
i
ALTERNATIVE SPLICING AND ADHESION PROPERTIES
OF A MOUSB CARCINOEMBRYONIC ANTIGEN GENE
FAMILY MEMBRR
Kimberly NcCuaig e
Department of Medicine, Division of
Experimental Medicine,
McGill University, Montreal, Quebec, Canada
April 1992
A thesis submitted t0 the Faculty of Graduate Studies and
Research in partial fulfillment of the requirements for
the degree of Master of Science.

Il
ABSTRACT
Carcinoembryonic antigen (CEA) is a tumor Illarker userl
clinically to assess post-operative recurrences of ln'east,
lung and colon cancers. The CEA gene famiIy, which j s part of
the immunoglobulin superfamily, is composed of several
proteins crossreative with anti-CEA antibodies.
Carcinoembryonic antigen appears ta function
development of the gastro-intestinal tract as weIl
in the
in tumor
formation; it is capable of mediating ceII-cell adhesion in
vitro which is consistent with its putative role of
maintaining tissue architecture in vivo. CEA gene fami Iy
members have also been identified in various tissues of the
mouse. Two of the mouse proteins, mmCGMla and mmCGMlb, have
been characterized. By sequence homology, rnrnCGMla and rnrnCGMlb
are the mouse homologues of human biliary glycoprotein and o[
rat hepatocyte ecto-ATPase. Both of the mouse CEA relaterl
proteins function as adhesion molecules when expressed on the
cell surface of transfectant cells; however, mmCGMld, unlike
mmCGMlb, mediates cellular aggregation irrespect ive of calcIum
concentration or temperature. Sequence comparison of rrunCGMla 1
mmCGMlb, and other cDNAs isolated by polymerase chain react ion
techniques, demonstrates that there are at least eight
possible transcripts encoding CEA-related proteins and that
these transcripts are aIl produced by alternative spI icinq of
one precursor messenger RNA.

111
RÉSUMÉ
L'antigène carcino-embryonnaire (ACE) est un marqueur de
tumeur employé en clinique pour évaluer les récurrences
postopératoires de cancer du sein, du poumon, et du côlon. La
fami Ile des gènes reliés à l'ACE, incluse dans Id superfamille
des immunoglobulines, code pour plusieurs protéines réagissant
avec des ant1corps dirigés contre l'ACE. L'antigène carcino
embryonnaire semble participer au développement du système
digestif ainsi qu'à la formation de tumeurs. Il sert
cl' intermédiaire lors d'adhésion intercellulaire in vitro,
réflétant bien son rôle présumé dans le maintien de
l'organisation tissulaire. Des membres de la famille de l'ACE
ont aussi été identifiés dans divers tissus chez ld souris.
Deux de ces
caractérisées.
protéines, rnmCGMla et mrnCGMlb, ont été
L'ana]yse de leur séquence fait d'elles les
homologues murins de la glycoprotéine biliaire humaine et de
l'ecto-ATPdse d' hepatocyte de rat. Elles agissent toutes deux
comme molécules d'adhesion lorsqu'exprimées à la surface de
celluJ es transfectantes, mais rnrnCGMla contrairement à mmCGMlb,
le fait indépendamment de la température d'incubation et de la
concent rat ion de calcium. La comparaison des séquences de
IlunCGMla, nunCGMlb, et d'autres ADN complémentaires isolés,
d~montre qu'au moins huit transcrits peuvent coder pour des
protéines reliées à l'ACE et que ces transcrits résultent tous
d'épissage alternatif d'un seul ARN messager précurseur.

..
ABSTRACT (English)
RESUME (french)
TABLE OF CONTENTS
LIST OF ABBREVIATIONS
ACKNOWLEDGEMENTS
PREFACE
INTRODUCTION
TABLE OF CONTENTS
Identification of CEA
Turnor Markers
Role of CEA in Cancer and Metastélsis
CEA SulJgroup Proteins
Pregnancy-Specific Glycoproteins
Gene Cluster Organization
Transcripts of the CEA Subgroup
protein Structure of CEA Subgroup Members
Cell Adhesion Molecules
Cadherins
Neural Cell Adhesion Molecule
Carcinoembryonic Antigen
Mouse Homologues of Human CEA Family Member s
Functional Analysis of Murine CEA
References
rrunCGMla: A MOUSE CARCINOEMBRYONIC ANTIGEN GENE FAMILY
MEMBER, GENERATED BY ALTERNATIVE SPLICING, FUNCTIOnS
iv
l l
1 l 1
IV
VIl
lX
X
4
7
H
9
1 1
12
13
14
le
lH

, 1
v
AS AN ADHESION MOLECULE.
Abst ract 37
Int roduct ion 38
Results
Characterlzation of the cDNA clones and
Comparison with other CEA Gene family members 40
Expression of mouse mmCGMla transcripts 43
Expression of the rnrnCGMla protein 45
mmCGM1a as a cell adhesion molecule 46
Discussion 48
Mat erials and Methods
Cell culture and transfections
Isolation of murine CEA cDNA clones
Reverse transcription and polymerase chain
ampl i fication
DNA sequence determination and analyses
\)P-labelled probes
RNA preparation and Northern analyses
Protein purification and generation of
antibodies
Western analyses
Immunoprecipitation
Immunofluorescence
Aggregation assays
Acknowledgements
References
53
53
54
55
55
55
56
58
59
59
60
61
62

vi
Legend to Figures
Fig.1 Restriction map and topology of nunl'UMlcl 74
Fig.2 Hornology of mmCGMla and nunCGMlb wi th
rat ecto ATPase and Human BGP
Fig.3 Expression of rnrnCGMla transcripts
in rnurine tissues
Fig.4 protein Expression
Fig.5 Adhesion Assays
Figures
DISCUSSION AND CONCLUSION
Structure and Alternative Splicing
Adhesion Molecule
Role in Turnor Formation
Homologues in the Rat
Blastocyst Implantation
Signal Transduction
Calmodulin Binding Protein
Virus Receptor
Conclusion
References
74
7S
'/ b
'1'/
'/ H
83
84
85
H9
H9
92
94

Al
A2
BI
BGP
CalI
CAM
cDNA
CGM
CEA
CML
DNA
EGF
FGF
19
1GF
kb
kDa
MHV
ml
MLCK
nunCGM
mRNA
N
NCA
LIST OF ABREVIATIONS
first halE of first inteInal Ig domain
second internal half 19 domain
second half of first internal 19 domain
biliary glycoprotein
calcium
cell adhesion molecule
complementary DNA
carcinoembryonic antigen gene family member
carcinoembryonic antigen
chronic myeloid leukemia
deoxyribonucleic acid
epidermal growth factor
fibroblast growth factor
immunoglûbulin
insulin-like growth factor
kilobase
kilodalton
mouse hepatitis virus
milliliter
myosin light chain kinase
Mus musculus CEA gene family member
messengér RNA
amino
pnnspGcific crossreacting antigen
vii

N-CAM
ng
p.C.
peR
pp120
PSG
RNA
snRNP
TSA
UT
neural cell adhesion molecule
nanogram
post. coitum
polymerase chain reaction
phosphoprotein of molecular weight 120 kDa
pregnancy-specific glycoproteln
ribonucleic acid
small nuclear ribonucleotide particles
tumor specifie antigen
untranslated sequence
VII l

IX
ACKNOWLEDGEMENTS
l would like ta acknawledge the following people for
thclr contrIbutions to my Master' s thesis: my supervisor Dr.
NIcol e BeauchemIn for her advice and technical help, as weIl
as [or preparation of the manuscript for publication and
(;n t ica l readlng of this thesis, Claire Turbide for technical
help wlth Western analyses, her contributions to the paper, as
well dS her help ln translat ing the abstract to French, Dr.
Mark Featherstone and Dr. Ron Baldassare for a cri tical
reading of this thesis.
l would also like to thank The Cancer Research Society
dnd 'J'he Medical Research Counci l of Canada for their support.

x
PREFACE
Chapter 2 of this dissertdt iO!1 was tmbmi l t- ('d fOI
publ;catlon July 19, 1991, and following sorne InlnOr rpvlsiom"
was accepted and published in Cell Growth and Dlffen--'nt"iat 10n,
vol. 3 , 165-174, March 1992. A note was added III proof to
acknowledge that, after the submission of this ['aper 1 tlH' cDNA
sequence of the mouse hepatitis VLLUS receptor l'Vas publ i [;lwd
(G.S. Dveksler et al., J. Viral., 65: 6881-6891, 1991). 'NilS
sequence is identical to that of the coding region of mmCC;M la.
Both the 5' and 3' untranslated regions are di f [ercmt f rom
those of the mmCGMla cDNA. However, the 3 ' unt l'dns lated
regions of both the hepatitis virus receptor and the mmCCMlb
are identical up to the first polyadenylat ion l:om;ennUf)
sequence, therefore reinforcing the statement thdt dl terndt ive·
splicing and alternate usage of pOlyadenylation signaIs
generate many transcripts from a single gene.
The submission of this paper wouid not have been posslblp
on my own. RNA isolation from mouse tissues, the pur j EiCd t j on
and preparation of the polyclonal antibody uned tu eVr..Iludt(!
the production of mrnCGMla protein by trdnsfectant c(~lln 1 t h(~
immunof luorescence assays 1 and the immunoprec ipi la t ion
experiments were done by Claire Turbide. The construct- ion of
the mouse colon cDNA library wh; ch was screened 1 and the
original screening using a human CEA cDNA prohe, was done hy
Nicole Beauchemin. The other work presented in Un 0 pdper was

(
Xl.
done hy myself. These experiments include the purification of
partial cDNA clones, reconstruction of a cDNA encoding the
full length mmCGMla protein, Northern blot analyses, PCR
analyses, sequencing, transfection of the rrunCGMla cDNA into
NIH 3T3 cells, evaluation of transfectant clones by Northern
and Wester~ analyses, anè the adhesion assays. The cioning
dnd sequencing of other alternatively spliced cDNAs from mouse
colon RNA, uSlng PCR techniques, as described in the
discussion, was aiso done on my own.

INTRODUCTION

1
INTRODUCTION
Identification of CHA:
Carcinoembryonic Antigen (CEA) was originally described
by Gold and Freedman in 1965 as a glycoprotein specific for
malignant turnors of endodermally derived tissues (Gold and
Freedman, .1965). The isolation of CEA was based on the theory
that turnor cells bear specifie antigens (TSAs) not found on
normal cells. An extract of hurnan tumor was inj ected into
animaIs with the assumption that the resuiting antiserum would
contain antibodies against tumor specifie antigens as weIl as
against normal hurnan tissue. The antibodies raised against
normal tissue could then be titrated out leaving an antiserum
recognizing tumor specifie antigens only.
Tumor Markers:
Turnor markers can potentially be used in screening for
cancer, either in the initial diagnosis and prognosis, or in
the assessment of the effectiveness of therapy (Gold and
Shuster, 1981). Many tl.lmor cells secrete substances into the
circulation which may reflect the state of the tumor, either
quantitatively or qualitatively.
In most cases, cancers are sufficiently differentiated to
permit the identification of the tissue of origin by routinE:'
microscopie sections. However, frequently tumor cells are
( poorly differentiated, and establishing the origin of a

2
metastasis becomes difficult. Since tumors wi th the same
origin often express the same tumor markers, these antigens
are useful in cellular identification by rddio-inununodssay
techniques (Shuster et al., 1980). An aeeurate morphologie
diagnosis is important due to the development of specifie
radiotherapeutic and chemotherapeutic protocols used for
particular cancers.
Tumor markers may be normal tissue constituents
inappropriately expressed, or may be tumor speeiflc antigens
which are never expressed at any time on normal cells (Shuster
et al., 1980). Tumor markers of the first type ean be
subdivided into groups (Robbins et al., 1984):
1) Products normally produced by plasma celis or
endocrine glands are expressed at elevated leveis. Examples
are: human chorionic gonadotropin in choriocarcinomas,
hydatidiform mole, seminoma, teratocarcinoma of the testis,
and ovarian carcinoma; human placental lactogen in
trophorlastic neoplasms, and calcitonin in thyroid medullary
carcinoma.
2) Ectopie production of hormones by tumors of
nonendocrine origin (paraneoplastic syndromes). For examp le,
the production of erythropoietin by renal cell carClnomas.
3) Change in production of enzymes such as acid
phosphatase in prostatic carcinoma and galactosyltransferase
II in nancreatic, gastric, and breast carcinomas.
4) Production of oncofetal antigens. These are proteins

