
Autoregulatory binding sites in the zebrafish six3apromoter region define a new recognition sequence forSix3 proteinsClotilde S. Suh, Staale Ellingsen*, Lars Austbø, Xiao-Feng Zhao, Hee-Chan Seo and Anders Fjose
Department of Molecular Biology, University of Bergen, Norway
Introduction
Vertebrate Six3 proteins have important roles during
the development of eyes and forebrain, and belong to
the Six ⁄Sine oculis family. This family represents a
divergent group of the homeodomain (HD) superfam-
ily of transcription factors [1,2]. The 60 amino acid
HD, which is a DNA-binding domain, has a conserved
global fold consisting of three a-helices and a flexible
N-terminal arm that becomes more ordered upon
DNA binding [3–6]. Binding to specific DNA
sequences is mediated by interactions between particu-
lar amino acids in the ‘recognition helix’ and bases in
the major groove, and specific contacts between the
N-terminal arm and the minor groove [4,7,8].
Specific base contacts in the minor groove involve
the first two nucleotides in the TAAT core, and are
achieved through interactions with residues at posi-
Keywords
chromatin; eye development; homeobox;
transcription factor; transgenic
Correspondence
A. Fjose, Department of Molecular Biology,
University of Bergen, PO Box 7803, N-5020
Bergen, Norway
Fax: +47 555 89683
Tel: +47 555 84331
E-mail: [email protected]
*Present address
National Institute of Nutrition and Seafood
Research, NIFES, PO Box 2029 Nordnes,
N-5817 Bergen, Norway
(Received 11 September 2009, revised
22 December 2009, accepted 29 January
2010)
doi:10.1111/j.1742-4658.2010.07599.x
The homeodomain (HD) transcription factor Six3, which is a member of
the Six ⁄Sine oculis family, is essential for development of the eyes and fore-
brain in vertebrates. It has recently been claimed that the HDs of Six3 and
other members of the Six family have a common recognition sequence,
TGATAC. However, a different recognition sequence including the typical
TAAT core motif, which has not yet been fully defined, has also been pro-
posed for the Six3 HD in mice. Our study of the zebrafish orthologue
six3a, which has an identical HD, shows that it binds in vitro to multiple
TAAT-containing sites within its promoter region. Comparison of the dif-
ferent binding affinities for these sequences identifies three high-affinity
sites with a common TAATGTC motif. Notably, this new recognition
sequence, which is supported by our analysis of the influence of single-
nucleotide substitutions on the DNA-binding affinity, is distinct from all
of the DNA-binding specificities previously described in surveys of HDs.
In addition, our comparison of Six3a HD binding to the novel TAATGTC
motif and the common recognition sequence of Six family HDs
(TGATAC) shows very similar affinities, suggesting two distinct DNA-
binding modes. Transient reporter assays of the six3a promoter in zebrafish
embryos also indicate that the three high-affinity sites are involved in auto-
regulation. In support of this, chromatin immunoprecipitation experiments
show enrichment of Six3a binding to a six3a promoter fragment containing
two clustered high-affinity sites. These findings provide strong evidence that
the TAATGTC motif is an important target sequence for vertebrate Six3
proteins in vivo.
Abbreviations
ChIP, chromatin immunoprecipitation; EGFP, enhanced green fluorescent protein; EMSA, electrophoretic mobility shift assay; GFP, green
fluorescent protein; GST, glutathione-S-transferase; HD, homeodomain; hpf, hours postfertilization.
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1761

tions 2, 3 and 5–8 in the N-terminal arm. Also, an
arginine at position 5 is important in most HDs
[4,5,9]. Similarly, the recognition helix makes specific
contacts with several nucleotides in the core motif, and
its residues at positions 47, 50 and 54 also specify two
adjacent nucleotides 3¢ of the TAAT core [5,10]. For
example, HDs containing Lys50 and Gln50 have
been shown to bind specifically to TAATCC and
TAATGG, respectively [11–13]. Recent studies indicate
that the sequence recognition also depends on a few
additional flanking nucleotides, and this variation in
specificity may include more than 60 distinct DNA-
binding activities [14].
The Six ⁄Sine oculis family proteins also have a con-
served Six domain of 115–119 amino acids involved in
protein–protein interactions [1,15], and can be subdi-
vided into three subfamilies, Six1 ⁄2, Six4 ⁄ 5, and
Six3 ⁄ 6, on the basis of their HD sequence divergence
and characteristic tetrapeptides in the N-terminal arm
[16]. The absence of Arg5 in their N-terminal arms
may explain, in part, why regulatory DNA sequences
that bind Six1 ⁄ 2 and Six4 ⁄ 5 do not contain the TAAT
core [17]. Although the Six3 ⁄ 6 proteins also lack Arg5,
their HDs are more distinct from those of the members
of the other two subfamilies, and various studies have
suggested that their DNA-binding specificity is differ-
ent [1,16,18]. In a previous investigation of murine
Six3, a common TAAT core motif was identified by
in vitro binding site selection from a randomized pool
of oligonucleotides, and autoregulatory binding sites
containing the TAAT core were also identified in the
promoter of the Six3 gene [18]. However, more recent
studies have provided evidence that Six3 ⁄ 6 proteins
have similar in vitro DNA-binding specificities to those
of the other Six family members [10,14]. Further analy-
sis of the binding affinities of functional Six3 target
sites and how they function in vivo may help to clarify
uncertainties regarding the recognition sequences of
these HD proteins.
Two orthologues of the murine Six3 gene, six3a and
six3b, are present in the zebrafish, Danio rerio, owing
to the extra genome duplication that occurred before
the teleost radiation [15,19]. An additional Six3-like
gene in zebrafish, six7, was probably generated by an
independent gene duplication event [20]. Several stud-
ies of Six3 homologues in mouse, fish and Xenopus
have demonstrated that these genes are essential for
forebrain and eye development, and their importance
is also reflected in human mutant phenotypes [21–25].
In these processes, Six3 proteins have been shown to
act both as transcriptional activators and repressors,
and as regulators of cell proliferation through interac-
tions with the cell cycle inhibitor Geminin [26,27].
Studies on Six3 proteins in zebrafish have contrib-
uted to our understanding of their functional roles in
forebrain and eye development [22,28–30], and how
they can act as transcriptional repressors through
interactions with members of the Groucho family of
corepressors [31]. Relatively little is known about the
regulation of zebrafish six3 genes during development
[32,33], but essential cis-regulatory elements have been
identified in one of the gene homologues in medaka
fish [34].
We have investigated the significance of the high
density of TAAT sequences present in the zebrafish
six3a promoter region. Our comparison of the relative
binding affinities of these potential target sites for the
Six3a HD identified several strong binding sites that
defined the sequence TAATGTC as a recognition
motif. Results from chromatin immunoprecipitation
(ChIP) experiments and transient reporter assays of
the six3a promoter in zebrafish embryos supported the
functional role of these high-affinity sites in mediating
autoregulation. Hence, it is also likely that many of
the target genes of vertebrate Six3 proteins are recog-
nized on the basis of high-affinity binding to sequence
elements containing this motif.
Results
A 3.6 kb promoter region of six3a recapitulates
early embryonic expression
The genomic region upstream of the translational start
site in zebrafish six3a is syntenic with a 4.5 kb pro-
moter region of the orthologous medaka (Oryzia latipes)
gene olSix3.2, which contains cis-regulatory elements
responsible for its spatiotemporal regulation in embryos
[34]. Additional evidence that the corresponding
promoter region of zebrafish six3a contains cis-acting
elements required for early expression in the eyes and
forebrain was obtained from transient expression assays
with injected reporter constructs [33].
To analyse the significance of the six3a promoter
region, we fused a 3.6 kb genomic fragment to the
ORF of an enhanced green fluorescent protein (EGFP)
reporter gene in a Tol2 vector (Fig. 1A), and used this
construct to establish transgenic lines of zebrafish (see
Experimental procedures). From 53 founders crossed
to wild-type fish, we identified three transgenic lines
with EGFP expression comparable to that of the
endogenous six3a gene (data not shown). The trans-
genic line Tg(3.6S3a:EGFP) was chosen for direct
comparison of EGFP expression with the spatial distri-
bution of endogenous six3a transcripts by in situ
hybridization. At 12 h postfertilization (hpf), EGFP
New recognition sequence for Six3 C. S. Suh et al.
1762 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS
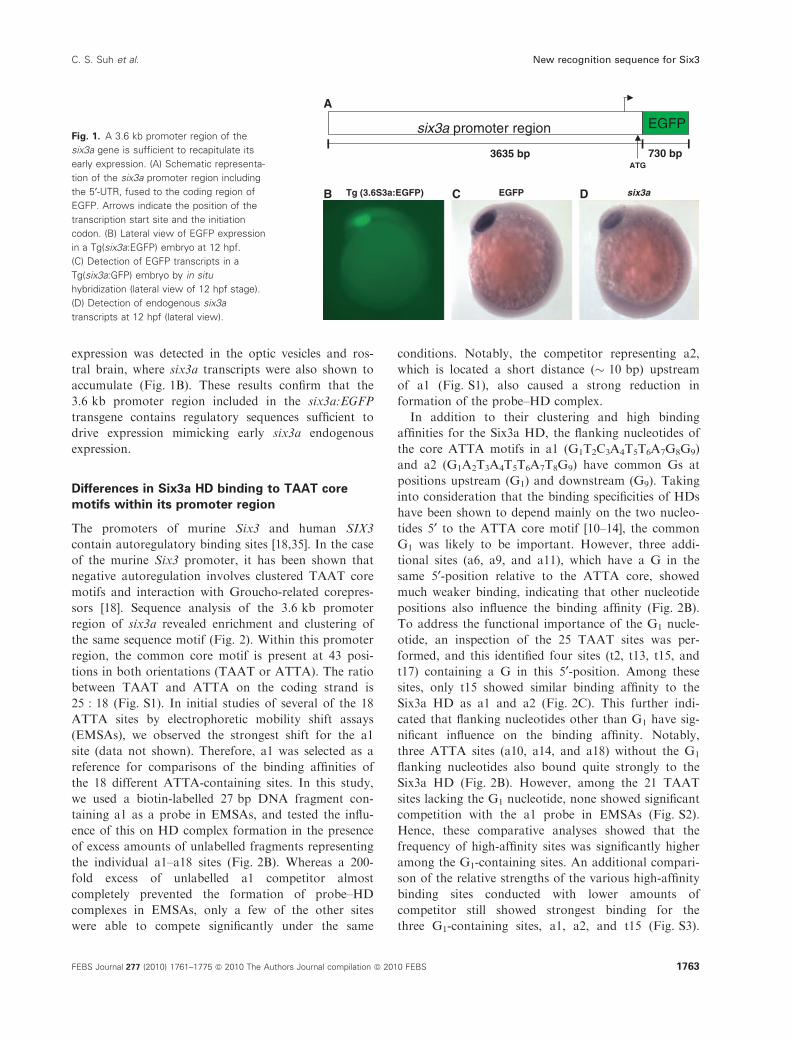
expression was detected in the optic vesicles and ros-
tral brain, where six3a transcripts were also shown to
accumulate (Fig. 1B). These results confirm that the
3.6 kb promoter region included in the six3a:EGFP
transgene contains regulatory sequences sufficient to
drive expression mimicking early six3a endogenous
expression.
