
1 1 2 1,3
1 Aquaculture & Fisheries Development Centre, School of Biological, Earth & Environmental Sciences, University College Cork,
Distillery Fields, North Mall, Cork, Ireland; 2 Kerry BioScience Ltd., Kilnagleary, Carrigaline, County Cork, Ireland;3 Department of Zoology, Ecology & Plant Science, School of Biological, Earth & Environmental Sciences, University College
Cork, Cork, Ireland
As part of a programme to develop sustainable diets for
macroalgivores, a 3-month experiment was conducted to
determine the effects of konjac glucomannan–xanthan gum
(KX) binder configuration on formulated feed stability, feed
palatability and growth performance of juvenile, hatchery-
reared, Haliotis discus hannai. This study was conducted in a
recirculation facility in which four KX binder configurations
were evaluated in a series of isonitrogenous experimental
feeds and freshly harvested Laminaria digitata was included
as a natural feed type. Dry matter leaching of the experi-
mental feed treatments was assessed with no significant dif-
ference in the dry matter leaching between treatments
observed. No differences (P > 0.05) were found in percent-
age survival, daily food consumption (DFC) and linear
growth rate (LGR) between treatments. Food conversion
efficiency (FCE), specific growth rate (SGR) and body
weight/shell length (BW/SL) ratio were significantly higher
when offered L. digitata. Trends showed that the best per-
forming KX feed in terms of FCE, LGR, SGR and BW/SL
ratio was produced with the 2% KX; 1 : 1 binder.
KEY WORDSKEY WORDS: abalone, binder, feed production, growth, Hali-
otis, nutrition
Received 4 March 2010, accepted 2 July 2010
Correspondence: Maria O�Mahoney, Aquaculture & Fisheries Development
Centre, University College Cork, Distillery Fields, North Mall, Cork,
Ireland. E-mail: [email protected]
It is widely accepted that aquaculture is pivotal to the future
maintenance of commercial fishery markets. Key challenges
that are faced by the industry include issues of sustainability,
feed availability and technological challenges to improve
production efficiency. For abalone culture, commercial feeds
exist, but the low uptake by producers in Ireland continues to
put pressure on local marine ecosystems because natural
harvests of marine kelp continue to be used as the primary
feed source. When harvested sustainably, i.e. a stump of
25 cm is left intact; regeneration can take 3–5 years (Werner
& Kraan 2004). However, the impact of locally intensive
harvesting of near-shore kelp beds can result in slow regen-
eration times and altered community structure (Kelly 2005).
Multiple stakeholders in the aquaculture sector in Ireland
have nominated feed development and availability as a
priority research focus for this sector. This research was
conducted as part of a feed development programme for high
value shellfish species in Ireland.
Recent FAO statistics indicate that world capture produc-
tion of abalone has declined in all producing countries. Max-
imal output of abalone aquaculture existed in just two of
the thirteen producing countries, China and Korea, in 2007
(FAO 2009). The high market value of abalone suggests that
intensification of commercial abalone culture is necessary to
sustain market demands. A diet that reduces the reliance on
non-sustainable feed sources is critical to commercial viability.
Target nutritional requirements for several commercially
lucrative species of abalone have been identified through
considerable research in recent decades. Regionally devel-
oped artificial feeds exist for locally produced species, and
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
� 2011 Blackwell Publishing Ltd
2011. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
doi: 10.1111/j.1365-2095.2010.00816.x
Aquaculture Nutrition

a number of feeds produced by extrusion cooking are
available. However, high importation costs have restricted
widespread use of currently available feeds in some regions.
The application of cost-effective, innovative feed production
technologies is necessary to achieve a locally dynamic and
internationally competitive aquaculture industry.
Binder configuration is important for abalone feed devel-
opment because of the perceived influence on feed palat-
ability. A review by Fleming et al. (1996) outlined the pivotal
role of feed binders in commercially focussed artificial feed
development for abalone. For abalone, previous studies have
shown that binder strength and feed palatability are nega-
tively correlated (Uki et al. 1985; Gorfine 1991). Similarly, a
negative relationship between consumption and feed tough-
ness has been demonstrated for Haliotis rubra and Haliotis
midae offered natural macroalgae species (Day & Cook 1995;
Fleming 1995). Coupled with this, leaching of feed attrac-
tants from artificial feeds is desirable in intensive culture
whereby the naturally passive feeding nature of abalone may
benefit from stimulation (Harada et al. 1996; Mai 1998;
Fermin 2003; Troell et al. 2006).
Hydrocolloids (e.g. gelatin, guar gum, sodium alginate,
agar, carrageenan, carboxymethylcellulose) as feed binders
are advantageous for artificial feed production because they
are easy to produce in small scale, and the effectiveness of
hydrocolloid-bound on-growing feeds has been demonstrated
for several aquatic species such as shrimp (De Muylder et al.
2008; Palma et al. 2008), sea urchins (Caltagirone et al. 1992;
Pearce et al. 2002; Mortensen et al. 2003), abalone (Uki et al.
1985; Knauer et al. 1993; Britz 1996a; Coote et al. 1996),
tilapia (Fagbenro & Jauncey 1995), crayfish (Ruscoe et al.
2005), sole (Liu et al. 2008) and rainbow trout (Storebakken
& Austreng 1987). However, some hydrocolloid binders such
as gelatin and sodium alginate are expensive, and their use in
formulated feed development may be prohibitive to overall
low-cost feed production goals. Other hydrocolloid binders
are more cost-effective as feed binders, and applications of
these binders have often been demonstrated in other areas.
Two such binders are konjac glucomannan and xanthan gum.
Konjac is a glucomannan polysaccharide obtained from the
tubers of Amorphophallus konjac and has a linear backbone
of (1 fi 4)-linked b-D-mannose and b-DD-glucose, in the ratio
1.6 : 1 (Kato & Matsuda 1969). Solubility is conferred by the
presence of acetyl groups (approximately, 1 acetyl group per
17 residues) at the C-6 position, and short oligosaccharide
side chains at the C-3 position of the mannoses. Xanthan
gum is an extracellular polysaccharide derived from the
bacterium, Xanthomonas campestris, which is a naturally
occurring pathogen of the brassicas (Kelco 2007). Xanthan
gum has a (1 fi 4)-linked linear backbone of b-D-glucose, as
in cellulose, and is solubilized by charged trisaccharide side
chains attached at O-3 of alternate glucose residues to give a
pentasaccharide repeating sequence (Melton et al. 1976). On
cooling and/or addition of salt, it undergoes a co-operative
conformational transition (Morris et al. 1977) from a disor-
dered coil to a rigid ordered structure (5-fold helix). Solutions
of xanthan gum in its ordered conformation show tenuous
‘‘weak gel’’ properties, which appear to arise from weak
association between the helical sequences, and are sensitive to
ionic environment. Xanthan gum alone, however, does not
give �true� (self-supporting) gels, but will form strong
and cohesive gels when mixed with konjac glucomannan
(Goycoolea et al. 1995). The exact mechanism of gel forma-
tion is not yet established, but probably involves association
of �smooth� regions of the (1 fi 4)-diequatorially linked of
konjac glucomannan to the cellulosic backbone of xanthan
(also (1 fi 4)-diequatorially linked). The formation of rec-
ognizable gels was found at total polysaccharide levels of only
0.02%, which is the lowest gelling concentration observed for
a carbohydrate system (Dea 1993). Both konjac glucomannan
and xanthan gum have been widely used in the food industry.
The formation of the gel network in the presence of the feed
physically entraps the material. Any interactions with the
components contained within the feed would result in the
compromised formation of this network. Applications of the
konjac glucomannan–xanthan gum binders have been dem-
onstrated in other areas (e.g. pharmaceutical and prosthetics)
(Eyo-okon & Hilton 2003; Alvarez-Mancenido et al. 2008;
Fan et al. 2008), but this binder complex has not previously
been evaluated for aquatic feed production.
Considerable development studies of the konjac gluco-
mannan–xanthan gum (KX) binder for use in aquatic feeds
has identified three major variables in the KX binder con-
figuration for artificial feeds (i) the use of seawater (SW) or
distilled water (DW) in binder preparation, (ii) overall binder
concentration and (iii) the ratio of K to X (O�Mahoney
2009). The aim of this study was to assess a formulated feed
with four forms of the KX binder on feed palatability and the
growth performance of cultured Haliotis discus hannai. The
results of this study were pivotal to further feed development
for H. discus hannai using the KX binder.
