Current Biology Vol 15 No 3R90
Aaron Johnson2 and Mike O’Donnell1,2
DNA ligation is the last step in amultitude of important DNAmetabolic reactions [1]. DNA nicksintroduced during replication,recombination and repair need tobe sealed — if left unchecked,they would lead to cell death. Soeven before its discovery, a DNAligase activity was proposed toexist. The discovery of ligase andthe elegant biochemical studies ofthe ligase reaction by the Lehmangroup and others revealed theamazing series of chemical stepsneeded to seal a nick in DNA(reviewed in [2]). The beauty ofligase and its clever enzymaticstrategy is highlighted by therecently reported crystal structureof human DNA ligase I (Lig1)bound to a nicked DNAsubstrate [3].
Sealing a nick in DNA mightappear a simple task, but thereaction is exceedingly complex,requiring three distinct catalyticsteps and two covalentintermediates (Figure 1A). The firststep, enzyme adenylation, isaccomplished using either NAD+(in eubacteria) or ATP, but bothresult in an AMP-linkage to theenzyme. In the second step, theAMP moiety is transferred to the 5′′phosphate at the site of a nick.This activates the 5′′ terminus forattack by the 3′′ OH in the third
and final phosphoryl transfer step,thereby sealing the nick.
Earlier ligase structures [4–7]were tremendously informativeabout the first step of the reactionand the AMP–ligase intermediate,and even suggested important
features of the subsequent steps.The structure of Lig1 in a complexwith nicked DNA [3] has now beendetermined, providing a three-dimensional snapshot of themoment before DNA ligation andgiving conclusive insight intoligase fidelity and the final steps ofthe reaction. To form this stablereaction intermediate, Pascal et al.[3] used a synthetic nicked duplexterminated with a 3′′dideoxynucleotide, therebyremoving the critical 3′′ OH groupnecessary to form thephosphodiester bond. When Lig1is reacted with this substrate in
15. de Belle, J.S., and Heisenberg, M. (1994).Associative odor learning in Drosophilaabolished by chemical ablation ofmushroom bodies. Science 263,692–695.
16. Dubnau, J., Grady, L., Kitamoto, T., andTully, T. (2001). Disruption ofneurotransmission in Drosophilamushroom body blocks retrieval but notacquisition of memory. Nature 411,476–480.
17. McGuire, S.E., Le, P.T., and Davis, R.L.(2001). The role of Drosophila mushroombody signaling in olfactory memory.Science 293, 1330–1333.
18. Waddell, S., Armstrong, J.D., Kitamoto,T., Kaiser, K., and Quinn, W.G. (2000).The amnesiac gene product is expressedin two neurons in the Drosophila brainthat are critical for memory. Cell 103,805–813.
19. Keene, A.C., Stratmann, M., Keller, A.,Perrat, P.N., Vosshall, L.B., and Waddell,S. (2004). Diverse odor-conditionedmemories require uniquely timed dorsalpaired medial neuron output. Neuron 44,521–533.
20. Schwaerzel, M., Monastirioti, M., Scholz,H., Friggi-Grelin, F., Birman, S., andHeisenberg, M. (2003). Dopamine and
octopamine differentiate betweenaversive and appetitive olfactorymemories in Drosophila. J. Neurosci. 23,10495–10502.
Department of Neurobiology, Universityof Massachusetts Medical School, 364Plantation Street, Worcester,Massachusetts 01605, USA. E-mail: [email protected]
DOI: 10.1016/j.cub.2005.01.024
DNA Ligase: Getting a Grip to Sealthe Deal
The crystal structure of human DNA ligase I catches the enzyme justbefore the last step of ligation and shows that the protein wrapscompletely around nicked DNA. The elegant structure explains howligase attains fidelity for the sealing operation.
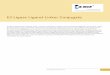
Figure 1. Comparison of prokaryotic and eukaryotic DNA ligase.