Differences in Six3a HD binding to TAAT core
motifs within its promoter region
The promoters of murine Six3 and human SIX3
contain autoregulatory binding sites [18,35]. In the case
of the murine Six3 promoter, it has been shown that
negative autoregulation involves clustered TAAT core
motifs and interaction with Groucho-related corepres-
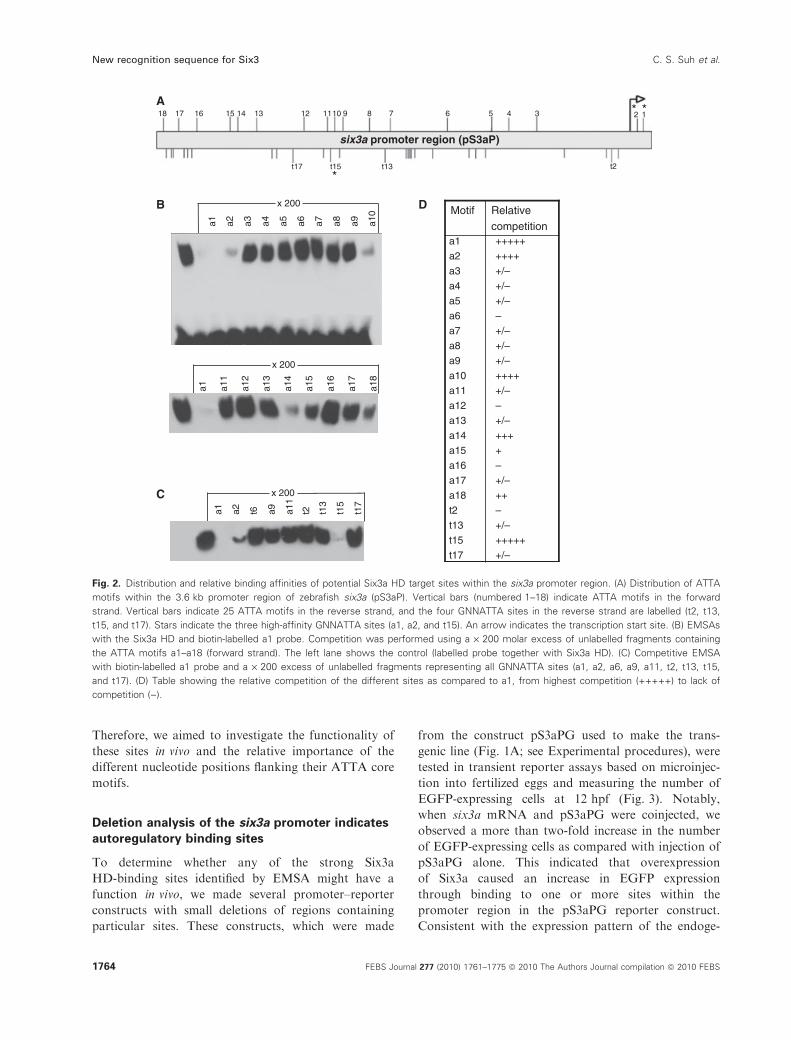
sors [18]. Sequence analysis of the 3.6 kb promoter
region of six3a revealed enrichment and clustering of
the same sequence motif (Fig. 2). Within this promoter
region, the common core motif is present at 43 posi-
tions in both orientations (TAAT or ATTA). The ratio
between TAAT and ATTA on the coding strand is
25 : 18 (Fig. S1). In initial studies of several of the 18
ATTA sites by electrophoretic mobility shift assays
(EMSAs), we observed the strongest shift for the a1
site (data not shown). Therefore, a1 was selected as a
reference for comparisons of the binding affinities of
the 18 different ATTA-containing sites. In this study,
we used a biotin-labelled 27 bp DNA fragment con-
taining a1 as a probe in EMSAs, and tested the influ-
ence of this on HD complex formation in the presence
of excess amounts of unlabelled fragments representing
the individual a1–a18 sites (Fig. 2B). Whereas a 200-
fold excess of unlabelled a1 competitor almost
completely prevented the formation of probe–HD
complexes in EMSAs, only a few of the other sites
were able to compete significantly under the same
conditions. Notably, the competitor representing a2,
which is located a short distance (� 10 bp) upstream
of a1 (Fig. S1), also caused a strong reduction in
formation of the probe–HD complex.
In addition to their clustering and high binding
affinities for the Six3a HD, the flanking nucleotides of
the core ATTA motifs in a1 (G1T2C3A4T5T6A7G8G9)
and a2 (G1A2T3A4T5T6A7T8G9) have common Gs at
positions upstream (G1) and downstream (G9). Taking
into consideration that the binding specificities of HDs
have been shown to depend mainly on the two nucleo-
tides 5¢ to the ATTA core motif [10–14], the common
G1 was likely to be important. However, three addi-
tional sites (a6, a9, and a11), which have a G in the
same 5¢-position relative to the ATTA core, showed
much weaker binding, indicating that other nucleotide
positions also influence the binding affinity (Fig. 2B).
To address the functional importance of the G1 nucle-
otide, an inspection of the 25 TAAT sites was per-
formed, and this identified four sites (t2, t13, t15, and
t17) containing a G in this 5¢-position. Among these
sites, only t15 showed similar binding affinity to the
Six3a HD as a1 and a2 (Fig. 2C). This further indi-
cated that flanking nucleotides other than G1 have sig-
nificant influence on the binding affinity. Notably,
three ATTA sites (a10, a14, and a18) without the G1
flanking nucleotides also bound quite strongly to the
Six3a HD (Fig. 2B). However, among the 21 TAAT
sites lacking the G1 nucleotide, none showed significant
competition with the a1 probe in EMSAs (Fig. S2).
Hence, these comparative analyses showed that the
frequency of high-affinity sites was significantly higher
among the G1-containing sites. An additional compari-
son of the relative strengths of the various high-affinity
binding sites conducted with lower amounts of
competitor still showed strongest binding for the
three G1-containing sites, a1, a2, and t15 (Fig. S3).
six3a promoter region
ATG
EGFP
730 bp3635 bp
A
B C DTg (3.6S3a:EGFP) six3aEGFP
Fig. 1. A 3.6 kb promoter region of the
six3a gene is sufficient to recapitulate its
early expression. (A) Schematic representa-
tion of the six3a promoter region including
the 5¢-UTR, fused to the coding region of
EGFP. Arrows indicate the position of the
transcription start site and the initiation
codon. (B) Lateral view of EGFP expression
in a Tg(six3a:EGFP) embryo at 12 hpf.
(C) Detection of EGFP transcripts in a
Tg(six3a:GFP) embryo by in situ
hybridization (lateral view of 12 hpf stage).
(D) Detection of endogenous six3a
transcripts at 12 hpf (lateral view).
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1763
Therefore, we aimed to investigate the functionality of
these sites in vivo and the relative importance of the
different nucleotide positions flanking their ATTA core
motifs.
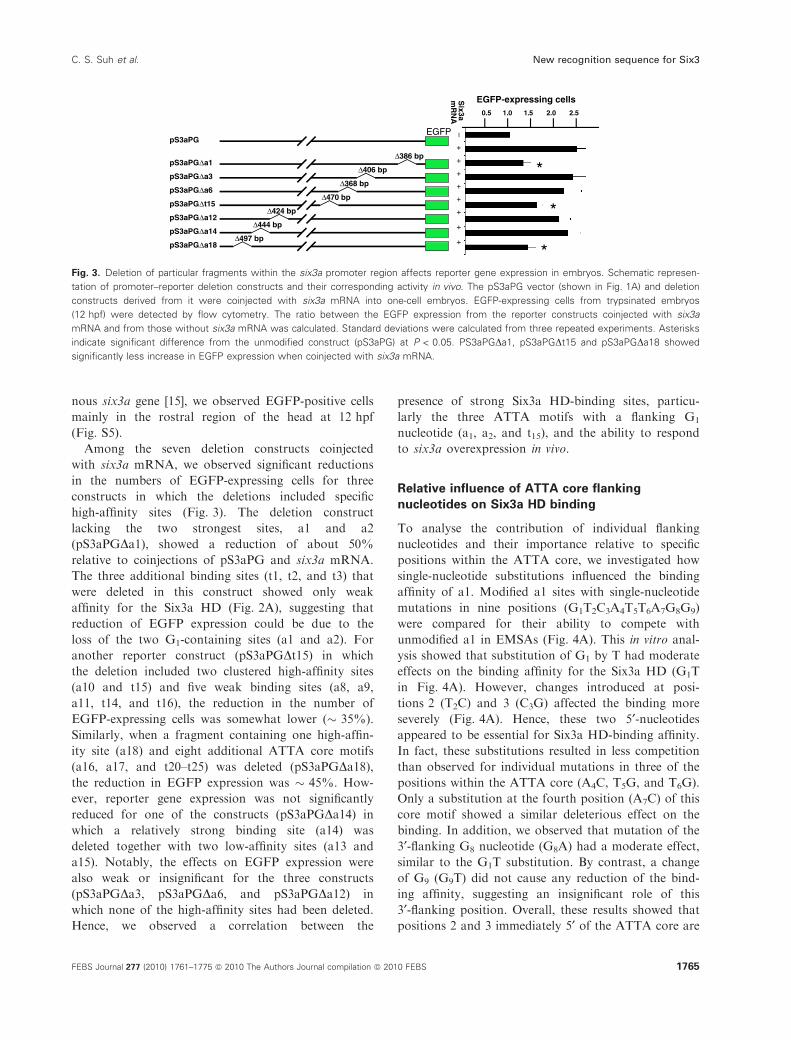
Deletion analysis of the six3a promoter indicates
autoregulatory binding sites
To determine whether any of the strong Six3a
HD-binding sites identified by EMSA might have a
function in vivo, we made several promoter–reporter
constructs with small deletions of regions containing
particular sites. These constructs, which were made
from the construct pS3aPG used to make the trans-
genic line (Fig. 1A; see Experimental procedures), were
tested in transient reporter assays based on microinjec-
tion into fertilized eggs and measuring the number of
EGFP-expressing cells at 12 hpf (Fig. 3). Notably,
when six3a mRNA and pS3aPG were coinjected, we
observed a more than two-fold increase in the number
of EGFP-expressing cells as compared with injection of
pS3aPG alone. This indicated that overexpression
of Six3a caused an increase in EGFP expression
through binding to one or more sites within the
promoter region in the pS3aPG reporter construct.
