
ORIGINAL ARTICLE
Comparison of different RT-qPCR assays for the detectionof human and bovine group A rotaviruses andcharacterization by sequences analysis of genes encodingVP4 and VP7 capsid proteinsP. Ward1, E. Poitras1, D. Leblanc1, C.A. Gagnon2, J. Brassard1 and A. Houde1
1 Agriculture and Agri-Food Canada, Food Research and Development Centre, Saint-Hyacinthe, QC, Canada
2 Groupe de recherche sur les maladies infectieuses du porc, Facult�e de m�edecine v�et�erinaire, Universit�e de Montr�eal, Saint-Hyacinthe, QC,
Canada
Keywords
capsid protein sequences, detection, group A
rotaviruses, phylogenetic analysis, RT-qPCR
assays.
Correspondence
Pierre Ward, Agriculture and Agri-Food
Canada, Food Research and Development
Centre, 3600 Casavant blvd West,
Saint-Hyacinthe, QC J2S 8E3, Canada.
E-mail: [email protected]
2013/1876: received 18 October 2012,
revised 11 January 2013 and accepted 23
January 2013
doi:10.1111/jam.12165
Abstract
Aims: The aim of this study was to compare the performance of four RT-qPCR
assays for the detection of human and bovine group A rotaviruses and to
characterize the positive samples by sequence analysis of VP4 and VP7 genes.
Methods and Results: RNA extracted from eight human rotavirus strains, and
a panel of 33 human and 25 bovine faecal samples was subjected to different
RT-qPCR detection systems. Among these assays, only RT-qPCR primers and
probe systems B and C were able to detect all human rotavirus strains from
cell culture solutions and faecal samples. However, the results showed that the
system C was generally more sensitive by one or two logs than the other
RT-qPCR assays tested. With the bovine faecal samples, the most efficient
RT-qPCR systems were B and A with the detection in 100 and 92% of samples
tested, respectively. Human group A rotavirus G1P[8] and bovine G6P[11]
were the most frequently used strains identified in this study. A G3P[9] strain,
closely related to a feline rotavirus isolated in the USA, was also discovered in
a human rotavirus infection.
Conclusion: The RT-qPCR system B was the only TaqMan assay evaluated in
this study able to detect rotavirus RNA in all positive human and bovine faecal
samples.
Significance and Impact of the Study: Utilization of only one RT-qPCR for
the detection of human and bovine group A rotaviruses and the possibility of
human infection by a feline rotavirus strain.
Introduction
Rotaviruses were first identified in 1973 as an important
cause of gastroenteritis in children (Bishop et al. 1973).
As is often the case with gastrointestinal diseases, rotavi-
ruses are transmitted through the faecal–oral route. Rota-viruses are the most common agents of diarrhoeal illness
in infants and young children worldwide, and in 2008,
diarrhoea attributable to rotavirus infection resulted in
453 000 deaths worldwide in children younger than five
years. More than half of all fatal rotavirus infections
occurred in five countries: Democratic Republic of the
Congo, Ethiopia, India, Nigeria and Pakistan (Tate et al.
2012). Rotaviruses are also an important cause of neona-
tal bovine diarrhoea (Maes et al. 2003; Swiatek et al.
2010) and are responsible for significant economic losses
(Alfieri et al. 2004).
Rotaviruses are nonenveloped icosahedral viruses
whose genome consists of 11 double-stranded RNA seg-
ments encoding six structural proteins (VP1, VP2, VP3,
VP4, VP6, VP7) and six nonstructural proteins (NSP1-
NSP6). The genome is enclosed in a triple-layered cap-
sid of 70 nm in diameter. The outer layer is composed
of VP4 and VP7 proteins; VP6 protein forms the inner
layer; and VP1, VP2 and VP3 proteins form the core
capsid layer (Min et al. 2006; Matthijnssens et al. 2008;
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1435
Journal of Applied Microbiology ISSN 1364-5072

Kottaridi et al. 2012). Viruses of the genus Rotavirus
belong to the family Reoviridae and are classified into
five defined species (A to E) and two tentative species
(F and G), which are recognized by the International
Committee on Taxonomy of Viruses (ICTV) (Raming
et al. 2005). A potential new rotavirus species,
ADRV-N, has been recently described and was tenta-
tively assigned to species H (Matthijnssens et al. 2012).
The rotavirus species are also commonly referred to as
rotavirus groups. Groups A, B and C have been
detected in human and animal samples, including swine
and bovine (Maes et al. 2003), but groups D, E, F and
G infect only animals (Rahman et al. 2005; Matthijns-
sens et al. 2012). Group A rotaviruses are the most
commonly isolated rotavirus strains. These strains can
be divided into three genogroups (Wa, DS-1 and
AU-1) and have been classified into 27 G types and 35
P types based on the sequence diversity of the genes
encoding the two outer capsid proteins, VP4 and VP7,
respectively (Matthijnssens et al. 2011). In human
group A rotavirus, G1P[8], G2P[4] and G4P[8] are
generally the most prevalent genotypes (Gentsch et al.
2005; van der Heide et al. 2005; Ahmed et al. 2006).
Among bovine rotavirus strains, the most prevalent
genotypes worldwide are G10P[11], G10P[5] and G10P
[1] (Steyer et al. 2010). Group C rotavirus has been
associated with sporadic diarrhoeal illness in different
parts of the world and could be an emerging pathogen
in humans (Abid et al. 2007).
Group A rotavirus is widespread in wild and domestic
animal species, and it has been suggested that zoonotic
transmission plays a substantial role in the introduction of
novel strains into the human population (Banyai et al.
2009). Evidence for zoonotic transmission of bovine rota-
virus strains to humans and genetic reassortment between
human and animal rotaviruses has been described in the
literature (Khamrin et al. 2006; Martella et al. 2010; Steyer
et al. 2010). Recently, a new complete genome classifica-
tion system was developed for group A rotavirus strains.
This nucleotide sequence-based system assigns a specific
genotype to each of the 11 genome segments and has
increased the recognition of homology between animal and
human rotavirus strains (Matthijnssens et al. 2011).
It is known that raw food, treated water, untreated
water and irrigation water can represent possible sources
of contamination by rotaviruses. These viruses are very
stable in the environment, can be spread by faecal mate-
rial from sick people or animals and may remain infec-
tious for many weeks (Brassard et al. 2005; Leung et al.
2005). Just a few viral particles appear to be sufficient to
trigger infection in humans. To put this into perspective,
there can be as many as 108 to 1011 particles ml�1 of
stool in infected patients (Koopmans and Duizer 2004).
Transmission electron microscopy (TEM) and antigen
detection kits, such as enzyme immunoassay and latex
agglutination, are the most frequently used methods for
the detection of rotavirus. However, these methods are
generally less sensitive than molecular techniques such as
RT-PCR or RT-qPCR. Approximately 106 viral particles
ml�1 of sample are required for detection by TEM
(Logan et al. 2006; Jothikumar et al. 2009). In the past
ten years, many RT-qPCR tests using SYBR Green or
hydrolysis probes targeting the NSP3, VP2, VP4, VP6 or
VP7 genes have been developed for the detection of rota-
virus RNA (Schwarz et al. 2002; Kang et al. 2004; Pang
et al. 2004, 2011; Logan et al. 2006; Min et al. 2006; Free-
man et al. 2008; Guti�errez-Aguirre et al. 2008; Zeng et al.
2008; Jothikumar et al. 2009; Plante et al. 2011; Kottaridi
et al. 2012). The sensitivity of these detection assays can
be affected by the quality of the extracted RNA, by RNase
contamination and by RT-PCR inhibitors in environmen-
tal and clinical samples, especially in faecal material
(Escobar-Herrera et al. 2006; Rutjes et al. 2007; Scipioni
et al. 2008), as well as by the genomic variability of rota-
virus strains in the target regions used in the detection
tests. Failure to amplify the viral RNA owing to these fac-
tors may result in false-negative results. The use of a
sample process control artificially added to the samples
prior to concentration of the viral particles and RNA
extraction can be extremely useful for monitoring the
quality of the extraction procedure and for identifying
the potential presence of RT-PCR inhibitors that interfere
with the amplification reactions (Jones et al. 2009; Matti-
son et al. 2009; Ward et al. 2009). In Canada, Health
Canada’s Technical Group on Virology has recommended
the use of feline calicivirus (FCV) as a sample process
control for detection methods (Houde et al. 2009).
Due to the low infectious dose, the zoonotic issues and
the possibility of recovering rotavirus not only from clini-
cal samples but also from environmental and food sam-
ples, it is important to have a sensitive detection method
that can detect both human and animal strains. The aim
of this study was to compare the performance of four
previously published RT-qPCR assays, targeting the VP6,
VP7 or NSP3 gene, for the detection of human and
bovine group A rotavirus strains and to characterize the
positive samples by sequence analysis of genes encoding
VP4 and VP7 outer capsid proteins.
Materials and methods
Strains and faecal samples collected between 2005 and
2009
Eight human rotavirus strains from the American Type
Culture Collection (ATCC) were used in this study [WA
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1436 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.