Konjac glucomannan (KJ2B) was obtained from Mannasol
Products Ltd (Cheshire, UK) and xanthan gum (Keltrol [E])
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

was obtained from CP Kelco (Surrey, UK). Laminaria digi-
tata meal was obtained from Arramara Teoranta (Co. Gal-
way, Ireland), soy protein isolate was obtained from Holland
and Barrett (Cork, Ireland), CaCO3 was obtained from
Sigma-Aldrich Ltd (Dorset, UK), and vitamins and minerals
were obtained from Inform Nutrition (Cork, Ireland).
Experimental feeds in this study were prepared according
to recently developed methodology for feed production
using the konjac glucomannan–xanthan gum binder
(O�Mahoney 2009). Dry feed ingredients were mixed for
10 min in a commercial food mixer. The konjac glucoman-
nan and xanthan gum powders were weighed out in separate
batches at the required concentration and mixed to com-
bine. The total volume of water (SW or DW) required was
placed in a stainless steel pot with a mixing paddle, and the
mixed powders were dispersed using a fine dredger. When
all the powders were dispersed, mixing was continued for
30 min. The viscous solution was placed in a hot water
bath (set temperature 85 �C) and heated for 30 min after a
minimum internal gel temperature of 72 �C was reached.
Dry feed ingredients and lipids were added to the hot gel
solution, and the composite feed was mixed in a commercial
food mixer for 3 min. The hot feed was poured into a rigid
container, cooled rapidly and refrigerated overnight. On the
following day, the feeds were sliced into strips and air-dried
in a drying room equipped with a convector heater and
dehumidifiers for 24–30 h (average temperature 30 �C;average relative humidity 28%). Air-dried feeds were stored
in air-tight containers.
To evaluate the key parameters of the KX binder config-
uration and the implications for H. discus hannai culture, an
experimental growth trial consisting of the following feed
treatments was conducted:
• Laminaria digitata
• 2% Distilled water K:X (1 : 3) (Diet A: DW 2% KX
1 : 3).
• 2% Seawater K:X (1 : 3) (Diet B: SW 2% KX 1 : 3).
• 2% Seawater K:X (1 : 1) (Diet C: SW 2% KX 1 : 1).
• 1.5% Seawater K:X (1 : 1) (Diet D: SW 1.5% KX 1 : 1).
The dietary treatments and feed ingredient composition
are shown in Table 1. Proximal composition and energy
composition of the feed treatments are shown in Table 2.
Table 1 Ingredient composition of the
experimental feeds (g dry matter kg)1)Ingredients
Diet A
(DW 2%KX; 1 : 3)
Diet B
(SW 2%KX; 1 : 3)
Diet C
(SW 2%KX; 1 : 1)
Diet D
(SW 1.5%KX; 1 : 1)
KX binder 174.7 174.7 174.7 174.7
Laminaria
digitata meal
319.7 319.7 319.7 319.7
Soy protein isolate 352.9 352.9 352.9 352.9
CaCO3 107.2 107.2 107.2 107.2
Vitamins1 14.9 14.9 14.9 14.9
Minerals2 30.6 30.6 30.6 30.6
DW, distilled water; KX, konjac glucomannan–xanthan gum; SW, seawater.1 Vitamin premix composition per kg of experimental feed (Mai et al. 1995a) : thiamine HCl
120 mg; riboflavin 100 mg; folic acid 30 mg; PABA 400 mg; pyridoxine HCl 40 mg; niacin 800 mg;
Ca pantothenate 200 mg; inositol 4000 mg; ascorbic acid 4000 mg; biotin 12 mg; vitamin E
450 mg; menadione 80 mg; vitamin B12 0.18 mg; vitamin A 100 000 IU; vitamin D 2000 IU;
ethooxyquin 400 mg.2 Mineral premix per kg of experimental feed (Mai et al. 1995a; Tan et al. 2001) : NaCl 0.4 g;
MgSO4.7H2O 6.0 g; NaH2PO4.2H2O 10.0 g; KH2PO4 20 g; Ca(H2PO4)2.H2O 8.0 g; ferric citrate
1.0 g; ZnSO4.7H2O 141.2 mg; MnSO4.2H2O 64.8 mg; CuSO4.5H2O 12.4 mg; CoCl2.6H2O 0.4 mg;
KIO3 1.2 mg; Na2SeO3 0.4 mg.
Table 2 Proximal composition of the
feed treatments (g kg)1 of dry matter)
Component
Diet A
(DW 2%KX;
1 : 3)
Diet B
(SW 2%KX;
1 : 3)
Diet C
(SW 2%KX;
1 : 1)
Diet D
(SW 1.5%KX;
1 : 1)
Laminaria
digitata
Protein1 387.6 361.0 361.3 357.0 148.4
Ash1 303.0 363.0 360.0 362.6 285.1
Carbohydrate1 306.4 275.2 278.3 276.5 516.3
Fat1 3.1 0.9 0.4 3.9 50.2
Energy2 11.74 10.68 10.72 10.75 13.02
DW, distilled water; KX, konjac glucomannan–xanthan gum; SW, seawater.1 Values given as g kg)1 of dry matter.2 Energy values in kJ g)1.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

Hatchery-produced H. discus hannai [20–30 mm shell length
(SL)], which were reared on L. digitata, were collected from
Brandon Bay Seafoods, County Kerry, Ireland in February
2008 and transported to the Aquaculture & Fisheries
Development Centre, University College Cork (AFDC,
UCC) in large, cable-tied plastic bags that were dampened
internally with SW and pumped with oxygen to reduce gill
desiccation and aerobic stress (Sales & Britz 2001). Abalone
were maintained on a ration of L. digitata at the AFDC for
2 weeks to acclimate to the change in conditions.
The experimental system in this study consisted of a recir-
culation culture system consisting of 15 independent rectan-
gular plastic tanks (320 mm L · 175 mm W · 150 mm H),
each containing a rearing unit composed of aquamesh lined
with corrugated plastic inserts. Each tank was aerated with a
centrally positioned airstone. The experimental system was
enclosed within a black surround such that abalone were
maintained in constant darkness throughout the study based
on evidence in Garcia-Esquivel et al. (2007). Outflow from
each tank was recirculated through a 2500- L SW reservoir
known as the Global Oceans System. Aeration in the SW
reservoir was maintained through a series of airlines lining
the base of the Global Oceans floor.
Water quality in the Global Oceans system was maintained
by weekly renewal of 50% of the SW capacity. Inflow into
the experimental tanks was monitored weekly using a
Palintest� photometer to measure levels of ammonia
(0.18 ± 0.048 mg L)1 N), nitrite (0.02 ± 0.005 mg L)1 N),
nitrate (0.22 ± 0.034 mg L)1 N), pH (8.23 ± 0.004) and
alkalinity (186.67 ± 4.495 mg L)1 CaCO3). Salinity was
monitored on a weekly basis using a hand-held refractometer
and averaged at 35.5 ± 0.230 g L)1. SW temperature and
dissolved oxygen was monitored using the OxyGuard System
(OxyGuard, Birkerod, Denmark) and averaged at 17.33 ±
0.321 �C and approximately 10.1 ± 0.08 mg L)1, respec-
tively, over the duration of the study.
After the two week acclimation period, the H. discus hannai
were starved for 7 days prior to the initiation of the experi-
ment. Individuals were drained of excess surface moisture
prior to measuring for individual SL (±0.1 mm) and weight
(±0.01 g) and randomly divided between 15 experimental
tanks (n = 15 tank)1). The initial stocking parameters of
each tank were 25.89 ± 0.18 mm SL and 2.79 ± 0.06 g live
weight (mean ± SE).
This study consisted of five experimental treatments of
which four feeds were novel formulated KX feeds. The
control treatment was fed locally harvested L. digitata. Three
replicates of each feed treatment were conducted. The
experimental set-up allowed interchanges of tank position at
each feeding interval for a completely randomized experi-
mental design.
Feed replenishment was conducted every 3/4 days and aba-
lone were fed ad libitum. Dry matter conversion factors for
the artificial feeds being fed throughout the trial were
obtained by drying five random samples of each diet in the
oven at 105 �C overnight at the beginning of the experiment.
