
XRB. UNIVERSIDAD DE BARCELONA
13 JULIO 2012
LUIS ENJUANES
CNB-CSIC. MADRID
MOLECULAR BASIS OF CORONAVIRUS VIRULENCE AND VACCINE DESIGN TO
PROTECT AGAINST SARS
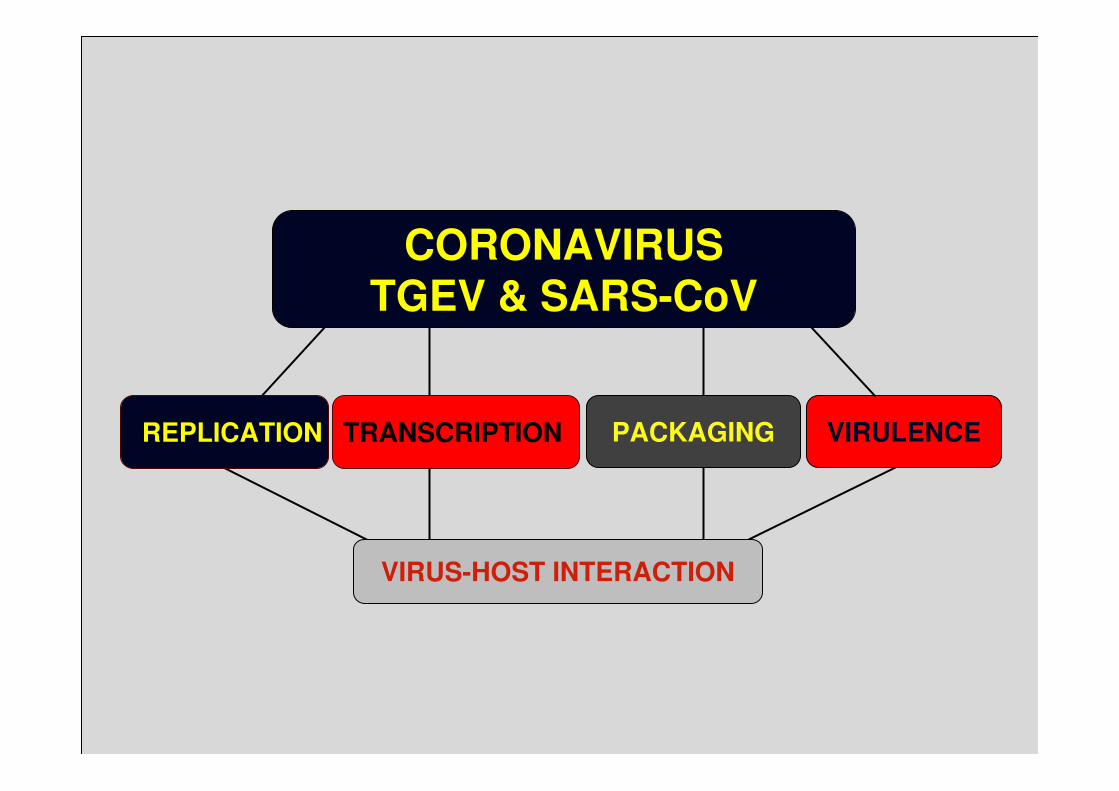
VIRULENCE
CORONAVIRUSTGEV & SARS-CoV
PACKAGINGTRANSCRIPTION REPLICATION
VIRUS-HOST INTERACTION
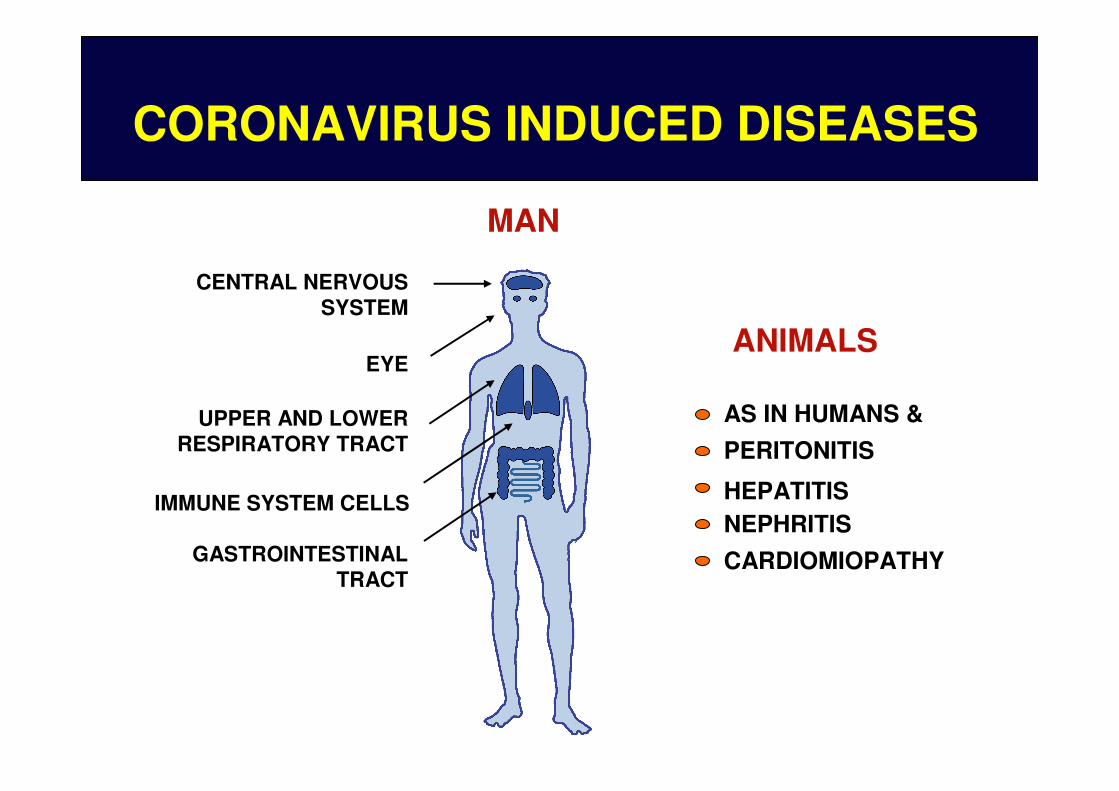
CORONAVIRUS INDUCED DISEASES
MAN
ANIMALS
AS IN HUMANS &
PERITONITIS
HEPATITISNEPHRITIS
CARDIOMIOPATHY
CENTRAL NERVOUSSYSTEM
EYE
UPPER AND LOWERRESPIRATORY TRACT
IMMUNE SYSTEM CELLS
GASTROINTESTINALTRACT
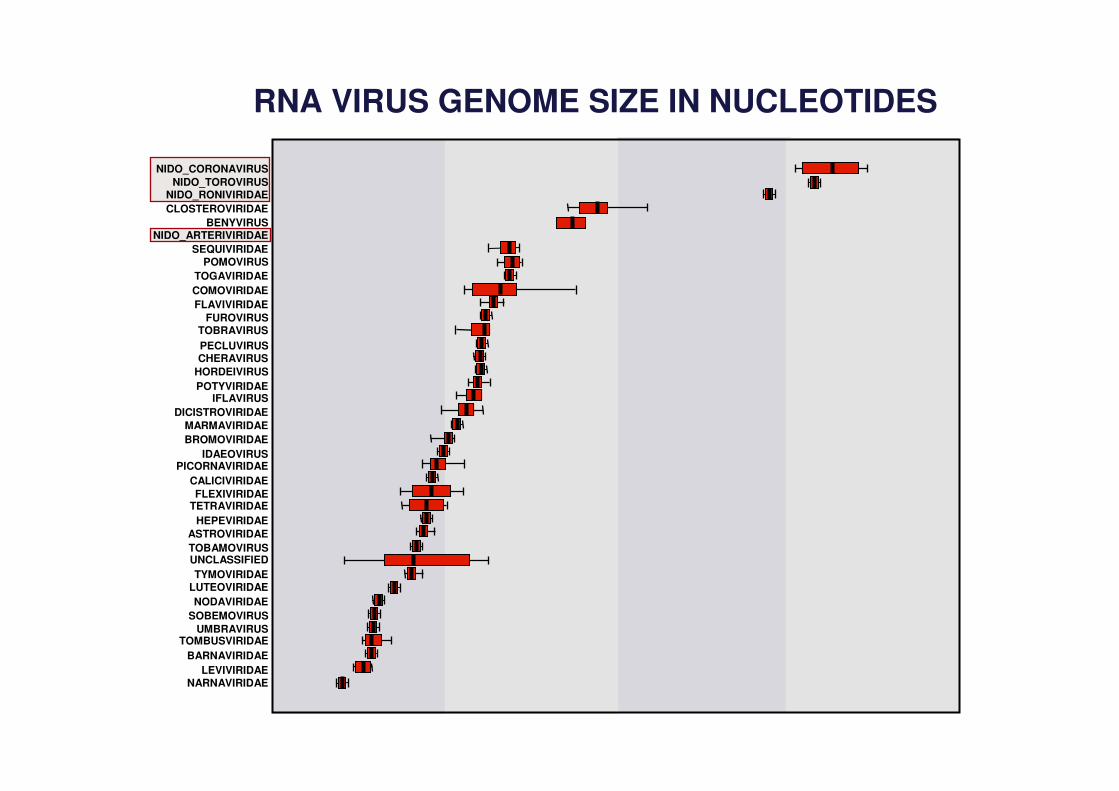
RNA VIRUS GENOME SIZE IN NUCLEOTIDES
NIDO_CORONAVIRUSNIDO_TOROVIRUS
NIDO_RONIVIRIDAECLOSTEROVIRIDAE
BENYVIRUSNIDO_ARTERIVIRIDAE
SEQUIVIRIDAEPOMOVIRUS
TOGAVIRIDAECOMOVIRIDAEFLAVIVIRIDAE
FUROVIRUSTOBRAVIRUSPECLUVIRUSCHERAVIRUS
HORDEIVIRUSPOTYVIRIDAE
IFLAVIRUSDICISTROVIRIDAE
IDAEOVIRUS
MARMAVIRIDAEBROMOVIRIDAE
PICORNAVIRIDAE
ASTROVIRIDAE
CALICIVIRIDAEFLEXIVIRIDAE
TETRAVIRIDAEHEPEVIRIDAE
TYMOVIRIDAE
TOBAMOVIRUSUNCLASSIFIED
LUTEOVIRIDAENODAVIRIDAE
SOBEMOVIRUS
TOMBUSVIRIDAEUMBRAVIRUS
BARNAVIRIDAELEVIVIRIDAE
NARNAVIRIDAE
GFDVEGC-111-RTCCLCDK-11-YACWNHS-3-DYVYNPFMIDVQ-23-HVASCDAIMTR
GMDVEGA-111-KHCQ-CGT-11-YCCFKHA-3-DYVYNPYVIDIQ-23-HVASGDAIMTR
GFDVEGA-111-QKCE-CGK-11-YACFKHA-3-DYLYNPYCIDIQ-23-HVASGDAIMTR
GFDVEAT-111-QVCS-CGS-11-YACWKHC-3-DFVYNPLLVDIQ-23-HVASVDAIMTR
AIDAEFV-103- VFVGHGLNNDFK-44-HDSIEDAHTAL
GFDAEGA-111-VVCSVCTK-11-YGCWRHS-3-DYLYNPLIVDIQ-23-HVASSDAIMTR
YEAST
SARS-CoV
TGEV
IBV
HCoV-229E
MHV-A59
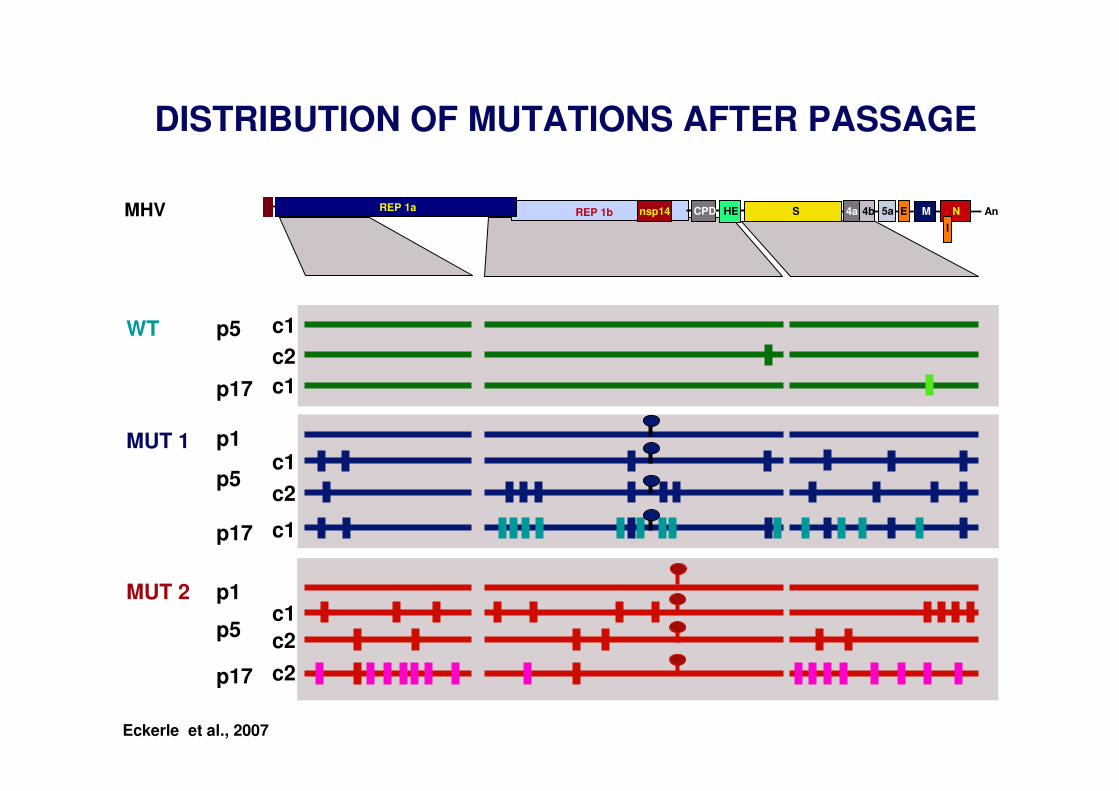
.. ..MUT 1 MUT 2
REP 1a REP 1b An4a 4b E M NSCPD
I
5aHEMHV
C1 D89 E91 ZN F D242 D272 Q521
nsp14
Zn F
Eckerle et al., 2007, 2010
MUTATION OF CORONAVIRUS EXON MOTIFS
c1
c1
c2
c1
c2
c2
c1
c1
c2
WT p5
p17
p1
p5
p17
p1
p5
p17
MUT 1
MUT 2
REP 1a REP 1b An4a 4b E M NSCPD
I
5aHEMHV nsp14
DISTRIBUTION OF MUTATIONS AFTER PASSAGE
Eckerle et al., 2007
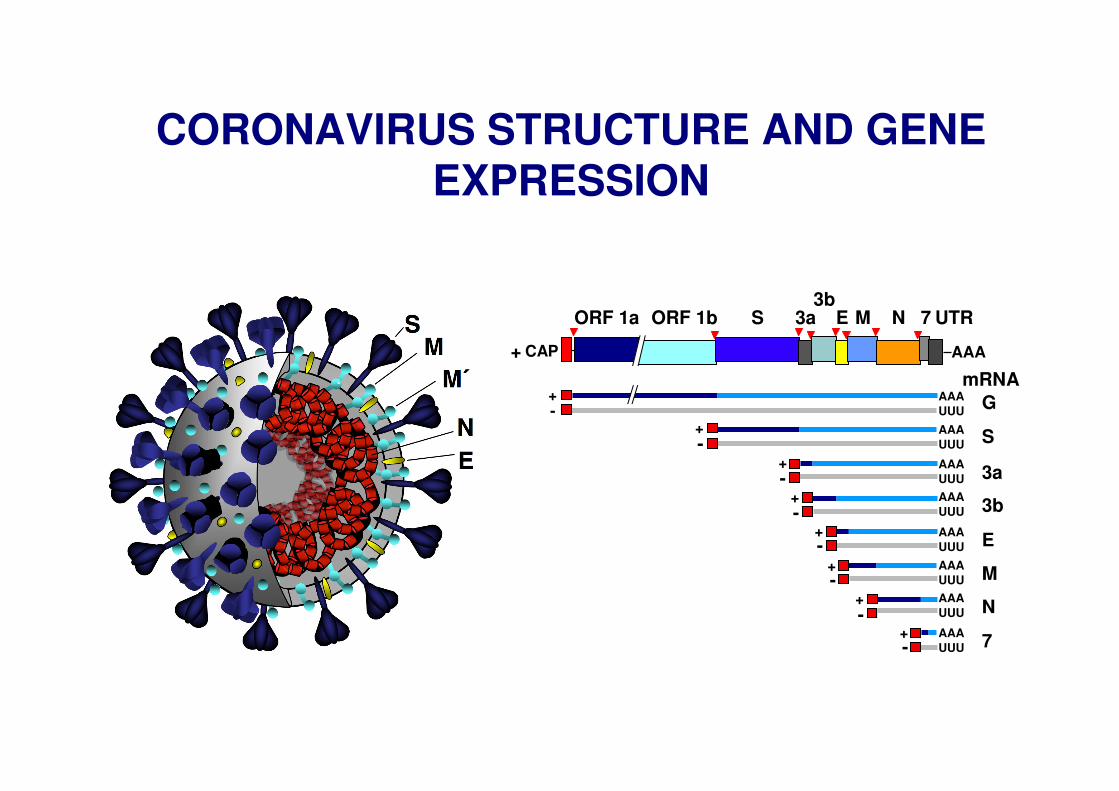
CORONAVIRUS STRUCTURE AND GENE EXPRESSION
G
S
3a
E
M
N
7
3b
mRNA
-+
-+
-+
-+
-+
-+
-+
-+
AAAUUU
AAAUUU
AAAUUU
AAAUUU
AAAUUU
AAAUUU
AAAUUU
AAAUUU
+
ORF 1a ORF 1b 3a 7 UTR3b
S M NE
AAACAP

