
Gene, 85 (1989) 391-402
Elsevier 391
GENE 03272
Divergence of the nucleotide sequences encoding xanthine dehydrogenase in Calliphora vicina and Drosophila melanogaster
(Recombinant DNA; rosy locus; protein structure; enzyme evolution; eye pigment)
Mario Houde *, Marie-Catherine Tiveron and Fraqois Br6gkg&re
Laboratoire de Gtnttique et Biologic Cellulaires du Centre National de la Recherche Scientifique, 13288 Marseille Ckdex 9 (France) Tel. 91269603
Received by J.-P. Lecocq: 18 May 1989 Revised: 4 July 1989 Accepted: 26 July 1989
SUMMARY
We present here two nucleotide sequences, from two different alleles encoding xanthine dehydrogenase in Culliphora vicina. One sequence covers the first exon with 1529 bp upstream from the initial ATG and 1737 bp downstream from the donor end of the first intron. The other sequence starts 2537 bp upstream from the acceptor site of the first intron, and ends 662 bp downstream from the putative polyadenylation site of the transcript. Comparison with the homologous gene from Drosophila melunoguster (rosy) reveals extensive divergence, with differences in the splicing patterns and no detectable homology between introns or flanking regions. Nevertheless, there is 76% identity between the amino acid (aa) sequences. The pattern of aa differences has been analysed and correlated with predicted three-dimensional (3-D) parameters. These studies consistently suggest that the evolution of the protein was strongly biased for conservation of its 3-D structure. Possible functional significance of the aa changes is discussed.
INTRODUCTION
Xanthine dehydrogenase (XDH; xanth- ine : NAD + oxidoreductase, EC 1.1.1.204) is active in the catabolism of purines in all organisms. In D. melunoguster, it is present in the developing eye
(Barrett and Davidson, 1975) where it catalyses the specific oxidation of pterin into xanthopterin, a pre- cursor of the eye pigment drosopterin (Dutton and Chovnick, 1988). In other Dipterans such as Culliphoru, however, the eye pigment essentially con- sists of xanthomatin, which comes from a separate
Correspondence to: Dr. F. Bregegere, at his present address: Institut Pasteur, Unite de Biochimie Cellulaire et Moltculaire, 28 rue du Dr. Roux, 75724 Paris Cedex 15 (France) Tel. (1)45688379; Fax (1)43069835. * Present address: Montreal General Research Institute, Neuroscience Unit, 1650 Cedar, Montreal, Quebec H3C lA4 (Canada) Tel. (514)937-8951.
Abbreviations: aa, ammo acid(s); bp, base pair(s); C. V. Xdh, gene encoding C. V. XDH; C. Y. XDH, C. vi&a XDH; 3-D, three-dimensional; D. m. Xdh, gene encoding D. m. XDH; D. m. XDH, D. melunogaster XDH; kb, kilobase or 1000 bp; nt, nucleotide(s); ORF, open reading frame; SDS, sodium dodecyl sulphate; SE., standard error; SSC, 0.15 M NaCl/O.OlS M Na, . citrate pH 7.0; wt, wild type; XDH, xanthine dehydrogenase.
0378-I 119/89/$03.50 0 1989 Elsevier Science Publishers B.V. (Biomedical Division)

392
biosynthetic pathway. Pteridins are virtually absent from the eyes in these species (Langer, 1975). Molecular features of XDH that are devoted to the biosynthesis of eye pigment in Drosophila are, thus, probably absent in Calliphora, and the Xdh gene, which has been analysed in considerable detail in D. melanogaster at both the genetic and molecular levels (Dutton and Chovnick, 1988), seems to be a favourable system for studying limited evolutionary changes in gene function.
The cloning and partial characterization of a wt Xdh allele from C. vicina was reported in a previous paper (Rocher-Chambonnet et al., 1987). Here we describe the general structure of the gene, and present its complete coding sequence. The contri- bution of polypeptide segments to the 3-D structure of the protein was predicted by computer analysis. These results were paralleled with the available data regarding the Xdh gene of D. melanogaster (Lee et al., 1987; Keith et al., 1987), and discussed with respect to the evolution of XDH in the two species.
MATERIALSANDMETHODS
(a) cDNA library construction and screening
Total RNA was extracted from late third&star larvae as described by Fyrberg et al. (1980). Poly(A) + RNA was selected by affinity chromato- graphy on oligo(dT) cellulose, and was used as tem- plate to produce double-stranded cDNA, using the cDNA synthesis system from Amersham. The cDNA molecules were cloned in vector ngtl0 (Hyunh et al., 1985) using the Amersham cDNA cloning system: internal EcoRI sites in the cDNA were methylated, then EcoRI linkers were added at the ends, and terminal EcoRI cohesive ends were created by subsequent EcoRI digestion. The pro- cessed cDNA was then inserted into the EcoRI site of 1gtlO and packaged in vitro.
The phages were plated, the plaques were trans- ferred to nitrocellulose and the filters were hybridised as previously described (Rocher-Chambonnet et al., 1987). To isolate cDNAs that are homologous to the first exon of D. m. Xdh, a PvuII fragment overlapping the 5’-end of the exon II of C. v. Xdh (fragment a in Fig. 1) was used as probe. To isolate cDNA clones
overlapping the 3’ moiety of the transcript, an AvaI-Hue11 fragment (fragment b in Fig. 1) was used.
(b) Computer and statistical methods
Nucleotide and aa sequence analyses were made using the PCGENE program package from Genofit. The alignment programs NUCALN and PRTALN were provided by Dr. D.J. Lipman. Substitution rates were calculated using the program LWL85, which was provided by Dr. W.-C. Li.
RESULTSANDDISCUSSION
(a) Cloning and sequencing the DNA fragments that
encode Cdliphora vicina XDH
We have previously isolated a recombinant phage, Lcvl (Rocher-Chambonnet et al., 1987), which contains a fragment of the C. vicina genome with high sequence similarity to the rosy gene of D. melanogas-
ter, here referred to as D. m. Xdh. The first 8062 bp of the 11.5-kb Hind111 fragment from Lcvl, which contain the region of interest, have been sequenced (Fig. 2). This sequence starts from the first Hind111 site, and ends at the second Hue11 site of Lcvl (Fig. 1). Obvious sequence similarity was found with exons II, III and IV (68 y0 sequence identity between homologous fragments), but not with exon I of D. m. Xdh. Still, a coding sequence corresponding to the first exon is expected in C. v. Xdh, because a stop codon in frame and an intron acceptor signal (Mount, 1982) are present just before the first codon of exon II.
A Calliphora cDNA which is homologous to exon I of D. m. Xdh was isolated from a cDNA library as describedin MATERIALS AND METHODS,
section a. The sequence of this cDNA clone, cXdh1, was determined. Its last 459 nt are identical to the first 459 nt of exon II in Lcvl. Immediately upstream from this block, a stretch of 27 bp displays 21 base identities with the terminal 27 bp of D. m. Xdh exon I. Further upstream, sequence similarity with D. m. Xdh disappears (Fig. 4).
