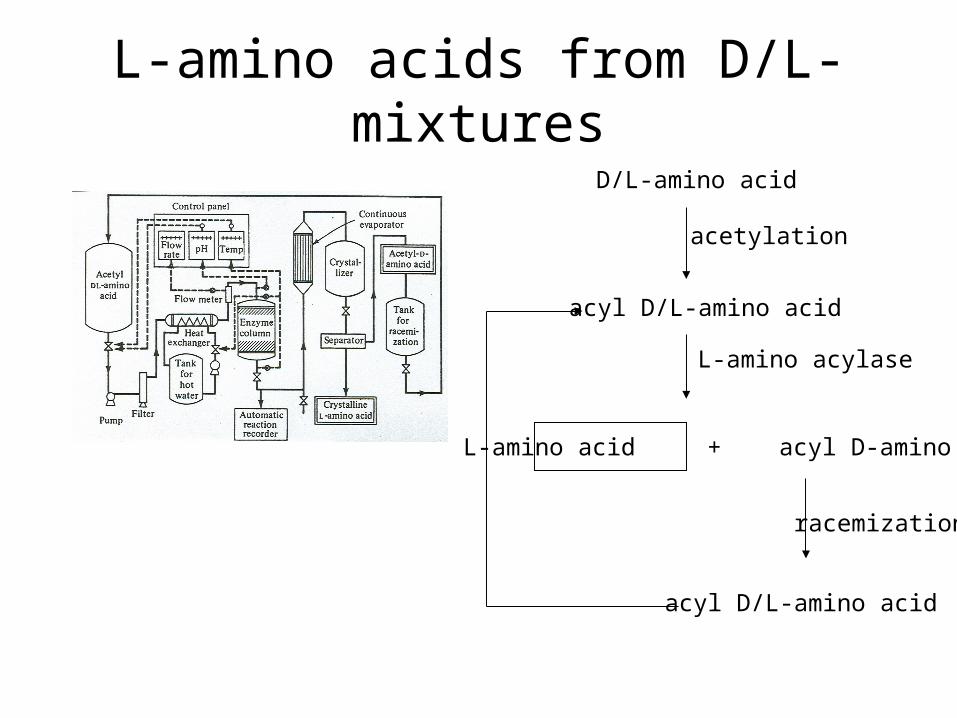
L-amino acids from D/L-mixtures
D/L-amino acid
acyl D/L-amino acid
L-amino acid + acyl D-amino acid
acyl D/L-amino acid
acetylation
L-amino acylase
racemization

Enzyme electrodes
Type Enzyme Sensor Stable Time
urea urease pH, NH3 3 weeks few min
glucose glu oxidase pH, O2 wk-months few min
L-glutamate glut dehydrog. various days-wks 1-2 min
lactic acid lact dehydrog. Pt(Fe(CN)) 1 week 3-10 m
alcohols alc oxidase Pt(O2) day-month >1 min
penicillin penicillinase pH 2-3 weeks 2 min
cholesterolchol oxidase Pt(H2O2) 2 min
