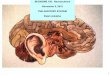
1. Auditory Physiology : Inner EarDr. Diptiman Baliarsingh1st
Year PG, Dept. of ENT,Hi-Tech Medical College &
Hospital,Bhubaneswar
2. Introduction Snail-shaped osseous structure Coiled 2 &
2/3 turns around a central axis Modiolus With in the bony cochlea
(osseous labyrinth) liesthe membranous labyrinth, consisting of
Central scala media cochlear duct Superiorly, Scala vestibuli
separated by Reissnersmembrane Inferiorly, Scala tympani separated
by basilarmembrane The connection of scala vestibuli with the
middleear occurs at Oval window, which is attached to
3. Cross-section of Cochlea
4. The Round window links the scala tympani tothe middle ear
& is covered by Round windowmembrane The scala vestibuli &
scala tympani merge at theapex of cochlea Helicotrema, & scala
mediaends blindly S.V. & S.T. are filled with perilymph,
extracellularfluid with High Na+ & Low K+ S.M. is filled with
endolymph, composed of HighK+ & Low Na+ Cochlear Endolymph has
electrical potential of+85mV This difference in ion composition and
electricalpotential difference provide energy for cochleas
5. Functional units of cochlea The Organ of Corti sensor of
cochlea, converts &lifies mechanical sound to electrical
signals(Mechano-electrical Transduction) Stria Vascularis cochleas
battery, generatesenergy (Endocochlear potential) The Spiral
Ganglion these are neurons featuringaxons, electrical wires,
transporting signals fromcochlea to CNS.
6. The Organ of Corti & surr. structures
7. THE ORGAN OF CORTI Named after Alfonso Giacomo Gaspare Corti
It consists of two types of sensory receptors inner and outer hair
cells There are about 3500 flask shaped Inner HairCells, lined up
in a single row, through out entirelength of S.M. Lateral to IHCs
lies 3 rows of Outer Hair Cells,cylindrical shaped. They contain
hair bundles consists of Actin-filledstereocilia, graded acc. to
height, with the mostlateral being the tallest and most medial
rowbeing the shortest.
8. The hair bundles of IHCs are organized as asmooth curved
line of 2-3 rows of stereocilia &OHCs stereocilia bundles are
arranged in ashallow V-shape. They are Mechano-sensitive organelles
of H.C.s Every H.C. sits over a phalangeal supporting cell,i.e.
Deiters Cells for OHCs The inner and outer pillar cells delienate
the areabetween IHCs & OHCs framing the Tunnel ofCorti Other
supporting cells embrace the hair cell-bearingpart of organ of
corti
9. Cochlear Stereocilia
10. Medially, Inner Marginal Cells Laterally, Hensens (Outer
Marginal) Cells Clauduis Cells Boettcher Cells The O.o.C. is
covered by Tectorial membranealong its entire length, an acellular
structure andis medially attached to spiral limbus &
connectshair bundles to OHCs.
11. The BASILAR MEMBRANE &TONOTOPY Sound eardrum vibration
ear ossicles inner ear Movement of stapes causes displacement
ofcochlear fluid in scala vestibuli The incompressibility of
perilymph causes apressure gradient between SV & ST, leading
tomovement of Basilar membrane & Organ of Corti Its like a
TRAVELLING WAVE moving from baseto apex along the basilar
memb.