(ATCC VR-2018)/G1, 1-9-12/77/S (ATCC VR-1546)/G2,
89-12C2 (ATCC VR-2272)/G3, 408 (ATCC VR-2273)/G1,
248 (ATCC VR-2274)/G4, WISC2 (ATCC VR-2417)/G1,
DS-1 (ATCC VR-2550)/G2, WI61 (ATCC VR-2551)/G9].
Viral strains were propagated to produce stock suspen-
sions, as recommended by ATCC. A panel of 65 faecal
specimens from Facult�e de m�edecine v�et�erinaire, Univer-
sit�e de Montr�eal, Saint-Hyacinthe (Canada), Chinook
Regional Hospital, Lethbridge (Canada) and the Hospital
for Sick Children, Toronto (Canada) was also used. This
panel included 13 rotavirus-positive faecal samples col-
lected from children, 10 human faecal samples from
patients of different age groups with fulminant gastroen-
teritis and negative for rotavirus, 10 human faecal samples
from healthy individuals and negative for rotavirus, 13
bovine faecal samples positive for rotavirus and 12 bovine
faecal samples negative for rotavirus. Positive samples
have been confirmed by transmission electron microscopy
or by RT-PCR. In addition, the assays were tested for any
cross-reactivity that may have occurred using a wide range
of viral and bacterial pathogens present in the stool. These
viral pathogens included strains of norovirus GI and GII,
swine hepatitis E virus, hepatitis A virus and adenovirus
40/41. Bacterial DNA from Escherichia coli O157:H7,
Campylobacter jejuni LSPQ 3234, Salmonella thyphimur-
ium ATCC 14028, Staphylococcus aureus ATCC 25923 and
Listeria monocytogenes ATCC 7644 were kindly provided
by Evelyne Gu�evremont (Agriculture and Agri-Food
Canada, Food Research and Development Center, Saint-
Hyacinthe, Canada).
Viral RNA extraction
Faecal samples were diluted 1 : 5 (w/v) in sterile PBS, pH
7�2 (Life Technologies Inc., Burlington, ON, CA) before
centrifugation for 20 min at 4000 g. The clarified stool
suspensions or the stock solutions of each rotavirus strain
were adjusted to reach 1% sodium dodecyl sulfate
(Sigma-Aldrich, Oakville ON, Canada) and 100 lg ml�1
of Proteinase K (QIAGEN, Mississauga, ON, Canada).
Mixtures were incubated at 37°C for 1 h. To monitor the
RNA extraction process, 3�2 9 103 PFU of feline calicivi-
rus (FCV) were added to 140 ll of the resulting suspen-
sions as sample process control (Ward et al. 2009). Viral
RNA was extracted with QIAamp� Viral RNA mini
(QIAGEN) protocols adapted for the QIAcube robotic
workstation (QIAGEN) using QIAamp� Viral RNA body
fluid: manual lysis protocol. To protect the extracted
RNA from exogenous RNases, RNase inhibitor (RNase-
Out, Life Technologies Inc.) was added to the final QIA-
GEN AVE elution buffer. Recovered RNA was frozen at
�80°C until further use.
Primers and probes
All primers (IDT, Coralville, IA, USA) and hydrolysis
probes (IDT and Life Technologies Inc. for MGB probes)
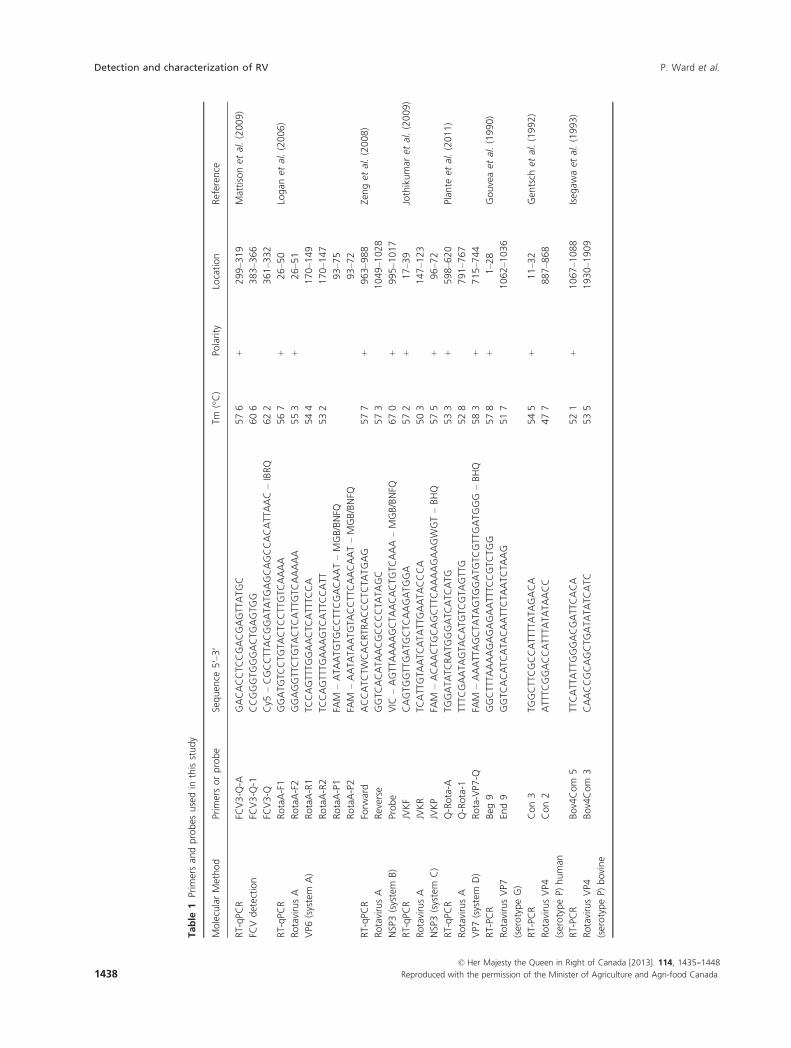
used in this study are listed in Table 1.
Conventional RT-PCR
The full-length VP7 segment from human and bovine
samples was amplified with Beg-9 and End-9 primers,
according to the parameters described by Gouvea et al.
(1990). The partial VP4 segment from human samples
was amplified with Con 2 and Con 3 primers with
cycling conditions described by Gentsch et al. (1992).
Primers used for bovine samples were Bov4Com 5 and
Bov4Com 3 with conditions described by Isegawa et al.
(1993). RT-PCR was performed using a QIAGEN� One-
Step RT-PCR Kit (QIAGEN), and the amplified products
were separated on a 1% agarose gel with amplicons
visualized with ethidium bromide staining.
Construction of plasmid DNA standards for RT-qPCR
reactions
Conventional RT-PCR were carried out in a total volume of
20 ll using the QIAGEN�OneStep RT-PCR kit (QIAGEN),
according to the manufacturer’s recommendations in an
Eppendorf Mastercycler gradient system (Brinkman Instru-
ments Canada Ltd., Mississauga, ON, Canada). Amplifica-
tions were performed using group A rotavirus strain WA
(ATCC VR-2018), and the different primer sets described in
Table 1. RT-PCR fragments of 145, 87, 131 and 194 bp cor-
responding to TaqMan amplification primer system A
(Logan et al. 2006), B (Zeng et al. 2008), C (Jothikumar
et al. 2009) and D (Plante et al. 2011), respectively, were
excised from the gel and purified using the QIAquick� Gel
Extraction kit (QIAGEN). PCR products were cloned into
pCR� 2�1 TOPO� vector using TOPO TA Cloning� kit
(Life Technologies Inc.) with One Shot� TOP10 electro-
competent cells in accordance with the manufacturer’s
recommendations. The recombinant plasmid stocks
were quantified using the NanoDrop spectrophotometer
ND–1000 according to the manufacturer’s instructions
(NanoDrop Technologies Inc., Wilmington, DE, USA) and
converted into copy number. The copy number of plasmid
was calculated as: copy number = [(concentration of linear-
ized plasmid)/(molar mass)] 9 (6�023 9 1023). These
DNA plasmids were used for the generation of standard
curves and as positive controls.