TRANSCRIPTION MECHANISM
TEMPLATE SWITCH
A HIGH FREQUENCY RECOMBINATION
ENGINEERED AN INFECTIOUS cDNA CLONE
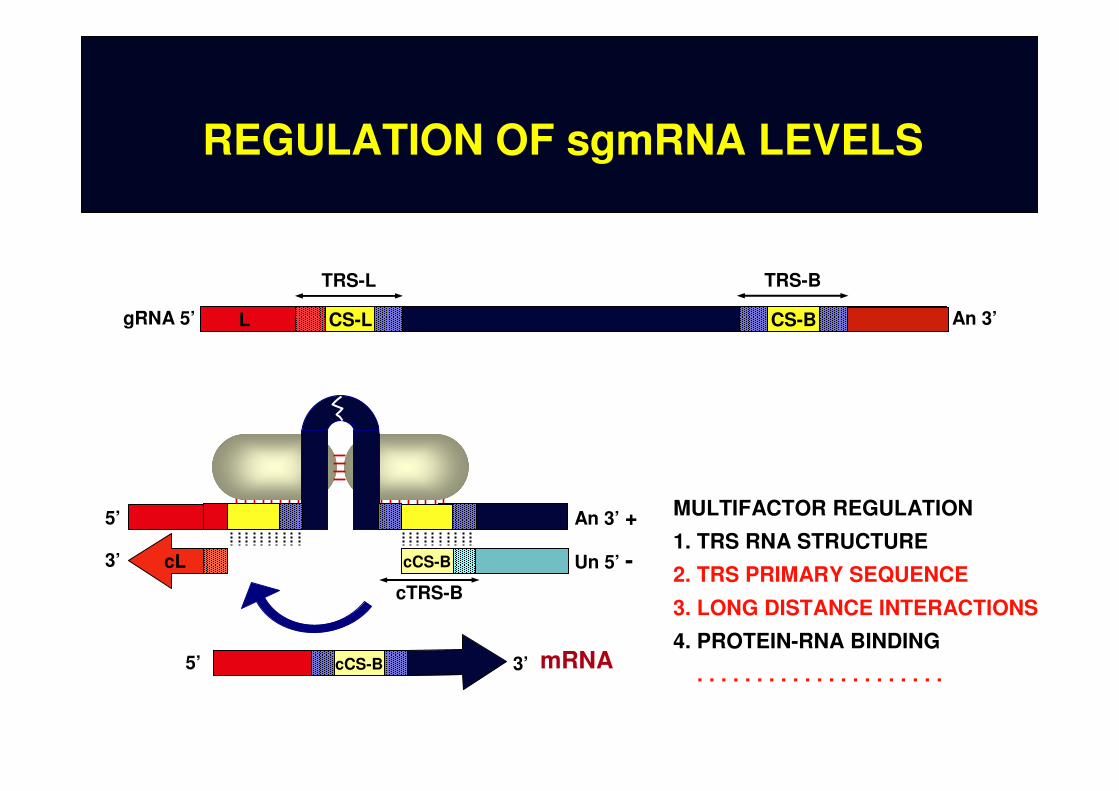
REGULATION OF sgmRNA LEVELS
CS-L CS-B An 3’
TRS-L TRS-B
gRNA 5’ L
5’ An 3’ +
Un 5’ -3’
cTRS-B
cCS-BcL
cCS-B5’ 3’ mRNA
MULTIFACTOR REGULATION
1. TRS RNA STRUCTURE
2. TRS PRIMARY SEQUENCE
3. LONG DISTANCE INTERACTIONS
4. PROTEIN-RNA BINDING
. . . . . . . . . . . . . . . . . . . . .
G A U
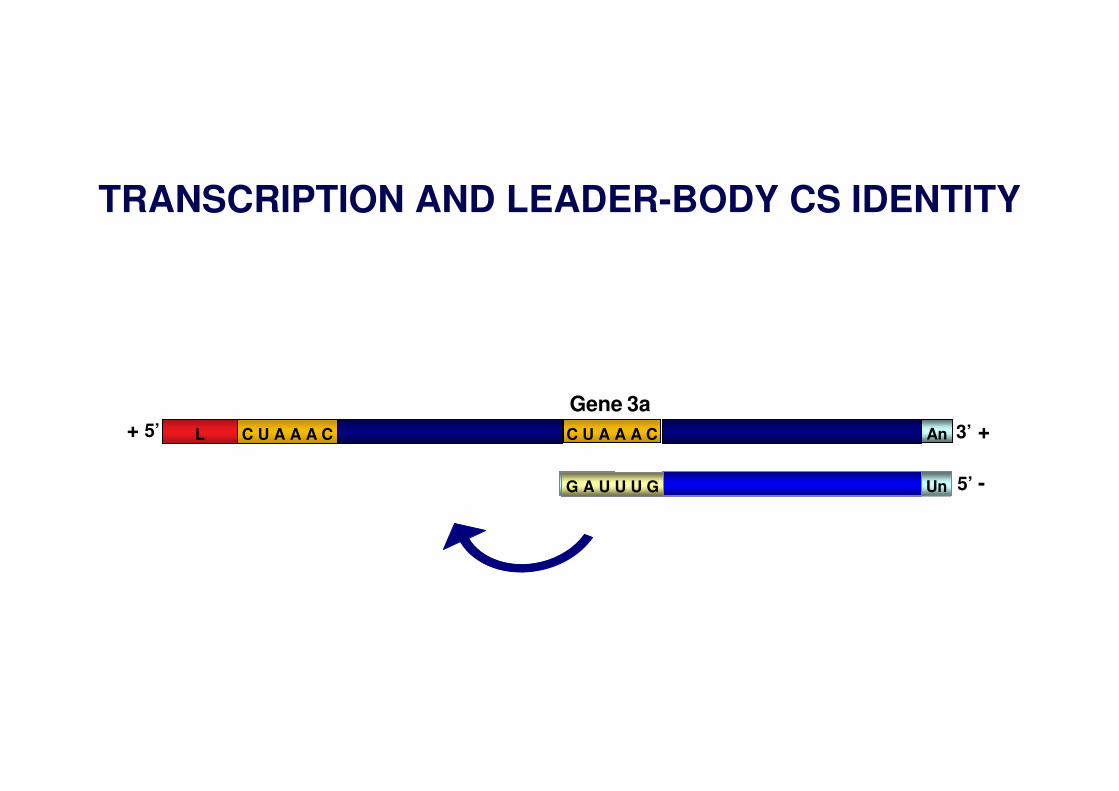
Gene 3a
TRANSCRIPTION AND LEADER-BODY CS IDENTITY
UnG A U U U G 5’ -
C U A A A CL AnC U A A A C+ 5’ 3’ +
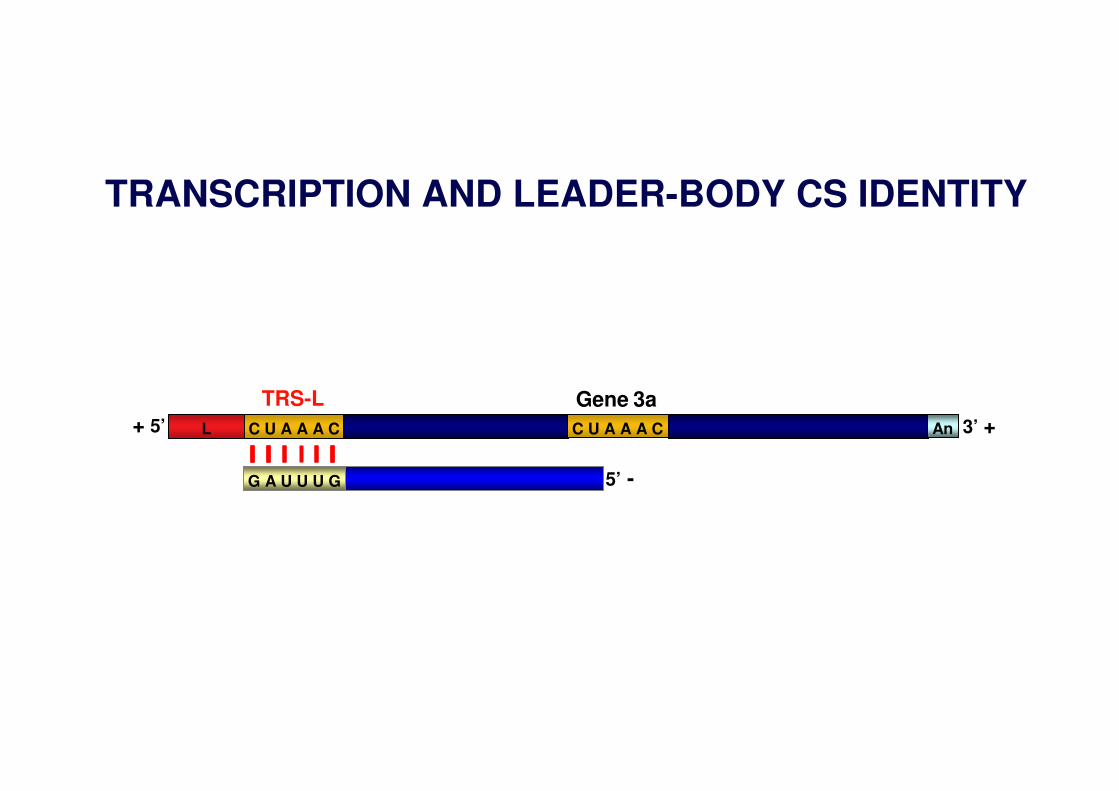
G A U U U G
Gene 3a
TRANSCRIPTION AND LEADER-BODY CS IDENTITY
C U A A A CL AnC U A A A C+ 5’ 3’ +TRS-L
5’ -