Consistent with the expression pattern of the endoge-
a1 a11
a12
a13
a14
a15
a16
a17
a18
x 200
a1 a2 t6 a9 a11
t2 t13
t15
t17
x 200
a1 a2 a3 a4 a5 a6 a7 a8 a9 a10
x 200
+++++
++++
+/–
+/–
+/–
–
+/–
+/–
+/–
++++
+/–
–
+/–
+++
+
–
+/–
++
–
+/–
+++++
+/–
a1
a2
a3
a4
a5
a6
a7
a8
a9
a10
a11
a12
a13
a14
a15
a16
a17
a18
t2
t13
t15
t17
Relative
competition
Motif
18A
B
C
D
17 16 15 14 13 12 1110 9 8 7 6 5 4 3 2
t2
1**
t13t15t17*
six3a promoter region (pS3aP)
Fig. 2. Distribution and relative binding affinities of potential Six3a HD target sites within the six3a promoter region. (A) Distribution of ATTA
motifs within the 3.6 kb promoter region of zebrafish six3a (pS3aP). Vertical bars (numbered 1–18) indicate ATTA motifs in the forward
strand. Vertical bars indicate 25 ATTA motifs in the reverse strand, and the four GNNATTA sites in the reverse strand are labelled (t2, t13,
t15, and t17). Stars indicate the three high-affinity GNNATTA sites (a1, a2, and t15). An arrow indicates the transcription start site. (B) EMSAs
with the Six3a HD and biotin-labelled a1 probe. Competition was performed using a · 200 molar excess of unlabelled fragments containing
the ATTA motifs a1–a18 (forward strand). The left lane shows the control (labelled probe together with Six3a HD). (C) Competitive EMSA
with biotin-labelled a1 probe and a · 200 excess of unlabelled fragments representing all GNNATTA sites (a1, a2, a6, a9, a11, t2, t13, t15,
and t17). (D) Table showing the relative competition of the different sites as compared to a1, from highest competition (+++++) to lack of
competition ()).
New recognition sequence for Six3 C. S. Suh et al.
1764 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS
nous six3a gene [15], we observed EGFP-positive cells
mainly in the rostral region of the head at 12 hpf
(Fig. S5).
Among the seven deletion constructs coinjected
with six3a mRNA, we observed significant reductions
in the numbers of EGFP-expressing cells for three
constructs in which the deletions included specific
high-affinity sites (Fig. 3). The deletion construct
lacking the two strongest sites, a1 and a2
(pS3aPGDa1), showed a reduction of about 50%
relative to coinjections of pS3aPG and six3a mRNA.
The three additional binding sites (t1, t2, and t3) that
were deleted in this construct showed only weak
affinity for the Six3a HD (Fig. 2A), suggesting that
reduction of EGFP expression could be due to the
loss of the two G1-containing sites (a1 and a2). For
another reporter construct (pS3aPGDt15) in which
the deletion included two clustered high-affinity sites
(a10 and t15) and five weak binding sites (a8, a9,
a11, t14, and t16), the reduction in the number of
EGFP-expressing cells was somewhat lower (� 35%).
Similarly, when a fragment containing one high-affin-
ity site (a18) and eight additional ATTA core motifs
(a16, a17, and t20–t25) was deleted (pS3aPGDa18),the reduction in EGFP expression was � 45%. How-
ever, reporter gene expression was not significantly
reduced for one of the constructs (pS3aPGDa14) in
which a relatively strong binding site (a14) was
deleted together with two low-affinity sites (a13 and
a15). Notably, the effects on EGFP expression were
also weak or insignificant for the three constructs
(pS3aPGDa3, pS3aPGDa6, and pS3aPGDa12) in
which none of the high-affinity sites had been deleted.
Hence, we observed a correlation between the
presence of strong Six3a HD-binding sites, particu-
larly the three ATTA motifs with a flanking G1
nucleotide (a1, a2, and t15), and the ability to respond
to six3a overexpression in vivo.
Relative influence of ATTA core flanking
nucleotides on Six3a HD binding
To analyse the contribution of individual flanking
nucleotides and their importance relative to specific
positions within the ATTA core, we investigated how
single-nucleotide substitutions influenced the binding
affinity of a1. Modified a1 sites with single-nucleotide
mutations in nine positions (G1T2C3A4T5T6A7G8G9)
were compared for their ability to compete with
unmodified a1 in EMSAs (Fig. 4A). This in vitro anal-
ysis showed that substitution of G1 by T had moderate
effects on the binding affinity for the Six3a HD (G1T
in Fig. 4A). However, changes introduced at posi-
tions 2 (T2C) and 3 (C3G) affected the binding more
severely (Fig. 4A). Hence, these two 5¢-nucleotidesappeared to be essential for Six3a HD-binding affinity.
In fact, these substitutions resulted in less competition
than observed for individual mutations in three of the
positions within the ATTA core (A4C, T5G, and T6G).
Only a substitution at the fourth position (A7C) of this
core motif showed a similar deleterious effect on the
binding. In addition, we observed that mutation of the
3¢-flanking G8 nucleotide (G8A) had a moderate effect,
similar to the G1T substitution. By contrast, a change
of G9 (G9T) did not cause any reduction of the bind-
ing affinity, suggesting an insignificant role of this
3¢-flanking position. Overall, these results showed that
positions 2 and 3 immediately 5¢ of the ATTA core are
EGFP
pS3aPGΔa1
pS3aPGΔa3
pS3aPGΔa6
pS3aPGΔt15
pS3aPGΔa12
pS3aPGΔa14
pS3aPGΔa18
pS3aPG
0.5
–+
++
++
++
+
1.0 1.5 2.0 2.5
EGFP-expressing cellsSix3a
mR
NA
*
*
*Δ386 bp
Δ406 bp
Δ368 bp
Δ470 bp
Δ424 bp
Δ444 bp
Δ497 bp
Fig. 3. Deletion of particular fragments within the six3a promoter region affects reporter gene expression in embryos. Schematic represen-
tation of promoter–reporter deletion constructs and their corresponding activity in vivo. The pS3aPG vector (shown in Fig. 1A) and deletion
constructs derived from it were coinjected with six3a mRNA into one-cell embryos. EGFP-expressing cells from trypsinated embryos
(12 hpf) were detected by flow cytometry. The ratio between the EGFP expression from the reporter constructs coinjected with six3a
mRNA and from those without six3a mRNA was calculated. Standard deviations were calculated from three repeated experiments. Asterisks
indicate significant difference from the unmodified construct (pS3aPG) at P < 0.05. PS3aPGDa1, pS3aPGDt15 and pS3aPGDa18 showed
significantly less increase in EGFP expression when coinjected with six3a mRNA.
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1765
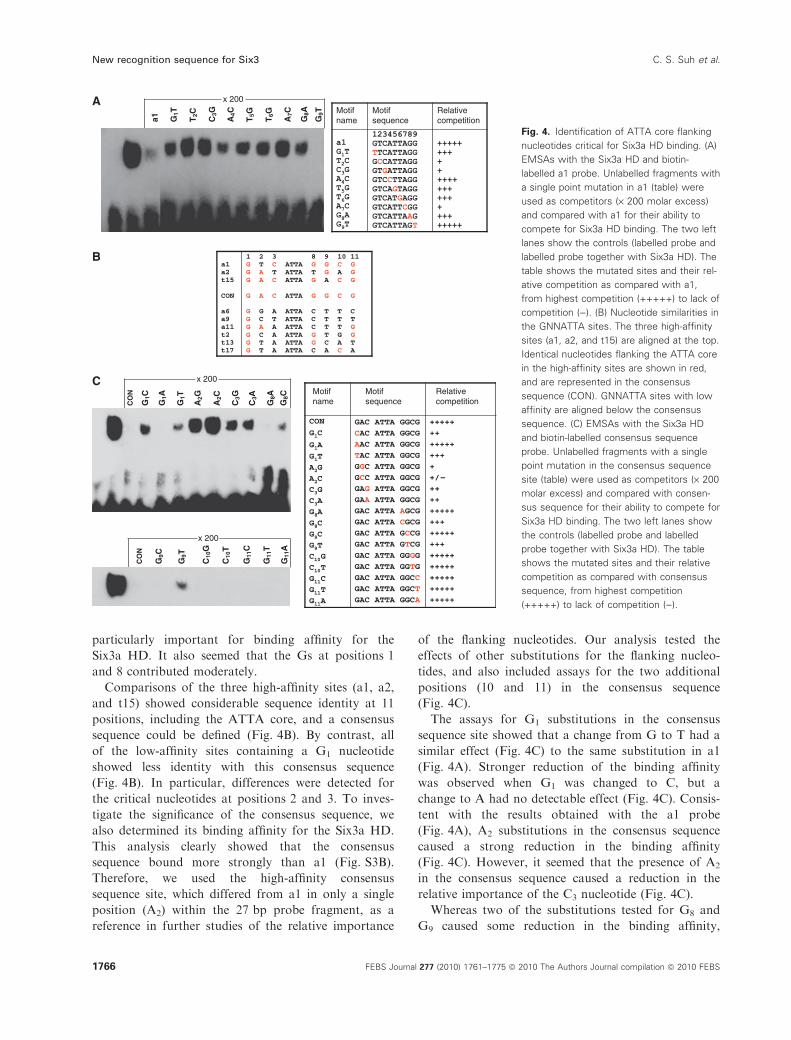
particularly important for binding affinity for the
Six3a HD. It also seemed that the Gs at positions 1
and 8 contributed moderately.