RT-qPCR assays
The RT-qPCR assays were carried out in 25 ll of a reactionmixture comprising 2�5 ll of extracted RNA and 22�5 ll of
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1437
P. Ward et al. Detection and characterization of RV
Table
1Prim
ersan
dprobes
usedin
thisstudy
MolecularMethod
Prim
ersorprobe
Sequen
ce5′–3′
Tm(°C)
Polarity
Location
Referen
ce
RT-qPC
R
FCVdetection
FCV3-Q
-AGACACCTC
CGACGAGTTATGC
57�6
+299–3
19
Mattisonet
al.(2009)
FCV3-Q
-1CCGGGTG
GGACTG
AGTG
G60�6
�383–3
66
FCV3-Q
Cy5
–CGCCTTACGGATA
TGAGCAGCCACATTAAC–IBRQ
62�2
�361–3
32
RT-qPC
R
RotavirusA
VP6
(system
A)
RotaA-F1
GGATG
TCCTG
TACTC
CTTGTC
AAAA
56�7
+26–5
0Logan
etal.(2006)
RotaA-F2
GGAGGTTCTG
TACTC
ATTGTC
AAAAA
55�3
+26–5
1
RotaA-R1
TCCAGTTTG
GAACTC
ATTTC
CA
54�4
�170–1
49
RotaA-R2
TCCAGTTTG
AAAGTC
ATTCCATT
53�2
�170–1
47
RotaA-P1
FAM
–ATA
ATG
TGCCTTCGACAAT–MGB/BNFQ
�93–7
5
RotaA-P2
FAM
–AATA
TAATG
TACCTTCAACAAT–MGB/BNFQ
�93–7
2
RT-qPC
R
RotavirusA
NSP3(system
B)
Forw
ard
ACCATC
TWCACRTR
ACCCTC
TATGAG
57�7
+963–9
88
Zenget
al.(2008)
Reverse
GGTC
ACATA
ACGCCCCTA
TAGC
57�3
�1049–1
028
Probe
VIC
–AGTTAAAAGCTA
ACACTG
TCAAA
–MGB/BNFQ
67�0
+995–1
017
RT-qPC
R
RotavirusA
NSP3(system
C)
JVKF
CAGTG
GTTGATG
CTC
AAGATG
GA
57�2
+17–3
9Jothikumar
etal.(2009)
JVKR
TCATTGTA
ATC
ATA
TTGAATA
CCCA
50�3
�147–1
23
JVKP
FAM
–ACAACTG
CAGCTTCAAAAGAAGWGT–BHQ
57�5
+96–7
2
RT-qPC
R
RotavirusA
VP7
(system
D)
Q-Rota-A
TGGATA
TCRATG
GGATC
ATC
ATG
53�3
+598–6
20
Plan
teet
al.(2011)
Q-Rota-1
TTTC
GAATA
GTA
CATG
TCGTAGTTG
52�8
�791–7
67
Rota-VP7
-QFA
M–AAATTAGCTA
TAGTG
GATG
TCGTTGATG
GG
–BHQ
58�3
+715–7
44
RT-PC
R
RotavirusVP7
(serotypeG)
Beg
9GGCTTTA
AAAGAGAGAATTTC
CGTC
TGG
57�8
+1–2
8Gouveaet
al.(1990)
End9
GGTC
ACATC
ATA
CAATTCTA
ATCTA
AG
51�7
�1062–1
036
RT-PC
R
RotavirusVP4
(serotypeP)
human
Con3
TGGCTTCGCCATTTTATA
GACA
54�5
+11–3
2Gen
tsch
etal.(1992)
Con2
ATTTC
GGACCATTTA
TATA
ACC
47�7
�887–8
68
RT-PC
R
RotavirusVP4
(serotypeP)
bovine
Bov4Com
5TTCATTATTGGGACGATTCACA
52�1
+1067–1
088
Iseg
awaet
al.(1993)
Bov4Com
3CAACCGCAGCTG
ATA
TATC
ATC
53�5
�1930–1
909
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1438 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.

master mix. Master mix was prepared using the OneStep
Brilliant II QRT-PCR core reagent kit (Agilent Technologies
Canada, Mississauga, ON, Canada) and contained
5�0 mmol l�1 of MgCl2, 600 nmol l�1 of both forward and
reverse primers and 250 nmol l�1 of hydrolysis probe for
system A (Logan et al. 2006); or 400 nmol l�1 of both for-
ward and reverse primers and 200 nmol l�1 of hydrolysis
probe for system B (Zeng et al. 2008); or 250 nmol l�1 of
forward and reverse primers and 100 nmol l�1 of hydrolysis
probe for system C (Jothikumar et al. 2009); or
300 nmol l�1 of forward and reverse primers and
200 nmol l�1 of hydrolysis probe for system D (Plante et al.
2011). For the FCV assay, 5�0 mmol l�1 of MgCl2,
300 nmol l�1 of forward and reverse primers and
200 nmol l�1 of hydrolysis probe were included in the mas-
ter mix (Mattison et al. 2009). RT-PCR amplifications were
run in a Stratagene Mx3005P system (Agilent Technologies
Canada) in a 96-well format under the following conditions:
30 min at 50°C for reverse transcription, 95°C for 10 min
for initial denaturation then followed by 45 cycles of ampli-
fication with denaturation at 95°C for 15 s and annealing
and extension at 60°C for 1 min. A standard curve for each
system was generated using 10-fold serial dilution (108 to
100 genomic equivalents) in a 5 ng ml�1 salmon sperm
DNA solution of appropriate purified DNA plasmid.
Cloning and sequencing of RT-PCR product
RT-PCR amplicons for VP4 and VP7 segments were excised
from the gel and purified using the QIAquick Gel Extraction
kit (Qiagen). Purified PCR products were cloned into pCR
2�1 TOPO vector using TOPO TA Cloning kit (Life
Technologies Inc.) with TOP10 electrocompetent cells in
accordance with the manufacturer’s recommendations.
Sequencing was performed on recombinant plasmids in
both directions using a CEQTM 8000 Genetic Analysis System
(Beckman Coulter, Fullerton, CA, USA) and a CEQ Dye
Terminator Cycle sequencing kit (Beckman Coulter) with
M13 forward and reverse primers. Nucleotide alignment
was undertaken with the CLUSTAL W (http://www.ebi.ac.
uk/clustalw) program. The phylogenetic tree was created by
the neighbour-joining method using CLC sequence viewer 6
software (http://www.clcbio.com). Bootstrap analysis was
employed to determine the statistical confidence of the phy-
logenetic relationships. All sequences were deposited in
GenBank under accession numbers JX470485 – JX470523.
Results
Efficiency evaluation of different RT-qPCR assays
A standard curve was established for each RT-qPCR
system using the corresponding cloned amplicon, which
was serially diluted from 1 9 108 to 1 9 100 copies and
amplified in triplicate. The quantification cycle number
values (Cq) were plotted against genomic equivalent cop-
ies. The standard curves obtained showed an efficiency of
99�1%, a regression coefficient of 0�993, a slope of
�3�344 and an intercept of 39�01 for system A; an effi-
ciency of 98�3%, a regression coefficient of 0�999, a slope
of �3�363 and an intercept of 39�79 for system B; an effi-
ciency of 99�1%, a regression coefficient of 0�996, a slope
of �3�343 and an intercept of 39�32 for system C; and an
efficiency of 100�2%, a regression coefficient of 0�996, aslope of �3�318 and an intercept of 40�99 for system D
(data not shown). These standard curves indicated that
the four assays could detect 2�5 copies per reaction.
Detection of rotavirus RNA by conventional RT-PCR
and RT-qPCR assays
All extracted RNA samples were first individually tested
for FCV. The 3�2 9 103 PFU of FCV added to the clari-
fied stool suspensions as a sample process control before
RNA extraction were detected in all samples with a mean
Cq of 25�75. These results were correlated with the Cq of
26�16 obtained for the extraction control with FCV alone
(data not shown). These results showed the efficiency of
the sample genome extraction process and showed that
the RT-PCR reactions were not affected by inhibitors.
