REVIEW
Biotechnological advances in guava (Psidium guajava L.): recentdevelopments and prospects for further research
Manoj K. Rai • Pooja Asthana • V. S. Jaiswal •
U. Jaiswal
Received: 3 January 2009 / Revised: 22 September 2009 / Accepted: 24 September 2009 / Published online: 6 October 2009
� Springer-Verlag 2009
Abstracts Guava (Psidium guajava L.), an important fruit
crop of several tropical and sub-tropical countries, is facing
several agronomic and horticultural problems such as sus-
ceptibility to many pathogens, particularly guava wilting
caused by Fusarium oxysporium psidii, low fruit growth,
short shelf life of fruits, high seed content, and stress sensi-
tivity. Conventional breeding techniques have limited scope
in improvement of guava owing to long juvenile period, self
incompatibility, and heterozygous nature. Conventional
propagation methods, i.e., cutting, grafting or stool layering,
for improvement of guava already exist, but the long juvenile
period has made them time consuming and cumbersome.
Several biotechnological approaches such as genetic trans-
formation may be effective practical solutions for such
problems and improvement of guava. The improvement of
fruit trees through genetic transformation requires an effi-
cient regeneration system. During the past 2–3 decades,
different approaches have been made for in vitro propagation
of guava. An overview on the in vitro regeneration of guava
via organogenesis, somatic embryogenesis, and synthetic
seeds is presented. Organogenesis in several different
genotypes through various explant selection from mature
tree and seedling plants has been achieved. Factors affecting
somatic embryogenesis in guava have been reviewed.
Production of synthetic seeds using embryogenic propa-
gules, i.e., somatic embryos and non-embryogenic vegeta-
tive propagules, i.e., shoot tips and nodal segments have also
been achieved. Development of synthetic seed in guava may
be applicable for propagation, short-term storage, and
germplasm exchange, and distribution. An initial attempt for
genetic transformation has also been reported. The purpose
of this review is to focus upon the current information on in
vitro propagation and biotechnological advances made in
guava.
Keywords Guava � Genetic transformation �In vitro propagation � Organogenesis �Somatic embryogenesis � Synthetic seeds
Introduction
Guava (Psidium guajava L., family Myrtaceae), ‘‘poor
man’s fruit’’ or ‘‘apple of tropics’’, is an important fruit
crop of tropical and sub-tropical regions of the world.
Guava fruit contains 2–5 times more vitamin C than orange
and is also good source of calcium, phosphorus, and iron
(Singh 2005). Traditionally, different parts of plants, i.e.,
fruits, leaves, roots, and bark are used in the treatment of
gastroenteritis, diarrheoa, and dysentery (Jaiswal and Amin
1992). High concentration of pectin in guava fruit may play
a significant role in the reduction of cholesterol and thereby
decrease the risk of cardiovascular disease (Singh 2005).
Despite these advantages, there are a number of problems
that affect guava production. Being a cross-pollinated
species, substantial variability is available in seedling
populations in different guava growing regions (Srivastava
2005). Guava wilting disease caused by Fusarium oxys-
porium psidii is a serious problem faced by guava growers,
Communicated by J. Carlson.
M. K. Rai � P. Asthana � V. S. Jaiswal � U. Jaiswal
Laboratory of Morphogenesis, Department of Botany,
Banaras Hindu University, Varanasi 221005, UP, India
Present Address:M. K. Rai (&)
Centre for Plant Biotechnology, CCSHAU Campus,
Hisar 125004, Haryana, India
e-mail: [email protected]
123
Trees (2010) 24:1–12
DOI 10.1007/s00468-009-0384-2

and loss due to this disease is substantial (Mishra 2005).
Other important field diseases of guava are anthracnose
(Gloeosporium psidii), and canker (Pestalotia psidii).
Guava suffers from different rot pathogens which cause
maximum loss (Mishra 2005).
Before initiation of any crop improvement program in
guava, priority needs to be given to the following: good fruit
quality, increasing yields, disease resistance, longer shelf life
of fruits, high vitamin C and pectin content, good aroma,
attractive skin, flesh color, and soft seeds (Dinesh and Iyer
2005). Such an ideal phenotype cannot be met by conven-
tional breeding. Floral structure (epigynous flower, with
abundant incurred stamens of various sizes), long juvenile
period, self incompatibility, and heterozygous nature limit
the scope of breeding programs for improvement of guava
(Jaiswal and Amin 1992). Despite these problems, a few
successful reports on guava breeding have also appeared
(Ribeiro and Pommer 2004; Pommer and Murakami 2008).
Guava is conventionally propagated through cutting, graft-
ing, stooling, or air layering, but these methods are time
consuming (Chandra et al. 2004). The planting of extensive
new orchards of vegetatively propagated clones of some
tropical fruits has some times been limited by pathogens
(Litz and Jaiswal 1991). In majority of trees, propagation by
root cutting is often characterized by a rapid loss of rooting
capacity of the cutting with increasing age of parent plant
(Thorpe et al. 1991). Clonal propagation using cell, tissue,
and organ culture techniques have considerable potential for
the improvement of economically important trees within a
limited time frame (Giri et al. 2004; Singh et al. 2004).
Generation of new and promising variability through
somaclonal variant selection, production of androgenic, and
gynogenic haploids to achieve homozygosity, freeing plants
from disease-causing organisms by shoot tip culture, pro-
duction of industrial compounds by cell culture, and deve-
lopment of stress-tolerant plants are some well-known
applications of plant tissue culture.
This review not only highlights the major biotechno-
logical advances made in guava during past years, but also
suggests how present technologies in tissue culture and
genetic engineering might affect the direction of future
research. The attempted and possible biotechnological
interventions in guava are presented in Fig. 1.
Achievements made in guava through tissue culture
The efficient regeneration of plants from cell, tissue, and
organ culture is recognized as prerequisite for application
of most modern genetic and biotechnological approaches
to crop improvement (Litz and Gray 1992). Several
workers have recognized that the two patterns of in
vitro differentiation, i.e., organogenesis and somatic
embryogenesis, are distinctly different process (Chris-
tianson 1987; Litz and Gray 1992). Successful regenera-
tion of plants from tissue culture offers excellent
opportunities for the improvement of guava. A report on
some morphological and cultural aspects of in vitro grown
guava tissue from fruit mesocarp was perhaps the first
attempt to manipulate somatic tissue (Schroeder 1961) of
this important fruit crop (Jaiswal and Amin 1992).
However, in recent years, several reports have been
published on regeneration of guava through organogenesis
and somatic embryogenesis (Tables 1, 2).
Problems associated with guava micropropagation
and their practical solutions
Morphogenesis from explants derived from mature trees is of
great commercial value because it can be applied in direct
cultivar improvement. However, there are several problems
associated with in vitro culture of explants obtained from
mature trees of guava such as browning or blackening of
medium and/or explants due to leaching of phenolics,
microbial contamination, and in vitro tissue recalcitrance etc.