Fresh L. digitata was obtained from Ballycotton Bay, Co.
Cork approximately every 7–10 days and stored in an aer-
ated SW tank at the AFDC. The L. digitata was removed
from the SW as required, and all epiphytes were removed. At
each feeding interval, a sample of the L. digitata frond used
in feeding was dried in an oven (n = 3) at 105 �C for 24 h to
obtain a dry matter conversion factor for that feed. All
conversion factors for day 0 feeds were calculated as follows:
C ¼ d=w
where C = conversion factor, d = dry weight of the feed (g)
and w = wet weight of the feed (g).
Control samples (triplicate) of fresh L. digitata were run at
each feeding interval to correct for loss in macroalgal bio-
mass in the absence of abalone. Correction factors for KX
feed consumption were determined from a dry matter
leaching assessment conducted at the end of the experiment.
Daily feed consumption rates (DFC) and food conversion
efficiencies (FCE) were calculated using the formula:
DFC ¼ ½ðFg � FuÞ=t�=ðW Þ
where the DFC was the daily food consumption (DFC)
(mgdry matter g)1
abalone day)1), Fg was the dry weight (g) of
food given during the experimental period, Fu was the dry
weight (g) of food uneaten during the experimental period,
W was the mean wet weight of abalone during the experi-
mental period (assuming linear growth) and t was the time in
days. FCE was calculated from the formula:
FCE ¼ 100ðWf � WiÞ=ðFg � FuÞ
where FCE was the food conversion efficiency (%), Wf and
Wi were the final and initial whole abalone wet weights (g),
Fg was the dry weight of food given (g) during the experi-
mental period, Fu was the dry weight of food uneaten (g)
during the experimental period.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

Initial and final biometric data (length; mm and weight; g)
were recorded during this study to minimize the impact of
sampling intervals on overall H. discus hannai health and
homeostasis (Sales & Britz 2001). Growth rate was calculated
in terms of (i) SL (linear growth rate; LGR) and (ii) total
weight (specific growth rate; SGR) using the following
equations:
LGR ðmm SL day�1Þ ¼ ðLf � LiÞ=t
SGR ð%day�1Þ ¼ 100ðlnWf � lnWiÞ=t
where Li and Wi are initial length and weight, respectively,
Lf and Wf are the final length and weight, respectively, t is
the time (days) and ln is the natural log.
The body weight to SL ratio (BW/SL) was calculated at
the beginning of this study and for all experimental treat-
ments at the end of the study using the equation:
BW : SL ðg mm�1Þ ¼Mean weight=mean length
Each tank was checked for mortalities at each feeding
interval. Overall percentage survival (St) was calculated as
the ratio of the number of individuals surviving at the end of
experiment to the number of individuals at the beginning of
the experiment using the equation:
St ¼ Nt=No�100
where Nt is the number of abalone surviving at the end of
the experiment, and No is number of abalone at the
beginning of the experiment.
Tests to determine the dry matter leaching of the feed
treatments in this study in the absence of abalone were
conducted over 3/4 days at the end of this study in the
experimental system vessels. The dry matter loss values of the
formulated KX feeds were used for corrected consumption
estimates in this study.
The aquaria were labelled with the diet treatments of this
study (n = 3). Into each tank, two pieces of the appropriate
feed were placed. Each feed piece was preweighed and
labelled with a twine tag corresponding to the days of the
stability test (1st and 2nd). On day 0 of the stability test, dry
matter conversion factors for the feeds in the stability test
were obtained using identical procedures to those outlined
earlier (see Feeding, consumption and growth). These values
were used to calculate the average dry matter (g) in each feed
piece used in the SW stability test. On each day, the feed
piece with the label corresponding to that day number (1–2)
was removed from the aquaria, drained on absorbent paper,
weighed and oven-dried at 105 �C for 24 h to determine the
dry weight (g) of the feed piece. The dry matter leaching
(g kg)1) on each day was calculated using the equation:
Dry matter loss ðg kg�1Þ ¼ ðDo � DmÞ=Do � 103
where, Do is the dry matter (g) in the original feed piece, Dm
is the dry matter (g) in the final feed piece after immersion in
SW.
Tank position was randomized throughout the experiment,
and the possibility of tank effects was assessed using a
randomized ANOVAANOVA at the end of the experiment. The
assumptions of normality were analysed using the
Kolmogorov–Smirnov or Shapiro–Wilks test. Comparisons
between groups of normal data were tested using ANOVAANOVA.
Non-normal data were compared using the non-parametric
Kruskal–Wallis test. Post hoc analyses were conducted using
the TUKEY multiple comparison test and the Kruskal–
Wallis multiple comparison test. All statistical analyses were
conducted using SPSSSPSS (Ver. 12.0.1 for Windows, SPSS Inc.,
Chicago, IL, USA). Significance was assumed when P < 0.05.
The initial abalone length and weight (mean ± SE) are
shown in Table 3. A one-way ANOVAANOVA indicated that there was
Table 3 Initial abalone stocking parameters (mean ± SE; n = 15)
Tanks Length (mm) Weight (g)
1 26.23 ± 0.796 2.97 ± 0.275
2 26.27 ± 0.846 2.97 ± 0.275
3 26.06 ± 0.759 3.01 ± 0.282
4 25.54 ± 0.821 2.68 ± 0.231
5 25.47 ± 0.683 2.57 ± 0.193
6 26.13 ± 0.539 2.80 ± 0.173
7 25.34 ± 0.818 2.75 ± 0.296
8 25.94 ± 0.772 2.94 ± 0.256
9 25.69 ± 0.668 2.59 ± 0.214
10 25.95 ± 0.729 2.75 ± 0.238
11 25.74 ± 0.729 2.81 ± 0.256
12 25.77 ± 0.533 2.63 ± 0.158
13 26.13 ± 0.626 2.87 ± 0.226
14 25.95 ± 0.752 2.69 ± 0.193
15 26.14 ± 0.831 2.79 ± 0.239
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd
no significant difference in the mean weight of each treatment
(ANOVAANOVA; d.f. = 4, 224; F = 0.722; P = 0.578).
The possibility of tank effect was tested at the end of the
experiment by randomized complete block ANOVAANOVA with tank
as a factor. Tank effect was non-significant (P > 0.05).
Immersion in SW resulted in dry matter loss in all experi-
mental treatments (Table 4).
On day 3, no significant differences in cumulative dry
matter leaching between treatments were observed (Kruskal–
Wallis; d.f. = 4; H = 8.433; P = 0.077). On day 4, dry
matter loss across all treatments was not significantly dif-
ferent (Kruskal–Wallis; d.f. = 4; H = 7.833; P = 0.098).
At the end of the experiment, mean per cent survival ranged
from 91.11 ± 5.9% for Diet A treatment to 100.00 ± 0.0%
for abalone fed L. digitata (Table 5). There were no signifi-
cant differences in the survival rates between treatments
(Kruskal–Wallis, d.f. = 4; H = 6.030, P = 0.197).