To isolate the genomic counterpart of cXdh 1, the original genomic library (Schenkel et al., 1985) was

393
c-xdhl c-xdh3
Calliphora vicina Y i EPA H AV ECAB P CVD
III I II IIII i I III LcVl
‘: . :
a b :.
: : Drosophila melanogaster -I I_ Carnegie 20
I 111 I II I D C E PPC P CP D
0 1 2 3 4 5
I I I I I 1
kb
Fig. 1. C. vicina and D. melunoguster Xdh genes. Solid boxes represent exons. The homologous regions of the Lcvl and Lcv2 inserts, which cover the Xdh locus of C. vicina, are aligned. The dotted lines indicate correspondence with D. m. Xdh exons, cloned in Carnegie 20 (Rubin and Spradling, 1983). The actual distance between the first and the second exons in Lcv2 is about 15 kb. At the top, cXdh1 and cXdh3 cDNAs (split, shaded boxes) are aligned with their genomic counterparts. Probes a and b, which were used to screen the cDNA library, are represented by heavy lines beneath the matching sequences in Lcvl. The restriction sites are abbreviated as follows: A, AvuI; B, BglII; C, &I; D, HindIII; E, EcoRI; H, HueII; I, HincII; P, ⅈ V, PvuI. Among the ZfincII sites, only those two that define the sequenced fragment (Fig. 3) are shown. The map of D. m. Xdh is derived from the sequence data of Lee et al. (1987) and Keith et al. (1987). The sites with boxed symbols are conserved in both species.
screened again, using a probe that was derived from cXdh 1, and overlapped only 4 nt of the second exon. Several positives were obtained. Their restriction maps suggested that they had overlapping inserts (not shown). One of them, called Lcv2, was studied in more detail (Fig. 1).
The exon-I specific probe hybridized with a 3354- bp H&II fragment which was sequenced (Fig. 3). There is no ORF over 200 bp in this fragment. Se- quence similarity with D. m. Xdh was searched using dot-matrix and alignment methods. There is only one region which is clearly similar in the two sequences: it overlaps the 3’ end of exon I and a few nt at the beginning of the fast intron in Fig. 4. While short fragments also display some sequence similarity, these are interspersed and often contain A,, or T, runs (the average A + T content is 74% in the non- coding parts of the HincII fragment).
(b) Xdlr polymorphism in Culliphoru vi&a
Phage Lcv2 hybridises with probes that are speci- fic for exon II as well as for exon I of C. v. Xdh (results not shown). As the Xdh gene is unique in C. vicina (Rocher-Chambonnet et al., 1987), this means that the Lcvl and Lcv2 inserts overlap within the Xdh locus. Homologous DNA fragments were identified by Southern blotting, and are aligned (Fig. 1); differences in their restriction maps suggest that they represent different alleles. This is consistent with the fact that several polymorphisms exist at the Xdh locus in our laboratory population (Rocher- Chambonnet et al., 1987).
The cXdh1 sequence is identical to that of Lcvl at the 459 homologous positions of exon II, and hence, represents the same allele. It differs from Lcv2 at only one of the 106 homologous positions in exon I

394
1
526
1051
1576
2101
2626
3151
3676
4201
15 30 45 60 75 90 105
AAGCTTCAACCTGTAA*ATAAATTTCAATXAAATGTTATAA**T~TTAT*TTTTGGTA*~AATTCC*TTTG*AT~*AACA*ATCTAG*C~AA*TTAT*CAT*C*~ TCAAATTTATGCACTTCAGATGTTGAAAACAGATTGGAAAATTACTTATTATACGTTTCTTAGTGTTCGATTTATTTCGCTTAGCAATAATTGCAAAATATTTGA TTGATTTGTAACCATAATAACATCGTTTTTAAATATGAGTACTTACTTGTCTTATCAACTTTGTTGTTTATATATCAATATATATGTATATTTCATTAAAAAACA ATAACAATTTTAAAATGGCAACTTTTTGTGTAACCTTAATGAAAAAAAAAAAATAACACATAAAACATCTTATCAATGACTGCTGTGAGACCCCCCTGTGACGTA CAGATAATAACAATCTTAAAACTGTTATACTGCACTTTTGTAGGACTAATGAATTAGTAAACAAATAAATAACTCTGTATAAATATAGAACATTTTATTATCCAC ATATZAGTTAAATTTGTCTTGGAGGTCTAATAGCTAGTTTCTGGGATAATAAAATCTACATTCTTCGTTTAAACCTAGATTAAGTAAACTACAAATTTTATTTCT CACTAATGGCGTAAAGAGTATATGTAATCTAGGTTTAACTGAGCATTATTGACGAGTTAAACCTATGAAAAAAATTATTCCAATTGATTTTGTTTCTTCAGATTC CATTFGATTTCGACAGTTCCCAAAATATTTTATAATTCTGAATTTCCAAATTTAATTTGGTTTTTGGTAACCCCCTGTTTATACTTGACAAATTTTGATCATAAT AATTAATGA>TTGTTGCCAAGACAAAGTTGTCCTGCAATAGCACTAACAATTGTGTCATAACGAAGCAATAATACTAATAAATGAAGACATTACCTGAAATTT TTTTTATCTTGACAACAATTTTGACTTTTATCATGATAAAAATTTATCAGGTATAGATAATTATTTAAAAATATCTGAAAAACAAATAATTACAAACTAATAACA