(A) Mechanism of DNA ligase. Note: bacterial cellular ligases use NAD+ and otherligases use ATP for self-adenylation. (B) DNA ligases have functionally similardomains, but they are scrambled in their linear sequence (top). Nevertheless, humanLig1 (left, in complex with DNA) and Thermus filiformis (Tfi) ligase (right) both adopt aring-shaped structure (bottom). The gray interaction domain (Int.) of Lig1 wasremoved for crystal structure analysis. The helix-hairpin-helix (HhH) domain ofT. filiformis ligase is analogous to the Lig1 DBD, but must undergo a large rotationwith the OBD relative to the AdD for T. filiformis ligase to similarly accommodate DNA.
ATPor
NAD+
PPior
NMN
AMPAMP
AMP
5′ 3′HO P
HO PLigase
Human DNA ligase I T. filiformis DNA ligase
N C N CInt. DBD AdD AdD ZnIa HhH BRCTOBD OBD
Interface
Ligation complex Ligase–AMP complex
Current Biology
A
B

the presence of ATP, the enzyme-and DNA-adenylation steps occur,but the ligase is frozen on DNA atthe last step.
The ‘frozen’ Lig1–DNA complexis a striking structure of a proteinring with DNA threaded throughthe middle. All three domains ofLig1 make intimate contacts withthe nicked DNA duplex. The mostextensive interactions are madeby the flat surface of the DNAbinding domain (DBD), mediatedthrough twelve αα helices. Theseprotein–DNA contacts occurmainly in the minor groove.Interestingly, the DBD binds DNAevenly on both sides of the nick,and this arrangement coincideswith an approximate two-fold axisof structural symmetry within theDBD (Figure 2A). This architectureprovides a ‘platform’ for bindingthe nicked DNA and for catalysisof the last step to seal the nick.The DBD platform, the adenylationdomain (AdD) and the OB-folddomain (OBD), togethercompletely encircle the DNA.
Many viral and phage ligasesare composed of only twodomains, AdD and OBD, and arenot expected to encircle DNAcompletely [4,6]. A secondcellular ligase structure, fromThermus filiformis, also shows athird main domain, like the Lig1structure (Figure 1B). Pascal et al.[3] note that the helix-loop-helixDNA binding domain of the T.filiformis ligase is structurallysimilar to Lig1 DBD, with a two-fold symmetry axis. Theprokaryotic ligase is not in aconformation that can readilyaccept DNA; the structuralsimilarity, however, suggests apermutation of sequencemodules in which functionallyhomologous domains of thesetwo cellular ligases are stitchedtogether in different ways,providing support for the earlierproposal that bacterial ligaseencircles DNA [7].
The OBD appears to aidsubstrate selection. Downstreamof the nick, the OBD interactswith B-form DNA, which mayexplain why Lig1 does not sealOkazaki fragments before theRNA primer is removed. But, in aseeming paradox, upstream DNAbound to the OBD is A-form,
making an induced-fit (Figure 2A).Additionally, the OBD distorts theDNA, leading to an underwoundhelix and a 5 Å shift in the helicalaxis between upstream anddownstream segments. Thesedistortions bring the nick close tothe active site and underliemultiple aspects of ligase fidelity.Ligases do not bind tightly toDNA containing a mismatchupstream of the nick [8], nor toDNA with a gap of even onenucleotide [9].
The Lig1–DNA structure impliesthat a dramatic conformationalchange between the OBD andAdD is necessary to proceed fromenzyme–AMP to DNA-boundligase. Large and dynamic rigidbody motions between these twodomains have been proposed formany ligases, based on thedistant locations of the enzyme-and DNA-adenylation catalyticresidues in the OBD [4–7]. Thesetwo motifs need access to theactive site at different steps in themechanism, necessitating rotation
of the OBD relative to the activesite in the AdD. Thisreorganization of catalyticresidues involves a gross changein structure that may explain howthe ligase ring opens.
The ring-shape of Lig1 bringsto mind the oligomericprocessivity factor rings, such asthe Escherichia coli ββ andeukaryotic PCNA proteins, whichact as sliding clamps on DNA.There is a clear distinction,however, in that Lig1 grips nickedDNA tightly through extensiveinteractions, whereas theprocessivity clamps have a largecentral channel and slide freelyalong the duplex.
Besides both being ring-shaped, Lig1 and PCNA are alsobinding partners [10,11], as are E.coli ligase and ββ proteins [12].Lig1 has an essential conservedPCNA-interacting motif shared bymany clamp-binding proteins[13], including FEN-1 nucleasewhich removes the RNA primer inOkazaki fragment maturation
Dispatch R91
Figure 2. Models of the Lig1–DNA complex and its proposed interaction with PCNA.
