
University of South FloridaScholar Commons
Graduate Theses and Dissertations Graduate School
2009
Cephalopods of the Broad Caribbean:Distribution, abundance, and ecologicalimportanceHeather L. JudkinsUniversity of South Florida
Follow this and additional works at: http://scholarcommons.usf.edu/etd
Part of the American Studies Commons
This Dissertation is brought to you for free and open access by the Graduate School at Scholar Commons. It has been accepted for inclusion inGraduate Theses and Dissertations by an authorized administrator of Scholar Commons. For more information, please [email protected].
Scholar Commons CitationJudkins, Heather L., "Cephalopods of the Broad Caribbean: Distribution, abundance, and ecological importance" (2009). GraduateTheses and Dissertations.http://scholarcommons.usf.edu/etd/2034

Cephalopods of the Broad Caribbean: Distribution, Abundance, and Ecological
Importance
by
Heather L. Judkins
A dissertation submitted in partial fulfillment of the requirements for the degree of
Doctor of Philosophy College of Marine Science University of South Florida
Co-Major Professor: Joseph Torres, Ph.D. Co-Major Professor: Michael Vecchione, Ph.D.
Susan Bell, Ph.D. David Mann, Ph.D. Mark Luther, Ph.D.
Date of Approval: June 10, 2009
Keywords: mollusca, Caribbean diversity, biogeography, latitudinal gradients, coastal Atlantic Ocean
©Copyright 2009, Heather L. Judkins

ACKNOWLEDGEMENTS I would like to acknowledge the team of people who have helped me through this
process. Dr. Joseph Torres, Dr. Michael Vecchione, and my entire committee, Dr. Susan
Bell, Dr. David Mann, and Dr. Mark Luther, have been instrumental in terms of advice,
pathways to follow, and excellent sounding boards for my many questions. I would like
to acknowledge Dr. Clyde Roper for his foresight in seeing that I had potential to add a
small part of research to the cephalopod scientific community and his words of wisdom
and encouragement throughout this entire venture. I would like to thank Dr. Nancy Voss
for her advice and guidance in understanding of cephalopod species of the Broad
Caribbean. Her assistance in my work is beyond compare. Dr. Ronald B. Toll and Dr.
Steven C. Hess were instrumental in documenting cephalopod specimen information
from the Hourglass Cruises that were conducted from 1965 through 1967, by the Marine
Research Laboratory of the Florida Board of Conservation on the continental shelf of the
Gulf of Mexico which was included in my dissertation. I also thank Teresa Greely and
Angela Lodge for their assistance throughout the two years of my GK12 Oceans
Fellowship and the opportunity to coordinate the regional competition for the National
Ocean Sciences Bowl that I have been so excited to be a part of for the College of Marine
Science at USF. Lastly, I wouldn’t be where I am without the unfailing support and
constant source of smiles from my family and friends who have been there through the
ups and downs of my journey. Thank you.

i
TABLE OF CONTENTS TABLE OF CONTENTS i
LIST OF TABLES iii
LIST OF FIGURES iv
ABSTRACT v
CHAPTER 1. INTRODUCTION 1
1.1 Broad Caribbean 1 1.2 Cephalopod Biology 3
1.3 Distribution of Cephalopods 4 1.4 Abundance of Cephalopods 4 1.5 Cephalopods in Food Webs 5 1.6 Overall Significance 7 1.7 Study Goals 8 1.8. Study Area 9
1.9 Materials and Methods 9 1.10 References 13
CHAPTER 2. BIOGEOGRAPHY OF CEPHALOPODS OF THE BROAD
CARIBBEAN 16 2.1 Introduction 16
Broad Caribbean 16 The Cephalopods 18 Biogeography of Cephalopods 18 Abundance of Cephalopods 21
Ecological Focus 21 Study Area 24
2.2 Materials and Methods 24 Statistical analysis 26 2.3 Results 27
Rapoport’s Rule and Species Richness 27 Cephalopod Biogeography 28
Distribution and Abundance 29 Range Extensions 29
New Species to Area 38 2.4 Discussion 38
Rapoport’s Rule and Species Richness 38 Cephalopod Biogeography 41

ii
Distribution and Abundance 43 Range Extensions 45
New Species 45 2.5 References 47
CHAPTER 3. CEPHALOPODA (MOLLUSCA: CEPHALOPODA) OF THE
GULF OF MEXICO 52 3.1 Introduction 52
3.2 Major Systematic Revisions 56 3.3 Comparative Assessment of the Group in the Gulf of Mexico 56 3.4 Explanation of Checklist 57 3.5 Acknowledgements 60 3.6 References 61 3.7 Gulf of Mexico Checklist 66 3.8 Taxonomic Summary for Cephalopods of the Gulf of Mexico 73
CHAPTER 4. FIRST RECORDS OF Asperoteuthis acanthoderma (Lu, 1977)
(CEPHALOPODA: OEGOPSIDA: CHIROTEUTHIDAE) FROM THE NORTH ATLANTIC OCEAN; STRAITS OF FLORIDA 74
4.1 Abstract 74 4.2 Introduction 75 4.3 Systematics 76 4.4 Description 76 4.5 Discussion 81 4.6 Acknowledgements 86 4.7 References 87 4.8 Figure Legends 88
CHAPTER 5. CONCLUSIONS 94 5.1 Summary 94 5.2 References 100
APPENDICES 102 Appendix A: Regional distribution point coordinates 103 Appendix B: Species richness comparison quadrats 104 Appendix C: Cephalopod distribution maps 105
ABOUT THE AUTHOR End page

iii
LIST OF TABLES
Table 2.1 Regional cephalopod locations 20
Table 2.2 Region, species number and sample number for hotspots 33
Table 2.3 Cephalopod range extensions to Broad Caribbean 35
Table 3.1 Checklist of the Cephalopods of the Gulf of Mexico 66
Table 3.2 Taxonomic summary for cephalopods of the Gulf of Mexico 73
Table 4.1 Measurements of Asperoteuthis acanthoderma specimens 89
Table 5.1 Gulf of Mexico checklist species not included in Ch. II 95

iv
LIST OF FIGURES
Figure 1.1 Current flow through the Caribbean Sea and Gulf of Mexico 2
Figure 1.2 A summary of the role of cephalopods in the world’s oceans 6
Figure 1.3 Study Area 9
Figure 2.1 Current flow through the Caribbean Sea and Gulf of Mexico 17
Figure 2.2 Study area 24
Figure 2.3 Species richness per 5˚ latitudinal band 30
Figure 2.4 Comparison of species richness and number of individuals 30
Figure 2.5 Rapoport’s Rule- Rarefaction curve 31
Figure 2.6 Rapoport’s Rule- Chao 1 estimator graph 31
Figure 2.7 Eight study sites for richness and diversity comparisons 32
Figure 2.8 Species richness regional comparison 32
Figure 2.9 Hotspot species observed- Rarefaction curve 33
Figure 2.10 Hotspot Chao 1 estimator of regional species richness 34
Figure 2.11 Broad Caribbean sample effort map 34
Figure 4.1 Key West Asperoteuthis acanthoderma specimen 91
Figure 4.2 Marathon Asperoteuthis acanthoderma specimen 92
Figure 4.3 Internal organs of the Key West specimen 93

v
CEPHALOPODS OF THE BROAD CARIBBEAN SEA: DISTRIBUTION, ABUNDANCE, AND ECOLOGICAL IMPORTANCE
Heather L. Judkins
ABSTRACT
The Broad Caribbean region is defined as the Gulf of Mexico, and the coastal and
marine areas of the Caribbean Sea, including the chain of islands forming the Greater and
Lesser Antilles, Turks and Caicos, the Bahamas, and the gulf coasts of the United States,
Central and South America (Stanley, 1995). The cephalopods of the Broad Caribbean
were examined in terms of distribution, abundance, and ecological importance. A suite
of 5190 preserved cephalopod specimens were identified and catalogued to produce
regional maps of cephalopod distribution within the Broad Caribbean. Eighteen range
extensions were noted for known species. Regional species richness was examined with
respect to Rapoport’s Rule with an eye toward possible cephalopod hotspots in the
region. Cephalopods of the Broad Caribbean within the latitudinal bands of 8˚N and
30˚N do not support Rapoport’s Rule as they exhibit increasing species richness with
increasing latitude. Eight subareas were chosen to compare species richness. Regionally,
species richness is patchy, with the largest concentration of cephalopods off the eastern
Florida coast. Areas of the southern Caribbean Sea are in need of more samples for
accurate assemblage counts and more meaningful comparisons with other Caribbean
regions. Rarefaction curves were used to normalize the variously sized samples
throughout the Broad Caribbean. A checklist of the Gulf of Mexico based on literature
developed a picture for the northern regions of the Broad Caribbean. This checklist

vi
provided an updated account of cephalopod species that were reported from smaller
literature works. Lastly, the first observation in the North Atlantic Ocean of the deep-sea
squid Asperoteuthis acanthoderma (family Chiroteuthidae) was described. The
description is based on two nearly intact, but damaged, specimens that were found
floating at the surface in the waters off Key West and Marathon, Florida in 2007. All
previously known records are recorded from a few specimens scattered in the western
Pacific Ocean. There is a need for increased sampling throughout the Broad Caribbean to
explore the systematics, life histories, distribution patterns, and potential fisheries for this
group of organisms.

1
CHAPTER 1
INTRODUCTION
Cephalopods of the Broad Caribbean have not been reviewed in recent years. The
unique features, currents, and coastlines of the Gulf of Mexico, Caribbean and the Straits
of Florida combine as a potentially unique ecotone for cephalopod distribution,
abundance, and diversity. Work conducted by G. Voss (1956), C.F. Roper (1984), N.
Voss (1998), M. Vecchione (2002), and others provide background for further
exploration of cephalopods in this area. The present study focuses on over 5000
specimens collected from the area to extend cephalopod research.
1.1 The Broad Caribbean
The Broad Caribbean region is defined as the Gulf of Mexico, and the coastal and
marine areas of the Caribbean Sea, including the chain of islands forming the Greater and
Lesser Antilles, Turks and Caicos, the Bahamas, and the Gulf coasts of North, Central
and South America (Stanley, 1995).
The region is influenced by waters that flow through the lower Caribbean islands,
originating from the Guiana Current that moves north along Brazil’s coast. The Guiana
Current is joined by the North Equatorial Current, which flows through the lower
Caribbean, veering north around western Cuba and into the Gulf of Mexico. Some
upwelling occurs along the southern region of the Caribbean Sea (Longhurst, 1998). The
majority of the flow moves around the Gulf coasts of the United States, flowing down
along the west Florida coast before moving through the Straits of Florida to become the

2
Gulf Stream which moves northward through the Bahamas and along the eastern coast of
Florida. The general movement of currents in the Broad Caribbean is from east to west
with gyres often spinning off the main water flow (Stanley, 1995).
Fig 1.1: Major currents of Broad Caribbean (from Carpenter et al. 2002)
The surface temperature of the ocean in the tropical half of the Broad Caribbean
region is averages 27˚C with little variation throughout the year. Temperatures in the
southern portion of the Gulf of Mexico also average near 27˚C but the northern Gulf of
Mexico has larger temperature fluctuations due to seasonal changes from 16˚C in winter
to 28˚C in summer (Stanley, 1995).
Salinity is relatively high in the surface waters between January and May, 36.36,
and lower between June and December in the region, 36.06 (Tomczak & Godfrey, 1994),
mainly due to the inflow in late fall of lower-salinity waters from the Orinoco and
Amazon Rivers as well as equatorial convergence coming through the region.
Geologically, the Caribbean Sea consists of two main basins separated by a broad,
submarine plateau. Two tectonic plates, the Caribbean plate and the North American

3
plate, potentially influence historical biogeography in the area. The Cayman Trench, a
trench between Cuba and Jamaica, contains the Caribbean’s deepest point (7,535 m
below sea level).
1.2 Cephalopod Biology
The cephalopods are molluscs with many unique features separating them from
other molluscan groups including a closed circulatory system, the reduction and in most
cases, absence of an external shell, a sophisticated nervous system, jet propulsion for
movement, many camouflage adaptations, and a predatory lifestyle (Brusca, 2003).
Cephalopods possess a biting apparatus resembling an inverted parrot beak, and
almost all have some form of radula, or rasping tongue. Food-capturing mechanisms
vary from sucker-bearing arms and tentacles to hooks modified from suckers for catching
prey. In particular, nektonic cephalopods are impressive predators.
Octopods are adept in stalking or hiding out to attack prey. Neritic octopods
home in visually on prey and they approach by partially raising an arm and gliding in that
direction. The attack is a pounce during which many of the arms and the web are thrown
over the prey to immobilize it. The octopods will then bite and poison the organism
(Vecchione, 2002).
Although research is scare in terms of the life history of many cephalopod
species, the consensus is that they possess a fast growth rate and relatively short lifespan.
They grow exponentially until mature and then they spawn and die. Cephalopods have
various spawning methods ranging from polycyclic spawning and multiple spawning to
intermittent terminal and continuous spawning. All spawning strategies end in the death
of the cephalopods after the spawning period (Jereb & Roper, 2005).

4
1.3 Distribution of Cephalopods
Past cephalopod studies have focused on small pockets of the Broad Caribbean
such as the Gulf of Mexico and Florida regions by Voss (1956), the Straits of Florida by
Cairns (1976), the eastern Gulf of Mexico, Passerella (1991), and regions surrounding
Colombia, by Gracia (2002). Cephalopods have various distribution patterns ranging
from coastal species to entirely pelagic. Coastal species include the economically
important lologinids and octopods. Offshore, the ommastrephids and histioteuthids are of
greater importance.
Cephalopod species exhibit considerable diversity in their depth distribution as
well. Squid, such as Doryteuthis are surface dwellers living within 500m of the ocean
surface while chiroteuthids are deep-sea organisms with ranges down to 4000m and
beyond. There are temperature tolerance patterns that exist among cephalopods; certain
cephalopods thrive in the warmer waters of the Caribbean proper (Octopus zonatus,
Octopus maya) while others have an affinity for the subtropical temperatures of the
northern sections of the Gulf of Mexico such as Ancistroteuthis lichensteinii.
Distribution patterns are important for researchers and fishery experts to understand as
conservation measures and management practices are created for cephalopod groups.
1.4 Abundance of Cephalopods:
Since their appearance in the Cambrian, cephalopods have evolved to include
some of the largest past and present invertebrates (over 20m) and to become common
predators in all shallow and deep seas. For part of life’s history, over 200 million years,
they were probably the top predators in the marine environment. After the Jurassic, it is
believed that fishes superseded their importance as predators. While fishes and

5
cephalopods both evolved to cope with the same physical marine environment, much
stimulus to cephalopod evolution also came from their interaction with fish and later with
other vertebrate predators, the reptiles, seals, cetaceans, and birds (Clarke, 1996).
Cephalopod species vary in abundance in the Broad Caribbean ranging from
schools of thousands in Illex species to solitary individuals of Discoteuthis discus and
Asperoteuthis acanthoderma. Cephalopods today are not as numerous as they once were
due to the constant struggle between them and all other marine species, including other
cephalopods. Their present success is apparent in their morphological diversity,
abundance, and major role they play in the ecology of the sea (Young et al. 1998).
1.5 Cephalopods in the food web
Cephalopods are important predators on micronekton and macrozooplankton and
a major food source for nektonic fish. Diets of cephalopods change as they mature. Most
begin as paralarvae preying upon small crustaceans, and as they grow, move onto larger
crustaceans, fish, and other mollusca. Small, immature longfin inshore squid (Doryteuthis
pealeii), for example, will feed on planktonic organisms while larger individuals feed on
larger crustaceans and small fish. Studies have also shown that the immature squid will
feed on euphausiids and arrow worms while adults feed on small crabs, polychaetes, and
shrimp (Cargnelli, 1999). Fish species preyed on by the longfin inshore squid include
silver hake, mackerel, herring, menhaden, sand lance, bay anchovy, weakfish, and
silversides. As a food source for other organisms, cephalopods have been found in large
numbers in the diets of seabirds, seals, whales, and fish (Cargnelli, 1999).
Prey preference varies among species. The congeners, Doryteuthis opalescens
(Pacific) and Doryteuthis pealeii (Atlantic) are important commercial squid with

6
different adult sizes and prey preferences. A study by Karpov and Cailliet (1978) found
that Doryteuthis opalescens fed mainly on fish, cephalopods, gastropods, and
polychaetes. Doryteuthis pealeii were found to eat more equal proportions of fish and
fellow squid (Amaratunga, 1983).
Figure 1.2 A summary of the role of cephalopods in the world’s oceans and seas as expressed by their position in the energetic hierarchy. (Clarke, 1996)
Cephalopods are born into the third trophic level and progress one to two stages
through their life. Research has not shown them achieving top-predator status because
there always seems to be some vertebrate that preys upon them (Summers, 1983). Diving
birds such as penguins and murres actively search for cephalopods as part of their diet as
do toothed whales and seals. Sei and Fin whales are the major baleen predators on
oceanic squid. Those whales seek copepods and euphausiids, which are probably also the
prey of squids. Examples of seabirds dependent on cephalopods would be the thick-

7
billed murres of the North Pacific and the emperor penguins of the Antarctic (Williams,
1995). Ogi lists squid as 72.6% of the diet of thick-billed murres (Summers, 1983).
1.6 Overall significance:
Cephalopods play an important role in the Broad Caribbean region in terms of
fisheries and overall biodiversity. At present, cephalopods contribute only 3% to the
tonnage of global fisheries, but their total value lies third, below only shrimps and tuna
(Clarke, 1996). Cephalopods are important as a food source for humans and well over 3
million tons are caught yearly. Fishing has become increasingly intense in the Orient and
the eastern Atlantic Ocean. Most of the cephalopods caught in the Broad Caribbean were
landed by Mexican fishermen, bringing in between 19,000 and 31,000 tons (Vecchione,
2002).
Ideally, a thorough knowledge of the systematics of a species is the required
foundation upon which all other biological and resource management studies must be
based. For example, in the Gulf of Campeche, Mexico, a traditional fishery was believed
to be based on Octopus “vulgaris” Lamarck, 1798, a ubiquitous octopus of broad
distribution. In the absence of local studies, knowledge about the biology of O.
“vulgaris” from other seas was applied to the Campeche octopus for fishery statistics
and management purposes. The discovery that the octopus was actually a new species,
described as O. maya, Voss and Solis, 1966, with a very different life history, explained
the problems that had plagued biologists assigned to study the fishery and develop
recommendations. This example underscores the need for sound systematic knowledge
of species and populations if we are to regulate fisheries for these forms (Roper, 1983).

8
Biodiversity studies are moving from a focus on individual species to the
ecosystem level. Similarly, fisheries management has progressed from single species to
multiple target species to ecosystems (such as the large marine ecosystem approach of
Sherman et. al. 1990). For an ecosystem management effort to have any chance of
success, information is needed on all abundant or ecologically important species
(Vecchione and Collette, 1996). Fisheries management is an integral part of ecosystem
management practices. Changes in both targeted and bycatch species need to be
monitored. An example is the fishery for bottom fish on Georges Bank. Of the fish
caught in 1963, 67% were the prized cod, hake, and flounders, whereas 24% was made
up of unwanted species. By 1986 the dominant catch had shifted dramatically, with 14%
wanted species and 74% “junk species”. Such changes in populations of large predators
cause profound effects throughout the food web (Vecchione and Collette, 1996).
1.7 Study Goals
The goals of the present study include elements of biogeography, distribution, and
ecology of cephalopods. They are:
1. To determine the distribution and abundance of cephalopod species in the Broad Caribbean region.
2. To discern general biogeographic patterns of the group in the Broad
Caribbean region. 3. To examine the distribution of the group for possible alignment to Rapoport’s
Rule (RR) as studied by other authors (Macpherson 2002, Fortes 2004, Rosa et al., 2008).
4. To create a comprehensive checklist of the major species found in the Gulf of
Mexico.
5. To describe new species to the region. This includes range extensions for currently known species in the area.

9
1.8 Study Area
The Broad Caribbean includes the Bahama Islands, Gulf of Mexico, and the
Caribbean Sea. The range for the study was between 8 ° N -30° N and 58° W – 97° W.
Figure 1.3: Study Area
1.9 Materials and Methods
The most helpful taxonomic information include examination of comparative
material from a variety of locations, including from the type locality when possible
(Vecchione; Collette, 2000). Based on current cephalopod dichotomous keys, type
specimens, literature, and expert opinions, the specimens analyzed were identified to the
species level. Once the specimen was identified, it was plotted on a distribution map of
the Broad Caribbean using the ArcView 9.2 mapping program.
An acceptable biogeographic study includes many specimens of each species
under consideration, including all the life-history stages, precise distribution within the

10
study area, and a complete geographic range of each species. This study included 4190
specimens that had known locales and 1000 secondary regional specimens that were
distributed throughout the Broad Caribbean for a total o 5190 specimens examined.
Many of the regional specimens were from the Caribbean Sea and were important to
include. The regional data was used in the analysis of the distributions of cephalopods
but not included for the application of RR.
A key component of biodiversity exploration is the discovery of new species.
Some factors that affect proper identification of cephalopods are the:
- Basis for identification: recent comprehensive reviews are more reliable. - Experience level of identifiers: some taxonomic groups are more difficult than
others to identify. - Difficulty of characters necessary for confident identification. - Life-history stages examined. - Conditions of specimens: quality can be greatly affected by methods used for
collection, fixation and preservation, potentially limiting or eliminating the usefulness of important characteristics.
- Possible presence of similar, easily confused species: confidence in identification is limited.
(Vecchione and Collette, 2000) The above six factors were taken into consideration while the study was
conducted. Identification guides (Voss 1956; Nesis 1975, 1987; Roper and Young, 1984;
N. Voss 1998; Vecchione, 2002) were used to identify each organism to species level.
The majority of the specimens examined had been preserved well and were in excellent
condition. The specimens date from 1898 to present.
Identification of specimens was conducted in the micronekton museum at the
College of Marine Science at the University of South Florida, the Rosenstiel School for
Marine and Atmospheric Science, University of Miami as well as the National Museum
of Natural History at the Smithsonian Institution in Washington, D.C.. A dissecting

11
microscope was used to identify small specimens and specific features of the animals.
Micrometer calipers were used for length measurements. The bulk of preserved
specimens analyzed were from two institutions, the Smithsonian Institution’s National
Museum of Natural History and the University of Miami’s Rosenstiel School of Marine
and Atmospheric Science. Smaller collections from the Florida Fish and Wildlife
Research Institute were also analyzed.
The methods used for identifying and cataloguing specimens were to identify the
individual using the cephalopod dichotomous keys, and to record its mantle length, sex,
number of gill lamellae (octopods), and species name into an excel file for statistical
analysis. If the specimen was in a mixed lot, individuals were separated according to
species. All station and cruise information was copied and put into the additional
collection jars as needed. Once all the organisms were identified, known specimens from
all institutions were also added to the excel file for analysis. Distribution was calculated
using the 9.2 ArcView software program.
Chapter II of the dissertation focuses on the biogeography of cephalopods in the
Broad Caribbean region. Species richness of cephalopods was analyzed for alignment to
Rapoport’s Rule which was proposed by Stevens (1989): that species richness tends to be
greater towards lower latitudes. Potential cephalopod “hotspots” are compared within 8
subareas of the Broad Caribbean and lastly, distribution of all species examined are
plotted on regional maps of the area. Eighteen range extensions are added to the
cephalopod database for the region. The third chapter of this study utilizes cephalopod
literature as the basis for a species checklist of the Gulf of Mexico. Comprehensive
studies of large oceanic regions are not present in current cephalopod research. The

12
chapter updates Voss (1956) and utilizes over 40 cephalopod studies of the area to
compile the checklist. Chapter IV describes a range extension of the chiroteuthid,
Asperoteuthis acanthoderma, a species newly found in the Broad Caribbean. It describes
two specimens discovered floating dead at the surface off the coast of Key West and
Marathon, Fl in 2007. Both specimens were examined by the author in collaboration
with experts in the field. Unique defining characteristics were used for identification
with emphasis on the Y-shaped funnel locking mechanism, sucker ring form and
dentition, beak morphology, photophore patch configuration on ventral surface of
eyeballs, and the numerous small cartilaginous tubercles that cover the mantle, head and
aboral surface of the arms. The concluding chapter is an overall summary of the Broad
Caribbean cephalopod assemblage.

13
REFERENCES
1. Amaratunga, T.1983. The Role of Cephalopods in the Marine Ecosystem. Advances in assessment of world cephalopod resources; 231: 379-415.
2. Brusa, Richard C., Gary J. Brusca. 2003. Invertebrates, Second edition; Sinauer Associates, Inc. Publishers; Sunderland, Ma; p. 714-760. 3. Cairns, S. 1976. Cephalopods collected in the Straits of Florida by the R/V Gerda.
Bulletin of Marine Science; 26(2): 233-272. 4. Cargnelli, L., Sara Griesbach, Cathy McBride,Christine A. Zetlin, and Wallace W.
Morse, 1999. Longfin Inshore Squid, Loligo pealeii, Life History and Habitat Characteristics, in NOAA Technical Memorandum NMFS-NE-146. NOAA: Northeast Region, Woods Hole, MA. 1-27.
5. Clarke, M.R. 1996. The role of cephalopods in the world’s oceans: an introduction.
Phil.Trans. R. Society London. B.; 351: 979-983. 6. Fortes, R.R., R.S. Absalao, 2004. The applicability of Rapoport’s rule to the marine
molluscs of the Americas. Journal of Biogeography; 31: 1909-1916. 7. Gracia, A. 2002. Cephalopods (Mollusca: Cephalopoda) of the Upper Colombian
Caribbean Shelf. Biol. Invest. Mar. Cos. 31, 219-238. 8. Jereb, P. and Roper, C.F.E. 2005. Cephalopods of the world. An annotated and
illustrated catalogue of cephalopod species known to date. FAO species catalogue for fisheries purposes, N4, v.1, FAO, Rome.
9. Karpov K.A., & G.M. Calliet. 1978. Feeding dynamics of Loligo opalescens. Calif. Dept. of Fish and Game Bulletin; 169: 45-65. 10. Longhurst, A.1998. Ecological geography of the sea. Academic Press; 146-150. 11. Macpherson, E. 2002. Large-scale species-richness gradients in the Atlantic Ocean.
Proceedings of the Royal Society of London; B; 269: 1715-1720.
12. Nesis K.N. 1975. Cephalopods of the American Mediterranean Sea; English Translations of Selected Publications on Cephalopods; Editor M. Sweeney; Smithsonian Institutions Libraries; 1; 318- 358.
------ 1987. Cephalopods of the world. (Edited by L. Burgess) T.F.H. Publication, Inc. Ltd. New Jersey; 351 pp.

14
13. Passarella, K.C., 1990. Oceanic Cephalopod Assemblage in the Eastern Gulf of Mexico, in Department of Marine Science. University of South Florida: St. Petersburg, Fl. 50pp.
14. Roper, C.F.E., 1983. An overview of cephalopod systematics: status, problems, and
recommendations. Memoirs of the National Museum Victoria.; 44: 13-27. 15. Roper, C. F. E., M. J. Sweeney, and C. Nauen. 1984. Cephalopods of the World. An
Annotated and Illustrated Catalogue of Species of Interest to Fisheries. FAO Fisheries Synopsis No.125, v.3, Food and Agricultural Organization of the United Nations, Rome. 277 pp.
16. Rosa, Rui, Heidi M. Dierssen, Liliana Gonzalez, and Brad Seibel. 2008. Ecological
biogeography of cephalopod molluscs in the Atlantic Ocean: historical and contemporary causes of coastal diversity patterns; Global Ecology and Biogeography; 17: 600-610.
17. Sherman K., L.M. Alexander & B.D. Gold. Editors.1990. Large marine ecosystems:
patterns, processes, and yields. American Association for the Advancement of Science, Washington D.C.
18. Stanley, Sonja (editor), 1995. A Global Representative System of Marine Protected
Areas: Vol 2, Marine Region 7: Wider Caribbean, Great Barrier Reef Marine Park Authority, The World Bank, The World Conservation Union, (IUCN); 15-42.
19. Stevens, G.C., 1989. The Latitudinal Gradient in Geographical Range: How So Many Species Coexist in the Tropics. The American Naturalist; 133(2): 240-256.
20. Summers, W.C., 1983. Physiological and Trophic Ecology of Cephalopods. The Mollusca. Ecology; 6: 261-279.
21. Tomczak, Mattias, J. Stuart Godfrey. 1994. Regional Oceanogrpahy: An
Introduction. Pergamon; 310-317. 22. Vecchione, M., Bruce B. Collette, 1996. Fisheries Agencies and Biodiversity. Ann.
Missouri Bot. Gard; 83: 29-36.
23. Vecchione, M., M.F. Mickevich, K. Fauchald, B.B. Collette, A.B. Williams, T.A. Munroe, and R.E. Young, 2000. Importance of Assessing Taxonomic Adequacy in Determining Fishing Effects on Marine Diversity. ICES Journal of Marine Science; 57: 677-681.

15
24. Vecchione, 2002. Cephalopods. Carpenter, K.E.(ed.) The living marine resources of the Western Central Atlantic. Volume 1: Introduction, molluscs, crustaceans, hagfishes, sharks, batoid fishes, and chimaeras. Cephalopods of Western Central Atlantic; FAO Species Identification Guide for Fisheries Purposes and American Society of Ichthyologists and Herpetologists Special Publication No. 5. Rome, FAO. 2002. 1-600.
25. Voss, G.L. 1956. A review of the cephalopods of the Gulf of Mexico. Bull Mar.
Sci. Gulf and Caribbean; 6:85-178.
26. Voss, N.A., Vecchione, M., Toll, R.B. & Sweeney, M.J.(eds) 1998. Systematics and biogeography of cephalopods. Smithson, Contrib. Zool.; 586: 1-599.
27. Williams, T.D. 1995. The penguins: Spheniscidae. Oxford University Press, Oxford.
28. Young, R. E, L. A. Burgess, C. F. E. Roper, M. J. Sweeney, and S. J. Stephen 1998. Classification of the Enoploteuthidae, Pyroteuthidae, and Ancestrocheiridae. Pp. 239–255 in N. A. Voss, R. B. Toll, and M. Vecchione, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology 586(1).

16
CHAPTER 2
BIOGEOGRAPHY OF CEPHALOPODS OF THE BROAD CARIBBEAN REGION
2.1 Introduction
Cephalopod studies in the Caribbean region have not provided a well-rounded,
comprehensive view of distribution and abundance in the group. Various island groups
or individual cephalopod species have been addressed, (Diaz, 2000, 2004; Gracia, 2002)
mainly in coastal waters, but to date, no study describes the Broad Caribbean species
complex as a whole. Rosa et al. (2008) and Smith et al. (2002) conducted literature-
based studies on latitudinal gradients of species richness but none have utilized large
numbers of specimens from the region to improve our understanding of cephalopod
ecology. This paper fills that need and based on 5190 specimens, reports on the
distribution, abundance, and diversity of cephalopods in the Broad Caribbean region.
The Broad Caribbean
The Broad Caribbean region is defined as the Gulf of Mexico and the coastal and
marine areas of the Caribbean Sea, including the chain of islands forming the Greater and
Lesser Antilles, Turks and Caicos, the Bahamas, and the gulf coasts of the United States,
Central and South America (Stanley, 1995). The Caribbean Sea proper covers
approximately 1,063,000 square miles while the Gulf of Mexico is smaller, covering an
area approximately 1,592,842 square kilometers (615,000 sq. miles).

17
The region is influenced by waters that flow through the lower Caribbean islands,
originating from the Guiana Current that moves north along Brazil’s coast. The Guiana
Current is joined by the North Equatorial Current, which flows through the lower
Caribbean, veering north around western Cuba and into the Gulf of Mexico. Some
upwelling occurs along the southern region of the Caribbean Sea (Longhurst, 1998).
Loop Current water moves through the Gulf of Mexico, flowing down the west Florida
coast before moving through the Straits of Florida. The water becomes the Gulf Stream,
which moves northward through the Bahamas and eastern coast of Florida. The general
movement of the Broad Caribbean is from east to west with gyres often spinning off the
main water flow. (Stanley, 1995) (Fig. 2.1)
Figure 2.1: current flow through Caribbean Sea and Gulf of Mexico (from Carpenter et al. 2002)
The surface temperature of the ocean in the tropical half of the Broad Caribbean
region averages 27˚C with little variation throughout the year. Temperatures in the
southern portion of the Gulf of Mexico also average near 27˚C but the northern Gulf of
Mexico has larger temperature fluctuations due to seasonal changes: from 16˚C in winter

18
to 28˚C in summer (Stanley, 1995). Salinity is relatively high between January and May
(36.39) and lower between June and December in the surface waters of the region (36.09)
(Tomczak & Godfrey, 1994), due mainly to the inflow in fall of lower-salinity waters
from the Orinoco and Amazon Rivers. Geologically, the Caribbean Sea consists of two
main basins separated by a broad, submarine plateau. The Cayman Trench, a trench
between Cuba and Jamaica, contains the Caribbean’s deepest point (7,535 m) and divides
two tectonic plates (Stanley, 1995).
The Cephalopods
Fewer than 1000 species of living cephalopods have been described worldwide;
over 720 are listed in the present catalogues (Jereb et al., 2005). Cephalopods occur in all
marine habitats, though none are found at salinities less than 17.5. Their depth range
extends from the intertidal to over 5,000 m. Due to their accessibility, many of the near
surface and coastal cephalopod species of the Greater Caribbean have been studied in
detail (Voss 1956, 1973; LaRoe, 1967; Lipka, 1975; Passarella, 1990). Deep-sea species
are more difficult to study because of net avoidance and other escape tactics by the
cephalopods (Passarella, 1990). A diverse cephalopod fauna is associated with the
bottom in both shallow and deep-seas.
Biogeography of the Region
Briggs (1995) divided the Broad Caribbean into four distinct regions: the
Brazilian Province, the Caribbean Province, the West Indian Province and the Carolina
Region. The Brazilian Province occupies the area between the Orinoco delta and Cape
Frio which has a distinct biota because of the habitat change. Almost the entire
northeastern coast of South America is virtually devoid of coral reefs and there are vast

19
stretches of mud bottom. Near the mouths of the rivers, salinity is greatly reduced
(Briggs, 1995).
The West Indian Province includes an extensive geographic area and consists
entirely of islands. Bermuda is an isolated northern outpost while the main portion is an
archipelago stretching from the Bahamas to Grenada. Recognition of a West Indian
Province means that the Straits of Florida- only 80 km wide, is an important barrier to the
dispersal of the marine shore biota. It seems clear that the barrier results not from
distance, but from the fast-flowing Florida Current. The Yucatan Channel is only 180km
wide and the passage between Grenada and Trinidad is about 100km wide (Briggs,
1995). The Caribbean Province extends southward from Cape Canaveral in eastern
Florida, Cape Romano in western Florida, and Cape Rojo, Mexico. From those points, it
follows the shore all the way to the northern edge of the Orinoco River delta in
Venezuela. The west coast of Florida supports a complex biotic assemblage. From north
to south, warm-temperate species reach their range limits and tropical species make their
appearance at various points. There is also considerable faunal change with depth, the
tropical species being more numerous on the outer edges of the shelf. The northern Gulf
of Mexico is included as part of the Carolina Region. Within this region, the warm-
temperature biota occupies an area north of the tropical boundaries at Cape Romano, Fl,
and Cape Rojo, Mexico (Briggs, 1995). Examples of Broad Caribbean cephalopod
regional locations (Table 2.1) are as follows:

20
Table 2.1: Examples of regional cephalopod locations (Clarke, 1996)
Region: Species:
The western central Atlantic (WCA) includes the world’s greatest concentration
of small countries including the full range of the world’s major political systems. All of
the Caribbean Sea and nearly all of the Gulf of Mexico are included within one or another
of the region’s 42 jurisdictional units, the largest number found in any ocean area of this
size. When the EEZ’s are compared to the geographic and ecological features of the
same area, it becomes clear that the countries of the region are faced with managing the
biological outcomes of oceanic and ecological processes that operate on a scale that is far
larger than any of the region’s individual management units (Smith et al., 2002).
Four commercially important squid species occur in the Caribbean: longfin
squid (Doryteuthis pealeii), arrow squid (Doryteuthis plei), brief squid (Lolliguncula
brevis), and southern shortfin squid (Illex coindeti). The sharptail shortfin squid (Illex
oxygonius) is found as well in the Caribbean region but is often unrecognized in the
catches. Octopus maya is a commercially important octopod species. It is important to
accurately identify and to determine the distribution of commercially harvested species.
Voss studied the seasonal distribution of the commercially harvested species and found
Inshore coastal region including continental shelf
Octopodidae- rocky or coral shores Sepiolids- sand or mud Loliginidae Seasonally- Ommastrephidae
Off slopes and island slopes
spawning aggregations
Surface water offshore Onychoteuthids, Argonautids, Ommastrephids
Midwater Histioteuthids and Enoploteuthids

21
that their distribution showed a relationship between squid occurrence and temperature,
bottom topography, and areas of high productivity (Voss & Brakoniecki, 1985).
Abundance of Cephalopods
Today, cephalopods are important in neritic waters although numerically they
only constitute a small part of the shelf fauna. In most nearshore regions they are
outnumbered by fish of similar size, except during certain seasons and in some localities.
In oceanic waters they are more diverse in size and play an important role in food webs
(Clarke, 1996).
Abundance of cephalopods varies (depending on group, habitat, and season) from
isolated territorial individuals (primarily benthic octopods) through small schools with a
few dozen individuals, to huge schools with millions of oceanic squids (Vecchione,
2002). Approximately 109 cephalopod species in 31 families occur in the Western
Central Atlantic Ocean and adjacent areas (Caribbean Sea and Gulf of Mexico). Both
decapods and octopods are common in those waters (Vecchione, 2002). Records of
cephalopod species in the Gulf of Mexico date back to Leseur (1821), but the modern
comprehensive systematics of the group begin with Voss, who reported 24 neritic and
oceanic species in 1954, and 42 species in 1956. Since that time, many species have been
added to the list (Passarella, 1990).
Ecological Focus
The ecological portion of this paper examines Rapoport’s Rule (RR) by focusing
on small-scale patterns within a region that had been described as an ecotone. Stevens
(1989) proposed that the greater biodiversity often seen in the tropics is explained by the
fact that tropical species have very narrow ranges while at higher latitudes there is a

22
higher proportion of species with wider ranges (Stevens, 1989). He explained the pattern
on the basis of differing tolerances of tropical and temperate species to climatic
variations. Organisms inhabiting lower latitudes are subject to less variation in climate,
and therefore their geographical distributions tend to be limited to a narrow climatic
range. Higher latitude species would be adapted to more marked climate variation
(Fortes, 2004).
Biodiversity for the purpose of this study is defined as species richness- numbers
of species per area examined. The species richness correlation is found in all higher taxa
whose geographical ranges are well known, both terrestrial and marine. Rapoport (1982)
had noted that the latitudinal ranges of individual species became smaller in lower
latitudes. Thus more species could be accommodated at lower latitudes because each
required less space.
Many studies have tested RR and the outcomes have been mixed. Some authors
(Steele, 1988, Macpherson, 2002; Roy et al. 1998) have found evidence in their studies
supporting RR while others (Clarke, 1992; Mokievsky & Azovsky, 2002) failed to find
such a relationship (Rosa, et al. 2008). Fortes (2004) examined selected bivalves and
gastropods along the Pacific and Atlantic coasts of the Americas and attempted to
evaluate the applicability of Rapoport’s rule in those regions; he concluded that
Rapoport’s Rule does apply in those cases (Fortes, 2004).
Other studies, such as one conducted by Rohde (1992), suggests that Rapoport’s
Rule does not apply to all taxonomic categories. The study focused on marine teleosts
using data collected from the Indo-Pacific and Atlantic oceans. Rohde (1992) found that
RR does not apply in all range areas within a taxon’s range and that it may be premature

23
to explain greater species numbers by narrower environmental tolerances of tropical
species (Rohde, 1992). The biogeographical pattern proposed by Stevens (1989) has
acquired increasing importance among researchers as an explanation for the biodiversity
gradient related to latitude (Fortes, 2004). Many studies have utilized a large latitudinal
range (ie. 80˚N to70˚S) for analysis while this study will examine a narrow range, 22˚ of
latitude, for comparison purposes. Understanding the application of Rapoport’s rule may
be essential for conservation and management (Fortes, 2004).
The systematics of cephalopods are rapidly changing, as research has increased in
the past 25 years. Phylogenetic relationships among families within the major groups
remain uncertain (Vecchione, 2002). Species that have been collected in the Caribbean
allow the opportunity to further describe the cephalopod assemblage as a whole as well as
addressing the ecological importance of the cephalopods within the region.
The goals of this study were:
1. To examine Rapoport’s Rule within the latitudinal range of 8˚ N to 30˚ N in the Broad Caribbean, looking for an increase in cephalopod diversity at lower latitudes following Stevens (1989) original predictions for RR.
2. To compare the cephalopod species richness to that of other studies
conducted (Smith et al., 2002). There are species “hot spots” reported for portions of the Broad Caribbean using a wide range of vertebrate and invertebrate species. The present study will address how the cephalopods fit into that picture.
3. To determine the species composition, distribution and abundance of
cephalopod species within the Broad Caribbean region. The specimens examined come from a variety of preserved materials from various institutions within the region. The study will expand on Voss’s work (1956, 1973, 1985) to include all cephalopod species and their importance in terms of abundance, distribution, and ecology.