The different rotavirus molecular detection assays were
evaluated and compared in parallel using the same RNA
extracts. Each molecular assay included a negative control
(RNAse-free water) and a positive control (cloned ampli-
con). All RT-PCR products obtained on ethidium bro-
mide-stained agarose gel were of the correct size, and no
bands were visible in the negative controls (data not
shown). The detection results obtained with the four dif-
ferent RT-qPCR assays performed on human and animal
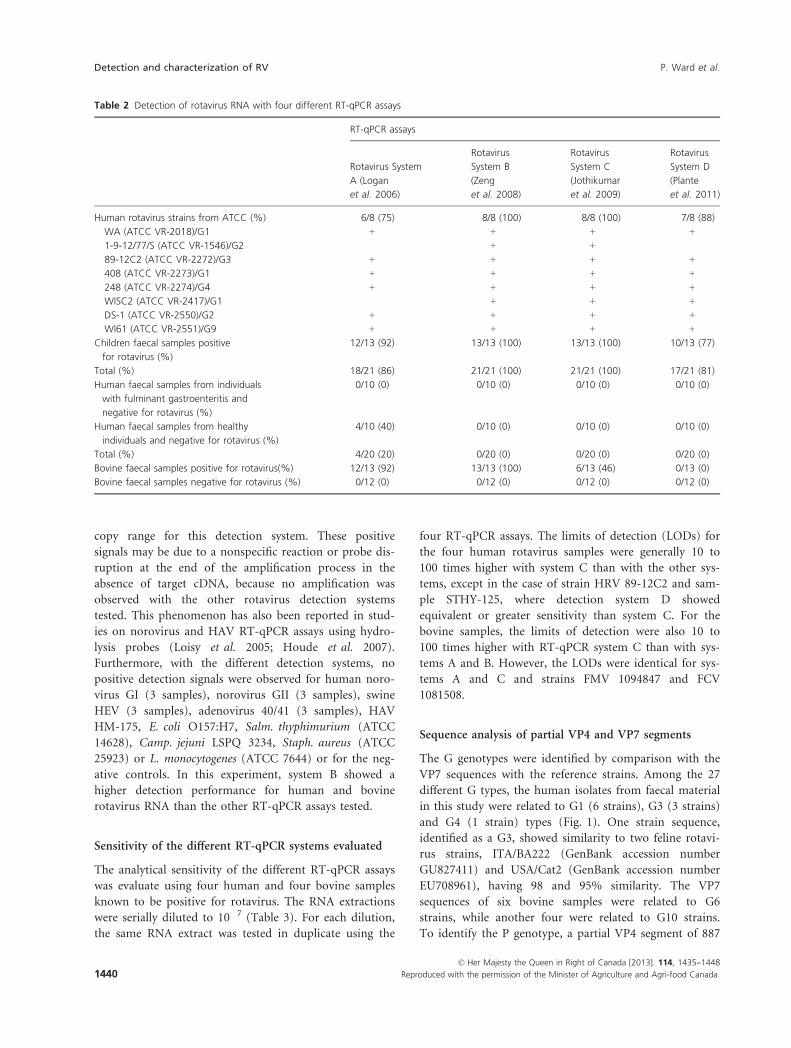
rotavirus strains are presented in Table 2. RT-qPCR sys-
tems B and C detected rotavirus RNA in 100% (21/21) of
the human rotavirus–positive samples, including eight
strains from the American Type Culture Collection
(ATCC) with G serotype, which are frequently detected
in clinical samples. Systems A and D detected rotavirus
RNA in 86% (18/21) and 81% (17/21) of the samples,
respectively. With the bovine rotavirus–positive samples,
system B was the best detection system, achieving 100%
detection (13/13) compared with 92% (12/13), 46% (6/
13) and 0% (0/13) for systems A, C and D, respectively.
All negative samples were found to be negative with the
different detection assays, except with system A, where a
positive amplification result was observed in 20% of
human rotavirus–negative samples. The positive fluores-
cence signal observed for these negative samples was in
the 35–39 Cq range, which corresponds to the single-
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1439
P. Ward et al. Detection and characterization of RV
copy range for this detection system. These positive
signals may be due to a nonspecific reaction or probe dis-
ruption at the end of the amplification process in the
absence of target cDNA, because no amplification was
observed with the other rotavirus detection systems
tested. This phenomenon has also been reported in stud-
ies on norovirus and HAV RT-qPCR assays using hydro-
lysis probes (Loisy et al. 2005; Houde et al. 2007).
Furthermore, with the different detection systems, no
positive detection signals were observed for human noro-
virus GI (3 samples), norovirus GII (3 samples), swine
HEV (3 samples), adenovirus 40/41 (3 samples), HAV
HM-175, E. coli O157:H7, Salm. thyphimurium (ATCC
14628), Camp. jejuni LSPQ 3234, Staph. aureus (ATCC
25923) or L. monocytogenes (ATCC 7644) or for the neg-
ative controls. In this experiment, system B showed a
higher detection performance for human and bovine
rotavirus RNA than the other RT-qPCR assays tested.
Sensitivity of the different RT-qPCR systems evaluated
The analytical sensitivity of the different RT-qPCR assays
was evaluate using four human and four bovine samples
known to be positive for rotavirus. The RNA extractions
were serially diluted to 10�7 (Table 3). For each dilution,
the same RNA extract was tested in duplicate using the
four RT-qPCR assays. The limits of detection (LODs) for
the four human rotavirus samples were generally 10 to
100 times higher with system C than with the other sys-
tems, except in the case of strain HRV 89-12C2 and sam-
ple STHY-125, where detection system D showed
equivalent or greater sensitivity than system C. For the
bovine samples, the limits of detection were also 10 to
100 times higher with RT-qPCR system C than with sys-
tems A and B. However, the LODs were identical for sys-
tems A and C and strains FMV 1094847 and FCV
1081508.
Sequence analysis of partial VP4 and VP7 segments
The G genotypes were identified by comparison with the
VP7 sequences with the reference strains. Among the 27
different G types, the human isolates from faecal material
in this study were related to G1 (6 strains), G3 (3 strains)
and G4 (1 strain) types (Fig. 1). One strain sequence,
identified as a G3, showed similarity to two feline rotavi-
rus strains, ITA/BA222 (GenBank accession number
GU827411) and USA/Cat2 (GenBank accession number
EU708961), having 98 and 95% similarity. The VP7
sequences of six bovine samples were related to G6
strains, while another four were related to G10 strains.
To identify the P genotype, a partial VP4 segment of 887
Table 2 Detection of rotavirus RNA with four different RT-qPCR assays
RT-qPCR assays
Rotavirus System
A (Logan
et al. 2006)
Rotavirus
System B
(Zeng
et al. 2008)
Rotavirus
System C
(Jothikumar
et al. 2009)
Rotavirus
System D
(Plante
et al. 2011)
Human rotavirus strains from ATCC (%) 6/8 (75) 8/8 (100) 8/8 (100) 7/8 (88)
WA (ATCC VR-2018)/G1 + + + +
1-9-12/77/S (ATCC VR-1546)/G2 � + + �89-12C2 (ATCC VR-2272)/G3 + + + +
408 (ATCC VR-2273)/G1 + + + +
248 (ATCC VR-2274)/G4 + + + +
WISC2 (ATCC VR-2417)/G1 � + + +
DS-1 (ATCC VR-2550)/G2 + + + +
WI61 (ATCC VR-2551)/G9 + + + +
Children faecal samples positive
for rotavirus (%)
12/13 (92) 13/13 (100) 13/13 (100) 10/13 (77)
Total (%) 18/21 (86) 21/21 (100) 21/21 (100) 17/21 (81)
Human faecal samples from individuals
with fulminant gastroenteritis and
negative for rotavirus (%)
0/10 (0) 0/10 (0) 0/10 (0) 0/10 (0)
Human faecal samples from healthy
individuals and negative for rotavirus (%)
4/10 (40) 0/10 (0) 0/10 (0) 0/10 (0)
Total (%) 4/20 (20) 0/20 (0) 0/20 (0) 0/20 (0)
Bovine faecal samples positive for rotavirus(%) 12/13 (92) 13/13 (100) 6/13 (46) 0/13 (0)
Bovine faecal samples negative for rotavirus (%) 0/12 (0) 0/12 (0) 0/12 (0) 0/12 (0)
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1440 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.
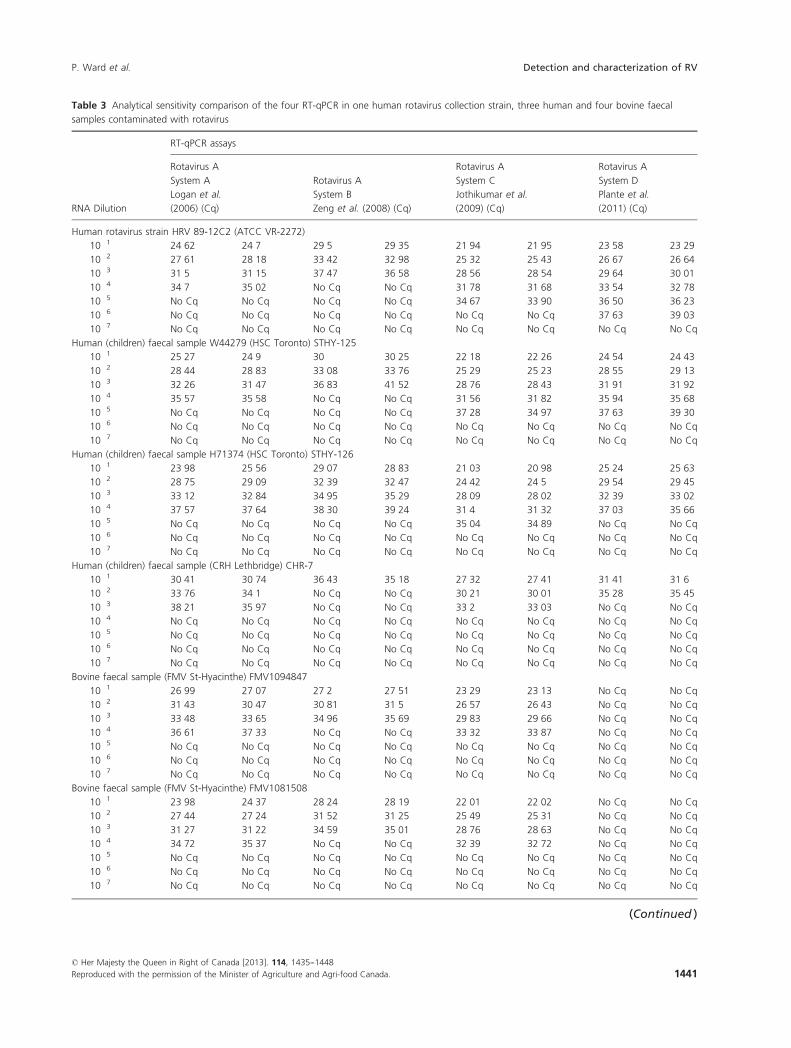
Table 3 Analytical sensitivity comparison of the four RT-qPCR in one human rotavirus collection strain, three human and four bovine faecal
samples contaminated with rotavirus
RNA Dilution
RT-qPCR assays
Rotavirus A
System A
Logan et al.