High phenolic exudation during the excision of plants,
explant browning, medium discoloration, and slow growth
response have made an ordeal for workers dealing with
several woody tree species including guava. Browning of
media occurred as a result of oxidation of polyphenols
exuded from explants (Rout et al. 2000). In order to reduce
phenolic exudation, Amin and Jaiswal (1987, 1988) sug-
gested the pretreatment of explants with antioxidant solu-
tions. Explants were agitated for 30–40 min in 0.5% (w/v)
solution of polyvinylpolypyrrolidone (PVPP) containing
2% sucrose followed by dip in an antioxidant solution
(75 mg citric acid and 50 mg ascorbic acid l-1 water) after
surface sterilization of explants. Besides, 2–3 changes of
medium for the initial 10–15 days were essential for
Fig. 1 Biotechnological interventions in guava
2 Trees (2010) 24:1–12
123

controlling phenolic exudation and establishment of cul-
tures (Amin and Jaiswal 1987, 1988; Chandra et al.
2005b). Mishra et al. (2005) also recommended the
supplementation of 500 mg citric acid in the MS media and
initial incubation of cultures in complete dark for 24 h
to reduce phenolic browning of media and explants.
Table 1 Summary of work on organogenesis in guava
Cultivar Explant Mature/
juvenile
Medium ? PGRs References
Shoot multiplication Rooting
Banaras local Nodal segments M MS ? BAP � MS ? IBA NAA ? AC Amin and Jaiswal (1987)
– Shoot tips M MS ? BAP � MS ? IBA NAA ? AC Jaiswal and Amin (1987)
Chittidar Nodal segments M MS ? BAP � MS ? IBA NAA Amin and Jaiswal (1988)
– Shoot tips, Nodal segments,
Hypocotyl, Leaf segments
(Seedling)
J MS ? BAP MS basal Loh and Rao (1989)
– Shoot tips from seedling J OM ? BAP OM ? NAA ? IBA Papadatau et al. (1990)
Red Indian Nodal segments from seedling J MS ? BAP MS ? IBA or AC Yasseen et al. (1995)
Mara-7 Stem shoot M MS ? BAP, NAA, IBA Fuenmayor and Montero
(1997)
Allahabad
Safeda
Hypocotyl from seedling J MMS ? TDZ ? NAA � MMS ? IBA ? AC Singh et al. (2002)
– Nodal segments from green
house grown plants (GHRP)
and in vitro harvested axillary
buds (IVDS)
M/J MS ? BAP MS ? IBA Ali et al. (2003)
Aneuploid no.
82
Nodal segments M WPM ? BAP MS ? IBA ? NAA Meghwal et al. (2003)
Sardar Apical shoot tips M MS ? BAP ? IBA � MS ? IBA ?NAA Chandra et al. (2005b)
Pant Prabhat Nodal segments M MS ? BAP ? IBA � MS ? IBA ? NAA Mishra et al. (2005)
Safeda Shoot tips M MS ? BAP ? L-
glutamine
MS ? IAA ? IBA Zamir et al. (2007)
Safeda Seedling explants J MS ? Zea ? GA3 � MS ? IBA ? NAA Shah et al. (2008)
Banarasi local Nodal segments J MS ? BAP MS ? IBA Rai et al. (2009b)
AC activated charcoal, BAP 6, benzylaminopurine, IAA indole-3- acetic acid, IBA indole-3- butyric acid, MS Murashige and Skoog (1962)
medium, MMS Modified MS medium, NAA a – naphthalene acetic acid, OM Rugini Olive medium, TDZ N-phenyl-1, 2, 3-thidiazol-5yl-urea,
WPM woody plant medium
Table 2 Summary of work on somatic embryogenesis in guava
Cultivar Explant Medium ? PGRs References
Induction of somatic embryos Germination of somatic embryos
Sardar Immature and mature
fruit mesocarp
MMS ? L-glutamine
(400 mg l-1) ? ascorbic acid
(100 mg l-1) ? 2, 4-D (2 mg l-1)
MMS ? L-glutamine
(400 mg l-1) ? ascorbic acid
(100 mg l-1) ? 2, 4-D (2 mg l-1)
Chandra et al.
(2004)
– Immature zygotic
embryo
GSEM ? 1.75 lM IAA ? 58.4 lM
L-glutamine
MS medium Biswas et al. (2007)
Allahabad Safeda Immature fruit
mesocarp
MS ? 1, 2, 4-1-H Triazol
(4 mg l-1)
– Chandra et al.
(2005a)
Cuban Red Dwarf
EEA
Immature zygotic
embryo
� MS ? L-glutamine
(400 mg l-1) ? ascorbic acid
(100 mg l-1) ? 6% sucrose ? 2,
4-D (1 mg l-1)
Liquid � MS ? BAP
(0.25 mg l-1) ? Biobras-6
(10 lg l-1)
Kosky et al. (2005)
Banarasi local Immature zygotic
embryo
MS ? 5% sucrose ? 2, 4-D
(1 mg l-1)
� MS ? 3% sucrose Rai et al. (2007)
BAP 6, benzylaminopurine, 2, 4-D 2, 4-dichlorophenoxyacetic acid, GSEM guava somatic embryogenesis medium, IAA indole-3-acetic acid, MSMurashige and Skoog (1962) medium, MMS Modified MS medium
Trees (2010) 24:1–12 3
123

Papadatau et al. (1990) explored the morphogenic potential
of shoot tip explants excised from seedlings grown in a
growth chamber. Blackening of medium and necrosis of
explants were not observed by Papadatau et al. (1990). This
is probably because the young seedlings do not synthesize
higher quantities of phenolics when grown in a growth
chamber (Chandra et al. 2005b).
Microbial contamination is one of the most important
limiting factors for culture initiation using vegetative plant
parts (Krishna and Singh 2007). Microbial contamination
can result in the death of cultures, growth retardation,
necrosis, and altered morphogenic potential such as reduced
rates of multiplication and rooting (George 1993). Acosta
et al. (2002) have identified several microbial contaminants/
pathogens, i.e., Alternaria, Aspergillus, Cladosporium,
Colletotrichum, Curvularia, Fusarium, Nigrospora, Peni-
cillium, and Trichoderma, during in vitro culture of guava cv.
Enana Roja cubana and all the pathogens, except Nigros-
pora, have been detected during the in vitro establishment of
nodal segments. They noticed that approximately 50% of the
pathogens were eliminated by 10-min exposure to 3%
hypochlorite or use of 0.05 and 0.1% HgCl2. Ali et al. (2007)
observed that 67–95% seeds of guava were contaminated by
bacteria and fungi using standard tissue culture methods.
They recommended the application of 10% HCl for 24–72 h
followed by a 30-min treatment with 10% bleach (NaOCl) to
guava seeds to reduce contamination load.
Recalcitrance of tissue is another major problem for the
culture initiation using mature plant parts (Krishna and
Singh 2007). In the case of guava, the donor plants may be
limited and successful regeneration was observed mostly
from seedling explants (Loh and Rao 1989; Papadatau
et al. 1990; Yasseen et al. 1995; Fuenmayor and Montero
1997; Singh et al. 2002; Shah et al. 2008). The selection of
explants at a specific responsive stage of a mature tree’s
life cycle is of great importance to overcome recalcitrance
(Benson 2000; Krishna and Singh 2007). New vegetative
growth that occurs from the base of the main stem (off-
shoots) serves as a reliable source of shoot tip and nodal
segments for guava tissue culture (Amin and Jaiswal 1987;
Jaiswal and Amin 1987; Singh et al. 2004).
Organogenesis
Organogenesis involves adventitious and axillary shoot
production. Organogenesis comprises the formation of
unipolar structure (either shoot or root meristems) from
callus or directly from organized tissues (Singh et al. 2004).