24
Study Area:
The Broad Caribbean region includes the Bahama Islands, Gulf of Mexico, and the
Caribbean Sea. The range for the present study is between 8 °-30° N and 60°– 95° W
(Figure 2.2).
Figure 2.2: Study Area
2.2 Materials and Methods
The present study used previously collected specimens, both identified and
unidentified, to determine the distribution of all cephalopods in the Broad Caribbean
region. The most reliable taxonomic information comes from examination of specimens,
in conjunction with, but not limited to, data compiled from sources in the literature. The
most helpful taxonomic studies include examination of comparative material from a
variety of locations, including specimens from the original type locality when possible
(Vecchione and Collette, 2000). Based on current cephalopod dichotomous keys, type
specimens, literature, and expert opinions, the specimens analyzed were identified to the

25
species level. Once a specimen was identified, it was plotted on a distribution map of the
Broad Caribbean region using the ArcView 9.2 mapping program.
The present study included 4190 specimens that were collected from known
locations, and 1000 specimens collected within less-specific regional locations within the
Broad Caribbean. Twenty coastal regions were arbitrarily assigned for inclusion into the
dataset (Appendix A). Many of the regional specimens were from the Caribbean Sea and
were important to include. The regional data were used in the analysis of the
distributions of cephalopods but not included for the application of RR or species
richness analysis.
Identification guides (Voss 1956; Roper and Young, 1984; Nesis 1987;
Vecchione, 2002) were used to identify each organism accurately to the species level.
The majority of specimens examined had been preserved well and were in excellent
condition. The specimens dated from 1898 to present. The bulk of preserved specimens
analyzed were from two institutions, the Smithsonian Institution’s National Museum of
Natural History and the University of Miami Rosenstiel School of Marine and
Atmospheric Science. Smaller collections from the Florida Fish and Wildlife Research
Institute were also analyzed. The Hourglass cruises were conducted from 1965-1967 in
the shelf waters off west central Florida where cephalopod species were identified and
documented by Dr. Ronald Toll and Dr. Steven Hess. Approximately 500 specimens
were included in the present study from those cruises.
Identification of specimens was conducted in the micronekton museum at the
College of Marine Science at the University of South Florida, the Rosenstiel School for
Marine and Atmospheric Science, University of Miami as well as the Smithsonian

26
Institution’s National Museum of Natural History. A dissecting microscope was used to
identify small specimens and specific features of the animals. Micrometer calipers were
used for length measurements.
The methods used for identifying and cataloguing specimens were to identify
each specimen using the cephalopod dichotomous keys and then to record its mantle
length, sex, number of gill lamellae (octopods), and species name into an excel file for
statistical analysis. Once all the organisms were identified, known specimens from all
institutions were also added to the excel file for analysis. Distribution was calculated
using the 9.2 ArcView software program.
Statistical Analysis
Rapoport’s Rule (RR) was evaluated by using species richness, which is defined
here as species number per 5˚ latitudinal bin within the study’s scope (5 bands). This was
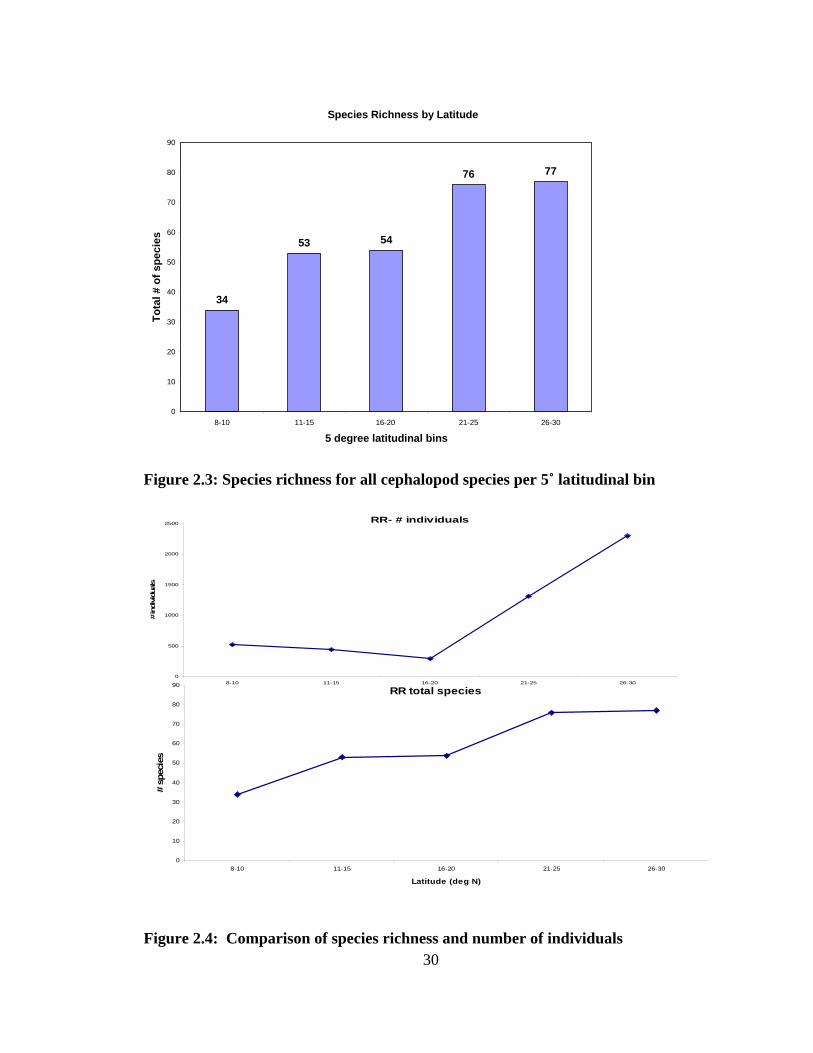
plotted as species richness versus latitude (Fig. 2.3). Figure 2.4 represents the two
variables addressed when comparing the 5 degree latitudinal bins- the number of species,
as well as total number of individuals used for calculations in each bin.
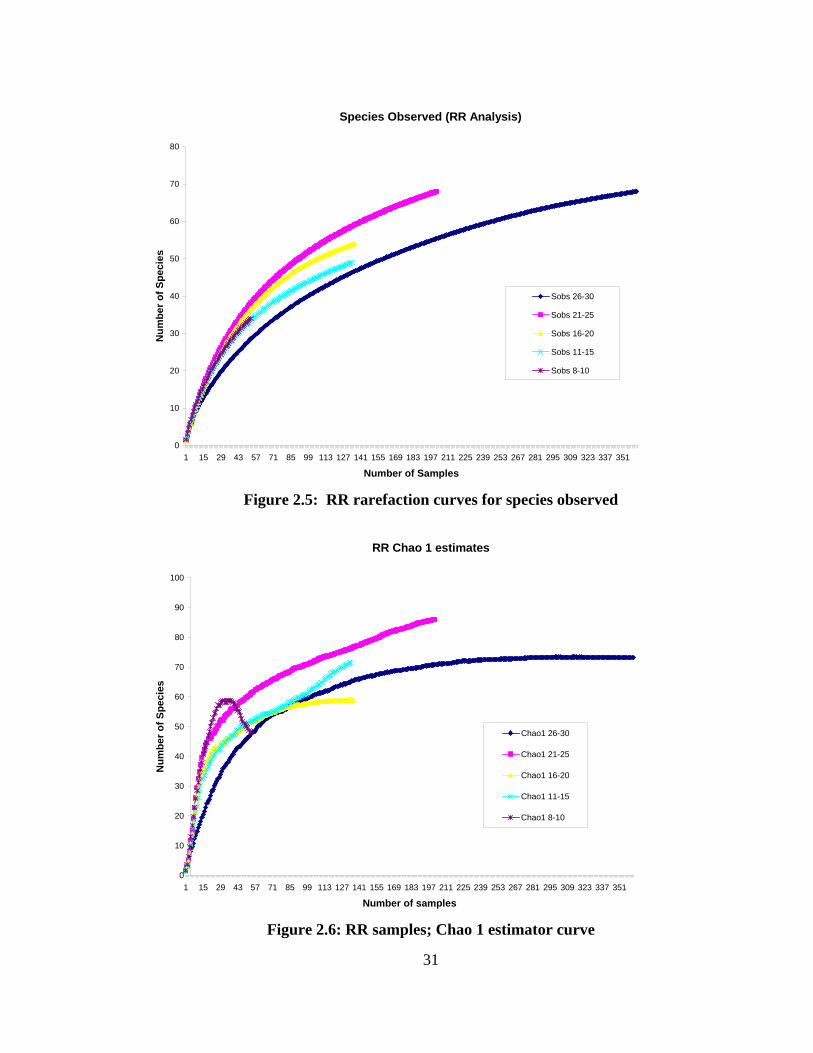
Rarefaction curves were created for all 5 latitudinal bands using Primer 6.2 for
estimates and graphic output (Fig. 2.5, 2.6). Rarefaction is a tool used to correct for
unbalanced sampling structure. The rarefaction curve is produced by repeatedly re-
sampling the pool of N individuals or N samples, at random, plotting the average number
of species represented by 1,2,….N individuals or samples (Gotelli & Colwell, 2001). It is
dependent on the shape of the species abundance curve rather than the absolute number
of specimens per sample. This method is valid when the same groups of organisms are
being compared and contrasted. Another requisite is that all the habitats sampled be

27
similar, in this case, coastal habitats. Methods of capture should be similar and lastly,
this method does not specify which species taken from the residue will be present, and it
can only be used to interpolate (Sanders, 1968). The rarefaction curve (Fig. 2.5) is
species observed (Sobs) compared to latitude. A second set of curves (Fig. 2.6) is the
Chao 1 estimator curve which gives a most likely total species estimate for each region
based on the actual sample provided. Anne Chao devised a non-parametric estimator
used for species richness comparisons. It is based on the number of rare species in a
sample and creates an estimate of total species for a region (Magurran, 2004). The Sobs
graph accounts for sample-size differences while the Chao 1 estimator gives an absolute
number for species richness in a region.
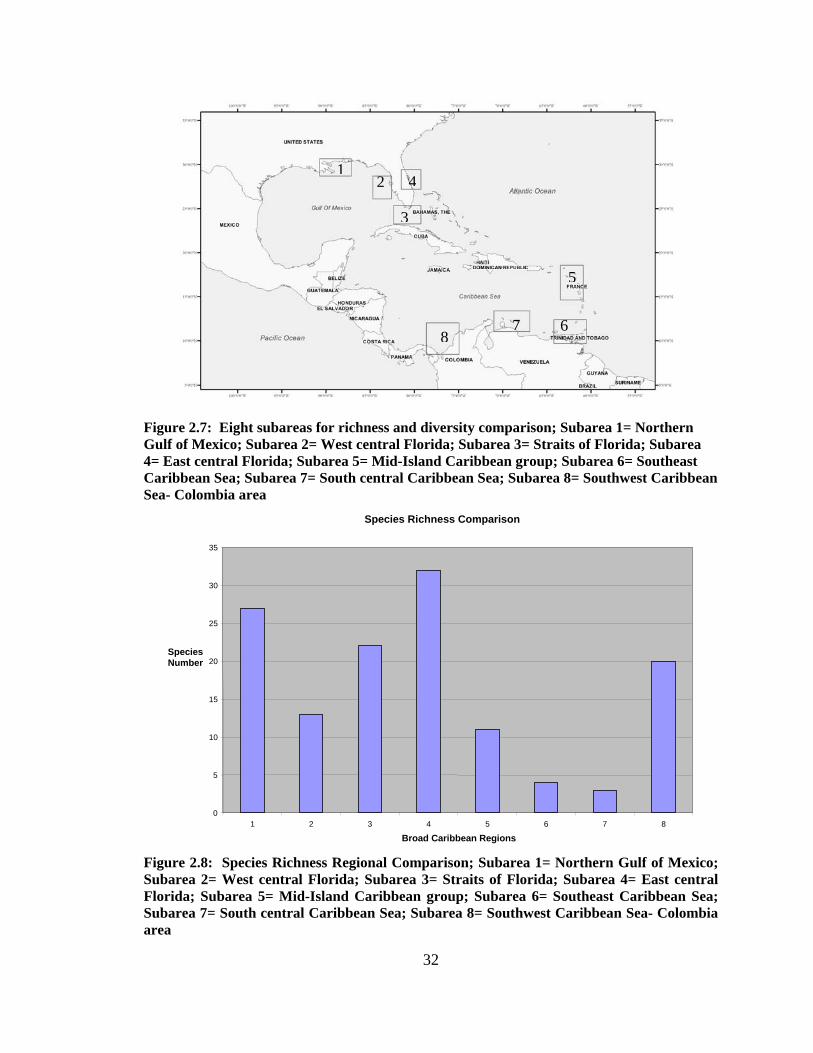
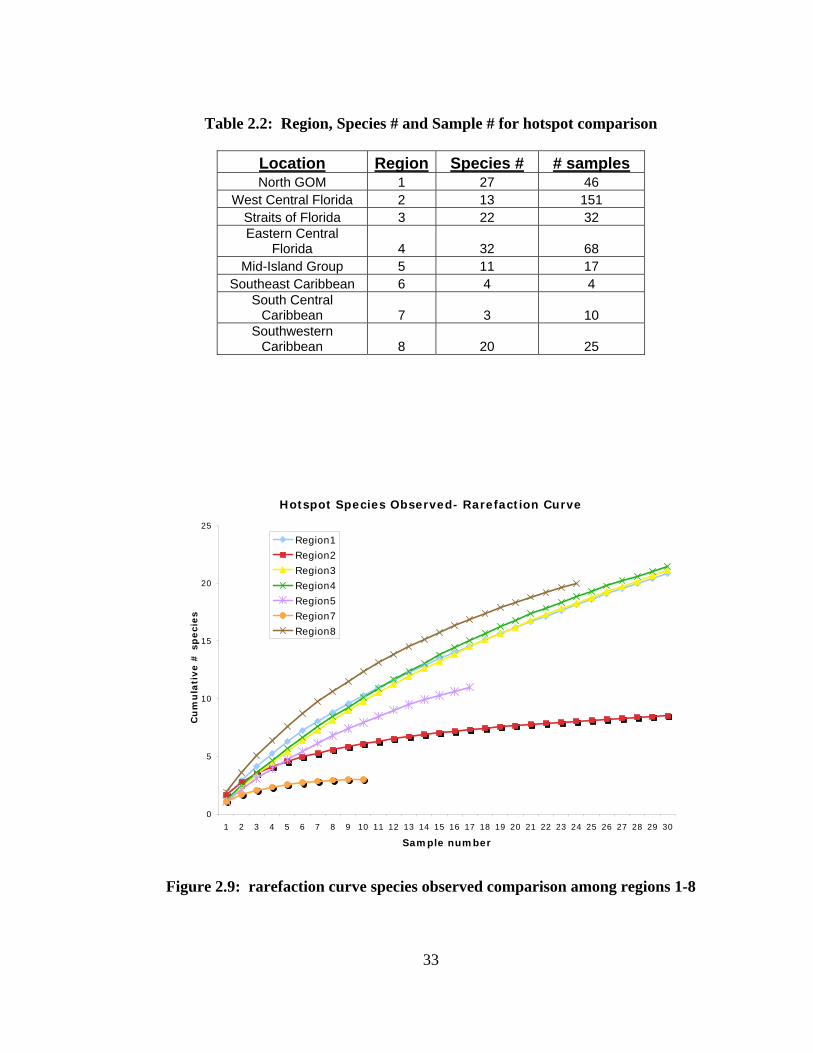
Hotspots were calculated using an excel spreadsheet for comparison of species
numbers in 8 subareas (Figs. 2.7, 2.8). The 8 sites were chosen by location in the Broad
Caribbean which also correlates with a species richness study conducted by Smith et al.
(2002). The 8 subarea coordinates are in Appendix B. Rarefaction curves for the sites
were created using Primer 6.2 for comparison of the species observed and the Chao 1
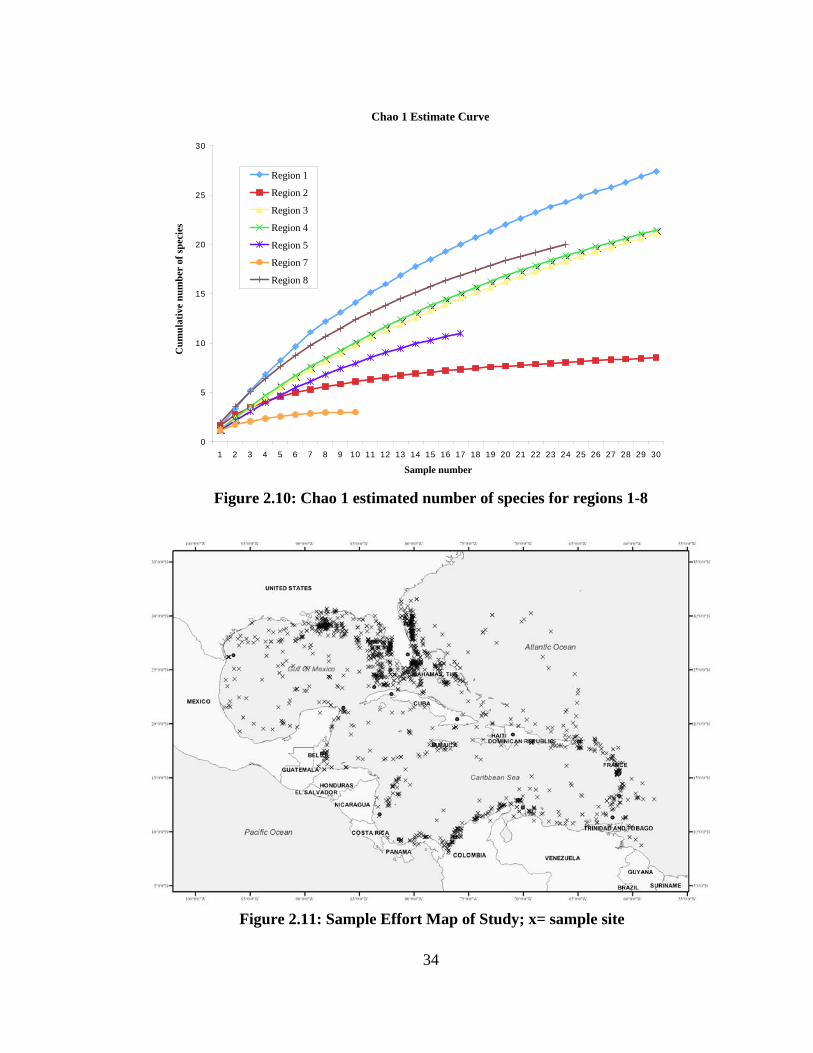
estimator (Fig. 2.9, 2.10).
2.3 Results Rapoport’s Rule and Species Richness
Species richness in the Broad Caribbean showed an increase with increasing
latitude (Fig. 2.3). The 8-10˚ band showed the lowest species richness, 34 species,
gradually increasing up through the highest latitudes to the 26-30˚ band with 77 species
represented. Figure 2.4 compares the number of individual cephalopods examined to the
number of species found in each latitudinal band. There was an increase in species found

28
per band while the number of individuals varied among bands. Latitudinal band 11-16˚
N had a decrease in individuals examined while species richness was increasing, which
indicates that sampling effort was not the sole factor for species richness increasing
northward. The species observed (Fig. 2.5) rarefaction curve shows a trend for all
latitudinal bands headed towards asymptote but only the higher latitudes close in on
approaching it. The Chao 1 estimator curve (Fig. 2.6) represents the expected number of
species found in each band.
Cephalopod Biogeography
Eight subareas (Fig. 2.7) were chosen to compare species richness within the
Broad Caribbean. Each of the regions incorporated features of biogeographical
significance; e.g. important current patterns or seafloor features, or exhibited potential as
human management areas. These subareas correspond to a biogeography study of
various marine organisms compared by Smith et al. (2002) analyzing invertebrates and
vertebrates for potential diversity hotspots in the region.
Figure 2.8 compares the 8 subareas and the number of species per region. It
shows that subarea 4 (Eastern central Florida) has the highest species richness (n=32),
followed by subarea 1, northern Gulf of Mexico (n=27), subarea 3 in the Straits of
Florida (n=22), subarea 8 in the southwestern Caribbean Sea (n=20), subarea 2 in the
West central Florida waters (n=13), subarea 5 with 11 different species, subarea 6 with 4
species, and lastly, subarea 7 with only 3 species types collected. Rarefaction curves
were used to determine the expected number of species per sample site as a function of
organisms sampled. Table 2.2 is the number of samples and species analyzed for the bar
graph analysis.

29
Figures 2.9 and 2.10 show the rarefaction curves derived for each region.
Subregion 6 was not included due to the low number of samples (n=4). The trendline is
similar with differences between subareas in close proximity of one another. Subarea 7
and 8 are both in the lower Caribbean Sea yet show a large difference in both species
observed and expected species. Another example of difference in species richness is
between subareas 2 and 4. Subarea 2, the eastern central Gulf of Mexico has a much
lower number of species observed and expected number of species than subarea 4 of
approximately the same latitude.
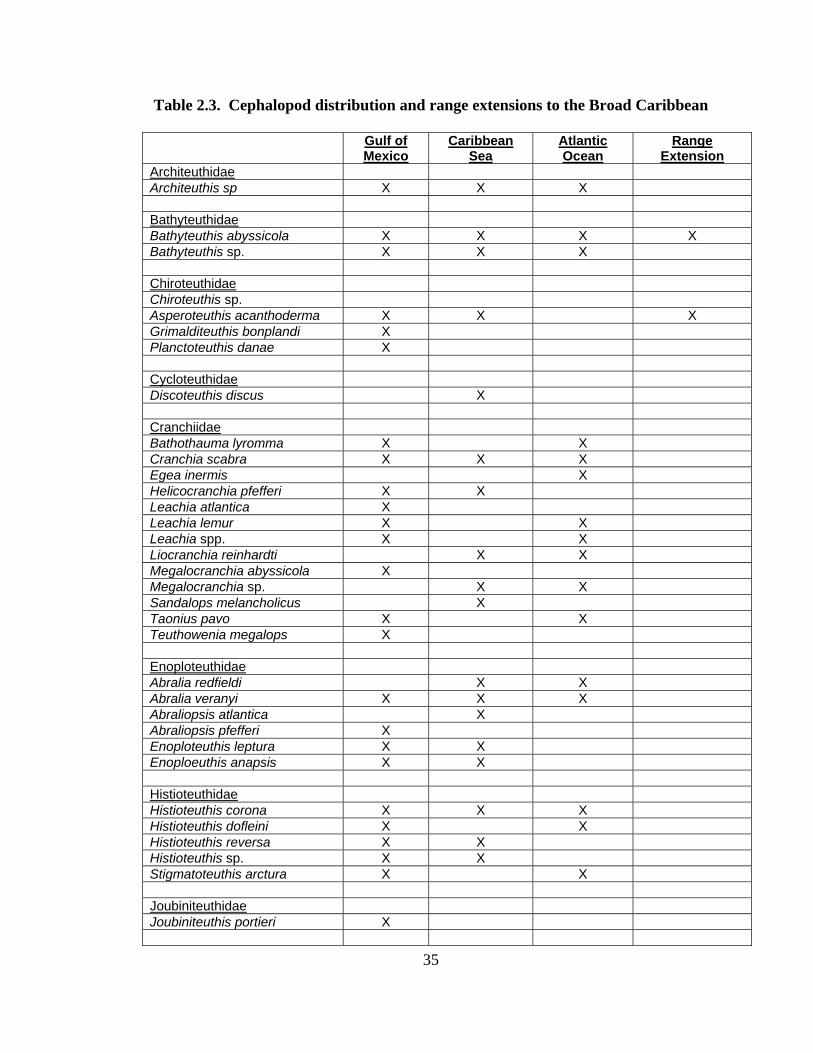
Distribution and abundance of Broad Caribbean Cephalopods Distribution maps for each cephalopod species found within the Broad Caribbean
are in Appendix C. Species maps are in order by family groups in most cases. All 5190
specimens were located within the distributional effort map shown in Figure 2.11. The
figure represents all of the known coordinate sampling sites for specimens as well as the
20 regions created to define areas throughout the Broad Caribbean, including the 1000
specimens that were collected with a regional location but lacked precise latitude and
longitude coordinates (Appendix A).
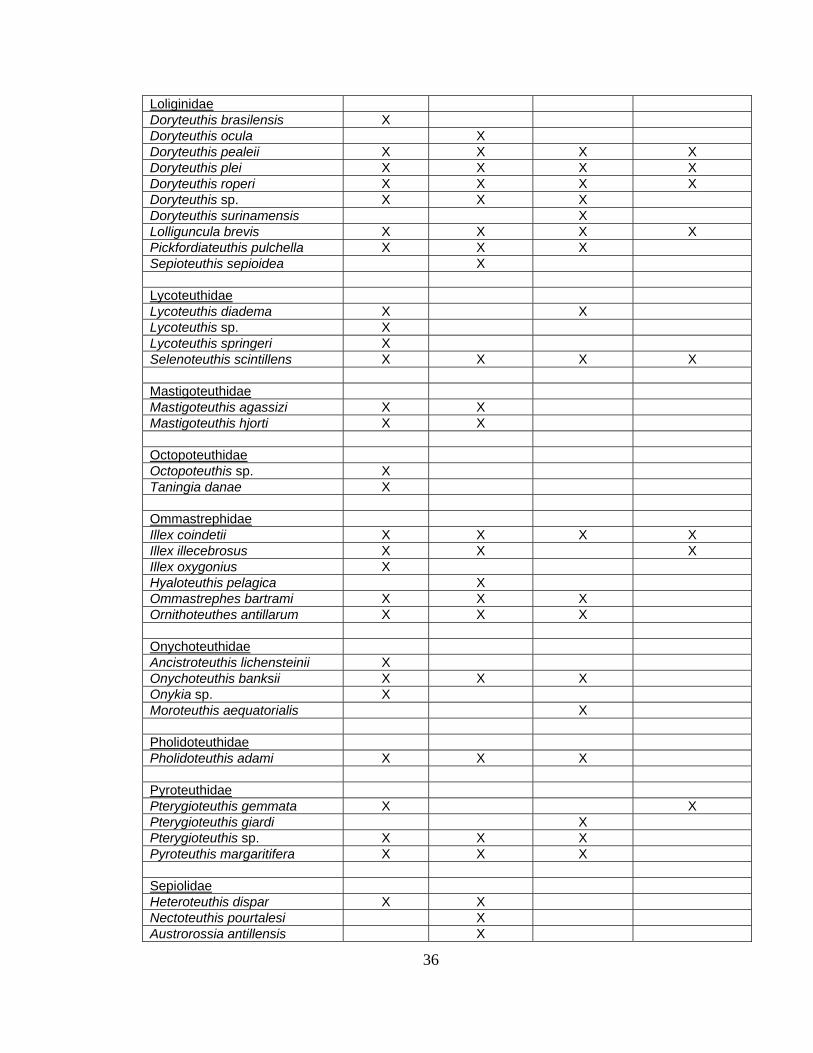
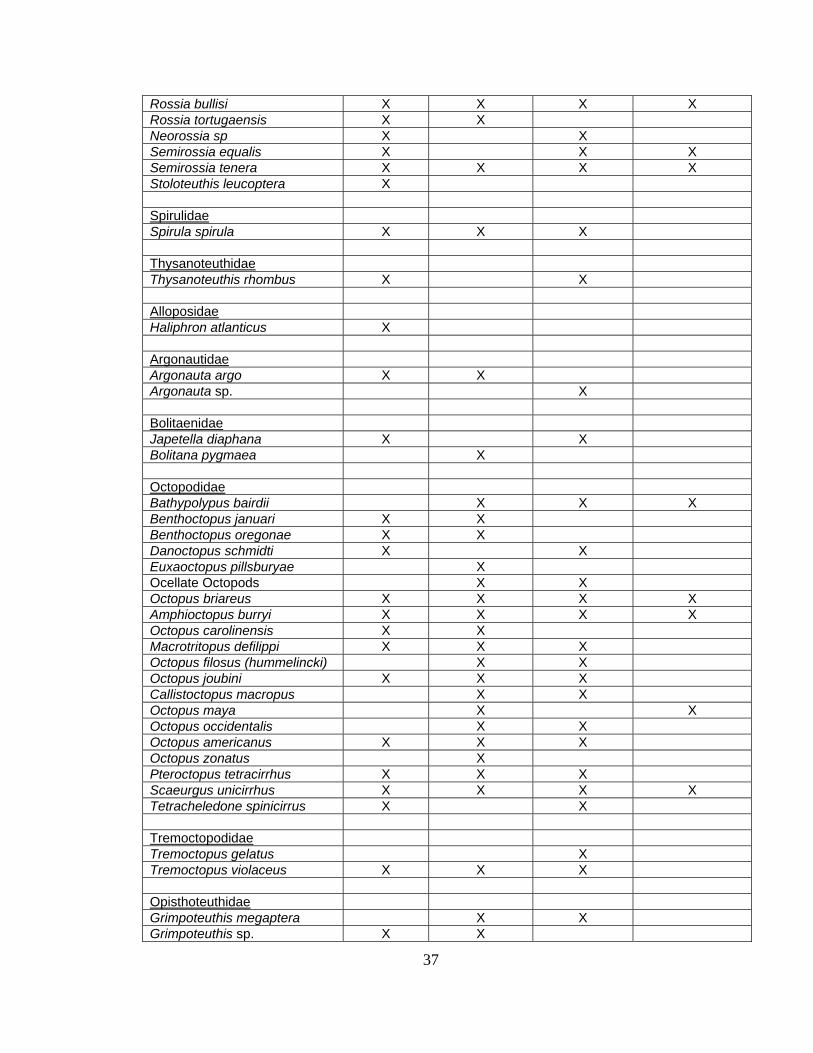
Cephalopod Range Extensions
Range extensions for several species of cephalopods in the Broad Caribbean
region were observed based on the dataset collected in the present study. Table 2.3
shows 18 extensions to previously known ranges based on the work of Roper and
Sweeney, 1984; Nesis, 1987; and Vecchione, 2002.
30
Species Richness by Latitude
34
53 54
76 77
0
10
20
30
40
50
60
70
80
90
8-10 11-15 16-20 21-25 26-30
5 degree latitudinal bins
Tota
l # o
f spe
cies
Figure 2.3: Species richness for all cephalopod species per 5˚ latitudinal bin
Figure 2.4: Comparison of species richness and number of individuals
RR total species
0
10
20
30
40
50
60
70
80
90
8-10 11-15 16-20 21-25 26-30
Latitude (deg N)
# sp
ecie
s
RR- # individuals
0
500
1000
1500
2000
2500
8-10 11-15 16-20 21-25 26-30
# in
divi
dual
s
31
Species Observed (RR Analysis)
0
10
20
30
40
50
60
70
80
1 15 29 43 57 71 85 99 113 127 141 155 169 183 197 211 225 239 253 267 281 295 309 323 337 351
Number of Samples
Num
ber o
f Spe
cies
Sobs 26-30
Sobs 21-25
Sobs 16-20
Sobs 11-15
Sobs 8-10
Figure 2.5: RR rarefaction curves for species observed
RR Chao 1 estimates
0
10
20
30
40
50
60
70
80
90
100
1 15 29 43 57 71 85 99 113 127 141 155 169 183 197 211 225 239 253 267 281 295 309 323 337 351
Number of samples
Num
ber o
f Spe
cies
Chao1 26-30
Chao1 21-25
Chao1 16-20
Chao1 11-15
Chao1 8-10
Figure 2.6: RR samples; Chao 1 estimator curve
32
Figure 2.7: Eight subareas for richness and diversity comparison; Subarea 1= Northern Gulf of Mexico; Subarea 2= West central Florida; Subarea 3= Straits of Florida; Subarea 4= East central Florida; Subarea 5= Mid-Island Caribbean group; Subarea 6= Southeast Caribbean Sea; Subarea 7= South central Caribbean Sea; Subarea 8= Southwest Caribbean Sea- Colombia area
Figure 2.8: Species Richness Regional Comparison; Subarea 1= Northern Gulf of Mexico; Subarea 2= West central Florida; Subarea 3= Straits of Florida; Subarea 4= East central Florida; Subarea 5= Mid-Island Caribbean group; Subarea 6= Southeast Caribbean Sea; Subarea 7= South central Caribbean Sea; Subarea 8= Southwest Caribbean Sea- Colombia area
Species Richness Comparison
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7 8
Broad Caribbean Regions
Species Number
12
3
4
5
6 78
33
Table 2.2: Region, Species # and Sample # for hotspot comparison
Location Region Species # # samples North GOM 1 27 46
West Central Florida 2 13 151 Straits of Florida 3 22 32 Eastern Central
Florida 4 32 68 Mid-Island Group 5 11 17
Southeast Caribbean 6 4 4 South Central
Caribbean 7 3 10 Southwestern
Caribbean 8 20 25
Hotspot Species Observed- Rarefaction Curve
0
5
10
15
20
25
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Sample number
Cu
mu
lati
ve #
sp
eci
es
Region1Region2Region3Region4Region5Region7Region8
Figure 2.9: rarefaction curve species observed comparison among regions 1-8
34
Chao 1 Estimate Curve
0
5
10
15
20
25
30
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
Sample number
Cum
ulat
ive
num
ber
of sp
ecie
sRegion 1
Region 2
Region 3
Region 4
Region 5
Region 7
Region 8
Figure 2.10: Chao 1 estimated number of species for regions 1-8
Figure 2.11: Sample Effort Map of Study; x= sample site
35
Table 2.3. Cephalopod distribution and range extensions to the Broad Caribbean
Gulf of Mexico
Caribbean Sea
Atlantic Ocean
Range Extension
Architeuthidae Architeuthis sp X X X Bathyteuthidae Bathyteuthis abyssicola X X X X Bathyteuthis sp. X X X Chiroteuthidae Chiroteuthis sp. Asperoteuthis acanthoderma X X X Grimalditeuthis bonplandi X Planctoteuthis danae X Cycloteuthidae Discoteuthis discus X Cranchiidae Bathothauma lyromma X X Cranchia scabra X X X Egea inermis X Helicocranchia pfefferi X X Leachia atlantica X Leachia lemur X X Leachia spp. X X Liocranchia reinhardti X X Megalocranchia abyssicola X Megalocranchia sp. X X Sandalops melancholicus X Taonius pavo X X Teuthowenia megalops X Enoploteuthidae Abralia redfieldi X X Abralia veranyi X X X Abraliopsis atlantica X Abraliopsis pfefferi X Enoploteuthis leptura X X Enoploeuthis anapsis X X Histioteuthidae Histioteuthis corona X X X Histioteuthis dofleini X X Histioteuthis reversa X X Histioteuthis sp. X X Stigmatoteuthis arctura X X Joubiniteuthidae Joubiniteuthis portieri X
36
Loliginidae Doryteuthis brasilensis X Doryteuthis ocula X Doryteuthis pealeii X X X X Doryteuthis plei X X X X Doryteuthis roperi X X X X Doryteuthis sp. X X X Doryteuthis surinamensis X Lolliguncula brevis X X X X Pickfordiateuthis pulchella X X X Sepioteuthis sepioidea X Lycoteuthidae Lycoteuthis diadema X X Lycoteuthis sp. X Lycoteuthis springeri X Selenoteuthis scintillens X X X X Mastigoteuthidae Mastigoteuthis agassizi X X Mastigoteuthis hjorti X X Octopoteuthidae Octopoteuthis sp. X Taningia danae X Ommastrephidae Illex coindetii X X X X Illex illecebrosus X X X Illex oxygonius X Hyaloteuthis pelagica X Ommastrephes bartrami X X X Ornithoteuthes antillarum X X X Onychoteuthidae Ancistroteuthis lichensteinii X Onychoteuthis banksii X X X Onykia sp. X Moroteuthis aequatorialis X Pholidoteuthidae Pholidoteuthis adami X X X Pyroteuthidae Pterygioteuthis gemmata X X Pterygioteuthis giardi X Pterygioteuthis sp. X X X Pyroteuthis margaritifera X X X
Sepiolidae Heteroteuthis dispar X X Nectoteuthis pourtalesi X Austrorossia antillensis X
37
Rossia bullisi X X X X Rossia tortugaensis X X Neorossia sp X X Semirossia equalis X X X Semirossia tenera X X X X Stoloteuthis leucoptera X Spirulidae Spirula spirula X X X Thysanoteuthidae Thysanoteuthis rhombus X X Alloposidae Haliphron atlanticus X Argonautidae Argonauta argo X X Argonauta sp. X Bolitaenidae Japetella diaphana X X Bolitana pygmaea X Octopodidae Bathypolypus bairdii X X X Benthoctopus januari X X Benthoctopus oregonae X X Danoctopus schmidti X X Euxaoctopus pillsburyae X Ocellate Octopods X X Octopus briareus X X X X Amphioctopus burryi X X X X Octopus carolinensis X X Macrotritopus defilippi X X X Octopus filosus (hummelincki) X X Octopus joubini X X X Callistoctopus macropus X X Octopus maya X X Octopus occidentalis X X Octopus americanus X X X Octopus zonatus X Pteroctopus tetracirrhus X X X Scaeurgus unicirrhus X X X X Tetracheledone spinicirrus X X Tremoctopodidae Tremoctopus gelatus X Tremoctopus violaceus X X X Opisthoteuthidae Grimpoteuthis megaptera X X Grimpoteuthis sp. X X