AT~GACATATAAGACATTCACACGATGAATTTTCAGTCATGCAATTGTAGCAATTCTCTTTTGAATTTTCTTTTACTCCACCCCTCTTGCCAACAAACTCATAA AATCACTTAGAGTGACATGACAAAACAAGACATCGTAAAAATTCCTTATTAAAAAAAAACAAGTTAGAGAGCTATATTCGGCAGTGCCGAATCTTATATACCCTT CACCAAATTATACTTCAAAATAAAAATTTTAAATATTTTTAAGTAAACAAATTTTTTTTTCCAAAGTTGT~TTTTAAGTTTTTAGGAAATTTTTTTTTTCGAAT TGTTATTTTAAAATTTTTAAACAGCTGTTACGAACAAAATCAACTATGTATTGTGAATGAAAAGTTCATGAGAATATTACGATAGCAATTTTCCCTTTGAGGGAA CATAAATT~CACGTATTATTGCCGATAGGGTCTATTGACTAGCATGTATTTTGTTGTTGCAAGAGTTAGGTTTTGGACAACAGACTTAGGTTTTTGACTCGTCTC GTCTTTTATTACACAGTTTTATTTTATATTTATTATTACAGAAGTTTAAAAGTTTCCATTGTTTTGCCAATGTTAAGCATTATATCCACATATACATGTATTTTT GTTAATTATTCCCTATATTTTAATAGAAAACCTGGTGAAAGGTCATATTTTTTAAAACTGCACAGAGAAAATGATTCGGTTGCAGACATAGACATTTTCGGTTCT ATCAACAGAAAATCAGTTGATAAAGAAGAATTTCCGTTGAAGCAACCAAACTTTTGTTACCTCTTCTAAAATGTTGTAGCCATAACTGTAAAATTCGGTCACTAA GAATCATTCGATTGGCAACAAATTCGGTTGCTATCACGAATCTGTTTTCTCTGTGTGAAAGTATGGAATATTGTTTTAAAAAAGTTTCCATTTTTTCGCTAGTAG GTCTGGGTTAATTAATAACTCGCGQCTTCGTATAACGATTCCTGTTCTACGTCGTTTACACCAACCGCATGTATTTAATTTGAACAAATCCTGGTGGAGACCATA AAAACCCTTCGATACTGGTGGAGTATCATTTCAAACTCAAATCGTAAATTGGTAGAGAACCATATAAAACTCGACGGATCAACTGGAGCAGTATCATTATCACTC TTGATTTTAATAACAAGGTTTAAGCATTCTCACTTCTATGAAGACATATGTAGGTCATTTAGGCTTACTTAAATAGACATGGGATAACCTGGAAAATTTGTGAGA CGTTATACTCAACCAACAAACTTTAAAACAATTTTATTGTCACTCTCTGTCTAAAGCATCGTAATCTATACATCACTTTACTTCACAAATGTTACAATACTCTCC GTAATTAATCTTAAATA CATCAATTACACATTTTGTTTAAGTAATTTAAAAATATTAAGTTCTGCATTCTTAAAATACAGCTTTGATTCAAGAAATATAACTAT
iexun TTGTCTTATTTTCCTAGGTAATTGATACCAATCCCGATCCAGAATGTACCCTTCTAACATATTTACGTGAAAAACTAAGATTATGTGGCACAAAATTGGGCTGTG
ValIleA~pThrAanProAspProGluCyslhrLeuLe”ThrTyrL~“ArgGl”Ly~LeuArgLeuCysGlyThrLy~LeuGlyCy~Gly
GTTTTTGTACACCCGGTATTGTCATGTCTATGTATGCACTGTTGAGAAATTTATCCCAGCCCTCGATGAAAGATTTGGAAATTGCTTTCCAGGGAAATTTGTGCA PheCy~ThrPraGlylleVel~etSerMetTyrAlaLe”Le”ArgA~“LeuSerGlnProS~rMetLy~A~pLe”GluIleAlaPheGlnGlyA~nLeuCy~Arg
GATGCACGGGCTATAGACCCATTTTAGAGGGTTATAAAACATTTACTAAAGAATTCGGCTGTGCTATGGGTGATAAATGTTGTAAGGTGAATGGCAATAAATGTG Cy~ThrGlyTyrArgProlleLe”GluGlyTyrLysrhrPh~ThrLy~Gl”PheGlyCysAlaMetGlyAspLysCy~Cy~LysValA~nGlyA~nLyaCy~Gly

4726
6301
6626
7351
7876
Fig. 2. C. vicina Xdh locus: nt sequence from Lcvl. This sequence starts within the large tirst intron and ends in the 3’4lanking region of C. v. Xdh. The intron boundaries are indicated by arrows. The putative polyadenylation site is indicated by a double slash and a line break, and the possible polyadenylation signals are underlined. The asterisk indicates the final stop codon. Sequencing methods: DNA fragments were subcloned into the plasmid vector Bluescript (Stratagene) and subjected to progressive digestion by exonuclease III (Yanisch-Perron et al., 1985; Henikoff, 1987). Sequencing was carried out as described by Sanger et al. (1977). All sequences were determined for both strands.

396
1
526
1051
1576
2.101
2626
3151
15 30 45 60 75 90 105
AAcTGTTcGAATAA~TTGAA*T~GccG*T~TTc***T**c****~*AccQ*TT**~~~G~G~cA*~TAAccGAT~A*ccGA*AT~cc~T~TTTT*GcQc~TT*A~ ATTTTTTTTTGTATTTATTTTTCCQTTTCAATTCAAATACAAATTAAAATTTGATATTTTTTGAACATCCAATAAACATQATTAATQAGTATQTATAACAACTGA CATGTTTTGAAACTTTGTTTGTATTTACATGTATAGACTAAACTTTTTTGAATTGAGTCTTTTATCATAACTAAATQAATCTATTAAATTAATCCAAATCTAAAA CAATCCAAAAAAGAATATTCTTTATCATGTTTTTQATTTCTCCAACATATAAAATCAQTGAQAGAQTTTTTTCCAAGAAAAQTTTTATTAAATCTGAAGAAAAAA ATCGTTTAAATTATATTCTTTTTATAAAAGATTATTTCAGAAGATCTCTTGCTGCAACAACQTAATAATTAAAAAAAAGTCGTTTCCAAAAAATGTCTTTGAATA TTTTATGACATTTTACTATTTCTTATATAAATTTTCAACAAATCATGCGATTTCAGAAATATAAATAGTAGATGTTAATATCTTTCTAATCTGTATTTAACCCAA ATTTTTACTTATQAAATATACGAGAAAACATGAATTTTAATTTTGATGTT~GCAATAATAAAAAACACTCTCACACAAAAAATAATTGACTTTTTTGAAAACTAT ATGAAAGATTATAGATCAGCATATTTTGTATTACTCAGTTCATAAAACACQGATTTTGAGTTATQACGTTGTTGCAQCAAATACAAAAAGATTACAAAACTGATC TCAAATACCTTATTTGCTGTTTTTTATGTTTTTGTACACAAAATTCGTTGTTGTATTTTCAATTTAAAATGAAAATAQAAATTATTCGGTTAATCGAATAATTTT AAATTAACCGATTATTAACCGAATAAATCTAAACCTCGATTAATTATTTGCTCQATTAACCQATTAAACCAGAGACCCQATTAATTGAATACCCTACCCCCATCT TTTTATGCAATTTTTGCCCTGTAAATQTCAATQCGACAACACTACCACTACTGTATATTTCTTATTQTAGCQCCCTCATGAQTATTAAGACATTTACTCTCGCCC TCGATTTTATTTGTATTATTCTCTGGT~TCACACTTGTTGAAAATTTATACACAAATATCTTAGCGACTCTCTACCGTAQCCAGCCGGTTGTACAATTAAT