In guava, organogenesis has been induced in vitro both from
mature tree explants (Amin and Jaiswal 1987, 1988; Jaiswal
and Amin 1987) and seedling explants (Loh and Rao 1989;
Papadatau et al. 1990; Yasseen et al. 1995; Fuenmayor and
Montero 1997; Singh et al. 2002; Shah et al. 2008).
Factors controlling organogenesis in guava
Success of in vitro regeneration depends on the control of
morphogenesis, which is influenced by several factors
namely kinds of tissue or explants, composition of med-
ium, plant growth regulators (PGRs), media additives,
culture environment etc.
Successful micropropagation, especially for difficult and
recalcitrant tree species, is mainly dependent on the quality
of explants and the response of explants is primarily deter-
mined by genotype, physiological state of the tissue, and
time of the year when the explants are collected and cultured
(Giri et al. 2004). Amin and Jaiswal (1987, 1988) have
compared the responses of nodal segments of mature tree and
nodal segments taken from in vitro proliferated shoots. The
performance of nodal segment taken from in vitro proliferated
shoots was better in comparison to that obtained from mature
tree. The probable reason for better response of in vitro nodal
segments as suggested by Amin and Jaiswal (1987) is the
absence of lag period between explanting and adaptation of
explants to in vitro conditions. Survival and growth response
of shoot tips were not as satisfactory as those of nodal seg-
ments (Amin and Jaiswal 1987). The greater responsiveness
of nodal segments over the shoot tips can be attributed to the
absence of apical dominance and presence of axillary buds at
a more advanced stage of development (Amin and Jaiswal
1987). Numerous studies have addressed the effect of season
on culture establishment in guava (Amin and Jaiswal 1987;
Mishra et al. 2005; Singh et al. 2005). The minimum phe-
nolic exudation and culture contamination, and maximum
survival and shoot proliferation were obtained from explants
harvested between April and June (Amin and Jaiswal 1987).
Ali et al. (2003) devised a protocol for regeneration of guava
using two different sources of explant, i.e., greenhouse-
grown plants (GHRP) and in vitro harvested axillary buds
(IVDS). The largest number of shoots and comparatively
better shoot growth was observed with IVDS.
In plant tissue culture, nutritional requirement for opti-
mal growth of a tissue in vitro may vary with species
(Bhojwani and Razdan 1996). Hence, media compositions
play a key role in morphogenesis and responses of
explants. In the case of guava, a number of media have
been used for initiation of culture during organogenesis.
But mostly MS (Murashige and Skoog 1962) medium was
used for shoot multiplication (Table 1). In a separate study,
Singh et al. (2002) reported the use of MS medium with
major salts reduced to one-half strength for shoot multi-
plication in cv. Allahabad safeda. In a few studies, other
media have also been used for optimum morphogenesis
(Papadatau et al. 1990; Meghwal et al. 2003).
Before exploiting plant tissue culture for commercial
purposes, detailed information regarding the requirement
of PGRs is necessary, and it has become a necessity to
4 Trees (2010) 24:1–12
123

standardize when dealing with tree species. The levels and
kinds of PGRs included in the culture medium largely
determine the success of tissue culture work. Root and
shoot initiations are closely regulated by the relative con-
centrations of auxin and cytokinin in the medium (Rout
et al. 2000). Cytokinin levels were shown to be the most
critical for multiplication of many tropical fruit tress. BAP
was the most common cytokinin used for guava propaga-
tion (Amin and Jaiswal 1987, 1988; Loh and Rao 1989;
Papadatau et al. 1990; Yasseen et al. 1995; Ali et al. 2003).
Superiority of BAP for shoot induction may be attributed to
the ability of plant tissues to metabolize BAP more readily
than other synthetic growth regulators or to the ability of
BAP to induce production of natural hormones such as
zeatin within the tissue (Malik et al. 2005). Singh et al.
(2002) were able to prompt shoot multiplication in cv.
Allahabad safeda after treatment with TDZ and NAA.
Other growth-enhancing medium additives including
sucrose (Amin and Jaiswal 1989a) and adenine sulfate
(Singh et al. 2002) had also significant effect on shoot
multiplication and elongation. Agar is most frequently used
as a gelling agent because of its desirable characteristics
such as clarity, stability, and its inertness (Pati et al. 2006).
Agar minimizes the water loss and allows good nutrient
diffusion (Amin and Jaiswal 1989a).
For any micropropagation protocol, successful rooting
of microshoots is a pre-requisite to facilitate their estab-
lishment in soil (Pati et al. 2006). Root initiation has been
encouraged in guava by incorporating either IBA alone or
with combination of NAA (Table 1), although Loh and
Rao (1989) were able to stimulate rooting only on MS
basal medium. Relatively low salt concentrations in med-
ium are known to enhance rooting of microshoots. Several
studies on guava indicate that half-strength MS medium
was adequate for root induction. Activated charcoal, often
used in plant tissue culture to improve cell growth and
differentiation, alone or sequentially after an auxin induced
rooting step of micropropagated shoots (Thomas 2008).
Rooting of shoots of guava has also been promoted by the
addition of activated charcoal (Amin and Jaiswal 1987;
Yasseen et al. 1995; Singh et al. 2002). Phloroglucinol was
found to inhibit in vitro rooting of shoots of guava and this
inhibition was more striking when it was used in combi-
nation with auxins (Amin and Jaiswal 1989b).
Somatic embryogenesis
Somatic embryogenesis is the process by which somatic
cells, under inductive conditions, generate embryogenic
cells, which undergo a series of morphological and bio-
chemical changes resulting in the formation of somatic
embryos (Zimmerman 1993; Komamine et al. 2005).
Somatic embryogenesis plays an important role in clonal
propagation. When integrated with conventional breeding
programs and molecular and cell biological techniques,
somatic embryogenesis provides a valuable tool to enhance
the pace of genetic improvement of commercial crop
species (Stasolla and Yeung 2003). As compared to
organogenesis, somatic embryogenesis provides an ideal
experimental process for investigation of plant differenti-
ation as well as a mechanism for expression of totipotency
in plant cells (Litz and Gray 1992). Many workers have
emphasized somatic embryogenesis as a preferred method
for genetic improvement and multiplication of valuable
germplasm of a number of woody perennials (Gupta and
Durzan 1987; Raj Bhansali 1990; Litz and Gray 1992).
In guava, considerable efforts have been made for in
vitro regeneration via somatic embryogenesis (Akhtar
1997; Akhtar et al. 2000; Chandra et al. 2004, 2005a;
Jaiswal and Jaiswal 2005; Biswas et al. 2007; Kosky et al.
2005; Rai et al. 2007). Immature zygotic embryos have
been utilized as the primary explants for the induction of
somatic embryogenesis by most of workers. Other explants
such as leaf, node, internode, petal, and mesocarp have also
been tested but failed to induce somatic embryos except
mesocarp with some success (Biswas et al. 2005; Chandra
et al. 2004, 2005a). Immature zygotic embryos have proved
to be an effective regenerable tissue for the many recalci-
trant tropical fruit species. Zygotic embryos are made up of
PEDCs (preembryogenic determined cells), in which, cells
have the embryogenic competence and can be easily
induced to follow the embryogenic pathways (Sharp et al.
1980).