3
normally expressed during embryonic or fetal development but
not present in adult tissues. Examples are: Carcinoembryonic
Antigen in colonie, pancreatic, bronchogenic, gastric, and
breast carcinomas, Alfa-fetoprotein in hepatocellular
carcinoma and germ cell tumors, and pancreatic oncofetal
antigen in pancreatic, bronchogenic, and gastric carcinomas.
Since all of these mo1ecules are norrnal1y expressed at
one time in the host, no immune response can be generated
against them. The hast is, however, capable of mounting an
anti-tumor immune response, indicating the presence of
specifie tumor antigens not seen prevl0us1y by the hest
(Shuster et al., 1980).
The second type of turnor rnarkers elicit an immune
response since they are foreign to the hosto Viral antigens
and proteins generated by gene rearrangements belong to this
class of tumor marker. These antigens ref1ect the
individua1ity of each tumor; each chemica1 carcinogen-induced
tumor expresses its own unique TSAs, while aIl tumors induced
by the same virus share the same TSAs. Detection of an
antiturnor immune response, in either case is useful in
diagnosing and managing patients with malignancy without
isolating the specifie turnor antigens (Shuster et al., 1980).
The possibility of using turnor markers in precise
localization and destruction of maligna.nt cells is under
investigation. Radiolabelling antibodies against tumor
mdrkers would specifically identify cancer cells, a110wing

1 4
localizat ion of primary and metastat ic tumors wi t Il
radioscanning techniques (Goldenberg et al., 1978). Also, if
antibodies of sufficient affinity and specificity were
developed, coupling them to antitumor agents would bring the
therapeutic agent directly to the site of action, sparing
normal tissues from destruction.
Role of CHA in Cancer and Netastasis:
At first, the usefulness of CEA as a tumor marker for
colon cancer looked promising. However, CEA or CEA-like
proteins have been detected in many tissues and in body fluids
of patients with other tumor types or without malignant
disease (Neville and Caurence 1974). It became apparent that
if CEA was to be used as a tumor marker, purificatlon and
characterization of these cross-reacting proteins was
essential.
Since then, evidence that CEA exists ln normal colon
tissue has been reported (Fritsche and Mach, 1978). However,
CEA mRNA levels are 6-10 times higher in colon carcinomas, and
corresponding protein levels are 2-100 times higher (Boucher
et al., 1989). No correlation exists between these mRNA and
protein levels. Gene rearrangement or amplification is not
evident, however, demethylation may be cl factor (Tran et al.,
1988; Boucher et al., 1989). These observations implicate
regulatory control of transcription as weIl as post
transcriptional mechanisms.

5
The seriaI CEA concentration in sera of cancer patients
(gastro-intestinal, breast, and lung cancer) has proven useful
in monitoring post-operative recurrences (Gold and Shuster
1981). AIso, patients exhibiting serum CEA levels exceeding
5 ng/ml preoperatively, have a poorer prognosis than those
having lower levels (Wanebo et al., 1978). Recently, Jessup
and Thomas (Jessup et al., 1988) have shown that 81% of
carcinomas expressing high levels of CEA were tumorigenic in
nude mice compared to 54 % of carcinomas expressing low levels.
Since 80% of patients who die of colorectal carcinomas
bear li ver metastases (Willis, 1973), the role of CEA in
metastasis was examined. CEA may be involved in several steps
of metastasis including the detachment of cells from the
primar.y tumor mass, the survival of tumor cells in the
circulation, and the implantation and invasion into secondary
sites. Redistribution of CEA on the cell surface, leads ta a
loss of polarity of tumor cells, and disrupts the regular
organization of colon tissue. This change in tissue
archi tecture affects intercell ular contacts, allowing tumor
cells to dissociate and disseminate intravascularly (Benchimol
et al., 1989). Once in the circulation, CEA may inhibit host
defense mechanisms by directly inhibiting lymphocyte function
or by lnducing lmmunosuppressi ve factors (Medoff et al., 1988;
Hakim, 1984). After entering the circulation, malignant cells
arres t in a capi llary bed or organ, implant, and invade
through the basement membrane into the parenchyma of the

(,
organ. The presence of CEA enhances the attdchment of
circulating cells, allowing them to implant. CEA injected
intravenously into nude mice causes both Kupffer cells and
hepatocytes to display CEA on their cell surface, resulting in
increased colony formation of a poorly metastatic colorectal
carcinoma ceilline in the liver (Hostetter et al. 1990).
Interestingly, this cellular attachment can be competitively
inhibited by a short peptide derived from a region of CEA
which has homology with protease sequences (Thomas et al.,
1991) . This homology suggests that an endogenous CEA
proteolytic activity may act to facilitate invasion.
CHA Subgroup Glycoproteins:
The CEA gene family can be divided into two subgroups.
The CEA subgroup genes encode CEA and its classical
crossreacting antigens, while the second gene subgroup
contains the pregnancy-specific glycoproteins (PSG) (Barnett
and Zimmermann, 1990). Two major crossreacting proteins in
the CEA subgroup have been identified through cross-reactivity
with anti-CEA antibodies. Non-specifie ~ross-reacting Antigen
(NCA) was first described in normal lung and spleen by von
Kleist et al. in 1972. ThIs protein was named NCA because lt
was found to be neither organ nor tumor specifie. Norrna 1
colon tissue, granulocytes, as weIl as breast carcinoffids dnd
leukocytes of CML patients, also conta in NCA (Cournoyer et
al., 1988).

7
A second cross-reactive protein identified was bilia1Y
glycoprotein (BGP). Its isolation was based on the
observation that a large number of patients with non
carcinomatous biliary obstruction had false positive CEA serum
tests (Moore et al., 1972). Svenberg, using polyclonal anti
CEA antibodies, purified a CEA-like protein, BGP1, from normal
hepatic bile (Svenberg, 1976). He also discovered that bile
from patients with an obstruction did not contain BGP1, but
had instead, two other proteins that he named BGPII and BGPIII
(Svenberg, 1976). BGP is also expressed in colon,
granulocytes, leukocytes, and in many tumor types such as
colon, breast, pancreas, gastric, hepatic and bladder
carcinomas (Hinoda et al., 1990; Kuroki et al., 1991a and
1991b) .
pregnancy-Specific Glycoproteins:
The second subgroup of the CEA gene family, the PSGs,
were ini tially labelled pregnancy-specific because they could
only be identified in pregnant women. The initial isolation
of PSG from placenta led investigators to believe it
const ituted a single protein. However, further analyses
revealed a number of related proteins differing in molecular
weight, electrophoretic mobility, and antigenicity (Sorensen,
1984) . However, these proteins possess common antigenic
determinants. Placental syncytiotrophoblasts constitute the
major production sites of PSGs (Bischof, 1984) .

8
Extraplacental production has been noted ln the testis,
intestine, uterus, submanèibular gland, fetal l ivt=>l" ,
hematopoeitic cells and bone marrow (Khan et al., 1989; Khall
and Harrunarstrom, 1989 and 1990; zinunermann et al., 1989;
Borjigin et al., 1990; Shupert et al., 1990; Tease and Chan,
unpublished data; Zoubir et al, 1990; Bischof, 1984).
Normal, non-pregnant women produce PSGs ln very low
levels. The PSG serum levels are greatly lncreased before
implantation and progressively rlse throughout pre~nancy
(Bischof, 1984). Lower than normal levels of PSGs correlate
with increased risk of abortion (Ho et al.,
possibility of ectopie pregnancy (Bischof,
proteins are also important in identifying
1988), and thp.
1984). These
f et al syndromes
such as Meckel's syndrome (Heikinheimo et al., 1982), Down's
syndrome (Bartels and Lindrnann, 1988), and fetal alcohoJ
syndrome (Heikinheimo et al., 1987), as weIl as in ident ifYlng
gestational trophoblastic tumors (Takayama et al., 1987) and
breast cancer (Bischof et al., 1984).
Gene Cluster Organization:
In humans, approximately 20 genes coding for CEA family
members are clustered on the long arm of chromosome 19, trom
19q13.1 to 19q13.3 (Zimmermann et al., 1988; Schonk et al.,
1990). Nine genes belong to the CEA subgroup and 13 genes to
the PSG subgroup. The CEA genes are in a tandem array,
separated by 12 kb (Thompson et al., 1990; Leslie et al.,

9
1990), while the PSG genes are more closely linked, being
separated by only 6 kb. The order of the genes on the
chromosome is CGM7, CGM2, CEA, NCA, and CGM1 closest to the
centroml're, BGP, CGM6, and CGM8 in the middle, and the PSG
c1uster closest to the telomere (Thompson et al., 1991).
C10ning and sequencing of the upstream regions of CEA, NCA
(Schrewe et al., 1990; Willcocks and Craig, 1990), and two
PSG (Watanabe and Chou, 1988; Thompson et al., 1990) subgroup
members, reveals the absence of 'rATA or CMT-box consensus
sequences. Transcriptional regulation must occur through
other elements not yet identified. The close linkage of the
PSG cluster may indicate coordinated expression through use of
common regulatory elements (Thompson et al., 1990; Leslie et
al., 1990). Relative position on the chromosome may also be
elemental in regulating CEA and NCA genes, since they are
almost always co-expressed (Cournoyer et al., 1988; Sato et
al., 1988: Boucher et al., 1989).
Transcripts of the CHA subgroup:
Complementary DNAs (cDNAs) for CEA, NCA, and BGP1 have
been isolated and sequenced (Beauchemin et al., 1987;
Neumaier et al., 1988: Oikawa et al., 1987; Barnett et al.,
1988; Tawaragi et al., 1988: Hinoda et al., 1988: Barnett
et al., 1989). A high degree of sequence homology exists
bptween the cDNAs. NCA lS 90% homologous to CEA at the
nucleotide level (Neumaier et al., 1988; Tawagari et al.,

1 10
1988), while BGP shows 80% homology respectively, in lts fn-st
two domains (Barnett et al., 1988; Hinoda et al., 1988). The
domain structure of these rnRNAs suggests that this gene fùmily
evolved from a common gene ancestor, shared with neurcll Cl'll
adhesion molecule and (lj!S-glycoprotein (Thompson et- al.,
1987) .
The domain structure of the transcripts encoding t'EA
family members is the following: a reglon encoding a 34 amHlO
acid hydrophobie leader sequence, a region encoding an N
terminal domain resembling an immunoglobulin varIable domain,
and a seri es of repeating units whose prctein sequenc:e
suggests a C2-type irnmunoglobulin structure. The CEA cDNA
contains three internaI repeating domains (Beauchemin et al.,
187; Oikawa et al., 1987; Zimmermann et al., 1987) whi le
that of NCA possesses only one (Neumaler et al., 19B7 ;
Tawaragi et al., 1988). BGP rnRNA encodes one complete C2-typ('~
domain followed by a BGP-specific region exhibiting
chacteristics of only half of a C2-type unit (Barnett et al.,
1988; Hinoda et al., 1988).
Although the coding regions for CEA and NCA are highly
conserved, there are differences ln the 3' untranslated
regions of the mature rnRNAs (Zimmermann et al., 1988). The
transcripts encoding BGP, however, show no significanl
homology to either CEA or NCA untranslated regions (Barnett et
;'lI., 1988). Since the greatest divergence between CEA, NCA,
and BGP rnRNAs lies in the 3' untranslated sequence, thesé

f
11
regions were used as specifie probes to identify the
corresponding trar.scripts. Northern blot analyses on RNA from
various tissues demonstrated that each family member presented
cl unique expression pattern and was encoded by distinct
transcripts. CEA was represented by a 3.0 kb and a 3.5 kb
mRNA (Cournoyer et al., 1988; z imme rmann et al., 1988),
di f fering only in the length of the 3' UT, and the use of
dlfferent polyadenylation sites. Using NCA as a probe, only
a 2.6 kb rnRNA was evident (Cournoyer et al., 1988 i Zimmermann
et al., 1988). 'rhe BGP 3 'U'!' probe identified a number of
transcripts (1.8, 2.2, 3.7, and 3.9 kb) (Barnett et al.,
1989) . Analyses of several BGP cDNA clones and of the BGP
gene, demonstrated that several different rnRNAs were produced
by alternative splicing of a precursor rnRNA (Barnett et al.,
1989) .
Prote in Structure of CEA Subgroup Hembers:
The protein structure of CEA family members classifies
them as part of the immunoglobulin (Ig) superfamily (Paxton et
al., 1987; Williams and Barclay, 1988). Proteins of the
immunoglobulin superfamily share the inununoglobulin doma.in
C'omposed of 90-100 amino acids arranged in a sandwich of two
sheets of antiparallel fS-strands, usually stabilized by a
disulfide bond at its centre (Williams annd Barclay, 1988).
The N-terminal domain of CEA, NCA, or BGP, exhibits an
iImnunoglobulin domain structure lacking the intrachain