12. A pure tone stimulus, the travelling wave movesfrom base to
apex, reaches a Maximum* at acharacteristic place along basilar
memb. andthen decays This precise location of this Maximum
dependson Frequency of stimulus the principle oftonotopic
organisation of cochlea The base is tuned for frequencies of 20 kHz
andapex for 20 Hz The tonotopic gradient is a continuous gradient
inbasilar membrane width and also with changes inhair cell height
and length of cellular structuresi.e. stereocilliary hair
bundles
13. Tonotopic organization of Organ of Corti
14. Inner Hair Cells &Mechanoelectrical Transduction The
IHCs are sensory cells which convertmechanical stimulation to
electrical signals andthe synaptic activity is transmitted to brain
This M-E Transduction occurs at tips ofstereocilia This apparatus
is present in all hiar cells &consists of mechanically gated
ion channels,that is closely asso. with elastic structures &
aTip-link, that connects the tip of the stereociliumto the side of
the next tallest stereocillium
15. Components of Stereocilia Cadherin 23 & Protocadherin
15 components ofTip-Link Mechano-Electrical Transduction channel
Insertional Plaque Myosin 1C Actin filaments Mechanical deflection
of hair cell bundles leadsto mechanical tension leading to
conformationalchange in transduction channel protein &increase
in channel opening USHER SYNDROME - Congenital Hearing Loss+
Progressive loss of vision due to RetinitisPigmentosa
16. On mechanical stimulation Towards the TALLEST row of
stereocillia K+ &Ca++ ions enter hair cell through open M-E
Tchannels, located near the tips leads toDEPOLARISATION of cell
Towards the SHORTEST stereocillia, the M-E Tchannels close leads to
HYPERPOLARISAITONof cell After a sustained excitatory deflection of
hairbundle, the initial large transduction currentADAPTS,
manifesting as decline of current correlated with closure of
transduction channels 2 distinct process involved in adaptation1.
Rapid reclosure of transduction channels2. Sliding of myosin based
motor asso. withtransduction appa.
17. Mechanoelectrical Transduction
18. 1st - Rapid Channel reclosure Fast Adaptation Ca++ binding
to intracellular site near ch. gate 2nd - Slow Adaptation 10 tmies
slower than Fast Adap. Upper tip-link slides down the stereocilum
During sustained stimulus, adaptation leads toresetting of restore
point, hence allowing thetransduction apparatus to function at
point ofhighest sensitivity The influx of Ca++ through open
transductionchannels leads to slippage of myosin basedadaptation
motor, which continuously strives tocrawl towards the
stereocilliary tip along actincore
19. The slippage of the myosin based motorchannels reduces the
tension in the tip-linkcomplex & lowers the open probability of
thetransduction channels, shutting off the Ca++influx At Low Ca++
levels myosins of the adaptationmotor will effectively move upwards
readjusting the tension in the tip-link complex toa pointwhere the
open probability ofthe M-E T ch. is close to theopen probability at
rest. Myosin 1C is crucial for thisadaptation process
20. OUTER HAIR CELLS & AMPLIFICATION OHCs have a key role
in amplification of Basilarmemb. motion Amplification is necessary
for detection of soundsat low pressure levels OHCs are mainly
responsible forAMPLIFICATION & SHARP TUNING of Auditorysystem
Mechanism of Amplification Somatic Electromotility the OHCs
changetheir length by 3-5% in response to
electricalstimulation
21. When Depolarized, they Contract & whenHyperpolarized
they Elongate The OHCs exert mechanical force causingmovements of
basilar memb. motion caused by thetravelling wave Prestin motor
protein responsible for somaticelectromotility in outer hair cells.
It belongs to SLC26anion transporter superfamily
mediateelectroneutral exchange of chloride & carbonateacross
plasma membrane Hypothesis* - intracellular anions act as