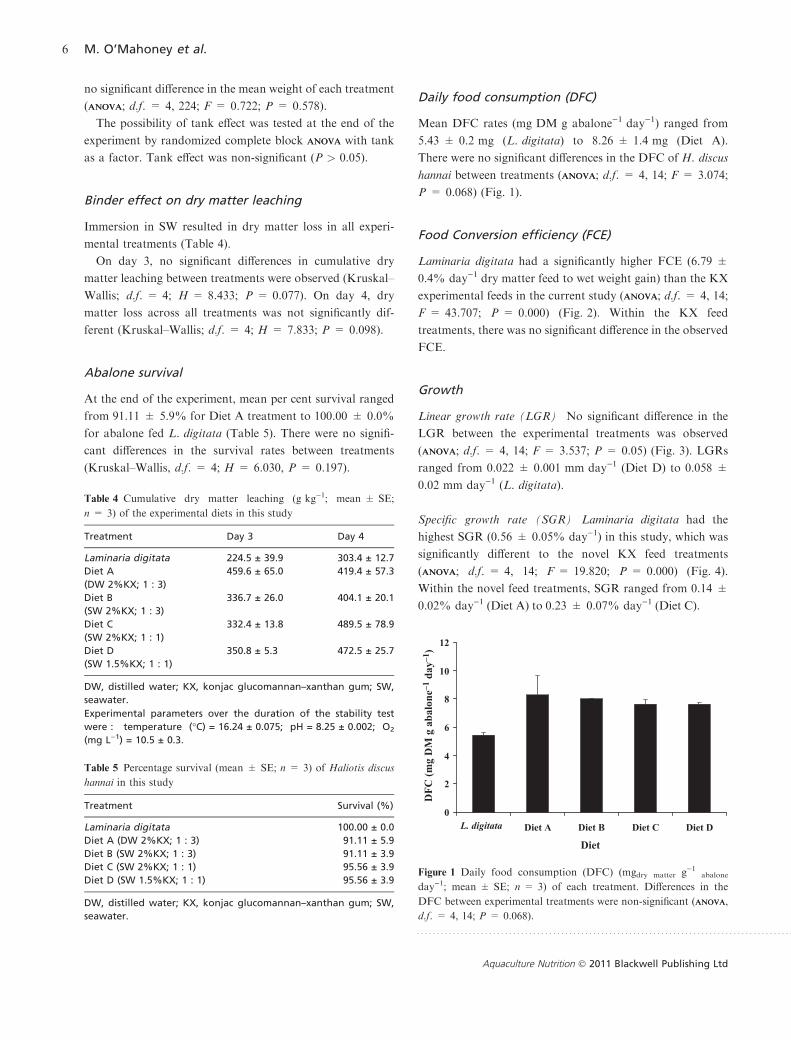
Mean DFC rates (mg DM g abalone)1 day)1) ranged from
5.43 ± 0.2 mg (L. digitata) to 8.26 ± 1.4 mg (Diet A).
There were no significant differences in the DFC of H. discus
hannai between treatments (ANOVAANOVA; d.f. = 4, 14; F = 3.074;
P = 0.068) (Fig. 1).
Laminaria digitata had a significantly higher FCE (6.79 ±
0.4% day)1 dry matter feed to wet weight gain) than the KX
experimental feeds in the current study (ANOVAANOVA; d.f. = 4, 14;
F = 43.707; P = 0.000) (Fig. 2). Within the KX feed
treatments, there was no significant difference in the observed
FCE.
Linear growth rate (LGR) No significant difference in the
LGR between the experimental treatments was observed
(ANOVAANOVA; d.f. = 4, 14; F = 3.537; P = 0.05) (Fig. 3). LGRs
ranged from 0.022 ± 0.001 mm day)1 (Diet D) to 0.058 ±
0.02 mm day)1 (L. digitata).
Specific growth rate (SGR) Laminaria digitata had the
highest SGR (0.56 ± 0.05% day)1) in this study, which was
significantly different to the novel KX feed treatments
(ANOVAANOVA; d.f. = 4, 14; F = 19.820; P = 0.000) (Fig. 4).
Within the novel feed treatments, SGR ranged from 0.14 ±
0.02% day)1 (Diet A) to 0.23 ± 0.07% day)1 (Diet C).
Table 5 Percentage survival (mean ± SE; n = 3) of Haliotis discus
hannai in this study
Treatment Survival (%)
Laminaria digitata 100.00 ± 0.0
Diet A (DW 2%KX; 1 : 3) 91.11 ± 5.9
Diet B (SW 2%KX; 1 : 3) 91.11 ± 3.9
Diet C (SW 2%KX; 1 : 1) 95.56 ± 3.9
Diet D (SW 1.5%KX; 1 : 1) 95.56 ± 3.9
DW, distilled water; KX, konjac glucomannan–xanthan gum; SW,
seawater.
Table 4 Cumulative dry matter leaching (g kg)1; mean ± SE;
n = 3) of the experimental diets in this study
Treatment Day 3 Day 4
Laminaria digitata 224.5 ± 39.9 303.4 ± 12.7
Diet A
(DW 2%KX; 1 : 3)
459.6 ± 65.0 419.4 ± 57.3
Diet B
(SW 2%KX; 1 : 3)
336.7 ± 26.0 404.1 ± 20.1
Diet C
(SW 2%KX; 1 : 1)
332.4 ± 13.8 489.5 ± 78.9
Diet D
(SW 1.5%KX; 1 : 1)
350.8 ± 5.3 472.5 ± 25.7
DW, distilled water; KX, konjac glucomannan–xanthan gum; SW,
seawater.
Experimental parameters over the duration of the stability test
were : temperature (�C) = 16.24 ± 0.075; pH = 8.25 ± 0.002; O2
(mg L)1) = 10.5 ± 0.3.
0
2
4
6
8
10
12
Diet A Diet B Diet C Diet D
Diet
DFC
(mg
DM
g a
balo
ne–1
day
–1)
L. digitata
Figure 1 Daily food consumption (DFC) (mgdry matter g)1abalone
day)1; mean ± SE; n = 3) of each treatment. Differences in the
DFC between experimental treatments were non-significant (ANOVAANOVA,
d.f. = 4, 14; P = 0.068).
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

Body weight/shell length ratio (BW/SL) Significant differ-
ences in the BW/SL ratios were evident in this study (Krus-
kal–Wallis; d.f. = 5; H = 15.822; P = 0.007) (Table 6).
The BW/SL ratio increased in all treatments when compared
with the initial BW/SL ratio (0.108 ± 0.001 g mm)1) but
Diet A (0.109 ± 0.002 g mm)1) did not show a significant
increase in BW/SL ratio over the duration of the study.
Laminaria digitata had the highest increase in BW/SL ratio
(0.155 ± 0.001 g mm)1) and was significantly higher than all
other experimental treatments at the end of this study. Diet C
(0.119 ± 0.007 g mm)1) had the highest BW/SL ratio of the
experimental KX feeds. This value was significantly different
to Diet A but not significantly different to Diet B
(0.114 ± 0.004 g mm)1) or Diet D (0.115 ± 0.001 g mm)1).
The results of the dry matter leaching assessment indicated
that dry matter leaching for the KX feeds did not differ
significantly to the control treatment, L. digitata. Varying the
concentration of KX and K to X ratios did not have a sig-
nificant effect on dry matter leaching in this study. It was
perceived that four days is the maximum duration for SW
stability for these feeds because removal of these feeds after
day 4 was hindered by feed disaggregating. Grazing is a slow
0
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
Diet A Diet B Diet C Diet D
–1
L. digitata
Figure 3 Linear growth rate (LGR) (mm shell length day)1;
mean ± SE; n = 3) of the experimental feeds in this study. There
was no significant difference in the observed LGR between treat-
ments (ANOVAANOVA; d.f. = 4, 14; P = 0.05).
b
b
bb
a
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Diet A Diet B Diet C Diet DDiet
SGR
(% d
ay–1
)
L. digitata
Figure 4 Specific growth rate (SGR) (% day)1, mean ± SE, n = 3)
of the experimental treatments in this study. SGR was significantly
different between treatments (ANOVAANOVA; d.f. = 4, 14; P = 0.000). The
results of the Tukey HSD are indicated by letters above the bars.
Where letters are not in common between bars indicates significant
differences between treatments (P < 0.05).
bb
bb
a
0
1
2
3
4
5
6
7
8
Diet A Diet B Diet C Diet D
FCE
(% d
ay–1
)
L. digitata
Figure 2 Food conversion efficiency (FCE) (% day)1 dry matter feed
to wet weight gain; mean ± S.E.; n = 3). Results showed that sig-
nificant differences in the FCE were evident between Laminaria
digitata and the experimental KX feeds (ANOVAANOVA; d.f. = 4, 14;
P = 0.000). The results of the Tukey HSD are indicated by letters
above the bars. Where letters are not in common between bars
indicates significant differences between treatments (P < 0.05).
Table 6 Body weight/shell length (BW/SL) ratios (mean ± SE;
n = 3) of the initial data and final dietary treatments of this study
Treatment BW : SL (g mm)1)
Initial 0.108 ± 0.001a
Laminaria digitata 0.155 ± 0.001b
Diet A (DW 2%KX; 1 : 3) 0.109 ± 0.002ac
Diet B (SW 2%KX; 1 : 3) 0.114 ± 0.004cd
Diet C (SW 2%KX; 1 : 1) 0.119 ± 0.007d
Diet D (SW 1.5%KX; 1 : 1) 0.115 ± 0.001d
DW, distilled water; KX, konjac glucomannan–xanthan gum; SW,
seawater.