12
disulfide bridge, but the formation of a salt bridge replacing
the disulfide bridge has been postulated (Thompson et clI.,
1989) . The internaI repeating units contain the necessary
conserved cysteine residues for disulfide bridge formation as
weIl as conserved motifs found in the Ig C2 type domains
(Williams and Barclay, 1988).
CEA, NCA, and BGP are highJy glycosylated proteins
(greater than 40% by weight) existing on the cell surface but
differing however, in their modes of attachment. CEA and NCA
are processed at the carboxyl terminus and linked by a
glycophospholipid moiety to the cell membrane (Takami et al. 1
1988; Hefta et al., 1988; Kolbinger et al., 1989). BGP is
anchored to the cell through a hydrophobic, putatlve
transmembrane domain. BGP is also unique in that it lS the
only family member expressing a cytoplasmic domain (Hi noda et
al., 1988; Barnett et al., 1989).
Cell Adhesion Molecules:
The amine acid sequence of CEA bears the most resemblance
to various mernbers of the immunoglobulin superfamily. TIns
large gene family includes immunoglobulin molecules, T cell
receptors 1 growth factor receptors, and intercellular adhesion
molecules such as N-CAM, T -CAM1, l -CAM2 1 and CD4 (Wi 1] iams and
Barclay, 1988; Springer, 1990). These molecules are aIl
involved, in sorne way, in cellular recognition. Since CEA is
expressed in large amounts by the cells of the déveloping

13
colon (Benchimol et al., 1989), an adhesion role during
embryogenesis has been hypothesized.
An important property of cells during morphogenesis, is
their ability to recognize identical cell types when mixed
with other cell types (Townes and Holtfreter, 1955). This
selectivity is due to specifie homophilic and heterophilic
adhesion molecules present on the cell surface. Two classes
of cellular adhesion molecules have been weIl characterized in
deve1opment: the immunoglobulin superfamily of which NCAM is
the most studied (Cunningham et al., 1987), and the cadherins
(Takeichi, 1988).
Cadherins:
The cadherins can be further di vided into subclasses,
including E-, N-, and p-cadhrrin (Yoshida and Takeichi, 1982;
Hatta et al., 1985; Hat ta and Takeichi, 1986; Nose and
Takeichi, 1986; Shirayoshi et al., 1986). Many more
cadherin-like molecules have been eharacterized across a range
of animal speeies. The fat tumor suppressor gene in
Drosophila encodes a cadherin-like protein containing 34
tandem cadherin domains (Mahoney et al., 1991).
Cadherins mediate homophilic, ealcium-dependent cellular
adhesion (Yoshida and Takeiehi, 1982 i Yoshida-Noro et al.,
1984). This adhesion mechanism is very specifie; E-eadherin
only binds to cells expressing E-cadherin, and the strength of
adhesion is directly proportional to the amount of E-cadherin

14
expressed on the cell surface (Takeichi et al., 1981). Most
tissue types contain at least one type of cadherin, with
multiple cadherin subclasses co-expressed in varying
combinations. As weIl, the pattern of expression within d
cell layer can change throughout development (Takeichi, 1988).
The organization of distinct cell layers is dependent on the
differential expression of cadherin subclasses during
morphogenesis (Damjanov et al., 1986; Vestweber et al.,
1987). 'l'wo well-studied processes, where different cell
groups originating from one cell layer exhibi t dist inct
patterns of cadherin expression when separating from each
other, are lens vesicle formation and development of the
neural tube (Takeichi, 1988).
Neural Cell Adhesion Molecule:
Neural Cell Adhesion Molecule (N-CAM) also plays a role
in neural development, mediating neuron-neuron and certain
neuron-muscle adhesions. Three different forms of N-CAM have
been cloned, and the encoderl. proteins can exist in two
glycosylation states (Cunningham et al., 1987). The proteins
with high carbohydrate content (N-CAM-H) are present during
development. A gradual decrease in sialic acid content occun.>
in the conversion to the adult form of N-CAM (N-CAM-L) i
however, N-CAM-L has also been detected in the early stages Ol
embryonic development (Rutishauser and .Jessel, 1988). In
early nervous system development, N-CAM-L could contribute to

"
15
the stability of neural epithelium during neurulation
(Rutishauser and Jessel, 1988). As development progresses,
decreased adhesiveness could facilitate cell migration and
axonal extension. Once the celis have reached their final
destination, re-expression of N-CAM-L stabilizes the position
and connections of fully differentiated neurons (Rutishauser
anù Jessel, 1988)
carcinoembry~nic Antigen:
Because many colon carcinoma cell lines grow as
aggregates (Rutzky et al. 1 1984), a cell adhesion function was
postulated for CEA. Human colon adenocarcinoma cells
expresslng high or low levels of CEA on the cell surface as
weIl as rodent cells transfected with CEA cDNA, were submitted
to aggregation assays (Benchimol et al., 1989). Celis with
CEA on their surface formed calcium-independent, homotypic
aggregates and the formation of such aggregates could be
specifically inhibited with anti-CEA antibodies. The extent
of aggregation was directly proportional to the CEA expression
level. Other CEA family members have been shown to function
as adhesion moleculesj however, BGP (Rojas et at., 1990),
unlike CEA and NCA (Benchimol et al., 1989; Oikawa et al.,
1989), requires calcium and physiologicai temperatures for its
adhesion function.

1
"
16
Nouse Homologues of Human CHA pami ly Nembers:
Further studies of the function(s) of CEA in development
as well as in tumor formation and metastasis, required animal
models. Animal models would allow approaches such as
transgenesis, controlled carcinogenesis, and in situ
assessment of expression during development. CEA-related
proteins had been detected in chemically-induced colonic
adenocarcinomas, embryonic tissue, and normal adult tissue in
rats (Abeyounis and Milgrom, 1976) Proteins cross-reactive
with anti-CEA antibodies had a] 50 been identified in the lung
and spleen of Macaca monkeys (Engvall et al., 1976) 1 as weIl
as in blood samples from other primates (Haagensen et al.,
1982) .
Murine CEA-like proteins, with a molecular weight of 120
kDa, were identified uSlng polyclonal anti-human CEA
antibodies (Beauchemin et al., 1989). A homologue of human
CEA family members was isolated from a mouse colon cDNA
library, using as a probe for screening, a restliction
fragment of the human CEA cDNA encoding the N-terminal domain.
The mouse N-terminai domain sequence showed 72.6% homology to
the N-terminai domain of human CEA, while the first internaI
repeating domain was 79.4% homologous (Beauchemin et al.,
1989) .
The spatial and temporal expression of murine CEA family
memben during mouse development, was examined us ing both
Northern blot analyses and in situ hybridization (Huang et

17
al., 1990). Mouse CEA probes detected transcripts in mouse
fetuses from 11.5 days ta 17.5 days post coitum (p.c.).
Northern analyses of RNA isolated from fetal intestine and
from colon of newborn pups, detected transcripts at 16.5 days
post coitum and continuing into adulthood. In order to more
accurately define the embryonic tissues expressing CEA family
members, in situ hybridization on sections of fetuses 10.5-
18.5 days p. c. and of newborn pups was performed. Many
tissues of the developing embryos and fetuses contained
transcripts encoding CEA family members; these tissues
included meninges, cartilage and bone, blood vessel walls,
placenta, dermis, muscle layers of the stomach and intestine,
and bronchioles.
Although the mouse CEA gene family members demonstrate a
striking homology to human CEA family members at the amine
acid level, their patterns of expression during development
are quite different. Human CEA is expressed in epithelial
tissues derived from the endoderm (von Kleist et al., 1986;
Nap et al., 1988); however, mouse CEA is found in tissues
derived from mesenchymal cells (Huang et al., 1990).
Developmental expression patterns of homologous genes are
usually well conserved between species. Therefore, the
different CEA expression patterns observed in mouse and in
humdns may indicate that these homologous proteins have
adopted different functions in different species. This
discrepancy may also result from the fact that several CEA

1 18
family members exist in mouse and ln ITtan èlnd these fdnlily
members may crossreact.
Pregnancy Specific-like family members have also been
identified in rat and in mouse. Unique to these proteins is
the expression of repeating Ig variable-like N-domains (3-5
copies) followed by one IgC-like region (Thompson et al.,
~989; Rebstock et al., 1990). Assignment to lhe PSG subgroup
is based on expression pattern alone; they are expressed in
the placenta and are directly secreted (Thompson et al.,
1989) .
Functional Analysis of Murine CBA:
As demonstrated by Northern blot analyses, many mouse CEA
gene family members exist (Beauchemin et al., 1989). One of
these family members, rnrnCGM2, possessing an N-terminal domain
and an A2 domain as well as putative transmembrane and short
cytoplasmic domains, has recently been characterized (Turbide
et al., 1991). Upon transfection of the cDNA into an LTA
mouse fibroblast cell line, these cells acquired the ability
to form aggregates in the presence of calcium at physiologic:al
temperatures.
In the same screening of the mouse colon cDNA li brary
which produced the full coding sequence of rnrnCGM2, a partial
cDNA clone of mmCGM1 was isolated. Chapter 2 of this
dissertation describes the isolation of a full length cDNA for
mmCGMl and the assessment of mmCGM1 protein as an

19
lntercellular adhesion molecule. As well, evidence that
mmCGMl is generated by alternative splicing of one gene is
presented.

l 20
References
Abeyounis, C.J. and Milgrom, F.A. A thermostc1ble dntigen
characteristic for carcinogen-induced rat intestinal turnors.
J. Immun. 116: 3 0 - 3 4, 197 6 .
Barnett, T., Goebel, S., Northdurft, M.A. and Elting, d.
Cloning and characterization of cDNAs for CEA and related
family members. Genornic 3: 59-66, 1988.
Barnett, T., Kretschmer, A., Austen, D.A., Goebel, S.J., Hart,
J.T., Elting, J.J. and Kamarck, M.E. Carcinoembryonic
antigens: Alternative splicing accounts for the mul t iple rnRNAs
that code for novel rnembers of the carcinoembryonic ant igen
family. J. Cell Biol. 108: 267-276, 1989.
Barnet t, T. and Zl.mmermann, W. Workshop report: Proposed
nomenclature for the carcinoembryonic antigen (CEA) gene
family. Turnor Biol. 11: 59 -63, 1990.
Bartels, I. and Lindmann, A. Maternal levels of pregnancy
specifie 1S1-glycoprotein (SPI) are elevated in pregnancies
affected by Down' s syndrome. Hum. Genet. 80: 46-48. 1988.
Beauchemin, N., Benchimol, S., Cournoyer, D. 1 Fuks, A. and
Stanners. C. Isolation and character ization of full length

21
functiona l cDNA clones for human carcinoembryonic antigen.
Mol. Cell. Biol. 7: 3221-3230, 1987.
Beauchemin, N., Turbide, C., Afar, D., Bell, J., Raymond, M.,
Stanners, C. P. and Fuks, A. A mouse analogue of the human
carcinoembryonic antigen. Cancer Res. 49: 2017-2021, 1989.
Benchimol, S., Fuks, A., Jothy, S., Beauchemin, N., Shirota,
K. and Stanners, C. P. Carcinoembryonic antigen, a human tumor
marker, functions as an intercellular adhesion molecule. Cell
57: 27-34, 1989.
Bischof, P. Placental Proteins. In "Contributions to
Gynecology and Obstetrics". P.J. Keller (ed), Karger, New
York, 12: 6-92, 1984.
Borj igin, J., Tease, L.A., Barnes, W. and Chan, W.Y.
Expression of the pregnancy-specific fS1-glycoprotein genes in
human testis. Biochem. Biophys. Res. Commun. 166: 622-629,
1990.
Boucher, D. , Cournoyer, D., Stanners, C . P . and Fuks, A.
Studies on the control of gene expression of the
carcinoembryonic antigen family in human tissue. Cancer Res.
49: 847-852, 1989.