Comparisons of the three high-affinity sites (a1, a2,
and t15) showed considerable sequence identity at 11
positions, including the ATTA core, and a consensus
sequence could be defined (Fig. 4B). By contrast, all
of the low-affinity sites containing a G1 nucleotide
showed less identity with this consensus sequence
(Fig. 4B). In particular, differences were detected for
the critical nucleotides at positions 2 and 3. To inves-
tigate the significance of the consensus sequence, we
also determined its binding affinity for the Six3a HD.
This analysis clearly showed that the consensus
sequence bound more strongly than a1 (Fig. S3B).
Therefore, we used the high-affinity consensus
sequence site, which differed from a1 in only a single
position (A2) within the 27 bp probe fragment, as a
reference in further studies of the relative importance
of the flanking nucleotides. Our analysis tested the
effects of other substitutions for the flanking nucleo-
tides, and also included assays for the two additional
positions (10 and 11) in the consensus sequence
(Fig. 4C).
The assays for G1 substitutions in the consensus
sequence site showed that a change from G to T had a
similar effect (Fig. 4C) to the same substitution in a1
(Fig. 4A). Stronger reduction of the binding affinity
was observed when G1 was changed to C, but a
change to A had no detectable effect (Fig. 4C). Consis-
tent with the results obtained with the a1 probe
(Fig. 4A), A2 substitutions in the consensus sequence
caused a strong reduction in the binding affinity
(Fig. 4C). However, it seemed that the presence of A2
in the consensus sequence caused a reduction in the
relative importance of the C3 nucleotide (Fig. 4C).
Whereas two of the substitutions tested for G8 and
G9 caused some reduction in the binding affinity,
a1 G1T
T2C
C3G
A4C
T5G
T6G
A7C
G8A
G9T
x 200A
B
C
Relativecompetition
Motifsequence
Motifname
CO
N
x 200
CO
N
x 200
Relativecompetition
Motifsequence
MotifnameG
1A
G1T
G1C
C3G
C3A
A2G
A2C
G8A
G8C
G9C
G9T
C10
G
C10
T
G11
C
G11
T
G11
A
Fig. 4. Identification of ATTA core flanking
nucleotides critical for Six3a HD binding. (A)
EMSAs with the Six3a HD and biotin-
labelled a1 probe. Unlabelled fragments with
a single point mutation in a1 (table) were
used as competitors (· 200 molar excess)
and compared with a1 for their ability to
compete for Six3a HD binding. The two left
lanes show the controls (labelled probe and
labelled probe together with Six3a HD). The
table shows the mutated sites and their rel-
ative competition as compared with a1,
from highest competition (+++++) to lack of
competition ()). (B) Nucleotide similarities in
the GNNATTA sites. The three high-affinity
sites (a1, a2, and t15) are aligned at the top.
Identical nucleotides flanking the ATTA core
in the high-affinity sites are shown in red,
and are represented in the consensus
sequence (CON). GNNATTA sites with low
affinity are aligned below the consensus
sequence. (C) EMSAs with the Six3a HD
and biotin-labelled consensus sequence
probe. Unlabelled fragments with a single
point mutation in the consensus sequence
site (table) were used as competitors (· 200
molar excess) and compared with consen-
sus sequence for their ability to compete for
Six3a HD binding. The two left lanes show
the controls (labelled probe and labelled
probe together with Six3a HD). The table
shows the mutated sites and their relative
competition as compared with consensus
sequence, from highest competition
(+++++) to lack of competition ()).
New recognition sequence for Six3 C. S. Suh et al.
1766 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS
single mutations of the other 3¢-nucleotides in the
consensus sequence (C10 and G11) did not have detect-
able effects (Fig. 4C). These results suggested marginal
roles for each of the four 3¢-flanking positions within
the consensus sequence. Information obtained from
investigations of the effects of swapping flanking
regions with sequences from a low-affinity binding site
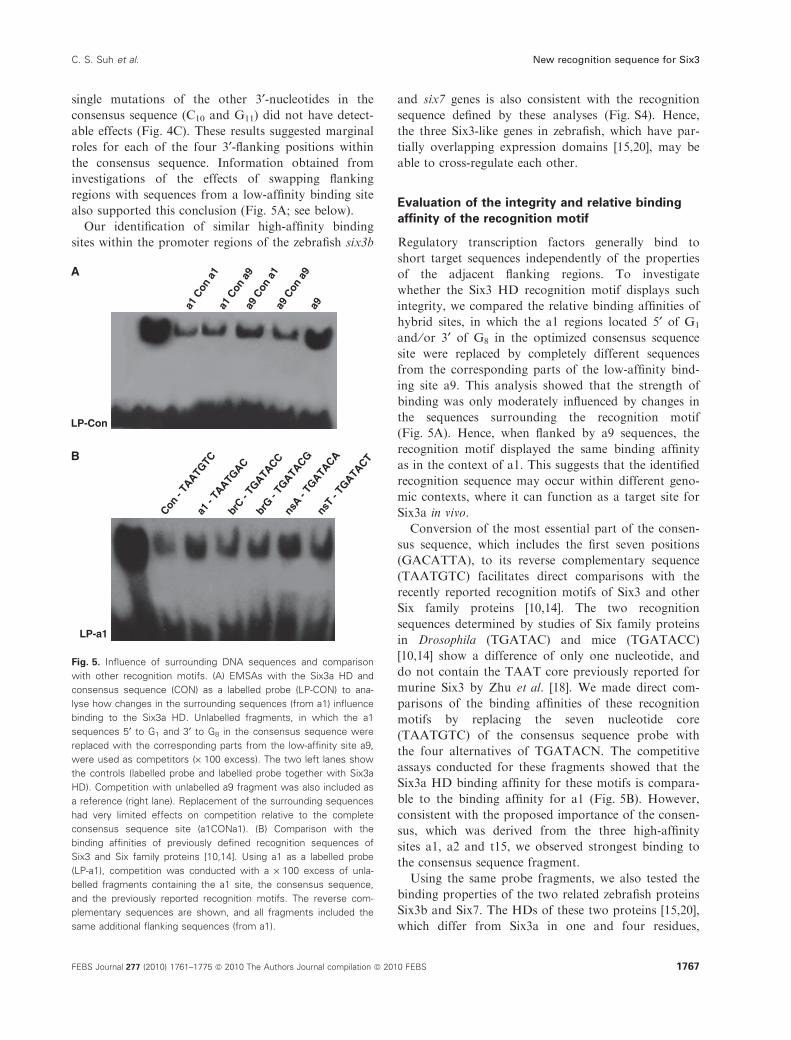
also supported this conclusion (Fig. 5A; see below).
Our identification of similar high-affinity binding
sites within the promoter regions of the zebrafish six3b
and six7 genes is also consistent with the recognition
sequence defined by these analyses (Fig. S4). Hence,
the three Six3-like genes in zebrafish, which have par-
tially overlapping expression domains [15,20], may be
able to cross-regulate each other.
Evaluation of the integrity and relative binding
affinity of the recognition motif
Regulatory transcription factors generally bind to
short target sequences independently of the properties
of the adjacent flanking regions. To investigate
whether the Six3 HD recognition motif displays such
integrity, we compared the relative binding affinities of
hybrid sites, in which the a1 regions located 5¢ of G1
and ⁄or 3¢ of G8 in the optimized consensus sequence
site were replaced by completely different sequences
from the corresponding parts of the low-affinity bind-
ing site a9. This analysis showed that the strength of
binding was only moderately influenced by changes in
the sequences surrounding the recognition motif
(Fig. 5A). Hence, when flanked by a9 sequences, the
recognition motif displayed the same binding affinity
as in the context of a1. This suggests that the identified
recognition sequence may occur within different geno-
mic contexts, where it can function as a target site for
Six3a in vivo.
Conversion of the most essential part of the consen-
sus sequence, which includes the first seven positions
(GACATTA), to its reverse complementary sequence
(TAATGTC) facilitates direct comparisons with the
recently reported recognition motifs of Six3 and other
Six family proteins [10,14]. The two recognition
sequences determined by studies of Six family proteins
in Drosophila (TGATAC) and mice (TGATACC)
[10,14] show a difference of only one nucleotide, and
do not contain the TAAT core previously reported for
murine Six3 by Zhu et al. [18]. We made direct com-
parisons of the binding affinities of these recognition
motifs by replacing the seven nucleotide core
(TAATGTC) of the consensus sequence probe with
the four alternatives of TGATACN. The competitive
assays conducted for these fragments showed that the
Six3a HD binding affinity for these motifs is compara-
ble to the binding affinity for a1 (Fig. 5B). However,
consistent with the proposed importance of the consen-
sus, which was derived from the three high-affinity
sites a1, a2 and t15, we observed strongest binding to
the consensus sequence fragment.
Using the same probe fragments, we also tested the
binding properties of the two related zebrafish proteins
Six3b and Six7. The HDs of these two proteins [15,20],
which differ from Six3a in one and four residues,
a1 C
on a
9
a1 C
on a
1
a9 C
on a
1 a9
Con
a9
a9
LP-Con
A
B
Con -
TAATG
TCa1
- TA
ATGAC
brG -
TGATA
CG
nsA -
TGATA
CA
nsT
- TG
ATACT
brC -
TGATA
CC
LP-a1
Fig. 5. Influence of surrounding DNA sequences and comparison
with other recognition motifs. (A) EMSAs with the Six3a HD and
consensus sequence (CON) as a labelled probe (LP-CON) to ana-
lyse how changes in the surrounding sequences (from a1) influence
binding to the Six3a HD. Unlabelled fragments, in which the a1
sequences 5¢ to G1 and 3¢ to G8 in the consensus sequence were
replaced with the corresponding parts from the low-affinity site a9,
were used as competitors (· 100 excess). The two left lanes show
the controls (labelled probe and labelled probe together with Six3a
HD). Competition with unlabelled a9 fragment was also included as
a reference (right lane). Replacement of the surrounding sequences
had very limited effects on competition relative to the complete
consensus sequence site (a1CONa1). (B) Comparison with the
binding affinities of previously defined recognition sequences of
Six3 and Six family proteins [10,14]. Using a1 as a labelled probe
(LP-a1), competition was conducted with a · 100 excess of unla-
belled fragments containing the a1 site, the consensus sequence,
and the previously reported recognition motifs. The reverse com-
plementary sequences are shown, and all fragments included the
same additional flanking sequences (from a1).