A3
G1
G2
C4
A5
U34
U32
U30
G31
C33
C10
A9
A8
C7
C6
A24
A25
U26
A27
U28
U29
G13
C12
C16
A15
A18
A19
A14 A20
U11
U17
A23
C21
G22
WT
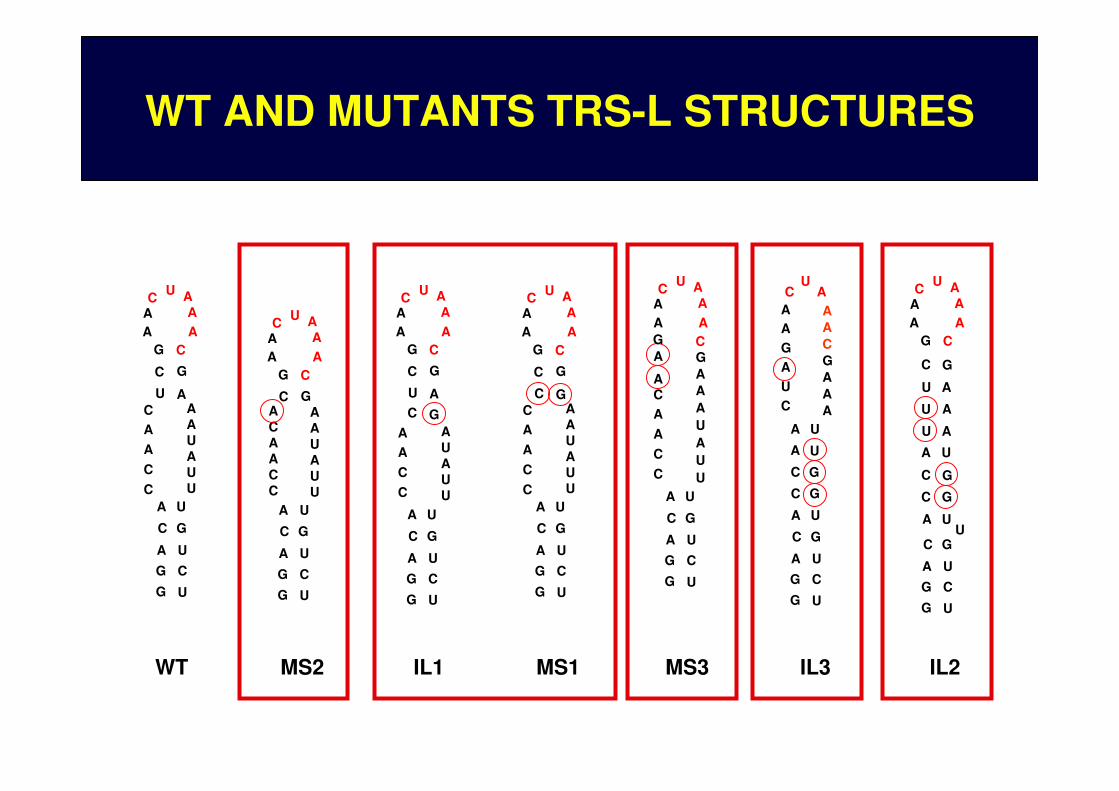
APICAL HEPTALOOP INCLUDING THE CS-L
MICRO STEM
INTERNAL LOOP
WILD TYPE TRS-L STRUCTURE PREDICTION
G
G
A
C
A
U
U
U
G
C
G
C
U A
C
G
A
A
C
C
AUAUU
C
G
CA
AA
A A
U
IL1
G
G
A
C
A
U
U
U
G
C
AAUAUU
ACAACC
G
C
C
G
CA
AA
A A
U
MS2WT
C
U A
G
G
G
A
C
A
U
U
U
G
C
C
A
A
C
C
AAUAUU
G C
CA
AA
A A
U
C
C G
G
G
G
A
C
A
U
U
U
G
C
C
A
A
C
C
AAUAUU
G C
CA
AA
A A
U
MS1
C
C G
U
G
A U
U A
C G
U A
U A
G
G
A
C
A
U
U
U
G
C
G C
CA
AA
A A
U
IL2
G
C
C
A
UA
G
U
C AU
G
G
A
C
A
U
U
U
G
C
AA
G
A
U
C
AACGAAA
IL3MS3
A
G
G
A
C
A
U
U
U
G
C
C
A
A
C
C
GAAAUAUU
G C
CA
AA
A
U
A
A
WT AND MUTANTS TRS-L STRUCTURES