Letters indicate the results of the Kruskal–Wallis multiple compar-
ison post hoc test. The values in the same column that do not have
the same superscript parameters were significantly different from
each other (P < 0.05).
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

feeding behaviour, and therefore abalone require that artifi-
cial feeds retain structural integrity in SW for at least 2 days
(Fleming et al. 1996). However, several studies report short
experimental periods of feed presentation ranging from 12 h
(Rivero & Viana 1996; Garcia-Esquivel et al. 2007) to 16 h
(Sales & Britz 2002a; b), whilst others report longer intervals
(3 days) between feeding to satiation (Mai 1998), and
therefore comparisons with this study are difficult to inter-
pret.
Previous studies have reported that blending and fine-
sieving of feed ingredients to <450 lm had a significant
influence on dry matter leaching of formulated feeds with
improved diet performance, measured as apparent digest-
ibility coefficients, for abalone diets produced within this
particle size category (Sales & Britz 2002b). By contrast,
however, Obaldo et al. (1999) reported poorer diet perfor-
mance for shrimp feeds at particle sizes <100 lm. Further
investigation to determine the optimal production method for
the novel KX feeds and factors that influence the property of
this binder for reduced dry matter leaching is required.
In this study, KX binder configuration did not significantly
affect diet performance on any parameter assessed. However,
the SGR achieved by the control treatment, L. digitata,
exceeded KX artificial diet performance in this parameter,
and overall, L. digitata had significantly higher FCE values
than all KX diet treatments.
Feed consumption rates in this study showed no significant
variability between all the experimental treatments. Although
non-significant, trends showed that the lowest DFC was
observed in the L. digitata treatment. The similarity of the
feeding rate, as measured by DFC, across all experimental
feed treatments suggests that feeding rate of H. discus hannai
for the feed treatments in this study was relatively constant
given the current feed formulations and experimental con-
ditions. Abalone feed by repeated rasping action of the
radula, and it is likely that the relatively soft texture of fully
hydrated KX feeds influenced feeding rates. As a result,
comparable daily feed consumption rates for the KX feeds
and L. digitata were achieved despite the abalone having
been predisposed to L. digitata. Previous studies have related
consumption in H. rubra to physical (e.g. toughness) and
chemical (e.g. presence of phlorotannins) feed characteristics
rather than nutritional quality (Fleming 1995), and indeed
the South African abalone, H. midae, has been found to
select for softer textured red and green macroalgae over kelp
species in feed preference experiments (Day & Cook 1995). In
Ireland, anecdotal evidence from producers pertaining to the
development of H. discus hannai culture have indicated that
the ecologically abundant species of kelp, Laminaria hyper-
borea, was unsuitable for culture of H. discus hannai
purported to result from the tougher texture of this algal
species (Hession et al. 1998).
Consumption rates in culture studies of H. discus hannai is
not a well-documented experimental parameter. Numerous
foundation studies on this species (for example: Mai et al.
1994, 1995a,b, 1996; Mai 1998; Tan et al. 2002a,b; Park et al.
2008) have measured food conversion ratio (FCR), growth,
survival and compositional tissue analyses as indicator of
the dietary requirements of the species. Mai et al. (1995b)
describes consumption in abalone as a very difficult para-
meter to evaluate in relation to artificial feeds because of the
highly particulate nature of the feed ingredients and the
feeding mechanism of the abalone. In essence, there is no
quantitative evaluation of ingestion as a result of the rasping
action of the radula, and there is no reason to assume that all
feed particles removed as a result of the feeding action are
consumed by the individual. In field studies, the proportion
of food consumed in abalone (Haliotis sp.) has been found
to be an indicator of feed palatability in situations where
the environmental availability allows access to more than
one feed type (Tutschulte & Connell 1988). Mercer et al.
(1993) found that feeding rates of H. discus hannai
(0.1–0.4% BW day)1) on six species of algae (Alaria escul-
enta, L. digitata, L. saccharina, Ulva lactuca, Palmaria
palmata and Chrondris crispus) and two mixed feeds levelled
off after 10 weeks, indicating a habitual acclimation to the
feed, and subsequent analyses of feeding rates were not
conducted in their study. When compared with Mercer et al.
(1993), the feeding rates of this study indicate comparatively
low feeding rates for H. discus hannai (0.5–0.8% BW day)1)
on all feed treatments. Our evaluation of consumption
throughout this study did not show any evidence of habitual
acclimation to the experimental feeds, and further study of
this aspect of abalone feeding behaviour may be necessary to
maximize feeding potential for experimental feeds. The
plastic responses of abalone to varied feed types (Garcia-
Esquivel & Felbeck 2008) coupled with adaptive physiolog-
ical processes suggests that, given time, abalone will consume
a range of feeds, and longer feeding trials would provide an
indication of adaptive feed responses.
The significantly higher FCE of the L. digitata treatment
in this study over all experimental KX feed treatments indi-
cated that the nutrient utilization of H. discus hannai for
fresh L. digitata was better than the KX feed formulation
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

ingredients. The proximal composition analyses, outlined in
Table 2, indicated notable differences in protein, carbohy-
drate and lipid concentrations between the natural and
experimental feed types. The success of a formulated feed
type may be dependent on a series of factors including feed
digestibility, gut physiology, gut transition times and inter-
active effects of nutrient composition that were not evaluated
in this study. The efficiency with which dry matter feed was
converted to wet weight gain despite comparable consump-
tion of L. digitata with the KX feeds may suggest an adaptive
response to this feed type. Haliotis discus hannai have asso-
ciative gut microflora for particular feed types, and the
composition of the gut microflora has been shown to change
over time and with changes in feed (Tanaka et al. 2003). In
addition, a 6- month experiment conducted by Garcia-
Esquivel & Felbeck (2006) found that digestive enzyme
activity in various parts of the gut of Haliotis rufescens al-
tered according to feed type.
It is suggested that the significantly higher FCE of
L. digitata in this study could have been as a result of a
history of feeding on L. digitata as this was the feed used at
the commercial hatchery from which the experimental
H. discus hannai were obtained. It is likely that the experi-
mental abalone were adapted to L. digitata as a feed type
through acquired optimal gut digestion and nutrient utiliza-
tion. It is therefore recommended that an adaptive period
should be allocated prior to growth assessment of artificial
feeds to remove any bias associated with previous feeding
history on growth assessment results (Pereira et al. 2007).
There are variable reports of acclimation periods for abalone
prior to experimental evaluation of formulated feeds in the
literature (Table 7). Further study is needed to determine the
optimal duration of acclimation period for experimental feed
types.
Although no significant differences in the LGR between
treatments were evident in this study, the SGR indicated
that a higher SGR was observed in the L. digitata treat-
ment. Haliotis discus hannai are slow growers achieving
approximately 1–2 cm growth in SL per year (Lee 2004).
Assuming linear growth, the LGR in this study of the
L. digitata treatment (0.058 ± 0.02 mm SL day)1) would
approximate to the maximum growth projected by Lee
(2004) in one year (2.1 cm year)1), and given that this
treatment was our experimental control, we evaluate the
growth achieved on the L. digitata treatment as a indication
that the experimental parameters were sufficient to promote
growth of H. discus hannai within the published range for
this species. Growth in the experimental KX feed treatments
in this study was below the projected annual growth range
for H. discus hannai as reported by Lee (2004) and lower
than published growth rate for this species in experimental
studies (Table 8).
BW/SL ratio is a useful determinant of profitability in a
commercial species such as abalone (Naidoo et al. 2006),
where the final market product is the whole animal tissue
weight. Moreover, the BW/SL ratio gives an indication of the
level of �fattening� achieved by on-growing feeds and as such
is a useful tool to collate the combined length and weight
achievements of a feed into a discernable parameter that
reflects commercial considerations. A commercially accept-
able feed will ideally achieve high BW/SL ratios more so than
extreme growth in either biometric dimension (e.g. length
and weight).