38
Opisthoteuthis agassizii X X X Vampyroteuthidae Vampyroteuthis infernalis X X X
New Species to Area
Two Asperoteuthis ancanthoderma specimens were examined demonstrating a
very significant range extension to the species; the only other specimens found previously
were off the coasts of Japan. Two nearly intact individuals were found floating dead on
the surface of the ocean and collected: one specimen was found off the coast of Key
West, FL and the other off the coast of Marathon, FL. Both specimens were dissected
and analyzed for identification. It was determined that they were both Asperoteuthis
acanthoderma. A description paper is in press with the Proceedings of the Biological
Society of Washington (Judkins et al. 2009).
2.4 Discussion Rapoport’s Rule and Species Richness
Rapoport’s Rule attributes the many observations of increasing diversity with
decreasing latitude to a reduction in size of species’ distributional ranges as you approach
the equator (Rosa et al., 2008). Stevens (1989) supported his claim with studies of
diverse taxa including North American trees, North American marine molluscs,
freshwater and coastal fishes, reptiles and amphibians.
Since that time, many scientists have studied the ecological patterns driving
biological diversity. There have been numerous hypotheses to explain diversity patterns
(Peet, 1974; Evans, 2005) and various groups of organisms have been examined to prove
or disprove RR. Many studies of marine groups have supported RR (Steele, 1988;

39
Stevens, 1996; Roy et. al. 1998, 2000; Rex et al. 2000, 2005; Macpherson 2002) while
others have failed to find such a relationship (Clarke, 1992; Lambshead et al. 2000;
Mokievsky and Azovsky, 2002; Rosa et al., 2008). The latter studies found a negative
correlation to RR as discussed below.
There have been a few molluscan species richness studies in the Atlantic Ocean to
date (Rosa et al., 2008, Fortes and Absalao, 2004, Macpherson, 2003). Fortes and
Absalao (2004) examined gastropods and bivalves using literature-based studies from
both the Pacific and Atlantic sides of the continental North and South American coasts.
After analyzing 4067 species they determined that RR applied to these organisms on both
coasts. They noted that regional features, such as the size of a biogeographical province,
seemed to affect the pattern strongly. They also found support for RR when they
incorporated depth into the study.
Macpherson (2003) studied the variability in size of species ranges in terms of
depth and latitude for various marine taxa, including cephalopods and fishes in the
Atlantic Ocean. The results showed that RR could hold true for those organisms but was
not solely responsible for latitudinal patterns in range sizes.
The research conducted by Rosa et al. (2008) examined cephalopod species of the
coastal Atlantic Ocean using primary literature, reports, and online databases. Their
results showed that latitudinal gradients of species richness were present along both
Atlantic coasts, but were distinct from one another. When the median latitudinal ranges
of the Western Atlantic neritic cephalopods were determined, it was evident that the size
of the distributional ranges did not decline with decreasing latitude which means that RR
may not explain the distribution patterns. Stevens, (1996) proposed that RR could extend

40
to elevation and water depth in terms of species richness. When species’ depths were
taken into consideration for the organisms in the western Atlantic, RR was exhibited
(Rosa et al. 2008).
The present specimen-based study showed that the cephalopods of the Broad
Caribbean do not exhibit the diversity patterns described originally by Stevens (1989).
Within the small latitudinal range of 8˚ to 30˚ N, cephalopods of the region increase in
species richness with increasing latitude. The lack of concordance with RR in the present
study agrees with Rosa et al. (2008). It should be noted that the lowest latitude band, (8˚-
10˚) includes only 3˚ latitude while the other 4 include 5˚ in each band. Therefore, there
may be more than the 34 species in the region that the present study suggests. However,
the species richness trend is still obvious as latitude increases.
One of the reasons for an increase in species number at higher latitudes could be
the convergence of the Florida current and the North Equatorial current in the middle to
northern end of the study area. The two currents converge to become the Gulf Stream
and may be transporting cephalopods northward. The Florida Current becomes the Gulf
Stream and then leaves the coast of the eastern United States at Cape Hatteras to head
across the Atlantic Ocean. This current has a profound influence on the distribution of
shore animals in the western Atlantic (Briggs, 1995).
Another factor possibly contributing to the northward increase in species richness
is the larger number of studies in the northern Broad Caribbean region. Numerous
studies have been conducted in Florida waters which may be contributing to the increased
richness of cephalopods in the northern portion of the study area. The RR rarefaction
curve (Fig. 2.5) showed a similar richness trend for all regions and pointed out the need

41
for more sampling in the lower latitudes, as the upper 2 bands were close to asymptote
while the lower 3 bands were still rising. The Chao 1 estimator analysis showed
differences among species richness between the bands. Note that the 8˚- 10˚ latitude
band approached 60 species expected and then decreased rapidly (Fig. 2.6). A possible
reason for the decline is that there was a smaller sample size for that latitudinal band in
conjunction with numerous singleton species counted, which caused a decrease in the
species richness curve for that region.
Cephalopod Biogeography Based on range map overlays, Smith et al. (2002) examined the distribution of
1172 vertebrate and invertebrate species and concluded that the area of highest species
richness was located in the waters surrounding southern Florida, the eastern Bahamas,
and northern Cuba. Secondary centers of diversity were located (in descending order of
richness) on the continental shelves of northern South America, Central America, and in
the northern Gulf of Mexico. Those patterns are apparently robust as they are repeated in
composite distributions for fishes and for other invertebrates taken separately (Smith et
al., 2002).
Analyzing the present study’s cephalopod species richness information, eastern
central Florida has the highest species richness in the Broad Caribbean (n=32), likely
because the Gulf Stream acts as a large transporter of larvae from the southern waters
feeding into it. Another possible, but less likely, factor for this subarea’s species richness
is that it is a well-traveled path for seasonally migrating cephalopods.
The cephalopods of the Broad Caribbean generally follow the pattern found by
Smith et al.(2002) in that the two regions exhibiting the highest richness were the same in

42
both studies. However, subarea 6, the southeastern Caribbean Sea, had too few samples
to include in the analysis (n=4). Subarea 7, south central Caribbean Sea, had 10 samples
and was included in the analysis. Smith et al. suggested that the southern edge of South
America was the second richest in terms of species whereas the present study does not
show that trend. The two areas of low sampling effort (6 & 7) over 111 years of
collecting indicate the need for further fieldwork in those regions.
The subarea rarefaction curve displays curious trends within the Broad Caribbean.
It was anticipated that there would be variations in species richness between the major
basins of the Caribbean and the Gulf of Mexico. This does not appear to be the case. For
example, subareas 7 and 8 were both located in the lower Caribbean and yet showed
significantly different trends in species richness (Fig. 2.9). Another example of a
variation within regions can be seen between subareas 2 and 4 in the northern sections of
the Broad Caribbean. A reason for the variation in this case could be the intense work
that was conducted by the Hourglass cruises in the mid 1960’s by the Marine Research
Laboratory of the Florida Board of Conservation, creating an artifact from sampling
efforts. Two experts identified and catalogued cephalopod shelf species which were
included in the present study. Although over 500 specimens were included in subarea 2,
the species richness of the subarea (n=13) was still lower than subareas 3 (n=22) and 4
(n=32). Another explanation for the increase in species richness in subarea 4 could be
the influence of the water depth changes and flow of the Gulf Stream. These two
examples of variation within regions (Gulf of Mexico and Caribbean basin) exceed the
variations between the northern and southern regions of the study’s scope.

43
Distribution and abundance There were 110 cephalopod species from 27 families examined in the present
study. This number is larger than that found in earlier studies (Voss, 1956- 42), but quite
similar to more recent work (Nesis, 1975-100), (Vecchione, 2002-109). This study
examined unidentified and identified specimens that were primarily from two large
institutions.
Broad Caribbean cephalopods are widely distributed, many finding niches within
coastal and island ecosystems. The abundance of organisms differs between species,
with Doryteuthis plei (n=1205) and Doryteuthis pealeii (n=702) being the most numerous
squid. Doryteuthis plei and D. pealeii move in large schools, are commercially harvested,
used in medical studies and are distributed throughout the region. Octopus joubini
(n=351) and Octopus americanus (vulgaris) (n=306) were the most abundant octopods
and are also commercially harvested throughout the region. Over 30 species were
represented by 3 or fewer specimens for the entire region. An example of a lesser known
cephalopod is Discoteuthis discus, found in deep water with little known about its
distribution or biology (Vecchione, 2002). Euaxoctopus pillsburyae inhabit small niches
surrounding a particular island group and only 1 specimen was recorded for this study.
Few deep-sea cephalopods have been collected, indicating the need for increased research
efforts in the area to uncover other unique organisms below 500m with regularity.
Few studies of cephalopod distribution have been reported in context for the
Broad Caribbean area. Two studies that focused on the region were completed by
Barrientos and Garcia-Cubas (1997) and Vecchione et al. (2001). Barrientos focused on
three loliginids, Doryteuthis (Loligo) plei, Doryteuthis (Loligo) pealeii, and Lolliguncula

44
brevis in the western Gulf of Mexico. The authors associated squid abundance and
distribution with currents, warm layers, fronts, water masses, and climate changes
(Barrientos and Garcia-Cubas, 1997). These results are concordant with the current
findings as D. plei were most abundant (n=1205) followed by D. pealeii (n=702) with L.
brevis (n=374) rounding out the top three most abundant cephalopods in this study.
Another study (Vecchione et al., 2001) centered on paralarval cephalopod
distribution and abundance in the western North Atlantic Ocean. The two most abundant
and frequently collected species were the neritic squids Doryteuthis pealeii and Illex
illecebrosus collected north of Cape Hatteras, both valuable fishery resources. The
highest abundance and diversity of planktonic cephalopods in the oceanic samples were
consistently found in the vicinity of the Gulf Stream. According to Vecchione (2001) the
most likely species other than D. pealeii to be present in the south is D. plei (Vecchione,
1981). Other Doryteuthis species such as D. ocula are restricted to Caribbean islands
whereas Doryteuthis roperi, once considered a Caribbean species, had a range extension
into the Gulf of Mexico and Atlantic Ocean, demonstrated in the present study.
Doryteuthis pealeii was abundant in the Broad Caribbean in the present study as
well as in Vecchiones’ work while Illex illecebrosus was only present in small numbers
(n=6). Another Illex species, I. coindetii was more abundant in the Gulf of Mexico and
the northern Caribbean (n=82) which suggests that it is better suited for tropical
temperatures than Illex illecebrosus. Findings here differ from Vecchiones’(2001) work
in that D. roperi were abundant (n=89) and found not only in the Caribbean but were
distributed through the Straits of Florida and up the Gulf Stream to the northernmost
region of the study. D. ocula is confined to the Caribbean as the 3 specimens revealed in

45
this study. It should be noted that there seem to be fewer species found in the western
regions of both the Gulf of Mexico and the Caribbean Sea. This could be attributed to
low sampling effort (Fig. 2.11).
Range Extensions The observed range extensions are extremely valuable. There have been recent
studies conducted (Rosa et al., 2008; Judkins et. al 2008) that compare organisms of the
region based on literature sources so to find 18 new extensions based on fresh
identification is helpful in studying the biology and habits of the organisms. It is also
important as the hunt for new fishery resources increases worldwide.
New Species to the Area
It is not surprising to find a new species in the Broad Caribbean as research
funding has decreased and cephalopod studies have been limited in past years. The
species described, Asperoteuthis acanthoderma, was found prior to this only in the West
Pacific Ocean. More studies targeting deep water species are needed to determine the
role these large and important species play in the oceans food webs as well as learning
their biology and habits to better understand cephalopod adaptations.
To summarize, the present study found that 110 cephalopod species of the Broad
Caribbean are dispersed throughout its waters, they occupy a myriad of niches along
coastlines and deep water, and there is no support for RR. Hotspots are patchy with the
most species richness occurring along the eastern edge of Florida in the Gulf Stream.
The range extensions based on data important to note for future conservation and fishery
ventures. Asperoteuthis acanthoderma is a fortunate discovery and begs the question

46
“what else is down there”? Areas of the southeastern Caribbean as well as the western
Gulf of Mexico appear to be understudied.
The present study is the first to examine the specimen-based species richness area
with rarefaction curves to support it, attempting to determine species richness of the
Broad Caribbean region. The study size range is unique in that it is not encompassing
numerous latitudes but focusing on 22˚ of a tropical/subtropical area important to a
variety of species. Studies surrounding all aspects of cephalopod life- diversity, biology,
ecology, and capture methods would improve the world database for these important
organisms.

47
REFERENCES
1. Barrientos, Gerardo, Antonio Garcia-Cubas. 1997. Disstribution and abundance of the Family Loliginidae (Mollusca: Cephalopoda) in Mexican waters of the Gulf of Mexico. Rev. Soc. Mex. Hist. Nat.; 47: 123-139.
2. Briggs, John C. 1995. Global Biogeography; Elsevier Science B.V.; 454 pp. 3. Cairns, S. 1976. Cephalopods collected in the Straits of Florida by the R/V Gerda.
Bulletin of Marine Science; 26(2): 233-272. 4. Clarke, A. 1992. Is there a latitudinal diversity cline in the sea? Trends in Ecology
and Evolution; 7: 286-287. 5. Clarke, M.R.1996. The Role of Cephalopods in the world’s oceans: general
conclusions and the future. Phil. Trans. R. Society London. B.; 351: 1105-1112. ------1996. The role of cephalopods in the world’s oceans: an introduction. Phil. Trans.
R. Society London. B.; 351: 979-983. 6. Diaz, J.M. 2004. First Record of an octopus species if the family Tremoctopodidae
(Cephalopda: Octopoda) for the Colombian Caribbean. Biol. Invest. Mar. Cost. 33; 283-286.
7. Diaz, J.M. and Gracia, N. 2000. Squids and octopuses (Mollusca: Cephalopoda) of the Colombian Sea. Biota Colombiana; 1(2) 195-201. 8. Fortes, R.R., R.S. Absalao, 2004. The applicability of Rapoport’s rule to the marine
molluscs of the Americas. Journal of Biogeography; 31: 1909-1916. 9. Gotelli, Nicholas, Robert K. Colwell. 2004. Quantifying biodiversity: procedures,
and pitfalls in the measurement and comparison of species richness. Ecology Letters; 4: 379-391.
10. Gracia, A. 2002. Cephalopods (Mollusca: Cephalopoda) of the Upper Colombian
Caribbean Shelf. Biol. Invest. Mar. Cos. 31, 219-238. 11. Jereb, P. and Roper, C.F.E. 2005. Cephalopods of the world. An annotated and
illustrated catalogue of cephalopod species known to date. FAO species catalogue for fisheries purposes, N4, v.1, FAO, Rome.
12. Judkins, H, M. Veccione, & C.F.E. Roper. In press. Cephalopods (Mollusca:
Cephalopoda) of the Gulf of Mexico. Gulf of Mexico Origin, Waters, and Biota Biodiversity, Volume 1 (Darryl L. Felder and David A. Camp, editors, Texas A&M University Press, College Station, Texas).

48
13. Judkins, H. Debra A. Ingrao, C.F.E. Roper. In press. First records of Asperoteuthis acanthoderma (Lu, 1977) (CEPHALOPODA: OEGOPSIDAE: CHIROTEUTHIDAE), from the North Atlantic Ocean, Straits of Florida. Proceedings of the Biological Society of Washington.
14. Lambshead, P.J.D., J. Tietjen, T. Ferraro, & P. Jensen. 2000. Latitudinal diversity
gradients in the deep sea with special refernce to North Atlantic nematodes. Marine Ecology Progress Series; 194: 159-167.
15. LaRoe, E.T. 1967. A contribution to the biology of the loliginidae (Cephalopoda:
Myopsida) of the Tropical Western Atlantic. Master of Science thesis; University of Miami, Coral Gables, Fl.
16. Lipka, D.A. 1975. The systematics and zoogeography of cephalopods of the Gulf of
Mexico. PhD. Dissertation, Texas A&M University; 204 p. 17. Longhurst, A.1998. Ecological geography of the sea. Academic Press; 146-150. 18. Macpherson, E. 2002. Large-scale species-richness gradients in the Atlantic Ocean.
Proceedings of the Royal Society of London; B; 269: 1715-1720. ------ 2003. Species range size distributions for some marine taxa in the Atlantic Ocean.
Effect of latitude and depth. Biological Journal of the Linnean Society; November; 80(3): 437-455.
19. Magurran, Anne E. 2004. Measuring Biological Diversity. Blackwell Publishing.
256p. 20. Mokievsky, V. and Azovsky, A.2002. Re-evaluation of species diversity patterns of
free living marine nematodes. Marine Ecology Progress Series; 238: 101-108. 21. Nesis K.N. 1975. Cephalopods of the American Mediterranean Sea
EnglishTranslations of Selected Publications on Cephalopods; Editor M. Sweeney; Smithsonian Institutions Libraries; 1; 318- 358.
------ 1987. Cephalopods of the world. (Edited by L. Burgess) T.F.H. Publication, Inc. Ltd. New Jersey; 351 pp.
22. Passarella, K.C., 1990. Oceanic Cephalopod Assemblage in the Eastern Gulf of
Mexico, Department of Marine Science. University of South Florida: St. Petersburg, Fl. 50pp.
23. Peet, R.K. 1974. The Measurement of Species Diversity. Annual Review of Ecological Systems; 5: 285- 307.
24. Rapoport, E.H. 1982. Areography:geographical strategies of species; 1st English ed.
B. Drausal, transl. Publ. Fundacion Bariloche. V.1; Pergamon, New York.

49
25. Rex, M.A., J.A. Crame, C.T. Stuart, & G. Coyne. 2000. Latitudinal gradients of species richness in the deep-sea benthos of the North Atlantic; Proceedings of the National Academy of Sciences; USA; 97: 4082-4085.
26. Rex, M.A., J.A. Crame, C.T. Stuart, & A. Clarke. 2005. Large-scale biogeographic
patterns in marine molluscs: a confluence of history and productivity? Ecology; 86: 2288-2297.
27. Rohde, K.1992. Latitudinal gradients in species diversity: the search for the primary
cause. Oikos; 65: 514-527. 28. Roper, C.F.E., 1983. An overview of cephalopod systematics: status, problems, and
recommendations. Memoirs of the National Museum Victoria.; 44: 13-27. 29. Roper C.F.E, M.J. Sweeney & C.E. Nauen, 1984. FAO species catalogue. v.3
Cephalopods of the world. An annotated and illustrated catalogue of species of interest to fisheries. FAO Fish. Synop. (125) v3: 277p.
30. Rosa, Rui, Heidi M. Dierssen, Liliana Gonzalez, and Brad Seibel. 2008. Ecological
biogeography of cephalopod molluscs in the Atlantic Ocean: historical and contemporary causes of coastal diversity patterns; Global Ecology and Biogeography; 17: 600-610.
31. Roy, Kaustuv, David Jablonski, James W. Valentine, and Gary Rosenberg. 1998.
Marine latitudinal diversity gradients: Tests of causal hypotheses. Proceedings of the National Academy of Science. USA. 95; March; 3699-3702.
32. Roy, K., D. Jablonski, J.W. Valentine. 2000. Dissecting latitudinal diversity
gradients: functional groups and clades of marine bivalves; Proceeding of the Royal Society of London; B267: 293-299.
33. Sanders, H.L. 1968. Marine benthic diversity: a comparative study. American
Naturalist. 102: 243-282. 34. Smith, M.L.,K.E. Carpenter, & R.W Waller; 2002. An Introduction to the
Oceangraphy, Geology, Biogeography, and Fisheries of the Tropical and Subtropical Western Central Atlantic. Carpenter, K.E.(ed.) The living marine resources of the Western Central Atlantic. Volume 1: Introduction, molluscs, crustaceans, hagfishes, sharks, batoid fishes, and chimaeras. FAO Species Identification Guide for Fisheries Purposes and American Society of Ichthyologists and Herpetologists Special Publication No. 5. Rome, FAO. 2002. pp.1-600.
35. Stanley, Sonja (editor), 1995. A Global Representative System of Marine Protected
Areas: Vol 2, Marine Region 7: Wider Caribbean, Great Barrier Reef Marine Park Authority, The World Bank, The World Conservation Union,(IUCN); 15-42.

50
36. Steele, D.H.1988. Latitudinal variations in body size and species diversity in marine decapod crustaceans of the continental shelf. International Review of Hydrobiology; 73: 235-246.
37. Stevens, G.C., 1989. The Latitudinal Gradient in Geographical Range: How So
Many Species Coexist in the Tropics. The American Naturalist; 133(2): 240-256.
38. Stevens, G.C. 1996. Extending Rapoport’s rule to the Pacific marine fishes. Journal of Biogeography; 23: 149-154.
39. Summers, W.C., 1983. Physiological and Trophic Ecology of Cephalopods. The
Mollusca. Ecology; 6: 261-279. 40. Sweeney, M.J., Clyde F.E. Roper, 1998. Classification, Type Localities, and Type
Repositories of Recent Cephalopoda, in Systematics and biogeography of Cephalopods, M.V. Nancy A. Voss, Ronald B. Toll, and Michael J. Sweeney, Editor. Smithsonian Institution Press: Washington, D.C. 562-599.
41. Tomczak, Mattias, J. Stuart Godfrey. 1994. Regional Oceanogrpahy: An
Introduction. Pergamon; 310-317. 42. Vecchione, M. 1981. Aspects of the early life history of Loligo pealeii
(Cephalopoda: Myopsida). Journal of Shellfish Resources; 1:171-180. 43. Vecchione, M., Bruce B. Collette, 1996. Fisheries Agencies and Biodiversity. Ann.
Missouri Bot. Gard; 83: 29-36. 44. Vecchione, M., and B.B. Collette 1996b. The central role of systematics in marine
biodiversity issues. Oceanography; 9(1): 44–49. 45. Vecchione, M., M.F. Mickevich, K. Fauchald, B.B. Collette, A.B. Williams, T.A.
Munroe, and R.E. Young, 2000. Importance of Assessing Taxonomic Adequacy in Determining Fishing Effects on Marine Diversity. ICES Journal of Marine Science; 57: 677-681.
46. Vecchione, M., C.F.E. Roper, Michael J. Sweeney, and C.C. Lu. 2001. Distribution,
relative abundance, and developmental morphology of paralarval cephalopods in the western North Atlantic Ocean. U.S. Department of Commerce, NOAA Tech. Rep. NMFS 152, 54p.
47. Vecchione, 2002. Cephalopods. Carpenter, K.E.(ed.) The living marine resources
of the Western Central Atlantic. Volume 1: Introduction, molluscs, crustaceans, hagfishes, sharks, batoid fishes, and chimaeras. Cephalopods of Western Central Atlantic; FAO Species Identification Guide for Fisheries Purposes and American Society of Ichthyologists and Herpetologists Special Publication No. 5. Rome, FAO. 2002. 1-600.

51
48. Voss, G.L. 1956. A review of the cephalopods of the Gulf of Mexico. Bull Mar. Sci. Gulf and Caribbean; 6:85-178.
------1973. The potentially commercial species of octopus and squid of Florida, the Gulf of Mexico and the Caribbean Sea. University of Miami Sea Grant Program (NOAA Sea Grant No. 04-3-158-27) Miami, Fl.
49. Voss, G., Thomas F. Brakoniecki, 1985. Squid resources of the Gulf of Mexico and
Southeast Atlantic coasts of the United States. Science Council Studies; 9: 27-37. 50. Voss, N.A., Vecchione, M., Toll, R.B. & Sweeney, M.J.(eds) 1998. Systematics and
biogeography of cephalopods. Smithson, Contrib. Zool.; 586: 1-599.

52
CHAPTER 3 CEPHALOPODS (MOLLUSCA: CEPHALOPODA) OF THE GULF OF MEXICO Heather L. Judkins, Michael Vecchione, and Clyde F.E. Roper (HJ) College of Marine Science, University of South Florida, St. Petersburg, Fl, 33701; USA (email: [email protected]) (MV) NMFS National Systematics Laboratory, National Museum of Natural History, Smithsonian Institution, Washington, DC 20013-7012 USA (email: [email protected]) (CR) Invertebrate Zoology, MRC 118, National Museum of Natural History, Smithsonian Institution; Washington DC 20013-7012 USA (email: [email protected])
3.1 Introduction
The cephalopods of the Gulf of Mexico have not been studied comprehensively
since Voss’ (1956) monograph. Several cephalopod studies have included this region in
broader studies (e.g., Vecchione 2002) or have examined distribution based on limited
geographic area (e.g. Nesis 1975, Passarella and Hopkins 1991). Collectively,
approximately 109 species of cephalopods in 31 families occur in the Western Central
Atlantic and adjacent areas, including the Caribbean Sea and the Gulf of Mexico. Both
squids and octopods are common in these waters (Vecchione 2002). The present paper
updates Voss’ works and summarizes the species found in the Gulf of Mexico to date.
Two major groups of cephalopods exist today: the Nautiloidea, with a few species
of the pearly nautilus confined to the Indo-West Pacific, and the Neocoleoidea, consisting

53
of squids, cuttlefishes, octopods, and their relatives. Neocoleoidea comprise more than
700 species worldwide:
(http://tolweb.org/tree?group=Cephalopoda&contgroup=Mollusca). The neocoleoids that
exist today evolved from forms that first appeared in the upper Triassic and Lower
Jurassic (between 200 and 150 million years ago). Although there are relatively few
species of living cephalopods, they occupy a great variety of habitats in all of the world’s
oceans. Individual species may be very abundant and are important in marine food webs.
Some species are major targets for commercial fisheries.
The Neocoleoidea contains two extant Superorders: the Octopodiformes
(octopods and vampire squid) and the Decapodiformes (squid and cuttlefishes). These
groups occupy all major habitats in the oceans from intertidal to great depths (deepest
record is 7279 m, Aldred et al. 1983, Voss 1988) and from southern to northern polar
seas. No cephalopods are found in salinities less than about 17.5 practical salinity units
(psu).
Many species of oceanic cephalopods undergo diel vertical migrations, wherein
they occur at depths of about 400 m to 1000 m during the day, then ascend to the
uppermost 200 m or so during the night (e.g., Enoploteuthidae, Ommastrephidae).
Abundance patterns vary (depending on group, habitat, and season) from isolated
territorial individuals (primarily octopods and sepioids), through small schools with a few
dozen individuals, to massive schools with millions of oceanic squids.
Although many cephalopods reach large sizes, generally they have a very short
life span. The life expectancy appears to be about 1–2 years in most cephalopods, but
large species of squids and octopods and those in cold water habitats may live longer.

54
Conversely, some small oceanic squids such as pyroteuthids may complete their life
cycles in less than six months. This is part of a life history strategy that seems designed
for rapid increase in population size. It has been suggested that this “life-strategy” may
guarantee survival against environmentally stressful conditions, including those by heavy
predation or over-fishing. However, as cephalopod fisheries experienced further
extensive development, parallel concern developed regarding potential overexploitation
(Jereb and Roper, 2005).
Cephalopods are important in terms of food web relationships, commercial
fisheries, and biomedical research. Cephalopods are born into the third trophic level and
progress one or two stages through their life. Research has not shown them as achieving
top-predator status because there always seems to be some vertebrate that preys upon
them (Summers 1983). Cephalopods are active predators that feed upon shrimps, crabs,
fishes, and other cephalopods, and, in the case of octopods, on other molluscs. In turn,
cephalopods are major food items in the diets of toothed whales, seals, pelagic birds and
both benthic and pelagic fishes as well as other cephalopods.
Cephalopod fisheries provide an important food source for humans across the
globe. Over three million metric tons are caught each year worldwide (Jereb and Roper,
2005). Squid fisheries also have existed in North America, historically principally to
provide bait for other fisheries, but recent decades have seen the development of
substantial fisheries for food production, as well. The total commercial catch of
cephalopods in the Western Central Atlantic varied during 1993 to 1998 between 19,000
tons and 31,000 tons, mostly landed in Mexico (Vecchione 2002). However, Voss
reported in 1973 that of the numerous species known on the coasts of Florida, the Gulf of

55
Mexico and the Caribbean Sea, “only about 12 species seem to have actual or potential
(fisheries) importance” (Voss et al. 1973; p.1). Of those, species that are commercially
important to the Gulf of Mexico include: Octopus maya, Illex coindetii, Doryteuthis
(Loligo) pealei, and Doryteuthis (Loligo) plei (Voss et al. 1985).
Squids also are important to biomedical research; for instance, much of what is
known about human neurophysiology results from experiments with the giant axon of
squids. Scientists culture squid in laboratories in order to study the giant axon. Lee et.
al. (1994) cultured the Indo-West Pacific species Sepioteuthis lessoniana (Ferussac,
1830) for this purpose because of its large hatchlings, and the quality of its large-diameter
axons for study (Lee 1994). LaRoe (1971) previously had worked with a Caribbean
species, S. sepioidea, for this purpose. Because of the highly developed brain and
sensory organs, cephalopods are valuable in behavioral and comparative neuroanatomical
studies as well.
The fauna of the Gulf of Mexico lacks the nautiloids and true cuttlefishes but does
include sepiolids, myopsid and oegopsid squids, octopods, and the vampyromorph,
Vampyroteuthis infernalis. Published records of cephalopod species in the Gulf of
Mexico date back to LeSueur (1821), but the modern, comprehensive systematic studies
begin with G.L. Voss, who reported 42 neritic and oceanic species in 1956(G. Voss
1956). Since then, many oceanic species have been added to the list (N. Voss and G.
Voss 1962, Roper 1964, Voss 1964, Roper et al. 1969, Lipka 1975, Passarella 1990).
Although the composition of the cephalopod fauna off southern Florida is known almost
exhaustively, the fauna of the rest of the Gulf of Mexico is less well studied. The
cephalopods of the Mexican waters of the Gulf were reviewed by Salcedo-Vargas (1991)

56
using specimens and past literature, he reported some questionable identifications. The
most recent compilation of the cephalopods in the Gulf of Mexico is that of Vecchione
(2002).
3.2 Major systematic revisions since 1954
The status of the systematics of cephalopods is rapidly changing, as research has
increased substantially world-wide, during the past 30 years. The families of living
cephalopods are, for the most part, well resolved and relatively well-accepted. Species-
level taxa can usually be placed in well-defined families (Jereb and Roper, 2005).
However, phylogenetic relationships among families within the major groups remain
uncertain (Vecchione 2002). Sweeney and Roper (1998, p.561) addressed the confusion,
stating, “We realize that numerous systematic problems exist at all taxonomic levels of
the Cephalopoda. For example, several higher taxa have been proposed (i.e., superorder
Pseudoctobrachia Guerra, 1992, and order Cirroctopodida Young, 1989).” The resolution
of these problems requires considerable research and review, as new cephalopod research
initiatives are being pursued globally.
3.3 Comparative assessment of group in GOM
One of the elements absent in current cephalopod research is comprehensive
studies of large oceanic or faunal regions. Numerous isolated island studies, studies from
a fisheries perspective, or those for biomedical advances do exist, but the need for
comprehensive systematics, abundance, distribution, and ecology requires significant
attention. The Broad Caribbean Realm, which includes the Caribbean Sea proper, the
Gulf of Mexico, and waters that extend through the Bahamas, is an area that is in need of
such comprehensive study. The first author of this study currently is researching this

57
area. Species that have been collected in the area allow the opportunity to further define
phylogenetic relationships within the cephalopods as a whole as well as to address the
trophic importance of cephalopods within the region, for example.
Because of the great diversity in the form, habitat, and behavior of cephalopods,
no single method is adequate to capture and/or sample the complex cephalopod fauna
(Rathjen 1991). The excellent vision and high mobility of most cephalopods traditionally
means that they have been undersampled with standard trawling and collecting protocols.
Despite their limitations, midwater trawls offer a starting point for population assessment
of pelagic species and provide minimum estimates of oceanic cephalopod abundance
(Passarella 1990). Numerous facilities around the Broad Caribbean house unidentified
cephalopods which when identified, will add further insight to the diversity of the
cephalopod families and ecology, and provide a better fisheries perspective about
potentially viable future catches in the region.
3.4 Explanation of Checklist
The classification and nomenclature used here follow that of Vecchione (2002), as
it is the most recent compilation for the Western Atlantic region. Orders and families are
arranged phylogenetically, and genera and species are arranged alphabetically.
Cephalopods are not exclusively benthic as are most other mollusca, and many are highly
mobile, pelagic/oceanic species. This habitat niche requires the use of the term “central”
in the Gulf of Mexico range column. Depth data and overall range for organisms are in
italics where they could not be determined exclusively for the Gulf of Mexico. Depth
ranges include paralarval through adult stages, so they represent the total known vertical
range for the species. However, most pelagic species exhibit several more specific

58
ranges during different phases of their life cycles: for example, paralarvae are epipelagic,
restricted to the upper 100–200 m; many then undergo ontogenetic descent, moving into
deeper waters with growth. Adults of many species then exhibit diel vertical migration
from around 400 m to 1000 m during the day into the upper 200–400 m at night to feed.
Very little information is available on biology and lifestyle for many of the deep sea
cephalopods in the Gulf of Mexico; this explains the “unknown” notation in some
columns.
The abbreviations used in the “Habitat-Biology” category are as follows: bat =
bathypelagic (> 1000 m); ben = benthic; cep = coastal surface and epipelagic; crr = coral
reef; cts = continental shelf; dps = deep sea; end = endemic; epi = epipelagic (0–200 m);
ins = insular; mes = mesopelagic (200–1000 m); ner = neritic; oce = oceanic; sft = mud,
sands, clays; sgr = seagrass; shw = shallow water; slp = continental slope. The
abbreviations used in the “Overall geographic range” category are as follows: AT =
Atlantic Ocean; BE = Bermuda; BH = Bahamas; BR = Brazil; CH = Cape Hatteras,
North Carolina; CT = Connecticut; CR = Caribbean; FL = Florida; IO = Indian Ocean;
ME = Mediterranean; N = North; NE = New England; PO = Pacific Ocean; S = South;
SA = South America; ST = Subtropical; T = Tropical; TWA = Tropical Western Atlantic;
UR = Uruguay; W = West; WA = Western Atlantic.
The abbreviations used in the “Gulf of Mexico Range” category vary because of
the high mobility of the species. In some cases, a specific region cannot be defined at
this point. Therefore, the term, “central” (cen) is indicative of the mid-Gulf of Mexico
species. The term, “entire” (ent) is used where the species is found throughout all regions
of the Gulf of Mexico. For those species that are found in more than one region, an

59
overall region is use. For example, instead of “se” and “ne” being used, the term “e”
(east) is used where appropriate.