TATTAATACCAGTATAAATAAAATTTTTATATTCAAATAAATAAATAATTTQTATTCGACTCTCAGTTCTCCAGTTTQCAACGTGTGATCATTGTG~ACGT ATCGTTTAAAAGTGCTTATAATATACGTACAATATTTCATATTQCGTTTATTATCGATTGTTTGATTAAAAAAATTGAATTGAAACGAAAATAAAGACATCATTG
E cXdh1
CAATTAGTTAAATAAAAATT~ATAAAA~AAGTG AA~~CAGACAGCTAAACGTTTAATATGACGCAAGAACATAA~GCAGCAGTCCTAQATTTAAA~CCAACATT
exon I t
MBTThrGlnQluHi~AsnAlaAlaValleuA~pLeuAsnProThrPhe
TTCTACATTGATATTTTTTGTTAATGG~GGTAATTAATGTAATAGTATTTAAAAATAATTTATGTATGAAAACAGTTAGTATAATATTTGQTTAAAACAT SerThrLeuIlePhaPheValAsnOlyLysbys
AACTATACATTGTGATTGAGGGGGGGTTTAGQQAACAAATTTTATTAACACACATATTTAAATACACGAAAATTTTATAAATAAAAQATATTTGTTGATTATTTA TTGAGTCAATTCCCTAATATTTGTTGTAATCGTACTCGCATTGTTATTGTCTGTCATACATAAAAACTTCTGTCTGTCTGTCTCAACTTCTCATCAAACGAGTAA TCATCATGAACACTTATACCTGQTTTGACQGCATAAATAATAAAATAGAGTTGCACAGTGAGGTCGATCGTAAAACATGCTAATAAATCTGAATCTTCTGACCTA TTTCGATTACAAAATGATQTATAAAAAACTAACTTTGAAAAAATAAATTTTAQTATAATTTGTATGCAATTAAAATCTGCTCCCCTQAQCTTGTTTTAGGACGGG TGATTAATGTTTAAATGTCAGATTAATATTCTAATTAATATGTTTATTATAATTTGTAAAAAAAATTTAAGAACTAGATGTGACACTTAGAATTTTTTTTTTTTT AAATCACTATTTTCTTCCTTTGCCTGAAQTAQGQTACATATCGATTTTAAAATCTTATCGAAAATGAAAAAATATTACAATTAAQCTACATTTTTGTCAAAATGC ATCCATTTTTATATCCTTCACCATGAGTGGCATAAGTCTQTCATTCCGATTGTAAAATCTACATTTTTCATTTTCGACCCCACAAAGAAAATATATTCTAGATCG TTATACATCAATATATCGGGAATTCTTCCGGCTCQGAAACTATTTAAAACCGAAAAAATCGGCTGAGATATAAGGAATAAACCQQQACAACCTCGACTCTTGGCC TATTTTTGATTTATATTTTTTTGTCATAAATTAAAAAAAGAATTAAAAATAAAATTTGGAAAAAACTTCTTTAAACAAAATTAAAAAACAAQTTAQAGAGATACA TATATTCGGCCQTGCATCTTATATACCCTTCACCAAATTATACTTCAAAATAAAAATTTTAAATAQTTTTAGGTAAACAAAATTTATTTTTTTTCCAAAGTAQTT TTTTTTTAATTTTTTGGAAAAAAAAATTTTCGAATTGTTTTTTAAAATTTTAATTTTTTTTTTGTTTTTTTAATATTTAGCGAAAAAAAATTTTTTTGTGAAAAA AAAATTAACAAAAAACTATTTACATGTCATTATAAATTTTTTGAAATAAGATTTTGACATTCTCGTAAAATCAGCCTAAGQTGTTCCAACATTTTGATTAAATAA
TATCTAGTTGATTTTTTTQTTQTTGTTQCTTTTQTCTATCATTCAGQTTCAGTTQTAAACATTAACGTQTTTTGTTTQCCTTATCACTTGTTTGCATTTTTAGAA
AATGTGTTTTAATTAAATTAGTTTAGAAAAQTTTTTTAAATGGAAATTATTTTAAGAGCATTGTCTGQACATTTATAGATATATATTATTGTTAATTGTATTAGT
TAAATTTTGAATAAATTGTGGAAAAATATCAQTTATTTTTTQGTTAAATTAAGQAATTAGTACATAAAAACGAAAQATAQCTGAGAATTTCCATATTAGCGAATG
CTTCTTQQCTTCCGAGTAAAAACTCQCCCGTTAGCGAATTCCATCATAQCGTTGTCAATGATTCTTACAGAAACATCAAGTATTAAATGTT
Fig. 3. C. vi&a Xdh locus: nt sequence from Ltd. The sequenced fragment is defined by two HincII sites; it starts 1529 bp upstream from the initial ATG, and ends in the first intron of C. Y. Xdh. The positions of the first nt of the cXdh1 cDNA, and the last nt of exon I, are shown by arrows. Possible transcription signals are underlined. Sequencing methods were as described in Fig. 2.
1540 1560 1560 GluHi~A~nAlaAlaValLeuA~~L~~A~nProThrPheS~rThrL~~llePhePh~V~l OAbCATAATGCAC~TCCTAGATTTAAATCCAACATTTTCTACATTGATATTTTTTGTT
: ::: - : :: : : :: ::: : ::::: :: CCGTGAGCTACAGATACATAGCACTTCACGATCTCTAACTCQGTTTTGQTGTTTTTCGTO
~BTSerAsnSerValLsvVslPhePhsVal 1500 1520 1540
1600 1620 A~oGlyLy~Lys AATGBCAAAAAG OTAATTAATGTAATAGTATTTAAAAATAAT... ::::: :: ::: ::: :::: : : AATGDAAADAAG OTACTTAAADATACTCGGA&CCTTGCOTG AmGlybysLya +,tron I
1560 1560
Fig. 4. Aligmnent of C. v. Xdh and D. m. Xdh in the region of exon I. The Calliphora sequence is from Lcv2. This alignment is the best fit when gaps are not allowed. Matching nt are indicated by colons. The position ofthe first nt ofcXdh1 cDNA is indicated by a downward arrow. The cXdh1 sequence is identical to that of Lcv2 in the fragment shown, except that an A is substituted by a T at position 1549. The nt at this position is boxed. A gap and an upward arrow indicate the beginning of the first exon.
(T instead of A at nt position 1549 of LcR, making a silent substitution and creating a PstI site in cXdh1).
(c) Overall structure of the gene
The first intron interrupts the coding sequence at the same position in C. v. Xdh and D. m. Xdh, but it is about 10.5 kb long in Calliphora, compared with 8 15 bp in Drosophila. While blocks of sequence simil- arity with D. m. Xdh exist in the first intron of D. pseudoobscura Xdh gene (Riley, 1989), such blocks were not found in the sequenced regions of the first intron of C. v. Xdh (1737 bp at the donor end and 2537 bp at the acceptor end were sequenced).