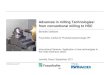
Development of different stages of somatic embryos on
zygotic embryo and germination of induced somatic
embryos of guava are presented in Fig. 2.
Induction of somatic embryogenesis
Induction of somatic embryogenesis in guava was affected
by nature of explants, physiological age of explants,
duration of treatment of 2, 4-dichlorophenoxyacetic acid
(2, 4-D) to explant, an interactive effect of 2, 4-D and
sucrose, different PGRs and their combination, and geno-
type (Akhtar et al. 2000; Jaiswal et al. 2005; Rai et al.
2007).
Requirement of auxin or other PGRs for the initiation of
somatic embryogenesis is largely determined by the
developmental stage of the explant tissue. The initiation of
the embryogenic pathway is restricted only to certain
responsive cells in the primary explant which have the
potential to activate those genes involved in the generation
of embryogenic cells. The competence for embryogenic
induction may be the result of varying auxin sensitivity to
these cells (Dudits et al. 1995; Arnold et al. 2002). Rai
et al. (2007) noted the zygotic embryos obtained from 10-
Trees (2010) 24:1–12 5
123

week-old fruits have the maximum ability to induce
somatic embryos in guava cv. Banarasi local. This is
probably due to their favorable physiological make-up at
that stage of development. Similarly, mesocarp explant
obtained from immature fruits has comparatively better
capacity to induce somatic embryos than those obtained
from mature fruits in cv. Sardar (Chandra et al. 2004).
In order to optimize the somatic embryo induction in
guava, a number of media are being used. However, the
most of successful cases recommended the use of MS
medium or derivative thereof (Chandra et al. 2004; Kosky
et al. 2005; Rai et al. 2007). Contrarily, Biswas et al.
(2005) did not obtain somatic embryos on MS, B5 or N6
culture media; thus they used a modified GSEM (guava
somatic embryogenesis medium) for the induction of
somatic embryogenesis. Moreover, all the above-said
media have high levels of ammonia and nitrate salts, which
played a key role in the induction of somatic embryogenesis
(Akhtar et al. 2000). Nitrogen in the form of amino acid
such as L-glutamine and organic compounds such as
ascorbic acid has been effective in induction of somatic
embryogenesis in guava (Chandra et al. 2004; Kosky et al.
2005; Biswas et al. 2005). The importance of reduced
nitrogen, certain amino acids, organic, and inorganic
nutrients has been reviewed in other tropical fruit species
by Akhtar et al. (2000). PGRs play an important role in the
induction of either unorganized callus growth or polarized
growth leading to somatic embryogenesis (Arnold et al.
2002; Rai et al. 2007). Auxin, particularly 2, 4-D is
required for the induction of somatic embryogenesis in
guava (Chandra et al. 2004; Kosky et al. 2005; Rai et al.
2007). Different auxins (IAA, IBA and NAA) and cyto-
kinins (BAP, Kin and TDZ) either alone or in combination
with 2, 4-D have been shown to be less effective than 2,
4-D alone for induction of somatic embryogenesis (Akhtar
et al. 2000; Jaiswal et al. 2005). Contrarily, Biswas et al.
(2005) obtained somatic embryos on IAA-containing
medium. The duration of explant exposure to a growth
regulator is an important factor for induction and deve-
lopment of somatic embryos. In most cases, auxin is
required only for induction of somatic embryogenesis and
is subsequently inhibitory for development of somatic
embryos. Rai et al. (2007) examined the treatment of
zygotic embryo for 8 days with 2, 4-D and observed that
this was effective for the induction of somatic embryo-
genesis. Continuous treatment for 60 days allowed differ-
entiation of somatic embryos only up to lower stage.
Various carbon sources were tested on zygotic embryo
explants for somatic embryo induction (Akhtar et al. 2000).
The rationale behind those experiments lay in the fact that
Fig. 2 Somatic embryogenesis
in guava. a Induction of somatic
embryogenesis on zygotic
embryo. b Development of
lower stages somatic embryos
on whole surface of zygotic
embryo after 4–5 weeks of
culture. c Development of
torpedo stages somatic embryos.
d Germination of somatic
embryos. e Development of a
well-developed plantlet from
germination of somatic embryo
6 Trees (2010) 24:1–12
123

the plant cannot use or metabolize all carbon sources
effectively, and thus those could be a limiting factor to
induction and development of somatic embryos. Sucrose
was shown to support somatic embryo induction and
addition of 5–6% sucrose in medium was found best for
induction of somatic embryogenesis (Kosky et al. 2005;
Rai et al. 2007). Somatic embryo induction in guava is
severely hampered by the presence of glucose, maltose,
lactose, fructose, sorbitol, and mannitol in medium (Akhtar
et al. 2000). Physical state (semisolid or liquid) and
strength of medium is often important for induction and
maintenance of somatic embryos in several tropical fruit
species including mango (Ara et al. 2000, 2004). In case of
guava, full-strength semisolid (solidified by 0.8% (w/v)
agar) medium was adequate for embryogenic response,
while in liquid medium much reduced level of induction
was obtained (Akhtar et al. 2000).
Development and maturation of somatic embryos
Synthetic auxins, particularly 2, 4-D, which are effective
for induction of somatic embryos, are usually less meta-
bolized by the cells than other auxins. Therefore, in order
to obtain development of somatic embryos it is necessary
to transfer the embryogenic cultures to medium lacking
auxin (Arnold et al. 2002). In guava, zygotic embryos
treated with 2, 4-D for 8 days were transferred to 2, 4-D
free medium for the development of somatic embryos (Rai
et al. 2007). Initial stage somatic embryos were formed
after 16–20 days. After 4–5 weeks of culture, entire sur-
face of zygotic embryos were covered with lower-stage
somatic embryos. Synchronization of embryogenic cultures
is difficult to achieve on development medium; it may be
due to establishment of polarity within culture relative to
the accessibility of 2, 4-D (Krishna and Singh 2007). In
order to achieve mature somatic embryos, lower-stage
somatic embryos induced on zygotic embryos transferred
to medium containing different concentrations of sucrose,
abscisic acid (ABA), two selected amino acids L-glutamine,
and L-proline or PEG (Rai et al. 2008a, 2009a). Among the
different concentrations of sucrose tried, a concentration of
5% was most effective for maturation of somatic embryos.
Rai et al. (2008a, 2009a) also suggested addition of ABA,
L-proline, or PEG to growth regulator free medium to
improve maturation of somatic embryos.
Germination of somatic embryos and plantlet development
Germination of somatic embryos and growth of regene-
rated plants depends on the conditions provided at earlier
stages when somatic embryos mature. In some cases, pre-
cocious germination of somatic embryos takes place
(Arnold et al. 2002; Jimenez 2005). Therefore, in order to
develop normal plants from somatic embryos, a dissection
of critical factors that might contribute to germination of
somatic embryos is required. In some cases, somatic
embryos develop into plants on culture medium without
PGRs, whereas there are some other cases where auxin or
cytokinin stimulates germination or an altered basal med-
ium was necessary (Jimenez 2005). In guava, lowering the
medium strength and sucrose concentration was necessary
for germination of somatic embryos. Germination of
mature somatic embryos was achieved on agar-solidified
half-strength MS medium containing 3% sucrose (Rai et al.