CONCLUSION
THE EXTENT OF sg mRNA SYNTHESIS:
CORRELATES WITH TRS-L STABILITY
REQUIRES A SPECIFIC SECONDARY STRUCTURE
WITH THE AVAILABILITY OF TRS-L

TRANSCRIPTION MECHANISM
TEMPLATE SWITCH
A HIGH FREQUENCY RECOMBINATION
ENGINEERED AN INFECTIOUS cDNA CLONE

G A U
Gene 3a
TRANSCRIPTION AND LEADER-BODY CS IDENTITY
UnG A U U U G 5’ -
C U A A A CL AnC U A C A C+ 5’ 3’ +C
G
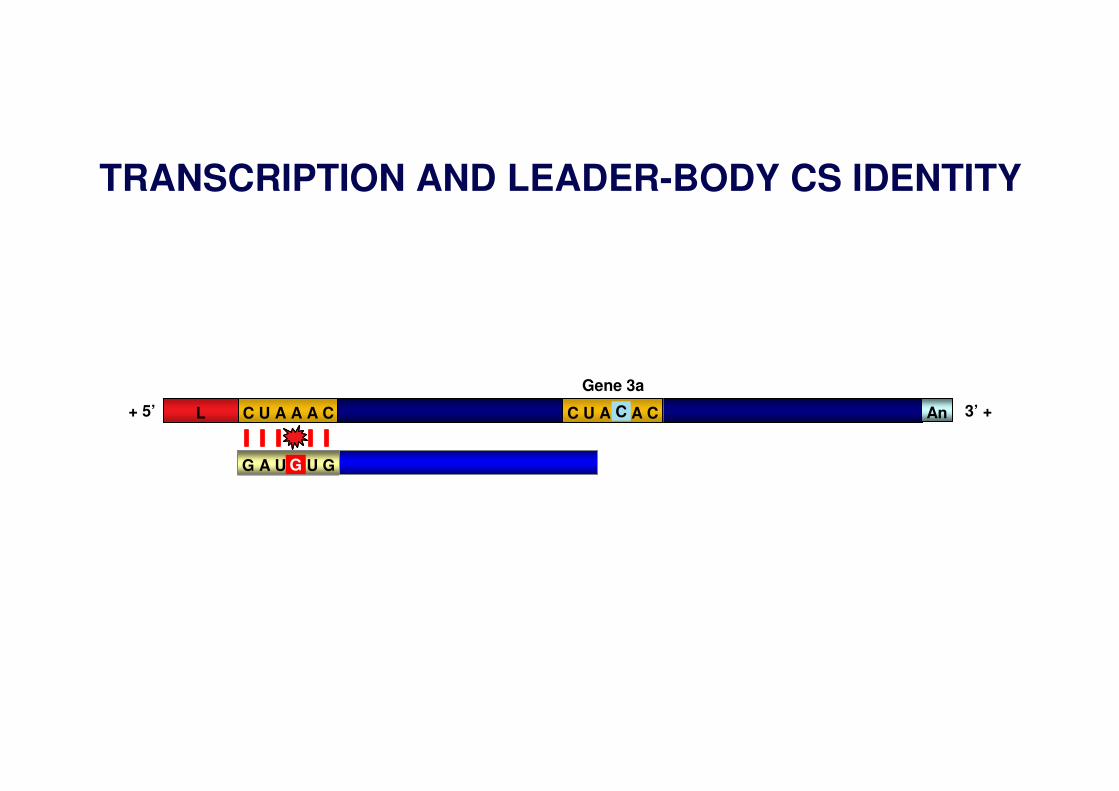
G A U U U GG
Gene 3a
TRANSCRIPTION AND LEADER-BODY CS IDENTITY
C U A A A CL AnC U A C A C+ 5’ 3’ +C
REQUIREMENT OF COMPLEMENTARITY BETWEEN CS-L AND cCS-B FOR sgmRNA SYNTHESIS
CC U A AA
GG
CC
CG
5’ 3’L-C1GL-U2GL-A3CL-A4CL-A5CL-C6G
TITER*
---
2x104
4x104
2x103
CC U A AA
GG
CC
CG
5’ 3’B-C1GB-U2GB-A3CB-A4CB-A5CB-C6G
TITER*
8x108
1x107
5x108
6x108
3x108
4x108
* pfu/ml
CS-B MUTANTS CS-L MUTANTS
5’ 3’D-C1G3’ 5’
5’ 3’D-C6G
3’ 5’
C
G
GU A AA
CA U UU
CG U A AA
GC A U UU
TITER*
---
2x105
4x105
2x103
COMPLEMENTARY MUTANTS
CC U A AA
U
UU
UU
5’ 3’L-C1U
L-A3UL-A4UL-A5UL-C6U
TITER*
2x103
5x108
3x103
5x104
2x104
1x108
WT
NON-WATSON-CRICK MUTANTS