The BW/SL ratio in this study showed that the highest
BW/SL ratio was achieved on the L. digitata treatment
(0.155 ± 0.001 g mm)1). This was a significantly higher BW/
SL ratio than that of the initial sample (0.108 ±
0.001 g mm)1) and was also significantly higher than the
Table 7 Acclimation periods allocated
prior to abalone growth trialsReference Species Interim feed type
Acclimation
period
Britz (1996b) Haliotis midae Plocamium corallorhiza 3 months
Guzman & Viana (1998) Haliotis fulgens Artificial feed (abalone viscera
silage, fish meal &
soybean meal)
75 days
Coote et al. (2000) Haliotis laevigata Experimental study feeds 22 days
Shipton & Britz (2001) H. midae Fish meal artificial feed 1 month
Sales et al. (2003) H. midae Artificial feed–reared abalone 16 months
Macey & Coyne (2005) H. midae Basal artificial feed 3 weeks
Garcia-Esquivel et al.
(2007)
H. fulgens No interim feed mentioned 2 weeks
Zhang et al. (2008) Haliotis discus
hannai
Basal artificial feed 1 week
This study H. discus hannai Laminaria digitata 2 weeks
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

BW/SL ratio of all experimental KX feed treatments at the
end of the study. The BW/SL ratio of Diet A was not sig-
nificantly different to the initial BW/SL ratio, and this was
the only experimental KX diet that showed no significant
increase in BW/SL ratio over the duration of the growth trial.
Although differences between KX feeds were non-significant,
trends showed that Diet C (0.119 ± 0.01 g mm)1) was the
best performing experimental KX diet in terms of the
BW/SL ratio increase of all novel feeds.
Survival in all treatments in this study was high and
L. digitata promoted maximum survival (100%) in this
study. The overall high survival rate (>91%) in all treatments
in this study may be an indication of the general suitability of
the KX feeds for abalone culture.
High survival rates have been reported forH. discus hannai
in previous studies (Table 9). The high percentage survival in
this study may also be linked to the husbandry protocols
maintained throughout the experimental period. Abalone
culturing success has been linked to a number of experi-
mental parameters including handling stress (Sales & Britz
2001), light intensity (Hahn 1989; Kim et al. 1997), water
quality (FAO, UNDP & Shallow Seafarming Institute 1990),
tank design and nutrition (Searle et al. 2006) and water
temperature (Britz et al. 1997). Frequent tank cleaning at
each feeding interval has also been attributed to survival
success with H. discus hannai (Mai et al. 1995b). Haliotis
discus hannai in this study were cultured within the recom-
mended ranges for this species (FAO, UNDP & Shallow
Seafarming Institute 1990) with the exception of water tem-
perature, which was maintained below the range outlined in
FAO, UNDP & Shallow Seafarming Institute (1990) but
above water temperature targets of previous studies (Mai
et al. 1995b; Park et al. 2008). The experimental system
utilized for this study entailed minimum handling of abalone
that were maintained in darkness during the growth trial.
Garcia-Esquivel et al. (2007) have demonstrated thatHaliotis
fulgens can be best cultured at 20 �C and 00 : 24 or 12 : 12
light : dark regimes.
Table 8 Comparative growth perfor-
mance of Haliotis discus hannai in
experimental growth trialsReference Feed type
Study
duration
(days)
Growth
(length)
(lm day)1)
Weight
gain
(% day)1)
Mai (1998) Artificial feed 100 – 0.74–0.88
Mai et al. (2001) Artificial feed 112 66.4–76.0 1.27–1.34
Tan & Mai (2001a) Artificial feed 120 38.66–59.60 0.39–0.49
Tan & Mai (2001b) Artificial feed 112 62.3–86.6 0.48–1.22
Tan et al. (2001) Artificial feed 112 67.4–86.9 0.76–1.24
Tan et al. (2002b) Artificial feed 112 36.87–55.07 0.37–0.58
Park et al. (2008)1 Undaria pinnatifida
Laminaria japonica
180 100.0 1.35
This study Laminaria digitata
KX feeds
84 58.0
22.0–26.0
0.56
0.14–0.23
KX, konjac glucomannan–xanthan gum.1 Commercial-scale experimental study.
Table 9 Mean survival rates (%) of Haliotis discus hannai in previous and the current on-growing studies
Reference
Study
duration
(days)
Length
(mm)
Weight
(g)
Stocking
parameters
(individuals tank)1)
Tank
volume
(cm3)
Survival
(%)
Mercer et al. (1993) 365 19.3 ± 0.10 0.817 ± 0.03 25 5301 84.0–95.0
Mai et al. (1995a) 100 0.389 20 10 000 87.5–98.3
Mai et al. (1995b) 100 – 0.378 ± 0.02 25 10 000 92.0–98.7
Mai (1998) 100 – 0.23 ± 0.01 30 10 000 86.7–96.7
Mai & Tan (2000) 112 16.11 ± 0.10 0.702 ± 0.02 25 8000 88.0–100.0
Mai et al. (2001) 112 10.92 ± 0.10 0.145 ± 0.001 40 19 600 85.8–94.2
Tan et al. (2001) 112 15.45 ± 0.03 0.62 ± 0.02 25 8000 94.7–100.0
Tan & Mai (2001a) 120 18.65 ± 0.18 1.18 ± 0.04 30 8000 75.0–83.3
Tan & Mai (2001b) 112 16.41 ± 0.04 0.74 ± 0.01 25 8000 94.7–100.0
Park et al. (2008)1 180 24.5 2.2 1667 192 · 104 94.5
This Study 84 25.89 ± 0.18 2.79 ± 0.06 15 8400 91.1–100.0
1 Commercial-scale experimental study.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

The aim of this study was to determine the most appropriate
feed binder for further KX feed development for juvenile
H. discus hannai. Based on the overall lack of a significant
difference between KX treatments in this study, further diet
development was based on the trends observed between
treatments. In terms of FCE, SGR, BW/SL ratio and sur-
vival, trends showed that the use of DW did not positively
benefit KX feed performance. There are subtle, although
non-significant, indications that a reduced K : X ratio was
beneficial to overall feed performance.
Based on the results of this study, further feed develop-
ment for abalone was conducted using the binder configu-
ration of Diet C (SW 2% KX; 1 : 1). Overall, the results of
this study indicated that feed and growth performance of
juvenile H. discus hannai was better with feeding fresh
L. digitata, and therefore an optimized diet formulation will
be utilized in the next phase of KX diet development for
juvenile H. discus hannai (O�Mahoney 2009).
The observed ability of the KX-produced feeds to retain
structural integrity in SW is an indication of the potential for
this binder in aquatic feed production where water quality
issues are paramount. Despite the good growth rates that
may be achieved with some existing commercially available
abalone feeds, we have observed that such feeds can create
severe problems with water quality maintenance. Such
aspects negate the positive benefit of these existing feeds for
commercial use. An all-encompassing sustainable feed
development programme requires consideration of the envi-
ronmental impact of the aquatic feeds in conjunction with
the input resources.
This study was conducted at the Aquaculture & Fisheries
Development Centre (AFDC), University College Cork. This
research was funded by Enterprise Ireland Commercializa-
tion Fund 2007–2008. The authors thank Chris O�Grady of
Brandon Bay Seafood�s Ltd for the supply of abalone.
Thanks also to the staff of the AFDC for technical assistance.
Thanks to Prof. Ed. Morris for comments on the manuscript
and also to the reviewers for insightful contributions. This
study is dedicated to James O�Mahoney.
Alvarez-Mancenido, F., Landin, M. & Martınez-Pacheco, R. (2008)
Konjac glucomannan/xanthan gum enzyme sensitive binary mix-
tures for colonic drug delivery. Eur. J. Pharm. Biopharm., 69, 573–
581.
Britz, P.J. (1996a) Effect of dietary protein level on growth perfor-
mance of South African abalone, Halitois midae, fed fishmeal-base
semi-purified diets. Aquaculture, 140, 55–61.
Britz, P.J. (1996b) The suitability of selected protein sources for
inclusion in formulated diets for the South African abalone,
Haliotis midae. Aquaculture, 140, 63–73.
Britz, P.J., Hecht, T. & Mangold, S. (1997) Effect of temperature on
growth, feed consumption and nutritional indices ofHaliotis midae
fed a formulated diet. Aquaculture, 152, 191–203.
Caltagirone, A., Francour, P. & Fernandez, C. (1992) Formulation
of an artificial diet for the rearing of the urchin Paracentrotus
lividus: I. Comparison of different binding agents. In: Echinoderm
Research (Scalera-Liaci, L. et al. eds.), pp. 115–119. A.A. Balk-
ema, Rotterdam.