uric acid uricase Pt(O2) 4 month 30 sec

YSI biochemistry analyzer•glucose, sucrose•lactose•L-lactate•galactose•L-glutamate•choline•L-glutamine•ethanol•hydrogen peroxide•starch
typical enzyme electrode
H5C2 CH2
O
C NH
O
CH
C
CH3
CH3
O
OHC
CH
N CH
CS
NH2
O
CH
C
CH3
CH3
O
OHC
CH
N CH
CS
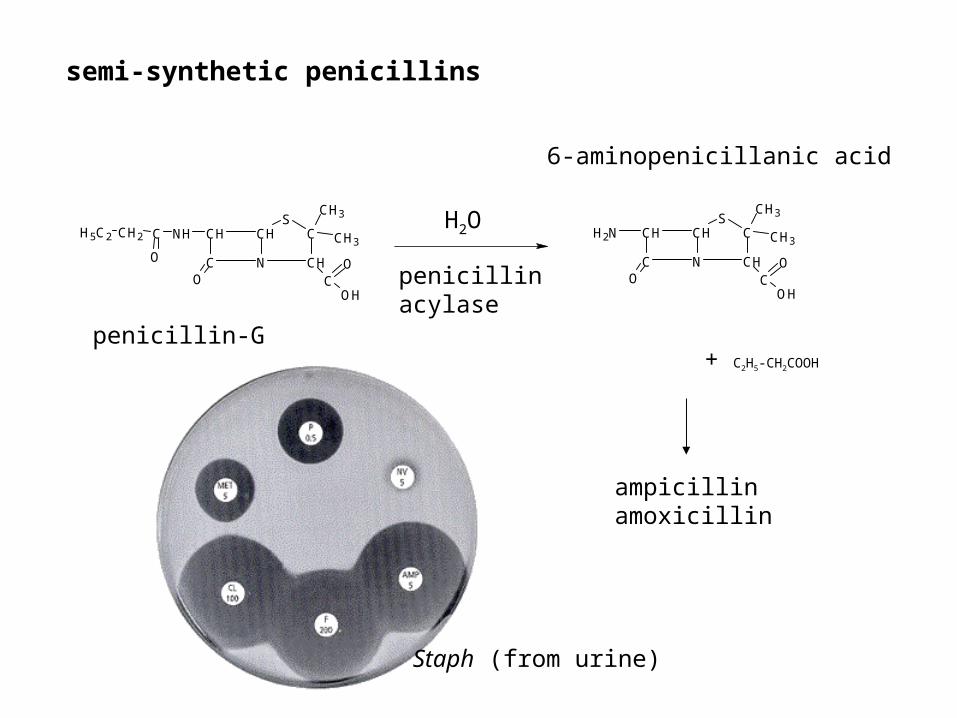
penicillinacylase
H2O
+ C2H5-CH2COOH
penicillin-G
6-aminopenicillanic acid
ampicillinamoxicillin
Staph (from urine)
semi-synthetic penicillins
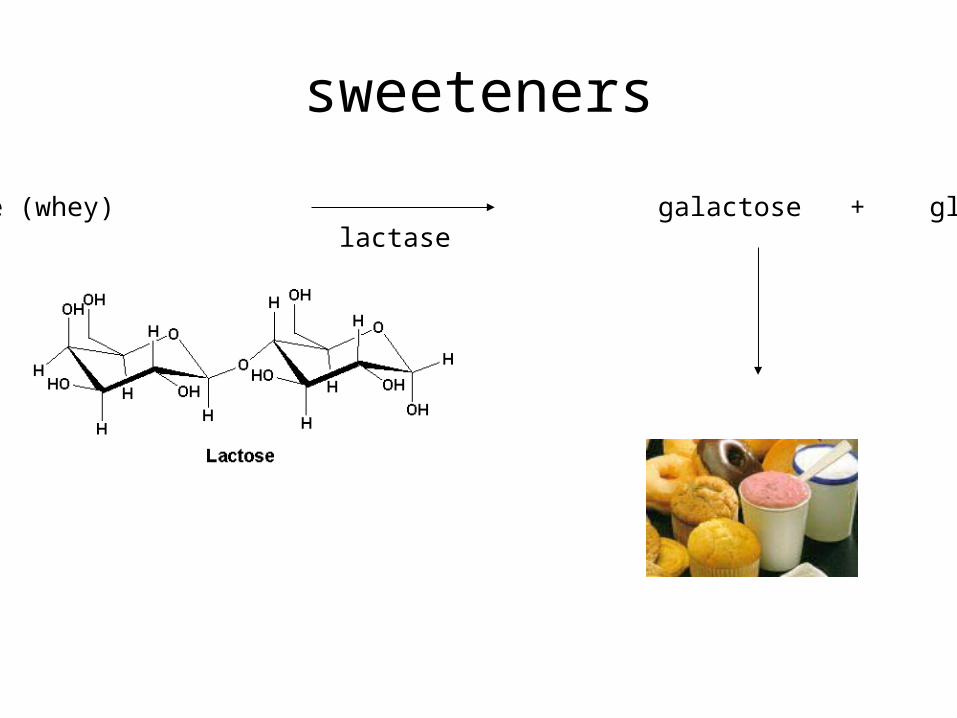
sweeteners