voltagesensors which bind to prestin and triggerconfirmational
changes Hyperpolarisation anion binding to prestin increase in
surface area of prestin cell elongation Depolarisation dissociation
of anion decrease inthe prestin surf area cell contraction At rest
anions are usually bound to prestin longer
22. TECTORIAL MEMBRANE It is an extracellular structure
overlying IHCs &OHCs & it changes its size from base to
apex Only the tallest stereocillia of OHCs are directlyembeded into
the undersurface of tectorialmembrane TM is attached on its inner
edge to spiral limbus& is loosely connected to the supporting
cells Hensens cells by trabeculae *Mutations in TM genes Alpha
& Beta Tectorin caused profound hearing loss It is more like a
resonant gel which is involved inenhancing the frequency
selectivity of cochlea
23. STRIA VASCULARIS Plays an important role in cochlear
hemostasisby generating endocochlear potential &maintaining the
unique ion compostion ofendolymph Highly vascularized,
multi-layered tissue & is apart of lateral wall of SM 3
distinct cell types Marginal Intermediate Basal
24. Stria Vascularis & K+ Circulation
25. Tight junction demarcate strial tissue & provideionic
barriers with marginal cells at one end &basal cells at other
end The extracellular space between these twobarriers is known as
intrastrial compartment Marginal cells separate endolymph filled
scalamedia from interstitial compartment that is filledwith
interstitial fluid Basal cells separate interstitial cells
fromperilymph that surrounds fibrocytes of the spiralligament The
intermediate cells as well as blood vesselsare embeded in
intrastrial compartment
26. Passage of ions from Perilymph to Endolymph in SV
27. Gap junctions connect basal cells with withintermediate
cells & with fibrocytes of spiralligament allowing Electric
coupling &exchange of ions and small molecules The regulation
of cochlear fluid homeostasisalso involves endolymphatic sac,
whichresponds to endolymph volume disturbance Malfunctions in
cochlear fluid homeostasis dueto disruptions of endocochlear
potential, ioniccomposition, or its volume regulatingmechanism
leads to varios forms of hearingimpariment
28. ENDOCOCHLEAR POTENTIAL& POTASSIUM HOMEOSTASIS Hair cell
mechanoelectrical transduction worksefficiently due to large
driving force for cations toenter cells cytoplasm from scala media
The +85 mV endocochlear potential ofendolymph & chemical
gradient of K+ are themain components of the driving force HEARING
THRESHOLD INCREASESAPPROXIMATELY BY 1dB PER 1mV LOSS OFENDOCOCHLEAR
POTENTIAL
29. K+ is the main cation of endolymph whichgenerates
endocochlear potential Movement of K+ in cochlea 1. K+ can enter
hair cells throughmechanoelectrical transduction channels &
isreleased through hair cells basolateralmembranes into
perilymphatic extracellularspace2. K+ can enter supporting cells
and move towardsthe spiral ligament by extensive gap
junctionnetwork3. Alternatively, K+ can diffuse extracellularly
viaperilymphatic space Spiral ligament composed of Type II &
Type Ifibrocytes take up K+ and provide anintracellular pathway
into basal & intermediatecells of stria vascularis
30. K+ flow through the Organ of Corti &Stria
Vascularis
31. K+ is released by intermediate cells via KCNJ10channels
into interstitial space from which it isactively pumped &
cotransported into marginalcells The marginal cells release K+ into
SM The K+ circulation is NOT a TRUERECYCLING* Malfunctions of
several K+ channels leads toperturbation of cochlear K+
homeostasis,resulting in hearing impairmentFor Ex loss of KCNE1
& KCNQ1 gene(encodes K+ ch subunits that allow secretion ofK+
from marginal cells to SM) leads to Jervelland Lange-Nielsen
Syndrome chatz by HearingLoss & Cardiac Arrhythmia
32. Passage of ions from Perilymph to Endolymph in SV
33. The most well known genes involved in cochlearhomeostasis
are the ones that encodeCONNEXIN Proteins Connexins form subunits