Coote, T.A., Hone, P.W., Kenyon, R. & Maguire, G.B. (1996) The
effect of different combinations of dietary calcium and phospho-
rous on the growth of juvenile Haliotis laevigata. Aquaculture, 145,
267–279.
Coote, T.A., Hone, P.W., Van Barneveld, R.J. & Maguire, G.B.
(2000) Optimal protein level in a semipurified diet for juvenile
greenlip abalone Haliotis laevigata. Aquac. Nutr., 6, 213–220.
Day, R.W. & Cook, P. (1995) Bias towards brown algae in deter-
mining diet and food preferences: the South African abalone
Haliotis midae. Mar. Freshw. Res., 46, 623–627.
De Muylder, E., Hage, H. & van der Velden, G. (2008) Binders:
gelatin as alternative for urea formaldehyde and wheat gluten in
the production of water stable shrimp feeds. Aquafeed Int., 11,
10–13.
Dea, I.C.M. (1993) Conformational origins of polysaccharide solu-
tion and gel properties. In: Industrial Gums (Whistler, R.L. &
BeMiller, J.N. eds.), pp. 21–52. Academic Press, London.
Eyo-okon, I.E. & Hilton, C.S. (2003) Use of Glucomannan Hydro-
colloid as Filler Material in Prostheses. Konjac Technologies, LLC,
South River, NJ. United States Patent 6537318.
Fagbenro, O. & Jauncey, K. (1995) Water stability, nutrient leaching
and nutritional properties of moist fermented fish silage diets.
Aquac. Eng., 14, 143–153.
Fan, J., Wang, K., Liu, M. & He, Z. (2008) In vitro evaluations of
konjac glucomannan and xanthan gum mixture as the sustained
release material of matrix tablet. Carbohydr. Polym., 73, 241–247.
FAO (2009) Fishery and Aquaculture Statistics 1950–2007. FAO,
Rome. http://www.fao.org/fishery/statistics/en. last accessed on
September 5, 2009).
FAO, UNDP & Shallow Seafarming Institute (1990) Training man-
ual on artificial breeding of abalone (Haliotis discus hannai) in
Korea D.P.R. In FAO/UNDP Regional Seafarming Development
and Demonstration Project, RAS/90/002, pp. 107. TrainingManual,
No. 7. FAO, UNDP & Shallow Seafarming Institute, Bangkok.
Fermin, A.C. (2003) Effects of alternate starvation and refeeding
cycles on food consumption and compensatory growth of abalone,
Haliotis asinina (Linnaeus). Aquac. Res., 33, 197–202.
Fleming, A.E. (1995) Growth, intake, feed conversion efficiency and
chemosensory preference of the Australian abalone,Haliotis rubra.
Aquaculture, 132, 297–311.
Fleming, A.E., Van Barneveld, R.J. & Hone, P.W. (1996) The
development of artificial diets for abalone: a review and future
directions. Aquaculture, 140, 5–53.
Garcia-Esquivel, Z. & Felbeck, H. (2006) Activity of digestive
enzymes along the gut of juvenile red abalone, Haliotis rufescens,
fed natural and balanced diets. Aquaculture, 261, 615–625.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

Garcia-Esquivel, Z. & Felbeck, H. (2008) Comparative performance
of juvenile red abalone, Haliotis rufescens, reared in laboratory
with fresh kelp and balanced diets. Aquac. Nutr., 15, 209–217.
Garcia-Esquivel, Z., Montes-Magallon, S. & Gonzalez-Gomez,
M.A. (2007) Effect of temperature and photoperiod on the growth,
feed consumption, and biochemical content of juvenile green
abalone, Haliotis fulgens, fed on a balanced diet. Aquaculture, 262,
129–141.
Gorfine, H.K. (1991) An Artificial Diet for Hatchery-reared Abalone
Haliotis rubra. Marine Science Laboratories, International Report
No. 190, Queenscliff, Australia.
Goycoolea, F.M., Richardson, R.K., Morris, E.R. & Gidley, M.J.
(1995) Stoichiometry and conformation of xanthan in synergistic
gelation with locust bean gum or konjac glucomannan. Macro-
molecules, 28, 8308–8320.
Guzman, J.M. & Viana, M.T. (1998) Growth of abalone Haliotis
fulgens fed diets with and without fish meal, compared to a com-
mercial diet. Aquaculture, 165, 321–331.
Hahn, K.O. (1989) Handbook of Culture of Abalone and Other
Marine Gastropods, pp. 368. CRC Press, FL.
Harada, K., Miyasaki, T., Kawashima, S. & Shiota, H. (1996)
Studies on the feeding attractants for fishes and shellfishes. XXVI.
Probable feeding attractants in allspice Pimenta officinalis for
black abalone Haliotis discus. Aquaculture, 140, 99–108.
Hession, C., Guiry, M.D., McGarvey, S. & Joyce, J. (1998) Mapping
and Assessment of the Seaweed Resources (Ascophyllum nodosum,
Laminaria spp.) off the West Coast of Ireland, pp. 1–89. Marine
Resource Series, No. 5, Marine Institute, Galway, Ireland.
Kato, K. & Matsuda, K. (1969) Studies on the chemical structure of
konjac mannan. Part I. Isolation and characterization of oligo-
saccharides from the partial acid hydrolyzate of mannan. Agric.
Biol. Chem., 33, 1446–1453.
Kelco (2007) Keltrol/Kelzan Xanthan Gum Book, pp. 32. CP Kelco,
Atlanta, GA.
Kelly, E. ed. (2005) The Role of Kelp in the Marine Environment. Irish
Wildlife Manuals No. 17, National Parks and Wildlife Service,
Department of Environment, Heritage and Local Government,
Dublin, Ireland.
Kim, B.L., Kim, J.W., Wom, S.H., Wi, C.H. & Park, H.Y. (1997)
Effects of complete dark conditions on the growth of four
species of juvenile abalones. Bull. Nat. Fish. Dev. Res. Inst., 53,
103–110.
Knauer, J., Britz, P.J. & Hecht, T. (1993) The effect of seven binding
agents on 24-hour water stability of an artificial weaning diet for
the South African abalone, Haliotis midae (Haliotidae, Gastro-
poda). Aquaculture, 115, 237–334.
Lee, S.-M. (2004) Utilization of dietary protein, lipid, and carbo-
hydrate by abalone Haliotis discus hannai: a review. J. Shellfish
Res., 23, 1027–1030.
Liu, F., Ai, Q., Mai, K., Tan, B., Ma, H., Xu, W., Zhang, W. &
LiuFu, Z. (2008) Effects of Dietary Binders on Survival and
Growth Performance of Postlarval Tongue Sole, Cynoglossus
semilaevis (Gunther). J. World Aquac. Soc., 39, 500–509.
Macey, B.M. & Coyne, V.E. (2005) Improved growth rate and
disease resistance in farmed Haliotis midae through probiotic
treatment. Aquaculture, 245, 249–261.
Mai, K. (1998) Comparative studies on the nutrition of two species
of abalone, Haliotis tuberculata L. and H. discus hannai Ino. VII.
Effects of dietary vitamin C on survival, growth and tissue con-
centration of ascorbic acid. Aquaculture, 161, 383–392.
Mai, K. & Tan, B. (2000) Iron methionine (FeMet) and iron sulfate
(FeSO4) as sources of dietary iron for juvenile abalone, Haliotis
discus hannai. J. Shellfish Res., 19, 861–868.
Mai, K., Mercer, J.P. & Donlon, J. (1994) Comparative studies on
the nutrition of two species of abalone, Haliotis tuberculata L. and
H. discus hannai Ino. II. Amino acid composition of abalone and
six species of macroalgae with an assessment of their nutritional
value. Aquaculture, 128, 115–130.
Mai, K., Mercer, J.P. & Donlon, J. (1995a) Comparative studies on
the nutrition of two species of abalone, Haliotis tuberculata L. and
H. discus hannai Ino. Aquaculture, 134, 65–80.
Mai, K., Mercer, J.P. & Donlon, J. (1995b) Comparative studies on
the nutrition of two species of abalone, Haliotis tuberculata L. and
H. discus hannai Ino. IV. Optimum dietary protein level for
growth. Aquaculture, 136, 165–180.