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1767
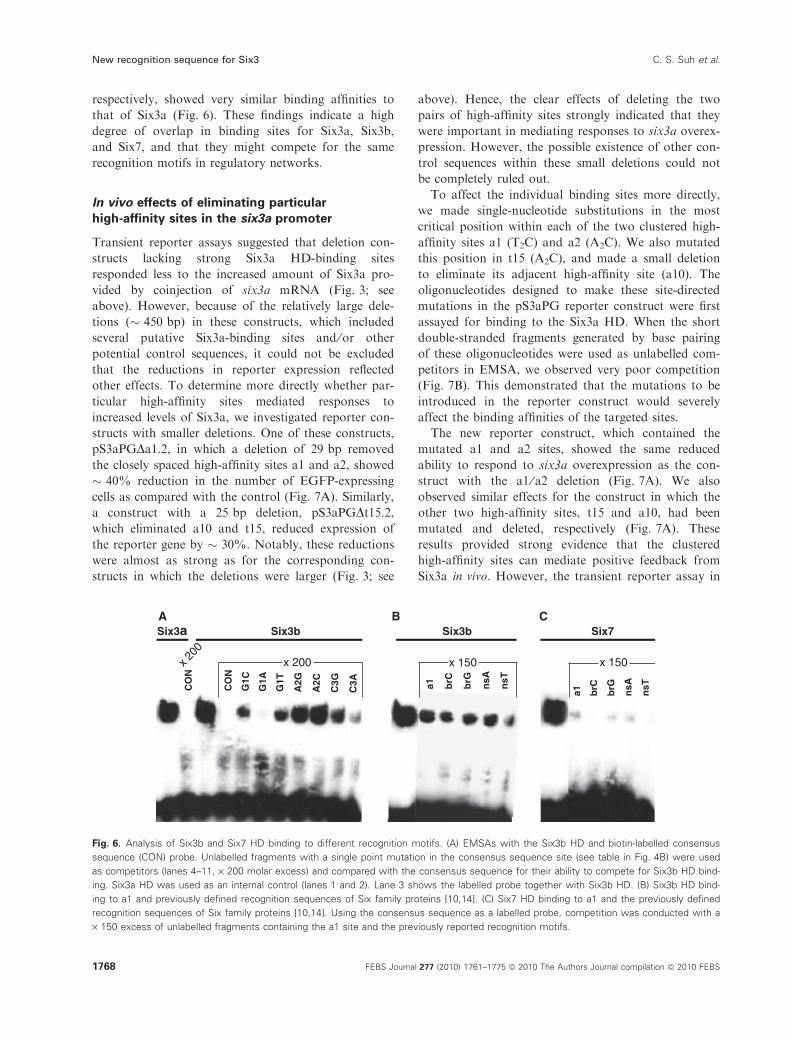
respectively, showed very similar binding affinities to
that of Six3a (Fig. 6). These findings indicate a high
degree of overlap in binding sites for Six3a, Six3b,
and Six7, and that they might compete for the same
recognition motifs in regulatory networks.
In vivo effects of eliminating particular
high-affinity sites in the six3a promoter
Transient reporter assays suggested that deletion con-
structs lacking strong Six3a HD-binding sites
responded less to the increased amount of Six3a pro-
vided by coinjection of six3a mRNA (Fig. 3; see
above). However, because of the relatively large dele-
tions (� 450 bp) in these constructs, which included
several putative Six3a-binding sites and ⁄or other
potential control sequences, it could not be excluded
that the reductions in reporter expression reflected
other effects. To determine more directly whether par-
ticular high-affinity sites mediated responses to
increased levels of Six3a, we investigated reporter con-
structs with smaller deletions. One of these constructs,
pS3aPGDa1.2, in which a deletion of 29 bp removed
the closely spaced high-affinity sites a1 and a2, showed
� 40% reduction in the number of EGFP-expressing
cells as compared with the control (Fig. 7A). Similarly,
a construct with a 25 bp deletion, pS3aPGDt15.2,which eliminated a10 and t15, reduced expression of
the reporter gene by � 30%. Notably, these reductions
were almost as strong as for the corresponding con-
structs in which the deletions were larger (Fig. 3; see
above). Hence, the clear effects of deleting the two
pairs of high-affinity sites strongly indicated that they
were important in mediating responses to six3a overex-
pression. However, the possible existence of other con-
trol sequences within these small deletions could not
be completely ruled out.
To affect the individual binding sites more directly,
we made single-nucleotide substitutions in the most
critical position within each of the two clustered high-
affinity sites a1 (T2C) and a2 (A2C). We also mutated
this position in t15 (A2C), and made a small deletion
to eliminate its adjacent high-affinity site (a10). The
oligonucleotides designed to make these site-directed
mutations in the pS3aPG reporter construct were first
assayed for binding to the Six3a HD. When the short
double-stranded fragments generated by base pairing
of these oligonucleotides were used as unlabelled com-
petitors in EMSA, we observed very poor competition
(Fig. 7B). This demonstrated that the mutations to be
introduced in the reporter construct would severely
affect the binding affinities of the targeted sites.
The new reporter construct, which contained the
mutated a1 and a2 sites, showed the same reduced
ability to respond to six3a overexpression as the con-
struct with the a1 ⁄ a2 deletion (Fig. 7A). We also
observed similar effects for the construct in which the
other two high-affinity sites, t15 and a10, had been
mutated and deleted, respectively (Fig. 7A). These
results provided strong evidence that the clustered
high-affinity sites can mediate positive feedback from
Six3a in vivo. However, the transient reporter assay in
CO
N
G1C
G1A
G1T
A2G
A2C
C3G
C3ACO
N
Six3aA B C
a1 brC
brG
nsA
nsT
Six3b Six3b
a1 brC
brG
nsA
nsT
x 200x 200
x 150x 150
Six7
Fig. 6. Analysis of Six3b and Six7 HD binding to different recognition motifs. (A) EMSAs with the Six3b HD and biotin-labelled consensus
sequence (CON) probe. Unlabelled fragments with a single point mutation in the consensus sequence site (see table in Fig. 4B) were used
as competitors (lanes 4–11, · 200 molar excess) and compared with the consensus sequence for their ability to compete for Six3b HD bind-
ing. Six3a HD was used as an internal control (lanes 1 and 2). Lane 3 shows the labelled probe together with Six3b HD. (B) Six3b HD bind-
ing to a1 and previously defined recognition sequences of Six family proteins [10,14]. (C) Six7 HD binding to a1 and the previously defined
recognition sequences of Six family proteins [10,14]. Using the consensus sequence as a labelled probe, competition was conducted with a
· 150 excess of unlabelled fragments containing the a1 site and the previously reported recognition motifs.
New recognition sequence for Six3 C. S. Suh et al.
1768 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS

injected embryos may not reflect the regulation of
six3a gene expression under normal conditions. To
gain further information about possible autoregulation
of the six3a gene during early embryonic development,
we investigated whether Six3a actually binds to some
of the high-affinity sites in the endogenous six3a
promoter (see below).
In vivo validation of Six3a-binding sites in the
six3a promoter region
The transient assays of reporter constructs, which were
conducted at the 12 hpf stage, did not provide any
information regarding the possible binding of Six3a to
the endogenous six3a promoter region in its natural
context. To facilitate such an investigation, which
required a specific antibody, we tagged Six3a with
EGFP and used an antibody against green fluorescent
protein (GFP) for ChIP assays on extracts from this
embryonic stage. Following injection of six3a–EGFP
mRNA into fertilized eggs (see Experimental proce-
dures), we detected high EGFP levels during embryo-
genesis (data not shown), and chromatin fragments
isolated from these embryos were immunoprecipitated
with the antibody against GFP. Using PCR primers
designed to amplify specific regions within the six3a
promoter (Fig. 8A), we investigated whether fragments
containing high-affinity sites were selectively amplified
(Fig. 8B). Consistent with the results obtained from
the transient reporter assays (Figs 3 and 7), a short
fragment (181 bp) containing the clustered a1 and a2
sites was amplified. In addition, we detected selective
amplification of a distal fragment, which included a
relatively strong binding site (a18) and five ATTA ⁄TAAT core motifs. This finding also correlates well
with results from transient reporter assays, where elim-
ination of a18 and eight adjacent core motifs by a
deletion caused a significant reduction in the response
to six3a overexpression (Fig. 3). Furthermore, cloning
and sequencing of the two amplified fragments con-
firmed their origins from the six3a promoter region
(data not shown).
Notably, amplification was not detected with the
primer pairs for regions containing only low-affinity
TAAT ⁄ATTA core motifs or lacking these motifs
completely. However, we also observed no amplification
0.5 1.0 1.5 2.0 2.5
–+
++
++
EGFP-expressing cells
Six3a
mR
NA
EGFP
Δ29 bppS3aPGΔΔΔΔa1.2
pS3aPG a1T2C, a2A2C Δ25 bppS3aPGΔΔΔΔt15.2
pS3aPGΔΔΔΔa10, t15A2C
a1/a
2
a10/
t15
Mu
t1
Mu
t2
x 200
pS3aPG
A
B
Fig. 7. Mutation of high-affinity sites within the six3a promoter region affects reporter gene expression in embryos. (A) Schematic represen-
tation of the in vivo activity of promoter–reporter constructs containing deletions (Da1 ⁄ a2 and Da10 ⁄ t15) or mutations of clustered high-affin-
ity sites. The plasmid constructs were coinjected with six3a mRNA into one-cell embryos. EGFP-expressing cells from trypsinated embryos
(12 hpf) were detected by flow cytometry. The ratio between the EGFP expression from the reporter construct coinjected with six3a mRNA
and from that without six3a mRNA was calculated. Standard deviations were calculated from three repeated experiments. Significant differ-
ence from the complete promoter–reporter construct (pS3aPG) was calculated at P < 0.05. (B) Double-stranded oligonucleotides with point
mutations at position 2 of the Six3a HD-binding sites were used as cold probes in competitive EMSA to confirm loss of binding affinity.