2007). Kosky et al. (2005) advocated the use of liquid
medium and addition of BAP, Biobras-6 (brassinosteroid
analog), and 2% sucrose in medium for germination of
somatic embryos.
Production of synthetic seeds
Encapsulation of somatic embryos or non-embryogenic
vegetative propagules to produce synthetic seeds could
possibly be utilized as means for germplasm storage and
transportation of elite germplasm (Krishna and Singh
2007). Guava is a cross-pollinated and vegetatively pro-
pagated crop (Doijode 2001). Therefore, it is a particularly
suitable candidate for synthetic seed technology. Success-
ful plantlet regeneration from encapsulated somatic
embryos of guava was reported by Akhtar (1997), Biswas
et al. (2007), Rai and Jaiswal (2008), and Rai et al. (2008a).
Torpedo stage somatic embryos were encapsulated in 2%
sodium alginate and 100 mM calcium chloride and
cultured on appropriate medium for plant regeneration
(Akhtar 1997). Maximum plantlet conversion from
encapsulated somatic embryos was obtained on growth
regulator free full-strength MS medium. Recently, Rai
et al. (2008b, c) have also employed the encapsulation of
vegetative propagules (shoot tips and nodal segments) for
the development of synthetic seeds in guava. A combina-
tion of 3% sodium alginate and 100 mM calcium chloride
was most suitable for formation of ideal synthetic seeds.
Maximum plantlet conversion from encapsulated shoot tips
was achieved on liquid MS medium (Rai et al. 2008b).
Plantlet conversion was also affected by medium strength
and sucrose concentrations in medium. These encapsulated
vegetative propagules could be potentially used in short-
term storage and germplasm exchange of elite genotype of
guava (Rai et al. 2008b, c).
In vitro storage of synthetic seeds
Two different approaches have been applied for storage of
synthetic seed of guava using slow-growth procedure (Rai
et al. 2008a, b). (1) Transferring the encapsulated somatic
embryos onto the full-strength MS medium containing
Trees (2010) 24:1–12 7
123

ABA (1 mg l-1) or 9% sucrose prior to culturing on ger-
mination medium (growth regulator free full-strength MS
medium ? 3% sucrose) resulted in extended storage of up
to 60 days (Rai et al. 2008a). The temporary suppression of
germination in encapsulated somatic embryos by ABA or
high sucrose offers a possibility of conservation of elite
genotype of guava for short period. (2) Encapsulated shoot
tips could be stored at low temperature (4�C) or room
temperature under minimal growth medium (sucrose
lacking medium) for different days (Rai et al. 2008b).
Results revealed that storage of encapsulated shoot tips
under minimal growth medium was better than storage at
low temperature (4�C) for conservation of guava.
In vitro selection
Development of improved variants obtained through in
vitro selection pressure technique is recommended for
increasing genetic diversity, both qualitatively and quan-
titatively inherited characters such as biotic and abiotic
stress tolerance, fruit quality, and yield etc. In vitro culture
of plant cells, tissues or organs on medium containing
selective agent offers the opportunity to regenerate and
select plants with desirable characteristics. The technique
has been effectively utilized to induce tolerance which
includes the use of some selective agents that permit the
preferential survival and growth of desired phenotypes
(Purohit et al. 1998). To create genetic variability for
selecting early bearing, short statured and less seeded
guava mutants, in vitro mutagenesis followed by micro-
propagation via shoot tips was carried out by Zamir et al.
(2003). Shoot tips were irradiated with gamma rays at
15–90 Gy using 60Co gamma cell source and cultured in
MS medium containing 3.0% sucrose, BAP and L-gluta-
mine. Sensitivity to radiation was evaluated by determining
the percentage shoot tip survival and shoot proliferation.
However, recovery of desired variants is still lacking.
Achievements made in guava through molecular
approaches
Molecular approaches are useful for characterizing the
genetic diversity among different cultivars or species, for
identifying genes of commercial interest and improvement
through genetic transformation technology. Some of the
important achievements made in guava through molecular
approaches are presented in Table 3.
Clonal identifications are traditionally based on various
morphological characters; however, morphological char-
acters may not be reliable to discriminate between closely
related guava genotypes (Chandra et al. 2005b). Most of the
cultivars grown on a commercial scale are seedling
selections from the well-known parent cultivars (Jaiswal
and Amin 1992). A close genetic relationship among
cultivars, somatic mutations, and changes due to environ-
mental alterations can create problems in correct identifi-
cation of germplasm. In recent years, different molecular
markers (RAPD, RFLP, AFLP, SSRs, ISSR, VNTRS) have
been employed for the investigations of cultivar origins and
taxonomic relationships of several plant species. Detection
of genetic variation is also important for micropropagation
and in vitro germplasm conservation to eliminate undesir-
able somaclonal variations. In guava, recently, a few reports
have been made on assessment of genetic diversity using
Random Amplified Polymorphic DNA (RAPD) markers
(Dahiya et al. 2002; Prakash et al. 2002; Chen et al. 2007;
Feria-Romero et al. 2009). Isolation of adequate quality of
genomic DNA for use in PCR-based DNA marker tech-
nology faces severe problems due to the presence of
inhibitors such as polysaccharides, which inhibit the enzy-
matic DNA processing or phenolics as inhibitors of PCR
reactions (Prakash et al. 2002). The well-established mod-
ified CTAB protocol (Porebski et al. 1997) yielded excel-
lent DNA templates for PCR amplification for guava
(Prakash et al. 2002). Prakash et al. (2002) analyzed
molecular diversity of 41 different genotypes of guava
collected from different parts of India by using RAPD
markers. The authors suggested that the genetic base of
Indian guava can be rated as low to moderate diversity and
various triploid seedless cultivars of guava are not gene-
tically identical and have independent origins. Dahiya et al.
(2002) also tried to determine genetic relationship in 13
north Indian cultivars of guava using RAPD markers. Chen
et al. (2007) using RAPD markers also attempted to deter-
mine phylogenetic relationship in 18 cultivars of Taiwan.
Apart from characterization and assessment of chemical
diversity, Feria-Romero et al. (2009) used RAPD amplifi-
cation method to identify molecular markers associated
with high quercetin accumulation in the leaves of guava
trees, selected from four different Mexican agronomic
regions. Simple sequence repeats (SSRs), also known as
microsatellites markers have been widely utilized in plant
genomic studies, and are reported to be more variable than
RFLPs and RAPDs (Krishna and Singh 2007). Microsatel-
lites markers to study genetic diversity in guava were
developed using a genomic library enriched for (GA)n and
(GT)n dinucleotide repeats and 23 nuclear SSR loci were
chosen to assess diversity in three guava species (Risterucci
et al. 2005). Hernandez-Delgado et al. (2007) studied the
amplified fragment length polymorphism (AFLP) analysis
of genetic relationship among 48 guava cultivars grown in
different parts of Mexico.
Genetic transformation opens the opportunity for
genetic manipulation of plants at cellular level and pro-
vides the means for modifying single horticultural traits
8 Trees (2010) 24:1–12
123

without significantly altering other aspects of the pheno-
type (Singh et al. 2004; Krishna and Singh 2007). The
main target of gene transfer techniques is to produce
improved varieties through the incorporation of horticul-
turally important genes into existing cultivars (Singh et al.