1 Cournoyer, D., Beauchemin, N., Boucher, D., BencIllmO l, S.,
Fuks, A. and Stanners, C. P. Transcription of genes of the
carcinoembryonic antigen family in malignant and nonmalignant
tissues. Cancer Res. 48: 3153-3157, 1988.
Cunningham, B.A., Hemperly, J.S., Murray, B .. Z\., Prediger,
E.A., Brackenbury, R. and Edelman, G.M. Neural cell adhesion
molecule: structure, immunoglobulin-like domains, cell
surface modulation, and alternative RNA splicing. Science 236:
799-806, 1987.
Damjanov, r" Damjanov, J. and Damsky, C.H. Developmentally
regulated expression of the cell-cell adhesion glycoprotein
cell-CAM 120/180 in peri-implantation mouse embryos and
extraembryonic membranes. Dev. Biol. 116: 194-202, 1986.
Engvall, E., Vuento, M. and Ruoslahti, E. A monkey antigen
cross-reacting with carcinoembryonic antigen, CEA. Br. lJ.
Cancer 34: 341-345, 1976.
Fritche, R. and Mach, J.P. Isolation and characterization of
carcinoembryonic antigen (CEA) extracted from normal human
colon mucosa. Immunochemistry, 14: 119-127, 1978.
Gold, P. , and Freedman, 5.0. Speclfic carcinoembryonlc
antigens of the human digestive system. J. Exp. Med., 122:

t 1
23
467-481, 1965.
Gold, P. and Shuster, J. Historical development and potential
uses of tumor antigens as markers of human growth. Cancer Res.
40: 2973-2976, 1981.
Goldenberg, D.M., Deland, F., Kim, E., Bennett, S., Primus,
F.lJ., van Nagell, J.R., Estes, R., DeSimone, P. andRayburnP.
Use of radiolabelled antibodies to CEA for the detection and
localization of diverse cancers by external phutoscanning N.
Engl. J. Med. 298: 1384-1388, 1978.
Haagensen, D. E., Metzgar, R. S., Swenson, B., Dilley, W.G.,
Cox, C.E., Davis, S., Murdoch, J., Zamchek, N. and Wells,
S.A., Carcinoembryonic antigen in nonhuman primates. J. Natl.
Cancer Inst. 69: 1073-1076, 1982.
Hakim, A. CEA antigen, a tumor associated glycoprotein induces
defective lymphocyte function. Neoplasma. 31: 385-397, 1984.
Hatta, K., Okada, T.S. and Takeichi, M. A monoclonal antibody
disrupting calcium dependent cell-cell adhesion of brain
tissues: possible role of its target antigen in animal pattern
formation. Proc. Natl. Acad. Sei. U.S.A. 82: 2789-2793, 1985.
Hat ta, K. and 'rakeichi, M. Expression of N-cadherin adhesion

! 24
, .l molecule associated with early morphogenetic events in chick
development. Nature 320: 447-449, 1986.
Hefta, S.A., Hefta, L.J.F., Lee, T.D., Paxton, R.LI. dnd
Shively, J.E. Carcinoembryonic antigen lS anchored to
membranes by covalent attachment to a glycophosphtidylinositol
moiety: identification of the ethanolamine linkage si te. Proc.
Natl. Acad. Sci. U.S.A. 85: 4648-4652, 1988.
Heikinheimo, M., Aula, P., Rapola, J., Wahlstrom, T., Jalanko,
H. and Seppala, M. Amniotic fluid pregnancy-specific e1-
glycoprotein (SP1 ) in Meckel' s syndrome: A new test (or
prenatal diagnosis? Prenat. Diag. 2: 103-108, 1982.
Heikinheimo, M., Jalanko, H., Renlund, M., Rapola, J. and
Wahlstrom, T. Studies on the amniotic fluid SPI in Meckel's
syndrome: modified glycosylation of SP1 • Placenta 8: 427-432,
1987.
Hinoda, Y., Neumaier, M., Hefta, S., Drzeniek, Z., Wagener,
C., Shive1y, L., Hefta, L., Shively, J. and Paxton, a.
Molecular cloning of a cDNA coding biliary glycocprotein 1:
primary structure of a glycoprotein immunologically
crossreactive with carcinoembryonic antigen. Proc. Natl. Acad.
Sei. U.S.A. 85: 6959-6963, 1988.

25
Hinoda, Y. rmai, K., Nakagawa, N., rbayashi, Y., Nakano, T.,
Paxton, R. J ., Shi vely, J. E. , Yachi, A. Transcription of
biliary glycoprotein 1 in malignant and non-malignant human
liver tissues. Int. J. Cancer 45: 875-878, 1990.
Ho, P.C., Chan, S.Y.W., and Tang, G.W.K. Diagnosis of early
pregnancy by enzyme immunoassay of Schwangerschafts prote in 1.
Fertil. Sterile 49: 76-80, 1988.
Hostetter, R.B., Campbell, D., Chi, K., Kerkhoff, S., Cleary,
K., Ullrich, S., Thomas, P. and Jessup, M. Carcinoembryonic
antigen enhances metastatic potential of human colorectal
carcinoma. Arch. Surg. 125: 300-304, 1990.
Huang, J .Q., Turbide, C., Daniels, E., Jothy, S. and
Beauchemin, N. Spatiotemporal expression of mur1ne
carcinoembryonic antigen (CEA) gene family members during
mouse ernbryogenesis. Development 110: 573-588, 1990.
Jessup, J.M, Giavazzi, R., Campbell, D., Cleary, R., Mo ri kawa ,
K. and Fidler, 1. J. Growth potential of human colorectal
carcinomas in nude mice: association with the preoperative
serum concentration of carcinoembryonic antigen in patients.
Cancer Res. 48: 1689-1692, 1988.

J6
Kahn, W. N. and Hammarstrom, S. Carcinoembryonic ant igen geI1l..'
family: molecular cloning of cDNA for a PSS/FL -NCA
glycoprotein with a novel domain arrangement. Biochem.
Biophys. Res. Commun. 161: 525-535, 1989.
Kahn, W.N. and Hammarstrom, S. Identification of a new
carcinoembryonic antigen (CEA) family member ln hUitian Eel al
liver: cloning and sequence determination of pregnancy
specifie glycoprotein 7. Biochem. Biophys. Res. Commun. 168:
214-225, 1990.
Kahn, W.N., Osterman, A. and Hammarstroffi, S. Molecular cloniwJ
and expression of cDNA for a carcinoernbryonic antigen-relatecl
fetal liver glycoprotein. Proc. Natl. Acad. Sei. U.S.A. 86:
3332-3336, 1989.
Kolbinger, F., Scharz, K., Brombacher, F., von Kleist, S. and
Grunert, F. Expression of an NCA cDNA in NIH/3T3 cells yields
a 110K glycoprotein, ~"hich is anchored lo the membrane Vid
glycosyl-phosphatidylinositol. Biochem. Biophys. Res. Commun.
161: 1126-1134, 1989.
Kuroki, M., Arakawa, F., Matsuo, Y., Oikawa, S., Misurm., Y.,
Nakazato, H. and Matsuoka, Y. Molecular cloning of nonspecific
cross-reacting antigens in human granulocytes. lJ. Biol. Chem.
266: 11810-11817, 1991a.

27
Kuroki, M., Arakawa, F., Matsuo, Y., Oikawa, S., Nakazoto, H.
and Matsuoka, Y. Three novel molecular forms of biliary
glycoprotein deduced from cDNA clones from a human leukocyte
library. Bioehem. Biophys. Res. Commun. 176: 578, 1991b.
Leslie, K.K., Watanabe, S., Lei, K-J., Chou, D.Y., Pl, utzek,
C.A., Deng, H-C., Torres, J. and Chou, J.Y. Linkage of two
pregnancy-specifie g1-g1ycoprotein genes: one is associated
with hydatidiform mole. Proc. Natl. Aead. Sei. U.S.A. 87:
5822-5826, 1990.
Mahoney, P .A., Weber, V., Onofrechuk, P., Biessmann, H.,
Bryant, P.J. and Goodman, C.S. The fat turnor suppressor gene
in Drosophila encodes a novel member of the cadherin gene
superfamily. Cell 67: 853-868, 1991.
Medoff, J.R., Clack, V.D. and Roche, J.K. Charaeterization of
irnrnunosuppressi ve factor from malignant asci ties that
resembles a factor induced in vitro by CEA in patients. Cancer
Res. 48: 1689-1692, 1988.
Moore, T. , Dhar, P. and Zarncheck, N. Careinoembryronic
antigen (s) in liver disease. Gastroenterology 63: 88-94, 1972.
Nap, M., Mollgard, K.,
Immunohistochemistry of
Burtin, P. and
carcinoernbryronic
Fleuren,
antigen in
G.J.
the

1 28
embryo, fetus, and adult. Tumor Biol. 9: 145-153, 1988.
Neumaier, M., Zimmermann, W., Shively, L., Hinoda, Y., Rigqs,
A.D. and Shively, J.E. Characterization of a cDNA clone for
the nonspecific cross-reacting antigen (NCA) and compdrison of
NCA and carcinoernbryonic antigen. J. Biol. Chem. 263: 3202-
3207, 1988.
Neville, A.M. and Caurence, D.J.R. The carcinoembryonic
antigen (CEA): the present position and proposaIs for future
investigation. Int. J. Cancer 14: 1-18, 1974.
Nose, A. and Takeichi, M. A novel cadherin adhesion molecule:
Its expression patterns associated with implantation and
organogensis ofmouseernbryos. J. Cel!. Biol.103: 2649-2658,
1986.
Oikawa, S., Nakazato, H. and Kosaki, G. primary structure of
human carcinoembryonic antigen (CEA) deduced from cDNA
sequence. Bioehem. Biophys. Res. Commun. 142: 511-518, 1987.
Oikawa, S., Inuzuka, C., Kuroki, M., Matsuoka, Y., Kosaki, G.
and Nakazato, H. Cell adhesion aetivity of non-specifie cross
reacting antigen (NCA) and careinoernbryronic antigen (CEA)
expressed on CHO cell surface: homophilic and heterophilic
adhesion. Bioehem. Biophys. Res. Commun. 164: 39-45, 1989.

29
Paxton, R.J., Mooser, G., Pande, H., Lee, T.D. and ShiveIy,
.J • R. Sequence analysis of carcinoembryonic antigen Proc. Nati.
Acad. Sei. U.S.A. 84: 920-924, 1987.
Rebstock, S., Lucas, K., Thompson, J.A. and Zimmermann, W.
cDNA and gene analyses imply a novel structure for a rat
carcinoembryonic antigen-related protein. J. Biol. Chem. 265:
7872-7879, 1990.
Robbins, S., Cotran, R. and Kumar, V., In Pathological Basis
of Disease, 3rd ed., W.B. Saunders Company, U.S.A. 268-269,
1984.
Rojas, M., Fuks, A. and Stanners, C.P. Biliary glycoprotein,
a member of the immunogiobulin supergene family, functions in
vitro as a Ca2+- dependent intercellular adhesion moiecuie.
Cell Growth Diff. 1: 527-533, 1990.
Rutishauser, U. and Jessel, T.M. Cell adhesion molecules in
vertebrate neural development. Biol. Rev. 68: 819-856, 1988.
Rutzky, L.P., Tom, B.H. and Kahan, B.D. Biological and
antigenic analysis of human colon cancer cell clones. Prog.
Cancer Res. Ther. 29: 135-145, 1984.
Sato, C., Miyaki, M., Oikawa, S., Nakazato H.and Kosaki, G.
Differential expression of carcinoembryonic antigen and

1 30
nonspecific crossreacting antigen genes in human colon
adenocarcinomas and normal colon mucosa. Jpn. J. Cancer Res.
79: 433-437, 1988.
Schonk, D., van Dijk, P., Riegmann, P., Trapman, J., Holm, C.,
Willcocks, T.C., Sillekens, P., van Venrooij, W., Wimmer, E.,
Geurts, E., van Kessel, A.,
Assignment of seven genes
Ropers, H-H. and Wieringa, B.
to distinct intervals on the
midportion of human chromosome 19q surrounding the myotonic
dystrophy gene region. Cytogenet. Cell Genet. 54: 15-19, 1990.
Schrewe, H., Thompson, J., Bona, M., Hefta, L.J.F., Marruya,
A., Hassauer, M., Shively, J.E., von Kleist, S. and
Zimmermann, W. Cloning of the complete gene for
carcinoembryonic antigen: analysis of its promotor indicates
a region conveying cell type specifie expression. Mol. Cell.
Biol. 10: 2738-2748, 1990.
Shirayoshi, Y., Nose, A., Iwasaki, K. and Takeichi, M. N
linked oligosaccharides are not involved in the function of
cell-cell binding glycoprotein. Cell Struct. Funct. 11: 245-
252, 1986.
Shupert, W.L. Pregnancy-specific 151-glycoprotein in human
intestine. Ph.D. dissertation, University of Oklahoma,
Oklahoma City, Oklahoma, 1990.

31
Shuster, J.D. Thompson, M.P., Fuks, A. and Gold, P.
Immunologie approaches to diagnosis of malignancies. Prog.
Exp. Tumor Res. 25: 89-139, 1980.
Sorensen, S. pregnancy "specifie" S1-glycoprotein (SP1):
purification, characterization, quantification and clinical
application in malignancies. Tumor Biol. 5: 275-302, 1984.
Springer T. A. Adhesion receptors of the immune system. Nature
346: 425-434, 1990.
Svenberg T. Carcinoernhryonic antigen-like substances of human
bile: isolation and partial characterization. Int. J. Cancer
17: 588-596, 1976.
Takami, N., Misumi, Y. 1 Kuroki, M., Matsuoka, Y. and Ikehara,
Y. Evidence for carboxyl-terminal processing and glycolipid
anchoring of human carcinoembryonic antigen. J. Biol. Chem.
263: 12716-12720, 1988.
Takayama, M., Soma, H., Isaka, K., Okudera, K., Ogawa, T. and
Kikuchi, K. Diagnostic reliability of simultaneous
measurements of beta human chorionic gonadotropin and
pregnancy-specific beta-1-glycoprotein in serum of patients
with trophoblastic disease. Gynecol. Obstet. Invest. 23: 151-
156, 1987.