(2006) (Cq)
Rotavirus A
System B
Zeng et al. (2008) (Cq)
Rotavirus A
System C
Jothikumar et al.
(2009) (Cq)
Rotavirus A
System D
Plante et al.
(2011) (Cq)
Human rotavirus strain HRV 89-12C2 (ATCC VR-2272)
10�1 24�62 24�7 29�5 29�35 21�94 21�95 23�58 23�2910�2 27�61 28�18 33�42 32�98 25�32 25�43 26�67 26�6410�3 31�5 31�15 37�47 36�58 28�56 28�54 29�64 30�0110�4 34�7 35�02 No Cq No Cq 31�78 31�68 33�54 32�7810�5 No Cq No Cq No Cq No Cq 34�67 33�90 36�50 36�2310�6 No Cq No Cq No Cq No Cq No Cq No Cq 37�63 39�0310�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
Human (children) faecal sample W44279 (HSC Toronto) STHY-125
10�1 25�27 24�9 30 30�25 22�18 22�26 24�54 24�4310�2 28�44 28�83 33�08 33�76 25�29 25�23 28�55 29�1310�3 32�26 31�47 36�83 41�52 28�76 28�43 31�91 31�9210�4 35�57 35�58 No Cq No Cq 31�56 31�82 35�94 35�6810�5 No Cq No Cq No Cq No Cq 37�28 34�97 37�63 39�3010�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
Human (children) faecal sample H71374 (HSC Toronto) STHY-126
10�1 23�98 25�56 29�07 28�83 21�03 20�98 25�24 25�6310�2 28�75 29�09 32�39 32�47 24�42 24�5 29�54 29�4510�3 33�12 32�84 34�95 35�29 28�09 28�02 32�39 33�0210�4 37�57 37�64 38�30 39�24 31�4 31�32 37�03 35�6610�5 No Cq No Cq No Cq No Cq 35�04 34�89 No Cq No Cq
10�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
Human (children) faecal sample (CRH Lethbridge) CHR-7
10�1 30�41 30�74 36�43 35�18 27�32 27�41 31�41 31�610�2 33�76 34�1 No Cq No Cq 30�21 30�01 35�28 35�4510�3 38�21 35�97 No Cq No Cq 33�2 33�03 No Cq No Cq
10�4 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�5 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
Bovine faecal sample (FMV St-Hyacinthe) FMV1094847
10�1 26�99 27�07 27�2 27�51 23�29 23�13 No Cq No Cq
10�2 31�43 30�47 30�81 31�5 26�57 26�43 No Cq No Cq
10�3 33�48 33�65 34�96 35�69 29�83 29�66 No Cq No Cq
10�4 36�61 37�33 No Cq No Cq 33�32 33�87 No Cq No Cq
10�5 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
Bovine faecal sample (FMV St-Hyacinthe) FMV1081508
10�1 23�98 24�37 28�24 28�19 22�01 22�02 No Cq No Cq
10�2 27�44 27�24 31�52 31�25 25�49 25�31 No Cq No Cq
10�3 31�27 31�22 34�59 35�01 28�76 28�63 No Cq No Cq
10�4 34�72 35�37 No Cq No Cq 32�39 32�72 No Cq No Cq
10�5 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
(Continued )
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1441
P. Ward et al. Detection and characterization of RV
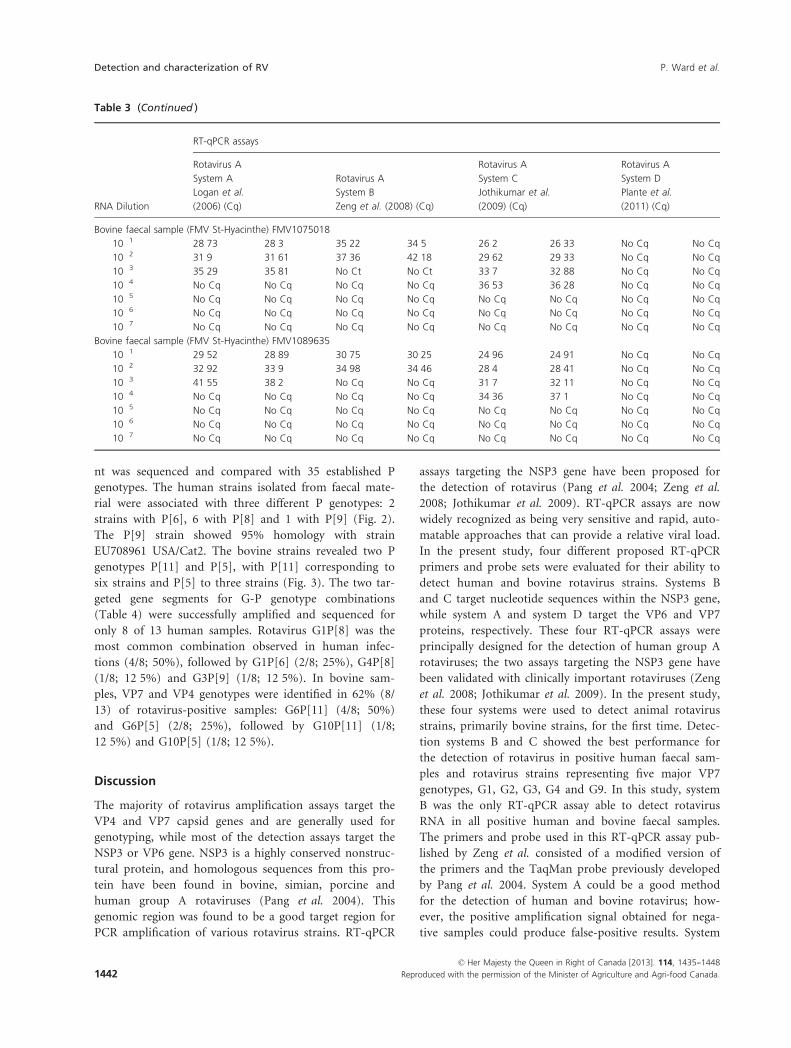
nt was sequenced and compared with 35 established P
genotypes. The human strains isolated from faecal mate-
rial were associated with three different P genotypes: 2
strains with P[6], 6 with P[8] and 1 with P[9] (Fig. 2).
The P[9] strain showed 95% homology with strain
EU708961 USA/Cat2. The bovine strains revealed two P
genotypes P[11] and P[5], with P[11] corresponding to
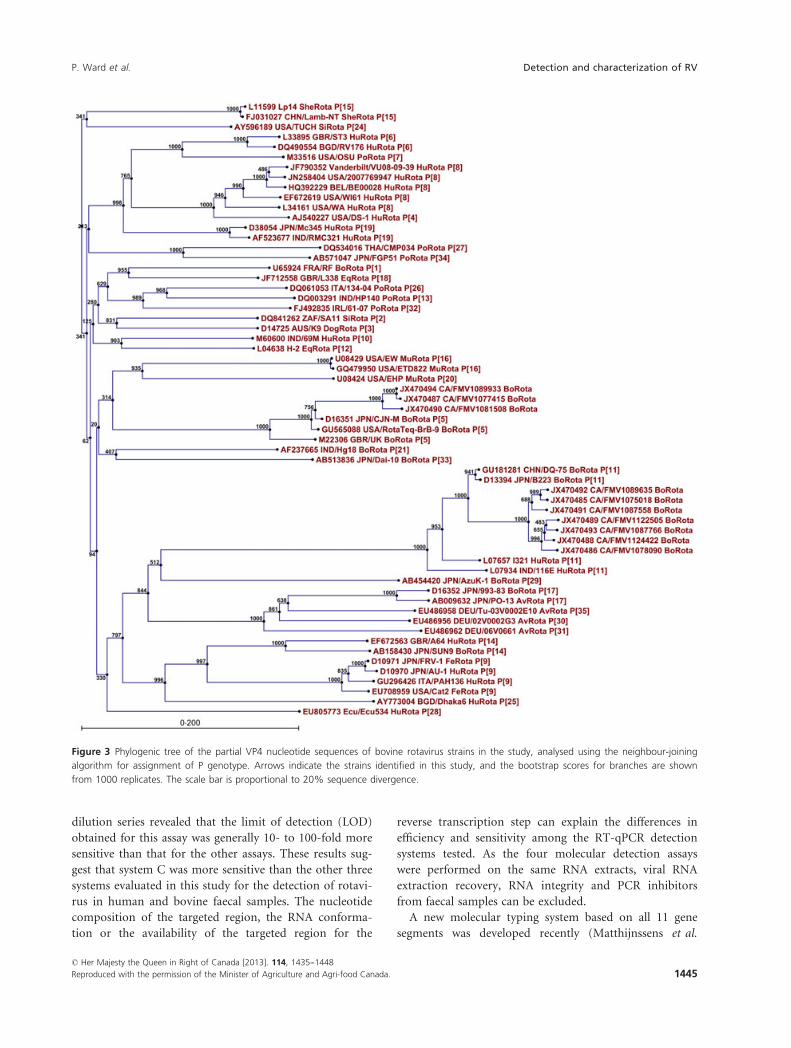
six strains and P[5] to three strains (Fig. 3). The two tar-
geted gene segments for G-P genotype combinations
(Table 4) were successfully amplified and sequenced for
only 8 of 13 human samples. Rotavirus G1P[8] was the
most common combination observed in human infec-
tions (4/8; 50%), followed by G1P[6] (2/8; 25%), G4P[8]
(1/8; 12�5%) and G3P[9] (1/8; 12�5%). In bovine sam-
ples, VP7 and VP4 genotypes were identified in 62% (8/
13) of rotavirus-positive samples: G6P[11] (4/8; 50%)
and G6P[5] (2/8; 25%), followed by G10P[11] (1/8;
12�5%) and G10P[5] (1/8; 12�5%).