Mut1 represents a1T2C,a2A2C, and Mut2 represents Da10,t15A2C.
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1769

of the fragment containing the high-affinity sites a10
and t15, despite strong evidence from the reporter
assays (Figs 3 and 7). Possibly, an unfavourable state
of the chromatin within this particular region of the
endogenous six3a promoter may prevent Six3a from
binding at this developmental stage (12 hpf). Hence, it
remains a possibility that the two sites may have
autoregulatory functions at later stages of development.
Discussion
In this study, we show that binding of zebrafish Six3a
to high-affinity sites in the promoter region of its own
gene may contribute to autoregulation. The Six3a HD,
which is identical to the corresponding DNA-binding
domain of orthologous Six3 proteins in other verte-
brates, displayed differential binding affinities for vari-
ous sites containing a TAAT core motif, and this
defined a specific recognition sequence (TAATGTC).
When we compared this with a previously reported
Six3-binding sequence (TGATAC), we observed simi-
lar binding affinity, suggesting that the Six3 HD has
several binding modes with distinct DNA-binding
specificities.
Using several experimental approaches, previous
investigations of the DNA-binding preferences of the
three Six protein subfamilies (Six1 ⁄ 2, Six3 ⁄ 6, and
Six4 ⁄ 5) have found a number of different target
sequences. Whereas analyses of native binding sites in
putative target genes identified a variety of sequences
[17,18,30,36–38], two independent studies based on dif-
ferent in vitro selection procedures established a com-
mon recognition sequence of the three subfamilies
[10,14,39]. These partially conflicting results may reflect
the fact that the HDs belonging to different subfami-
lies have almost identical recognition helices but diver-
gent N-terminal arms [16].
The HDs in the Six3 ⁄ 6 subfamily proteins show a
higher degree of divergence relative to the other two
subfamilies [16]. Consistent with this sequence diver-
gence, early studies of the Six3 ⁄6 proteins indicated
clear differences in DNA-binding specificity relative to
the Six1 ⁄ 2 and Six4 ⁄ 5 family members [1]. Hence,
murine Six3 did not bind to the ARE regulatory
element of the Na+ ⁄K+-ATPase a1-subunit gene,
GGTGTCAGGTTGC, which showed specific binding
to Six4 and other members of the Six1 ⁄ 2 and Six4 ⁄5subfamilies [17]. In further investigations of the DNA-
binding properties of murine Six3, in vitro binding site
selection demonstrated high-affinity binding to
sequences containing the TAAT core motif, and clus-
ters of these tetranucleotide sites were found in regula-
tory elements within the promoter regions of both
Six3 and Wnt1 [18,37]. Although these analyses dem-
onstrated an involvement of the clustered TAAT
motifs in mediating negative regulation by Six3,
–357
9 –3
290
–289
3 –2
704
–209
3 –1
872
–118
3 –9
80
–656
–47
7
–25
3 –7
2
six3a promoter region (pS3aP)
IP (+Ab, injected)
Input control
IP (+Ab, uninjected)
300 bp200 bp
300 bp200 bp
300 bp200 bp
18 17
A
B
16 15 14 13 12 1110 9 8 7 6 5 4 3 2
t2
1**
t13t15t17*
Fig. 8. Detection of Six3a binding to high-affinity sites in the six3a promoter in vivo. (A) Schematic representation of the promoter region of
six3a (see legend to Fig. 2A) and the fragments that were selected for PCR amplification. Filled boxes indicate the regions that were ampli-
fied, and empty boxes represent regions that showed no amplification. (B) ChIP PCR assay on 12 hpf embryos. The upper panel represents
PCR amplification of different regions (indicated by the boxes) within the six3a promoter following immunoprecipitation (IP) of DNA bound to
Six3a–EGFP fusion protein (Experimental procedures). The middle panel represents PCR amplification from uninjected zebrafish embryos
immunoprecitated with GFP antibody (Ab). The lower panel represents PCR amplification from total embryonic DNA.
New recognition sequence for Six3 C. S. Suh et al.
1770 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS

additional features specifically favouring binding to
Six3 proteins were not identified.
Our examination of the interaction of zebrafish
Six3a with 43 TAAT-containing sequences, distributed
within the promoter region of its own gene, identified
three high-affinity sites that defined the recognition
sequence TAATGTC. Notably, this sequence prefer-
ence includes three adjacent nucleotides 3¢ to the
TAAT core, corresponding to the positions generally
involved in discriminating between different HDs.
The importance of this novel recognition motif was
also supported by comparisons of the relative binding
affinities of mutated sites and by in vivo assays of
reporter constructs. In addition, a direct comparison
with the recently reported generic recognition
sequence of Six family HDs, TGATAC [10,14],
showed the same strong binding affinity. Further-
more, major changes of the surrounding sequences
did not affect the binding affinity of the recognition
sequence, indicating that it is generally independent
of the sequence context. Importantly, the TAATGTC
motif is also distinct from all the different DNA-
binding specificities that were recently described in
surveys including the majority of the Drosophila and
mouse HDs [10,14]. This unique sequence preference
is likely to facilitate binding of Six3 monomers to
regulatory sites in their target genes without requiring
cooperating factors to achieve sufficient sequence
specificity.
It remains to understand how it is possible for the
Six3 HD to bind strongly to two distinct types of
recognition sequences. A complete elucidation of this
issue can only be achieved by analysing the crystal
and ⁄or solution structures of the different complexes
formed when the Six3 HD binds to each motif.
Although such structures have not been determined for
any of the Six family proteins, some clues may be
obtained by considering the divergent features of their
HDs. In contrast to the majority of HDs, which recog-
nize various sequences containing the TAAT core, the
N-terminal arms of Six family HDs lack several resi-
dues, such as Arg5, known to be involved in specifying
the first two positions in this tetranucleotide motif
[1,15]. Another characteristic of the Six family HDs,
which they share with several other atypical HDs, is
the presence of Arg55 in the recognition helix that
may specify a G in position 2 of the core motif [10].
Consistent with this, the HDs of all Six family proteins
in mouse and Drosophila were shown to recognize
sequences containing the core motif TGAT [10,15].
Although the sequence divergence in the specifying
residues of the Six family HDs is not consistent with
our identification of the TAATGTC motif, it is impor-
tant to consider that intramolecular interactions
involving residues outside of the protein–DNA binding
interface can also influence DNA recognition [10,40].
In particular, variations in the position of the N-termi-
nal arm have been shown to influence its recognition
properties [6,10]. Investigations of the binding proper-
ties of the Ultrabithorax protein have also demon-
strated that the N-terminal arm can strongly affect the
topological binding mode of the HD and influence the
nucleotide contacts made by the recognition helix [6].
Notably, these different binding modes seemed to
depend on the binding site sequence, and the binding
affinities were similar [6]. Currently, it is not known
whether such transitions between different binding
modes are common among HDs. However, multiple
binding modes have been proposed as an explanation
for the identification of several recognition sequences
for some of the mouse HDs [14].
Consistent with the previous report of autoregula-
tion of the murine Six3 gene [18], we found that the
high-affinity sites recognized by the Six3a HD in vitro
were clustered within the promoter region of its own
gene. Our investigation of the functional significance
of these clustered high-affinity sites included in vivo
assays of various promoter–reporter constructs in
zebrafish embryos and ChIP. In transient transgenic
assays, which provide more natural conditions than
cell culture transfections, the 3.6 kb promoter fragment
of the six3a gene showed a clear response to coinjec-
tion of six3a mRNA by increasing expression of the
EGFP reporter by more than 100%. The strong effect
of overexpressing Six3a was possibly due to direct
binding to high-affinity sites within the promoter
region. Consistent with this interpretation, we observed
reduced responses to Six3a overexpression when
regions containing the clustered high-affinity sites were
deleted from the reporter constructs. To rule out the
possible involvement of other control sequences, we
assayed the effects of removing the high-affinity sites
by introducing single-nucleotide substitutions that
severely affected their ability to bind the Six3a HD.
Remarkably, reporter constructs with these minimal
changes showed almost the same reduced ability to
respond to higher levels of Six3a in injected embryos
as the constructs with larger deletions.
Our results clearly demonstrated that the clustered
high-affinity sites mediate positive autoregulation of
the zebrafish six3a gene under experimental conditions
in vivo. In contrast, autorepression of the murine Six3
gene has been detected in cotransfection assays in
cultured cells [18]. The opposite effects observed for
the zebrafish six3a and murine Six3 genes may reflect
differences in the experimental conditions and ⁄or
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1771

involvement of different cofactors. The autorepression
of the murine Six3 gene was shown to depend on
interaction with members of the Groucho-related
family of corepressors [18]. Additional studies in
medaka fish (O. latipes) have provided evidence that
Six3 ⁄ 6 family proteins bind differentially to these core-
pressors and the related cofactor Aes, which may lead
to transcriptional activation [41]. Similar mechanisms
of derepression may have occurred when we
experimentally overexpressed Six3a in our transient
reporter assays of the six3a promoter in zebrafish
embryos. However, irrespective of the cofactors that
may have contributed, the activation was dependent
on the Six3a HD-binding sites shown to bind strongly
in vitro.
Other members of the Six3 ⁄ 6 family, which have simi-
lar or identical DNA-binding specificity to that of Six3a,
have partially overlapping expression at early stages of
eye and forebrain development [15,20]. Hence, tran-
scriptional control of these genes is likely to involve
both autoregulation and cross-regulation. Differences in
their interactions with Groucho-related cofactors may
be important for this regulation and may contribute to
the control of eye and forebrain development. This pos-
sibility is supported by previous studies in medaka fish
that showed different effects on eye size when the Six3
and Six6 activities were modulated by Aes and the
Groucho family corepressor Tle1 [41].
The chromatin structure of the injected reporter con-
struct may differ considerably from the native state of
the endogenous six3a gene, which could also influence
the responses to increased levels of Six3a. To obtain
more direct evidence that Six3a binds to the clustered
high-affinity sites in a native context, we investigated
the chromatin of the endogenous six3a promoter
region in embryos. Our ChIP analysis detected Six3a
binding specifically to a small fragment (� 180 bp)
containing the two strong binding sites that were
shown to mediate autoregulation of the reporter con-
struct. This finding, which is also consistent with the
importance of the Six3 HD recognition sequence sug-
gested by the in vitro studies, provides a basis for
future identification of Six3 target genes.