32
Takeichi, M., Atsumi, T., Yoshida, C., Uno, K. and Okada, T. S.
Selective adhesion of embryonal carcinoma cells and
differentiated cells by Ca2+-dependent sites. Dev. Biol. 87:
340-350, 1981.
Takeichi, M. The cadherins: Cell-cell adhesion molecules
controlling animal morphogenesis. Development 102: 639-655,
1988.
Tawaragi, Y. , Oikawa, S. , Matsuoka, Y., Kosaki, G. and
Nakazato, H. primary structure of nonspecific crossreacting
antigen (NCA), a member of the carcinoembryonic antigen (CEA)
gene family, deduced from cDNA sequence. Biochem. Biophys.
Res. Commun. 150: 89-96, 1988.
Thomas, P., Toth, C.A., Elting, J.J. and Steele, G. A peptide
involved in CEA binding to a 80kD protein on Kupffer cells is
common to stromelysin and complement Cls. Washington
International PSG/CEA Workshop #19, June 1991.
Thompson, J., Pande, H., Paxton, R., Shively, L., Padma, A.,
Simmer, R., Todd, C., Riggs, A. and Shively, .1. Molecular
cloning of a gene belonging to the carcinoembryonic antigen
gene family and discussion of a domain model. Proe. Natl.
Acad. Sci. U.S.A. 84: 2965-2969, 1987.

33
'rhompson, J., Barnert, S., Berling, B., von Kleist, S.,
Kodelja, V., Lucas, K., Mauch, E-M., Rudert, F., Schrewe, H.,
Weiss, M. and Zimmermann, W. Structure, expression and
evolution of the human and rat carcinoembryonic antigen (CEA)
gene families. In The Carcinoembryonic Antigen Gene Family, A
Yachi, JE Shively eds. Elsevier Science Publishers, BV
Amsterdam, 65-74, 1989.
Thompson, J . , Koumari, R. , Wagner, K. , Barnert, S. ,
Schleussner, C., Schrewe, H., Zimmermann, W., Muller, G.,
Schemp, W., Zaninetta, D., Ammaturo, D. and Hardrnan, N. The
human pregnancy specifie glycoprotein genes are tightly linked
on the long arm of chromosome 19 and are coordinately
expressed. Biochem. Biophys. Res. Commun. 167: 848-859, 1990.
Thompson, J., Grunert, F. and zimmermann, W. The
carcinoembryonic antigen gene family: IT.olecular biology and
clinical perspectives. J. Clin. Lab. Anal. (submitted)
Townes, P.L. and Holtfreter, J. Directed movements and
selective adhesion of embryonic amphibian cells. J. Exp. Zool.
128: 53-120, 1955.
Tran, R., Kashrniri, S.V.S., Kantor, J., Greiner, J.W., Pestka,
S., Shively, J.E., Schlom, J. Correlation of DNA
hypomethylation with expression of carcinoembryonic antigen in

1 34
human colon carcinoma cells. Cancer Res. 48: 5674-5679, 1988.
Turbide, C., Rojas, M., Stanners, C.P. and Beauchemin, N. A
mouse carcinoembryonic antigen gene family member is a calcium
dependent cell adhesion molecule. J. Biol. Chem. 266: 309 -315,
1991.
Vestweber,
Expression
D., Gossler, A.,
and distribution
BolIer, K. and Kemler, R.
of cell adhesion molecule
uvomorulin in mouse preimplantation embryos. Dev. Biol. 124:
451-456, 1987.
von Kleist, S., Chavanel, G. and Burtin, P. Identification of
an antigen from normal human tissue that crossreacts with the
carcinoembryonic antigen Proc. Nat!. Acad. Sci. U. S. A. 69:
2492-2494, 1972.
von Kleist, S., Winkler, J., Migule, 1. and Bohm, N.
Carcinoembryonic antigen (CEA) expression ln early
embryogenesis: a study of the first trimester of gestation.
Anticancer Res. 6: 1265-1272, 1986.
Wanebo, H.J., Rao, B., Pinsky, C.M. et al. Preoperative
carcinoembryonic antigen level as a prognostic indicator in
colorectal cancer. N. Engl. J. Med. 299: 448-451, 1978.

35
Watanabe, S. and Chou, J.Y. Isolation and characterization of
complementary DNAs encoding human pregnancy-speci fic $51-
glycoprotein. J. Biol. Chem. 263: 2049-2054, 1988.
Willcocks, T.C. and Craig, I.W. Characterization of the
genomic organization of human carcinoembryonic antigen (CEA):
Comparison with other family members and sequence analysis of
the 5' controlling region. Genomics 8: 492-500, 1990.
Williams A.F. and Barclay, A.N. The immunoglobulin
superfamily: domains for cell surface recognition. Ann. Rev.
Immun. 6: 381-405, 1988.
Willis, R.A. Spread of tumors in the human body. Stoneham
Mass: Butterworths 417-450, 1973.
Yoshida, C. and Takeichi, M. Teratocarcinoma cell adhesion:
identification of a cell surface protein involved in calcium
dependent cell aggregation. Cell 28: 217-224, 1982.
Yoshida-Noro, C., Suzuki, N. and Takeichi, M. Molecular nature
of the calcium-dependent cell-cell adhesion system in mouse
teratocarcinoma and embryonic cells studied with a monoclonal
antibody. Dev. Biol. 101: 19-27, 1984.

36
Zimmermann, w., Ortleib, B., Freidrich, R. and von Kleist, S.
Isolation and characterization of cDNA clones encoding the
human carcinoembryronic antigen reveal a highly conserved
repeating structure. Proc. Natl. Acad. Sci. 84: 2960-2904.
1987.
Zimmermann, W., Weber, B., Ortljeb, B., Rudert, F., Werner,
S., Fiebig, H-H., Shively, J., von Kleist, S. and Thompson,
J.A. Chromosomal localization of the carcinoembryonic antigen
gene family and differential expression in various tumors.
Cancer Res. 48: 2550-2554, 1988.
Zimmermann, W., Weiss, M. and Thompson, J .A. cDNA cloning
demonstrates the expression of pregnancy-specific glycoprotein
genes, a subgroup of the carcinoembryonic antigen gene family,
in fetal liver. Biochem. Biophys. Res. Commun. 163: 1197-1209,
1989.
Zoubir, F., Khan, W.N. and Hammarstrom, S. Carcinoembryonic
antigen gene family members in submandibular sali vary gland:
demonstration of pregnancy-specific glycoproteins by cDNA
cloning. Biochem. Biophys. Res. Commun. 169: 203-216, 1990.

mmCGMla: A MOUSB CARCINOBMBRYOHIC AHTIGBN GBNE FAMILY
MBMBBR, GBNERATBD BY ALTBRNATIVE SPLICING,
FURCTIONS AS AH ADHBSION MOLECULB
This rnanuscript was prepared for publication. McCuaig, R., Turbide, C., and Beauchemin, N. Cell Growth and Differentiation vol. 3, 165-174.

37
AB S TRACT
Carcinoembryonic antigen (CEA) is a human tumor marker and the
prototype of a large family of irnmunoglobulin-like proteins.
We have been developing a mouse modei for this large protein
family and have cloned a complementary DNA for a mouse CEA
gene family member (rnmCGM1a). Two transcripts expressed in
several different adult mouse tissues hybridize to this cDNA,
a 1.8 kb and a 4.6 kb mRNA. Sequences of many related cDNA
clones indicate that they are most likely encoded by a single
gene which undergoes alternative splicing. The protein encoded
by the rnmCGM1a cDNA shares 69% of the amino acid residues in
the N-terminal domain with a rat liver ecto-ATPase and with
the human biliary glycoprotein. Mouse fibroblast transfectant
ceoiis expressing the mmCGM1a protein on their cell surface
exhibit calcium- and temperature-independent adhesion in vitro
which can be specifically inhibited by an antibody raised
against a CEA-related 120 kD proteine

38
INTRODUCTION
Carcinoembryonic antigen (CEA1) is a heavily glycosylated
protein used clinically as a tumor marker to detect
recurrences of many types of tumors (1,2). This 180 kD
glycoprotein belongs to the immunoglobulin superfamily (3,4)
and is the prototype of the large CEA family of proteins (5).
Following the cloning of CEA (6-9), many other related
proteins have been cloned: the CEA-related group (CEA, NCA,
BGP, CGM1, CGM6, CGM7) and the pregnancy-specific group (PSG1-
Il) (for references, see 10). AlI of these human proteins
display a similar structure where the N-terminal domain
resembles a variable immunoglobulin domain and the internaI
repeating domains are C2-set Ig domains (11). These proteins
are, however, different in their cell membrane attachment
mechanism and in their expreSSlon pattern. Biliary
glycoprotein (BGP) bears a transmembrane domain and a long or
short intracytoplasmic tail (12,13) while CEA, NCA and CGM6
are at tached to the cell membrane by a glycophospholipid
anchor (14-17) . The pregnancy-specific glycoproteins (PSG) are
generally secreted (18). Most CEA-related family members are
expressed at the apical membrane of Epithelial cells of many
IThe abbreviations used are: CEA, carcinoembryonic antigeni NCA, normal cross-reacting antigen; BGP, biliary glycoprotein; CGM, CEA gene family member; PSG, pregnancy-specific glycoproteini Ig, immunoglobulin; TCA, trichloroacetic acid; EDTA, ethyJenediaminetetraacetic acid; EGTA, ethylenebis(oxyethylenenitrilo)-tetraacetic acid; PBS, phosphate-buffered saline; MOPS, morpholinopropanesulfonic acid.

1 l
39
normal tissues and tumour cells (1) while the pregnancy
specific-like proteins are usually found in placenta (19),
testis (20) and choriocarcinomas (18).
The functions of these family members have been
investigated. CEA, NCA and BGP are cell adhesion molecules as
demonstrated by in vitro aggregation assays (21-23)i CEA Inùy
also play a role in intestinal tissue organization during
development (21). Both CEA and NCA have also been shown to
recognize Escherichia coli extracted from human gut or trachecl
(24) • PSG has been postulated to function as an
immunomodulator during pregnancy (25).
In order to develop a mouse model for the human CEA gene
family, we first constructed and screened a mouse colon cDNA
library using a human CEA cDNA restriction fragment. Several
clones were isolated from this cDNA librarYi a partial
sequence of mmCGM1 (Mus musculus CEA gene family member) has
previously been reported (26). A second cDNA clone (~nCGM2)
has recently been characterized as a cell adheslon molecule
(27). We now wish to report on the full length ~nCGMl cDNAi
we also present evidence that these mouse CEA gene famjly
members are splice variants of the same gene and that they
share tissue specificity as weIl as function, although rnrnCGM1,
unlike mmCGM2, encodes a protein that behaves as a calcium-
and temperature-independent cell adhesion molecule. In order
to comply with the adopted nomenclature for these gene family
members (10), we shall henceforth refer to mmCGMl as mmCGM1Q

( 40
and mmCGM2 (27) as mrnCGMlQ.
RBSULTS
Characterization of the cDNA Clones and Comparison with Other
CHA Gene Family Hembers:
A first screening of the mouse colon cDNA library
produced 92 positive clones, the majority of which enclosed
inserts of either 1.42 (mrnCGMla) or 1.31 kb (mmCGM1b).
Initial nucleotide sequence analyses indicated that clone 46
was composed of an incomplete N-terminal domain, followed by
C2-set immunoglobulin domains (11). This cDNA clone also
enclosed a transmembrane and a cytoplasmic domain followed by
306 bp of 3' untranslated region (Fig.lA). To obtain the 5'
end of this cDNA clone, the same library was rescreened with
two probes: a 5' EcoRl-Sstl and a 3' Accl-EcoRl restriction
fragments of the clone 46. Several cDNA inserts were
analyzed, one of which (clone 23) contained an identical
nucleotide sequence overlapping the partial N-terminal
sequence of clone 46. Clone 23 also contained a 5'
untranslated region (104 bp), a putative leader sequence (102
bp) and the complete mrnCGMla N-terminal domain homologous to
mmCGMlb. In addition, clone 23 contained an A2 domain, a
t ransmembrane and a cytoplasrnic domain as weIl as a 3'
untranslated region identical to that of mmCGMlb (27) but
extended this 3' untranslated region by 200 bp and terminated
with a polyadenine tail. A full length mrnCGMla cDNA (Fig. lA)