60
3.5 Acknowledgements
(HJ) would like to acknowledge Dr. Joseph Torres and the College of Marine
Science at the University of South Florida for providing invaluable guidance, resources,
and facilities for furthering her doctoral research.

61
REFERENCES 1. Aldred, R. G., M. Nixon and J. Z. Young. 1983. Cirrothauma murrayi Chun,
finned octopod. Philosophical Transactions of the Royal Society B 301: 1–54.
2. Clarke, M., and C. Lu. 1974. Vertical distribution of cephalopods at 30°N 23°W in the North Atlantic. Journal of the Marine Biological Association, U.K. 54(4): 969–984. 3. Clarke, M., and C. Lu. 1975. Vertical distribution of cephalopods at 18°N 25°W in the North Atlantic. Journal of the Marine Biological Association, U.K. 55(1): 165–182. 4. DeBose, J. L., and M. Vecchione. 2005. First documentation of the Roper inshore squid, Loligo roperi (Cohen 1976) in the Gulf of Mexico. Gulf of Mexico Science 23(1): 132–135. 5. Hochberg, F. G., M. Nixon and R. B. Toll. 1992. Order Octopoda Leach, 1818, Pp. 213–279 in Sweeney, M. J., C. F. E. Roper, K. M. Mangold, M. R. Clarke and S. v. Boletzky, eds. “Larval” and juvenile cephalopods: A manual for their identification., Smithsonian Contributions to Zoology 513: 1-282. 6. Jereb, P. and C.F.E. Roper, eds. 2005. Cephalopods of the World. An Annotated and Illustrated Catalogue of Species Known to Date. Volume 1. Chambered nautiluses and sepioids (Nautilidae, Sepiidae, Sepiolidae, Sepiadariidae, Idiosepiidae and Spirulidae). FAO Species Catalogue for Fishery Purposes. No. 4, Vol. 1 Rome, Italy. 7. Kubodera, T., U. Piatkowski, T. Okutani, and M. R. Clarke. 1998. Taxonomy and zoogeography of the family Onychoteuthidae (Cephalopoda: Oegopsida). Pp. 277–291 in N. A. Voss, R. B. Toll, and M. Vecchione, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology 586. 8. LaRoe, E. T. 1971. The culture and Maintenance of the Loliginid Squids Sepioteuthis sepioidea and Doryteuthis plei. Marine Biology; v. 9, 9-25. 9. Lee, P. G., P. E. Turk, W. T. Yang, and R. T. Hanlon. 1994. Biological characteristics and biomedical applications of the squid Sepioteuthis lessoniana cultured through multiple generations. Biological Bulletin 186: 328–341.

62
10. Lesueur, C.A. 1821. Descriptions of Several New Species of Cuttle-fish. Journal of the Academy of Natural Sciences of Philadelphia, 2(1):86-101. 11. Lipka, D. A. 1975. The Systematics and Zoogeography of Cephalopods from the Gulf of Mexico. Ph.D. dissertation, Texas A&M University, College Station, Texas. 351 pp. 12. Nesis, K. N. 1975. Cephalopods of the American Mediterranean Sea. English Translations of Selected Publications on Cephalopods. Smithsonian Institution Libraries, 2003, 1(1): 319–358. 13. Nesis, K. N. 1987. Cephalopods of the World. Squids, Cuttlefishes, Octopuses, and Allies. (trans. B. S. Levitov). T.F.H. Publications, Neptune City, New Jersey. 351 pp. 14. Nixon, M., and J. Z. Young. 2003. The Brains and Lives of Cephalopods. Oxford University Press, Oxford, U.K. 448 pp. 15. Okutani, T. 2005. Cuttlefishes and Squids of the World; 253 Pp. National Cooperative Association of Squid Processors, Tokyo, Japan. 254 pp. 16. Passarella, K. C. 1990. Oceanic Cephalopod Assemblage in the Eastern Gulf of Mexico. Masters thesis, University of South Florida, St. Petersburg, Florida. 50 pp. 17. Passarella, K. C., and T. L. Hopkins. 1991. Species composition and food habits of the micronektonic cephalopod assemblage in the Eastern Gulf of Mexico. Bulletin of Marine Science 49(1–2): 638–659. 18. Rathjen, W. F. 1991. Cephalopod Capture Methods: An Overview. Bulletin of Marine Science. 49(1-2): 494–505. 19. Roper, C. F. E. 1964. Enoploteuthis anapsis, a new species of enoploteuthid squid (Cephalopoda: Oegopsida) from the Atlantic Ocean. Bulletin of Marine Science of the Gulf and Caribbean 14(1): 140–148. 20. Roper, C. F. E., C. C. Lu, and K. Mangold. 1969. A new species of Illex from the Western Atlantic and distributional aspects of other Illex Species (Cephalopoda: Oegopsidae). Proceedings of Biological Society of Washington 82: 295–322.

63
21. Roper, C. F. E., C.C. Lu, and M. Vecchione. 1998. A revision of the systematics and distribution of Illex species. Pp. 405-423 in N.A. Voss, M. Vecchione R. B. Toll and M. J. Sweeney, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology, 586(I-II). Tokai University Press. Tokyo, Japan. 22. Roper, C. F. E., M. J. Sweeney and M. Clarke. 1985. FAO Species Identification Sheets for Fishery Purposes. Pp. 117–205 in W. Fischer, and J. C. Hureau, eds. Southern Ocean. Food and Agricultural Organization of the United Nations, Rome, vol. 1. 23. Roper, C. F. E., M. J. Sweeney, and C. Nauen. 1984. Cephalopods of the World. An Annotated and Illustrated Catalogue of Species of Interest to Fisheries. FAO Fisheries Synopsis No.125, v.3, Food and Agricultural Organization of the United Nations, Rome. 277 pp. 24. Roper, C. F. E., and M. Vecchione. 1993. A geographic and taxonomic review of Taningia danae Joubin, 1931(Cephalopoda: Octopoteuthidae), with new records and observations on bioluminescence. Pp. 441–456 in T. Okutani, R. K. O’Dor and T. Kubodera, eds. The Recent Advances in Cephalopod Fishery Biology. 25. Roper, C. F. E., and R. E. Young. 1975. Vertical distribution of pelagic cephalopods. Smithsonian Contributions to Zoology 209: 1–51. 26. Salcedo-Vargas, M. A. 1991. Checklist of the cephalopods from the Gulf of Mexico. Bulletin of Marine Science 49(1-2): 216–220. 27. Summers, W. C. 1983. Physiological and trophic ecology of cephalopods. Pp. 261-279 in W. D. Russell-Hunter, ed. The Mollusca, Volume 6. Ecology, Academic Press, Orlando, Florida. 28. Sweeney, M. J., and C. F. E. Roper, 1998. Classification, type localities, and type repositories of Recent Cephalopoda. Pp. 561–595 in N. A. Voss, M. Vecchione, R. B. Toll and M. J. Sweeney, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology 586(I-II). 29. Vecchione, M. 2002. Cephalopods. Pp. 150-244 in K. E. Carpenter, ed. The Living Marine Resources of the Western Central Atlantic. Volume 1. Introduction, mollusks, crustaceans, hagfishes, sharks, batoid fishes and chimaeras. FAO Identificatiion Guide for Fisheries Purposes. The Food and Agriculture Organization of the United Nations, Rome, Italy.

64
30. Vecchione, M. and C. F. E. Roper. 1991. Cephalopods observed from submersibles in the Western North Atlantic. Bulletin of Marine Science 49(1-2): 433–445. 31. Vecchione, M., C. F. E. Roper and M. J. Sweeney. 1989. Marine flora and fauna of the Eastern United States. Mollusca: Cephalopoda. NOAA Technical Report NMFS 73. U.S. Department of Commerce. 23 pp. 32. Vecchione, M., and R. E. Young. In press. The squid family Magnapinnidae (Mollusca: Cephalopoda) in the Atlantic Ocean, with descriptions of new species. 33. Villanueva, R., M. A. Collins, P. Sanchez and N. A. Voss. 2002. Systematics, distribution, and biology of the cirrate octopods of the genus Opisthoteuthis (Mollusca, Cephalopoda) in the Atlantic Ocean, with description of two new species. Bulletin of Marine Science 71(2): 933–985. 34. Voss, G. L. 1956. A review of the cephalopods of the Gulf of Mexico. Bulletin of Marine Science of the Gulf and Caribbean 6(2): 85–178. 35. Voss, G. L. 1964. A note on some cephalopods from Brazil with a description of a new species of octopod, Eledone massyae. Bulletin of Marine Science of the Gulf and Caribbean 14(3): 511–516. 36. Voss, G. L. 1988. Evolution and phylogenetic relationship of deep-sea cephalopods (Cirrata and Incirrata). Pp. 253–276 in M. R. Clarke, and E. R. Trueman, eds. The Mollusca, Vol. 12. Paleontology and neontology of cephalopods. Academic Press, San Diego, California. 37. Voss, G. L., and T. F. Brakoniecki. 1985. Squid resources of the Gulf of Mexico and Southeast Atlantic coasts of the United States. Pp. 27-37 in V. M. Hodder, ed. Northwest Atlantic Fisheries Organization: Scientific Council Studies, Special Session on Squids 9. 38. Voss, G. L., L. Opresko, and R. Thomas. 1973. The potentially commercial species of octopus and squid of Florida, the Gulf of Mexico and the Caribbean Sea. University of Miami Sea Grant Field Guide Series, No. 2: 1-33. 39. Voss, G. L., and R. R. Toll. 1998. The systematics and nomenclatural status of the octopodinae described from the Western Atlantic Ocean. Pp. 457–474 in N. A. Voss, R. B. Toll, and M. Vecchione, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology 586(I-II).

65
40. Voss, N. A., and G. L. Voss. 1962. Two new species of squids of the genus Calliteuthis from the Western Atlantic with a redescription of Calliteuthis reversa Verrill. Bulletin of Marine Science of the Gulf and Caribbean 12(2): 169–200. 41. Voss, N. A. 1980. A generic revision of the Cranchiidae (Cephalopoda; Oegopsida). Bulletin of Marine Science 30: 365–412. 42. Voss, N. A. 1988. Systematics and zoogeography of cephalopods. Malacologia 29(1): 209–214. 43. Voss, N. A., K. N. Nesis, P. Rodhouse. 1998. The cephalopod family Histioteuthidae (Oegopsida): systematics, biology, and biogeography. Pp. 293–372 in N. A. Voss, R. B. Toll, and M. Vecchione, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology 586(II). 44. Young, R. E., C. F. E. Roper. 1969. A monograph of the Cephalopoda of the North Atlantic: The family Cycloteuthidae. Smithsonian Contributions to Zoology 5: 1–24. 45. Young, R. E. 1975. Leachia pacifica (Cephalopoda, Teuthoidea): Spawning habitat and function of the brachial photophores. Pacific Science 29: 19–25. 46. Young, R. E. 1978. Vertical distribution and photosensitive vesicles of pelagic cephalopods from Hawaiian Waters. Fishery Bulletin 76: 583–615. 47. Young, R. E., Vecchione, M., and Mangold K. 1995. Coleoidea Bather, 1888. Octopods, squids, cuttlefishes and their relatives. Version 01 January 1995 (under construction). http://tolweb.org/Coleoidea/19400/1995.01.01 in The Tree of Life Web Project, http://tolweb.org/ 48. Young, R. E, L. A. Burgess, C. F. E. Roper, M. J. Sweeney, and S. J. Stephen 1998. Classification of the Enoploteuthidae, Pyroteuthidae, and Ancestrocheiridae. Pp. 239–255 in N. A. Voss, R. B. Toll, and M. Vecchione, eds. Systematics and Biogeography of Cephalopods. Smithsonian Contributions to Zoology 586(1).
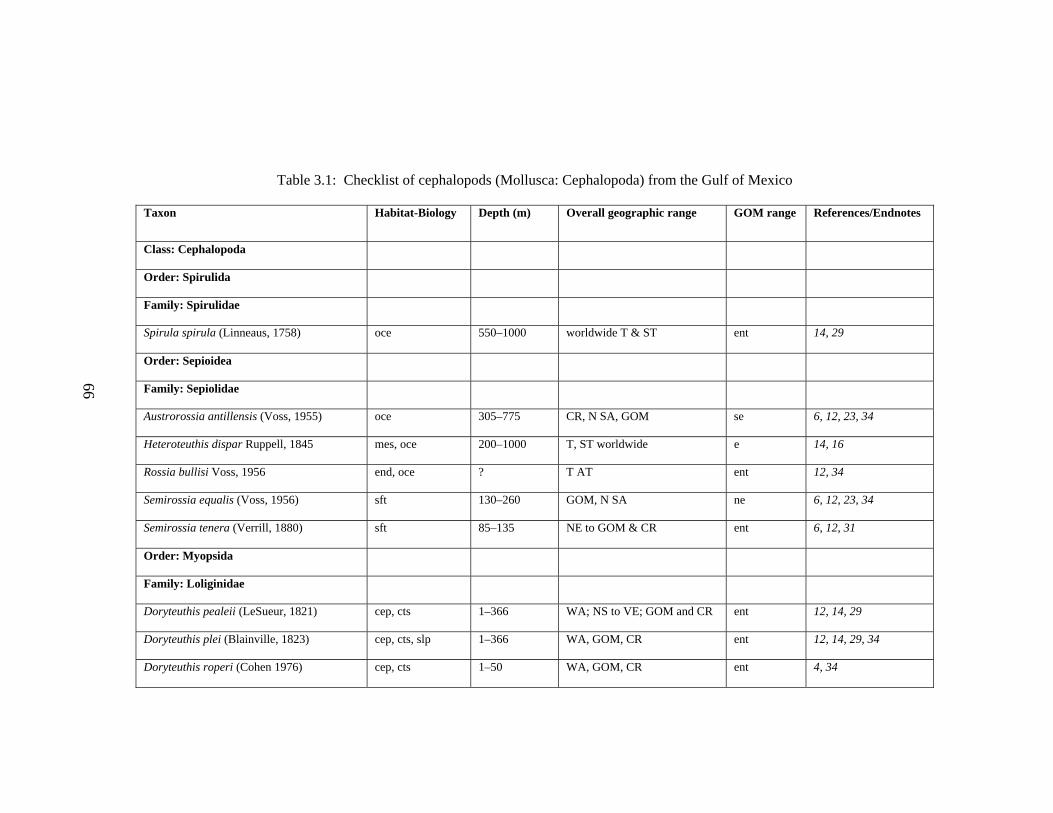
66
Table 3.1: Checklist of cephalopods (Mollusca: Cephalopoda) from the Gulf of Mexico
Taxon Habitat-Biology Depth (m) Overall geographic range GOM range References/Endnotes
Class: Cephalopoda
Order: Spirulida
Family: Spirulidae
Spirula spirula (Linneaus, 1758) oce 550–1000 worldwide T & ST ent 14, 29
Order: Sepioidea
Family: Sepiolidae
Austrorossia antillensis (Voss, 1955) oce 305–775 CR, N SA, GOM se 6, 12, 23, 34
Heteroteuthis dispar Ruppell, 1845 mes, oce 200–1000 T, ST worldwide e 14, 16
Rossia bullisi Voss, 1956 end, oce ? T AT ent 12, 34
Semirossia equalis (Voss, 1956) sft 130–260 GOM, N SA ne 6, 12, 23, 34
Semirossia tenera (Verrill, 1880) sft 85–135 NE to GOM & CR ent 6, 12, 31
Order: Myopsida
Family: Loliginidae
Doryteuthis pealeii (LeSueur, 1821) cep, cts 1–366 WA; NS to VE; GOM and CR ent 12, 14, 29
Doryteuthis plei (Blainville, 1823) cep, cts, slp 1–366 WA, GOM, CR ent 12, 14, 29, 34
Doryteuthis roperi (Cohen 1976) cep, cts 1–50 WA, GOM, CR ent 4, 34
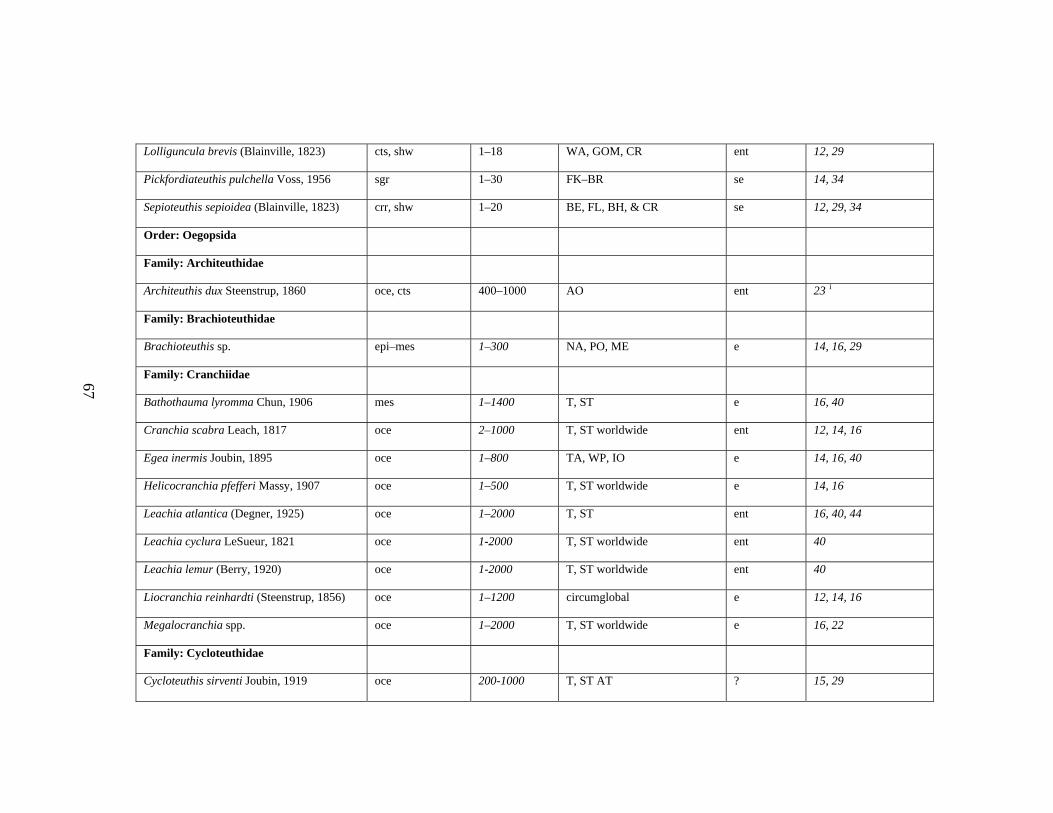
67
Lolliguncula brevis (Blainville, 1823) cts, shw 1–18 WA, GOM, CR ent 12, 29
Pickfordiateuthis pulchella Voss, 1956 sgr 1–30 FK–BR se 14, 34
Sepioteuthis sepioidea (Blainville, 1823) crr, shw 1–20 BE, FL, BH, & CR se 12, 29, 34
Order: Oegopsida
Family: Architeuthidae
Architeuthis dux Steenstrup, 1860 oce, cts 400–1000 AO ent 23 i
Family: Brachioteuthidae
Brachioteuthis sp. epi–mes 1–300 NA, PO, ME e 14, 16, 29
Family: Cranchiidae
Bathothauma lyromma Chun, 1906 mes 1–1400 T, ST e 16, 40
Cranchia scabra Leach, 1817 oce 2–1000 T, ST worldwide ent 12, 14, 16
Egea inermis Joubin, 1895 oce 1–800 TA, WP, IO e 14, 16, 40
Helicocranchia pfefferi Massy, 1907 oce 1–500 T, ST worldwide e 14, 16
Leachia atlantica (Degner, 1925) oce 1–2000 T, ST ent 16, 40, 44
Leachia cyclura LeSueur, 1821 oce 1-2000 T, ST worldwide ent 40
Leachia lemur (Berry, 1920) oce 1-2000 T, ST worldwide ent 40
Liocranchia reinhardti (Steenstrup, 1856) oce 1–1200 circumglobal e 12, 14, 16
Megalocranchia spp. oce 1–2000 T, ST worldwide e 16, 22
Family: Cycloteuthidae
Cycloteuthis sirventi Joubin, 1919 oce 200-1000 T, ST AT ? 15, 29
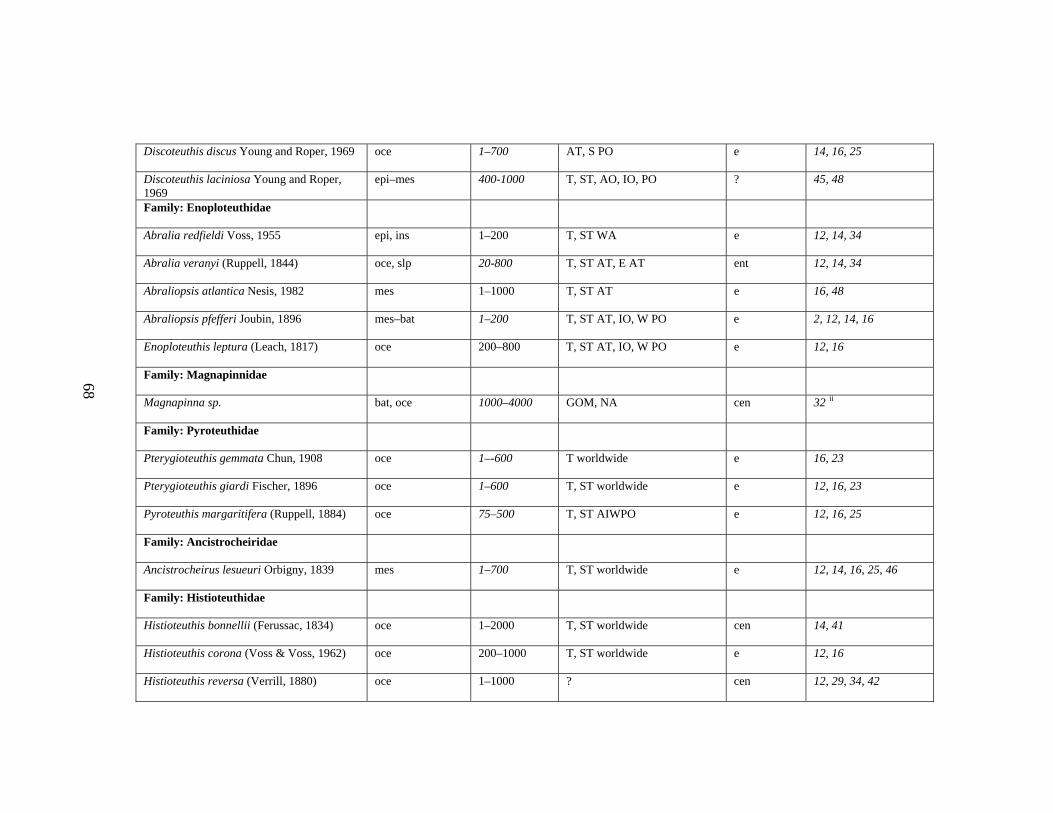
68
Discoteuthis discus Young and Roper, 1969 oce 1–700 AT, S PO e 14, 16, 25
Discoteuthis laciniosa Young and Roper, 1969
epi–mes 400-1000 T, ST, AO, IO, PO ? 45, 48
Family: Enoploteuthidae
Abralia redfieldi Voss, 1955 epi, ins 1–200 T, ST WA e 12, 14, 34
Abralia veranyi (Ruppell, 1844) oce, slp 20-800 T, ST AT, E AT ent 12, 14, 34
Abraliopsis atlantica Nesis, 1982 mes 1–1000 T, ST AT e 16, 48
Abraliopsis pfefferi Joubin, 1896 mes–bat 1–200 T, ST AT, IO, W PO e 2, 12, 14, 16
Enoploteuthis leptura (Leach, 1817) oce 200–800 T, ST AT, IO, W PO e 12, 16
Family: Magnapinnidae
Magnapinna sp. bat, oce 1000–4000 GOM, NA cen 32 ii
Family: Pyroteuthidae
Pterygioteuthis gemmata Chun, 1908 oce 1–-600 T worldwide e 16, 23
Pterygioteuthis giardi Fischer, 1896 oce 1–600 T, ST worldwide e 12, 16, 23
Pyroteuthis margaritifera (Ruppell, 1884) oce 75–500 T, ST AIWPO e 12, 16, 25
Family: Ancistrocheiridae
Ancistrocheirus lesueuri Orbigny, 1839 mes 1–700 T, ST worldwide e 12, 14, 16, 25, 46
Family: Histioteuthidae
Histioteuthis bonnellii (Ferussac, 1834) oce 1–2000 T, ST worldwide cen 14, 41
Histioteuthis corona (Voss & Voss, 1962) oce 200–1000 T, ST worldwide e 12, 16
Histioteuthis reversa (Verrill, 1880) oce 1–1000 ? cen 12, 29, 34, 42
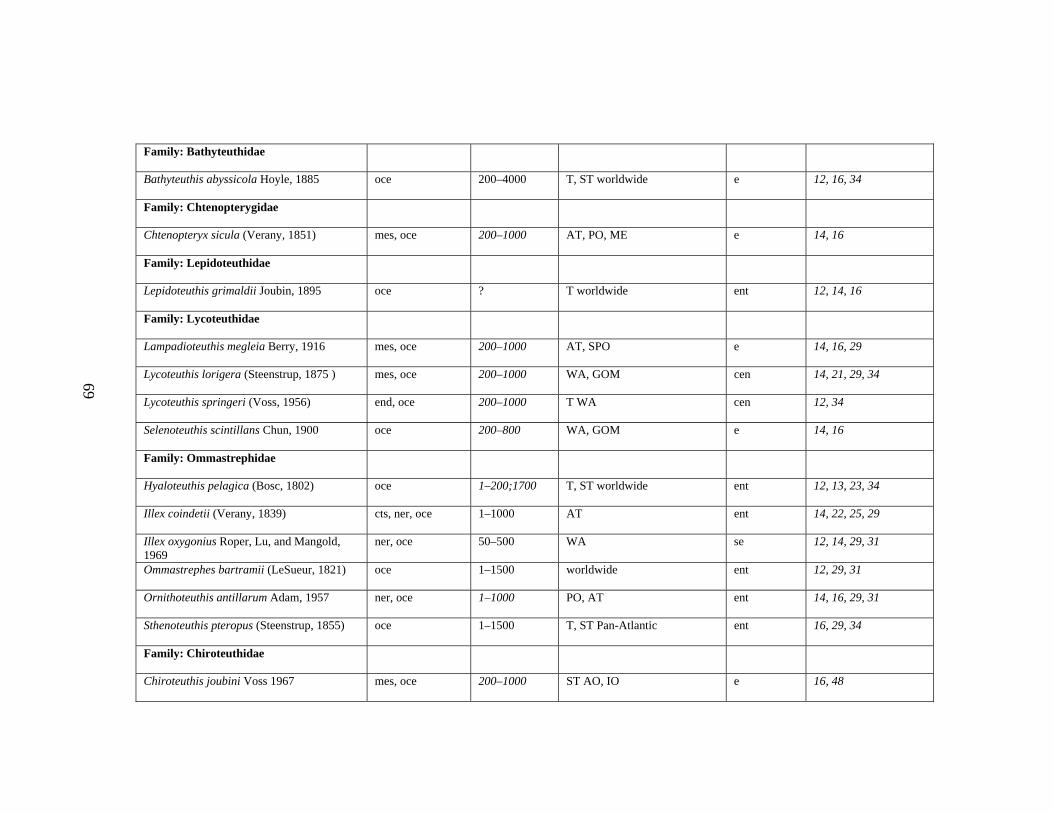
69
Family: Bathyteuthidae
Bathyteuthis abyssicola Hoyle, 1885 oce 200–4000 T, ST worldwide e 12, 16, 34
Family: Chtenopterygidae
Chtenopteryx sicula (Verany, 1851) mes, oce 200–1000 AT, PO, ME e 14, 16
Family: Lepidoteuthidae
Lepidoteuthis grimaldii Joubin, 1895 oce ? T worldwide ent 12, 14, 16
Family: Lycoteuthidae
Lampadioteuthis megleia Berry, 1916 mes, oce 200–1000 AT, SPO e 14, 16, 29
Lycoteuthis lorigera (Steenstrup, 1875 ) mes, oce 200–1000 WA, GOM cen 14, 21, 29, 34
Lycoteuthis springeri (Voss, 1956) end, oce 200–1000 T WA cen 12, 34
Selenoteuthis scintillans Chun, 1900 oce 200–800 WA, GOM e 14, 16
Family: Ommastrephidae
Hyaloteuthis pelagica (Bosc, 1802) oce 1–200;1700 T, ST worldwide ent 12, 13, 23, 34
Illex coindetii (Verany, 1839) cts, ner, oce 1–1000 AT ent 14, 22, 25, 29
Illex oxygonius Roper, Lu, and Mangold, 1969
ner, oce 50–500 WA se 12, 14, 29, 31
Ommastrephes bartramii (LeSueur, 1821) oce 1–1500 worldwide ent 12, 29, 31
Ornithoteuthis antillarum Adam, 1957 ner, oce 1–1000 PO, AT ent 14, 16, 29, 31
Sthenoteuthis pteropus (Steenstrup, 1855) oce 1–1500 T, ST Pan-Atlantic ent 16, 29, 34
Family: Chiroteuthidae
Chiroteuthis joubini Voss 1967 mes, oce 200–1000 ST AO, IO e 16, 48
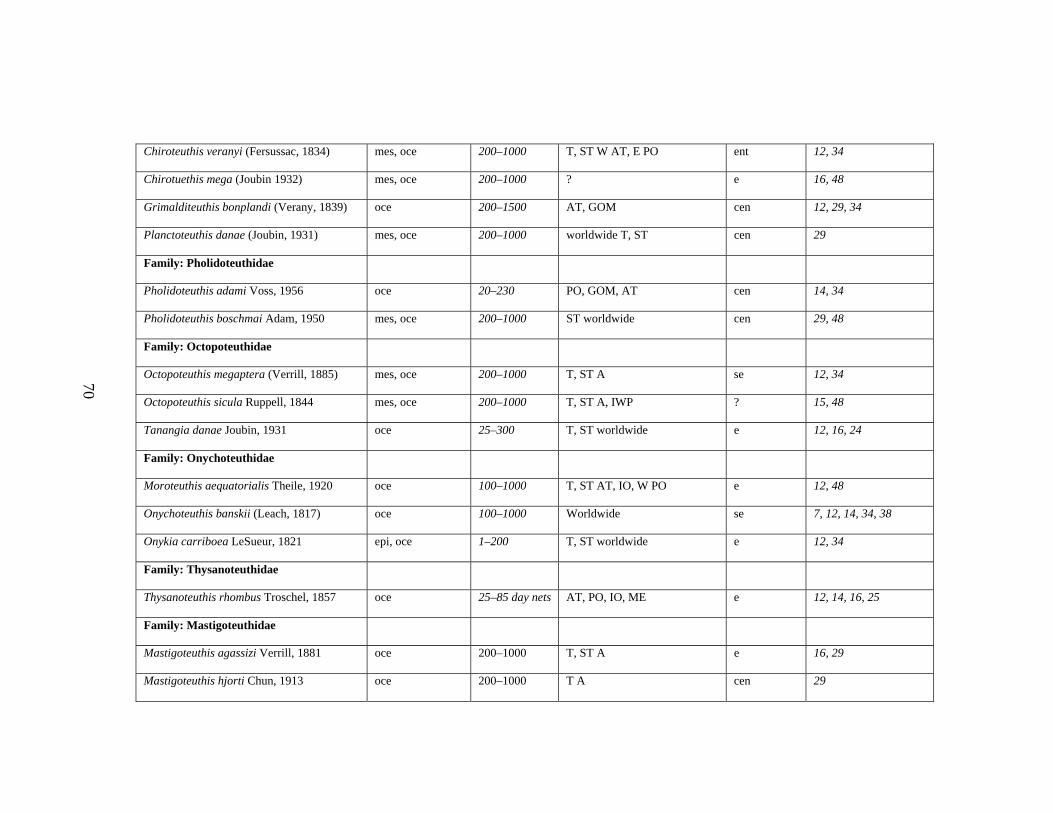
70
Chiroteuthis veranyi (Fersussac, 1834) mes, oce 200–1000 T, ST W AT, E PO ent 12, 34
Chirotuethis mega (Joubin 1932) mes, oce 200–1000 ? e 16, 48
Grimalditeuthis bonplandi (Verany, 1839) oce 200–1500 AT, GOM cen 12, 29, 34
Planctoteuthis danae (Joubin, 1931) mes, oce 200–1000 worldwide T, ST cen 29
Family: Pholidoteuthidae
Pholidoteuthis adami Voss, 1956 oce 20–230 PO, GOM, AT cen 14, 34
Pholidoteuthis boschmai Adam, 1950 mes, oce 200–1000 ST worldwide cen 29, 48
Family: Octopoteuthidae
Octopoteuthis megaptera (Verrill, 1885) mes, oce 200–1000 T, ST A se 12, 34
Octopoteuthis sicula Ruppell, 1844 mes, oce 200–1000 T, ST A, IWP ? 15, 48
Tanangia danae Joubin, 1931 oce 25–300 T, ST worldwide e 12, 16, 24
Family: Onychoteuthidae
Moroteuthis aequatorialis Theile, 1920 oce 100–1000 T, ST AT, IO, W PO e 12, 48
Onychoteuthis banskii (Leach, 1817) oce 100–1000 Worldwide se 7, 12, 14, 34, 38
Onykia carriboea LeSueur, 1821 epi, oce 1–200 T, ST worldwide e 12, 34
Family: Thysanoteuthidae
Thysanoteuthis rhombus Troschel, 1857 oce 25–85 day nets AT, PO, IO, ME e 12, 14, 16, 25
Family: Mastigoteuthidae
Mastigoteuthis agassizi Verrill, 1881 oce 200–1000 T, ST A e 16, 29
Mastigoteuthis hjorti Chun, 1913 oce 200–1000 T A cen 29
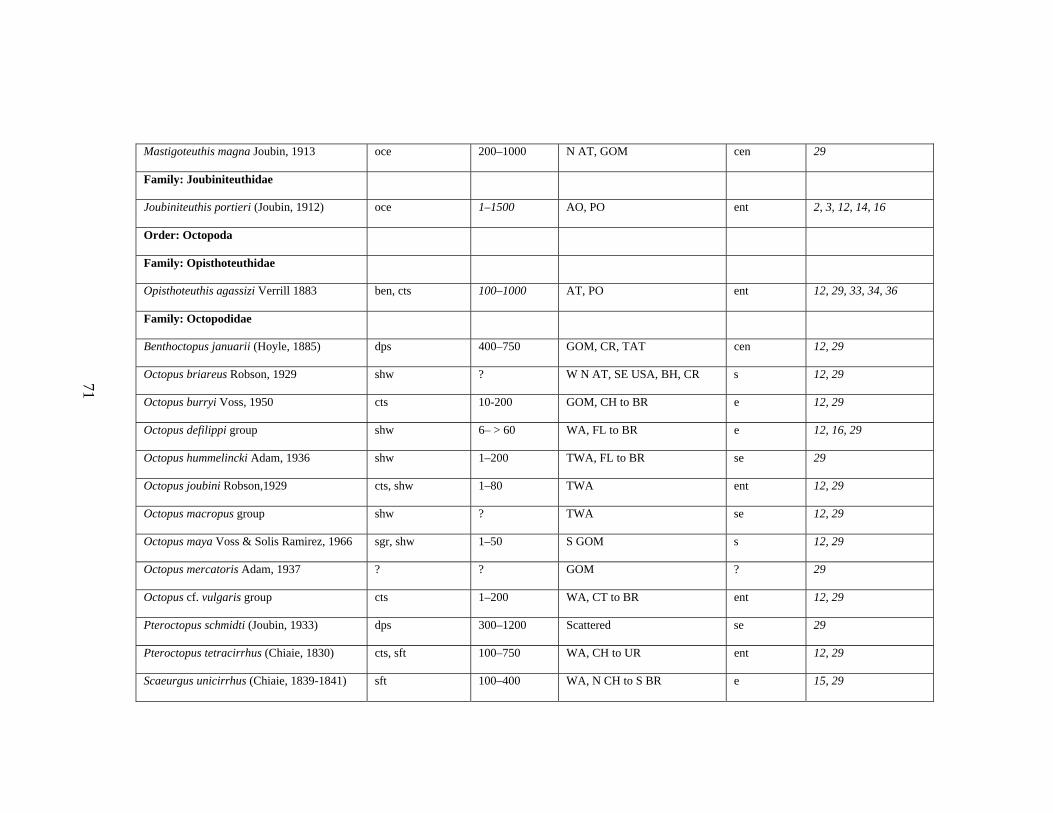
71
Mastigoteuthis magna Joubin, 1913 oce 200–1000 N AT, GOM cen 29
Family: Joubiniteuthidae
Joubiniteuthis portieri (Joubin, 1912) oce 1–1500 AO, PO ent 2, 3, 12, 14, 16
Order: Octopoda
Family: Opisthoteuthidae
Opisthoteuthis agassizi Verrill 1883 ben, cts 100–1000 AT, PO ent 12, 29, 33, 34, 36
Family: Octopodidae
Benthoctopus januarii (Hoyle, 1885) dps 400–750 GOM, CR, TAT cen 12, 29
Octopus briareus Robson, 1929 shw ? W N AT, SE USA, BH, CR s 12, 29
Octopus burryi Voss, 1950 cts 10-200 GOM, CH to BR e 12, 29
Octopus defilippi group shw 6– > 60 WA, FL to BR e 12, 16, 29
Octopus hummelincki Adam, 1936 shw 1–200 TWA, FL to BR se 29
Octopus joubini Robson,1929 cts, shw 1–80 TWA ent 12, 29
Octopus macropus group shw ? TWA se 12, 29
Octopus maya Voss & Solis Ramirez, 1966 sgr, shw 1–50 S GOM s 12, 29
Octopus mercatoris Adam, 1937 ? ? GOM ? 29
Octopus cf. vulgaris group cts 1–200 WA, CT to BR ent 12, 29
Pteroctopus schmidti (Joubin, 1933) dps 300–1200 Scattered se 29
Pteroctopus tetracirrhus (Chiaie, 1830) cts, sft 100–750 WA, CH to UR ent 12, 29
Scaeurgus unicirrhus (Chiaie, 1839-1841) sft 100–400 WA, N CH to S BR e 15, 29
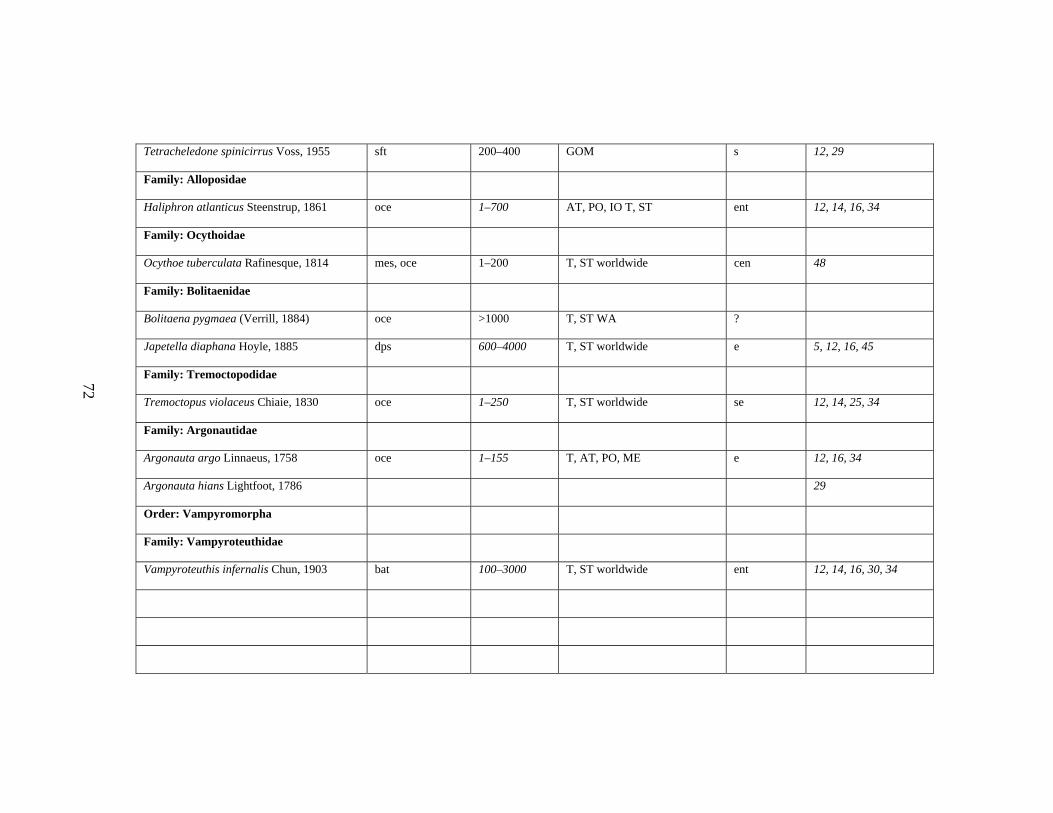
72
Tetracheledone spinicirrus Voss, 1955 sft 200–400 GOM s 12, 29
Family: Alloposidae
Haliphron atlanticus Steenstrup, 1861 oce 1–700 AT, PO, IO T, ST ent 12, 14, 16, 34
Family: Ocythoidae
Ocythoe tuberculata Rafinesque, 1814 mes, oce 1–200 T, ST worldwide cen 48
Family: Bolitaenidae
Bolitaena pygmaea (Verrill, 1884) oce >1000 T, ST WA ?
Japetella diaphana Hoyle, 1885 dps 600–4000 T, ST worldwide e 5, 12, 16, 45
Family: Tremoctopodidae
Tremoctopus violaceus Chiaie, 1830 oce 1–250 T, ST worldwide se 12, 14, 25, 34
Family: Argonautidae
Argonauta argo Linnaeus, 1758 oce 1–155 T, AT, PO, ME e 12, 16, 34
Argonauta hians Lightfoot, 1786 29
Order: Vampyromorpha
Family: Vampyroteuthidae
Vampyroteuthis infernalis Chun, 1903 bat 100–3000 T, ST worldwide ent 12, 14, 16, 30, 34