lactose (whey) galactose + glucoselactase

heparinase I fromFlavobacterium heparinum
neutralization of heparin in blood and plasma

enzyme kinetic considerations
• rate of reaction (catalytic activity)
• extent of reaction (equilibrium constant)
• duration of usable activity (stability)
• cost

Michaelis Menten kinetic model (1913)
enzyme + reactant enzyme-reactant complex enzyme + product
E + R ER E + Pk1 k2
k-1
Assumptions:• steady state [ER] = constant, d[ER]/dt = 0• single reactant R• [R] >> [E] and [R] >> [ER]• neglect k-2

E + R ER E + Pk1 k2
k-1
from assumption 1 (steady state)
[R] K
][R]E[ ER][
dt
d[P] v rate
[R] K
][R][E [ER]
[ER] - ][E [E]
constant [E] [ER] [E]
K[ER]
[E][R]
[ER])( [E][R]
0 [ER])( - [E][R]dt
d[ER]
m
o22
m
o
o
o
m1
21
211
211
kk
k
kk
kkk
kkk
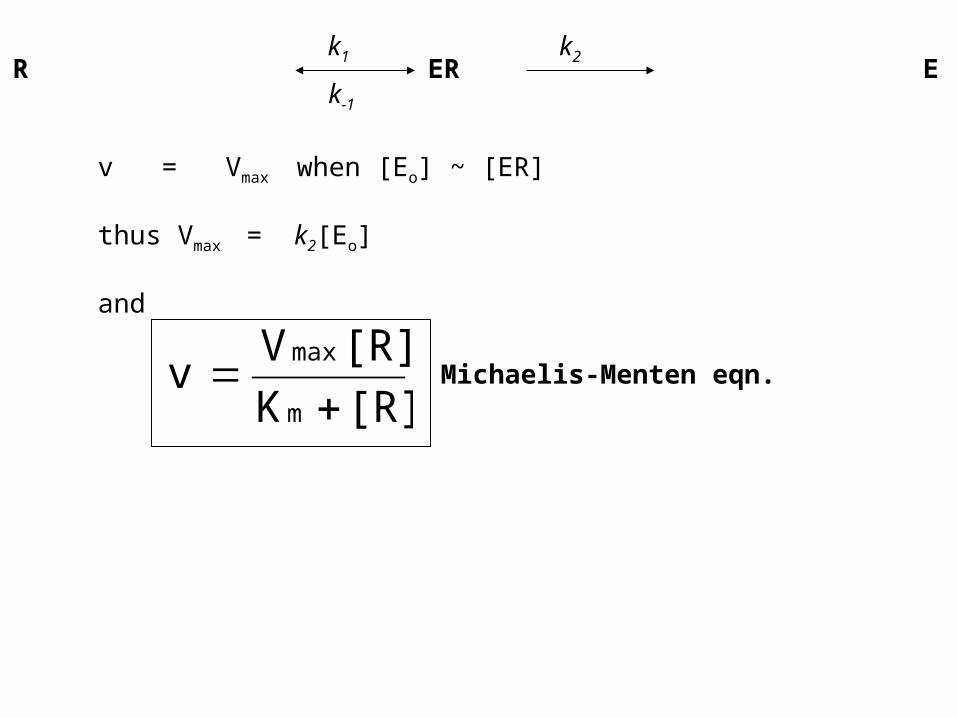
E + R ER E + Pk1 k2
k-1
v = Vmax when [Eo] ~ [ER]
thus Vmax = k2[Eo]
and
[R] K
[R] V v
m
max
Michaelis-Menten eqn.

ctc
t
k
kkK
ER
REK
Kk
kK
kk
K
m
s
sm
m
11
1
1
21
1
1
21
-1
Units,
][
]][[ where
complex ERfor constant on dissociati the,~then
reactions, somein
substrate)for enzyme of(affinity ~