of gap junctionchannels, which underlie K+ circulation
networksdescribed for supporting supporting cells of theOrgan of
Corti, the spiral ligament & striavascularis Mutations
involving Connexins 26, 30, 31 & 43are responsible for majority
of non-syndromichereditary hearing loss
34. GENE PROTEIN PR. LOCATION PR. FUNCTION DISEASEKCNE1 KCNE1
Marginal Cells K+ Ch Jervell/Lange-Nielsen Synd.KCNQ1 KCNQ1
Marginal Cells K+ Ch Jervell/Lange-Nielsen Synd.KCNQ4 KCNQ4 OHCs
& IHCs K+ Ch DFNA2GJB2 Cx26 Fibrocy. in SL & SLiEpi. on BM,
I & B ClGap Junction Protein DFNB1/DFNA3GJB6 Cx30 Fibrocy. in
SL & SLiSupp Cells of OoCGap Junction Protein DFNA3GJB3 Cx31
Fibrocy. in SL & SLiEpithelia on BMGap Junction Protein DFNA2,
AR nonsynd. deafGJB1 Cx32 Fibrocy. in SL & SLiEpithelia on
BMGap Junction Protein X-linked CharcotMarie-Tooth
&DeafnessGJA1 Cx43 Fibrocy. in SL & SLiEpi. on BM, I &
B ClGap Junction Protein AR nonsynd.deafnessBSND Barttin Marginal
Cells Cl- Ch Ty 4 BarttersSyndrome
35. COCHLEAR FLUIDHOMEOSTASIS Perilymph, Endolymph &
Interstitial Fluid are 3types of fluids found in the cochlea &
itsmetabolic support system The proper ionic composition of these
fluids isessential for generation of endocochlear potential
Perilymph & Interstitial fluid have High Na+ & LowK+
Endolymph have Low Na+ & High K+ , Low Ca++
37. In stria vascularis , influx of Na+ accompaniesK+ from the
interstitial compartment intomarginal cells Cotransporter NKCC1
uses strong Na+ gradientto bring Na+, K+ & 2Cl- ions into
marginal cells Na+/K+ ATPase sets up this gradient by bypumping Na+
into intrastrial space in exchangefor K+ Lastly, K+ leaves marginal
cells to enterendolymphatic space This process maintains a High Na+
& Low K+concerntration of intrastrial fluid, which
facilitatesK+ replenishment into intrastrial space
38. Cl- is transported back to intrastrial space
byCIC-K/Barttin Channels Inhibition of NKCC1 & Na+/K+ ATPase by
LoopDiuretic Furosemide & Ouabain leads tosupression of E-C
Potential Mut. of Barttin gene or Mut. of both CIC-Ka &CIC-Kb
subunits of basolateral Cl- Ch. leads toBartters Synd Ty 4 chatz by
Deafness & Renalsalt wasting Na+ is reabsorbed from endolymph
by outersulcus & Reissners Memb cells, which play arole in
maintaining Low Na+ conc of SM
39. Ca++ regulation the tip-links break at very lowCa++ conc.
& the mechanoelectical transductionchannels are blocked at high
Ca++concerntrations Ca++ carries part of transduction current &
playscritical roles in Adaptation & CochlearAmplification Ca++
permeable channels Ca++ ATPases &Na+/K+ exchangers are also
found & regulateCa++ efflux & influx into Endolymph
40. Cochlear Fluid VolumeRegulation It is equally important for
cochlear funtion Previously, 2 popular theories Longitudinal
&Radial Flow patterns1. Longitudinal Flow of endolymph is the
secretionalong the membranous labyrinth withreabsorption in the
endolymphatic duct & sac2. Radial Flow is based on local
secretion &reabsorption, via stria vascularis Today, the
prevailing thought is that there is nosignificant volume flow under
physiologicalconditions
41. A low volume flow inside the cochlea hasconsequences for
intracochlear drug application,where under physiological
conditions, diffusionof compounds inside the fluid filled
compartmentappears to limit the equal dosing of potentialdrugs from
base to apex At cellular level, transmembranous watermovement
depends on aquaporins Aquaporin 2 found in Endolymph lining
epitheliumof endolymphatic sac & is regulated byVasopressin
Vasopressin also increases the activity ofepithelial Na+ ch &
NKCC1 cotransporters foundin strial marginal cells & type II
fibrocytes ofspiral ligament
42. Glucocorticoids have opposite effects supresssymptoms of
Menieres disease, by decreasingvasopressin production &
altering expression ofcertain aquaporins Cochlear fluid
homeostasis, Ion transport &Endocochlear potential are all req.