1 41
was reconstructed by combining the EcoRl-BamHl of clone 23
with the BarnHl-EcoRl portion of clone 46. To ascertaln that
such a full length clone existed in the mRNA population,
reverse transcript ion of colon RNA
chain amplification (peR) of the
followed by polymerase
first strand cDNA WdS
performed using oligonucleotides within the N-terminal and A2
domains, as described in Materials and Methods. 'l'Wo PCR
fragments were obtained and sequenced: one band exhibi ted
identical nucleotide sequence to the N-terminal, Al, 81 and A2
domains of the reconstructed mmCGM1a while the second band
represented the sp1icing of tht: N-terminal to the A2 domain
and was identical to cDNA clone 23, confirming that both RNA
species are expressed in mouse adult colon.
The protein sequence encoded by rrunCGMla and the
structural features common to CEA gene family members are
shown in Fig. 1Bi this protein exhibits a 34 amino aei.d region
corresponding to a signal sequence as defined by von Heijne
(28). The mature protein contains a 108 amino acid N-terminaJ
domain with characteri stics of an immunogiobul in variable
region (11). This domain supports three N-I inked
glycosylation sites, two of whieh are eornmonly found in aIl
CEA gene famiIy members of all species studied so far (huma.n,
rat and mouse). It is thought that a salt bridge can forrn
using the charged residues (GREIl} and (KDMGV} of the N
terminal domain (29). The N-terminal domain is followed by
three C2-set irnmunoglobulin domains (Al, B1, A2) (11)

42
exhibiting two cysteine residues per domain (identified ~n
Fig. lB) which are postulated to form intrachain disulphide
bonds. Contrary to the human BGP protein, a third cysteine
thought to be invol ved in dimer formation (13) is absent from
the A2 domain. A short 10 amine acid linker region (Li in
Fig. lA) hooks these immunoglobulin domains to a transmembrane
domain and a 10 amine acid putative intracytoplasmic tail rich
in serines and glycines. A stop codon is found at nucleotide
1478. Sixteen N-linked glycosylation consensus sequences are
spread throughout the N-terminal and C2 -set inununoglobulin
domains. The predicted molecular weight of the mature protein
(before post-translational modifications) is estimated to be
46.5 kD.
We have compdred the mmCGM1a DNA and protein sequences
with those of mouse mmCGM1b, rat hepatocyte ecto-ATPase (30)
and human biliary glycoprotein (BGP, 12,13) (Fig. 2). The
signal sequences of mmCGM1a and mmCGM1b display only two
conservative replacements. The first thirty-seven residues of
the N-terminal domains of these two clones are identical while
significant substitutions characterize the core of this N
domain. The Al and B1 domains found in rnmCGM1a are absent and
possibly spliced out of rnrnCGMlb (indicated by dashes in Fig.
2). The mouse cDNAs share 85% of their DNA sequences in the
signal sequence and N-terminal domain (87% at protein level)
while the rat ecto-ATPase is 75% homologous (69% at protein
level) to rnmCGM1a and the human BGP is 69% homologous (69% at

'f , 43
protein level). The internaI repeat structures (Al, BI, A2)
of the mouse, rat and human cDNAs share 99%, 86% and 73% of
their bases (98%, 87% and 73% of their amIno acids)
respectively. Three nucleotide changes leading to three amino
acid substitutions are seen in the mouse A2 domains of rnrnCGMld
and mmCGMlb. The linker region as weIl as the transmembrdne
and putative intracytoplasmic domains of the two mouse ('Innes
are identical. However, the 3' untranslated regions are
radically different.
Bxpression of Nouse mmCGMla Transcripts:
Hybridization with a probe consisting of clone 46 (Fig.
lA) revealed several transcripts which react or crossreact
with this probe in many tissues, as can be seen in Fig. 3A.
A predominant transcript measured as 4.6 kb was the most.
abundant in colon, uterus, kidney and liver (Fig. 3A) as weIl
as in gallbladder and small intestine (data not shown).
Underexposure of the autoradiogram demonstrated that the same
discrete transcripts seen in uterus were also expressed in
colon where transcript expression was most abundant. Several
other less abundant transcripts were apparent ; n colon,
uterus, li ver, ovary and placenta. Crossre,3ct i vit y wi th the
mouse probe was also observed with human colon carcinoma RNA
(L8180) where four transcripts were detected. As noticed
previously (27), upon Northern analyses, many mouse tissues

44
did not express any detectable transcripts: brain, pancreas,
heart, lung, bladder, testes, spleen and breast (data not
shown) as weIl as fibroblasts (LTA) (Fig. 3A). Equal loading
of the samples was assessed by rehybridization with a fS-actin
probe (Fig. 3B).
Assignment of a specifie transcript to mmCGMla was done
using either restriction fragments or oligonucleotides. The
104 bp 5' untranslated region (EcoRl-Ddel) detected only a 4.6
kb mRNA (Fig. 3C, panel A) while a longer EcoRl-BamHl
restriction fragment containing the same 5' untranslated
region, the signal sequence and two thirds of the N-terminal
domain detected both a 1. 8 and 4.6 kb rnRNA (Fig. 3C, panel B).
A 19-mer oligonucleotide corresponding to the 5' end of clone
46 was also used as a probe as its sequence was different from
that of the corresponding oligonucleotide in mmCGM1b: at low
stringency (Fig. 3C, panel Cl, this oligonucleotide recognized
a 1.8, a 4.1 and a 4.6 kb transcript. However, raising the
stringency of the washes retained only the 4.6 kb signal (data
not shawn). A 60 bp Accl-EcoRl restriction fragment at the 3'
terminus (Fig. 3C, panel D) or an Sspl-EcoRl probe including
the transmembrane and short intracytoplasmic domain as weIl as
the specifie 3' untranslated region (data not shown) detected
only the 4.6 kb transcript. Therefore, two main transcripts
respond to hybridization with specifie mmCGMla probes: a 1.8
kb and a 4.6 kb rnRNA.

i 45
Expression of the mmCGNla protein:
The rnrnCGM1a cDNA was inserted in the sense and a~tisense
orientation into the p91023B vector (31) where transcription
of the cDNA is insured by the adenovirus major Idte' promoter.
These contructs were coprecipitated with the pSV2neo plasrnid
into a mouse embryonic fibroblast cell line (NIH3T3). Several
sense and antisense transfectant cell clones were selected and
evaluated for their production of a mmCGMla CEA-related
protein.
'l'wo antibodies were used: a polyclonal anti-hurndn CEA
antibody raised against purified human CEA extracted from
liver metastases of primary colonie adenocarcinomas and d
polyclonaJ antibody raised aga:i nst a p120 CEA-related protein
purified from mouse colon as described in Materials and
Methods. The purified mouse p120 protein used as antigen did
not react with antibodies raised against other cell adhesion
molecules known to be present in mouse colon, such as E
Cadherin (results not shown) (32). As (~an be seen in Fig. 4A,
both mouse colon membrane proteins (lanes 1, 7 and 12) and the
purified p120 protein (lane 2) contained proteins
immunoprecipitated by either the anti-hurnan CEA antibody
(lanes 1, 2, 12) or the anti-mouse p120 antibody (lane 7).
The affinity of the anti-hurnan CEA antibody for the p120
protein is less, however, than that of the anti -mouse p120
antibody as can be seen in the immunoblots. Sense-dri ven
transfectant cells S6 and 511 express a 110-120 kD protein

(
(
46
that is recognized by both antibodies (lanes 4, 5 and 9, 1D),
while antisense-driven transfectant cells A4 or normal rabbit
serum did not express this protein (lanes 3 and 8, 6 and 11) .
This result demonstrates that the sense-driven transfectant
cell s, contrary to the antisense-dri ven transfectant or
parental cells, are expressing a new protein on this cellular
background which corresponds to the glycosylated forro of the
mmCGM1a protein product.
S11 sense transfectant cells grown to 90% confluency were
incubated in the presence of the anti-mouse p120 antibody and
then reacted with a fluorescein-conjugated secondary antibody.
The CEA-related protein produced by these cells was expressed
at the cell surface as can be judged in Fig. 4B. Reaction of
either parental cells with the same antibody or Sll cells with
pre-immune serum did not show any significant fluorescent
signal (data not shown).
mmCGMla as a Cell Adhesion Molecule:
Since many of the human CEA gene family members function
in vitro (21-23, 27, 33-35) and in vivo (21, 35) as cell
adhesion molecules, we wished to assess the adhesion
properties of the mmCGM1a transfectant cells. Por this
purpose, parental NIH3T3 cells, sense and antisense-driven
transfectant cells were submitted to in vitro aggregation
assays. When either parental or antisense-driven transfectant
cells were incubated in any conditions (in complete medium or
saI ine, at physiological or cold temperatures) little or no

1 47
formation of cell clumps was observed (Fig. 5A, 8, C).
However, when sense S6 or SIl t:r ansfectant cells were
incubated in complete medium, cells formed clurnps even after
a time interval of 30 min (Fig. 5A) and proceeded to aggregate
during a 2 h period at which time 60% of the cells were found
in aggregates of 5-6 cells each. The adhesion of these cells
is specifically due to the presence of the rnrnCGMla protein dt
the cell surface since aggregation assays done in the presence
of Fab fragments of an antibody recogni zing this protein (Fig.
4A) did not reveal any significant formation of cell clumps
(Fig. 5A). When complete medium was substituted by d saI ine
solution containing or lacking Ca 2 +, no difference was seen in
the kinetics of aggregation (Fig. 5B) Incubation of sense
transfectant cells in the presence of EGTA demonstrated
similar aggregation (data not shown). The temperature of the
assay was shifted from 37°C to 4°C with only slight
modi fications to the aggregation kinetics (Fig. 5C) when
compared to aggregation at 37°C (Fig. SA) or to aggregation of
rnrnCGMlb at 4°C (Fig. 58 in ref. 27). We conclude that the
rnrnCGMla protein confers Ca 2+- and temperature-independent cell
adhesion properties onto the parental cells.

'.
48
DISCUSSION
The present paper describes the characterization.
expression and adhesJ.on function of the first major mouse CEA
related cDNA to be cloned (26). The mmCGMla cDNA encodes a 34
amine acid signal sequence ~hat is followed by a 108 amine
acid N-terminal domain. ..s domain, by comparison to the
mmCGMlb N-terminal domain, is the one that most distinguishes
these two coding regions since 87% of the residues are
conserved versus 99% of amino acids in the C2-set (A2) domain.
In fact, two features differentiate these two N-terminal
domains: 1. mmCGMla contains an extra N-linked glycosylation
consensus sequence. 2. many amino acid substitutions are not
well conserved (i.e. K~A, F~Q, M~K, Q~P, Y~H) indicating that,
if these residues bear any functional significance, they would
impri.nt on these two proteins a different structure and
possibly a different function. This hypothesis is presently
being evaluated.
Furthermore, we now present strong evidence that many of
the mouse CEA-related gene family members are produced
through alternative splicing of one gene. cDNA clone 23. used
in the reconstruction of the hybrid mmCGMla clone, displayed
the N1-terminal domain which is different from the N2-terminal
domain encoded by mmCGM1b. However, clone 23 encoded in its
downstream region identical A2 , transmembrane and
intracytoplasmic domains and the same 3' untranslated region

..
as nunCGM1b, but did not contain the
immunoglobulin domains found in clone 46.
49
Al and Bl C2-set
These results hdve
also been confirmed by performing reverse transcription dnd
PCR amplification with NI and N2 -terminal oligonucleotides (K.
McCuaig, manuscript in preparation). The genomic structure of
aIl CEA/PSG family members, whether human or rodent,
encompasses the 5' untranslated region and a half-leader as
exon 1 and another half-Ieader associated with an N-terminal
domain as exon 2 (36, 37). The structures of the many mouse
BGP-like cDNA clones, however, indicate that the mouse Bgp
gene would thus encode two N-terminal domains which are
aiternatively spliced to either the Al domain (as mmCGMla) or
to the A2 domain (as clone 23). Alternative splicing has been
reported to occur in the human BGP gene (12, 13) but this
splicing involves addition or removai of the A2 domain or
inclusion of a short 60 bp exon to produce a long
intracytoplasmic tail. Recent characterization of three new
human BGP splice variants has aiso shown that exons otherwise
represented by intron sequences are spliced to the Bl domain
to produce proteins with a hydrophilic structure (38).
However, only one N-terminal domain has so far been found for
human BGP (12, 13, 38).
We have aiso isolated and sequenced a great number of
cDNA clones which indicate that variability of CEA-related
gene transcripts and proteins is observed in the mouse and
that this variability is ensured through both al ternati ve