73
Table 3.2: Taxonomic summary for Cephalopods of the Gulf of Mexico. Component Subgroups Spirulida Sepiodea Myopsida Oegopsida Octopoda Vampyromorpha
Total Species 1 5 6 58 22 1
Number Endemic Not enough study yet to give good account
Number Non-indigenous species Only 2 known at this time 1 1

74
CHAPTER 4
FIRST RECORDS OF Asperoteuthis acanthoderma (Lu, 1977) (CEPHALOPODA:
OEGOPSIDAE: CHIROTEUTHIDAE), FROM THE NORTH ATLANTIC OCEAN,
STRAITS OF FLORIDA
Heather Judkins, Debra A. Ingrao and Clyde F. E. Roper (HJ) College of Marine Science, University of South Florida, 140 7th Ave South, St.Petersburg, FL 33701; [email protected] (DI) Mote Marine Laboratory, 1600 Ken Thompson Parkway, Sarasota, FL 34236; [email protected] (CR) Musuem of Natural History, Smithsonian Institution, Washington D.C. 20013-7012 USA; [email protected] 4.1 Abstract
The first observation in the North Atlantic Ocean of the deep sea squid
Asperoteuthis acanthoderma (family Chiroteuthidae) is reported here from off the coast
of Key West, Fl in the Straits of Florida. We describe the morphology of the two nearly
complete, but damaged, specimens. A third record is based on photographs of a specimen
from off Grand Cayman Island; this specimen was not available for examination. The
multiple occurrences of this species, heretofore unknown in the North Atlantic Ocean,
within a 10-month period are so unusual that we attempt to hypothesize an explanation
for these events. All previously known records are recorded from a few specimens

75
scattered from Hawaii to the Philippines. The present specimens were identified by the
following characteristics unique to the species: Y-shaped funnel locking apparatus,
sucker ring form and dentition, beak morphology, photophore patch configuration on
ventral surface of eyeballs, and numerous, small cartilaginous tubercles that cover the
mantle, head and the aboral surface of the arms.
4.2 Introduction
A mature female Asperoteuthis acanthoderma was discovered approximately 90
miles southwest of Key West, Florida on February 20, 2007. It was found floating at the
surface above a bottom depth of approximately 259 meters. The specimen was retrieved
by a charter boat captain, Clint Moore, who recognized the uniqueness of the squid and
donated it to Mote Marine Laboratory in Sarasota, Florida. This specimen is now located
in the cephalopod collection of the Smithsonian Institution’s National Museum of Natural
History in Washington D.C. A second specimen was found by Captain Jack Carlson in
the Straits of Florida, 16 miles east of Marathon, Florida in June 2007, floating at the
surface above a water depth of approximately 355m (24˚43’N 81˚06W). This specimen
is deposited in the Marine Invertebrate Museum at the Rosenstiel School of Marine and
Atmospheric Sciences, University of Miami, in Miami, Florida (UMML 31.3212).
The posterior portion of the primary fin is missing in the Key West specimen, so
the mantle length (ML) is measured from the anterior tip of the mantle, along the dorsal
mid-line, to the posterior end of the existing portion of the fins. Also missing are both
tentacular clubs and stalks, with the exception of a very short section of the left tentacle.
Internal structures are in relatively good condition, including the female reproductive

76
organs, so that the principal specific characters are present except for those found on the
tentacular clubs.
The Marathon specimen compliments the Key West specimen by having more
complete tentacles that show the photophore patches along their lengths, and it retains the
majority of the pair of flaps known as the “secondary fin” or “tail flap” along the sides of
the tail. The tentacular clubs and eyes are missing. Although the internal organs are
damaged, sufficient evidence remains to indicate that the specimen is a spent female.
4.3 Systematics
Asperoteuthis acanthoderma (Lu, 1977)
Diagnosis.---A chiroteuthid with mantle, head and arms covered with numerous, minute
cartilaginous tubercles; photophore patch present on ventral surface of eyes; 3-4 broad,
rounded teeth on arm-sucker rings; funnel locking apparatus an inverted Y-shape. (Lu,
1977)
Material examined.---Specimen 1. (Fig. 1) Female, 620+ ML, 90 miles southwest of
Key West, FL. Found floating dead at surface. The posterior tip of the mantle, primary,
and secondary fins are missing. When approximate measures are used, the TL is
estimated to have been 1250 mm. Specimen 2.(Fig.2) Female, 1630 mm ML, and 3420
TL; measured to tip of tail posterior to “tail flap”.. Found floating dead at surface, 16
miles off Marathon, FL.
4.4 Description
External anatomy.---The description is based primarily on the Key West
specimen with additional details included from the Marathon specimen as appropriate.
The mantle is long, narrow, semigelatinous, with dark purple pigmentation. Numerous,

77
minute cartilaginous tubercles cover the mantle, head and arms. The mantle is tapered
posteriorly towards the fins. The fins together form an elongate oval shape and are
estimated to have been 220 mm long (including estimated length of missing portion). The
fin length is therefore approximately 50% of ML. The width of the fins is 340 mm at their
widest part. The tail with its posterior extension of the gladius and lateral structures is
missing altogether. The Marathon specimen had a nearly complete fin assemblage with
only the very tip of the tail missing. The fin length measures 460 mm, the tail is 610 mm
long. The fin width measures 330 mm and the secondary fin width is 235 mm. The fins
and tail flaps are each elongate oval-shaped. The fins become proportionately narrower
posteriorly. The tail flaps are very thin, while the fins are noticeably thicker.
The funnel is large in comparison to the head. It measures 173 mm from the
anterior funnel opening to the posterior border along the ventral midline. The diameter of
the anterior funnel aperture measured (flattened) is 86 mm. The funnel locking apparatus
is ovoid and is in an inverted Y-shape; the mantle component matches the funnel
counterpart. The dorsal component of the funnel organ is roughly diamond shaped with
one triangular flap on each side. The ventral pads are present and oval shaped. The flaps
form a skirt-like sheath that is attached to the dorsal surface of the funnel organ
suggesting that this organ hangs ventrally open while the animal is alive. The funnel
valve is very large.
The head is elongate, narrow; it is deep dorsoventrally, narrow laterally.
Olfactory papillae are present on each side of the postero-lateral surface of the head in the
form of a slender stalk with a bulbous terminal head. The eyes are large with a ventral
photophore patch that extends toward the posterior surface of each eye. Diameter of right

78
eye, 28mm; horizontal opening of the eyes: left, 44 mm and right, 37 mm. The nuchal
cartilage is long and narrow; the central groove is distinct.
The arms are very long and attenuate. Little difference is apparent among the
lengths of arms, but only two are completely intact. The intact AL of right arm I is 880
mm and of intact left arm IV, 870 mm. All arms have approximately the same semi-
gelatinous consistency and appear to be closely equal in diameter at their bases. Right
arm III has a distinct keel distally, but no other keels are omm. able. All arms have
trabeculae along the oral bases from which the sucker stalks arise. The arm suckers are
biserial and evenly spaced. The diameters of the suckers range from 4 mm to 12 mm on
each arm, varying slightly between arms but consistently patterned small- enlarged- small
as they progress distally. On the arms that are intact, the tips have minute suckers on
small stalks, clustered together much more closely than along the rest of arms. The arm
sucker rings have 3 to 4 broad, rounded teeth around the rim.
The remnants of the tentacles of the Key West specimen have only short,
extremely thin stalks that extend from the arm crown. The longest remaining portion is
on the right side and it measures 100 mm with no visible photophore patches. The
Marathon specimen has larger tentacle remanants measuring 1560 mm on the left side
and 780 mm on the right side. No tentacular clubs are present. The tentacles have
photophore patches scattered along their length in no discernable pattern. Forty such
patches occur on the most intact (left) tentacle of the Marathon specimen.
The gladius was not extracted from the Key West specimen because it is broken
in many places. The gladius of the Marathon specimen is in much better condition. The
gladius is approximately 1630 mm long and 74 mm wide at its widest point. The most

79
anterior portion of the gladius is flattened and becomes rounded as it extends to the
posterior end. The gladius could not be completely reconstructed because pieces are
missing.
The buccal membrane has 7 connectives that attach on the dorsal sides of the
bases of arms I and II and on the ventral sides of arms III and IV. The lappets are too
indistinct to be counted.
The beaks were extracted from the Key West specimen for examination and
preservation. The anterior tip of the upper mandible is narrow, sharply pointed. It has a
deep, dark brown pigmentation. The hood is normal in size and has a brownish
coloration that lightens toward the edges, which are translucent in appearance. The curve
of the dorsal half of the hood margin of the beak is irregularly shaped, not smooth. The
anterior tip of the lower beak also is pointed with its inner edges curved. It is somewhat
broader than the upper mandible, with dark brown pigmentation and the edges of the
wings are a lighter brown in tone.
The radula has 7 teeth per transverse row. The width at the dorsal side of the
radula measures 4 mm, while the ventral portion of the set measures 3.5 mm. The length
of the radula is 14 mm. The rhachidian tooth has a long central cusp with shorter, lateral
cusps. The first lateral tooth is bicusped, with a long, pointed cusp and a shorter, blunt
lateral cusp. The second lateral tooth is pointed with a broad base but no lateral cusp.
The third lateral tooth is longer than the other lateral teeth and is pointed and curved.
Most of the surface of the squid is covered with cartilaginous tubercles. They are
very numerous and thickly-concentrated on the head, mantle and aboral surfaces of arms.
There appear to be fewer on the oral surfaces of arms. Each tubercle has a wide base that

80
ends with a pointed cartilaginous tip. Numerous dark purple chromatophores cover the
squid, as well. Histological sections of the tubercles of the mantle wall indicate that they
are similar to a published description (Roper and Lu, 1990), as well as in the original
description of the mantle by Lu (1977) which states that there is an outer vacuolated
dermal layer followed by a longitudinal muscle layer, an exceptionally thick vacuolated
medullary layer, an inner circular muscle layer and the lining of the mantle cavity.
Internal anatomy.---The internal organs of the Key West specimen are in
relatively good condition, while the organs from the Marathon squid are significantly
deteriorated. Key West specimen (Fig. 3): The gladius extends the length of the mantle
on the internal dorsal side. The gills extend approximately half the length of the mantle
cavity on both sides, lateral to the digestive gland. The ink sac is small and appears to be
empty; it was difficult to locate during dissection. The nidamental glands lay medial to
the gills. They are paired, oval, enlarged and milky white in appearance; the right gland
is damaged by a tear. The branchial hearts are small, white, and round; they are located
posterior to the kidney at the base of each gill with the systemic heart lying along the
central midline, dorsal to the nidamental glands. The digestive organs are intact; the
stomach, thin walled, and collapsed, leads to the caecum, which is creamy white in color,
and thick with an ovoid spongy, reticulated mass. The ovary is posterior to the digestive
organs, translucent and appears empty of oocytes and eggs. All organs appear
proportional in size compared with mantle length, with the exception of the substantially
enlarged nidamental glands.

81
4.5 Discussion
The first species of Asperoteuthis was originally desribed as Chiroteuthis
acanthoderma by Lu (1977) from the tropical western Pacific. Nesis (1980) moved this
species to the new genus Asperoteuthis. He considered C. acanthoderma Lu, 1977 to be
a junior synonym of Aspreoteuthis famelica (Berry 1909), described from a damaged
specimen caught near Hawaii. Subsquent studies found that A. famelica was actually
Mastigotuthis famelica (Young, 1978), so the valid name for Lu’s specimens is
Asperoteuthis acanthoderma (see Young et al. 2007, Arkhipkin et. al, 2008). We identify
the present specimens as Asperoteuthis acanthoderma because of the unique
characteristics that exist based on Lu’s description of the species.
The genus Asperoteuthis is distinguished from the other genera of the family
Chiroteuthidae by the presence of a terminal photophore on the tentacular clubs and an
elongate oval photophoric patch on the ventral surface of each eyeball; the absence of
intestinal photophores and of a tragus/antitragus in the funnel component of the locking
apparatus.
Other features that distinguish Asperoteuthis from other chiroteuthids include 1)
the structure of arm IV is similar to that of arms I-III, not significantly enlarged in
advanced subadults; 2) absence of characteristic pairs of adjacent club suckers drawn
together with an intermediate widening on the stalk and absence of an enlarged, central
tooth on the club suckers as occurs in other chiroteuthids; and 3) the forked (Y-shaped)
funnel cartilage
Asperoteuthis acanthoderma (Lu, 1977) is characterized by the possession of
numerous, minute, sharply pointed, conical cartilaginous tubercles distributed over the

82
entire surface of the mantle, head, and arms. The tubercles measure 1 mm in diameter
and 0.4 mm in height from a specimen of 144 mm ML. The tubercles are discrete
structures, and their bases are embedded in the dermis of the mantle (Roper & Lu 1990,
as C. acanthoderma). The small number of rounded teeth on the arm suckers (3-4) and
the prominence if the funnel valve (Tsuchiya & Okutani 1993) are additional specific
characteristics.
The morphological characteristics of our specimens, that are about 4 times larger
than the holotype and paratype, conform closely with the original description, which was
based on females of 188 mm and 144.5 mm ML. Exceptions are minor, such as the
inconspicuousness of the dermal tubercles on the smaller specimens, and the number of
knobs on the tentacular stalks. We concur with Tsuchiya and Okuntani, (1993) that these
and other minor differences are attributable to variation due to the size and stages of
maturity of specimens.
Distribution.---Asperoteuthis acanthoderma has been recorded in the central and
western Pacific and Indo-Pacific waters as follows: in the southern part of Philippine Sea
and in the Celebes Sea to the west, southeast, and east of the Philippine Islands by Lu
(1977) and Nesis (1980), (as Chiroteuthis acanthoderma and Mastigoteuthis famelica,
respectively). A specimen of A. acanthoderma was recorded near the bottom by a
SERPENT-Project ROV off western Australia (18˚30’S, 115˚30’E) at 580 m depth in
April, 2005 (Vecchione pers. omm..).
A recent survey of literature of the cephalopods of the Gulf of Mexico revealed
that only three genera of Chiroteuthids previously have been recorded there: Chiroteuthis
joubini, Chiroteuthis veranyi, and Chiroteuthis capensis, as well as Grimalditeuthis

83
bonplandi (Verany, 1839) and Plancoteuthis danae (Joubin, 1931) (Judkins et. al, in
press). The present report of A. acanthoderma is therefore a dramatic range extension
into the North Atlantic.
Information on the vertical distribution of Asperoteuthis acanthoderma is limited,
but the following capture data indicate that this is a deep-sea species (as also
demonstrated by its morphology and rarity of capture):
Holotype.---914-0 m (Lu 1977).
Paralarvae.---Daytime 600-1100 m and night 700-925 m (Roper & Young 1975).
Night on horizon of 200-300 m, (Lu 1977, Nesis 1980). Based on the limited
information, we believe that the vertical movements of A. acanthoderma may not be
ontogenetic descent but diel vertical migrations as Nesis (1980) suggested. This is
similar to other chiroteuthids (Roper & Young 1975).
The state of knowledge about the chiroteuthids is expanding as teuthologists
discover and report new findings of genera and species. For example, Young et. al.
(2007) published information of a new Asperoteuthis species found off the coast of
Hawaii. Asperoteuthis mangoldae (Young, Vecchione, & Roper, 2007) shares some A.
acanthoderma characters but differs in the absence of tubercles in the skin; a more
gelatinous consistency of tissues; the absence of an anti-tragus in the funnel locking
apparatus; 8-10 slender, truncated teeth on the arm sucker rings; club and club
photophore structure and fin length characteristics. Arkhipkin and Laptikhovsky (2008)
introduced a fourth species to the Asperoteuthis genus, Aspreoteuthis nesisi which shares
many characteristics of A. acanthoderma, such as the funnel locking apparatus, the
numerous tubercles on head and mantle, and the photophore patches on eyes.

84
Asperoteuthis acanthoderma may be a circumglobal species whose broad
distribution has just recently been recognized because of fortuitous findings of specimens
and intensified research in previously under-sampled study areas. The global
understanding of the oceanic environment and its inhabitants is enhanced as those who
participate in maritime activities, deep sea fishermen, for example, become more
cognizant of their surroundings and the organisms that inhabit their specific regions. Our
knowledge is advanced when they report their observations and findings to marine
scientists.
The two specimens we examined from off Florida and the one record via photos
of A. acanthoderma in Grand Cayman suggest that certain physical parameters of the
region may be different from earlier years. All three specimens were found within a 10-
month period from mid-2006 to May 2007, rare discoveries that seem to be more than
coincidental. Even though they appear to be spent females that could have drifted to the
surface, a typical event for many deep sea squids and octopods, the total absence of
specimens in the North Atlantic Ocean until now could suggest additional causative
effects.
Based on our detailed examination of the two large, nearly intact specimens, and
our comparisons with descriptions in the literature, we conclude that our specimens are
indeed A. acanthoderma. Consequently, this species is now shown to occur not only in
the tropical western Pacific Ocean, but also in the Caribbean/Gulf of Mexico/Straits of
Florida region.

85
4.6 Acknowledgments
Photos were taken by Chuck Stevenson at Mote Marine Laboratory, for which we are
grateful. We acknowledge the enormous help of the faithful Mote Marine lab volunteers
as the dissections were conducted. We also thank the two captains from the fishing
vessels that made the original finds and contacted us. Captain Clint Moore found the Key
West specimen on February 20, 2007, while Captain Jack Carlson discovered the
specimen off Marathon, FL on May 17, 2007; He coordinated with Gabe Delgado of the
Florida Wildlife Research Institute to ensure retrieval of the specimen. We appreciate the
help of Mike Vecchione, NMFS Systematics Laboratory, Natural Museum of Natural
History, Washington D.C., and Nancy Voss, Rosenstiel School of Marine and
Atmospheric Science, and their expertise during the examination of material and the
preparation of this manuscript. Heather Judkins acknowledges the College of Marine
Science at the University of South Florida for the opportunity to pursue her scientific
goals. Clyde Roper acknowledges with appreciation the association with and support of
Mote Marine Laboratory as an adjunct scientist.

86
REFERENCES
1. Judkins, H, M. Vecchione, & C. F. E Roper.In press. Cephalopods (Mollusca: Cephalopoda) of the Gulf of Mexico. Gulf of Mexico Origin, Waters and Biota Biodiversity, Volume 1 (Darryl L. Felder and David A. Camp, editors, Texas A&M University Press, College Station, Texas).
2. Lu, C. C. 1977. A new species of squid, Chiroteuthis acanthoderma, from the
Southwest Pacific (Cephalopoda: Chiroteuthidae). Steenstrupia, Zoological Museum of University of Copenhagen; 4:179-188.
3. Nesis, K. N. 1980. Taxonomic Position of Chiroteuthis famelica Berry
(Cephalopods, Oegopsida). Byulleten Moskovskogo Obsestva Ispytatelei Prirody (Otdel Biologicheskii); 85 (4): 59-66.
------. 1982. Cephalopods of the World, Squids, Cuttlefishes, Octopuses, and Allies. T.F.H. Publications, Inc. 385 pp.
4. Roper, C. F. E. and R. E. Young, 1975. Vertical Distribution of Pelagic Cephalopods.
Smithsonian Contributions to Zoology, no. 209: 51pp. ------ & C. C. Lu, 1990. Comparative Morphology and Function of Dermal Structures in
Oceanic Squids (Cephalopoda). Smithsonian Contribution to Zoology, no. 493: 41 pp.
5. Tsuchiya, K. and T. Okutani. 1993. Rare and interesting squids in Japan, recent
occurrences of big squids from Okinawa. Venus, 52 (4): 299-311. 6. Young, R. E., 1991. Chiroteuthid and related paralarvae from Hawaiian waters.
Bulletin of Marine Science, 49(1-2): 162-185. ------ & C. F. E. Roper. 1999. Asperoteuthis acanthoderma (Lu, 1977). Version 01
January 1999. (under construction). http://tolweb.org/Asperoteuthis acanthoderma/19466/1999.01.01 in the Tree of Life Web Project, http://tolweb.org/
------, M. Vecchione, & C. F. E. Roper. 2007. A new genus and three new species of decapodiform cephalopods (Mollusca: Cephalopoda). Review of Fish Biology and Fisheries, 17:353-365.

87
4.8 FIGURE LEGENDS
Fig. 1. Key West Asperoteuthis acanthoderma specimen.
Fig. 2. Marathon Asperoteuthis acanthoderma specimen.
Fig 3. Internal organs of Key West specimen: A. digestive gland B. gill; C. nidamental glands; D. branchial heart; E. stomach; F caecum; G. ovary.
Manuscript Note.---After the manuscript was completed, we received information that an additional specimen of Asperoteuthis had been found floating at the surface by a fisherman five miles south off Little Cayman Island in the northern Caribbean Sea on 18 May 2008. It was donated to the Little Cayman Research Center for identification by station manager, Jon Clamp, who in turn, contacted us for positive identification via photographs. The damaged specimen had a mantle length of approximately 152.4 cm when collected. The specimen will be deposited at the Smithsonian Institution along with the Key West specimen once additional studies have been conducted. We thank Jon Clamp, Judie Clee, and the fisherman who discovered the specimen for their efforts to insure that the specimen was made available for study and permanent deposition.
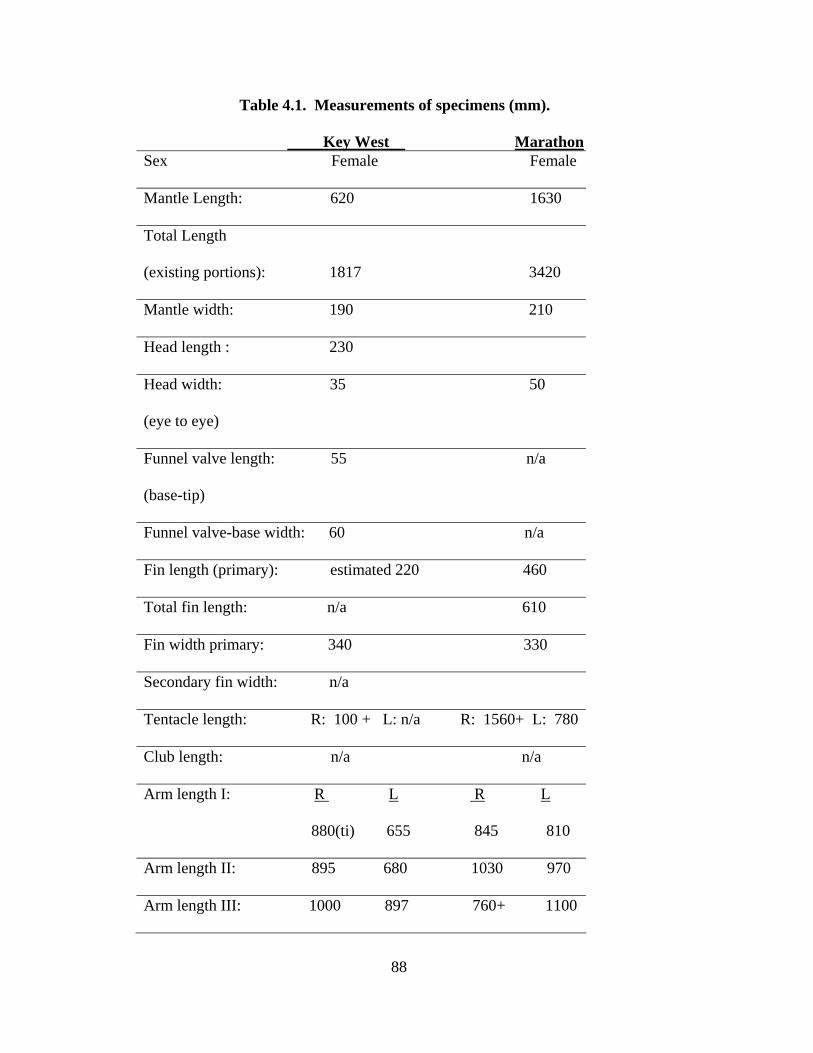
88
Table 4.1. Measurements of specimens (mm).
Key West Marathon Sex Female Female
Mantle Length: 620 1630
Total Length
(existing portions): 1817 3420
Mantle width: 190 210
Head length : 230
Head width: 35 50
(eye to eye)
Funnel valve length: 55 n/a
(base-tip)
Funnel valve-base width: 60 n/a
Fin length (primary): estimated 220 460
Total fin length: n/a 610
Fin width primary: 340 330
Secondary fin width: n/a
Tentacle length: R: 100 + L: n/a R: 1560+ L: 780
Club length: n/a n/a
Arm length I: R L R L
880(ti) 655 845 810
Arm length II: 895 680 1030 970
Arm length III: 1000 897 760+ 1100
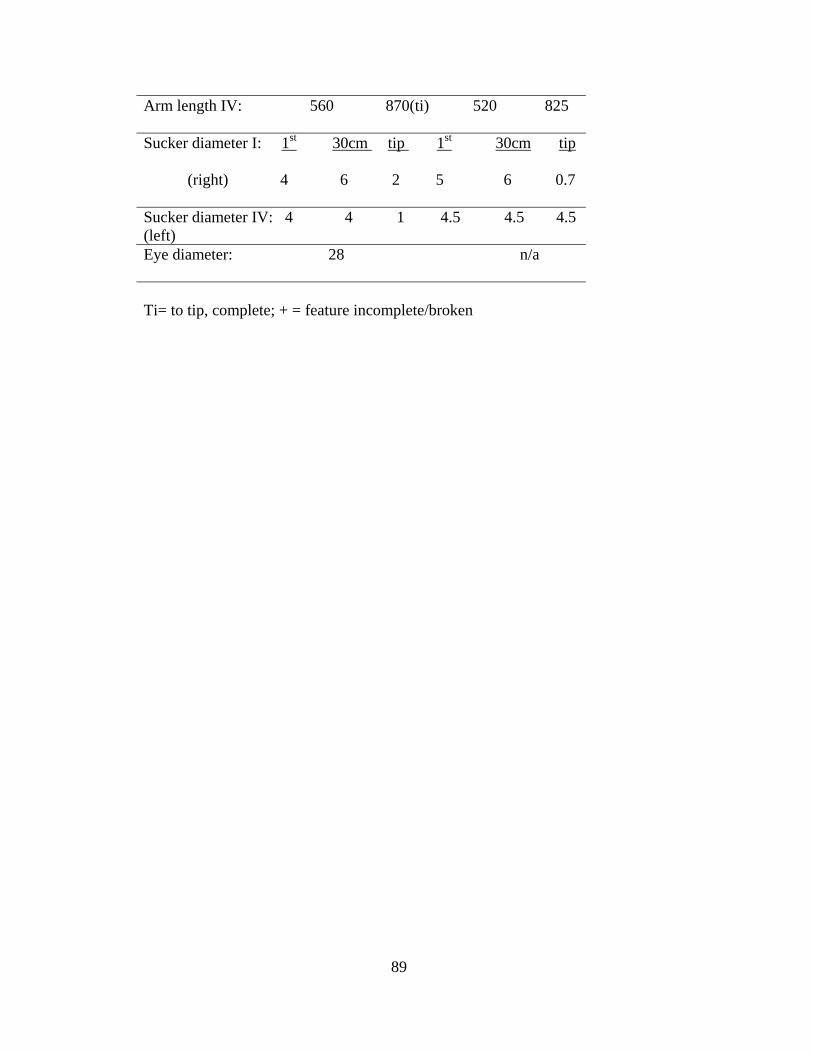
89
Arm length IV: 560 870(ti) 520 825
Sucker diameter I: 1st 30cm tip 1st 30cm tip
(right) 4 6 2 5 6 0.7
Sucker diameter IV: 4 4 1 4.5 4.5 4.5 (left) Eye diameter: 28 n/a
Ti= to tip, complete; + = feature incomplete/broken

90
Figure 4.1. Key West Asperoteuthis acanthoderma specimen.

91
Figure 4.2. Marathon Asperoteuthis acanthoderma specimen.

92
Figure 4.3. Internal organs of Key West specimen: A. digestive gland B. gill; C. nidamental glands; D. branchial heart; E. stomach; F caecum; G. ovary.

93
CHAPTER 5
CONCLUSIONS Summary
The cephalopods of the Broad Caribbean are widely distributed with
concentrations along the continental shelves. There are gaps in the collections from the
western Gulf of Mexico and the Caribbean Sea, both nearshore and offshore, which are
important to investigate further. Distribution maps (Appendix C) display a wide variety
of localities that the cephalopods occupy with island nations hosting many octopod
species.
According to the present study, Rapoport’s Rule does not apply to the
cephalopods of the Broad Caribbean. This finding is supported by a literature survey
conducted by Rosa in 2008 (Rosa et al., 2008) describing species richness of cephalopods
along the Atlantic coast. Cephalopod species richness increased with increasing latitude
contrary to Stevens’ (Stevens, 1989) belief that species richness increases towards the
equator. The search for diversity “hotspots” of the Broad Caribbean was not conclusive
in most instances as the sample sizes were too small to allow the rarefaction curves to
reach asymptote. According to the analysis, region 4, the eastern coast of Florida
exhibited the highest species richness (n=32). Species richness may be higher here
because of increased nutrient mixing in the Gulf Stream, patterns of current transport in
the region and presence of both shallow and deep water habitats existing along the
eastern Florida coast. Collection intensity could have been an attributing factor to

94
increased richness although the majority of the rarefaction curves were headed towards
an asymptote. The low sample size in the southeastern Caribbean Sea (n=4) displays the
need for increased study to enrich the cephalopod knowledge base in that region.
The Gulf of Mexico species checklist exposes the advantages and pitfalls of
creating species checklist studies today based on literature alone. The literature provided
an excellent collection of species that are found within the region, dating back to the
1800’s. However, when it is compared to the actual data collected in chapter 2 of this
study, discrepancies were discovered.
Table 5.1: Gulf of Mexico Checklist Species not included in Ch II:
1. Brachioteuthis sp. 10. Chiroteuthis veranyi 2. Leachia cyclura 11. Chiroteuthis mega 3. Cycloteuthis sirventyi 12. Pholidoteuthis boschmai 4. Discoteuthis laciniosa 13. Octopoteuthis sicula 5. Histioteuthis bonnelleii 14. Mastigoteuthis magna 6. Chtenopteryx sicula 15. Octopus mercatoris 7. Lepidoteuthis grimaldii 16. Pteroctopus schmidti 8. Lampadioteuthis megaleia 17. Ocychoe tuberculata 9. Chiroteuthis joubini
Seventeen species (Table 5.1) were found in the Gulf of Mexico checklist were
not found in the biogeography study described in chapter 2. If the biogeographical
results are combined with the checklist, the result is a total of 127 cephalopod species that
are found in the Broad Caribbean region. When the species from Vecchione’s work
(2002) is merged in, a total of 131 species exist in the Broad Caribbean. Although the
study presented a few discrepancies in the Gulf of Mexico species totals, the overall
picture is one of large species diversity throughout the region. A factor to consider is that
some of the checklist species are based on one or two specimens which were badly

95
damaged animals. The paucity of species samples demonstrates the need to investigate
the Broad Caribbean thoroughly for additional samples to ascertain the validity,
abundance, and range limits of species distribution. The 18 range extensions reported
here are of value for understanding how cephalopods fit into the ecosystems of this area.
Many facets are needed to create the “complete” picture in terms of marine ecosystems
and the role that the cephalopods play in them. Neither data nor literature alone will
complete the picture for any diverse marine group.
The deep-dwelling squid, Asperoteuthis acanthoderma was an exciting new find
for the Broad Caribbean and illustrates how little is known about the distribution of deep-
water cephalopods. The species was only found off the coast of Japan prior to this
discovery. Further research may reveal that A. acanthoderma’s distribution is
circumglobal. To find and describe a new species of large squid after extensive studies
have been conducted in the region (G. Voss 1956, Lipka 1975, Nesis 1975, Roper et al.
1984, N. Voss et al. 1988, Passarella, 1991, Vecchione 2002) reveals the likelihood of
finding new cephalopods in the Broad Caribbean.
A recent paper (Norman & Hochberg, 2005) examined the status of octopods
worldwide. There have been new discoveries in many regions of the world including
New Zealand, New Caledonia, South Africa, and Hawaii. There is an obvious absence of
octopod findings in the Broad Caribbean region. G. Voss (1977) calculated the projected
amount of octopod species that would be found on an annual basis, and suggested that an
average of 6.7 species being described annually worldwide. He also stated that “it seems
clear that we are still in the descriptive stage of systematics”. These statements seem to
hold true over 25 years later (Norman & Hochberg, 2005). The study went on to describe

96
the progress and impediments to taxonomic studies today. Progress is visible in
advanced electronic communication, better technology, and better international links.
Areas that are in need of improvement: poor retention of materials, few replicates, poor
curation, little support for taxonomic research, very few understudies to continue
progress, and museum curators are not being replaced as they retire, collections being
maintained but not active or growing, and lastly fewer primary field studies are being
undertaken (Norman & Hochberg, 2005).
Proper identification and collection will also provide better understanding of the
potential for cephalopod fisheries. Over 50% of the global cephalopod catch recorded by
the Food and Agricultural Organization (FAO) is not segregated into single species
categories, and this significantly reduces any value the data may have for population
assessment (Boyle & Boletzky, 1996) Collaboration between scientists throughout the
Caribbean would further quantify and identify important and scarcely known species.
Improved collection methods and new species specific research cruises are sorely
needed to discover what else lies in the deeper waters of the Caribbean Sea and Gulf of
Mexico. There is a demand for the information as finfish harvests collapse, the fisheries
will turn to cephalopods for commercial exploitation. An immediate priority for the
ocotopods is production of detailed and accurate descriptions of all species as the octopod
taxonomy needs major revision and stabilization (Norman & Hochberg 2005)
Bottom depth and vertical migration can be important elements in the cephalopod
lifestyle as many (e.g. Selenoteuthis scintillans, Spirula spirula) ascend nightly for
feeding (Roper and Young, 1975). Deep-sea cephalopod collection has been traditionally
difficult (Wormuth & Roper, 1983). Also, connections in food webs are sometimes

97
difficult to determine as the cephalopods need to break down their prey into small parts
prior to digestion and stomach contents are therefore difficult to identify. Vertical
migration of cephalopods and the role they play in the transfer of energy between the
layers are not well known. Less than half of the specimens with known locales of the
5190 examined had depth of collection recorded. The depth information available was
not adequate to describe specific depth ranges for the cephalopods in the region.
Another challenge in this study was the lack of detail on capture locations for
many specimens. Many had only regional information associated with the sample and
over 200 specimens were not used due to a complete absence of information (e.g.
“collected in the Caribbean Sea”). A universal standard for recording sample information
throughout the Broad Caribbean could improve sample details and would glean
consistent accuracy for research studies. The sample information should include the
following: latitude/longitude, date, time, depth, water temperature, identification (if
possible) and collection method used for capture.
A technique that has been successful in solidifying relationships between
cephalopod species is DNA analysis. An example of an organism to examine in the
Broad Caribbean for genetic relatedness could be Doryteuthis plei and determine the
genetic linkages between the northern and southern populations. Also, DNA
comparisons with cephalopod species from the eastern tropical Pacific could lead to
insights concerning the origins of the fauna. Past geological occurrences provide an
excellent backdrop to examine the origins for cephalopods in the region.
There have not been any previous comprehensive cephalopod studies of the Broad
Caribbean since the early work of Voss (1956) and there is little literature from other

98
similar-sized regions for comparison. There are many smaller scaled studies, and it may
be because cephalopod taxonomy is in flux that large biogeography studies have been
neglected. Cephalopods have been the focus of many studies in the Broad Caribbean
region and the results from the present research paper build and highlight additional
advances in the comprehensive overview of the group. Much work is still needed to
attain a better understanding of this complex cephalopod assemblage.