2004). Fruit trees are considered to be recalcitrant mate-
rial for genetic transformation studies and the main
impediment for genetic transformation is the regeneration
of transformed plantlets. Choice of explants having
competence for transformation and regeneration is a
crucial factor. Hence, efficient tissue culture techniques
become the base for genetic transformation studies (Giri
et al. 2004). The successful regeneration of genetically
transformed plants has been achieved in several tropical
fruit plant species (Gomez-Lim and Litz 2004). An
engineered Agrobacterium tumefaciens strain LBA 4404
(harboring binary vector pBI121 having selectable mark-
ers (nptII and GUS) with CaMV 35S promoter gene) has
been used for transformation of guava (Biswas et al.
2007). Recently, preliminary work on genetic transfor-
mation of guava with cold hardiness genes (CBF1, CBF2
and CBF3) also demonstrated by Biswas et al. (2005,
2007), however, complete regeneration of transformed
plants could not be achieved.
The development of recombinant DNA technology has
not only extremely impacted on our understanding of gene
structures, functions, and regulations, but also greatly
facilitated gene cloning, characterization, and their
expression into target species. Guava fruit was identified as
a particularly rich source of hydroperoxide lyase (HPL)
activity. HPL catalyzes the cleavage of 13- and 9-hydro-
peroxides of linoleic and linolenic acid into volatile C6- or
C9-aldehydes and C12- or C9-oxoacids, respectively (Kim
and Grosch 1981). The C6 and C9 volatile compounds have
a commercial value in the production of natural flavor in
the food industry, and are potentially important in plant
defense against pathogens (Croft et al. 1993). The HPL
enzyme purified from guava fruits was cloned by poly-
merase chain reaction with 30 and 50 rapid amplification of
cDNA ends (Tijet et al. 2000). The sequence shows
approximately 60–70% identity to known 13-hydroperox-
ide lyases. The cDNA was expressed in Escherichia coli.
Concluding remarks and future prospects
In the past 2–3 decades, encouraging progress has been
made regarding in vitro propagation of guava via
Table 3 Achievements made in
guava through molecular
approaches
Biotechnological tool Achievement References
DNA markers
RAPD Estimation of molecular diversity of 41
genotype of guava
Prakash et al. (2002)
Determination of genetic relationship in
13 north Indian guava cultivars
Dahiya et al. (2002)
Molecular identification of 18 guava
cultivars of Taiwan.
Chen et al. (2007)
Assessment of genetic relationship among
four Mexican guava cultivars to
estimate chemical (quercetin) diversity
Feria-Romero et al. (2009)
SSR Construction of (GA)n and (GT)nmicrosatellite-enriched library and
characterization of 23 nuclear simple
sequence repeat (SSR) loci in three
guava species for cultivars identification
and linkage mapping
Risterucci et al. (2005)
AFLP Genetic characterization of Mexican
native 48 guava cultivars
Hernandez-Delgado et al.
(2007)
Gene cloning Purification, molecular cloning, and
expression of the gene encoding 13-
hydroperoxide lyase from guava fruit
Tijet et al. (2000)
Genetic transformation Agrobacterium tumefaciens mediated
genetic transformation of guava,
resultants plants showed kanamycin
resistance
Biswas et al. (2007)
Introduction of cold tolerance genes
(CBF1, CBF2 and CBF3) to
organogenic and embryogenic explants
Biswas et al. (2007)
Trees (2010) 24:1–12 9
123

organogenesis and somatic embryogenesis by manipulation
of growth media and culture conditions as well as by
testing a variety of explant sources. However, some of the
long-standing problems such as guava wilt disease, short
shelf life of fruits, and abiotic stress sensitivity requires
urgent attention of researchers. There is need to exploita-
tion of modern tools of biotechnology in improvement of
guava. An increase in genetic transformation studies aimed
at improving visual and growth characteristics of the plants
has been hindered by low transformation efficiencies and
genotype dependence of protocols. As a result, guava
regeneration studies have once again emerged as an
essential complement of transformation studies. Since
genetic transformation system for guava is not yet well
developed, efforts need to be made to develop an efficient
transformation system for guava. For instance, insertion of
genes controlling ethylene biosynthesis could be helpful in
increasing shelf life of fruits of guava. Transformation of
genes encoding hydrolytic enzymes such as chitinase and
glucanase (which can degrade fungal cell wall) could also
be beneficial in development of wilt resistant plant of
guava (Chandra et al. 2005b). Such efforts will ultimately
provide the most rapid advances in guava.
Acknowledgments Financial assistance provided by Council of
Scientific & Industrial Research (CSIR), New Delhi, to the authors
(MKR and PA) is gratefully acknowledged. Suggestions by the
anonymous reviewers for improving the manuscript are also very
much appreciated.
References
Acosta M, Caballero I, Alvarado Y, Leiva M (2002) Epiphytic
microbiota and fungal contaminants from in vitro establishment
of guava (Psidium guajava L). Biotecnologia Vegetal 2:67–71
Akhtar N (1997) Studies on induction of somatic embryogenesis and
production of artificial seeds for micropropagation of a tropical
fruit tree guava (Psidium guajava L.). Ph.D. thesis, Banaras
Hindu University, Varanasi, India
Akhtar N, Kumari N, Pandey S, Ara H, Singh M, Jaiswal U, Jaiswal VS,
Jain SM (2000) Somatic embryogenesis in tropical fruit trees. In:
Jain SM, Gupta PK, Newton RJ (eds) Somatic embryogenesis in
woody plants, vol 6. Kluwer, Dordrecht, pp 93–140
Ali N, Mulwa RMS, Norton MA, Skirvin RM (2003) Micropropa-
gation of guava (Psidium guajava L.). J Hort Sci Biotechnol
78:739–741
Ali N, Mulwa RMS, Norton MA, Skirvin RM (2007) Radical
disinfestation protocol eliminates in vitro contamination in
Guava (Psidium guajava L.) seeds. Plant Cell Tissue Organ
Cult 91:295–298
Amin MN, Jaiswal VS (1987) Rapid clonal propagation of guava
through in vitro shoot proliferation on nodal explants of mature
trees. Plant Cell Tissue Organ Cult 9:235–243
Amin MN, Jaiswal VS (1988) Micropropagation as an aid to rapid
cloning of a guava cultivar. Sci Hort 36:89–95
Amin MN, Jaiswal VS (1989a) In vitro propagation of guava
(Psidium guajava L.): effects of sucrose, agar and pH on growth
and proliferation of shoots. Bangladesh J Bot 18:1–8
Amin MN, Jaiswal VS (1989b) Effects of phloroglucinol, sucrose pH
and temperature in vitro rooting of guava (Psidium guajava L.)
microcutting. Bangladesh J Bot 18:129–139
Ara H, Jaiswal U, Jaiswal VS (2000) Plant regeneration from
protoplasts of mango (Mangifera indica L.) through somatic
embryogenesis. Plant Cell Rep 19:622–627
Ara H, Jaiswal U, Jaiswal VS (2004) An improved method of
proliferation of proembryogenic calli of Mangifera indica L. var.