Discussion
The majority of rotavirus amplification assays target the
VP4 and VP7 capsid genes and are generally used for
genotyping, while most of the detection assays target the
NSP3 or VP6 gene. NSP3 is a highly conserved nonstruc-
tural protein, and homologous sequences from this pro-
tein have been found in bovine, simian, porcine and
human group A rotaviruses (Pang et al. 2004). This
genomic region was found to be a good target region for
PCR amplification of various rotavirus strains. RT-qPCR
assays targeting the NSP3 gene have been proposed for
the detection of rotavirus (Pang et al. 2004; Zeng et al.
2008; Jothikumar et al. 2009). RT-qPCR assays are now
widely recognized as being very sensitive and rapid, auto-
matable approaches that can provide a relative viral load.
In the present study, four different proposed RT-qPCR
primers and probe sets were evaluated for their ability to
detect human and bovine rotavirus strains. Systems B
and C target nucleotide sequences within the NSP3 gene,
while system A and system D target the VP6 and VP7
proteins, respectively. These four RT-qPCR assays were
principally designed for the detection of human group A
rotaviruses; the two assays targeting the NSP3 gene have
been validated with clinically important rotaviruses (Zeng
et al. 2008; Jothikumar et al. 2009). In the present study,
these four systems were used to detect animal rotavirus
strains, primarily bovine strains, for the first time. Detec-
tion systems B and C showed the best performance for
the detection of rotavirus in positive human faecal sam-
ples and rotavirus strains representing five major VP7
genotypes, G1, G2, G3, G4 and G9. In this study, system
B was the only RT-qPCR assay able to detect rotavirus
RNA in all positive human and bovine faecal samples.
The primers and probe used in this RT-qPCR assay pub-
lished by Zeng et al. consisted of a modified version of
the primers and the TaqMan probe previously developed
by Pang et al. 2004. System A could be a good method
for the detection of human and bovine rotavirus; how-
ever, the positive amplification signal obtained for nega-
tive samples could produce false-positive results. System
Table 3 (Continued )
RNA Dilution
RT-qPCR assays
Rotavirus A
System A
Logan et al.
(2006) (Cq)
Rotavirus A
System B
Zeng et al. (2008) (Cq)
Rotavirus A
System C
Jothikumar et al.
(2009) (Cq)
Rotavirus A
System D
Plante et al.
(2011) (Cq)
Bovine faecal sample (FMV St-Hyacinthe) FMV1075018
10�1 28�73 28�3 35�22 34�5 26�2 26�33 No Cq No Cq
10�2 31�9 31�61 37�36 42�18 29�62 29�33 No Cq No Cq
10�3 35�29 35�81 No Ct No Ct 33�7 32�88 No Cq No Cq
10�4 No Cq No Cq No Cq No Cq 36�53 36�28 No Cq No Cq
10�5 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
Bovine faecal sample (FMV St-Hyacinthe) FMV1089635
10�1 29�52 28�89 30�75 30�25 24�96 24�91 No Cq No Cq
10�2 32�92 33�9 34�98 34�46 28�4 28�41 No Cq No Cq
10�3 41�55 38�2 No Cq No Cq 31�7 32�11 No Cq No Cq
10�4 No Cq No Cq No Cq No Cq 34�36 37�1 No Cq No Cq
10�5 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�6 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
10�7 No Cq No Cq No Cq No Cq No Cq No Cq No Cq No Cq
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1442 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.

D, targeting the VP7 protein and previously tested only
with the human rotavirus strain WA (ATCC CRL-2018)
(Plante et al. 2011), was the least effective system, detect-
ing only 81% of human rotavirus–positive samples, and
none of the bovine rotavirus RNA material (0/13)
included in this study. These results confirm that the
Figure 1 Phylogenic tree of the VP7 nucleotide sequences of rotavirus strains in the study, analysed using the neighbour-joining algorithm for
assignment of G genotype. Arrows indicate the strains identified in this study, and the bootstrap scores for branches are shown from 1000
replicates. The scale bar is proportional to 22% sequence divergence.
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1443
P. Ward et al. Detection and characterization of RV

VP7 capsid gene may not be a good target for the
detection of a large diversity of rotavirus strains.
Serially diluted plasmid DNA standard was previously
used to validate the primers and probes for amplification
systems A, B and C and to estimate the limits of detection
(Logan et al. 2006; Zeng et al. 2008; Jothikumar et al.
2009). The resulting limits of detection, ranging from 3 to
10 genome equivalent copies, are consistent with the
results obtained in the present study. The sensitivity of
system C, developed by Jothikumar et al. 2009, was esti-
mated to be between 2 and 4 viral particles. In the present
study, system C showed Cq values that were consistently
lower than those for the other detection systems, for every
sample. In addition, the analysis of rotavirus RNA
Figure 2 Phylogenic tree of the partial VP4 nucleotide sequences of human rotavirus strains in the study, analysed using the neighbour-joining
algorithm for assignment of P genotype. Arrows indicate the strains identified in this study, and the bootstrap scores for branches are shown
from 1000 replicates. The scale bar is proportional to 25% sequence divergence.
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1444 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.
dilution series revealed that the limit of detection (LOD)
obtained for this assay was generally 10- to 100-fold more
sensitive than that for the other assays. These results sug-
gest that system C was more sensitive than the other three
systems evaluated in this study for the detection of rotavi-
rus in human and bovine faecal samples. The nucleotide
composition of the targeted region, the RNA conforma-
tion or the availability of the targeted region for the
reverse transcription step can explain the differences in
efficiency and sensitivity among the RT-qPCR detection
systems tested. As the four molecular detection assays
were performed on the same RNA extracts, viral RNA
extraction recovery, RNA integrity and PCR inhibitors
from faecal samples can be excluded.
A new molecular typing system based on all 11 gene
segments was developed recently (Matthijnssens et al.
Figure 3 Phylogenic tree of the partial VP4 nucleotide sequences of bovine rotavirus strains in the study, analysed using the neighbour-joining
algorithm for assignment of P genotype. Arrows indicate the strains identified in this study, and the bootstrap scores for branches are shown
from 1000 replicates. The scale bar is proportional to 20% sequence divergence.
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1445
P. Ward et al. Detection and characterization of RV

2008). This system is based on sequencing and phyloge-
netic analysis of all fragments and uses the notations
Gx-P[x]-Ix-Rx-Cx-Mx-Ax-Nx-Tx-Ex-Hx for the VP7-
VP4-VP6-VP1-VP2-VP3-NSP1-NSP2-NSP3-NSP4-NSP5/6
genes. The system offers good discriminatory power and
allows a very fine description of each strain’s genotype.
However, sequencing of the complete rotavirus genome is
a time-consuming process, and this may limit the applica-
tion of the new typing system. Rotavirus characterization
based on G and P genotypes is still widely used. Four G
and P genotypes were identified in this study in human
samples, including G1P[8], G1P[6], G4P[8] and G3P[9],
with G1P[8] being the most commonly detected strain.
The G1P[8] genotype is predominant in various countries
around the world (Gentsch et al. 2005; van der Heide
et al. 2005; Ahmed et al. 2006; Pietruchinski et al. 2006;
Lee et al. 2009) and is targeted by the two commercial
vaccines approved in Canada, RotaTeqR and RotarixTM
(Lamhoujeb et al. 2010). The emerging rotavirus G9 and
the rare G12 serotype were not detected in faecal samples
from Canadian children included in this study.
The human rotavirus strain CHR-120 with the G3P[9]
genotype was found to be similar to a feline rotavirus
strain isolated in the United States. In a study by Tsuga-
wa and Hoshino (2008), homology between human and
feline rotavirus strains was demonstrated in all genome
segments. Close contact between animals and humans in
the domestic environment is a factor that promotes inter-
species transmission and mixed infections. The G3 strain
has not only been identified in humans but also in
several other animal species, such as pigs, monkeys, dogs,
cats, horses, mice, lambs, birds and rabbits (Khamrin
et al. 2006; Martella et al. 2010).