:r \
50
splicing and differential usage of polyadenylation signaIs
(manuscript in preparation). The functional significance of
this diversity is presently being evaluated.
The number of transcripts detected in colon, uterus and
liver aiso provides proof of the existence of a large family
of related prot.eins ln the mouse. When clone 46 was used as
a probe, it detected at least four transcripts cloned from the
human colon carcinoma cell line (see Fig. 3A) i. e. a 1.8 kb
transcript known as human BGP (12,13), a 2.6 kb transcript
known to be human NCA (39-42), two transcripts of 3.4 and 3.9
kb winch are human CEA transcripts using different
polyadenylation signaIs (6, 7, 43). In mouse placenta, three
distinct transcripts are foundi this tissue is known to
express many of the pregnancy-specific-like transcripts (36,
37). In fact, in a collaborative effort with Dr. Wolfgang
zimmermann (Freiburg, Germany) to decipher the extent of the
rodent family, we have determined that the 2.65 kb transcript
in placenta corresponds to rnrnCGM5, a mouse pregnancy-speci fic-
like product.
We have assigned a size of 4.6 kb to the major transcript
found in colon using ribosomal RNA as markers. We had
previously determined, in a different buffer system, the size
of this transcript as being 3.8 kb (26). However, by
comparison with the documented size of the human CEA larger
sized transcript (3.9 kb), the major mouse transcript would be
approximately 4.0 kb. 'rhe rnrnCGM1a cDNA sequence is 1.786 kb.

l 51
Primer extension analyses (data not shown) have determilled
that there is at most an additional 120 bp of DNA at the 5'
terminus of this clone. We have also found using the RACE
Polymerase Chain Reaction technique (44) on first strand cDNA
that there exists a much longer 3' untranslated region at the
3' terminus. This complete cDNA would account for the 4.6 kb
(4.0 kb) transcript. This assignment is reinforced by the
hybridization of the Accl-EcoRl restriction fragment of the
mmCGMla cDNA (Fig. 3B, panel D): because there is no sequence
homology between the 3' untranslated regions of mmCGMla and
mmCGMlb, this probe is undoubtedly the most specifie for this
particular mmCGMla cDNA. On the other hand, Northern analyses
using fragments corresponding to a long intracytoplasmic tail
or a 3' untranslated region found beyond that of mmCGMlb (data
not shown) also indicate that at least three other transcripts
co-migrate wi th the major 4.0 kb transcript in colon. The 1.8
kb transcript detected in Fig. 3B most probably represents
mRNAs where the Al and B1 domains are spliced out or where
alternatively, differential polyadenylation signaIs are used
in the extended 3 ' untranslated region. Obviously, more
cloning is required to understand the complexity of this mouse
CEA-related gene family.
The pl20 protein, likely corresponding ta mmCGMla, is
localized at the cell surface after transfection of the
mmCGM1a sense-driven cDNA. Since human CEA, NCA, BGP and
mouse mmCGMlb function in aggregation assays as cell adhesion

52
molecules, we wished to assess whether the rrrrnCGMla protein
would also sel f -adhere in a hornotypic fashion. Our resul ts
establish that the mmCGMla protein functions as an adhesion
molecule in vitro and that inhibition of this interaction
occurs in the presence of a polyclonal antibody specifically
recognizin3 this proteine However, this interaction does not
require physiological temperatures or the presence of the
divalent cation Ca2+ as was the case with romCGMlb (27). The
mmCGMlb protein was expressed on a different cellular
background (L-cells) which may account for this discrepancy.
Contrary to what has been demonstrated with the cadherins,
another major adhesion family which strickly depends on
calcium for the interactions of its members (45), the calcium
dependency of this mouse CEA-related farnily member does not
require the presence of a long intracytoplasmic tail. As
mentioned above, the N-terminal domain confers the major amino
acid sequence di f ferences between the two cloned mouse family
members. In human CEA and NCA homophilic and heterophilic
interactions, this domain is thought to be of significant
importance (46). The mmCGMla protein behaves like the cell
CAM 105 protein which has been shown by Aurivillius et al.
(47) to be the rat equivalent of the human BGP, and, by
inference, of the mouse CEA-related gene family members. The
cell-CAM 105 protein has been well characterized as a Ca2+
independent cell adhesion molecule involved in the aggregation
of rat hepatocytes (35). Similar]y, the expression of mmCGMla

1 53
transcripts i s consistent wi th that of a cell adhesion role III
vivo (48). Experiments are presently under way to verify
these hypotheses.
MATBRIALS AND METHODS
Cell culture and trans fect ions : Cells of the human colonie
adenocarcinoma line LS-180 (49) 1 a line of L-strain mouse
fibroblasts (LTA) and a mouse embryonic fibroblast cell line
(NIH3T3) were grown at 37°C in monolayer cultures in (X-MEM
medium (GIBCO) supplemented with 10% fetal bovine serum
(GIBCO) and 2 mM glutamine. rnrnCGMla-producing transfectant
cells were obtained by calcium phosphate-mediâted
coprecipitation (50) into 5xl0 5 NIH3T3 cells of 5 J..Ig of the
rnrnCGM1a cDNA inserted in either the sense or antisense
orientation of the p91023B expression vector (31) 1 âlong with
o .5 Jlg of the dominant selectable marker pSV2Neo (51), and 10
Jlg of mouse carrier genomic DNA . Transfectant clones were
selected with medium containing 500 J..lg/ml of active geneticin
(G418, Gibco) and evaluated for production of mmCGM1a mRNA and
protein by Northern and Western analyses.
Isolation of murine CEA cDNA clones: cDNA was synthesi zed as
previously described (6, 27). To isolate the 5' end of the
rnrnCGM1a cDNA clone, the library was rescreened using an EcoHl
Sst1 and dn Acc1-EcoR1 restriction fragment of clone 46. A
full coding cDNA was reconstructed by ligating over lapping

54
fragments at the BamHl site.
Reverse Transcription and Polymerase Chain Amplification:
Reverse transcription was performed using AMV reverse
transcriptase (Pharmacia) and 10 ~g of total mouse colon RNA
as template (44). The oligonucleotide primer used in this
reaction was a (dT) 17 adaptor with three restriction sites
(GACTCGAGTCGACGGTACCCTJ7 ). The reaction was incubated for 2
hrs at 37 Ù C in 50 mM TrisCI pH 8.3 (at 42°C), 8 mM MgCI 2 , 30 mM
KCI, 10 mM DTT, 20 U RNasin (Promega) and 1. 25 mM of each
deoxynucleotide (dATP, dCTP, dGTP, dTTP). The reverse
transcription product was desalted and purified from excess
dNTPs using a Centricon-100 microconcentrator (Amersham). One
tenth of the eluted DNA was used for PCR amplification. The
primers used for amplification were located in the N-terminal
domain of mmCGMla (nt 430-448) (CCAAATGATCACCATGMG) and in
the A2 domain (nt 1240-1273:antisense)
(CCGGCATCTTCCCTCTTAATAGGGTCTATTCTG) . The N-terminal
oligonucleotide was previously shown to be specific to the Nl
terminal domain versus the N2-terminal domain. The reaction
was incubated in 20 mM TrisCI pH 8.8 (at 24°C), 10 mM KCI, 2
mM MgS04 , la mM ammonium sulphate. 0.1% Triton X-IOO, 0.1 ~g
BSA, 0.2 mM of each dNTP, 40 pmoles of each primer using Vent R
DNA polymerase (New England Biolabs). The peR products were
agaln desalted on Centricon-100 microconcentrators and
submi t ted to sequencing using 5' labelled oligonucleotides
(52) •

1 55
DNA eequence determination and analyses: Mouse mmCGMl a cDNA
restriction fragments were subcloned into unique sites of the
BlueScript SK+ plasmid (Stratagene). 'l'Wo J.lg of plc1smi d DNA
were denatured by a 10 min incubation ln 0.2 N NdOH followed
by neutralization with 0.3 N Na-acetate pH 5.2 dnd eUlc1nol
precipitation. 30 ng uf either T7 or T3 promoter primers were
annealed to the DNA to sequence both st rands by the dideoxy
chain termination method (53) using T7 DNA polym~rdse
(Pharmacia). Sequences were analyzed using the DNAsis, Prosis
(Pharmacia) and Devereux (54) programs.
32P-labelled probes: Restriction fragments used as
hybridization probes for Northern analyses were ext racted (rom
0.8% low mel ting point agarose gels and labelled wi th (1- 1)[>
dATP (Arnersham or NEN Dupont) using the random primer
technique (55). The fS-actin probe used is cl 1.5 kb Pst l
fragment of chicken fS-actin (56). Oligonucleotjdes were
synthesized on a Pharmacia synthesizer, purified and 5' end
labelled using y'j/P-ATP (Amersham) and T4 polynucleot ide kirldSû
(Boehringer-Mannheim) .
RNA preparation and Horthern analyses: RNA was prepdred from
CD-l mice as previously described (27). Total RNA WdS
electrophoresed ln 1.5% agarose gels contc1inlng 1.1 M
formaldehyde, 10 mM MOPS buffer, pH 7.4, and 1 mM EDTA,
stained with acridine orange and transferred te Hybond-N
membranes (Amersham). Hybridization with cDNA restrlctio[l
fragments was performed for 18 h at 42°C in 5X SSPE (lX SSPE

56
is 0.18 M NaCI, 1 mM EDTA, 10 mM sodium phosphate, pH 7.7), lX
Denhardt' s solution, 50% formaTilide, 150 ~g/ml heat-denatured
salmon testis DNA, 2 mM SO(ÜUll1 ph0~phc:te, 0.1% SDS, and 10%
dextran sulphate with 1.0x106 cpmiml of the 32P-labelled
probes. The filters were washed at a final stringency of O.lX
SSC and 0.1% SDS at 65°C. Alternatively, for hybridization
with oligonucleotides, the gels were transferred to GeneScreen
plus ~embranes (NEN Dupont) and hybridized at 42°C for 18 h in
5X SSPE, 10X Denhardt's solution, 7% SDS, 10% dextran
sulphate, 100 J.1g/ml of carrier tRNA and 2X10 6 cpm/ml ot 32p_
labelled oligonucleotide (39). The filters were washed to a
final stringency of 0.1 SSC and 0.1% SDS at either 20°C or
50°C. 18S rRNA (1.86 kb) and 28S rRNA (4.71kb) were used as
markers.
Protein purification and generation of antibodies: Sixt Y
remale CD-l mice (retired breeders, approximately 40 9
supplied by Charles River) were sacrificed by cervical
dislocation. Colons were collected on ice in PBS (10 mM sodium
phosphate pH 7.4, 154 mM NaCl) containing protease inhibitors
(aprotinin 2 Ilg/ml, leupeptin 5 Jlg/ml and pepstatin 0.4 Jlg/ml;
Sigma and Boehringer Mannheim). The tissue was coarsely cut
up with scissors, then swollen in ten times its weight of 1 mM
Znel; for 10 min at 20°C followed by 10 min on ice. It was
then disrupted by 5 X 3ü-second bursts of a Polytron
homogenizer (Brinkmann) set at 60% maximum intensity,
interspersed by 30 s cooling periods on ice. The homogenate

1 57
was centrifuged at 1000g for 5 mIn and the pellets were
reextracted twice in the same conditions. Superndrants were
po01ed and centrifuged at 31, OOOg for 1 h. The membrane
pellets were pooled, wdshed with 50 mM TrisCl pH 7.4,
centrifuged at 31,000g for 30 min and the proteins were
solubilized in 10% Lubrol PX (Sigma) by stirring on lce for 30
min. Undissolved material was removed by d 30 min
centrifugation at 31, OOOg and the supernatants were stored
fro?;en at -80°C.
Protein extracts from ten colon membrane preparat ions
(equivalent to 600 mice) were pooled and assayed for protein
concentration according to Peterson (57). The 400 ml solution
containing 800 mg of proteins was acidi f ied to pH 3.0 by
addition of concentrated HCl and swirled dt 20°C for 3 mIn
with 32 ml of Polybuffer exchanger 94 gel (Pharmdcia)
previously equilibrated to pH 3.0 with Polybuffer 74
(dilution 1:8), 1 mM CaCl 2 and 0.1% Lubrol dccording to Lln
and Fain (58). The gel pellets were pooled, washed 5 times
with the equilibrating solution (approx. 1 1), then with 50 mM
TrisCl pH 7.4 containing 0.1% Lubrol untII the pH of the
supernatant reached 7.3 (approximately 500 ml of buf fer). The
proteins were eluted fram the polybuffer exchanger colurnn (1
cm x 50 cm) at 20°C with a 0 ta 0.65 M linear gradient of NaCl
in the equilibrating solution at 14 ml/ho The mouse p120 CEA
related protein was localized by Western analyses of each
fifth fraction with a rabbit anti-hurnan CEA antibody (:.;ee