Sequence similarity between the first exons of C. v. Xdh and D. m. Xdh ceases upstream from the sixth codon of D. m. Xdh (Fig. 4), so that the un- translated leader sequences, as well as the sequences further upstream, seem unrelated. In the first intron, only a few bases are conserved at the donor site. At least one important rearrangement (i.e., insertion, deletion or transposition) must have contributed to

391
these differences. This results in different N termini for the two XDH proteins, and different sequences upstream and downstream from the coding segment of exon I. As the 5’-flanking region and the first intron are important to the regulation of the D. m. Xdh gene (Dutton and Chovnick, 1988), these results suggest that Xdh regulation may be different in Calliphora.
Typical polymerase II-promoter sequences were not found at the expected positions in D. m. Xdh
(Lee et al., 1987). In C. v. Xdh, the only typical TATA box (Bucher and Trifanov, 1986), within the first 900 bp upstream from the initial ATG, resides at position 1273 (Fig. 3). Additional experiments are needed to determine the actual extent of the trans- cript.
A cDNA clone overlapping the last exons of C. v. Xdh, cXdh3, was isolated as described in MATERIALSANDMETHODS, section a, and sequenc- ed. Its cDNA insert extends from the second EcoRI site of exon II into the 3’-untranslated region (Fig. 1). Its sequence is identical to that of the ho- mologous fragments of Lcv 1. Its structure reveals the splicing pattern indicated in Fig. 1: while exons I and IV match, the second introns are found at differ- ent positions in the two species. Thus, exons II and III are 3762 bp and 168 bp in C. v. Xdh, and 2601 and 1314 bp in D. m. Xdh, respectively. The sizes of the introns are not conserved, and no se- quence similarity was found between homologous introns of the two species.
Interestingly, another Drosophila species, D. pseudoobscura, contains an additional intron in its Xdh gene (Riley, 1989) which sits exactly at the same position, and has the same size (67 bp), as the second intron of C. v. Xdh. This suggests that this intron was present in a common ancestor species and has been lost in D. melanogaster.
The 3’-untranslated region of the Calliphora Xdh transcript extends until at least position 7400 (Fig. 2). At this point, the genomic and cDNA se- quences diverge, with an A, run in the cDNA. The polyadenylation signal, AATTAA (Bimstiel et al., 1985), is found twice, 30 bp and 9 bp upstream from position 7400. Hence, this position is likely to be a polyadenylation site of the transcript. The 3’- untranslated region is 119 bp long and contains 85.5% of A + T. No obvious sequence similarity with D. m. Xdh can be seen.
(d) Comparison of C.v.Xdlr- and D.m.X&-coding sequences
The overall A + T content is 58% in the coding fragments and 72% in the introns (compared to 44% and 63 y0 in D. m. Xdh, respectively). Codon usage is markedly different, reflecting the greater A + T con- tent in C. v. Xdh. The base frequencies at the third position of the codons in C. v. XdhlD. m. Xdh are: A, 24%/14x; T, 38x/21%; G, 19x/33%; C, 19%/32X.
When the two sequences are aligned over the 1336 homologous codons, 1272 nt differences (32%) are found. The rate of synonymous substitutions (Li et al., 1985) is largely superior to 1, indicating that nt substitutions occurred at least once at most sites that allowed synonymous changes. The calculated rate of nonsynonymous substitutions is 0.178 (S.E. = 0.012)persite. Ifameanrateof0.88 x 10m9 per site per year is assumed (Li et al., 1985), this corresponds to a divergence time of 2.0 x lo8 years, consistent with our previous minimal estimate of 0.8 x 10’ years, derived from paleontological data (references in Rocher-Chambonnet et al., 1987). The contrast between the frequencies of synonymous and nonsynonymous substitutions reflects the role of na- tural selection to maintain the functional structure of XDH.
(e) Comparison of aa sequences
The aa sequence of C. v. XDH is shown in Fig. 5. The complete polypeptide chain contains 1353 aa, corresponding to 150207 Da. Its theoretical p1 is 6.18. Like D. m. XDH, C. v. XDH does not contain a typical signal sequence (von Heijne, 1986). The two sequences are colinear, except at the N terminus, where C. v. XDH has an extension of 13 aa, and at position 203, where it has five additional aa. Excluding these two gaps, the number of aa differ- ences between the aligned sequences is 322 (24.5 %).
The aa replacements were examined with respect to the chemical classes of the residues. Table I shows that conservative replacements are favoured. Lower replacement frequencies are found for glycines (6.9%), prolines (11.6%) and cysteines (13.1%), which have key positions in the peptidic backbone (Richardson, 1981). This suggests that aa replace- ments were restricted to maintain the 3-D structure

398
1
101
201
301
401
501
601
701
801
901
1001