Amrapali for scale-up of somatic embryo production. Indian J
Biotechnol 3:229–234
Arnold SV, Sabala I, Bozhkov P, Dyachok J, Filonova L (2002)
Developmental pathways of somatic embryogenesis. Plant Cell
Tissue Organ Cult 69:233–249
Benson EE (2000) In vitro recalcitrance: an introduction. In Vitro
Cell Dev Biol Plant 36:141–148
Bhojwani SS, Razdan MK (1996) Plant tissue culture: theory and
practice. Elsevier, Amsterdam
Biswas BK, Yadav A, Joshee N, Yadav AK (2005) In vitro plant
regeneration and genetic transformation to enhance cold hardiness
in guava: a nutraceutical fruit. In: Proceeding of 1st international
guava symposium. CISH, Lucknow, India, (abst.) pp 31–32
Biswas BK, Joshee N, Yadav A, Yadav AK (2007) Development and
application of biotechnology in guava: a nutraceutical fruit. In:
Desjardins Y (ed) ISHS Acta Horticulturae 744: Ist international
symposium on human health effects of fruits and vegetables,
pp 267–276
Chandra R, Bajpai A, Gupta S, Tiwari RK (2004) Embryogenesis and
plant regeneration from mesocarp of Psidium guajava L.
(guava). Indian J Biotech 3:246–248
Chandra R, Mishra M, Abida M, Singh DB (2005a) Paclobutrazol
mediated somatic embryogenesis in guava. In: Abstract of 1st
international guava symposium. CISH, Lucknow, India, p 34
Chandra R, Mishra M, Bajpai A (2005b) Biotechnological interventions
for improvement of guava (Psidium guajava L.). In: Kishun R,
Mishra AK, Singh G, Chandra R (eds) Proceeding of 1st
international guava symposium. CISH, Lucknow, India, pp 19–25
Chen TW, Chang-Chai NG, CY Wang, Shyu YT (2007) Molecular
identification and analysis of Psidium guajava L. from indige-
nous tribes of Taiwan. J Food Drug Anal 15:82–88
Christianson ML (1987) Causal events in morphogenesis. In: Green
CE, Somers DA, Hackett WP, Biesboer DD (eds) Plant tissue
and cell culture. A.R. Liss., New York, pp 44–55
Croft KPC, Juttner F, Slusarenko AJ (1993) Volatile products of the
lipoxygenase pathway evolved from Phaseolus vulgaris (L.)
leaves inoculated with Pseudomonas syringae pv. phaseolicola.
Plant Physiol 101:13–24
Dahiya KK, Archak S, Karihaloo JL (2002) DNA fingerprinting of
guava (Psidium guajava L.) cultivars using RAPD markers.
Indian J Plant Genet Resour 15:112–115Dinesh MR, Iyer CPA (2005) Significant research achievement in
guava—improvement and future needs. In: Kishun R, Mishra
AK, Singh G, Chandra R (eds) Proceeding of 1st international
guava symposium. CISH, Lucknow, India, pp 7–16
Doijode SD (2001) Guava: Psidium guajava L. In: Doijode SD (ed)
Seed storage of horticultural crops. Haworth Press, New York,
pp 65–67
Dudits D, Gyorgyey J, Bogre L, Bako L (1995) Molecular biology of
somatic embryogenesis. In: Thorpe TA (ed) In vitro embryogen-
esis in plants. Kluwer Academic Publisher, Dordrecht, pp 267–308
Feria-Romero IA, Astudillo-Dela HV, Chavez-soto MA, Rivera-arce
E, Lopez M, Serrano H, Lozoya X (2009) RAPD markers
associated with quercetin accumulation in Psidium guajava. Bio
Plant 53:125–128
Fuenmayor MED, Montero NJM (1997) In vitro clonal propagation of
guava (Psidium guajava L.) from stem shoot of cv. Mara-7. Acta
Hort 452:47–51
10 Trees (2010) 24:1–12
123

George EF (1993) Plant propagation by tissue culture: part 1. The
technology, Exegetics, Edington, England
Giri C, Shyamkumar B, Anjaneyulu C (2004) Progress in tissue
culture, genetic transformation and applications of biotechnol-
ogy to trees: an overview. Trees Struct Funct 18:115–135
Gomez-Lim MA, Litz RE (2004) Genetic transformation of perennial
tropical fruits. In Vitro Cell Dev Biol Plant 40:442–449
Gupta PK, Durzan DJ (1987) Biotechnology of somatic polyembry-
ogenesis and plantlet regeneration in loblolly pine. Biotechnol
5:147–151
Hernandez-Delgado S, Padilla-Ramirez JS, Nava-Cedillo A, Mayek-
Perez N (2007) Morphological and genetic diversity of Mexican
guava germplasm. Plant Genet Resour 5:131–141
Jaiswal VS, Amin MN (1987) In vitro propagation of guava from
shoot cultures of mature trees. J Plant Physiol 130:7–12
Jaiswal VS, Amin MN (1992) Guava and jackfruit. In: Hammerschlag
FA, Litz RE (eds) Biotechnology of perennial fruit crops,
Biotechnology in agriculture, vol 8. CAB International, Wal-
lingford, pp 421–431
Jaiswal U, Jaiswal VS (2005) Psidium guajava. In: Litz RE (ed)
Biotechnology of fruit and nut crops. CAB International,
Wallingford, pp 394–401
Jaiswal VS, Akhtar N, Rai MK (2005) Role of different factors on
somatic embryogenesis and synthetic seed production in guava.
In: Abstract of 1st international guava symposium. CISH,
Lucknow, India, p 36
Jimenez VM (2005) Involvement of plant hormones and plant growth
regulators on in vitro somatic embryogenesis. Plant Growth
Regul 47:91–110
Kim IS, Grosch W (1981) Partial-purification and properties of a
hydroperoxide lyase from fruits of Pear. J Agric Food Chem
29:1220–1225
Komamine A, Murata N, Nomura K (2005) Mechanisms of somatic
embryogenesis in carrot suspension cultures—morphology,
physiology, biochemistry, and molecular biology. In vitro Cell
Dev Biol Plant 41:6–10
Kosky RG, Perozo JV, Valero NA, Penalver DA (2005) Somatic
embryo germination of Psidium guajava L. in the Rita�temporary immersion system, on semisolid medium. In: Hvos-
lef-Eide KA, Preil W (eds) Liquid culture systems for in vitro
plant propagation. Springer-Verlag, New York, pp 225–229
Krishna H, Singh SK (2007) Biotechnological advances in mango
(Mangifera indica L.) and their future implication in crop
improvement—a review. Biotech Adv 25:223–243
Litz RE, Gray DJ (1992) Organogenesis and somatic embryogenesis.
In: Hammerschlag FA, Litz RE (eds) Biotechnology of perennial
fruit crops. CAB International, Wallingford, pp 3–34
Litz RE, Jaiswal VS (1991) Micropropagation of tropical and
subtropical fruits. In: Debergh PC, Zimmerman RH (eds)
Micropropagation. Kluwer Academic Publishers, Dordrecht, pp
247–263
Loh CS, Rao AN (1989) Clonal propagation of guava (Psidiumguajava L.) from seedlings and grafted plants and adventitious
shoot formation in vitro. Sci Hort 39:31–39
Malik SK, Chaudhury R, Kalia RK (2005) Rapid in vitro multipli-
cation and conservation of Garcinia indica: a tropical medicinal
tree species. Sci Hort 106:539–553
Meghwal PR, Singh SK, Sharma HC (2003) Micropropagation of
Aneuploid guava. Indian J Hort 60:29–33
Mishra AK (2005) Important diseases of guava in India with special
reference to wilt. In: Kishun R, Mishra AK, Singh G, Chandra R
(eds) Proceeding of 1st international guava symposium. CISH,
Lucknow, India, pp 75–90
Mishra DS, Tiwari JP, Lal S (2005) In vitro cloning of guava (Psidiumguajava L.) cv. Pant Prabahat. In: Abstract of 1st international
guava symposium. CISH, Lucknow, India, pp 32–33
Murashige T, Skoog FA (1962) Revised medium for rapid growth and
bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Papadatau P, Pontikis CA, Ephtimiadou E, Lydaki M (1990) Rapid
multiplication of guava seedlings by in vitro shoot tip culture.