MAIN CONCLUSION
BASE-PAIRING (ΔΔΔΔG) IS A DRIVING FORCE IN CoV TRANSCRIPTION
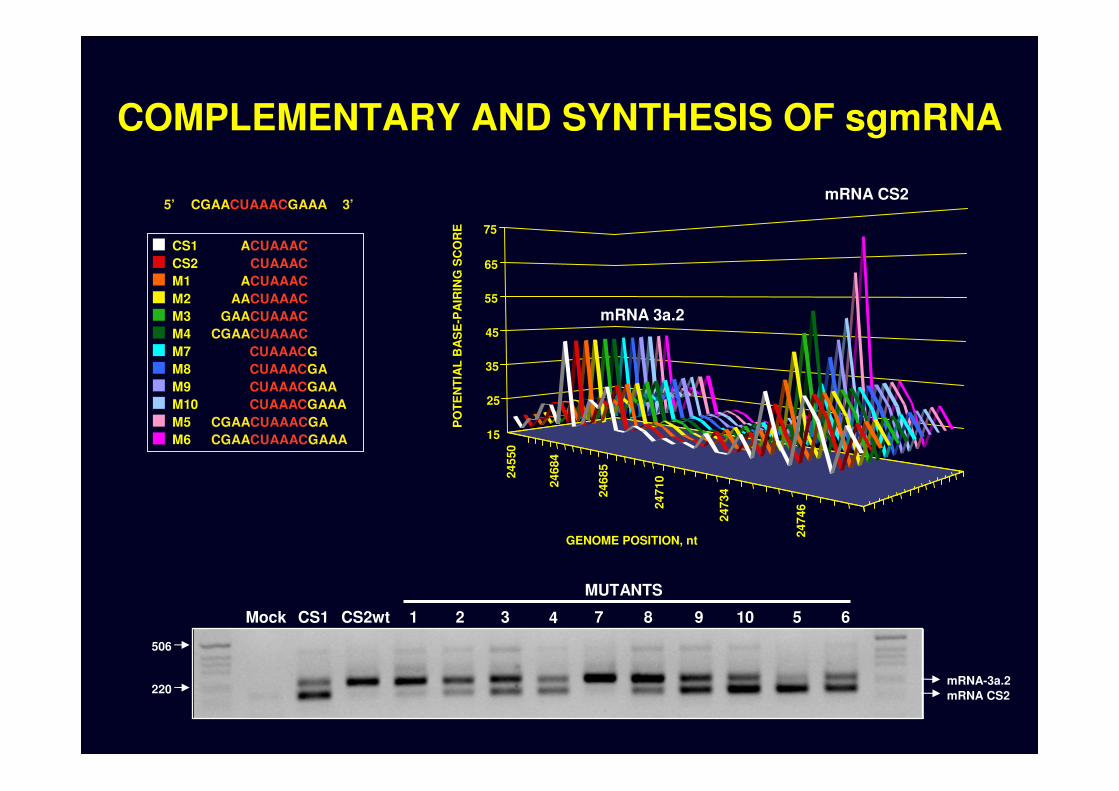
COMPLEMENTARY AND SYNTHESIS OF sgmRNA
2455
0
2468
4
2468
5
2471
0
2473
4
2474
6
15
25
35
45
55
65
75
PO
TE
NT
IAL
BA
SE
-PA
IRIN
G S
CO
RE
GENOME POSITION, nt
CS1CS2M1M2M3M4M7M8M9M10M5M6
CUAAACACUAAAC
ACUAAACAACUAAAC
GAACUAAACCGAACUAAAC
CGAACUAAACGAAACGAACUAAACGA
CUAAACGAAACUAAACGAACUAAACGACUAAACG
CGAACUAAACGAAA5’ 3’
506
220 mRNA CS2
Mock CS1 CS2wt 1 62 3 4 597 8 10
MUTANTS
mRNA-3a.2
mRNA 3a.2
mRNA CS2
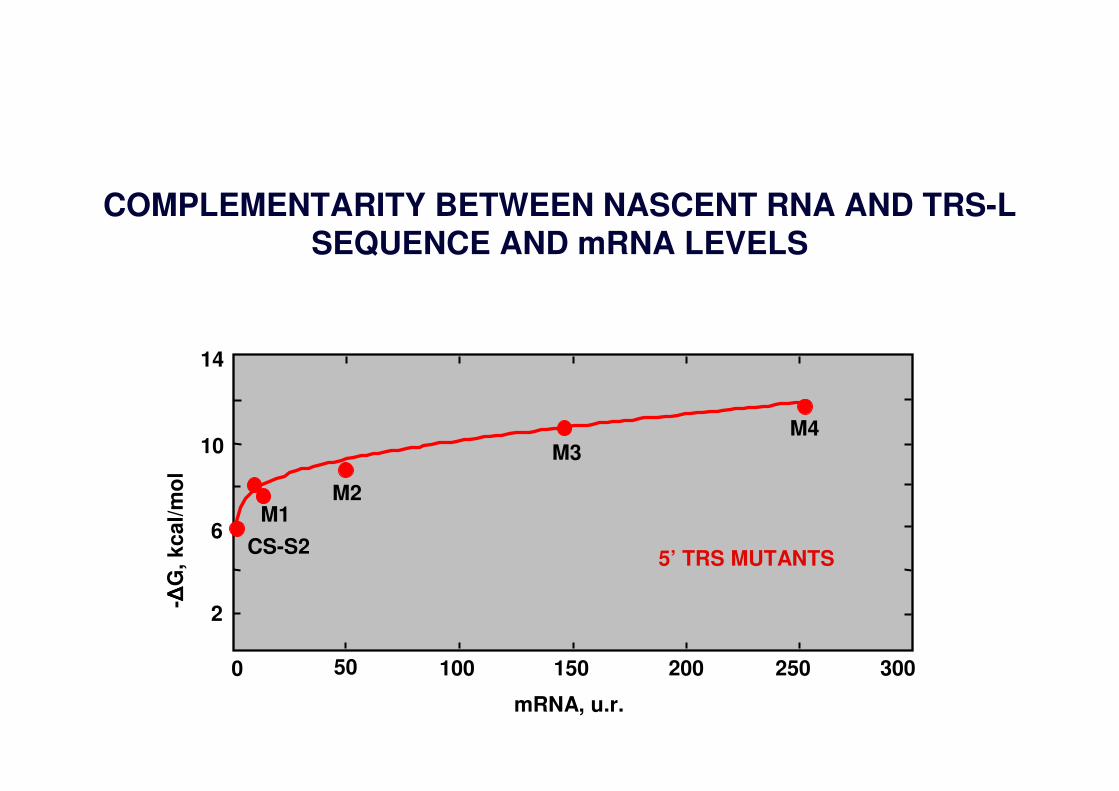
5’ TRS MUTANTS
- ΔΔ ΔΔG
, kca
l/mo
l
2
6
10
14
0 50 100 150 200 250 300
M4
M1
M3
M2
CS-S2
mRNA, u.r.
COMPLEMENTARITY BETWEEN NASCENT RNA AND TRS-LSEQUENCE AND mRNA LEVELS
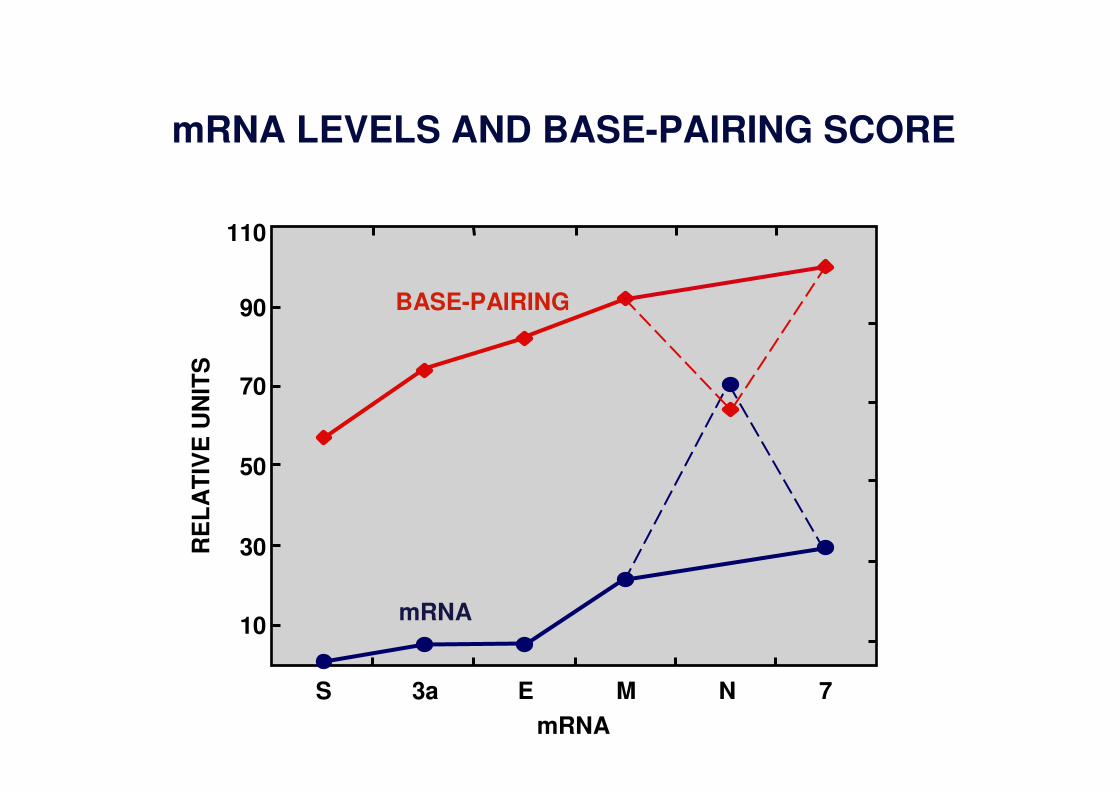
mRNA LEVELS AND BASE-PAIRING SCORE
10
30
50
70
90
110
S 3a E M N 7
mRNA
BASE-PAIRING
RE
LA
TIV
E U
NIT
S
mRNA
REGULATION OF sg mRNA-N LEVELS
CS-N An 3’
TRS-L TRS-N
gRNA 5’ L CS-L dE pE
Un 5’ -
3’
cTRS-N
cCS-N
cL
5’
An 3’ +
7nt460nt
ATTACATAT
TAATGTATA
N-GENE
COMPLEMENTARITY AND ENHANCER ACTIVITY
50
100
150
200
10 20150 5 25∆∆∆∆G, Kcal/mol
sgm
RN
A-N
, RE
LA
TIV
E U
NIT
S
REQUIREMENT OF pE AND dE FLANKING SEQUENCES
TRS-N
dE-113-158
dE-173-20
E2-TRS-N
pE-20
dE-45-158
pEdEMUTANT
sgmRNA-N, %
AD 0 20010050 150
OPTIMIZED TRM
ORF 1aORF 1bORF 1a ORF 1b AAA7 AAAN 7
dE pE CS
20 7 620173 699
250 ntOriginal TRM 642 nt ����
ENHANCEMENT MECHANISM
CS-NpE
dE
3’
5’
5’
AD
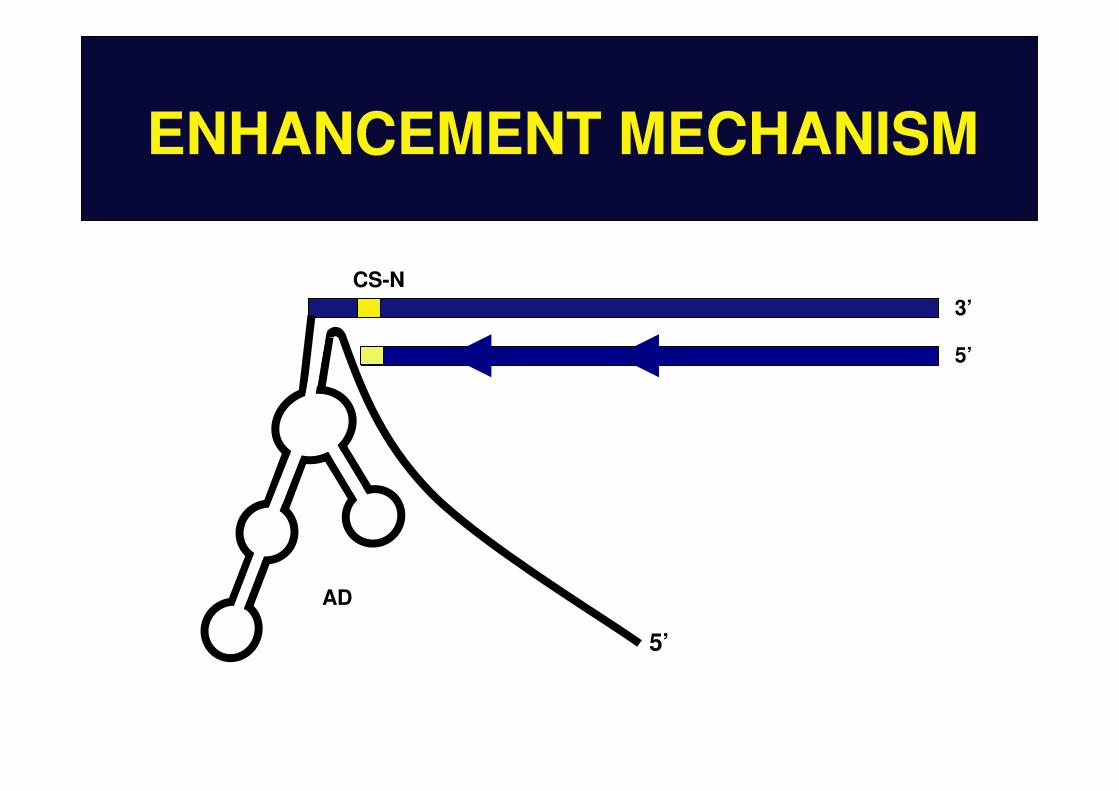
ENHANCEMENT MECHANISM
CS-N3’
5’
5’
AD
REQUIREMENT OF LONG DISTANCE INTERACTION
REP-1
sgmRNA-N/gRNA
0
AD-TRS-N
TRMopt-N
TRS-N
5 10 15
TRMpEdE CS-N
REP-1
TRS-N
AD-TRS-N
NAD
TRMopt-NTRMopt
MUTANT
AD
ENHANCEMENT MECHANISM
CS-NpE
dE
3’
5’
5’
AD
80% ENHANCEMENT
ACTIVE DOMAIN REGIONS REQUIREMENT
A
B
CΔΔΔΔA ΔΔΔΔB ΔΔΔΔC
0
5
10
15
20
25
+ -
sgm
RN
A-N
, RE
LA
TIV
E U
NIT
S
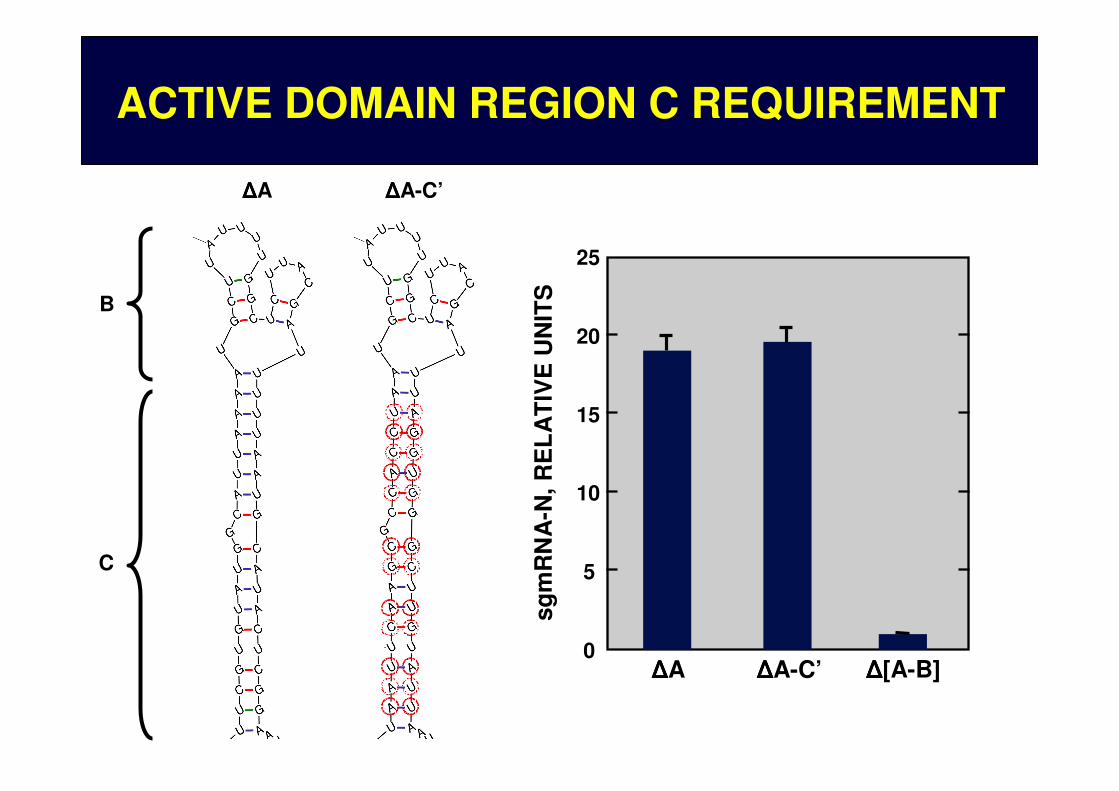
ACTIVE DOMAIN REGION C REQUIREMENT
ΔΔΔΔA-C’ΔΔΔΔA
B
C
0
5
10
15
20
25
ΔΔΔΔA-C’ΔΔΔΔA
sgm
RN
A-N
, RE
LA
TIV
E U
NIT
S
ΔΔΔΔ[A-B]
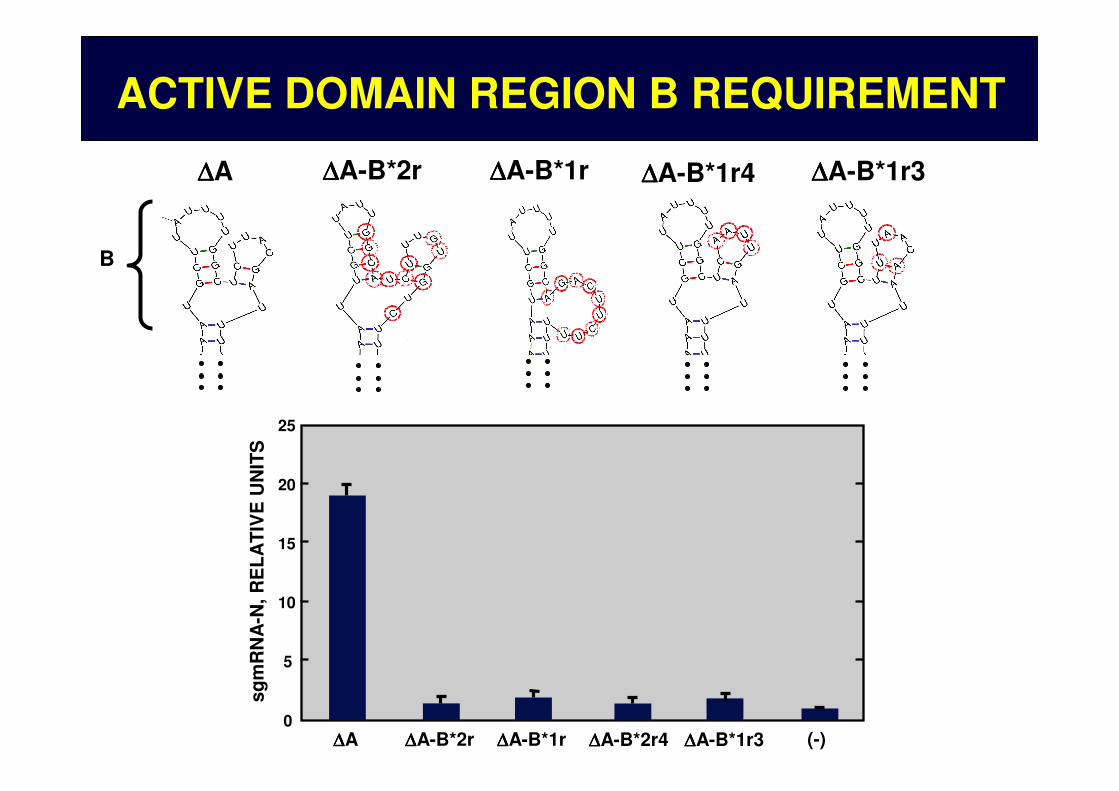
B
∆∆∆∆A-B*2r ∆∆∆∆A-B*1r ∆∆∆∆A-B*1r4 ∆∆∆∆A-B*1r3∆∆∆∆A
ACTIVE DOMAIN REGION B REQUIREMENT
0
5
10
15
20
25
∆∆∆∆A ∆∆∆∆A-B*2r ∆∆∆∆A-B*1r (-)
sgm
RN
A-N
, RE
LA
TIV
E U
NIT
S
∆∆∆∆A-B*2r4 ∆∆∆∆A-B*1r3
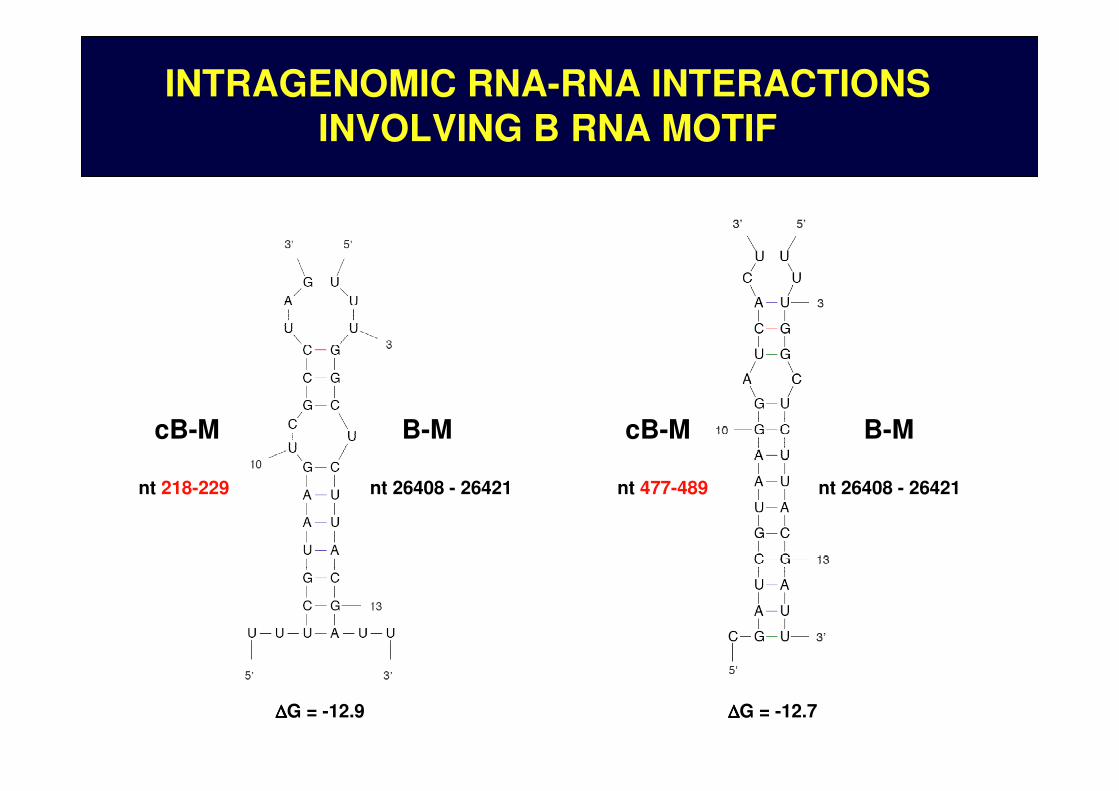
nt 218-229
INTRAGENOMIC RNA-RNA INTERACTIONS INVOLVING B RNA MOTIF
∆∆∆∆G = -12.7 ∆∆∆∆G = -12.9
cB-M
nt 477-489
cB-M
nt 26408 - 26421
B-M
nt 26408 - 26421
B-M