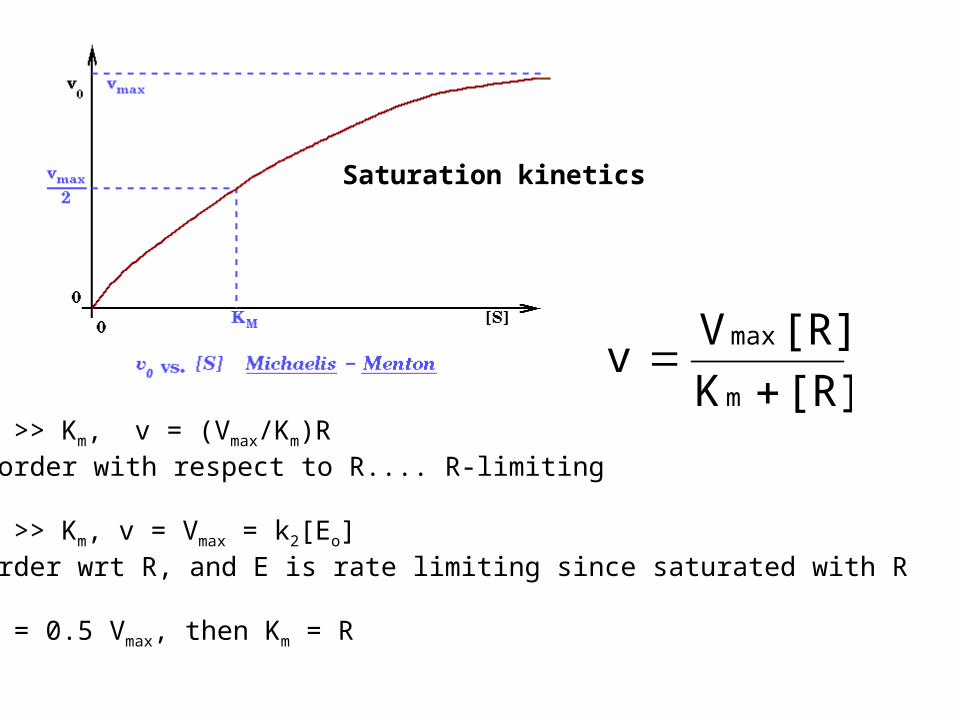
Saturation kinetics
When R >> Km, v = (Vmax/Km)Rfirst order with respect to R.... R-limiting
When R >> Km, v = Vmax = k2[Eo]zero order wrt R, and E is rate limiting since saturated with R
When v = 0.5 Vmax, then Km = R
[R] K
[R] V v
m
max
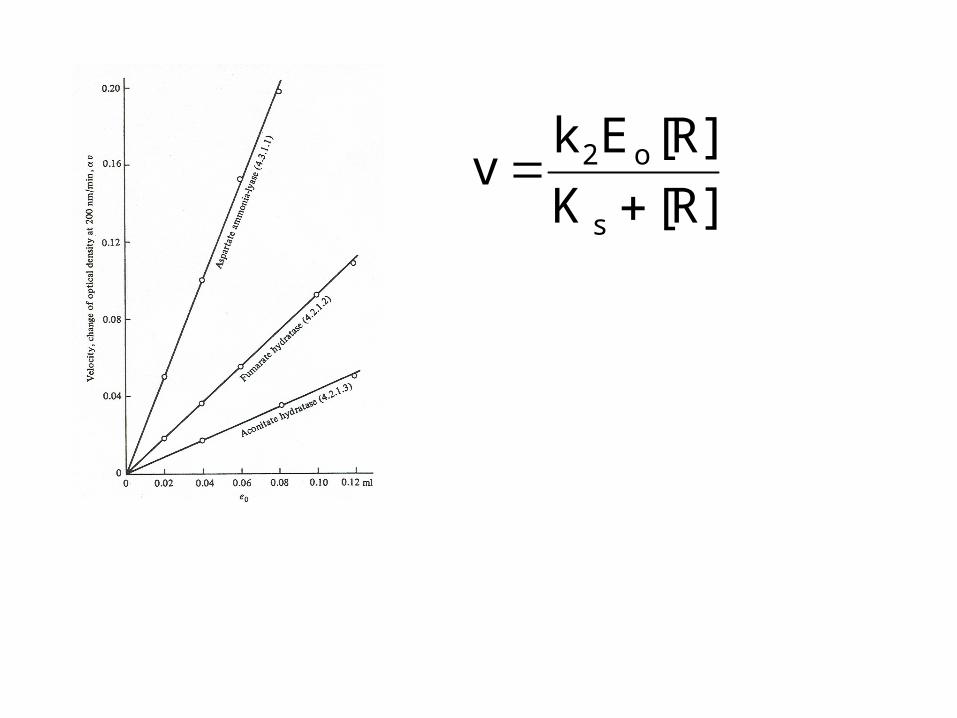
R][K
R][Ekv
s
o2
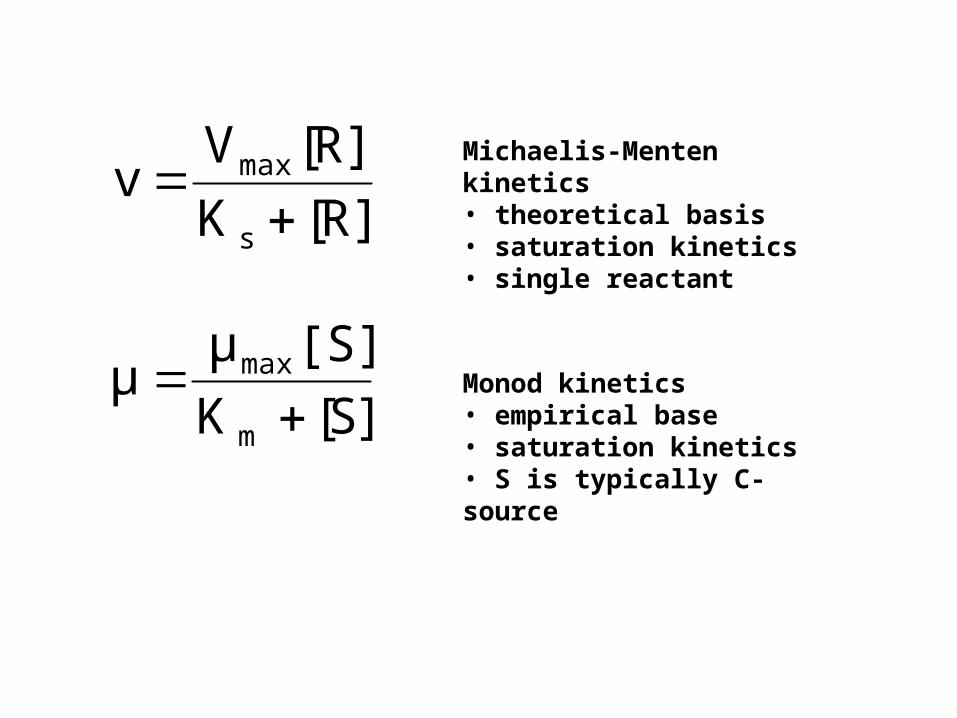
Michaelis-Menten kinetics• theoretical basis• saturation kinetics• single reactant
Monod kinetics• empirical base• saturation kinetics• S is typically C-source]S[K