Experimental procedures
Maintenance of fish stock and in situ
hybridization
Zebrafish were maintained and bred at 28 �C. Whole
mount in situ hybridization on zebrafish embryos was
performed with digoxigenin-labelled probes as described
previously [15].
Generation of Tg(six3a:EGFP) line
One-cell-stage embryos were coinjected through the cho-
rion with 1 nL of a solution containing plasmid DNA
()3.6 kb six3a promoter in Tol2 vector) at a concentration
of 10 ngÆlL)1, along with Tol2 transposase mRNA at
15 ngÆlL)1, KCl at 200 mm, and phenol red at 0.05%, as
described previously [42]. Injected F0 embryos were raised
and crossed with the wild-type TAB strain [43]. F1 prog-
eny were screened for EGFP expression at 12 hpf.
Embryos and larvae exhibiting tissue-restricted EGFP fluo-
rescence were selected and raised to obtain stable trans-
genic lines.
Cloning, expression and purification of
glutathione-S-transferase (GST)–Six3aHD
fusion protein
The HD coding sequence of six3a was amplified by PCR,
using 5¢-TCAGGTCGGATCCATGGTTTTCAGA-3¢ as
forward primer and 5¢-CTGTGTGGAATTCATACGTCG
CATTC-3¢ as reverse primer, cloned in-frame into the
BamHI and EcoRI site of the pGEX-2T expression vector
(Pharmacia Biotech Inc., Uppsala, Sweden), and transformed
into Escherichia coli [BL21 (DE3) pLysS Competent Cells]
cells. E. coli cells transformed with the pGEX-2T–Six3-
aHD vector were grown in 250 mL of LB medium prior
to induction with 2 mm isopropyl thio-b-d-galactoside.The bacteria were pelleted after a 3 h induction period,
and whole cell extracts were prepared by using a French
press. The GST–Six3aHD fusion protein was purified by
batch adsorption onto glutathione–Sepharose 4B beads
(Pharmacia Biotech Inc.). After several washes of the
beads with NaCl ⁄Pi with Tween-20, the GST fusion pro-
teins were eluted with 50 mm reduced glutathione solution
and subsequently used in EMSAs.
EMSAs
EMSAs were performed with 5¢-biotin-labelled oligonucle-
otides in 20 lL reactions containing 80 mm Tris ⁄HCl,
240 mm KCl, 2 mm dithiothreitol, 0.2% NP40, 10%
Ficoll, and 50 ng of poly(dI.dC), and incubated with
50 ng of purified GST–Six3aHD fusion protein. Each
27 bp DNA fragment contained the core ATTA motif
flanked by 13 nucleotides at the 5¢-end and 10 nucleotides
at the 3¢-end. For competition reactions, a · 200 molar
excess of unlabelled double-stranded oligonucleotide was
added to the binding reaction [40]. After incubation at
room temperature for 20 min, protein–DNA complexes
were separated on a 6% polyacrylamide gel in 0.5 · TBE
buffer. A chemiluminescent nucleic acid detection kit
(Pierce, Rockford, IL, USA) was used for the detection of
protein–DNA interactions, according to the manufacturer’s
instructions.
New recognition sequence for Six3 C. S. Suh et al.
1772 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS

Generation of reporter constructs for transient
analysis
A 3.6 kb fragment spanning positions )3616 to +19 of the
cis-regulatory region of the six3a gene was amplified by
PCR, and cloned into the pT2AL200R150G vector in front
of EGFP [44], generating the construct pS3aPG. Deletions
in the 3.6 kb six3a promoter region were performed by
PCR [45]. Primers were designed to add a noncomplemen-
tary 5¢-sequence consisting of the restriction enzyme
sequence XmaI. The following deletion constructs were
made: pS3aPG, pS3aPGDa1,a2,t2 (or pS3aPGDa1),pS3aPGDa3–a5 (or pS3aPGDa3), pS3aPGDa6,a7,t13(or pS3aPGDa6), pS3aPGDa8–a11,t15 (or pS3aPGDt15),pS3aPGDa12,t17 (or pS3aPGDa12), pS3aPGDa13–a15(or pS3aPGDa14), pS3aPGDa16–a18 (or pS3aPGDa18),pS3aPGDa1.2, and pS3aPGDat15.2. Following PCR ampli-
fication of the whole plasmid except the region to be
deleted, the PCR product was digested with XmaI and
DpnI (to eliminate the template DNA). The plasmid was
self-ligated and transformed into E. coli cells. Specific
nucleotide mutations on pS3aPG were generated using the
quick-change site-directed mutagenesis kit (Stratagene, La
Jolla, CA, USA), according to the manufacturer’s instruc-
tions, to make pS3aPGa1T2C,a2A2C and pS3aPGDa10,t15A2C. All deletion and mutation constructs were verified
by DNA sequencing.
Generation of mRNA
The Six3a coding sequence was amplified from zebrafish
cDNA (10–12 hpf) and cloned into the pEGFP-N1 vector
(Clontech, Mountain View, CA, USA). PstI and XbaI were
used to cut out the six3a–EGFP fusion and subclone it
into the expression vector pCS2+ [46]. For six3a mRNA
transcription, the six3a coding sequence was cloned directly
into the pCS2+ vector. The six3a–GFP and six3a mRNAs
were generated with the mMessage mMachine kit (Ambion,
Austin, TX, USA). All RNA products were analysed by
agarose gel electrophoresis and stored at )80 �C until use.
Detection of promoter activity by flow cytometry
Zebrafish embryos at the one-cell stage were coinjected with
25–30 pg of reporter constructs (as described above) and
100 pg of six3a mRNA per embryo. Embryos were examined
for EGFP expression at 12 hpf. One hundred EGFP-positive
embryos at 10–12 hpf were selected and washed twice in
NaCl ⁄Pi. Preparation of embryos for cell cytometry was per-
formed as previously described [47]. Single-cell suspensions
from the embryos were then analysed using a FACSCalibur
flow cytometer equipped with cellquest from BD Bio-
sciences (San Jose, CA, USA). During flow cytometric analy-
sis, the cell suspensions were kept on ice. The time between
dissociation of cells and flow cytometry was minimized to
2 h. Using forward and side scatter, yolk cells and debris
were eliminated in the analysis. EGFP fluorescence was
detected using a 530 ⁄ 30 nm bandpass filter in the FL1 chan-
nel. For each sample, information on 25 000 events was
acquired. The calculation of significant differences between
mean values was determined using Student’s t-test. Each
value is the mean difference ± standard deviation from
three experiments. The level accepted for statistical signifi-
cance in all cases was P < 0.05.
ChIP PCR
Zebrafish embryos at the one-cell stage were injected with
six3a–EGFP mRNA in the yolk at 100 pg per embryo. The
embryos were dechorionated at 12 hpf. ChIP was performed
as described in [48], using mouse antibody against GFP from
Invitrogen (Carlsbad, CA, USA). Briefly, 12 hpf embryos
were crosslinked with formaldehyde, and chromatin was iso-
lated. The isolated chromatin was sonicated to an average
size of about 300 bp. Incubation with normal goat IgG
precleared the chromatin. Protein G magnetic beads were
incubated with antibody against GFP at 4 �C overnight.
Immunoprecipitation reactions were performed in duplicate
by incubating 10 lg of antibody against GFP with the chro-
matin overnight at 4 �C. Parallel controls with uninjected
zebrafish embryos were incubated with antibody against
GFP as a negative control. The immunoprecipitated chro-
matin complexes were washed several times and eluted with
50 mm Tris (pH 8). In each ChIP experiment, a portion of
the chromatin solution corresponding to 1% of that used in
the ChIP reaction was used as input DNA control. The
immunoprecipitated and uninjected control immunoprecipi-
tated samples were purified by phenol ⁄ chloroform extraction
and ethanol precipitation after the protein–DNA crosslinks
had been reversed by incubation at 65 �C. Immunoprecipi-
tated DNA and input DNA were used as templates for PCR
amplification with the five primer pairs (Table S1).
Acknowledgements
We thank H. Savolainen, R. Aanesen and G. Merkin
for their expert technical help in our zebrafish facility.
This study was funded by the Research Council of
Norway (Grant 174979 ⁄ I30), and the Faculty of Mathe-
matics and Natural Sciences at the University of Bergen.
References
1 Kawakami K, Sato S, Ozaki H & Ikeda K (2000) Six
family genes: structure and function as transcription
factors and their roles in development. Bioessays 22,
616–626.
2 Christensen KL, Patrick AN, McCoy EL & Ford HL
(2008) The six family of homeobox genes in
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1773

development and cancer. Adv Cancer Res 101, 93–
126.
3 Qian YQ, Billeter M, Otting G, Muller M, Gehring WJ
& Wuthrich K (1989) The structure of the Antennape-
dia homeodomain determined by NMR spectroscopy in
solution: comparison with prokaryotic repressors. Cell
59, 573–580.
4 Kissinger CR, Liu BS, Martin-Blanco E, Kornberg TB
& Pabo CO (1990) Crystal structure of an engrailed
homeodomain–DNA complex at 2.8 A resolution: a
framework for understanding homeodomain–DNA
interactions. Cell 63, 579–590.
5 Gehring WJ, Qian YQ, Billeter M, Furukubo-Tokuna-
ga K, Schier AF, Resendez-Perez D, Affolter M, Otting
G & Wuthrich K (1994) Homeodomain–DNA recogni-
tion. Cell 78, 211–223.
6 Frazee RW, Taylor JA & Tullius TD (2002) Inter-
change of DNA-binding modes in the deformed and
ultrabithorax homeodomains: a structural role for the
N-terminal arm. J Mol Biol 323, 665–683.
7 Wolberger C, Vershon AK, Liu B, Johnson AD & Pabo
CO (1991) Crystal structure of a MAT alpha 2 homeod-
omain–operator complex suggests a general model for
homeodomain–DNA interactions. Cell 67, 517–528.
8 Billeter M, Guntert P, Luginbuhl P & Wuthrich K
(1996) Hydration and DNA recognition by homeodo-
mains. Cell 85, 1057–1065.
9 Banerjee-Basu S & Baxevanis AD (2001) Molecular
evolution of the homeodomain family of transcription
factors. Nucleic Acids Res 29, 3258–3269.