Studies carried out in Argentina, the Netherlands,
Australia, Brazil and Canada showed that the G and P
genotypes commonly identified in bovine rotavirus
strains are G6, G10, P[11] and P[5] (Alfieri et al. 2004;
van der Heide et al. 2005; Lamhoujeb et al. 2010; Swiatek
et al. 2010; Badaracco et al. 2012). A similar pattern was
observed in the present study; the genotypes identified
were G6P[11], G6P[5], G10P[11] and G10P[5]. No simi-
larity has been observed between these bovine rotavirus
strains and human strains.
Results obtained in this study indicate that the
RT-qPCR assay developed by Jothikumar et al. 2009
(system C) performed very well for the detection of
human rotavirus; it was generally more sensitive than
the other RT-qPCR assays tested. However, system B is
the only assay evaluated in the study that was able to
detect rotavirus RNA in all positive human and bovine
faecal samples. Among the P and G genotypes identi-
fied in this study, G1P[8] and G6P[11] were predomi-
nant in the human and bovine rotavirus strains,
respectively. Sequence analysis of the VP4 and VP7
genes revealed that the human rotavirus strain CHR-
120 was similar to a feline rotavirus strain isolated in
the United States.
Acknowledgement
This research was supported by Agriculture and
Agri-Food Canada Research Branch Project RPBI # 1485.
References
Abid, I., Guix, S., Aouni, M., Pinto, R. and Bosch, A. (2007)
Detection and characterization of human group C
rotavirus in the pediatric population of Barcelona, Spain.
J Clin Virol 38, 78–82.
Ahmed, H.E., Coulter, J.B.S., Nakagomi, O., Hart, C.A., Zaki,
J.M., Al-Rabaty, A.A., Dove, W. and Cunliffe, N.A. (2006)
Molecular characterization of rotavirus gastroenteritis
strains, Iraqi Kurdistan. Emerg Infect Dis 12, 824–826.
Table 4 Rotavirus G and P genotypes identified in human and bovine
faecal samples
VP7 VP4
Human faecal samples positive for rotavirus
Sample 8081 - STHY-21 G1 P[8]
Sample S66825 (HSC Toronto) – STHY-119 ND * ND
Sample T54210 (HSC Toronto) – STHY-120 G3 ND
Sample W45010 (HSC Toronto) – STHY-121 G3 ND
Sample T57043 (HSC Toronto) – STHY-122 G1 P[8]
Sample M43529 (HSC Toronto) – STHY-123 G4 P[8]
Sample F76031 (HSC Toronto) – STHY-124 G1 P[6]
Sample W44279 (HSC Toronto) – STHY-125 G1 P[8]
Sample H71374 (HSC Toronto) – STHY-126 G1 P[6]
Sample H73726 (HSC Toronto) – STHY-127 ND ND
Sample M45804 (HSC Toronto) – STHY-128 ND P[8]
Sample CHR-7 G1 P[8]
Sample CHR-120 G3 P[9]
Bovine faecal samples positive for rotavirus
Sample FMV 1124422 G10 P[11]
Sample FMV 1122505 ND P[11]
Sample FMV 1094847 G10 ND
Sample FMV 1089933 G6 P[5]
Sample FMV 1089635 G6 P[11]
Sample FMV 1087558 G6 P[11]
Sample FMV 1081508 G6 P[5]
Sample FMV 1081283 ND ND
Sample FMV 1078090 G6 P[11]
Sample FMV 1077415 G10 P[5]
Sample FMV 1075017 G10 ND
Sample FMV 1075018 G6 P[11]
Sample FMV 1087766 ND P[11]
*ND: Not Determined.
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1446 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.

Alfieri, A.F., Alfieri, A.A., Bacellar Barreiros, M.A., Gagliardi
Leite, J.P. and Richtzenhain, L.J. (2004) G and P
genotypes of group A rotavirus strains circulating in calves
in Brazil, 1996–1999. Vet Microbiol 99, 167–173.
Badaracco, A., Garaicoechea, L., Rodriguez, D., Uriarte, E.L.,
Odeon, A., Galarza, R., Abdala, A., Fernandez, F. et al. (2012)
Bovine rotavirus strains circulating in beef and dairy herds in
Argentina from 2004–2010. Vet Microbiol 158, 394–399.
Banyai, K., Esona, M.D., Mijatovic, S., Kerin, T.K., Pedreira,
C., Mercado, J., Balmaseda, A., Perz, M.C. et al. (2009)
Zoonotic bovine rotavirus strain in diarrheic child,
Nicaragua. J Clin Virol 46, 391–393.
Bishop, R.F., Davidson, G.P., Holmes, I.H. and Ruck, B.J.
(1973) Virus particles in epithelial cells of duodenal
mucosa from children with acute non-bacterial
gastroenteritis. Lancet 2, 1281–1283.
Brassard, J., Seyer, K., Houde, A., Simard, C. and Trottier, Y.-
L. (2005) Concentration and detection of hepatitis A and
rotavirus in spring water samples by reverse transcription-
PCR. J Virol Methods 123, 163–169.
Escobar-Herrera, J., Cancio, C., Guzman, G.I., Villegas-
Sepulveda, N., Estrada-Garcia, T., Garcia-Lozano, H.,
Gomez-Santiago, F. and Gutierrez-Escolano, A.L. (2006)
Construction of an internal RT-PCR standard control for
the detection of human caliciviruses in stool. J Virol
Methods 137, 334–338.
Freeman, M.M., Kerin, T., Hull, J., McCaustland, K. and
Gentsch, J. (2008) Enhancement of detection and
quantification of rotavirus in stool using a modified real-
time RT-PCR assay. J Med Virol 80, 1489–1496.
Gentsch, J.R., Glass, R.I., Woods, P., Gouvea, V., Gorziglia,
M., Flores, J., Das, B.K. and Bhan, M.K. (1992)
Identification of group A rotavirus gene 4 types by
polymerase chain reaction. J Clin Microbiol 30, 1365–1373.
Gentsch, J.R., Laird, A.R., Bielfelt, B., Griffin, D.D., Banyai, K.,
Ramachandran, M., Jain, V., Cunliffe, N.A. et al. (2005)
Serotype diversity and reassortment betweem human and
animal rotavirus strains: implication for rotavirus vaccine
programs. J Infect Dis 192, S146–S159.
Gouvea, V., Glass, R.I., Woods, P., Taniguchi, K., Clark, H.F.,
Forrester, B. and Fang, Z.-Y. (1990) Polymerase chain
reaction amplification and typing of rotavirus nucleic acid
from stool specimens. J Clin Microbiol 28, 276–282.
Guti�errez-Aguirre, I., Steyer, A., Boben, J., Gruden, K.,
Poljsak-Prijatelj, M. and Ravnikar, M. (2008) Sensitive
detection of multiple rotavirus genotypes with a single
reverse transcription-real-time quantitative PCR assay.
J Clin Microbiol 46, 2547–2554.
van der Heide, R., Koopmans, M.P.G., Shekary, N., Houwers,
D.J., van Duynhoven, T.T.H.P. and van der Poel, W.H.M.
(2005) Molecular characterizations of human and animal
group A rotaviruses in the Netherlands. J Clin Microbiol
43, 669–675.
Houde, A., Gu�evremont, E., Poitras, E., Leblanc, D., Ward, P.,
Simard, C. and Trottier, Y.-L. (2007) Comparative
evaluation of new TaqMan real-time assays for the
detection of hepatitis A virus. J Virol Methods 140, 80–89.
Houde, A., Mattison, K., Ward, P. and Plante, D. (2009)
Molecular detection and characterization of food and
waterborne viruses. In Bacterial DNA, DNA Polymerase
and DNA Helicases ed. Knudsen, W.D. and Bruns, S.S. pp.
137–185. New York, USA: Nova Publishers.
Isegawa, Y., Nakagomi, O., Nakagomi, T., Ishida, S., Uesugi, S.
and Ueda, S. (1993) Determination of bovine rotavirus G
and P serotypes by polymerase chain reaction. Mol Cell
Probes 7, 277–284.
Jones, T.H., Houde, A., Poitras, E., Ward, P. and Johns, M.W.
(2009) Development and evaluation of a multiplexed real-
time TaqMan RT-PCR assay with a sample process
control for detection of F-specific RNA coliphage
genogroup I and IV. Food Environ Virol 1, 57–65.
Jothikumar, N., Kang, G. and Hill, V.R. (2009) Broadly
reactive TaqMan assay for real-time RT-PCR detection of
rotavirus in clinical and environmental samples. J Virol
Methods 155, 126–131.
Kang, G., Iturriza-Gomara, M., Wheeler, J.G., Crystal, P.,
Monica, B., Ramani, S., Primrose, B., Moses, P.D. et al.
(2004) Quantitation of group A rotavirus by real-time
reverse-transcription-polymerase chain reaction:
correlation with clinical severity in children in South
India. J Med Virol 73, 118–122.