99
REFERENCES
1. Boyle, P.R. & Boletzky, S.V. 1996. Cephalopod populations, definition, and dynamics. Philosophical Transactions of the Royal Society, London. Series B. Biological Sciences; B351:985-1002.
2. Lipka, Douglas. 1975. The systematics and zoogeography of cephalopods from the
Gulf of Mexico. Dissertation, Texas A&M. August 1975; 1-347.
3. Nesis K.N. 1975. Cephalopods of the American Mediterranean Sea; English Translations of Selected Publications on Cephalopods; Editor M. Sweeney; Smithsonian Institutions Libraries; 1; 318- 358.
4. O’Dor, R.K., J.A. Hoar, D.M. Webber, F.G. Carey, S. Tanaka, H.R. Martins and F.M.
Porteiro. 1994. Squid (Loligo forbesi) performance and metabolic rates in nature. Physiology of Cephalopod Molluscs- Lifestyle and Performance Adaptations. Gordon and Breach Publishers. 163-177.
5. Passarella, K.C., 1990. Oceanic Cephalopod Assemblage in the Eastern Gulf of Mexico, Department of Marine Science. University of South Florida: St. Petersburg, Fl. 50pp.
6. Roper, Clyde F. E. and Richard E. Young. 1975. Vertical distribution of pelagic cephalopods. Smithsonian Contribution to Zoology; 209; 1-51.
7. Roper C.F.E, M.J. Sweeney & C.E. Nauen, 1984. FAO species catalogue. v.3 Cephalopods of the world. An annotated and illustrated catalogue of species of interest to fisheries. FAO Fish. Synop. (125) v3: 277p.
8. Rosa, Rui, Heidi M. Dierssen, Liliana Gonzalez, and Brad Seibel. 2008. Ecological
biogeography of cephalopod molluscs in the Atlantic Ocean: historical and contemporary causes of coastal diversity patterns; Global Ecology and Biogeography; 17: 600-610.
9. Semmens, Jayson M., Gretta T. Peel, Bronwyn M. Gillanders, Claire M. Walunda,
Elizabeth K. Shea, Didier, Jouffre, Taro Ichii, Kartsen Zumhotz, Oleg N. Katugin, Stephen C. Leporati and Paul W. Shaw. 2007. Approaches to resolving cephalopod movement and migration patterns. Review of Fish and Biological Fisheries; 17: 401-423.
10. Stevens, G.C., 1989. The Latitudinal Gradient in Geographical Range: How So
Many Species Coexist in the Tropics. The American Naturalist; 133(2): 240-256.

100
11. Vecchione, 2002. Cephalopods. Carpenter, K.E.(ed.) The living marine resources of the Western Central Atlantic. Volume 1: Introduction, molluscs, crustaceans, hagfishes, sharks, batoid fishes, and chimaeras. Cephalopods of Western Central Atlantic; FAO Species Identification Guide for Fisheries Purposes and American Society of Ichthyologists and Herpetologists Special Publication No. 5. Rome, FAO. 2002. 1-600.
12. Voss, G.L. 1956. A review of the cephalopods of the Gulf of Mexico. Bulletin of
Marine Science: Gulf and Caribbean; 6:85-178. 13. Wormuth, John H. and Clyde F.E. Roper. 1983. Quantitative sampling of oceanic
cephalopods by nets: problems and recommendations. Biological Oceanography; 2 (2-4) 357- 377.

101
APPENDICES
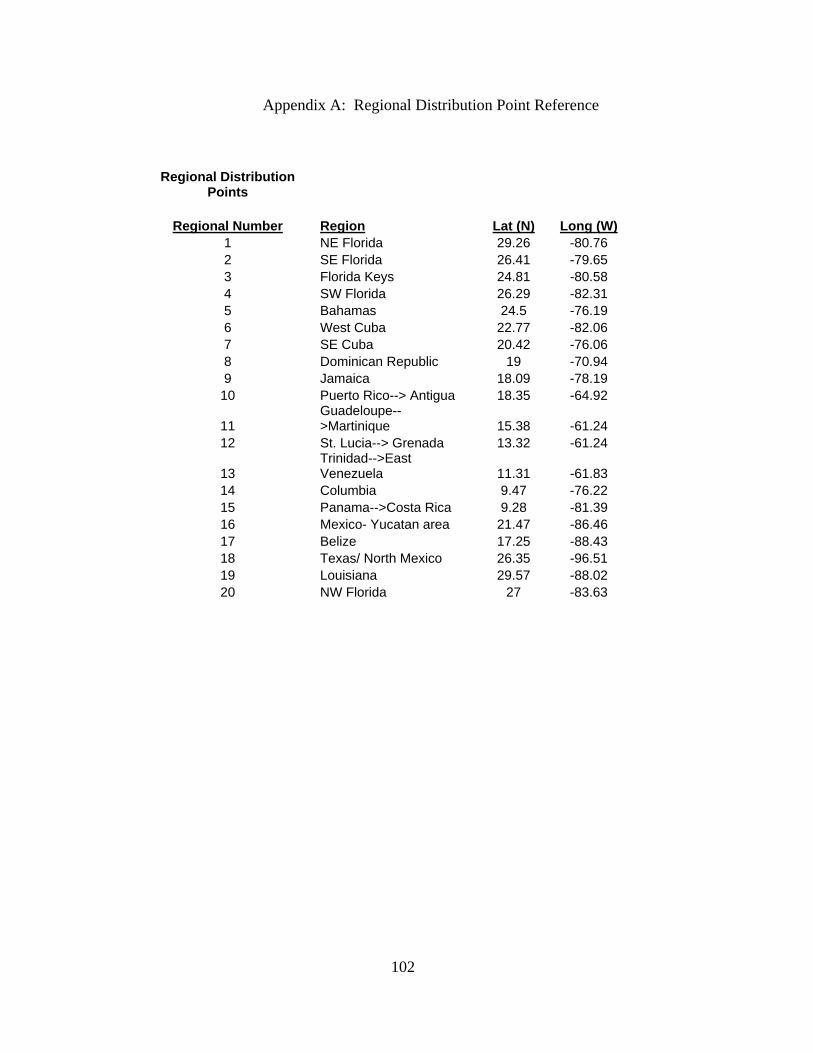
102
Appendix A: Regional Distribution Point Reference
Regional Distribution Points
Regional Number Region Lat (N) Long (W)
1 NE Florida 29.26 -80.76 2 SE Florida 26.41 -79.65 3 Florida Keys 24.81 -80.58 4 SW Florida 26.29 -82.31 5 Bahamas 24.5 -76.19 6 West Cuba 22.77 -82.06 7 SE Cuba 20.42 -76.06 8 Dominican Republic 19 -70.94 9 Jamaica 18.09 -78.19
10 Puerto Rico--> Antigua 18.35 -64.92
11 Guadeloupe-->Martinique 15.38 -61.24
12 St. Lucia--> Grenada 13.32 -61.24
13 Trinidad-->East Venezuela 11.31 -61.83
14 Columbia 9.47 -76.22 15 Panama-->Costa Rica 9.28 -81.39 16 Mexico- Yucatan area 21.47 -86.46 17 Belize 17.25 -88.43 18 Texas/ North Mexico 26.35 -96.51 19 Louisiana 29.57 -88.02 20 NW Florida 27 -83.63
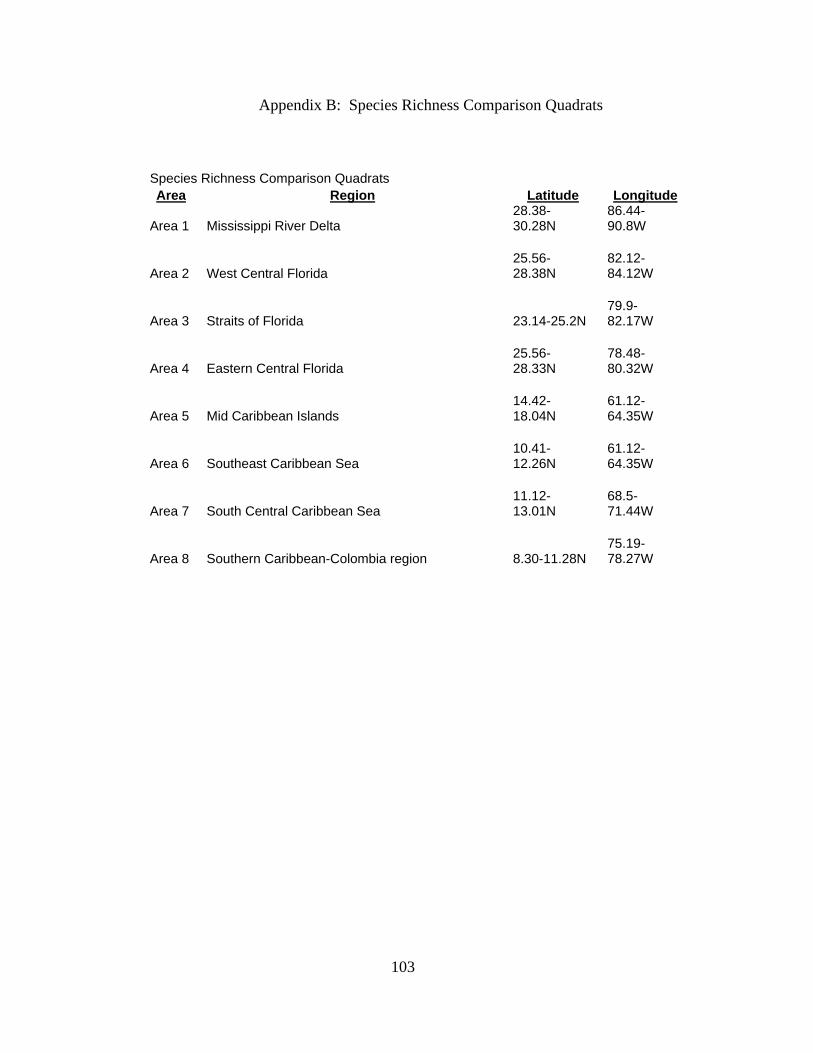
103
Appendix B: Species Richness Comparison Quadrats
Species Richness Comparison Quadrats Area Region Latitude Longitude
Area 1 Mississippi River Delta 28.38-30.28N
86.44-90.8W
Area 2 West Central Florida 25.56-28.38N
82.12-84.12W
Area 3 Straits of Florida 23.14-25.2N 79.9-82.17W
Area 4 Eastern Central Florida 25.56-28.33N
78.48-80.32W
Area 5 Mid Caribbean Islands 14.42-18.04N
61.12-64.35W
Area 6 Southeast Caribbean Sea 10.41-12.26N
61.12-64.35W
Area 7 South Central Caribbean Sea 11.12-13.01N
68.5-71.44W
Area 8 Southern Caribbean-Colombia region 8.30-11.28N 75.19-78.27W
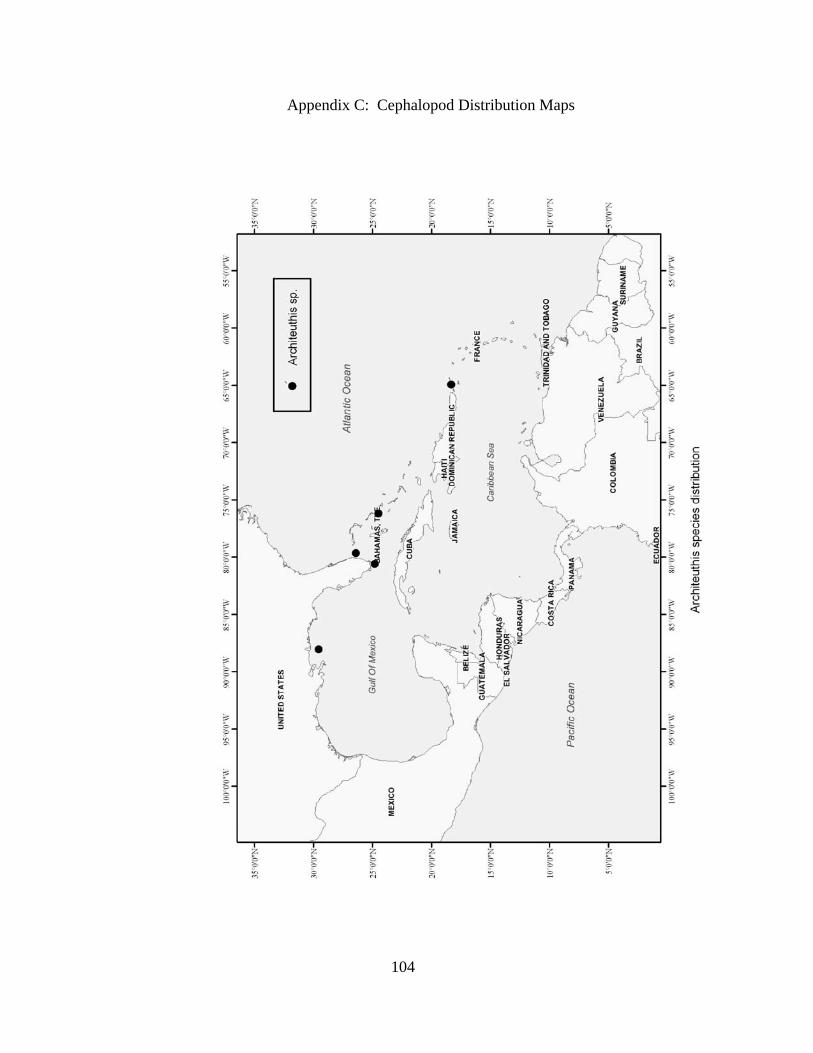
104
Appendix C: Cephalopod Distribution Maps
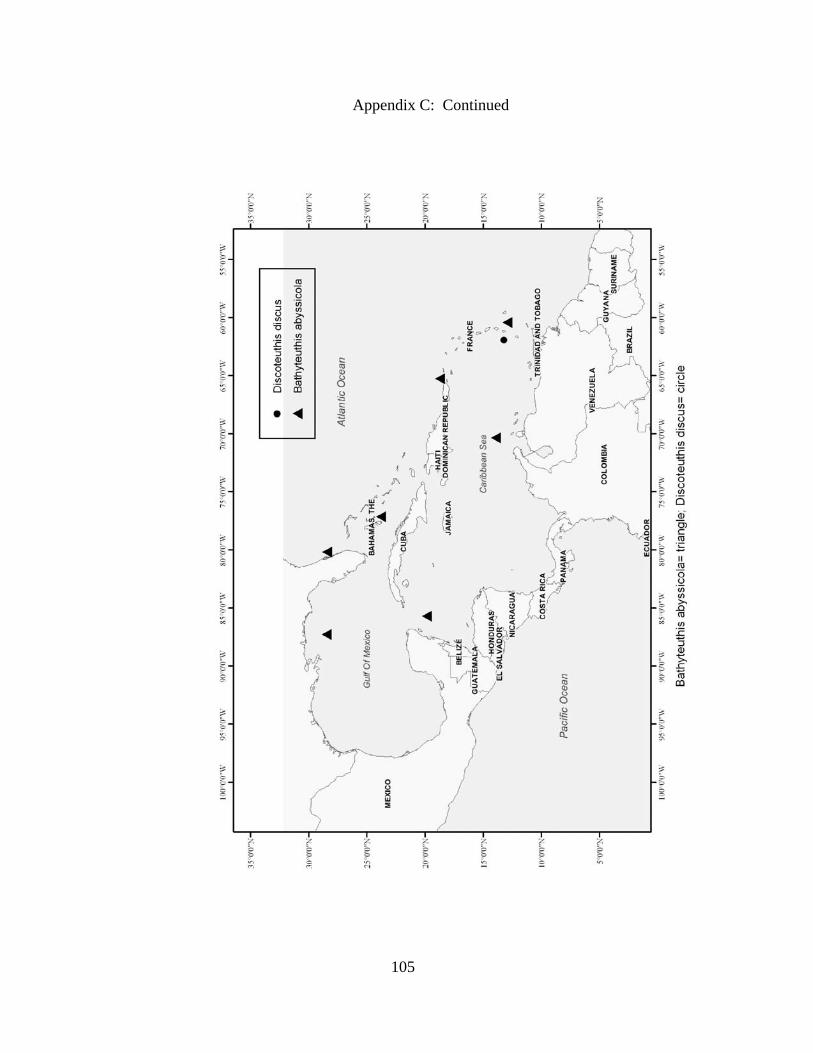
105
Appendix C: Continued
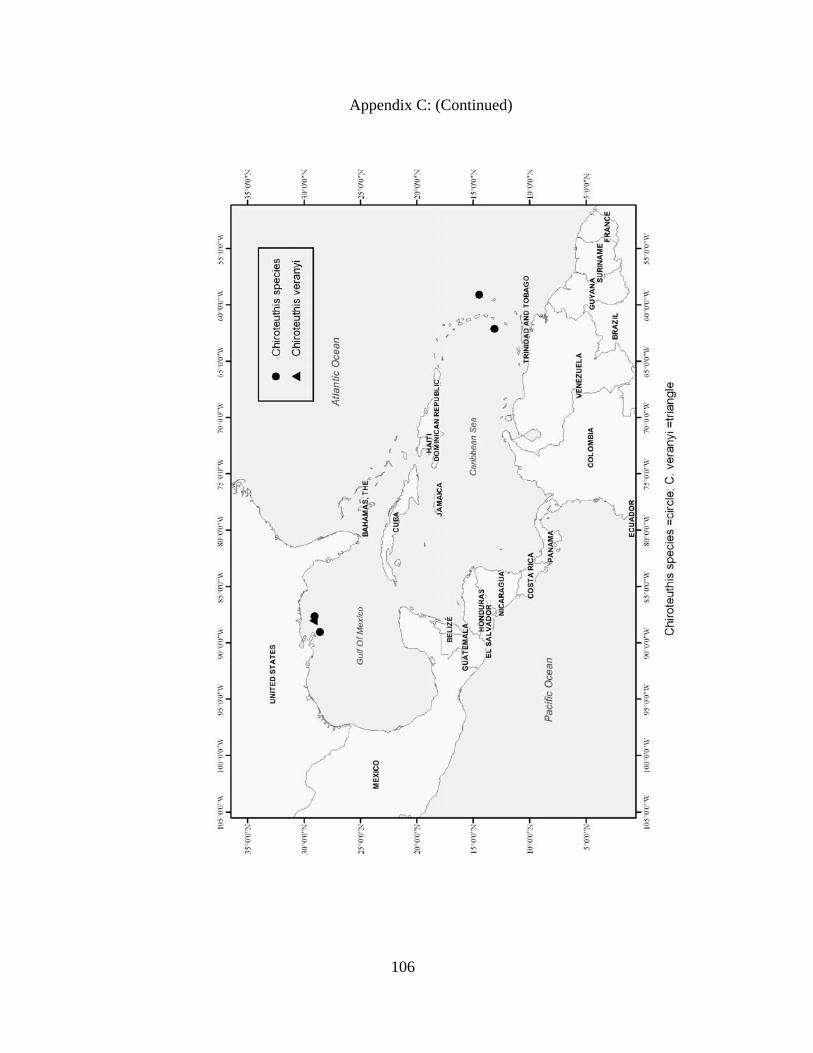
106
Appendix C: (Continued)
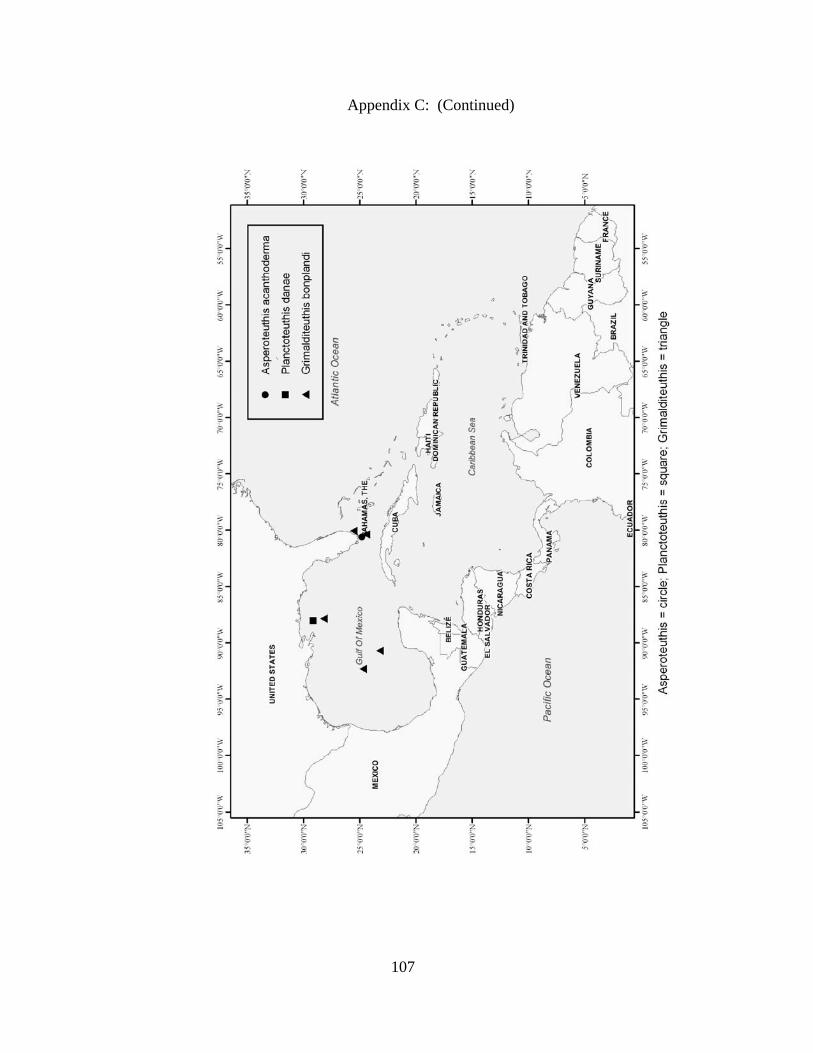
107
Appendix C: (Continued)

108
Appendix C: (Continued)
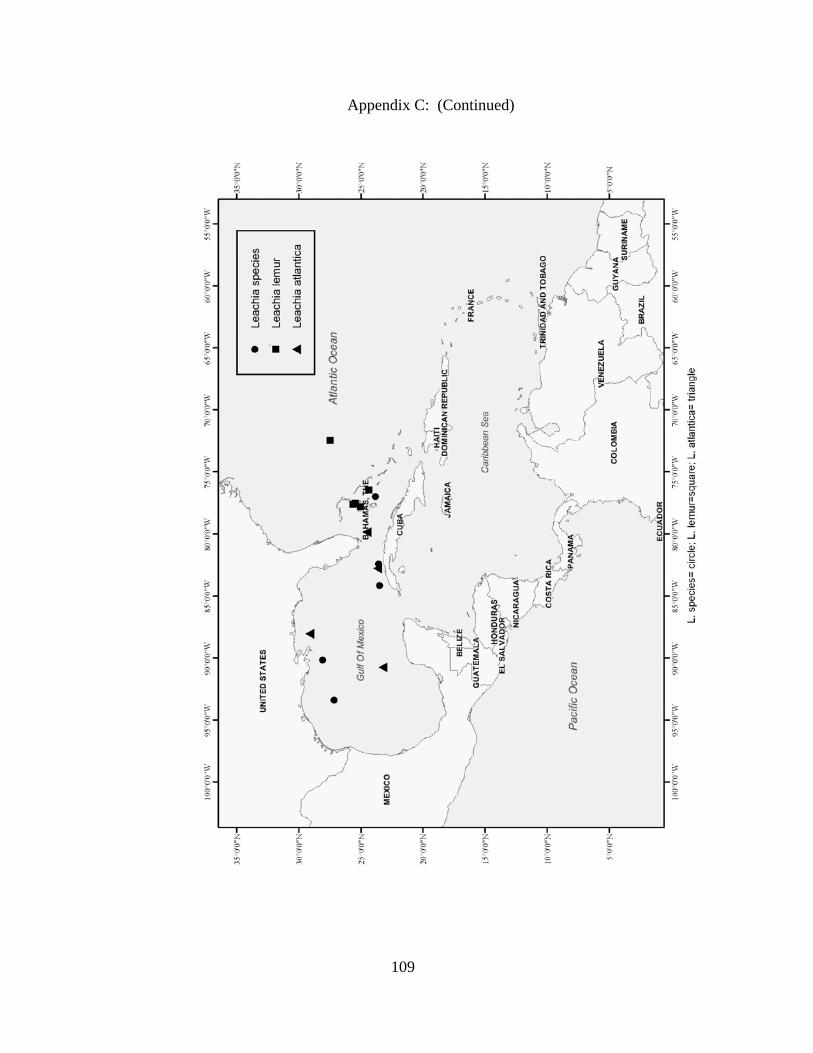
109
Appendix C: (Continued)
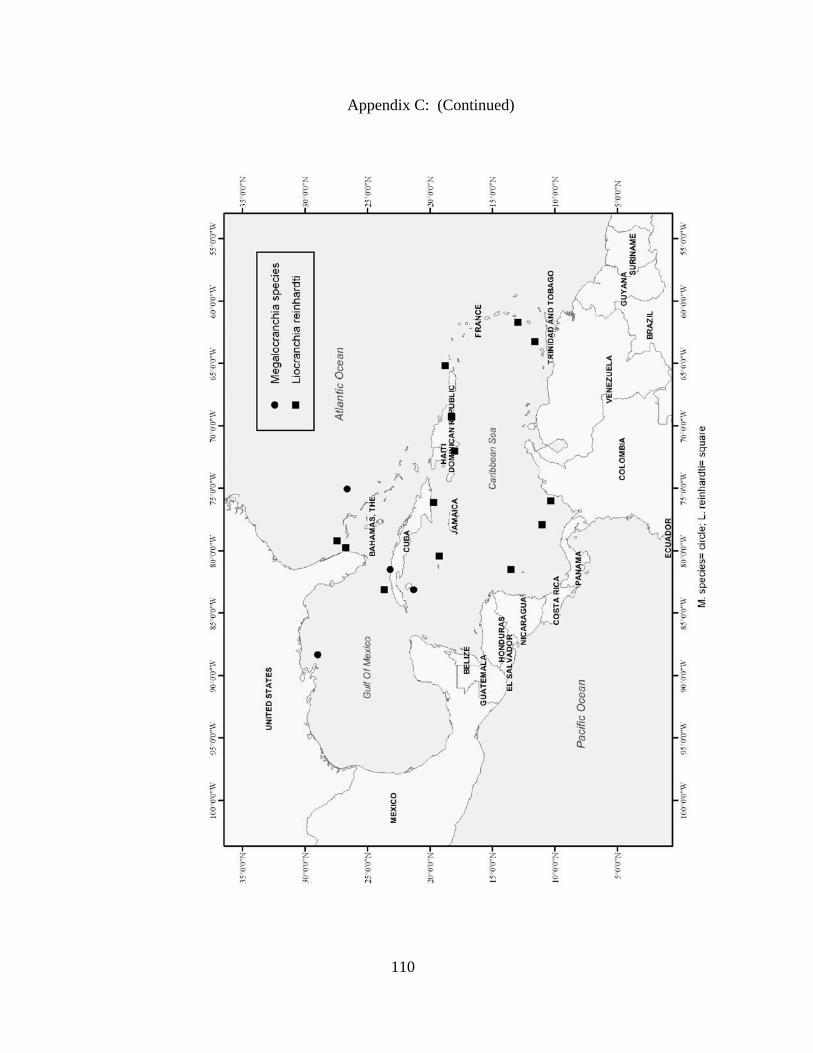
110
Appendix C: (Continued)
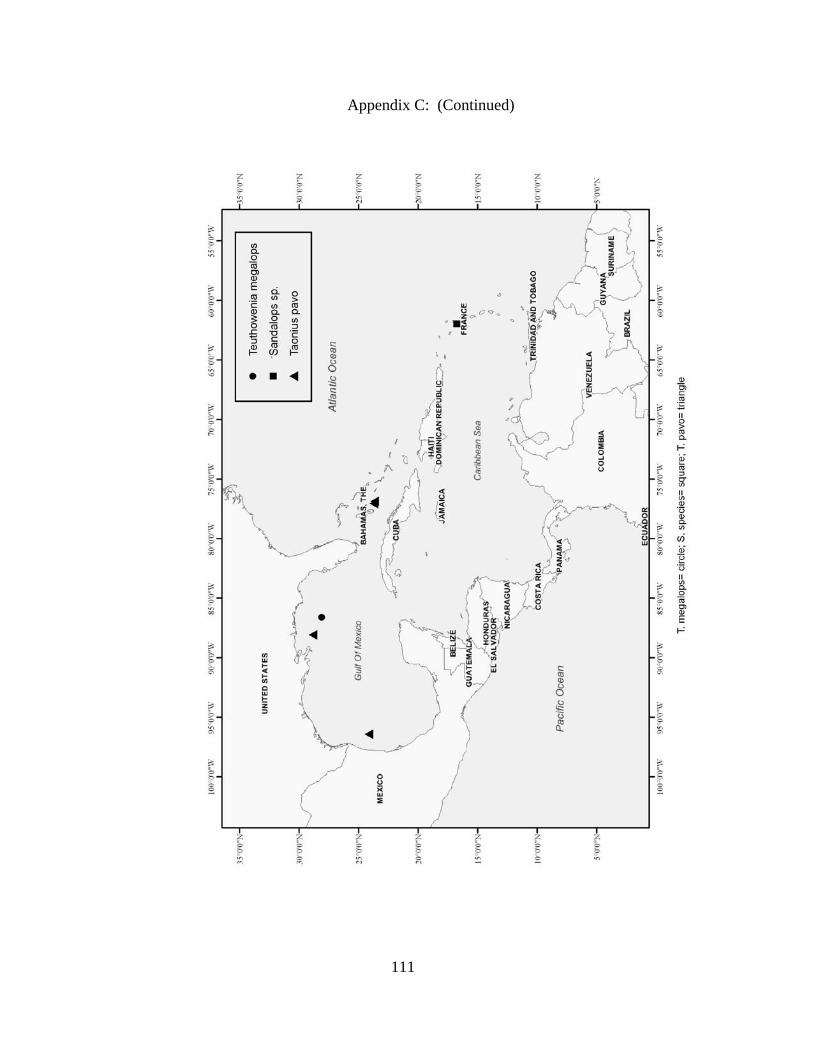
111
Appendix C: (Continued)
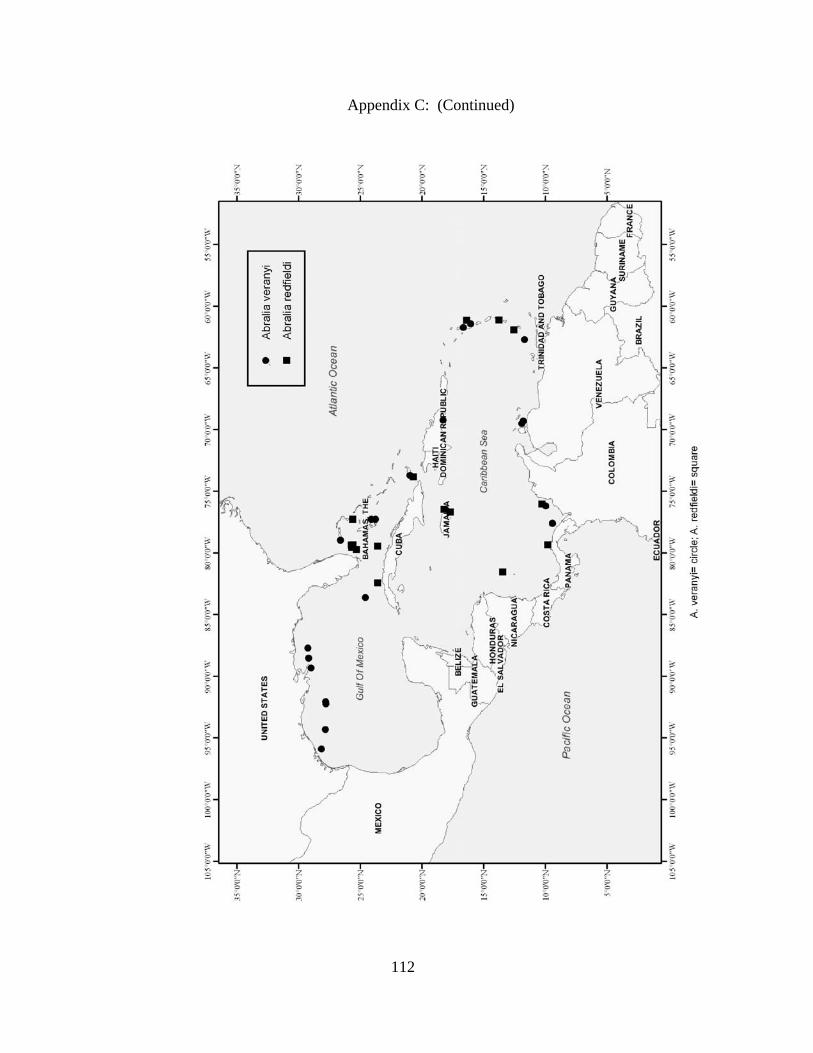
112
Appendix C: (Continued)
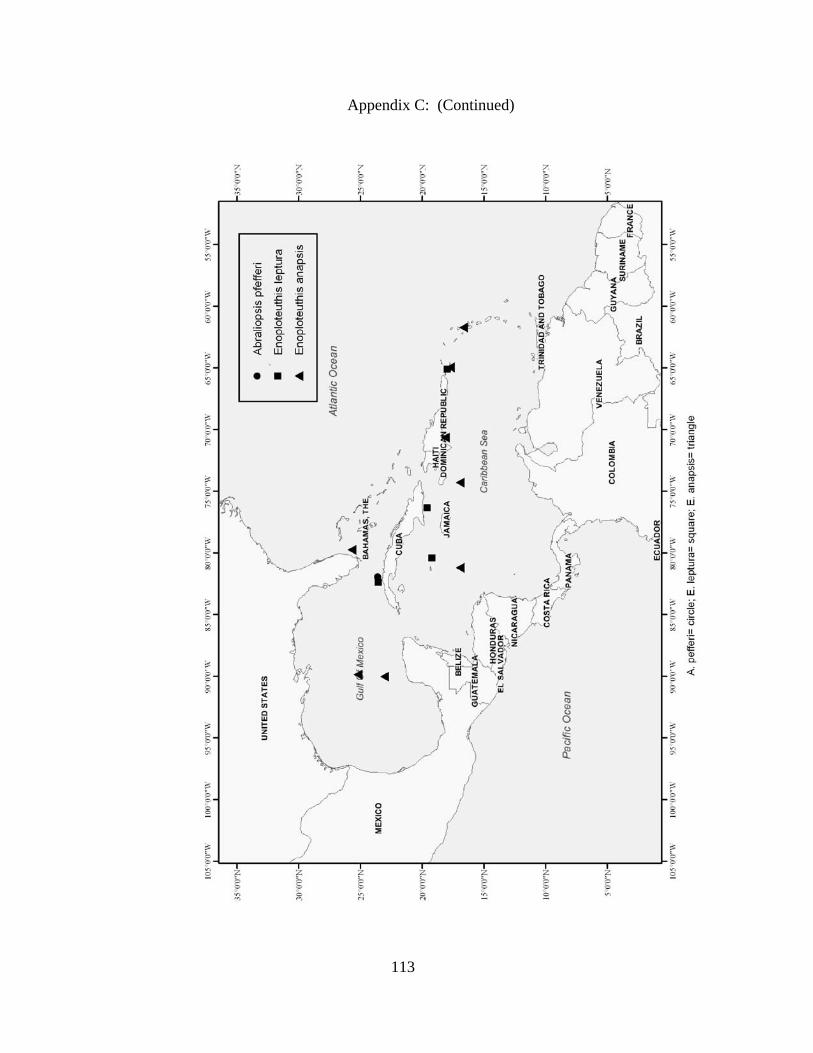
113
Appendix C: (Continued)