Sci Hort 45:99–108
Pati PK, Rath SP, Sharma M, Sood A, Ahuja PS (2006) In vitro
propagation of rose—a review. Biotechnol Adv 24:94–114
Pommer CV, Murakami KRN (2008) Breeding guava (Psidiumguajava L.). In: Jain SM, Priyadarshan PM (eds) Breeding
plantation tree crops: tropical species. Springer, New York, pp
83–119
Porebski S, Bailey LG, Baurn BR (1997) Modification of a CTAB
DNA extraction protocol for plants containing high polysaccha-
ride and polyphenol components. Plant Mol Biol Reprod 15:8–15
Prakash DP, Narayanaswamy P, Sondur SN (2002) Analysis of
molecular diversity in Guava using RAPD markers. J Hort Sci
Biotechnol 77:287–293
Purohit M, Srivastava S, Srivastava PS (1998) Stress tolerant plants
through tissue culture. In: Srivastava PS (ed) Plant tissue culture
and molecular biology: application and prospects. Narosa
Publishing House, New Delhi, pp 554–578
Rai MK, Jaiswal VS (2008) Synthetic seeds of guava (Psidiumguajava L) from somatic embryos and plant regeneration.
In: Arya ID, Arya S (eds) Utilization of biotechnology in
plant sciences. Microsoft Printech (I) Pvt Ltd, Dehradun,
pp 217–222
Rai MK, Akhtar N, Jaiswal VS (2007) Somatic embryogenesis and
plant regeneration in Psidium guajava L. cv. Banarasi local. Sci
Hort 113:129–133
Rai MK, Jaiswal VS, Jaiswal U (2008a) Effect of ABA and sucrose
on germination of encapsulated somatic embryos of guava
(Psidium guajava L.). Sci Hort 117:302–305
Rai MK, Jaiswal VS, Jaiswal U (2008b) Encapsulation of shoot tips of
guava (Psidium guajava L.) for short-term storage and germ-
plasm exchange. Sci Hort 118:33–38
Rai MK, Jaiswal VS, Jaiswal U (2008c) Alginate-encapsulation of
nodal segments of guava (Psidium guajava L.) for germplasm
exchange and distribution. J Hort Sci Biotechnol 83:569–573
Rai MK, Jaiswal VS, Jaiswal U (2009a) Effect of selected amino
acids and polyethylene glycol on maturation and germination of
somatic embryos of guava (Psidium guajava L.). Sci Hort
121:233–236
Rai MK, Jaiswal VS, Jaiswal U (2009b) Shoot multiplication and
plant regeneration of guava (Psidium guajava L.) from nodal
explants of in vitro raised plantlets. J Fruit Ornam Plant Res
17:29–38
Raj Bhansali R (1990) Somatic embryogenesis and regeneration of
plantlets in pomegranate. Ann Bot 66:249–253
Ribeiro IJA, Pommer CV (2004) Breeding guava (Psidium guajava)
for resistance to rust caused by Puccinia psidii. Acta Hort
632:75–78
Risterucci AM, Duval MF, Rohde W, Billotte N (2005) Isolation and
characterization of microsatellite loci from Psidium guajava L.
Mol Ecol Notes 5:745–748
Rout GR, Samantaray S, Das P (2000) In vitro manipulation and
propagation of medicinal plants. Biotechnol Adv 18:91–120
Schroeder CA (1961) Some morphological aspects of fruit tissues
grown in vitro. Bot Gaz 122:198–204
Shah ST, Zamir R, Ahmad J, Ali H, Lutfullah G (2008) In vitro
regeneration of plantlets from seedling explants of guava
(Psidium guajava L.) cv. Safeda. Pak J Bot 40:1195–1200
Sharp WR, Sondahl MR, Caldas LS, Maraffa SB (1980) The physiology
of in vitro asexual embryogenesis. Hortic Rev 268–310
Singh G (2005) Strategies for improved production in guava. In: Kishun
R, Mishra AK, Singh G, Chandra R (eds) Proceeding of 1st
international guava symposium. CISH, Lucknow, India, pp 26–39
Trees (2010) 24:1–12 11
123

Singh SK, Meghwal PR, Sharma HC, Singh SP (2002) Direct shoot
organogenesis on explants from germinated seedlings of Psidiumguajava L. cv. Allahabad Safeda. Sci Hort 95:213–221
Singh M, Jaiswal U, Jaiswal VS (2004) In vitro regeneration and
improvement in tropical fruit trees: an assessment. In: Srivastava
PS, Narula A, Srivastava S (eds) Plant biotechnology and molec-
ular markers. Anamanya Publishers, New Delhi, pp 228–243
Singh D, Mohammed S, Shukla AK (2005) Micropropagation of
guava as influenced by season, source and pretreatment to
explant. In: Abstract of 1st international guava symposium.
CISH, Lucknow, India, p 33
Srivastava U (2005) Genetic resources management in guava. In:
Kishun R, Mishra AK, Singh G, Chandra R (eds) Proceeding
of 1st international guava symposium. CISH, Lucknow, India,
pp 17–18
Stasolla C, Yeung EC (2003) Recent advances in conifer somatic
embryogenesis: improving somatic embryo quality. Plant Cell
Tissue Organ Cult 74:15–35
Thomas TD (2008) The role of activated charcoal in plant tissue
culture. Biotechnol Adv 26:618–631
Thorpe TA, Harpy IS, Kumar PP (1991) Application of microprop-
agation to forestry. In: Debergh PC, Zimmerman RH (eds)
Micropropagation. Kluwer Academic Publishers, Dordrecht,
pp 311–336
Tijet N, Waspi U, Gaskin DJH, Hunziker P, Muller BL, Vulfson EN,
Slusarenko A, Brash AR, Whitehead IM (2000) Purification,
molecular cloning, and expression of the gene encoding fatty
acid 13-hydroperoxide lyase from guava fruit (Psidium guajava).
Lipids 35:709–720
Yasseen M, Barringer SA, Schnell RJ, Splittoesser WE (1995) In
vitro shoot proliferation of guava (Psidium guajava L.) from
germinated seedlings. Plant Cell Rep 14:525–528
Zamir R, Khattak GSS, Mohammad T, Ali N (2003) In vitro
mutagenesis in guava (Psidium guajava L.). Pak J Bot 35:825–
828
Zamir R, Ali N, Shah ST, Muhammad T, Shah SA (2007) In vitro
regeneration of guava (Psidium guajava L.) from shoot tips of
mature trees. Pak J Bot 39:2395–2398
Zimmerman JL (1993) Somatic embryogenesis: a model of early
development in higher plants. Plant Cell 5:1411–1423
12 Trees (2010) 24:1–12
123
Recommended