(A) The three domains of the Lig1 ring interact with the minor groove of DNA (left). TheOBD distorts DNA at the nick (right), offsetting the helical axis by 5 Å and convertingthe upstream DNA to A-form while recognizing B-form DNA downstream. (B) A modelof the Lig1–DNA (red and gray) and PCNA–p21 [17] (blue and green) structures togetheron DNA. The amino-terminal domain of Lig1, not present in the crystal structure, inter-acts with the PCNA homotrimer. The interaction is presumed to occur via the PIPsequence in similar fashion to the corresponding peptide of p21WAF1/CIP1 (green). Lig1and PCNA have been aligned to position the amino terminus of Lig1 in close proximityto one of the p21 peptides. (Reproduced with permission [3].)
A
B
AMPP
P AMP
HO
HO
OBD
DBD
AdD A-formDNA
B-formDNA
3′
3′5′
5′
PAMP OH
3′
5′
5Å
Upstream Downstream
PCNA-p21
Lig1–DNA
Position ofPCNA-interaction
domain
NH2
90o
Current Biology

[14]. This motif resides in the first231 amino acids of Lig1, whichwere removed for crystalstructure analysis, but Figure 2Billustrates how these two proteinswould be juxtaposed while onDNA. From studies with E. coli[15], the PCNA clamp ispresumed to be left behind on anOkazaki fragment after the DNApolymerase has finishedextending it to a nick. Theabandoned PCNA clamp may actas a marker to recruit Lig1 andmay help orient it as well. Lig1may also employ the slidingclamp to track along DNA until itlocates a nick.
Provided Lig1 binds PCNAwhile both proteins encircle DNA,as modeled in Figure 2B, Pascalet al. [3] make the point that Lig1would effectively mask all threeprotein binding sites on the PCNAtrimer, thereby excluding otherproteins from binding the clamp.This steric exclusion arguesagainst the ‘toolbelt’ model inwhich trimeric PCNAsimultaneously binds threedifferent replication proteins [16].For example, instead of FEN1 andLig1 both binding PCNAsimultaneously during Okazakifragment maturation, FEN1 wouldneed to be dislodged from PCNAin order for Lig1 to interact withthe clamp. How FEN1 and Lig1switch places on PCNA, andwhether Lig1 has a specificmechanism to displace FEN1 fromPCNA must await future studies.
The structural snapshot of theligase ring about to seal the nickbrings to mind new questions.How does a protein encirclingDNA dissociate after sealing thenick? Perhaps the rigidity of thefully double-stranded DNAproduct provides energy to openthe Lig1 ring. How does Lig1assemble onto DNA? Does theenzyme adenylation stepdestabilize and open the ring?Alternatively, ligase may not be astable ring without DNA. Indeed,the T. filiformis ligase–AMPstructure requires largeconformational changes to bindDNA as a ring. Finally, how doesligase integrate its actions withother binding partners? Thesequestions and many more suggestthat exciting studies of this
fascinating enzyme will continuewell into the future.
References1. Kornberg, A., and Baker, T. (1992). DNA
replication, 2nd Edition (New York: W.H.Freeman).
2. Lehman, I.R. (1974). DNA ligase:structure, mechanism, and function.Science 186, 790–797.
3. Pascal, J.M., O’Brien, P.J., Tomkinson,A.E., and Ellenberger, T. (2004). HumanDNA ligase I completely encircles andpartially unwinds nicked DNA. Nature432, 473–478.
4. Subramanya, H.S., Doherty, A.J.,Ashford, S.R., and Wigley, D.B. (1996).Crystal structure of an ATP-dependentDNA ligase from bacteriophage T7. Cell85, 607–615.
5. Singleton, M.R., Hakansson, K., Timson,D.J., and Wigley, D.B. (1999). Structureof the adenylation domain of an NAD+-dependent DNA ligase. Structure FoldDes. 7, 35–42.