Khamrin, P., Maneekarn, N., Peerakome, S., Yagyu, F., Okitsu,
S. and Ushijima, H. (2006) Molecular characterization of
rare G3P[3] human rotavirus reassortant strain reveals
evidence for multiple human-animal interspecies
transmissions. J Med Virol 78, 986–994.
Koopmans, M. and Duizer, E. (2004) Foodborne viruses: an
emerging problem. Int J Food Microbiol 90, 23–41.
Kottaridi, C., Spathis, A.T., Ntova, C.K., Papaevangelou, V.
and Karakitsos, P. (2012) Evaluation of a multiplex real
time reverse transcription PCR assay for the detection and
quantification of the most common human rotavirus
genotypes. J Virol Methods 180, 49–53.
Lamhoujeb, S., Cook, A., Pollari, F., Bidawid, S., Farber, J. and
Mattison, K. (2010) Rotavirus from Canadian farms
samples. Arch Virol 155, 1127–1137.
Lee, S.Y., Hong, S.K., Lee., S.G., Suh, C.I., Park, S.W., Lee,
J.H., Kim, J.H., Kim, D.S. et al. (2009) Human rotavirus
genotypes in hospitalized children, South Korea, April
2005 to March 2007. Vaccine 27S, F97–F101.
Leung, A.K., Kellner, J.D. and Davis, H.D. (2005) Rotavirus
gastroenteritis. Adv Ther 22, 476–487.
Logan, C., O’Leary, J.J. and O’Sullivan, N. (2006) Real-time
reverse transcription-PCR for detection or rotavirus and
adenovirus as causative agents of acute viral gastroenteritis
in children. J Clin Microbiol 44, 3189–3195.
Loisy, F., Atmar, R.L., Guillon, P., Le Cann, P., Pommepuy,
M. and Le Guyader, F.S. (2005) Real-time RT-PCR
for norovirus screening in shellfish. J Virol Methods 123,
1–7.
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
Reproduced with the permission of the Minister of Agriculture and Agri-food Canada. 1447
P. Ward et al. Detection and characterization of RV

Maes, R.K., Grooms, D.L., Wise, A.G., Han, C., Ciesicki, V.,
Hanson, L., Vickers, M.L., Kanitz, C. et al. (2003)
Evaluation of a human group A rotavirus assay for on-site
detection of bovine rotavirus. J Clin Microbiol 41, 290–
294.
Martella, V., Banyai, K., Matthijnssens, J., Buonavoglia, C. and
Ciarlet, M. (2010) Zoonotic aspects of rotaviruses. Vet
Microbiol 140, 246–255.
Matthijnssens, J., Ciarlet, M., Rahman, M., Attoui, H., Banyai,
K., Estes, M.K., Gentsch, J.R., Iturriza-Gomara, M. et al.
(2008) Recommendations for the classification of group A
rotaviruses using all 11 genomic RNA segments. Arch
Virol 153, 1621–1629.
Matthijnssens, J., Ciarlet, M., McDonald, S.H., Attoui, H.,
Banyai, K., Brister, J.R., Buesa, J., Esona, M.D. et al.
(2011) Uniformity of rotavirus strain nomenclature
proposed by the Rotavirus Classification Working Group
(RCWG). Arch Virol 156, 1397–1413.
Matthijnssens, J., Otto, P.H., Ciarlet, M., Desselberger, U., Van
Ranst, M. and Johne, R. (2012) VP6-sequence-based cutoff
values as a criterion for rotavirus species demarcation.
Arch Virol 157, 1177–1182.
Mattison, K., Brassard, J., Gagn�e, M.-J., Ward, P., Houde, A.,
Lessard, L., Simard, C., Shukla, A. et al. (2009) The feline
calicivirus as a sample process control for the detection of
food and waterborne RNA viruses. Int J Food Microbiol
132, 73–77.
Min, B.-S., Noh, Y.-J., Shin, J.-H., Baek, S.-Y., Min, K.-I., Ryu,
S.-R., Kim, B.-G., Park, M.-K. et al. (2006) Assessment of
the quantitative real-time polymerase chain reaction using
a cDNA standard for human group A rotavirus. J Virol
Methods 137, 280–286.
Pang, X.L., Lee, B., Boroumand, N., Leblanc, B. and
Preiksaitis, K. (2004) Increased detection of rotavirus
using a real time reverse transcription-polymerase chain
reaction (RT-PCR) assay in stool specimens from children
with diarrhea. J Med Virol 72, 496–501.
Pang, X., Cao, M., Zhang, M. and Lee, B. (2011) Increased
sensitivity for various rotavirus genotypes in stool
specimens by amending three mismatched nucleotides in
the forward primer of a real-time RT-PCR assay. J Virol
Methods 172, 85–87.
Pietruchinski, E., Benati, F., Lauretti, F., Kisielius, J., Ueda, M.,
Volotao, E.M., Soares, C.C., Hoshino, Y. et al. (2006)
Rotavirus Diarrhea in children and adults in a Southern
city of Brazil in 2003: distribution of G/P types and
finding of rare G12 strain. J Med Virol 78, 1241–1249.
Plante, D., B�elanger, G., Leblanc, D., Ward, P., Houde, A. and
Trottier, Y.-L. (2011) The use of bovine serum albumin to
improve the RT-qPCR detection of foodborne viruses
rinsed from vegetable surfaces. Lett Appl Microbiol 52,
239–244.
Rahman, M., Banik, S., Faruque, A.S.G., Taniguchi, K., Sack,
D.A., Van Ranst, M. and Azim, T. (2005) Detection and
characterization of human group C rotaviruses in
Bangladesh. J Clin Microbiol 43, 4460–4465.
Raming, F., Ciarlet, M., Mertens, P.P.C. and Dermody, T.S.
(2005) Rotavirus. In Virus Taxonomy; eight report of the
international committee on taxonomy of viruses ed. Fauquet,
C.M., Mayo, M.A., Maniloff, J., Desselberger, U. and Ball,
L.A. pp. 484–496. London: Elsevier Academic Press.
Rutjes, S.A., Lodder, W.J., Bouwknegt, M. and de Roda
Husman, A.M. (2007) Increased hepatitis E virus
prevalence on Dutch pig farms from 33 to 55% by using
appropriate internal quality controls for RT-PCR. J Virol
Methods 143, 112–116.
Schwarz, B.-A., Bange, R., Vahlenkamp, T.W., Johne, R. and
M€uller, H. (2002) Detection and quantitation of group A
rotaviruses by competitive and real-time reverse
transcription-polymerase chain reaction. J Virol Methods
105, 277–285.
Scipioni, A., Bourgot, I., Mauroy, A., Ziant, D., Saegerman, C.,
Daube, G. and Thiry, E. (2008) Detection and
quantification of human and bovine noroviruses by a
TaqMan RT-PCR assay with a control for inhibition. Mol
Cell Probes 22, 215–222.
Steyer, A., Bajzelj, M., Iturriza-Gomara, M., Mladenova, Z.,
Korsun, N. and Poljsak-Prijatelj, M. (2010) Molecular
analysis of human group A rotavirus G10P[14] genotype
in Slovenia. J Clin Virol 49, 121–125.
Swiatek, D.L., Palombo, E.A., Lee, A., Coventry, M.J., Britz,
M.L. and Kirkwood, C.D. (2010) Detection and analysis of
bovine rotavirus strains circulating in Australian calves
during 2004 and 2005. Vet Microbiol 140, 56–62.
Tate, J.E., Burton, A.H., Boschi-Pinto, C., Steele, A.D., Duque,
J. and Parashar, U.D. (2012) 2008 estimate of worldwide
rotavirus-associated mortality in children younger than
5 years before the introduction of universal rotavirus
vaccination programmes: a systematic review and meta-
analysis. Lancet Infect Dis 12, 137–141.
Tsugawa, T. and Hoshino, Y. (2008) Whole genome sequence
and phylogenetic analyses reveal human rotavirus G3P[3]
strains Ro1845 and HCR3A are examples of direct virion
transmission of canine/feline rotaviruses to humans.
Virology 380, 344–353.
Ward, P., Poitras, E., Leblanc, D., Letellier, A., Brassard, J., Plante,
D. and Houde, A. (2009) Comparative analysis of different
TaqMan real-time RT-PCR assays for the detection of swine
Hepatitis E virus and integration of Feline calicivirus as
internal control. J Appl Microbiol 106, 1360–1369.
Zeng, S.-Q., Halkosalo, A., Salminen, M., Szakal, E.D.,
Puustinen, L. and Vesikari, T. (2008) One-step
quantitative RT-PCR for the detection of rotavirus in
acute gastroenteritis. J Virol Methods 153, 238–240.
© Her Majesty the Queen in Right of Canada [2013]. 114, 1435--1448
1448 Reproduced with the permission of the Minister of Agriculture and Agri-food Canada.
Detection and characterization of RV P. Ward et al.
Recommended

