TGEV REPLICONS WITH MUTATED B AND cB RNA MOTIFS
wt/∆∆∆∆B
wt/B
5’ 3’
5’ 3’B dE pE N
pE N
5’ 3’
5’ 3’B*
*
5’ 3’B**
cB-477*/B*-14 5’ 3’B**
pE N
dE pE N
dE pE N
dE pE N
cB-218*/∆∆∆∆B
cB-218*/B
5’ 3’
5’ 3’B*
cB-218*/B* 5’ 3’B**
pE N
dE pE N
dE pE N
*
cB-218
cB-477
cB-477*/∆∆∆∆B
cB-477*/B
cB-477*/B*
dE
dE
dE
5’ 3’
218-229 477-486 26,212-26,221
TRANSCRIPTION AND COMPLEMENTARY LEVELS
0
5
10
15
20
25
30
35
40
45
wt/ ∆∆ ∆∆
B
wt/B
cB-2
18*/ ∆∆ ∆∆
B
cB-2
18*/B
cB-2
18*/B
*
sgm
RN
A N
, RE
LA
TIV
E U
NIT
S
cB-4
77*/B
*-14
cB-4
77*/ ∆∆ ∆∆
B
cB47
7*/B
cB-4
77*/B
*
5’ 3’B dE pE N
218 477 26,212

EVOLUTION OF VIRUSES WITH MUTATED B AND cB RNA MOTIFS
RELEVANCE OF THE INTERACTION BETWEEN B AND cB RNA MOTIFS ON sgmRNA AND VIRAL TITERS
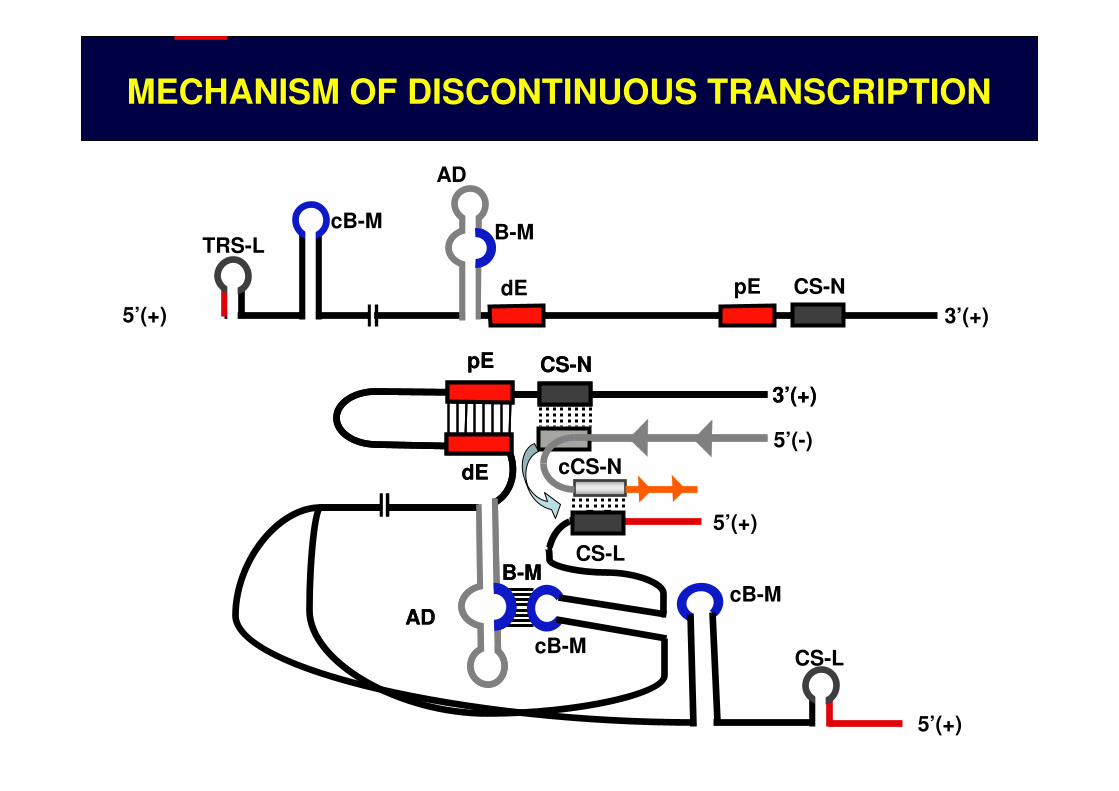
MECHANISM OF DISCONTINUOUS TRANSCRIPTION
5’(+) 3’(+)CS-NpEdE
AD
B-McB-MTRS-L
pE
dE
AD
5’(+)
CS-N3’(+)
CS-LB-M
cB-M
5’(-)cCS-NdE
AD
5’(+)
CS-N3’(+)
CS-L
B-McB-M
pE