10 Noyes MB, Christensen RG, Wakabayashi A, Stormo
GD, Brodsky MH & Wolfe SA (2008) Analysis of home-
odomain specificities allows the family-wide prediction of
preferred recognition sites. Cell 133, 1277–1289.
11 Hanes SD & Brent R (1989) DNA specificity of the
bicoid activator protein is determined by homeodomain
recognition helix residue 9. Cell 57, 1275–1283.
12 Treisman J, Gonczy P, Vashishtha M, Harris E &
Desplan C (1989) A single amino acid can determine
the DNA binding specificity of homeodomain proteins.
Cell 59, 553–562.
13 Tucker-Kellogg L, Rould MA, Chambers KA, Ades
SE, Sauer RT & Pabo CO (1997) Engrailed (Gln50–
>Lys) homeodomain–DNA complex at 1.9 A resolu-
tion: structural basis for enhanced affinity and altered
specificity. Structure 5, 1047–1054.
14 Berger MF, Badis G, Gehrke AR, Talukder S, Philippa-
kis AA, Pena-Castillo L, Alleyne TM, Mnaimneh S,
Botvinnik OB, Chan ET et al. (2008) Variation in
homeodomain DNA binding revealed by high-resolution
analysis of sequence preferences. Cell 133, 1266–1276.
15 Seo HC, Drivenes O, Ellingsen S & Fjose A (1998a)
Expression of two zebrafish homologues of the murine
Six3 gene demarcates the initial eye primordia. Mech
Dev 73, 45–57.
16 Seo HC, Curtiss J, Mlodzik M & Fjose A (1999) Six
class homeobox genes in Drosophila belong to three
distinct families and are involved in head development.
Mech Dev 83, 127–139.
17 Kawakami K, Ohto H, Ikeda K & Roeder RG (1996)
Structure, function and expression of a murine homeo-
box protein AREC3, a homologue of Drosophila sine
oculis gene product, and implication in development.
Nucleic Acids Res 24, 303–310.
18 Zhu CC, Dyer MA, Uchikawa M, Kondoh H, Lagutin
OV & Oliver G (2002) Six3-mediated auto repression
and eye development requires its interaction with mem-
bers of the Groucho-related family of co-repressors.
Development 129, 2835–2849.
19 Jozefowicz C, McClintock J & Prince V (2003)
The fates of zebrafish Hox gene duplicates. J Struct
Funct Genomics 3, 185–194.
20 Seo HC, Drivenes O, Ellingsen S & Fjose A (1998b)
Transient expression of a novel Six3-related zebrafish
gene during gastrulation and eye formation. Gene 216,
39–46.
21 Oliver G, Mailhos A, Wehr R, Copeland NG, Jenkins
NA & Gruss P (1995) Six3, a murine homologue of the
sine oculis gene, demarcates the most anterior border of
the developing neural plate and is expressed during eye
development. Development 121, 4045–4055.
22 Kobayashi M, Toyama R, Takeda H, Dawid IB &
Kawakami K (1998) Overexpression of the forebrain-
specific homeobox gene six3 induces rostral forebrain
enlargement in zebrafish. Development 125, 2973–2982.
23 Loosli F, Winkler S & Wittbrodt J (1999) Six3 overex-
pression initiates the formation of ectopic retina. Genes
Dev 13, 649–654.
24 Wallis DE, Roessler E, Hehr U, Nanni L, Wiltshire T,
Richieri-Costa A, Gillessen-Kaesbach G, Zackai EH,
Rommens J & Muenke M (1999) Mutations in the
homeodomain of the human SIX3 gene cause holopros-
encephaly. Nat Genet 22, 196–198.
25 Zhou X, Hollemann T, Pieler T & Gruss P (2000) Clon-
ing and expression of xSix3, the Xenopus homologue of
murine Six3. Mech Dev 91, 327–330.
26 Del Bene F, Tessmar-Raible K & Wittbrodt J (2004)
Direct interaction of geminin and Six3 in eye develop-
ment. Nature 427, 745–749.
27 Gestri G, Carl M, Appolloni I, Wilson SW, Barsacchi
G & Andreazzoli M (2005) Six3 functions in anterior
neural plate specification by promoting cell proliferation
and inhibiting Bmp4 expression. Development 132,
2401–2413.
28 Ando H, Kobayashi M, Tsubokawa T, Uyemura K,
Furuta T & Okamoto H (2005) Lhx2 mediates the
activity of Six3 in zebrafish forebrain growth. Dev Biol
287, 456–468.
29 Inbal A, Kim SH, Shin J & Solnica-Krezel L
(2007) Six3 represses nodal activity to establish
New recognition sequence for Six3 C. S. Suh et al.
1774 FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS

early brain asymmetry in zebrafish. Neuron 55, 407–
415.
30 Jeong Y, Leskow FC, El-Jaick K, Roessler E, Muenke
M, Yocum A, Dubourg C, Li X, Geng X, Oliver G et al.
(2008) Regulation of a remote Shh forebrain enhancer by
the Six3 homeoprotein. Nat Genet 40, 1348–1353.
31 Kobayashi M, Nishikawa K, Suzuki T & Yamamoto M
(2001) The homeobox protein Six3 interacts with the
Groucho corepressor and acts as a transcriptional
repressor in eye and forebrain formation. Dev Biol 232,
315–326.
32 Drivenes O, Seo HC & Fjose A (2000) Characterisation
of the promoter region of the zebrafish six7 gene.
Biochim Biophys Acta 1491, 240–247.
33 Wargelius A, Seo HC, Austbø L & Fjose A (2003)
Retinal expression of zebrafish six3.1 and its regulation
by Pax6. Biochem Biophys Res Commun 309, 475–481.
34 Conte I & Bovolenta P (2007) Comprehensive charac-
terization of the cis-regulatory code responsible for the
spatio-temporal expression of olSix3.2 in the developing
medaka forebrain. Genome Biol 8, R137.
35 Westefield M (2000) The Zebrafish Book. A Guide for
the Laboratory Use of Zebrafish (Danio rerio), 4th edn.
University of Oregon.
36 Lengler J & Graw J (2001) Regulation of the human
SIX3 gene promoter. Biochem Biophys Res Commun
287, 372–376.
37 Spitz F, Demignon J, Porteu A, Kahn A, Concordet
JP, Daegelen D & Maire P (1998) Expression of myoge-
nin during embryogenesis is controlled by Six ⁄ sineoculis homeoproteins through a conserved MEF3
binding site. Proc Natl Acad Sci USA 95, 14220–14225.
38 Lagutin OV, Zhu CC, Kobayashi D, Topczewski J,
Shimamura K, Puelles L, Russell HR, McKinnon PJ,
Solnica-Krezel L & Oliver G (2003) Six3 repression of
Wnt signaling in the anterior neuroectoderm is essential
for vertebrate forebrain development. Genes Dev 17,
368–379.
39 Affolter M, Slattery M & Mann RS (2008) A lexicon for
homeodomain–DNA recognition. Cell 133, 1133–1135.
40 Pauli T, Seimiya M, Blanco J & Gehring WJ (2005)
Identification of functional sine oculis motifs in the
autoregulatory element of its own gene, in the eyeless
enhancer and in the signalling gene hedgehog. Develop-
ment 132, 2771–2782.
41 Lopez-Rıos J, Tessmar K, Loosli F, Wittbrodt J &
Bovolenta P (2003) Six3 and Six6 activity is modulated
by members of the groucho family. Development 130,
185–195.
42 Kawakami K & Shima A (1999) Identification of the
Tol2 transposase of the medaka fish Oryzias latipes that
catalyzes excision of a nonautonomous Tol2 element in
zebrafish Danio rerio. Gene 240, 239–244.
43 Amsterdam A, Nissen RM, Sun Z, Swindell EC,
Farrington S & Hopkins N (2004) Identification of 315
genes essential for early zebrafish development. Proc
Natl Acad Sci USA 101, 12792–12797.
44 Urasaki A, Morvan G & Kawakami K (2006)
Functional dissection of the Tol2 transposable element
identified the minimal cis-sequence and a highly
repetitive sequence in the subterminal region essential
for transposition. Genetics 174, 639–649.
45 Perez-Pinera P, Menendez-Gonzalez M & Vega JA
(2006) Deletion of DNA sequences of using a
polymerase chain reaction based approach. Electron J
Biotechnol 9, 604–609.
46 Kawakami K, Takeda H, Kawakami N, Kobayashi M,
Matsuda N & Mishina M (2004) A transposon medi-
ated gene trap approach identifies developmentally reg-
ulated genes in zebrafish. Dev Cell 7, 133–144.
47 Chan PK & Cheng SH (2002) Cadmium-induced ectopic
apoptosis in zebrafish embryos. Arch Toxicol 77, 69–79.
48 Wardle FC, Odom DT, Bell GW, Yuan B, Danford
TW, Wiellette EL, Herbolsheimer E, Sive HL, Young
RA & Smith JC (2006) Zebrafish promoter microarrays
identify actively transcribed embryonic genes. Genome
Biol 7, R71, 1–14.
Supporting information
The following supplementary material is available
online:
Fig. S1. Overview of the DNA sequence of the six3a
promoter region showing the positions of sites contain-
ing ATTA (and TAAT) core sequences.
Fig. S2. Sites within the six3a promoter containing
oppositely oriented TAAT cores that showed weak
binding as compared with the a1 site in competition
EMSAs.
Fig. S3. Comparison of relative binding affinities of
the consensus sequence (CON) and the strongest bind-
ing sites located within the six3a promoter region.
Fig. S4. Comparison of high-affinity GNNATTA sites
located within the promoter regions of the zebrafish
genes six3a, six3b and six7 indicates a common recog-
nition motif.
Fig. S5. Autoregulatory activity of the six3a promoter
assayed in zebrafish embryos.
Table S1. Primer sequences used in ChIP assay.
This supplementary material can be found in the
online version of this article.
Please note: As a service to our authors and readers,
this journal provides supporting information supplied
by the authors. Such materials are peer-reviewed and
may be re-organized for online delivery, but are not
copy-edited or typeset. Technical support issues arising
from supporting information (other than missing files)
should be addressed to the authors.
C. S. Suh et al. New recognition sequence for Six3
FEBS Journal 277 (2010) 1761–1775 ª 2010 The Authors Journal compilation ª 2010 FEBS 1775
Recommended

