6. Odell, M., Sriskanda, V., Shuman, S., andNikolov, D.B. (2000). Crystal structure ofeukaryotic DNA ligase-adenylateilluminates the mechanism of nicksensing and strand joining. Mol. Cell 6,1183–1193.
7. Lee, J.Y., Chang, C., Song, H.K., Moon,J., Yang, J.K., Kim, H.K., Kwon, S.T., andSuh, S.W. (2000). Crystal structure ofNAD(+)-dependent DNA ligase: modulararchitecture and functional implications.EMBO J. 19, 1119–1129.
8. Tomkinson, A.E., Tappe, N.J., andFriedberg, E.C. (1992). DNA ligase I fromSaccharomyces cerevisiae: physical andbiochemical characterization of theCDC9 gene product. Biochemistry 31,11762–11771.
9. Shuman, S. (1995). Vaccinia virus DNAligase: specificity, fidelity, and inhibition.Biochemistry 34, 16138–16147.
10. Levin, D.S., Bai, W., Yao, N., O’Donnell,M., and Tomkinson, A.E. (1997). Aninteraction between DNA ligase I andproliferating cell nuclear antigen:
implications for Okazaki fragmentsynthesis and joining. Proc. Natl. Acad.Sci. USA 94, 12863–12868.
11. Montecucco, A., Rossi, R., Levin, D.S.,Gary, R., Park, M.S., Motycka, T.A.,Ciarrocchi, G., Villa, A., Biamonti, G., andTomkinson, A.E. (1998). DNA ligase I isrecruited to sites of DNA replication byan interaction with proliferating cellnuclear antigen: identification of acommon targeting mechanism for theassembly of replication factories. EMBOJ. 17, 3786–3795.
12. Lopez de Saro, F.J., and O’Donnell, M.(2001). Interaction of the beta slidingclamp with MutS, ligase, and DNApolymerase I. Proc. Natl. Acad. Sci. USA98, 8376–8380.
13. Warbrick, E. (2000). The puzzle ofPCNA’s many partners. Bioessays 22,997–1006.
14. Bambara, R.A., Murante, R.S., andHenricksen, L.A. (1997). Enzymes andreactions at the eukaryotic DNAreplication fork. J. Biol. Chem. 272,4647–4650.
15. Stukenberg, P.T., Turner, J., andO’Donnell, M. (1994). An explanation forlagging strand replication: polymerasehopping among DNA sliding clamps. Cell78, 877–887.
16. Maga, G., and Hubscher, U. (2003).Proliferating cell nuclear antigen (PCNA):a dancer with many partners. J. Cell Sci.116, 3051–3060.
17. Gulbis, J.M., Kelman, Z., Hurwitz, J.,O’Donnell, M., and Kuriyan, J. (1996).Structure of the C-terminal region ofp21(WAF1/CIP1) complexed with humanPCNA. Cell 87, 297–306.
1Howard Hughes Medical Institute, 2TheRockefeller University 1230 YorkAvenue, Box 228, New York, New York10021-6399, USA.E-mail: [email protected]
DOI: 10.1016/j.cub.2005.01.025
Current Biology Vol 15 No 3R92
Seth S. Blair
The proteoglycans are a majorcomponent of cell surfaces andthe extracellular matrix [1]. Theyare made from a core proteindecorated with one or moreglycosaminoglycan side chains,unbranched carbohydratepolymers made of disaccharidesubunits. The two major familiesof cell surface proteoglycans arethe transmembrane Syndecans,which are decorated with theglycosaminoglycans heparan
sulfate and chondroitin sulfate,and the Glypicans, which aredecorated with heparan sulfateand are anchored to the cellsurface via a glycosylphos-phatidylinositol (GPI) linkage.
Proteoglycans play a number ofdifferent roles, but one of the mostintriguing is the regulation ofsignaling between cells. Specificglycosaminoglycans can recognizeand bind members of severaldifferent families of signals and, inhumans, defects in Glypicansinduce Simpson-Golabi-Behmel
Cell Signaling: Wingless andGlypicans Together Again
The role of the Glypican proteoglycans in Wingless signaling has beencontroversial. New studies show that the Glypican Dally-like can haveboth positive and negative effects on Wingless signaling; moreover,signaling can be regulated by removing Dally-like from the cell surface.
Recommended