TRS-L STABILITY AND SECONDARY STRUCTURE
COMPLEMENTARITY BETWEEN NASCENT MINUS RNAs AND TRS-L
CoV TRANSCRIPTION IS REGULATEDAT THREE LEVELS
LONG DISTANCE RNA-RNA INTERACTIONS
ORF 1a ORF 1b
N
3a 3b
N A
N 7MS Ewt-TGEV
TRS-3a
dE pE CS
TRMopt-3a
CS
AD
dE pE
sgmRNA-3a/gRNA
TRMopt-19-3a
0 2 4 6
TRMopt*-3a
TRM ACTIVITY WITHIN INFECTIOUS VIRUSES
GEOGRAPHICAL AREA ORIGIN OF SARS
SEVERE ACUTE RESPIRATORY SYNDROME
Neumonia High fever
Lymphopenia Death

SPREADING OF SARS
…JULY 5th, 03 END EPIDEMY BY WHO
Beijin Apr F 26y, BSL-4 Beijin Apr M 44y, BSL-4
NOV 02 JAN 03 FEB 03 DEC03 to JAN04FOSHAN GUANGZHOU HONG KONG
MOST RECENT COMMON ANCESTOR
FIRSTMAJOROUTBREAKHOSPITAL
Dr. ANephro-logist
HOTEL M
VIETNAMCANADASINGAPOREUSATWO ADDDITIONALSSE IN HK (>100 CONTACTS)
FOUR NON-LABORATORYISOLATED CASES
FOUR LABORATORY CASES

RACOON
CIVET CATFERRETS
GENUS αααα
GENUS ϒϒϒϒGENUS ββββ
CORONAVIRUS EVOLUTION
ENGINEERING OF A SARS-CoV INFECTIOUS cDNA CLONE AS A BAC
EM
NREP 1a REP 1b SL
An3a
3b
6
7a
7b
8a8b 9b
CMV
T by C10338
T by A11163
pBAC-SARS-CoV-URB*

BASIS OF SARS-CoV VIRULENCE
Structural: E
Accessory: 6, 7a, 7b, 8a, 8b, 9b
Combinations: E, 6, 7a, 7b, 8a, 8b, 9b
GENEDELETED
8a
L REP 1a REP 1b S 3a3b
EM
67a
7b8b
N9b
An

BSL-3 CNB. CSIC. MADRID
GROWTH KINETICS OF SARS-CoV DELETION MUTANTS
CaCo-2
GENES: E, 6, 7a, 7b, 8a, 8b, 9b ARE NON ESSENTIAL
TIME POST-INFECTION, HOUR
VERO E6
0 24 48 72
101
103
105
107
VIR
US
TIT
ER
, p
fu/m
l
ΔΔΔΔE,6-9b
0 24 48 72
wt
ΔΔΔΔ6-9b
ΔΔΔΔE
MORPHOGENESIS OF SARS-CoV IN ERGIC
SARS-CoVSARS-CoV-∆∆∆∆ [6-9b]
SARS-CoV-∆∆∆∆ESARS-CoV-∆∆∆∆ [E,6-9b]

ACTIVITY WHEEL USED TO QUANTIFY CLINICAL ILLNESS
DAY
INFECTION
0
200
400
600
800
1000
1200
-7 -6 -5 -4 -3 -2 -1 0 1 2 3 4
RE
VO
LU
TIO
NS
/ hr
MOCK
∆∆∆∆E
wt
SARS-CoV-∆∆∆∆E-INFECTED HAMSTER WHEEL ACTIVITY
2 4 6 8
25
50
75
100
SU
RV
IVA
L, %
TIME POSTINOCULATION, days
0
ΔΔΔΔE
ΔΔΔΔE,6-9b
ΔΔΔΔ6-9b
ATTENUATION OF SARS-CoV DELETION MUTANTSIN TRANSGENIC MICE EXPRESSING hACE-2
2 4 6 870
80
90
110
ST
AR
TT
ING
WE
IGH
T, %
100
wt
ΔΔΔΔE
ΔΔΔΔE,6-9b
ΔΔΔΔ6-9b
TIME POSTINOCULATION, days
12000 pfu intranasally
CLINICAL DISEASE LETHALITY