1101
1201
1301
MTaBHNAAVLDLNPTFSTLIFFVNGKKVIDTNPDPgCTLLTYLREKLRLCGTKLGCGEGGCGACTVMISRIDTLTNRIKHIAVNACLTPVCAMHGSAVTT eeeeeeeee///..*//////eee^^^////...^^//eeeeeeeeee//-^///^^^"^^^ /////////eeeeeee///////^//////ee.e/////
iiiiiiiiiiiiiiiiiiiiii iiiiiiii i iiiiiiiiiiiiiiiiiiiiiii iiiiiiiiiiiiiiiiiiiiiii SN_--___-______ V V TKVS F A V L RRA K R L s c
VBGIGSTRTRLHPVaKRLAKAHGSaCGFCTPGIVMSHYALLRNLSaPSMKDLEIAFaGNLCRCTGYRPILEGYKTFTKEFGCAMGDKCCKVNGNKCGBGM ///...////...eeeereee^^^^*"^///^//////eeeee.....eeeeeeeee^^^^^*^^///////^eeeeeeeeee~eNee////~~~~~./// iiiii i iiiiiiiiiiiiiiiiiiiii iiiiiiiiiiiiiiiiiii iii iiiiiiiiii
K AE R V AC E S KG TDA
BNGGDMVDDKLFBKSBFVPFDPSaKPIFPPBLOLNKDWDSQTLVYKGERATWYRPGNL~DLLKIKAaFPEAKLVVGNTEIGVBVKFKRFLYPVLVNPTKV eeeeeeeeeeeeeeee//-*..-..^.//eeeee^^^^ee/////////////..^ .eeeNeeeeeeeeeeeee//eeeeeeeeee////////~~eeee
ii “’ T-----l NC R Q1;l
iiii iiiiiii iiiiiiiiiiiiiiiiiiiiiiiiiiiii
SDAF S IFSSD V T E QL KH A V HI a
KBMIDVaBLBDSIYFGASVSLMDIDRILRSSIEKL~EHaTRFFaCAVNMLHYFAGKaIRNVASLGGNIMTGSPISDMNPVLMAGAVKLKVAKYVKGaIKY eeeeeeeeeeNeeeeeee///eeeeeeeee.. . . ..eeeeeeeeeeeeeeeeeeeee////j/.. ^////////..////eeeeeeeeeeeeeeeseeeee/
iiiiiii iiiiiiiiiiiiiiiiiiiiii iiiiiiiiiiiii i iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii
LLBIK Na G A 8 AL QR L SE L T D C S AGAQ E SF D KLQK
RBVCMASGFFTGYRKNVIBPTEILVGLYFPKTLBHaYVVAFKaAKRRDDDIAIVNAAINVFIDPRSITVDKVYMAFGGMAPTTVLATRTADIMVKaaWNK //////ee///^^^..////////////.. *eeeeeeeeeeeeeeeeeeee//eeee//////.^////////////..../////eeeeeee/eeeeeee
iiiiiiiiii i iiiiiiiiiiii iiiiiii iiiiiiiiiiiiiiiii iiiiiiiiiiiiiiiiiiiiiiiiii
S H GT R AR V L IR R TPD I R RFKEK NI AKIS P SPL G B SH
VLMBRVVENLCAKLPLAPSAPGGMIAYRRSLVVSLFFKAYLTITOGLIKSGILPGDSLPGKELSGSDVFHTPALKSAGLFEKVSNKGSECDPIGRPKIHA e//eeeeNeeeeeee//e...^/////////////eeeeeeeee/////////.^^..//eee^^.ee///eeeeeeeeeeee^^^^^///--~/eeeee
iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii iiiiiiiiiiiiiiiiii i ii QV AS T A A A SLK S TSS A P R AKT V R CSD PI V
SALKQATGBAIYCDDMPRMENBLYLALVLSTKAHAKILSIDASEALAMPGVHAFFSSKDIT~HENEVGPVFHDKEVFASDMVYCOG~VIGAIAADNPNFS eeeeeeeee////^e.eeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeeee~~~,ee/..^^ .eeeeeeeeeeeeee////////////...*^e iiiiiiiiiiiiiii iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii iiiiiiii iiiiiiiiiiiiiiiiiii
A T I DGV F PR TKL LD a CY LE H AGKHY IV KALA
SKTARKVTIEYKDIKPVIITlE~AIKHKSYFPDYPRFTEIGDVEKAFSRADHVYEGSCRMGG~KHFYLETHASLAVPRDSDEIEIFCST~HPSEV~KLVA eeeee/////eeeeee///eeeeeee~~~~.~~..-///eeeeeeeeeeeeeeee^^^^^^.eee/eeeeeeee//e.eeeeeeeee~~...^eeeeeee
i iiiiiiiiii iiiiii ii iiiiiiii iiiiiii iiiii QRA L KV ELS v L VTK N BLQ TF T A IL
RVLSTSARRVVCRAKRLGGGFGGKESRAIAVALPVALACHRLRRPIRCMLDRDEDMMITGTRHPFLFKYKIAFTSEGRLTGCYIECYNNAGWSMDLSFSV eeeeeeee/////////^^.... .eeeeeeeeeeeeeeeee^^/^^/////eeeeeeeeeee..eeeeeeee///.^^^^^^//////^^^^/////eeee iiiiiiiiiii iii iiii iiiiiiiiiiiii ii iiiiiii iiiiiiiiiiiiiiiiiiiiii iiiiiiii
TALP GS AYMG V L VG K LIAD
LBRAMFHFKNCYKIPNIKVGGWVCKTNLPSNTAFRGFGGPQGMFAGEHIIRDVARILGKDYLEIMK~NFYKEGDITHY~~KLDNFPIEKCFYDCL~~SNY eeeeeeeeee^^^//////^//////...^/////^^.. . ../eeeeeeeeeee//eeeeeeeeeeeeee^^^^eee///^~~./e~e~~-///~~~~~~ iiiii iii iiiiiiiiiiiii iiiiii iiiiiiiiiiiiiiiiiiiiiii 1 iiiiiiii
R VR Y V R VVDV RL T Y H a KH RIB K R
YaKRKBIEEFNRNHRWRKRGISLVPTKYGIAFGVSHLNaAGALINIYADGSVLLSHGGVEIGaGLHTKMIaCCARALaIPIKFIHISKTATDKVPNTSPT *~.//eeeeeeeee~~~~~~ ///////////////..eeee//////////////.. .///eeeee*/////eeeee//e////eeeeee^^/..^//i/
iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii i
DE QD AR. EN MAV M S G N A G SL
AASSGSDLNGMAVLDACKKLNKRLAPIKRANPNGSWTKWINKAYFERVSLSATGFYRMPDIGYDPVQNPNALMYNYFTNGVGSSIVKIDCLTGDHGVLST
////. . ..//eeeeeeeeeeeeeee/////..^.^.//ee...eee//ee.^^//////^^^///^/////////////~~...///////eeeee///// iiiiiiiiiiiiiiiiii i i iiiiiiiiiiiiii i ii iiiiiiiiiiiiiiiiiiiiiiii ii
V LGTK D A G H ET RT S Y VTV
DIVMDIGSSLNPAIDIGQIKGAFMGGYGLFTLEKMIYSPGGVLYSRGPGMYKLPGFADIPGKFNVTILTGAANPRAVYSSKAVGEPPLFIGCSVFFAIKE
//////^..*. ./////////////^^ ..eeeeeeee./*//////. ^^//////////.^eei/////.. ..////////////////////eeeeeee iiiiiiiiiiiiiiiiiiiiiiiiiiiiijiiiiiii iiiiiiiiii iiiiiiiiiiiiiiiiiiiiiiilii iii iiiiiiiiiiiiiiiii
LM M SL P S A
AITSARLMNGLSBDFKLKSPATSARIRMACPDEFTNLIBQPPAGSYVPWNIVP eeeeeeee..eeeeeeeeeeeeeeee////eeere././/.^^^^///////// iiiiiiiiiii iiiiiiiiii i ii iiii iiiii
AA BDQ G P A S I K 6 L I FP FT
Fig. 5. C. Y. XDH aa sequence, 3-D structure predictions and aa differences with D. m. XDH. First line: aa sequence in the one-letter code, deduced from the nt sequence. Second line: secondary-structure predictions by the method of Garnier et al. (1978): a helices, /I sheets, turns and coils are represented by: a circled a, a slash, a caret, and a full stop, respectively. Third line: interior positions, predicted according to Kyte and Doolittle (1982), are marked by ‘i’. Fourth line: aa that are diierent in D. mehnoguster, in the one-letter code. Gaps in alignment are indicated by a dash.