114
Appendix C: (Continued)
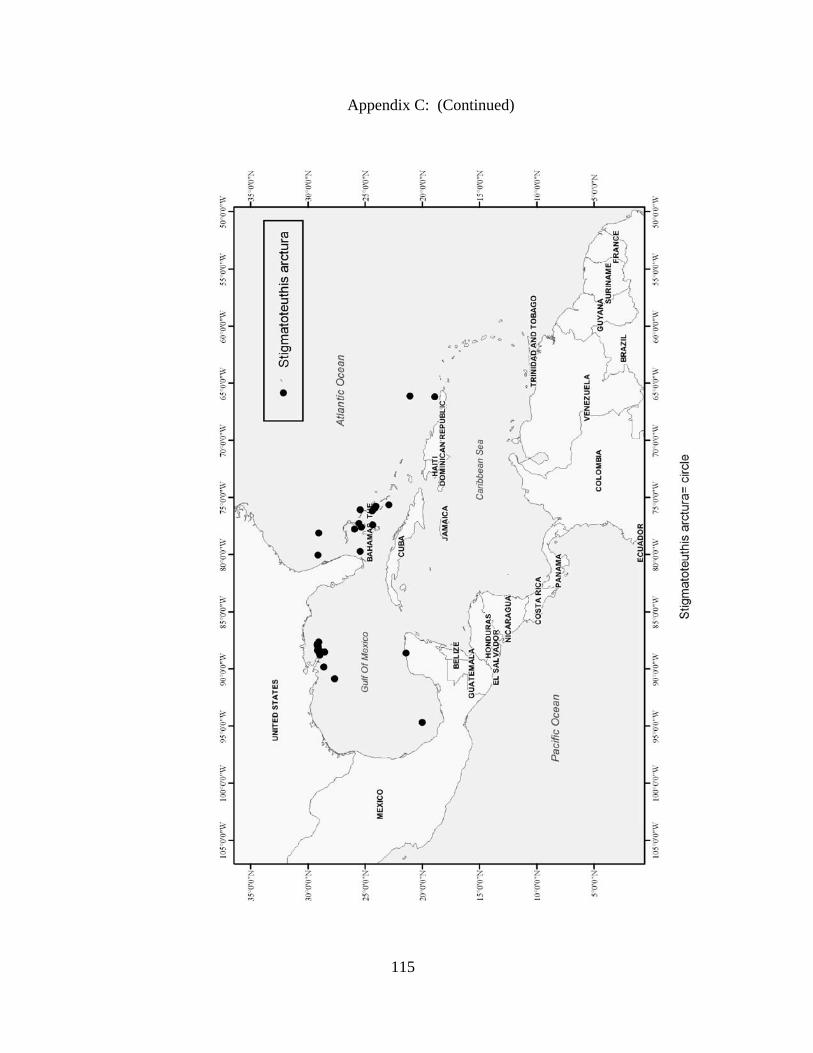
115
Appendix C: (Continued)
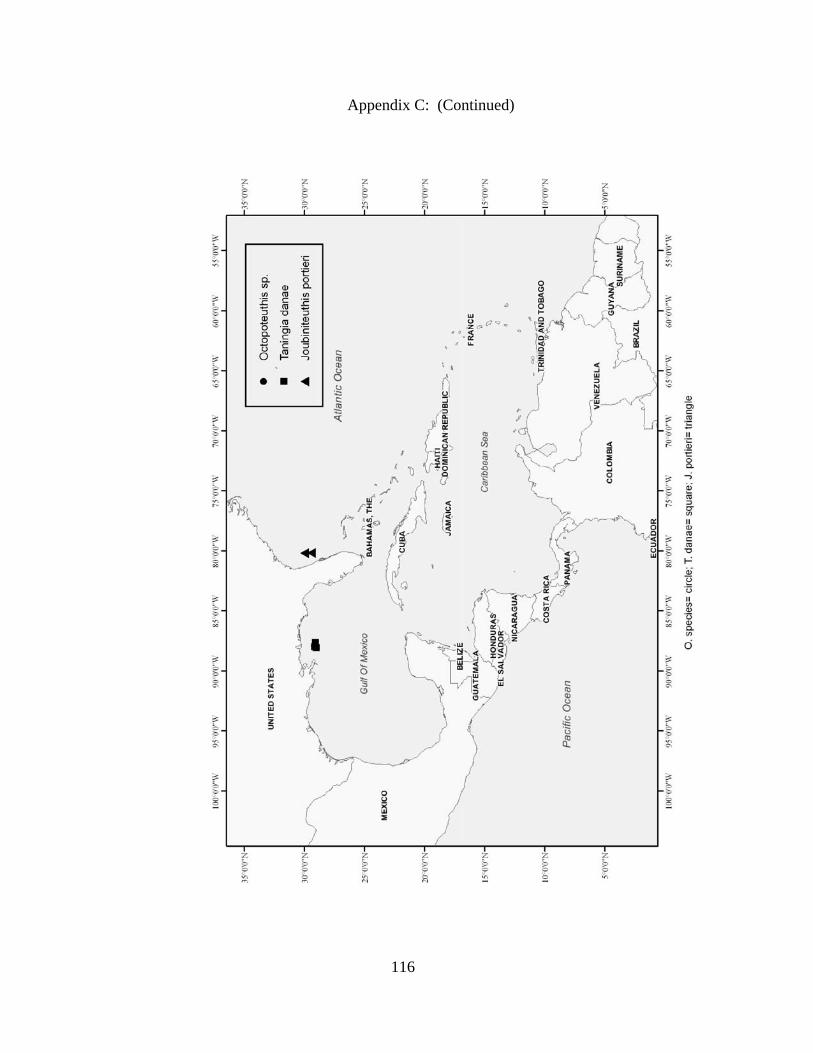
116
Appendix C: (Continued)
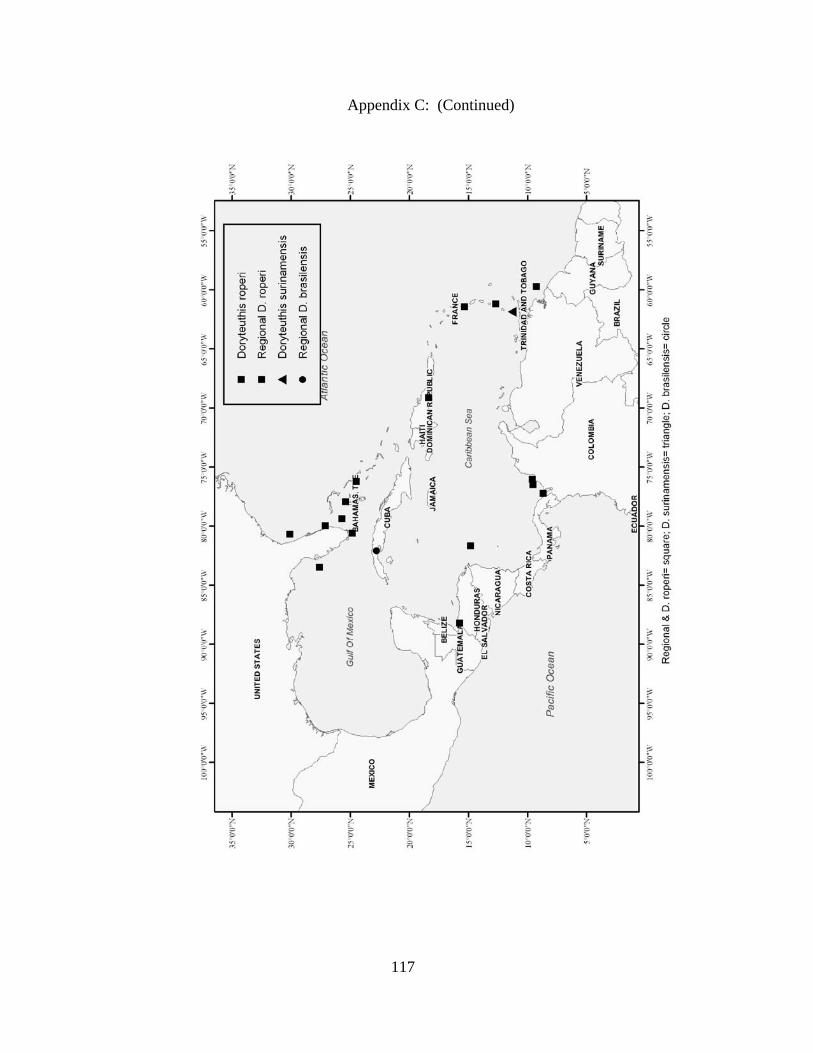
117
Appendix C: (Continued)

118
Appendix C: (Continued)
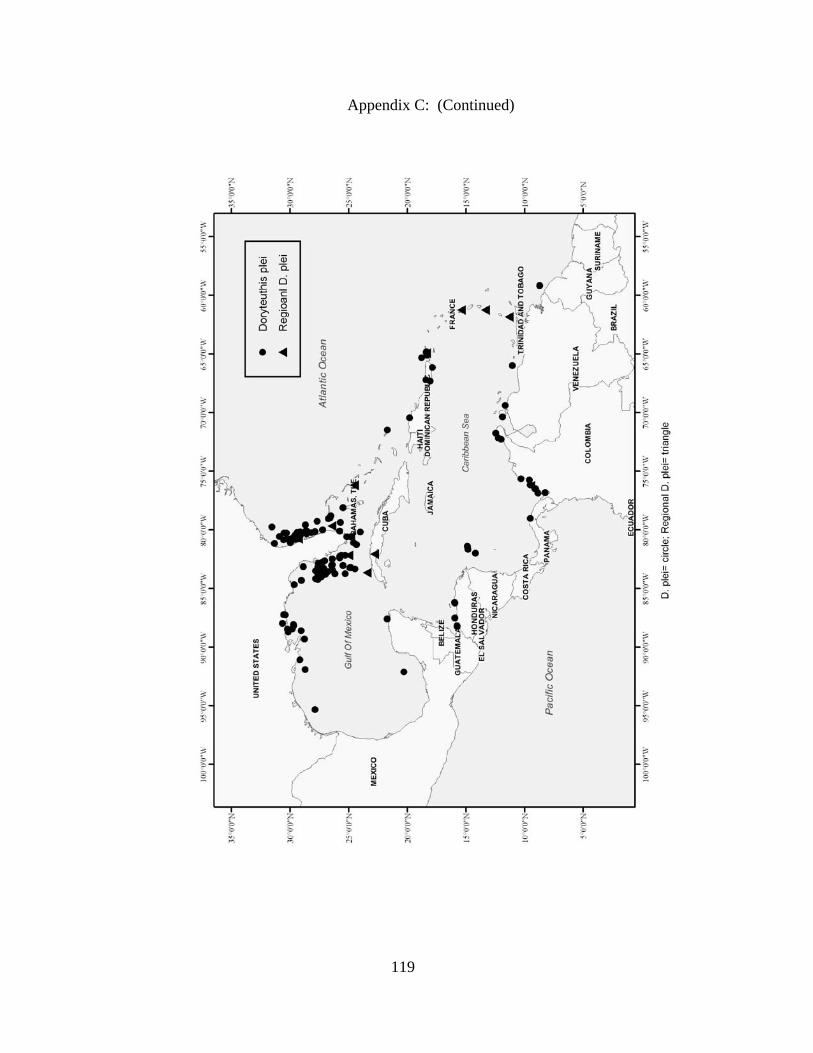
119
Appendix C: (Continued)
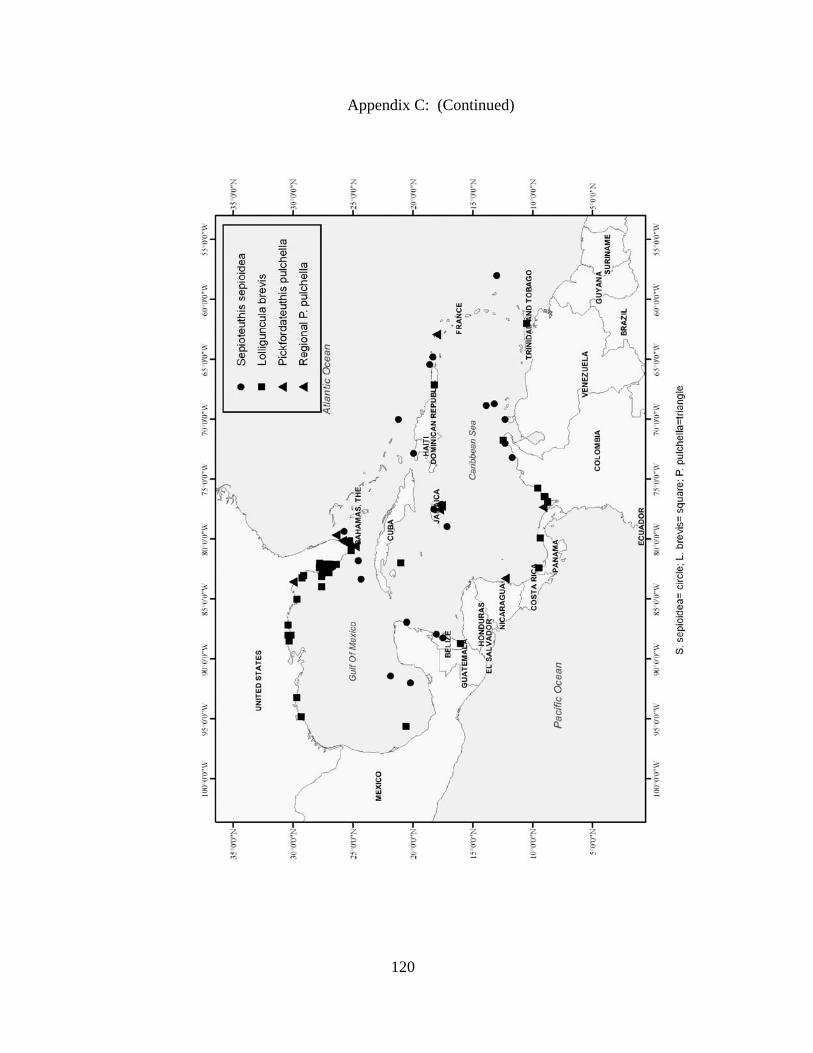
120
Appendix C: (Continued)
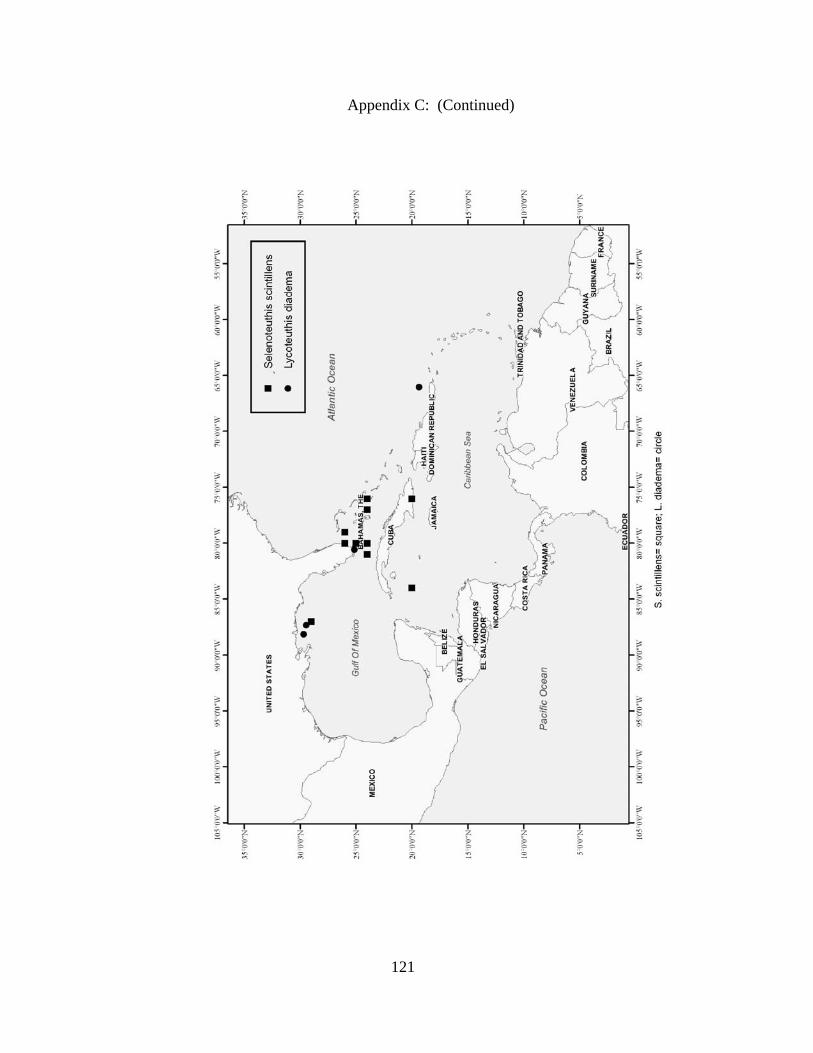
121
Appendix C: (Continued)
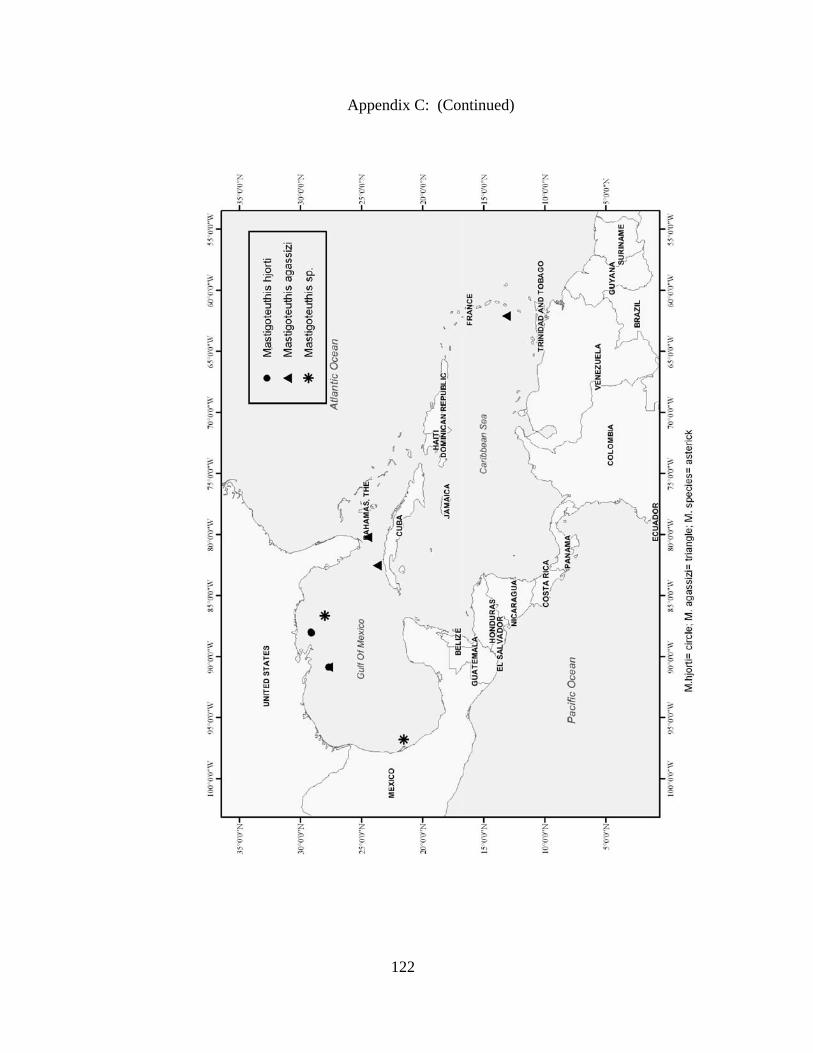
122
Appendix C: (Continued)
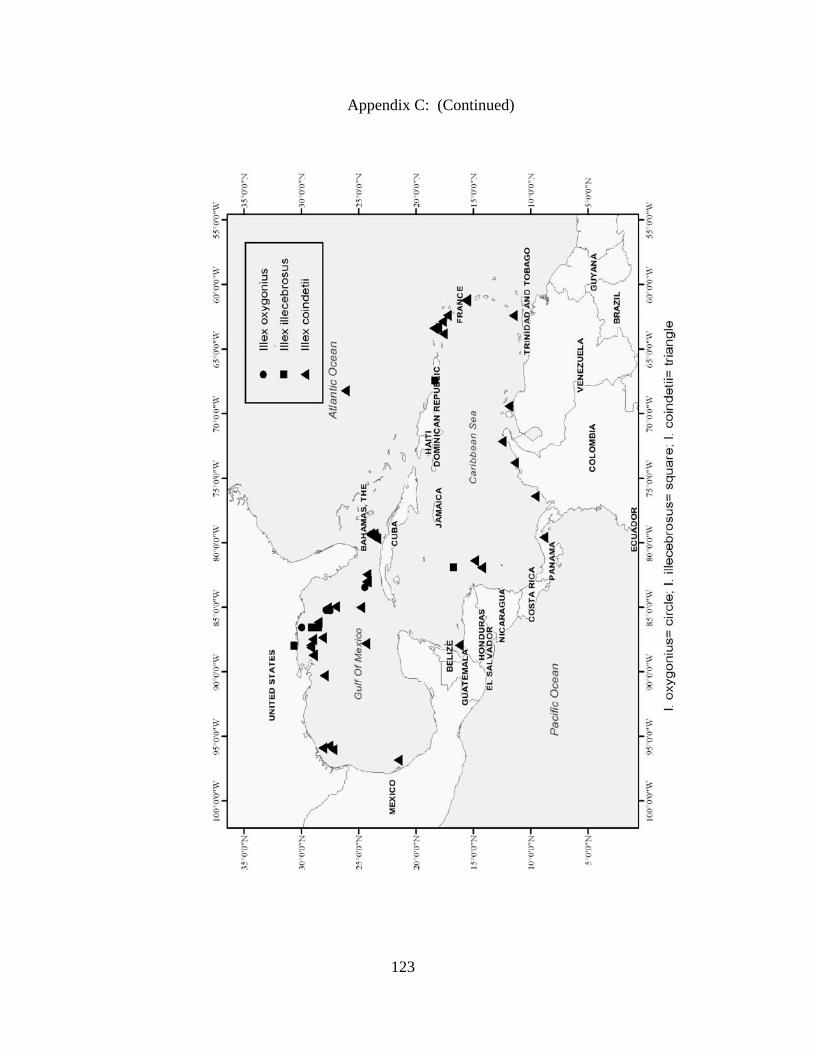
123
Appendix C: (Continued)
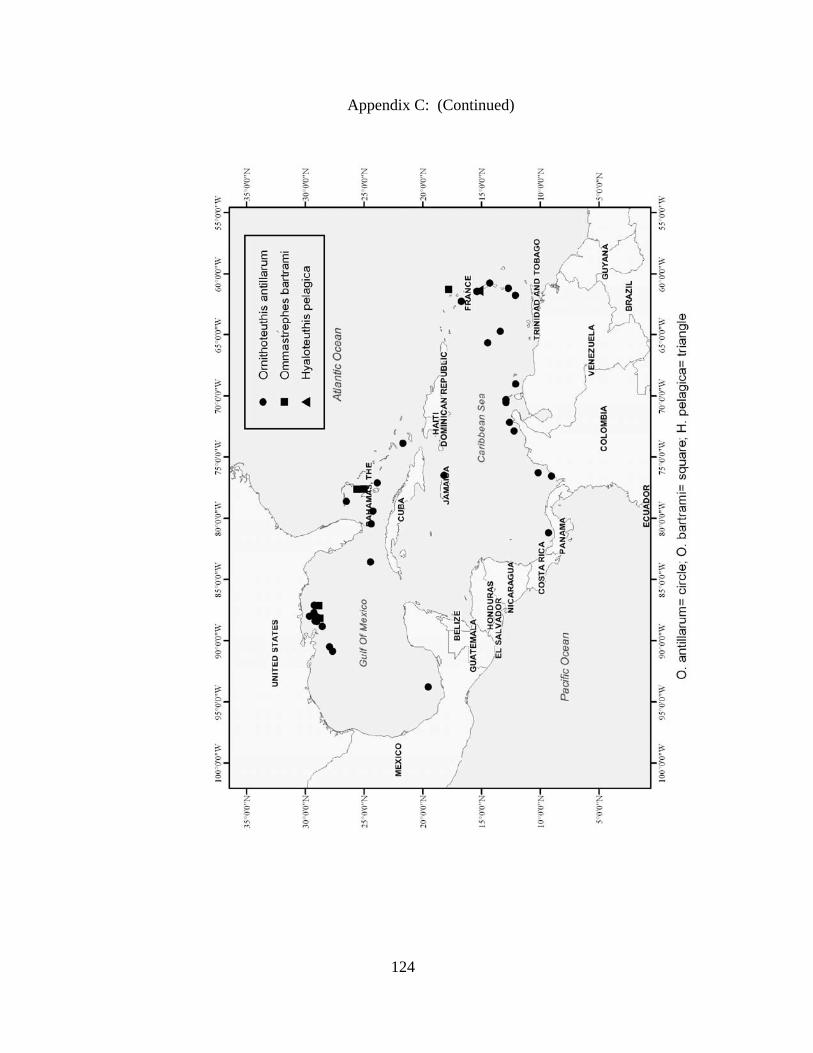
124
Appendix C: (Continued)

125
Appendix C: (Continued)
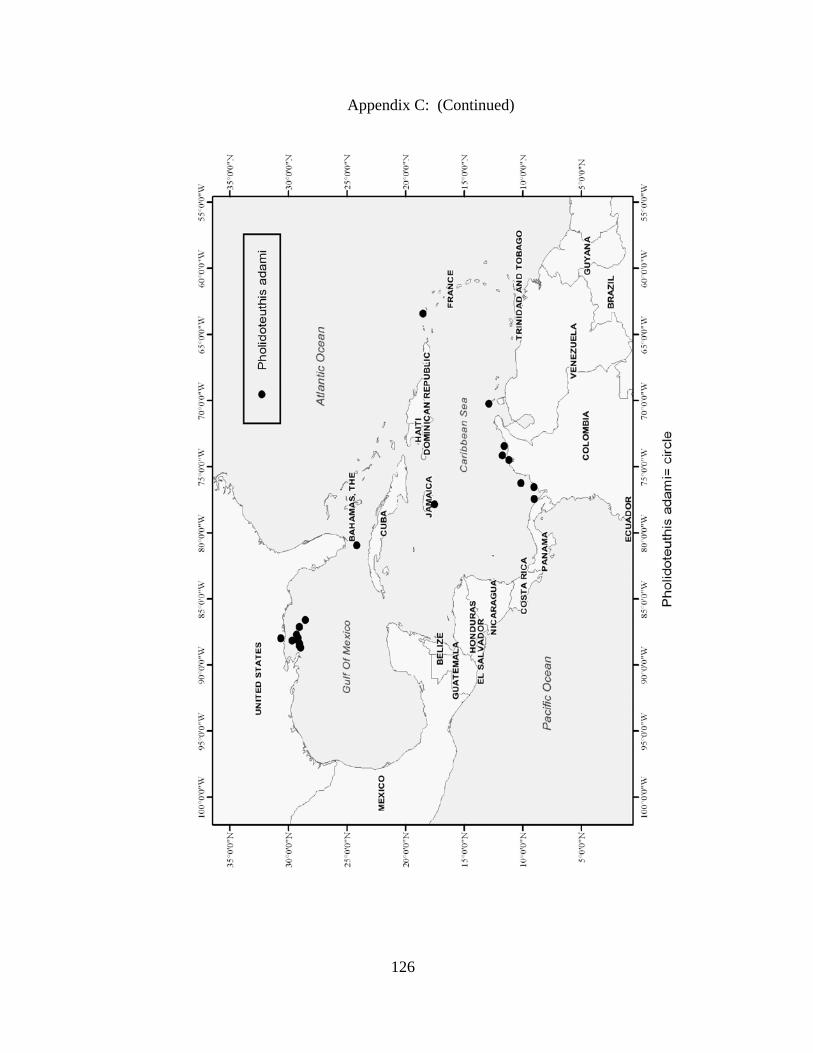
126
Appendix C: (Continued)
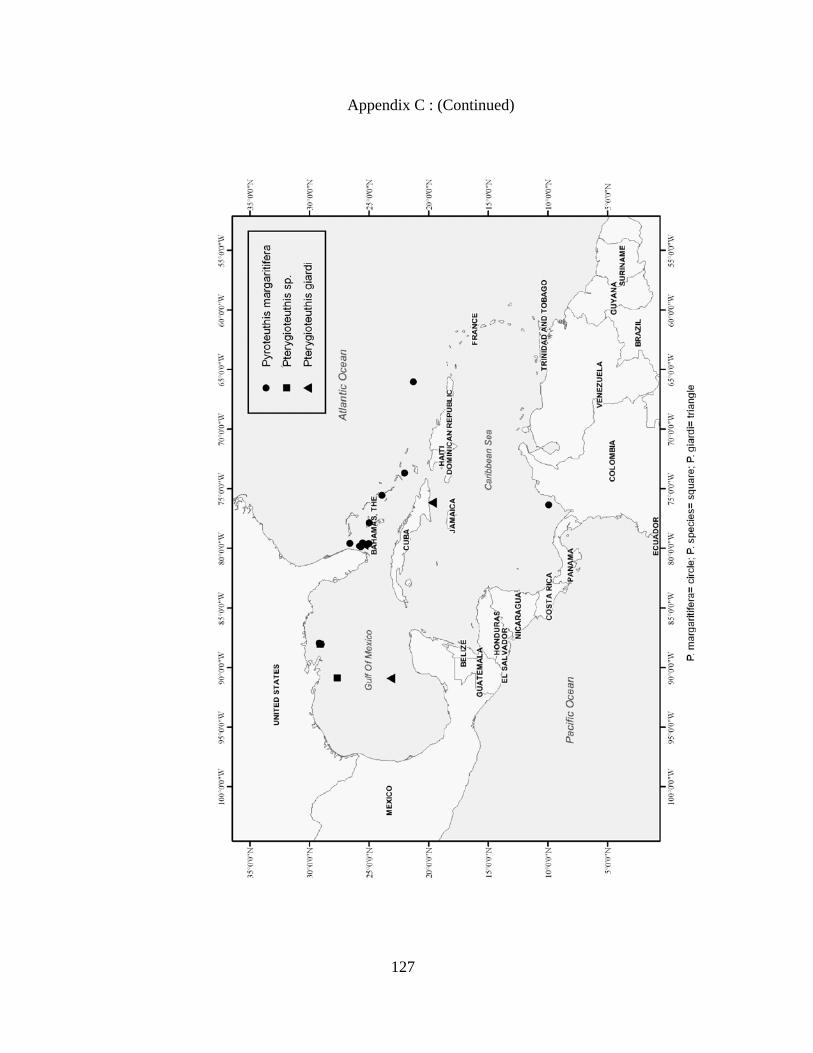
127
Appendix C : (Continued)