SUMMARY
SARS-CoV-∆∆∆∆E IS ATTENUATED
E GENE CONDITIONS TISSUE SPECIFIC TROPISM
GENE E IS A VIRULENCE FACTOR
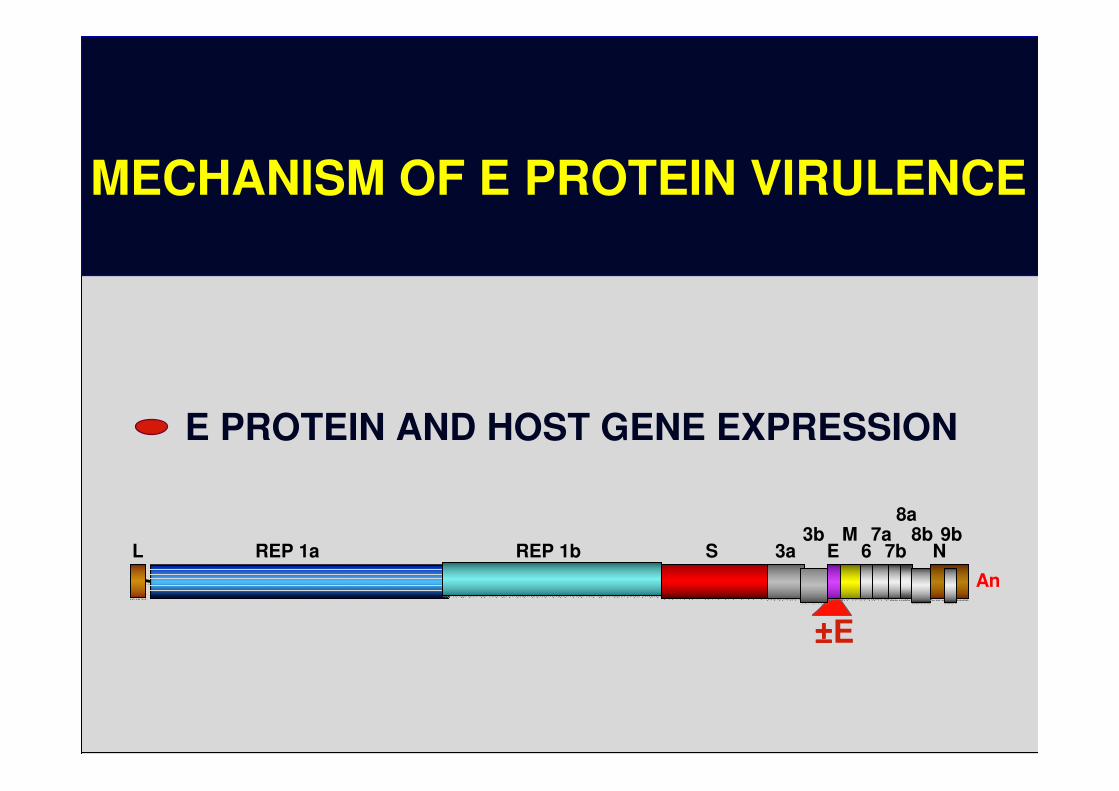
MECHANISM OF E PROTEIN VIRULENCE
E PROTEIN AND HOST GENE EXPRESSION
±±±±E
8a
L REP 1a REP 1b S 3a3b
EM
67a
7b8b
N9b
An
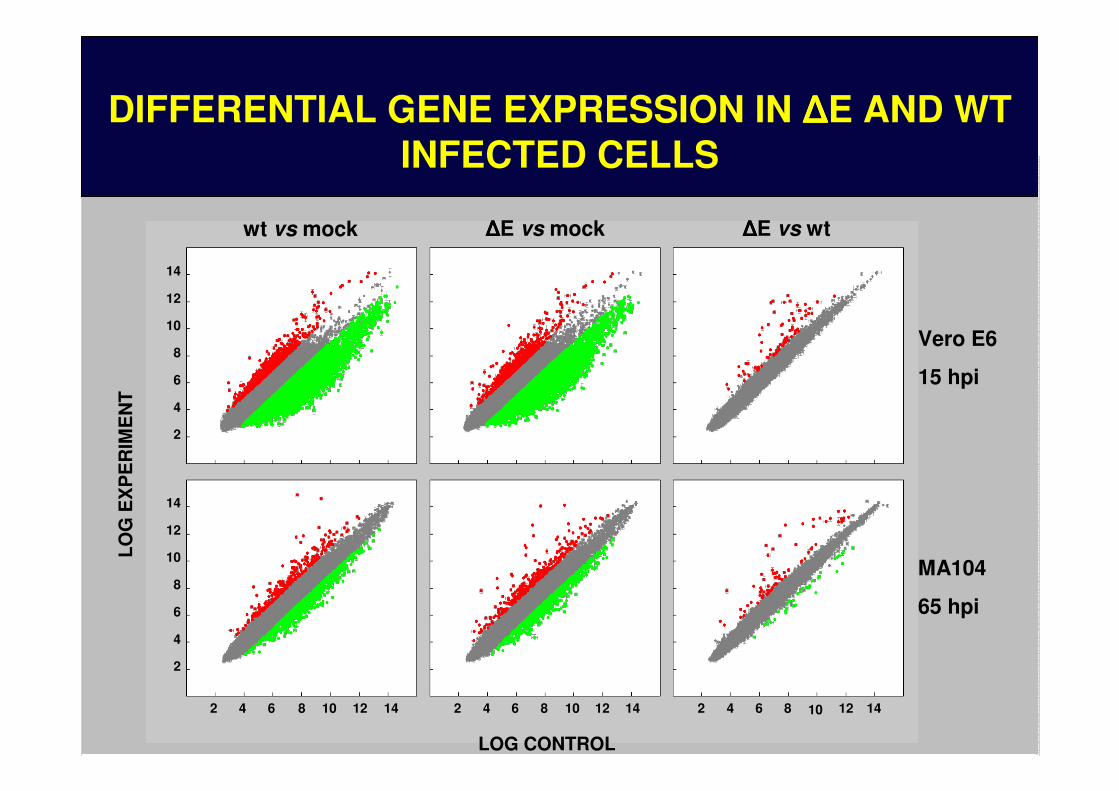
10 12 144 6 8210 12 144 6 82 10 12 144 6 82
10
12
14
4
6
8
2
10
12
14
4
6
8
2
MA104
65 hpi
wt vs mock ΔΔΔΔE vs mock ΔΔΔΔE vs wt
LOG CONTROL
LO
G E
XP
ER
IME
NT
Vero E6
15 hpi
DIFFERENTIAL GENE EXPRESSION IN ∆∆∆∆E AND WT INFECTED CELLSDIFFERENTIAL GENE EXPRESSION IN ΔΔΔΔE AND WT
INFECTED CELLS
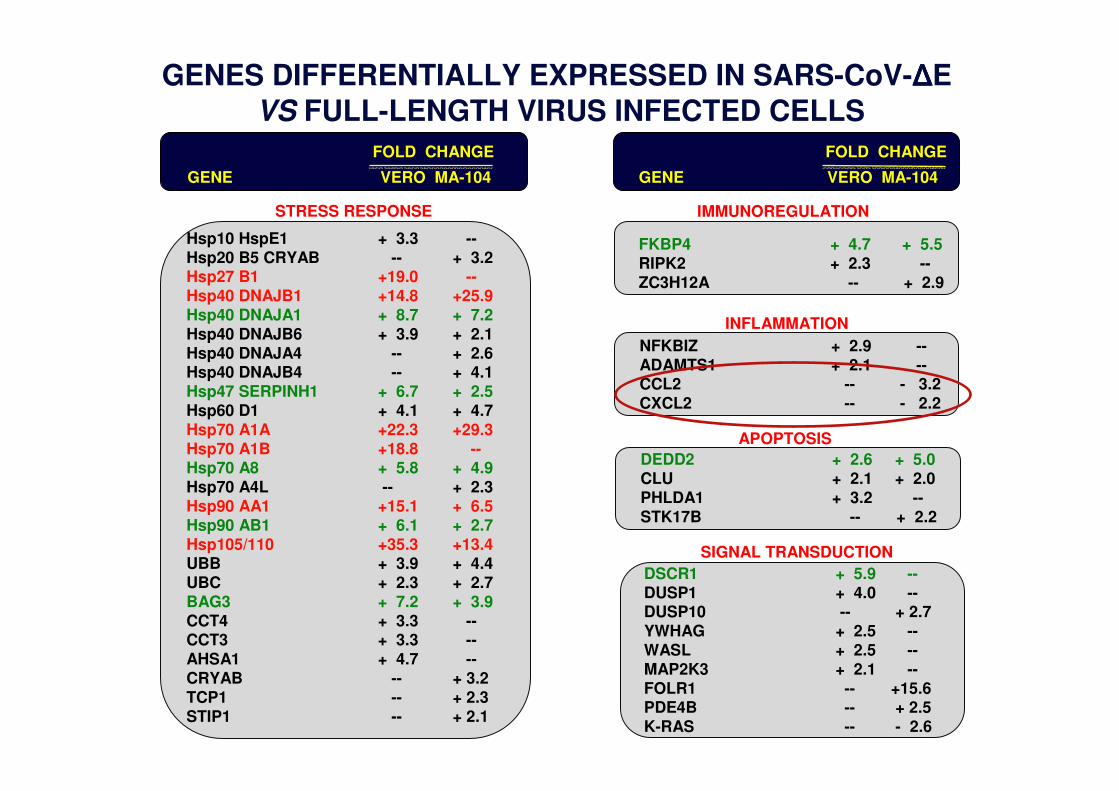
GENES DIFFERENTIALLY EXPRESSED IN SARS-CoV-ΔΔΔΔE VS FULL-LENGTH VIRUS INFECTED CELLS
INFLAMMATION
APOPTOSIS
SIGNAL TRANSDUCTION
IMMUNOREGULATION
DSCR1 + 5.9 --DUSP1 + 4.0 --DUSP10 -- + 2.7YWHAG + 2.5 --WASL + 2.5 --MAP2K3 + 2.1 --FOLR1 -- +15.6PDE4B -- + 2.5K-RAS -- - 2.6
DEDD2 + 2.6 + 5.0CLU + 2.1 + 2.0PHLDA1 + 3.2 --STK17B -- + 2.2
NFKBIZ + 2.9 --ADAMTS1 + 2.1 --CCL2 -- - 3.2CXCL2 -- - 2.2
GENE VERO MA-104
FOLD CHANGE
FKBP4 + 4.7 + 5.5RIPK2 + 2.3 --ZC3H12A -- + 2.9
GENE VERO MA-104
FOLD CHANGE
STRESS RESPONSE
Hsp10 HspE1 + 3.3 --Hsp20 B5 CRYAB -- + 3.2Hsp27 B1 +19.0 --Hsp40 DNAJB1 +14.8 +25.9Hsp40 DNAJA1 + 8.7 + 7.2Hsp40 DNAJB6 + 3.9 + 2.1Hsp40 DNAJA4 -- + 2.6Hsp40 DNAJB4 -- + 4.1Hsp47 SERPINH1 + 6.7 + 2.5Hsp60 D1 + 4.1 + 4.7Hsp70 A1A +22.3 +29.3Hsp70 A1B +18.8 --Hsp70 A8 + 5.8 + 4.9Hsp70 A4L -- + 2.3Hsp90 AA1 +15.1 + 6.5Hsp90 AB1 + 6.1 + 2.7Hsp105/110 +35.3 +13.4UBB + 3.9 + 4.4UBC + 2.3 + 2.7BAG3 + 7.2 + 3.9CCT4 + 3.3 --CCT3 + 3.3 --AHSA1 + 4.7 --CRYAB -- + 3.2TCP1 -- + 2.3STIP1 -- + 2.1
UNFOLDED PROTEIN RESPONSE
RE
PERK
GR
P78
eIF2 TRANSLATIONALBLOCK
ATF6CLEAVED
XBP1mRNA
sXBP1mRNA
sXBP1
CYTOPLASM NUCLEUS
ACTIVATION IN ΔΔΔΔE VS WT-
INFECTED CELLS
GENEEXPRESSION
TRANSLATION ATTENUATIONCHOP
CORRECT PROTEINFOLDINGCHOP
PROTEIN DEGRADATIONp58IPK
-
+
GRP78
GRP78
GR
P78
GR
P78
GR
P78
GR
P78
GR
P78
PERK
eIF2
ATF6
PATF4
IRE1
IRE1
-
EFFECT OF E PROTEIN ON THE STRESS PRODUCED BY SARS-CoV-ΔΔΔΔE INFECTION
FO
LD
CH
AN
GE
, rel
ativ
e u
nit
s
2
4
6
8
10
12
E+
E-
SERPIN H1HSP90 AA1 HSP H1 HSP E1HSP A1A 18S
E PROTEIN PROVIDED IN TRANS
SARS-CoV E PROTEIN AND STRESS PRODUCEDBY RSV INFECTION
TIME POST-INFECTION, hours
FO
LD
CH
AN
GE
, rel
ativ
e u
nit
s
1
2
3
4
5
6
7
50
100
150
200
250
300
350
2 24 48 2 24 48
HSP90 AA1 UBB
E+
E-
2 24 48 2 24 48
HSP H1 SERPIN H1
2 24 48
HSP E1 18S
2 24 48
E PROTEIN PROVIDED IN TRANS

CONCLUSION
E PROTEIN REDUCED CELLULAR STRESSINDUCED BY:
SARS-CoV INFECTION
THAPSIGARGIN - Ca++
TUNICAMYCIN - GLYCOSYLATION
RSV INFECTION
SARS-CoV-ΔΔΔΔE INFECTION AND INFLAMMATION
INFLAMMATORYRESPONSE
-E
+E
STRESS RESPONSE

MOUSE ADAPTED SARS-CoV
High titers in lungs
Viremia, extrapulmonary spread
Neutrophilia
Pathological changes in lungs
Death
K. SUBBARAO AND R. BARIC LABS
SARS-CoV-MA15
CONSTRUCTION OF A MOUSE ADAPTED SARS-CoV
nsp510384
nsp510793
nsp912814
nsp1316117
S (RBD)22797
M26428
ENGINEERED MUTANTS wt
ΔΔΔΔEΔΔΔΔ[E,6-9b]
ΔΔΔΔ[6-9b]
8a
L REP 1a REP 1b S 3a3b
EM
67a
7b8b
N9b
An

CONCLUSION
SARS-CoV-MA15-ΔΔΔΔE IS ATTENUATED AND FULLYPROTECTS BOTH YOUNG AND OLD MICE
THEREFORE IT IS A PROMISING VACCINECANDIDATE, BETTER THAN SARS-CoV-ΔΔΔΔE

REPLICATIONFERNANDO ALMAZANAITOR NOGALESSILVIA MARQUEZ
TRANSCRIPTION AND ASSEMBLYISABEL SOLAPEDRO A. MATEOSSONIA ZUÑIGAMARTINA BECARES
VIRUS-HOST INTERACTION: SARSMARTA L. DeDIEGOJOSE LUIS NIETOJOSE MANUEL JIMENEZJOSE ANGEL REGLARAUL FERNANDEZ
PROTEOMICSSILVIA JUAREZ. CNB-CSICALEXANDER AKOULICHEV. OXFORD
CNB. CSIC. MADRID COLLABORATORS
LUIS ENJUANES
ANIMAL MODELSKANTA SUBBARAO. NIHSTANLEY PERLMAN. UNIV. IOWA
ELECTRON MICROSCOPYMARIA TERESA REJAS. CBM, CSICCRISTINA PATIÑO. CNB. CSIC
CARLOS M. SANCHEZSARHAY ROSMARGARITA GONZALEZ
GENOMICSIRENE LOPEZ-VIDRIEROMARTA GODOYJUAN CARLOS OLIVEROS
Recommended

