Mai, K., Mercer, J.P. & Donlon, J. (1996) Comparative studies on
the nutrition of two species of abalone, Haliotis tuberculata L. and
H. discus hannai Ino. V. The role of polyunsaturated fatty acids of
macroalgae in abalone nutrition. Aquaculture, 139, 77–89.
Mai, K., Wu, G. & Zhu, W. (2001) Abalone, Haliotis discus hannai
Ino, can synthesis myo-inositol de novo to meet physiological
needs. J. Nutr., 131, 2898–2903.
Melton, L.D., Mindt, L. & Rees, D.A. (1976) Covalent structure of
the extracellular polysaccharide from Xanthomonas campestris:
evidence from partial hydrolysis studies. Carbohydr. Res., 46, 245–
257.
Mercer, J.P., Mai, K. & Donlon, J. (1993) Comparative studies on
the nutrition of two species of abalone, Haliotis tuberculata L. and
H. discus hannai Ino. I. Effects of algal diet on growth and bio-
chemical composition. Invertebr. Reprod. Dev., 23, 75–88.
Morris, E.R., Rees, D.A., Young, G., Walkinshaw, M.D. & Darke,
A. (1977) Order-disorder transition for a bacterial polysaccharide
in solution. A role for polysaccharide conformation in recognition
between Xanthomonas pathogen and its plant host. J. Mol. Biol.,
110, 1–16.
Mortensen, A., Siikavuopio, S.I. & Raa, J. (2003) Use of transglu-
taminase to produce a stable sea urchin feed. In: Sea Urchin
Fisheries and Ecology (Lawrence, J.M. & Guzman, O. eds.),
pp. 203–213. Destech Publications Inc., Puerto Varas, Chile.
Naidoo, K., Maneveldt, G., Ruck, K. & Bolton, J.J. (2006) A
comparison of various seaweed-based diets and formulated feed on
growth rate of abalone in a land-based aquaculture system.
J. Appl. Phycol., 18, 437–443.
Obaldo, L.G., Dominy, W.G., Terpstra, J.H., Cody, J.J. & Behnke,
K.C. (1999) The impact of ingredient particle size on shrimp feed.
J. Appl. Aquac., 8, 55–67.
O�Mahoney, M. (2009) Development of a Novel Binder for Aquacul-
ture: application of a Konjac glucomannan-xanthan Gum Binder to
Formulated Feed Development for the Sea Urchin Paracentrotus
lividus and abalone Haliotis discus hannai, pp. 297. PhD Thesis,
National University of Ireland, Cork.
Palma, J., Bureau, D.P. & Andrade, J.P. (2008) Effects of binder type
and binder addition on the growth of juvenile Palaemonetes vari-
ans and Palaemon elegans (Crustacea: Palaemonidae). Aquac. Int.,
16, 427–436.
Park, J., Kim, P.-K. & Jo, J.-Y. (2008) Growth performance of disk
abalone Haliotis discus hannai in pilot- and commercial-scale
recirculating aquaculture systems. Aquac. Int., 16, 191–202.
Pearce, C.M., Daggett, T.L. & Robinson, S.M.C. (2002) Effect of
binder type and concentration on prepared feed stability and
gonad yield and quality of the green sea urchin, Strongylocentrotus
droebachiensis. Aquaculture, 205, 301–323.
Pereira, L., Riquelme, T. & Hosokawa, H. (2007) Effect of three
photoperiod regimes on the growth and mortality of the Japa-
nese abalone Haliotis discus hannai Ino. J. Shellfish Res., 26,
763–767.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd

Rivero, L.E. & Viana, M.T. (1996) Effect of pH, water stability and
toughness of artificial diets on the palatability for juvenile abalone
Haliotis fulgens. Aquaculture, 144, 353–362.
Ruscoe, I.M., Jones, C.M., Jones, P.L. & Caley, P. (2005) The
effects of various binders and moisture content on pellet stability
of research diets for freshwater crayfish. Aquac. Nutr., 11,
87–93.
Sales, J. & Britz, P.J. (2001) Research on abalone (Halitois midae L.)
cultivation in South Africa. Aquac. Res., 32, 863–874.
Sales, J. & Britz, P.J. (2002a) Evaluation of the reference diet sub-
stitution method for determination of apparent nutrient digest-
ibility coefficients of feed ingredients for South African abalone
(Haliotis midae L.). Aquaculture, 207, 113–123.
Sales, J. & Britz, P.J. (2002b) Influence of ingredient particle size and
inclusion level of pre-gelatinised maize starch on apparent digest-
ibility coefficients of diets in South African abalone (Haliotis midae
L.). Aquaculture, 212, 299–309.
Sales, J., Truter, P.J. & Britz, P.J. (2003) Optimum dietary crude
protein level for growth in South African abalone (Haliotis midae
L.). Aquac. Nutr., 9, 85–89.
Searle, T., Roberts, R.D. & Lokman, P.M. (2006) Effects of tem-
perature on growth of juvenile blackfoot abalone, Haliotis iris
Gmelin. Aquac. Res., 37, 1441–1449.
Shipton, T.A. & Britz, P.J. (2001) The effect of animal size on the
ability of Haliotis midae L. to utilise selected dietary protein
sources. Aquac. Res., 32, 393–403.
Storebakken, T. & Austreng, E. (1987) Binders in fish feeds. II: effect
of different alginates on the digestibility of macronutrients in
rainbow trout. Aquaculture, 60, 121–131.
Tan, B. & Mai, K. (2001a) Effect of dietary vitamin K on survival,
growth and tissue concentrations of phylloquinone (PK) and
menaquinone-4 (MK-4) for juvenile abalone, Haliotis discus
hannai Ino. J. Exp. Mar. Biol. Ecol., 256, 229–239.
Tan, B. & Mai, K. (2001b) Zinc methionine and zinc sulfate as
sources of dietary zinc for juvenile abalone, Haliotis discus hannai
Ino. Aquaculture, 192, 67–84.
Tan, B., Mai, K. & Liufu, Z. (2001) Response of juvenile abalone,
Haliotis discus hannai, to dietary calcium, phosphorous and
calcium/phosphorous ratio. Aquaculture, 198, 141–158.
Tan, B., Mai, K. & Liufu, Z. (2002a) Dietary phosphorous
requirement of juvenile abalone, H. discus hannai Ino. Chin. J.
Oceanol. Limnol., 20, 22–31.
Tan, B., Mai, K. & Xu, W. (2002b) Availability of phosphorous
from selected inorganic phosphates to juvenile abalone, Haliotis
discus hannai Ino. Chin. J. Oceanol. Limnol., 20, 118–128.
Tanaka, R., Sugimura, I., Sawabe, T., Yoshimizu, M. & Ezura, Y.
(2003) Gut microflora of abalone Haliotis discus hannai in culture
changes coincident with a change in diet. Fish. Sci., 69, 951–958.
Troell, M., Robertson-Andersson, D., Anderson, R.J., Bolton, J.J.,
Maneveldt, G., Halling, C. & Probyn, T. (2006) Abalone farming
in South Africa: an overview with perspectives on kelp resources,
abalone feed, potential for on-farm seaweed production and socio-
economic importance. Aquaculture, 257, 266–281.
Tutschulte, T.C. & Connell, J.H. (1988) Feeding behavior and algal
food of three species of abalones (Haliotis) in southern California.
Mar. Ecol. Prog. Ser., 49, 57–64.
Uki, N., Kemuyama, A. & Watanabe, T. (1985) Development of
semipurified test diets for abalone. Bull. Japanese Soc. Sci. Fish.,
15, 1825–1833.
Werner, A. & Kraan, S. (2004) Review of the Potential Mechanisation
of Kelp Harvesting in Ireland. Marine Environment and Health
Series, No. 17, pp. 52. Marine Institute, Galway, Ireland.
Zhang, W., Mai, K., Xu, W., Liufu, Z., Tan, B., Ai, Q., Ma, H. &
Wang, X. (2008) Effects of dietary guaiacol on shell biomineral-
ization of juvenile abaloneHaliotis discus hannai, Ino. Aquac. Res.,
39, 954–961.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Aquaculture Nutrition � 2011 Blackwell Publishing Ltd
Recommended

