[S]μμ
m
max
R][K
R][Vv
s
max

double reciprocal plot (Lineweaver Burk plot)
maxmax
m
V
1
R
1
V
K
v
1
max
m
max
maxm
V
K
V
R
v
R
VR
vKv
Eadie-Hofstee plot
Hanes plotLineweaver-Burkplot
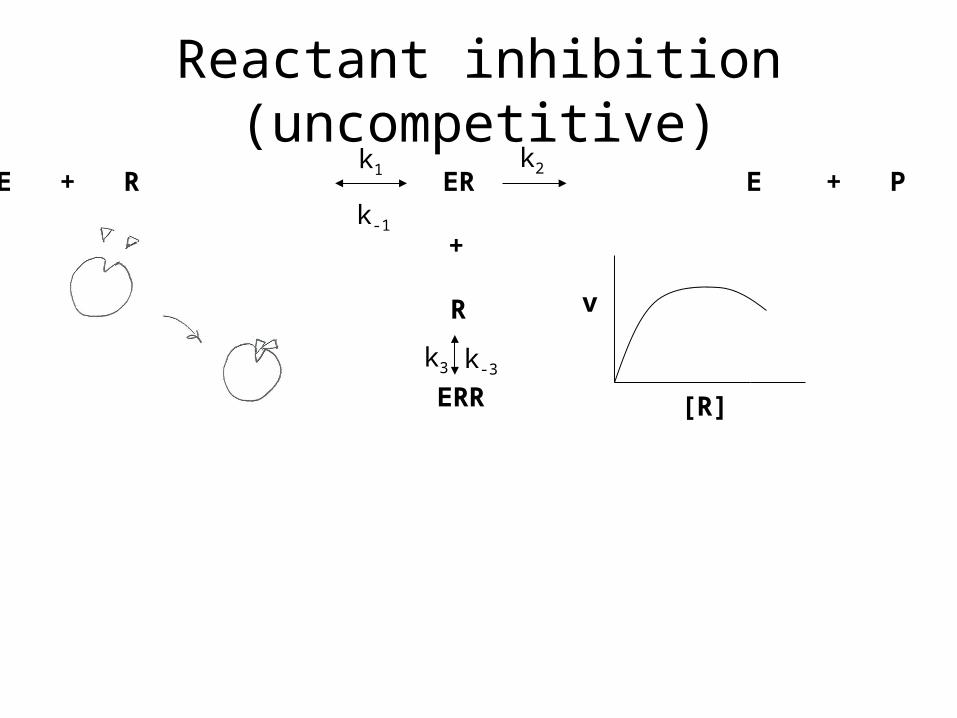
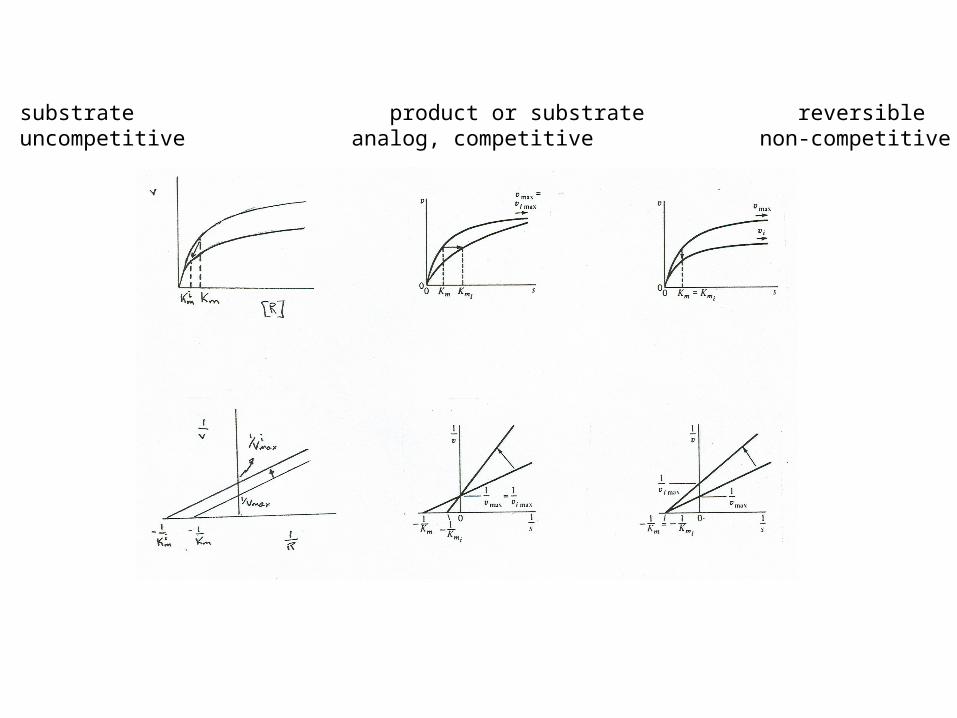
Reactant inhibition (uncompetitive)
E + R ER E + P
+
R
ERR
k1
k-1
k2
k3 k-3
v
[R]
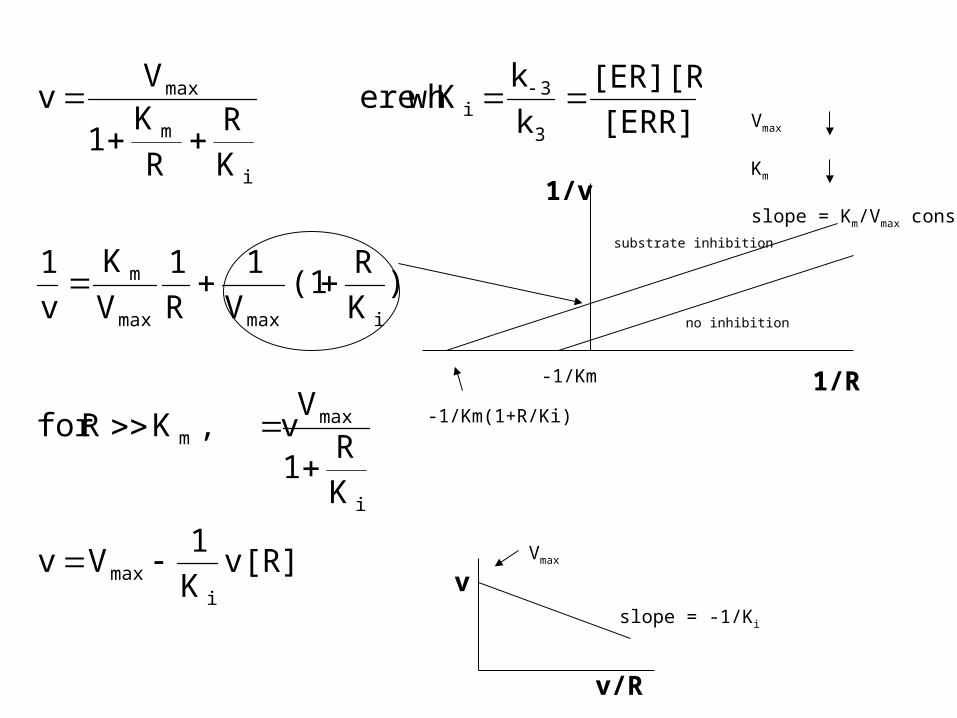
v[R]K
1Vv
K
R1
V v,K Rfor
)K
R(1
V
1
R
1
V
K
v
1
[ERR]
[ER][R]
k
kK ere wh
K
R
R
K1
Vv
imax
i
maxm
imaxmax
m
3
3i
i
m
max
1/v
1/R
no inhibition
substrate inhibition