128
Appendix C: (Continued)
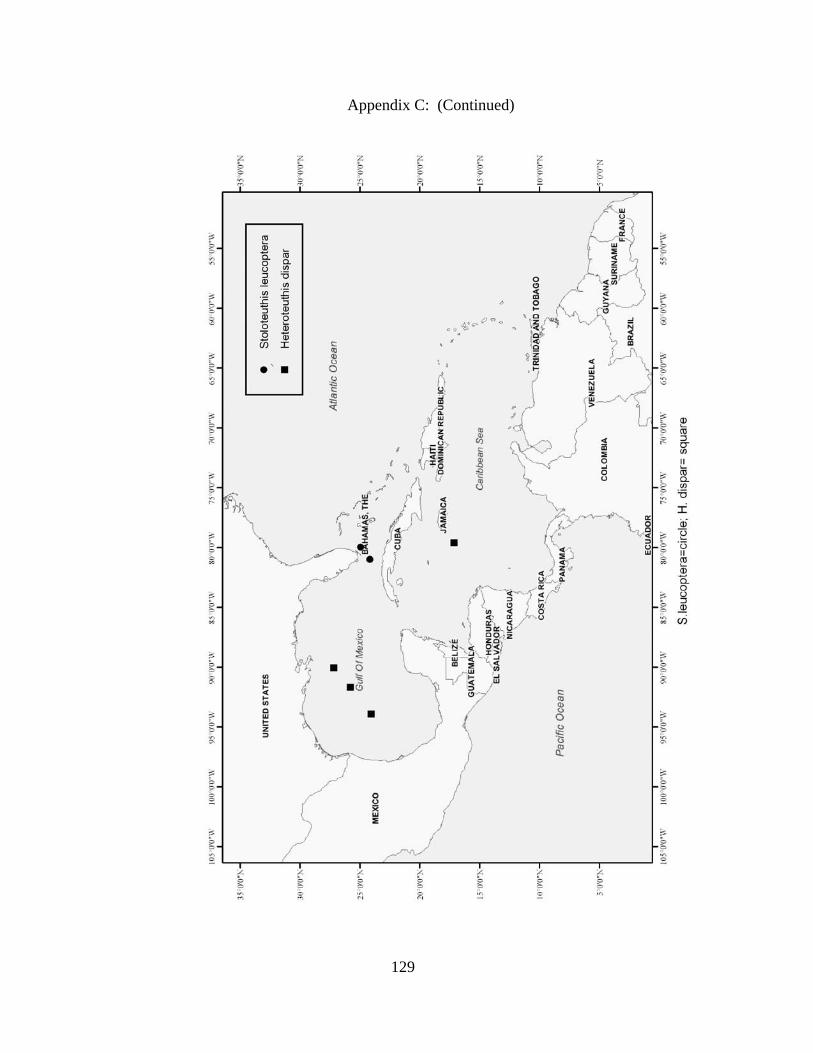
129
Appendix C: (Continued)
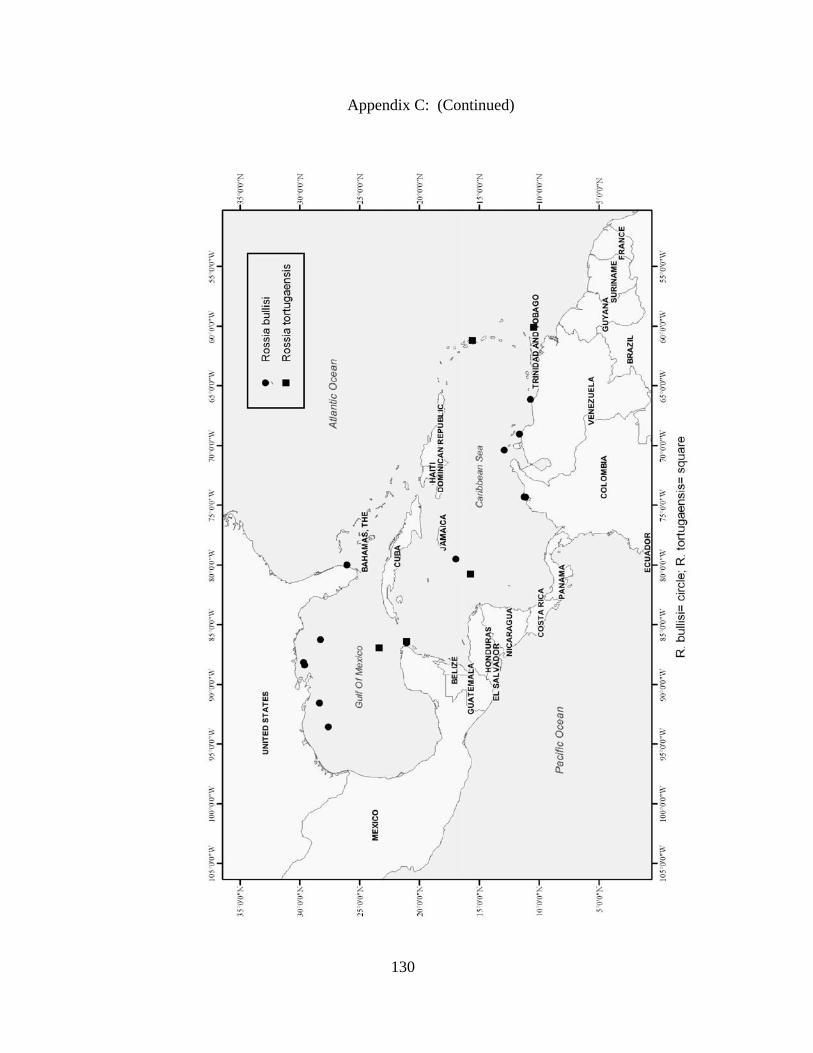
130
Appendix C: (Continued)
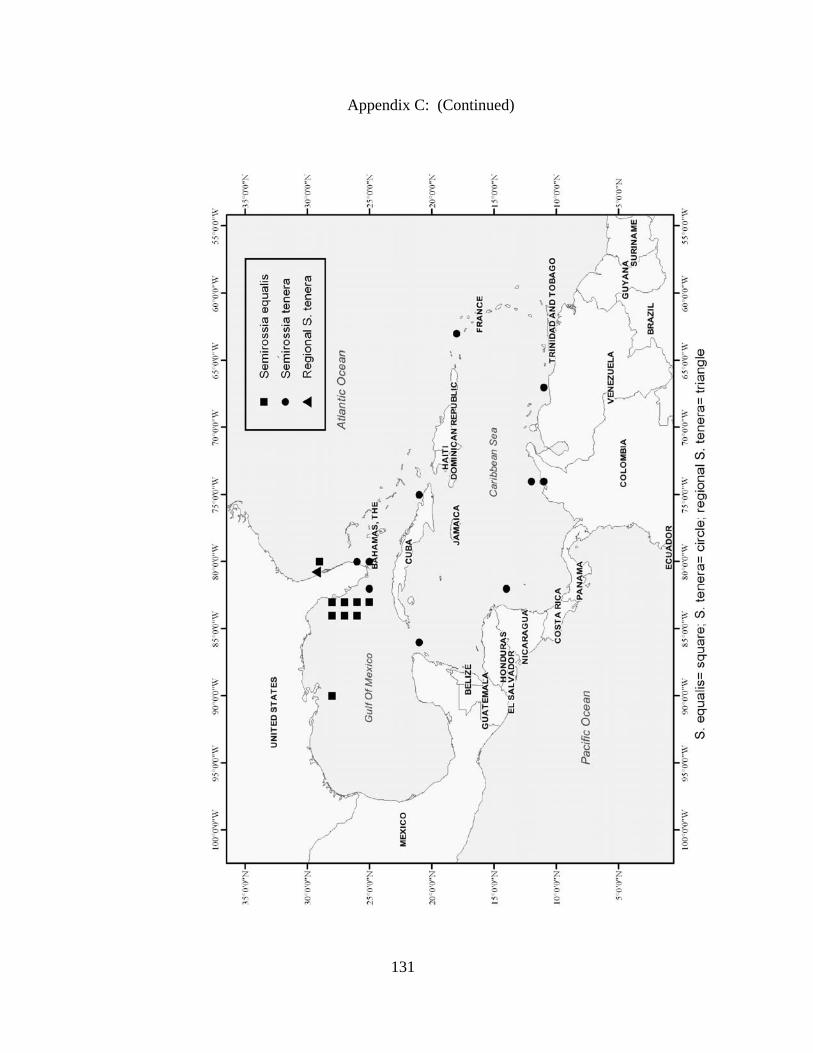
131
Appendix C: (Continued)
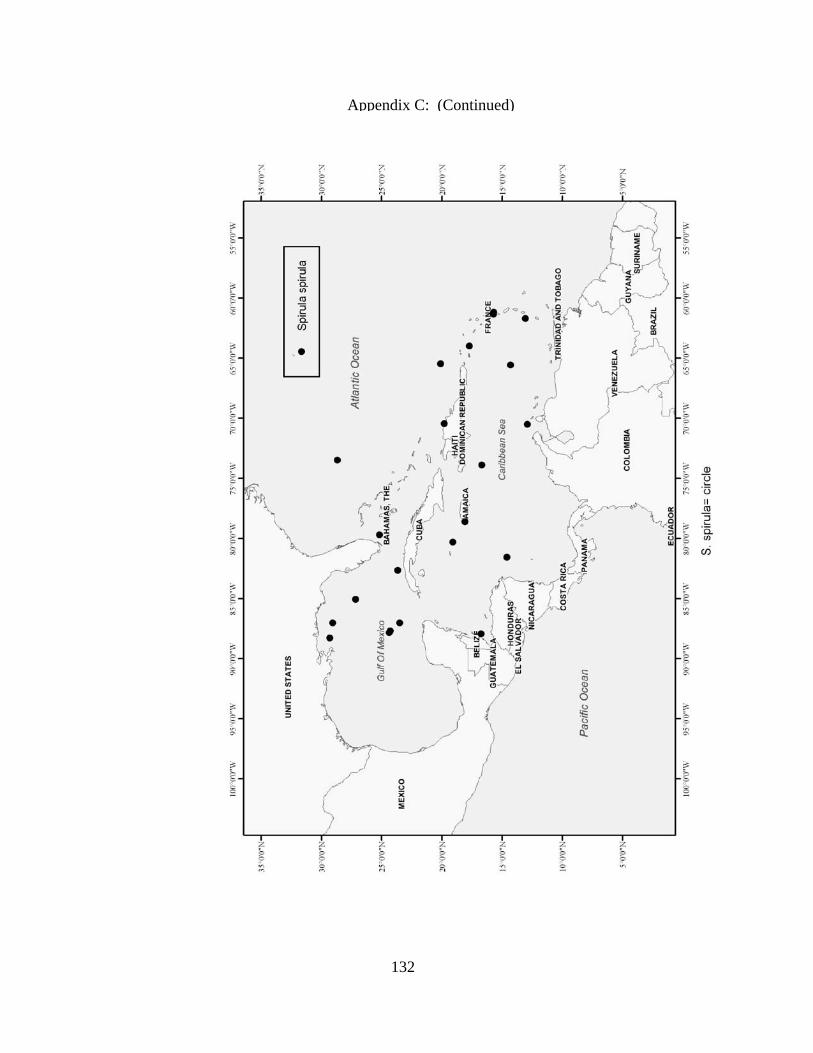
132
Appendix C: (Continued)

133
Appendix C: (Continued)
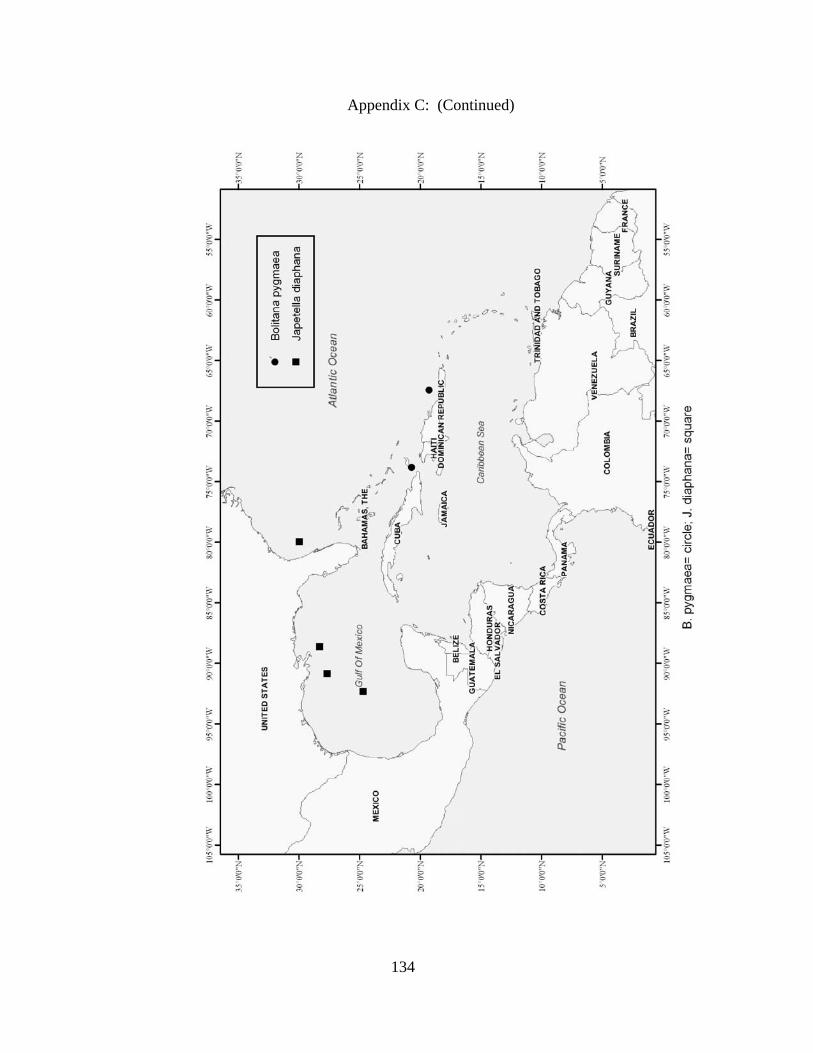
134
Appendix C: (Continued)
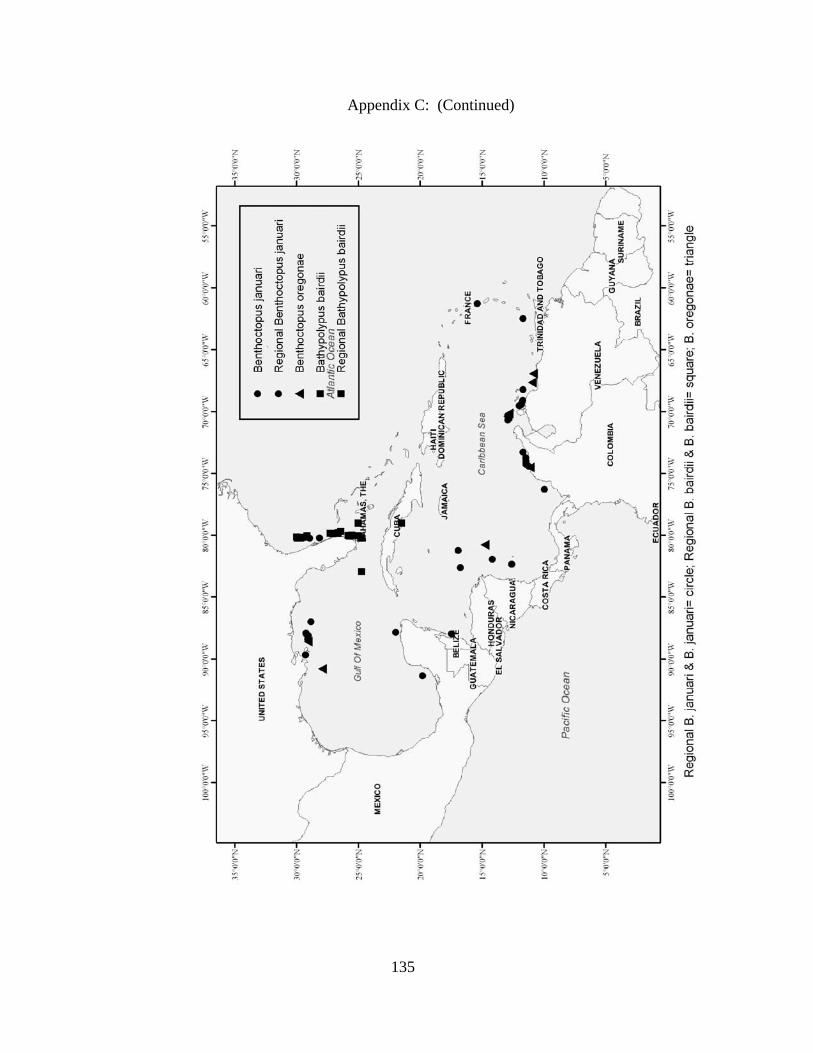
135
Appendix C: (Continued)
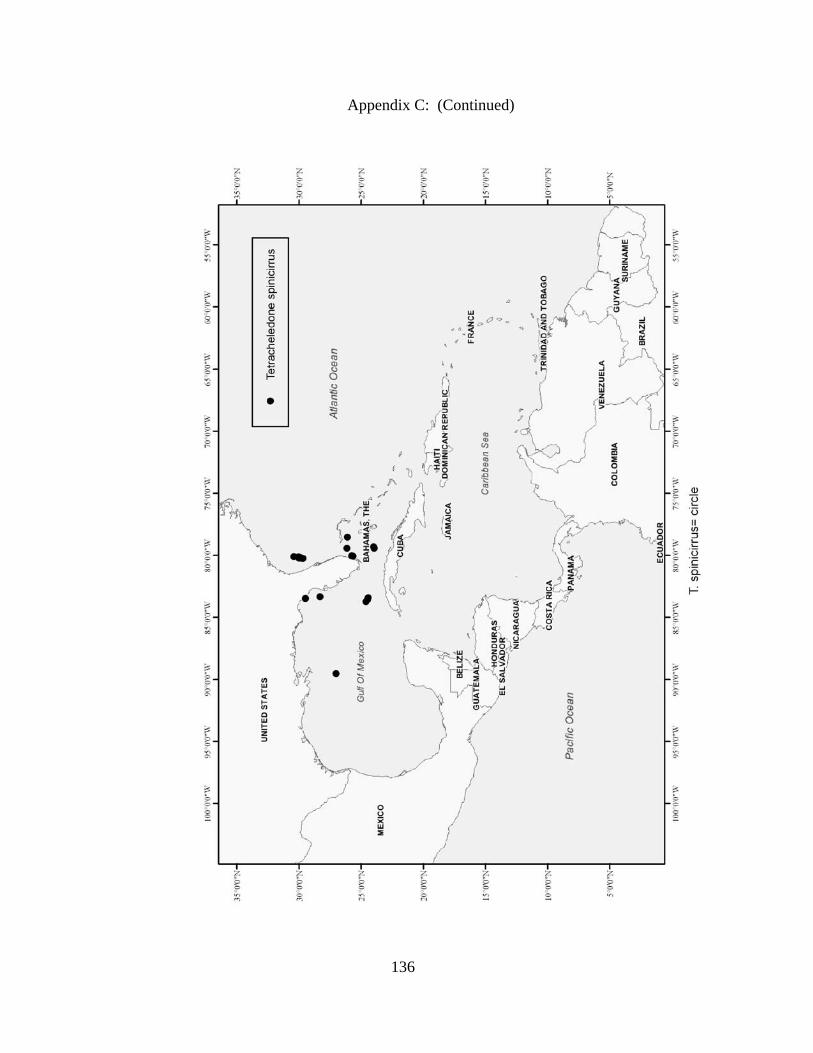
136
Appendix C: (Continued)
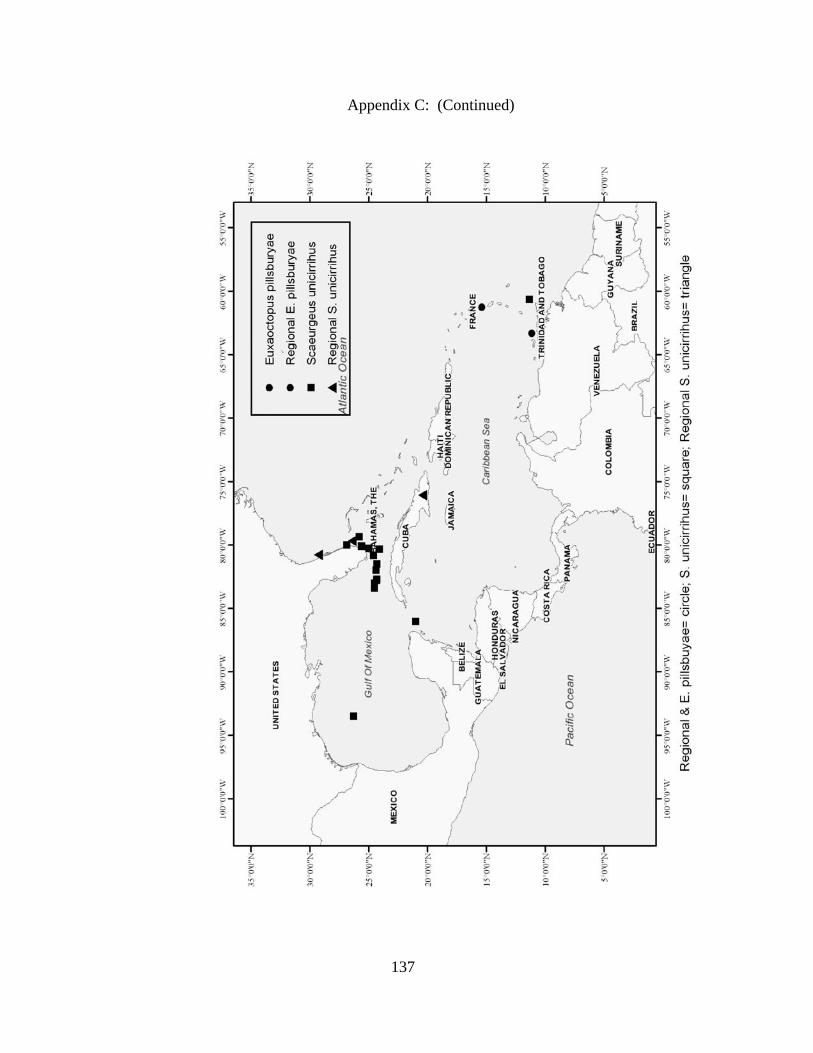
137
Appendix C: (Continued)
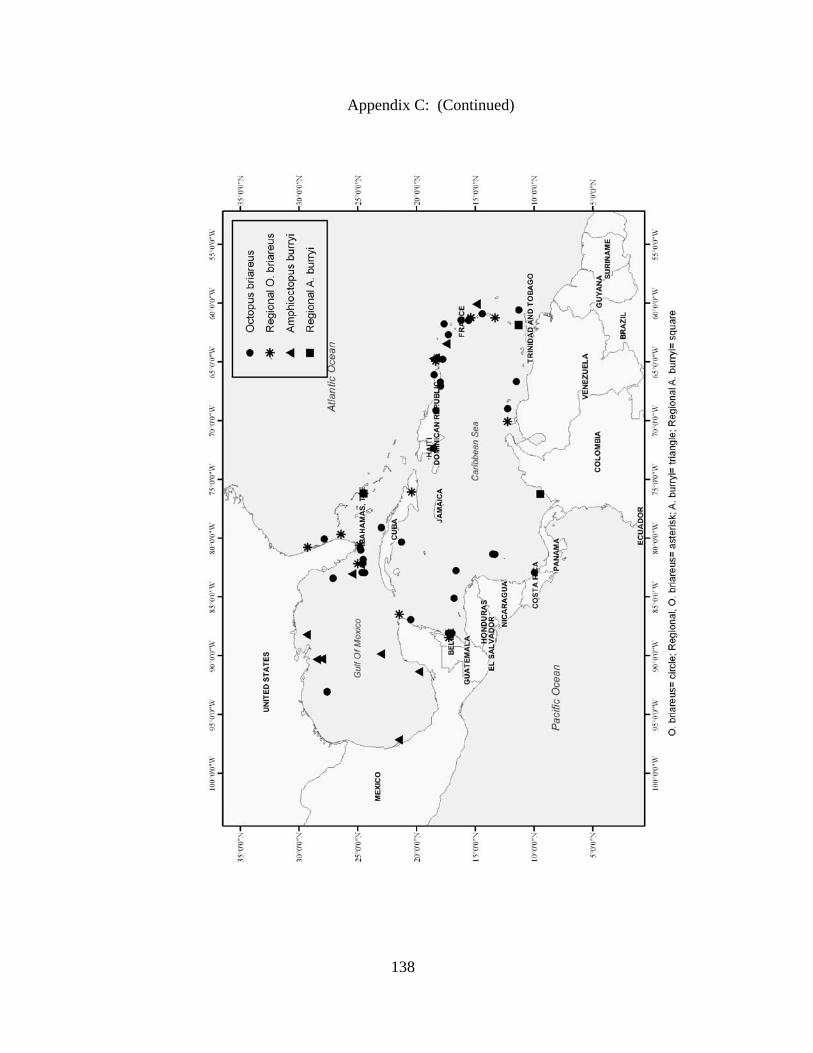
138
Appendix C: (Continued)
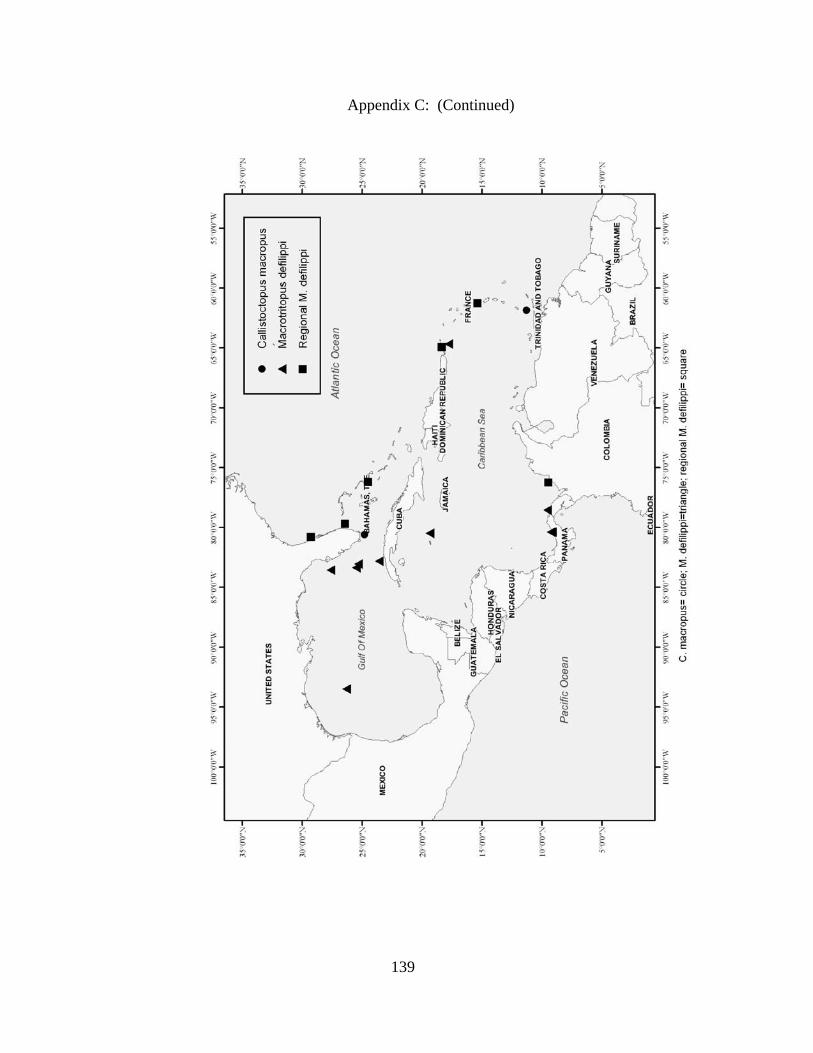
139
Appendix C: (Continued)

140
Appendix C: (Continued)
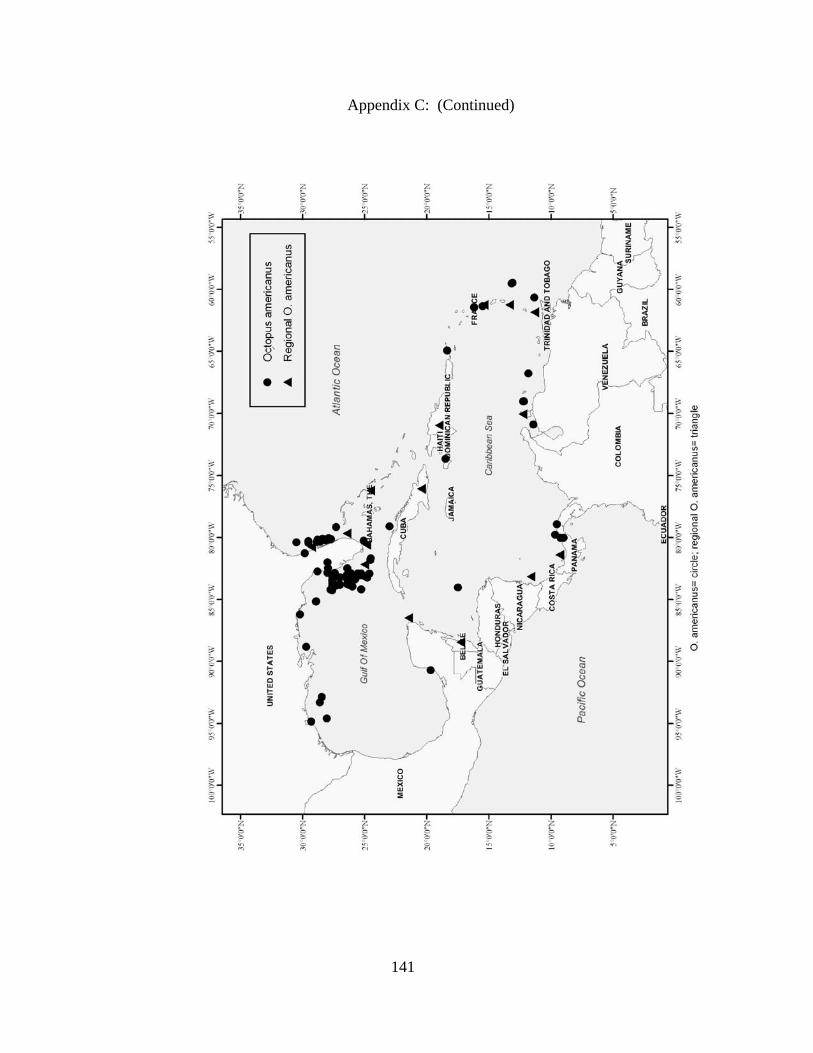
141
Appendix C: (Continued)
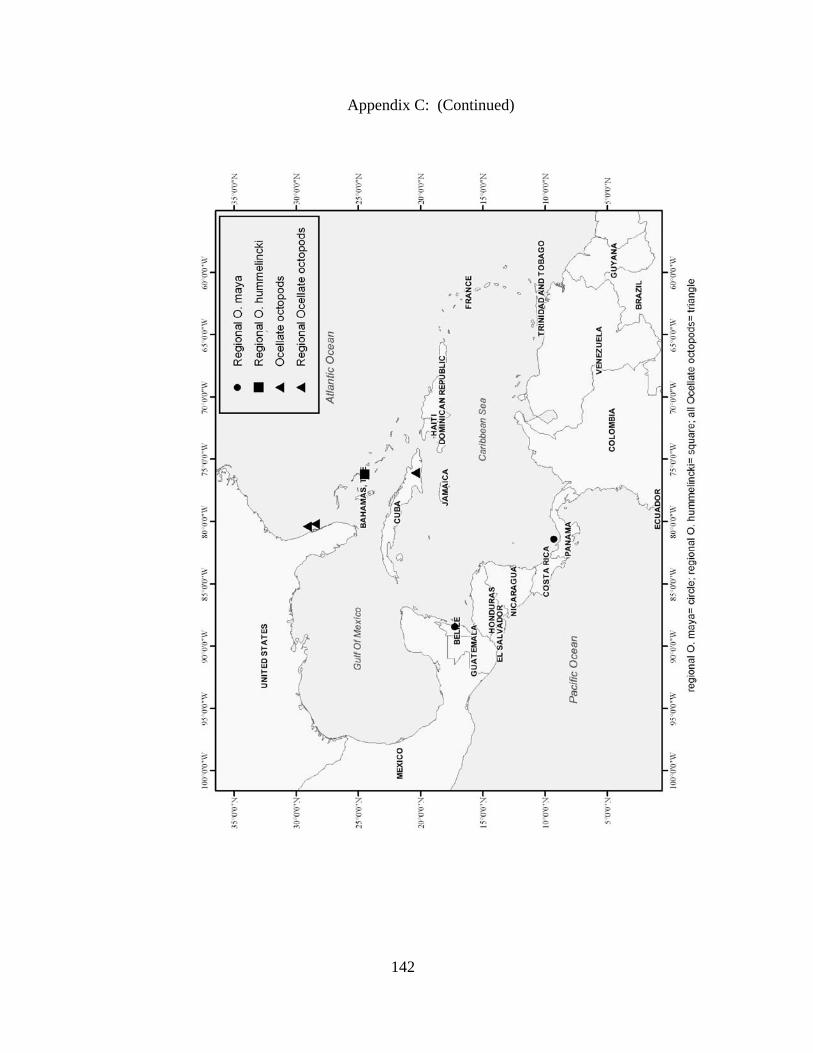
142
Appendix C: (Continued)
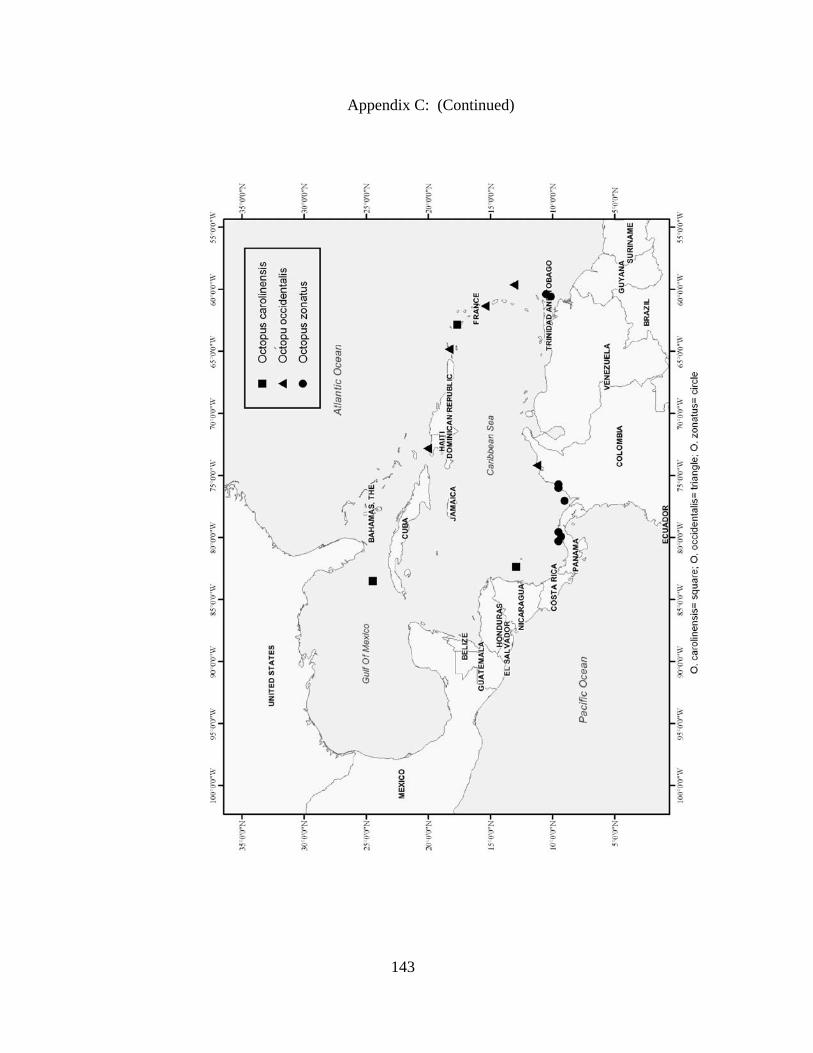
143
Appendix C: (Continued)
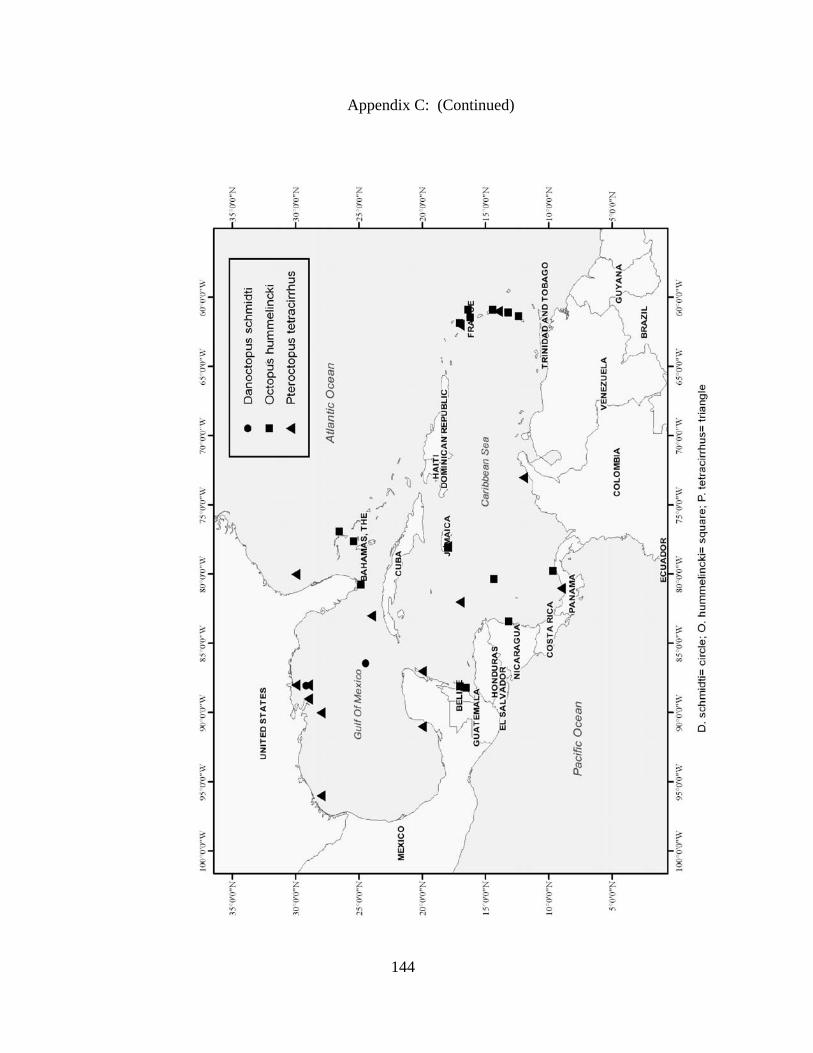
144
Appendix C: (Continued)
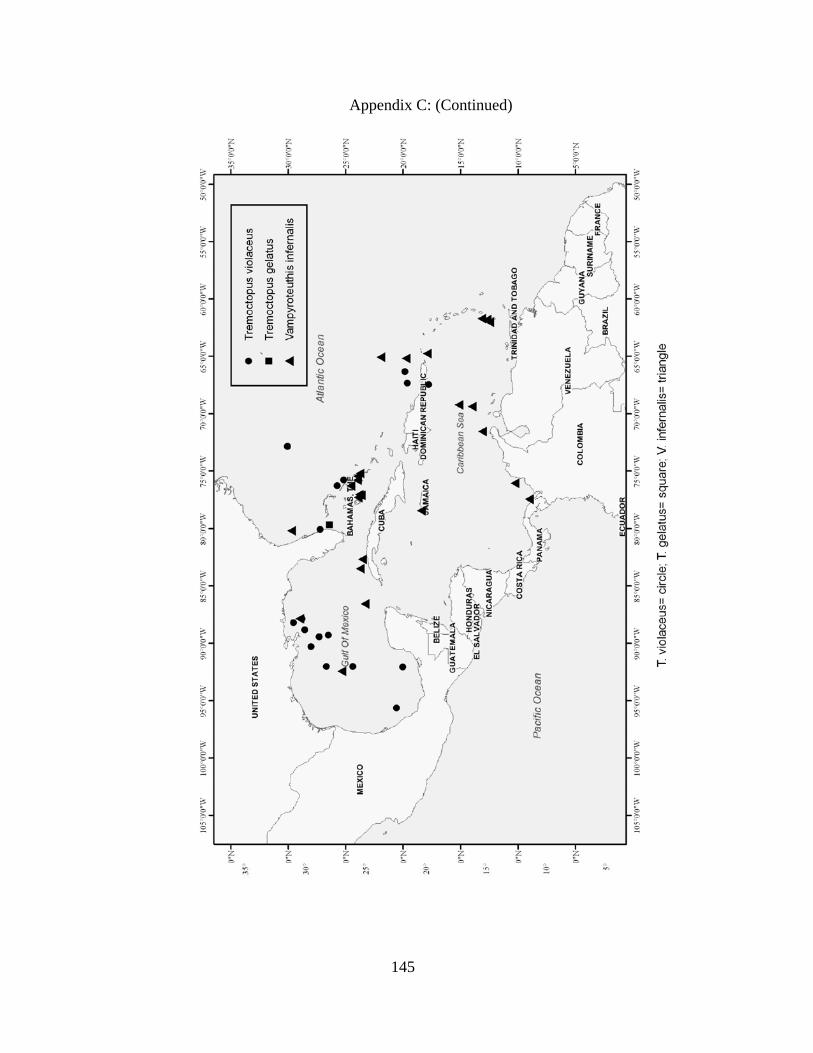
145
Appendix C: (Continued)

146
ABOUT THE AUTHOR
Heather Judkins received her Bachelor’s Degree in Marine Affairs from the University of
Rhode Island in 1993. She moved to Florida and received her Master’s Degree in
Secondary Science Education while teaching high school marine science classes. In
2003, she entered the Marine Science graduate program through the University of South
Florida pursuing a biological track focusing on cephalopods. While enrolled, she
received two fellowships through the GK12 Oceans Program which was sponsored by the
National Science Foundation. She also coordinated the Regional National Ocean
Sciences Bowl, the Spoonbill Bowl, as a result of her fellowship project from 2004-
present. She has continued teaching full time while tackling her degree. She has
coauthored two publications regarding cephalopods of the Broad Caribbean to date.

147
Recommended

