for propercochlear function Ageing, affects long-term maintainance
ofendocochlear potential & lowered metabolicrates of stria
vascularis plays a role in age rel.hearing loss
43. THE SPIRAL GANGLION Located in Rosenthals canal within the
Modiolus ofthe cochlea It contains cell bodies of afferent neurons,
itsdendrites are excited by neurotransmitter releasedby Organ of
Corti, hair cells & the axons of whichare projects centrally
into the cochlear nucleus, inbrain stem. Majority (95%) of afferent
fibers are thick &myelinated, & originate from type I
ganglionneurons, exclusively innervate IHCs Remaining (5%) of
afferent fibers are thin,unmyelinated & originate from type II
ganglionneurons, innervate OHCs
44. Spiral ganglion innervations
45. A dozen of type I ganglion neurons innervateeach IHCs
Converging innervation pattern The type II afferent nerve fibers
divide intomultiple branches & contact multiple OHCs Diverging
innervation pattern All auditory information is carried to
brainstemby afferent system, the auditory & vestibularnerves
join each other forming vestibulocochlearnerve Efferent fibers
originate in brain stem fromneurons located in Superior Olivary
complex &send information to cochlea by synapsing withOHCs
& with the afferent fibers beneath IHCs
46. NEURAL PROCESSING OF AUDITORYINFORMATION & I.H.C.
SYNAPSES Afferent NT release by IHCs is initiated at their
5-30ribbon type synapses, where local influx of Ca++through voltage
gated Ca++ channels leads tofusion of synaptic vesicles at
presynaptic sites Each ribbon synapse is composed of
presynapticdense body & is surrounded by vesicles
containingNTs, thick plasma memb, synaptic cleft & postsynaptic
region containing AMPA-type glutaminergicreceptors of afferent
nuerons
47. The tonotopic organisation of the organ of cortiis also
maintained within the afferent system,where the depolarisation of
IHCs at specificlocation leads to excitation of connected
afferentspiral ganglion neurons Each afferent is characterized by a
specificturning curve, that decribes the sound pressurelevel of
stimulus needed to elicit a response at agiven frequency The
feature of turning curve is that they showthe frequency at which
the nerve fibers displayshighest sensitivity its characteristic
frequency Loss of cochlear amplification, i.e. by OHCsloss, leads
to broadening of turning curve &increase in fibers response
thresholds
48. Tonotopic organisation of cochlea is the basisfor frequency
coding in auditory nerve fibres Place Coding Frequency is coded by
auditory nerve fibresdischarge characteristics Phase Locking,
ithappens only at low frequencies Tonotopic organisation &
Phase Locking areboth important for frequency discrimination
Discharge rates are determined by frequency &intensity of
stimulus
49. As intensity increases, the discharge rate with ina single
auditory nerve fibre increases, & the no.of auditory nerve
fibers activated at a givencharacteristic frequency increases
withintensifying stimuli The recruitment of more fibers is because
ofauditory nerve fibers of same characteristicfrequency have
different response thresholds With increased stimulus intensity,
other afferentnerve fibres of nearby characteristic frequencyare
also activated.
50. EFFERNET INNERVATION OFCOCHLEA 2 different types of
efferent fibers originate inbrain stem1. Myelinated Medial
Olivocochlear (MOC)Efferents arise from neurons located
aroundMedial Superior Olivary Nucleus. They project toC/L & I/L
cochlea, where they form cholinergicsynapses with OHCs2.
Unmyelinated Lateral Olivocochlear (LOC)Efferents arise from
Lateral Superior OlivaryNucleus. Fibers project predominantly into
I/Lcochlea, where they terminate on dendrites ofAfferent type I
neurons beneath IHCs LOC efferent synapses are neurochemically
51. The MOC system Stimulation of MOC system leads to
increasedthresholds, due to decrease in the degree ofcochlear
amplification by OHCs This sound-evoked feedback,
decreasessensitivity of hearing apparatus whenmetabolically
expensive amplifications systemsare not needed The LOC system Their
direct input on afferent neurons, suggeststhey regulate afferent
activity affects dynamicrange LOC feedback systems are slow &
req. minutes tobecome effective It also additionally functions to
perform slowintegration & adjustment of binaural inputs
neededfor accurate binaural function & sound localization
52. The activity of MOC & LOC efferent systemsseems to have
protective effects againstacoustic injury, important in loud
noiseenvironments.
53. Thank You...REFERENCES Surgery of the Ear
Glasscock-Shambaugh - 6thEd. Scott Browns Otorhinolaryngology &
Head andNeck Surgery - 7th Ed. Cummings Otolaryngology & Head
and NeckSurgery - 5th Ed. Mohan Bansal 2nd Ed.














![Inner Ear Anatomy[1]](https://img.pdfslide.us/doc/110x75/5528566b4979591c048b47a6/inner-ear-anatomy1.jpg)