-1/Km
-1/Km(1+R/Ki)
Vmax
Km
slope = Km/Vmax const
v
v/R
slope = -1/Ki
Vmax
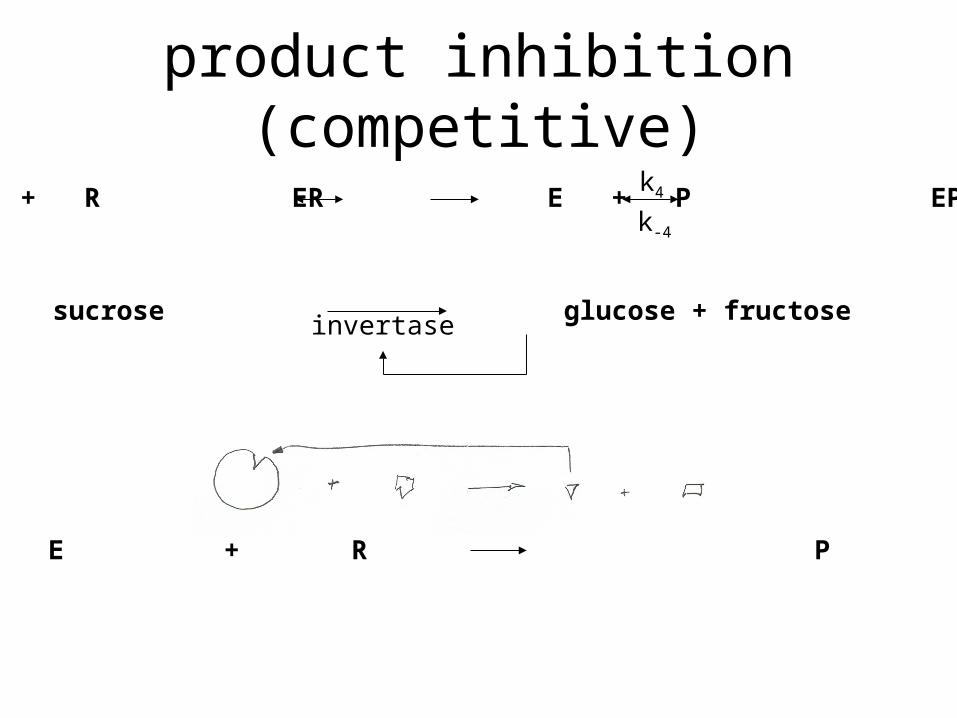
product inhibition (competitive)
E + R ER E + P EPk4
k-4
sucrose glucose + fructoseinvertase
E + R P
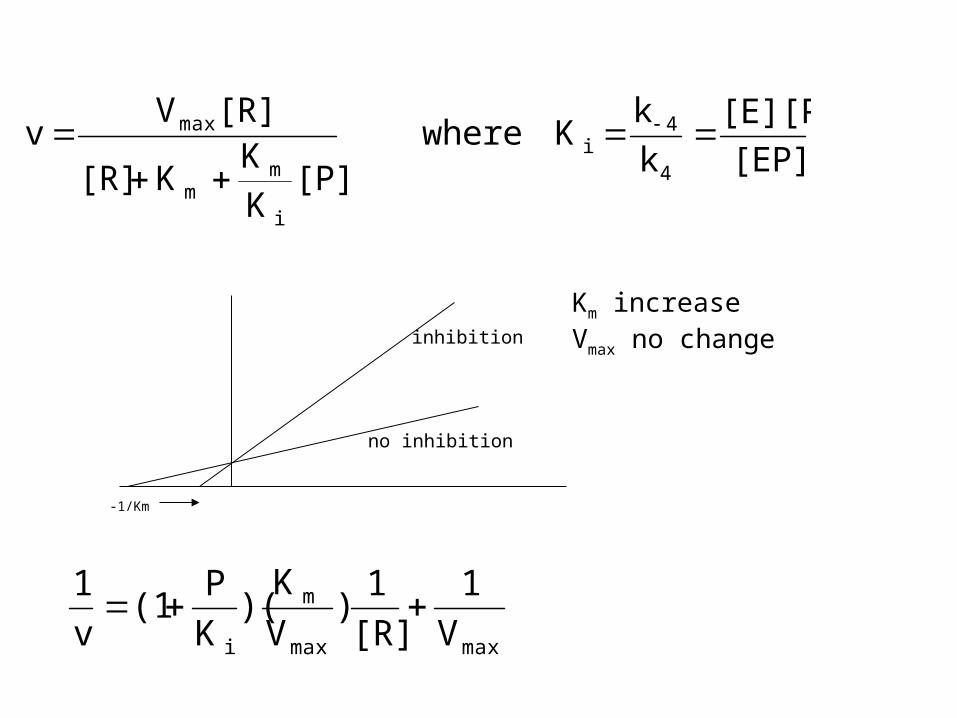
[EP]
[E][P]
k
kK where
[P]K
KK[R]
[R]Vv
4
4i
i
mm
max
no inhibition
inhibition
-1/Km
Km increaseVmax no change
maxmax
m
i V
1
[R]
1)
V
K)(
K
P(1
v
1

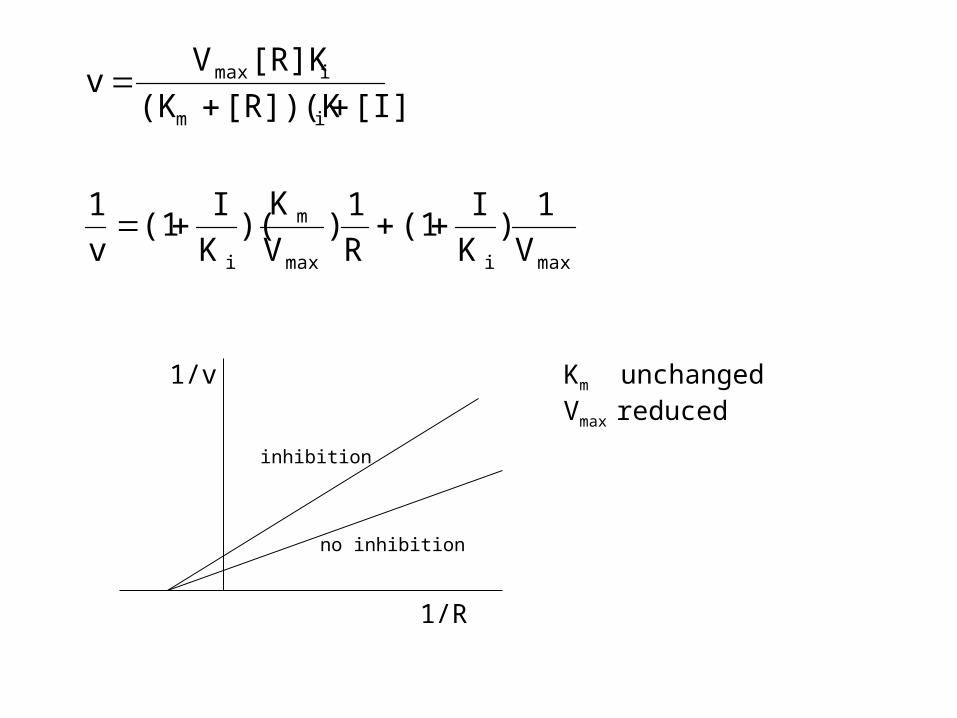
non-competitive inhibition
• urease-Ni + EDTA (chelator)
• heavy metal toxicity, binding to disulfide bridges (cysteine)
• organic acids (acetate, propionate, lactate on hydrolytic enzymes
• inhibitor acts at another site on the enzyme, but changes configuration
maximax
m
i
im
imax
V
1)
K
I(1
R
1)
V
K)(
K
I(1
v
1
[I][R])(K(K
[R]KVv
no inhibition
inhibition
1/v
1/R
Km unchangedVmax reduced
substrate product or substrate reversibleuncompetitive analog, competitive non-competitive

T-effects
RT
EAln kln
eqn. Arrhenius )RT
EAexp(k
][EkV
2
2
o2max
activation
inactivation
ln vorln Vmax
orln k2
1/T (kelvin)
Recommended

