399
TABLE I
Distribution of the aa replacements per chemical class a
Changed from b: Gl Y Aliph Hydr cys Acid Amide Basic Arom Pro
Changed to b:
Gl Y Aliph
Hydr W Acid Amide Basic Arom Pro
-
4 2 0 1 0 0 0 0
6
59 19
1 4 5 5 3 3
1 0 3 3 3 24 2 5 5 6
8 2 4 10 4 4 - 0 0 0 1 0 15 I 6 2 0 4 2 9 I 1 4 10 17 1 0 0 1 1 3 0 1 1 1
0
2 1 0 3 0 2 0
-
ChangedC I 105 51 5 36 39 41 24 8 Unchanged c 94 322 101 33 113 68 122 99 61 Total” 101 427 152 38 149 107 169 123 69
a In these statistics, the aa replacements found at 322 among 1335 homologous positions of C. v. XDH and D. m. XDH were distributed according to the chemical class of the replaced aa. Changes are understood to be from Cdphoru to Drosophila.
b aa chemical classes: Gly, glycine; Aliph, alanine, valine, leucine, isoleucine, methionine; Hydr, serine, threonine; Cys, cysteine; Acid, aspartate, glutamate; Amide, asparagine, glutamine; Basic, lysine, arginine, histidine; Arom, phenylalanine, tyrosine, tryptophan; Pro, proline. c Changed, number of replaced aa per class; Unchanged, number of conserved aa per class; Total, total number of aa per class.
a
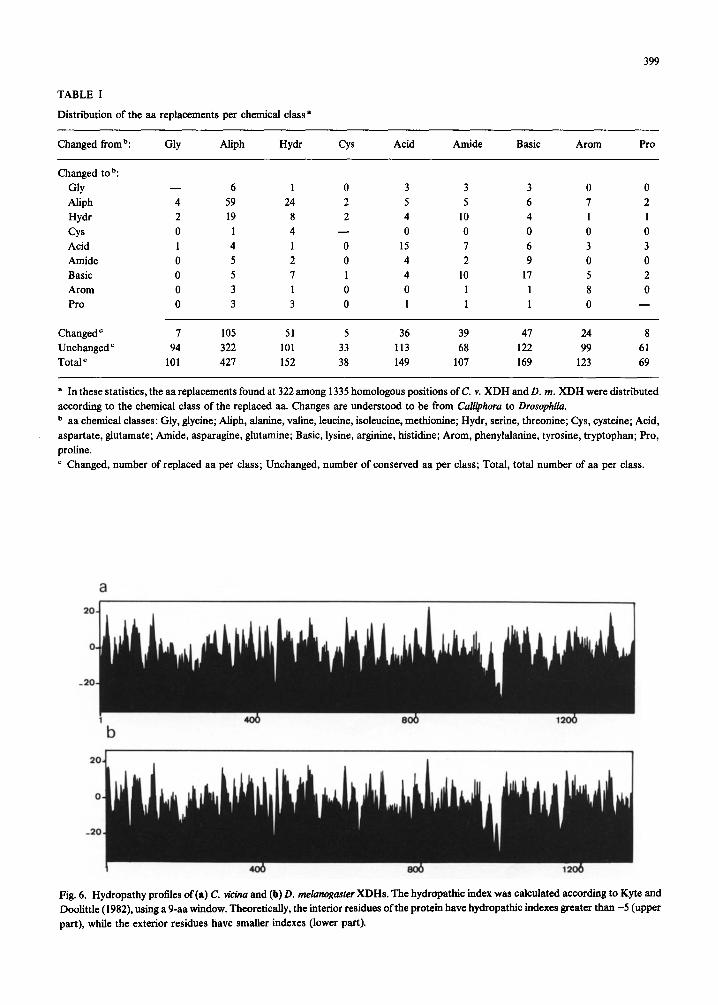
Fig. 6. Hydropathy profiles of (a) C. vi&a and (b) D. melunogaster XDHs. The hydropathic index was calculated according to Kyte and Doolittle (1982), using a 9-aa window. Theoretically, the interior residues ofthe protein have hydropathic indexes greater than -5 (upper part), while the exterior residues have smaller indexes (lower part).

400
of the enzyme. The obvious similarity of the hydro- pathy profiles (Fig. 6) suggests that the pattern of interior and exterior segments, and hence the overall 3-D structure, may be very similar in the two proteins (Kyte and Doolittle, 1982).
(f) Correlation of aa differences with predicted 3-D parameters
When the distribution of aa differences is examined, some regions proved to be extremely con- served (e.g., 4% difference over 80 aa at po- sitions 1186-1265), while others are more divergent (e.g. 47% difference over 91 residues at po- sitions 382-473). To look for a possible correlation of aa differences with parameters of 3-D structure, we made secondary-structure predictions on C. v. XDH, using the method of Garnier et al. (1978). The accuracy of this method, when applied to proteins whose tertiary structure is known, seems
TABLE II
to be 56 to 57%, and we assumed that the inexacti- tudes are distributed at random with respect to the actual 3-D structure. Then, significant association or exclusion between predicted secondary structures and other parameters may be detected, using statisti- cal methods.
Fig. 5 displays the output of the Gamier analysis of C. v. XDH, along with the hydropathy status of each aa position, and with the aa differences in D. m. XDH. Table II shows our statistical analysis of these data. It leads to the following conclusions.
(1) Replacements of aa preferentially occurred at predicted exterior residues. Hence mutations seem to be fared more often at the surface of this soluble protein than in the more hydrophobic, putatively buried peptidic fragments, where structural constraints may be tighter.
(2) Replacements of aa occur more frequently in the predicted o! helices than in the predicted /I sheets, consistent with the current assumption that a-helical
Statistical association of predicted 3-D structures with aa replacements”
Exterior Helices j sheets Turns Coils
Pinf(z)” 99.96% 99.24% 1.13% 68.24% 7.88% Psup(z) a 0.03% 0.53% 98.41% 25.32% 88.28% Concl. ’ associated associated exclusive - -
a The predictions used for these calculations are those of Fig. 5. The correlations were assessed using the following method. Let us consider a set of N positions. Let a fast property, A, be verified at a positions, and a second property, B, be verified at b positions. The number of possible distributions of A and E is:
nl = C: x Cr
where 2 is the number of subsets of q elements in a set of p elements. Let us now consider z, the number of positions at which both properties A and B are verified together. The number of possible distributions that are defined by (I, b and z is:
n2(z) = CN x CNeZ x CNea Z a-z b-z
Hence, given a and b, the probability of having z is:
P(z) = n2(z)/nl
The correlation between A and B is given by the probabilities to find a lower and a higher coincidence between them (i.e., a lower and a higher I, respectively), assuming that the a and b positions were distributed at random. These probabilities are:
x-1-1 X - in@*b) finf(z) = C rQ(x)/nl
x-o fiuP(z) = x _T+ , ~W/nl
b Non-randomness was assumed for values of Pinf(z) or Psup(z) superior to 95%. With increasing values of Pinf(z), the two properties tend to be associated, and with increasing values of Psup(z) they tend to be exclusive. The results of this test are consistent with the results of the x2 test for random distribution. This method is preferred because it provides direct estimates of association and exclusion.

401
structures accommodate mutations more easily than do /.I sheets (Hubbard and Blundell, 1987).
(3) No significant conclusions are obtained for residues predictably engaged in turns or coils. The five additional aa at positions 203-207 are predicted to fall in an external a helix.
(g) How do aa replacements affect XDH function?
The aa sequences of C. v. XDH and D. m. XDH were kept 76% identical in spite of the extensive divergence of nt sequences at the sites of syno- nymous substitutions. Since conservative aa
changes were favoured, and since the aa that have key positions in the peptidic backbone were prefer- entially conserved, it is clear that aa replacements were strongly restricted to prevent 3-D structure divergence, as illustrated by the hydropathy profiles. Recent data on the Xdh gene of D. pseudoobscura
(Riley, 1989) reveal that among 141 aa positions that are substituted between D. melanogaster and D.pseudoobscura, 102 (72%) also differ between C. v. XDH and D. m. XDH. This suggests that the same minority of positions is permissive to aa changes in the three species. But it is not obvious to identify those positions that may be responsible for functional differences, or even to know what pro- portion they may represent. To answer this question, the results of both structural and functional studies must be considered in parallel.
This kind of work was done with hemoglobins from a large range of species, and some differences of biochemical properties were correlated with parti- cular aa replacements (Perutz, 1983). In that study, however, the functional parameters could only be assayed in vitro. Genetic transformation now offers the opportunity to assay the function of a gene by in vivo complementation (Scavarda and Hartl, 1984). The transgene can then be artificially altered to identify functional determinants. Experiments are now in progress, aiming to complement rosy mu- tations by transferring Calliphora DNA into Droso-
phila hosts.
ACKNOWLEDGEMENTS
We thank Drs. P. Berreur and J.A. Lepesant for stimulating discussions and for their encouragement,
Drs. D.J. Lipman and W.-H. Li for sending com- puter programs, Dr. M.A. Riley for sending the D. pseudoobscura Xdh sequence before publication and for positive criticism, Mr. Dominique Brandli for expert help in computer programming and Mrs. C. Doumeng for excellent technical assistance. This work was supported by a C.N.R.S. fellowship to M.H. as ‘Chercheur Associt’ and a fellowship of the Ministere de la Recherche to M.C.T.
REFERENCES
Barrett, D. and Davidson, N.A.: Xanthine dehydrogenase accu- mulation in developing Drosophila eyes. J. Insect Physiol. 21
(1975) 1447-1452. Bimstiel, M.L., Busslinger, M. and Strub, K.: Transcription ter-
mination and 3’ processing: the end is in site! Cell 41 (1985) 349-359.
Bucher, P. and Trifanov, E.N.: Compilation and analysis of eukaryotic POL II promoter sequences. Nucleic Acids Res. 14 (1986) 10009-10026.
Dutton, F.L. and Chovnick, A.: Developmental regulation of the rosy locus in Drosophila melanogaster. In L.W. Browder (Ed.), Developmental Biology, Vol. 5. Plenum, New York, 1988, pp. 267-3 16.
Fyrberg, E.A., Kindle, K.L. and Davidson, N.: The actin gene of Drosophila: a dispersed multigene family. Cell 19 (1980) 365-378.
Gamier, J., Osguthorpe, D.J. and Robson, B.: Analysis of the accuracy and implications of simple methods for predicting the secondary structure ofglobular proteins. J. Mol. Biol. 120 (1978) 97-120.
Henikoff, S.: Unidirectional digestion with exonuclease III in DNA sequence analysis. Methods Enzymol. 155 (1987) 156-165.
Hubbard, T.J.P. and Blundell, T.L.: Comparison of solvent- inaccessible cores of homologous proteins: definitions useful for protein modeling. Prot. Engin. 1 (1987) 159-171.
Hyunh, T.V., Young, R.A. and Davis, R.W.: Constructing and screening cDNA libraries in QtlO and lgtll. In Glover, D. (Ed.), DNA Cloning Techniques. A Practical Approach, Vol. I. IRL Press, Oxford, 1985, pp. 49-78.
Keith, T.P., Riley, M.A., Kreitman, M., Lewontin, R.C., Curtis, D. and Chambers, G.: Sequence of the structural gene for xanthine dehydrogenase (rosy locus) in Drosophila melano-
gaster. Genetics 116 (1987) 67-73. Kyte, J. and Doolittle, R.F.: A simple method for displaying the
hydrophobic character of a protein. J. Mol. Biol. 157 (1982) 105-132.
Langer, H.: Properties and functions of screening pigments in insect eyes; Calliphora erythrocephala Meig. In Snyder, A.W. and Menzel, R. (Eds.), Photoreceptor Optics. Springer- Verlag, New York, 1975, pp. 445-455.

402
Lee, C.-S., Curtis, D., McCarron, M., Love, C., Gray, M., Bender, W. and Chovnick, A.: Mutations affecting the expression of the rosy locus in Drosophila melanogaster.
Genetics 116 (1987) 67-73. Li, W.-H., Wu, C.-I. and Luo, C.-C.: A new method for estimating
synonymous and nonsynonymous rates of nucleotide substi- tution considering the relative likelihood of nucleotide and codon changes. Mol. Biol. Evol. 2 (1985) 150-174.
Mount, SM.: A catalogue of splice junction sequences. Nucleic Acids Res. 10 (1982) 459-472.
Peru& M.F.: Species adaptation in a protein molecule. Mol. Biol. Evol. 1 (1983) l-28.
Richardson, J.: The anatomy and taxonomy of protein structure. Adv. Prot. Chem. 34 (1981) 167-339.
Riley, M.A.: Nucleotide sequence of the Xdh region in Drosophila
pseudoobscura and an analysis ofthe evolution of synonymous codons. Mol. Biol. Evol. 6 (1989) 33-52.
Rocher-Chambonnet. C., Berreur, P., Houde, M., Tiveron, M.C., Lepesant, J.A. and Bregegere, F.: Cloning and partial charac- terization of the xanthme dehydrogenase gene of Calliphora
vicina, a distant relative of Drosophila melanogaster. Gene 59 (1987) 201-212.
Rubm, G.M. and Spradling, A.C.: Vectors for P element- mediated gene transfer in Drosophila. Nucleic Acids Res. 11 (1983) 6341-6351.
Sanger, F., Nicklen, S. and Coulson, A.R.: DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74 (1977) 5463-5467.
Scavarda, N.J. and Hart& D.L.: Interspecific DNA transfor- mation in Drosophila. Proc. Natl. Acad. Sci. USA 81 (1984) 2983-2990.
Schenkel, H., Kejzlarova-Lepesant, J., Berreur, P., Moreau, J., Scheller, K., Bregtgdre, F. and Lepesant, J.A.: Identification and molecular analysis of a multigene family encoding calli- phorin, the major larval serum protein of Calliphora vi&a.
EMBO J. 4 (1985) 2993-2990. von Heijne, G.: A new method for predicting signal sequence
cleavage sites. Nucleic Acids Res. 14 (1986) 4683-4690. Yanisch-Perron, C., Vieira, J. and Messing, J.: Improved Ml3
phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 33 (1985) 103-l 19.
Recommended

