58
below). Positive fractions were pooled, TCA-precipitated as
described below and resolved by SDS-PAGE on a 5-15%
SDS-acrylamide gel. The band migrating at 120 kD was revealed
in the Coomassie blue-stained gel, excised and electroeluted
for 5 h. The eluate was collected, dialysed against H20 for
24 h at 4°C and finally lyophilized. 'l'he protein powder was
suspended in sterile H/O. Aliquots of 150 ~g were mixed with
150 ~ l of RIBI ad] UVdnt (containing TDM: trehalose dimycolate,
MPL: monophosphoryl lipl.d A, and CWS: cell wall skeleton, from
Cedar Lane) and injected into the popliteal lymph nodes of
rabbits. The rabbits were boosted at two week intervals with
75 ~g of the p120 preparation in Freund' s incomplete adjuvant.
Western analyses: Diluted protein samples were precipitated
with 0.2% Lubrol and 12% TCA on ice for 30 min and centrifuged
at 12,OOOg for 15 min at 4°C. The pellets were washed twice
with ice cold acetone, dried in a SpeedVac evaporator and
boiled in SDS-PAGE Laemmli sample buffer for 10 min. The
proteins were resoIv8J by electrophoresis in 5 to 15%
SDS-acrylamide gels (59), transferred to a ni trocellulose
membrane according to Towbin (60) and then probed with a
rabbit anti-human CEA antibody (MCC: generously provided by
Dr. A. Fuks, McGill Cancer Centre) at a 1/100 dilution or with
d rabbit antl.-mouse p120 antibody at a 1/1000 dilution (serum
231). The purified p120 protein was aiso probed by Western
analysis with an anti-E-c;ldherin antibody (DECMA, Sigma).
Alternatively, transfectant cells were grown in a

1 59
monolayer to confluency, washed twice with PBS and lysed by
sonication in PBS with protease inhibitors at 4°C. After
determination of total protein concentration (57), aliquot 5 of
300 fig of proteins were precipitated in 0.05% Triton X-IOO and
12% TCA and resolved as above. Specifie glycoproteins were
visualized by inununoelectroblotting (60) using the rabbit
anti-mouse p120 antibody at a 1/1000 dilution (serum 231).
Alkaline phosphatase-conjugated secondary antibodies were
used. Fab fragments of the anti-mouse p120 antibody were
prepared by digestion with papain using an ImmunoSelect kit
from Gibco-BRL. These Fab fragments had the same specificity
as complete antibodies as judged by Western analyses.
zmmunoprecipitation: Proteins were immunoprecipitated for 18
h at 4° C to ensure aIl desired proteins precipitate, with
either 90 fig of an IgG fraction of a rabbit anti-human CEA
antibody or alternatively with a rabbit anci-mouse pJ20
antibody (serum 231). After gel electrophoresis and
immunoblotting, the membranes were processed for Western
analyses. The immunoprecipitated proteins were revealed by
reaction with l2SI-labelled protein A and exposure to Kodak
XAR film for 18 h.
zmmunofluorescence: Transfectant cells or parental NIH3T3
cells were lifted in a PBS-citrate solution without trypsin
and washed twice 1n PBS solution. Cell suspensions were
incubated for 1 h on ice with either immune serum (serum 231),
purified IgG from the same serum or pre-immune serum at

60
dilutions of 1/200 or 500 and washed four times with a PBS
solution. Cell pellets collected after centrifugation were
resuspended in 50 ~l of a 1:10 dilution (6.4 ~g/sample) of
dichlorotriazinylaminofluorescein (DTAF) -conjugated goat anti
rabbit IgG (Jackson Immuno Research Laboratories, Mississauga)
and the suspensions were gently mixed in the dark for 1 h.
After three washes in a PBS solution, the cell pellets were
resuspended in 20 ~l of a glycerol-PBS (9:1) solution and
observed with a Leitz Orthoplan flu~rescence microscope.
Aggregation assays: Parental, transfectant sense (86 and 811)
and antisense (A3 and A4) cell lines were grown ta confluency
and harvested by a light trypsin treatment (0.12%) in
phosphate-buffered saline lacking Mg Î+ and Ca2
+ and containing
15 mM sodium citrate. Suspensions at a concentration of 2x10 6
cells/ml were rendered single cells by three passes through a
27-gauge needle and then stirred at 100 rpm at 37°C. Samples
were taken over a 2 h incubation period and the percentage of
total cells remaining as single cells was measured using a
hemocytometer (21). The cells were suspended in 3 ml of
ei ther (X-MEM (Ca21 concentration, 2 mM) containing 0.8% fetal
bovine serum and 10 I1g/ml DNaseI, or Puck' s saline (21)
containing 0.8% of fetal bovine serum and 10 ~g/ml of DNaseI
with or without 10 mM Ca2+ added. Human CEA transfectant cell
clone 17-16 was used as a control in these experiments (21).
To test for p120 aggregation specificity, inhibition of
adhesion was performed with cell suspensions of mmCGM1a sense

61
and antisense-driyen transfectants resuspended dt 4xlO~
cells/ml in (X-MEM containing 0.5 mg/ml of Fab fragments of the
rabbit anti-mouse p120 antibody described aboye. The
suspensions were mixed at 4°C for 10 min, diluted to 2xlO"
eells/ml with a-MEM containing 0.8% of FBS and 10 J..lg/ml of
DNaseI and aggregation kinetics were measured as aboye. Each
aggregation asssay in aIl conditions was repeated a minimum of
three times.
ACItNOWLBDGBMBNTS
The authors wish to thank Drs. A. Herscovics dnd A. Fuks
for critical reading of the manuscript. We are also greatly
indebted to Dr. Abe Fuks for his kind gi ft of the Mec
antibody, Dr. C.P. Stanners for the human CEA transfectant
cell clone and Dr. André Veill~tte for the NIH3T3 cells. This
research is supported by the Medical Research Council of
Canada and the Cancer Research Society Ine. K.M. is a
recipient of a studentship from the Cancer Research Society
Ine. and the Medical Research Council of Canada. N.B. is a
Seholar from the Fonds de la Recherche en Santé du Québec.

62
RBFBRBNCBS
1. Gold, P. and Freedman, S .0. Specifie careinoembryonie
antigens of the human digestive system. J. Exp. Med. 122:
467-481, 1965.
2. Shuster, J.D., Thompson, M.P., Fuks, A. and Gold, P.
Immunologie approaches to diagnostic of malignancies. Prog.
Exp. '.'umor Res. 25: 89-139, 1980.
3. Paxton, R.J., Mooser, G., Pande, H., Lee, T.D. and Shively,
J.E. Sequence analysis of carcinoembryonic antigen:
Identification of glycosylation sites and homology with the
immunoglobulin supergene family. Proc. Nat!. Acad. Sei.
U.S.A. 84: 920-924 1987.
4. Oikawa, S. , Imajo, S., Noguchi, T. , Kosaki 1 G. and
Nakazato, H. The earcinoembryonic antigen (CEA) contains
multiple immunoglobulin-like domains. Biochem. Biophys. Res.
Comm. 142: 634-642, 1987.
5. Shively, J.E.and Beatty, J.D. CEA-related antigens:
molecular biology and clinical signifieance. Crit. Rev. Oncol.
Hemdtol. 2: 355-399, 1985.
6. Beauchemin, N., Benchimol, S., Cournoyer, D., Fuks, A. and

1 63
Stanners, C.P. Isolation and characterization of full-length
functional cDNA clones for human carcinoembryonic dntigen.
Mol. Cell. Biol. 7: 3221-3230, 1987.
7. Oikawa, S., Nakazato, H. and Kosaki, G. primary structure
of human carcinoembryonic antigen (CEA) deduced from cDNA
sequence. Biochem. Biophys. Res. Comm. 142: 511-518, 1987.
8. Zirmnermann, W., Ortlieb, B., Friedrich, R. and von Kleist,
S. Isolation and characterization of cDNA clones encoding the
human carcinoembryonic antigen reveal a highly conserved
repeating structure. Proc. Natl. Acad. Sei. U.S.A. 84: 2960-
2964, 1987.
9. Kamarck, M.E., Elting, J.J., Hart, .J.T., Goebnel, S .. J.,
Rae, P.M.M., Nothdurft, M.A., Nedwin, J.J. arJd Barnett, T. R.
Carcinoembryonic antigen family: expression in a mouse L-cell
transfectant and characterization of a partial eDNA ln
bacteriophage lambda gt11. Proe. Natl. Aead. Sei. U.S.A. 84:
5350-5354, 1987.
10. Barnett, T. and I,immermann, W. Workshop report: Proposed
nomenclature for the carcinoembryonic antigen (CEA) gene
family. Turnor Biol. Il: 59-63, 1990.
11. Williams, A.F. and & Barclay, A.N. The immunoglobulin

64
superfamily: Domains for cell surface recognition. Ann. Rev.
Immunol. 6: 381-405, 1988.
12. Hinoda, Y., Neumaier, M., Hefta, S.A., Drzeniek, Z.,
Wagener, C., Shively, L., Hefta, L.J.F., Shively, J.E. and
Paxton, R.J. Molecular cloning of a cDNA coding biliary
glycoprotein I: primary structure of a glycoprotein
immunologically crossreactive with carcinoembryonic antigen.
Proc. Natl. Acad. Sei. U.S.A. 85: 6959-6963, 1988.
13. Barnett, T., Kretschmer, A., Austen, D.A., Goebel, S.J.,
Hart, J.T., Elting, J.J., and Kamarck, M.E. Carcinoembryonic
antigens: Alternative splicing aC'counts for the multiple rnRNAs
that code for novel members of the carcinoernbryonic antigen
family. J. Cell Biol. 108: 267-276, 1989.
14. Hefta, S.A., Hefta, L.J.F., Lee, T.D., Paxton, R.J. and
Shively, J.E. Carcinoembryonic antigen is anchored to
membranes by covalent attachment to a
glycosylphosphatidylinositol moiety: Identification of the
ethanolamine linkage site. Proc. Natl. Acad. Sei. U.S.A. 85:
4648-4652, 1988.
15. Takami, N., Misumi, Y., Kuroki, M., Matsuoka, Y. and
lkehara, Y. Evidence for carboxyl-terminal processing and
glycol ipid-anchoring of human carcinoembryonic antigen. J.

1 65
Biol. Chem. 263: 12716-12720, 1988.
16. Kolbinger, F., Schwarz, K., Brombacher, F" Von Kleist, S.
and Grunert, F. Expression of an NCA cDNA in NIH/3T3 cells
yields a 110K glycoprotein, which is anchored into the
membrane via glycosyl-phosphatidylinositol. Biochem. Biophys.
Res. Comm. 161: 1126-1134. 1989.
17. Berling, B., Kolbinger, F., Grunert, F., Thompson, LT.A.,
Brombacher, F., Buchegger, F., von Kleist, S., Zimmermann, W.
Cloning of a carcinoembryonic antigen gene family member
expressed in leukocytes of chronic rnyeloid leukemia pat ient s
and bone marrow. Cancer Res. 50: 6534-6539, 1990.
18. Watanabe, S. and Chou, J.Y. Isolation and characterization
of complementary DNAs encoding human pregnancy-specific lS,
glycoprotein. J. Biol. Chem. 263: 2049-2054, 1988.
19. Streydio, C. and Vassart, G. Expression of human pregnancy
specifie S1 glycoprotein (PSG) genes during placenta]
development. Biochem. Biophys. Res. Comm. 166: 1265-1273,
1990.
20. Borj igin, J., Tease, L .A., Barnes, W. and Chan W. Y.
Expression of the pragnancy-specific beta 1-g1ycoprotein genes
in human testis. Biochem. Biophys. Res. Comm. 166: 622-629,

'.
66
1990.
21. Benchimol, S., Fuks, A., Jothy, S., Beauchemin, N.,
Shirota, K. and Stanners, C.P. Carcinoembryonic antigen, a
human tumor marker, functions as an intereeIIuIar adhesion
molecule. Cell 57: 27-34, 1989.
22. Oikawa, S., Inuzuka, C., Kurokj, M., Matsuoka, Y., Kosaki,
G. and Nakazato, H. Cell adhesion activity of non-specifie
cross-reacting antigen (NCA) and careinoembryonie antigen
(CEA) expressed on CHO celi surface: homophilic and
heterophilic adhesion. Bioehem. Biophys. Res. Comm. 164: 39-
45, 1989.
23. Zhou, H., Fuks, A. and Stanners, C.P. Specificity of
intercellular adhesion mediated by various members of the
immunogiobulin supergene famiIy. Cell Growth and Diff. 1: 209-
215, 1990.
24. Leusch, H.G., Hefta, S. A., Drzeniek, Z., Hummel, K.,
Markos-Pusztai, Z. and Wagener, C. Escherichia coli of human
origin binds to careinoembryonie antigen (CEA) and non
specifie erossreacting antigen (NCA). FEBS Letters 261: 405-
409, 1990.
25. Cerni, C., Tatra, G. and Bohm, H. Immunosuppression by

67
human placenta lactogen (HPL) and the pregnancy-specific bet.-1
1-g1ycoprotein (SP1). Inhibition of mitogen-induced lymphocyt fi
transformation. Arch. Gynak. 223: 1-7, 1977.
26. Beauchemin, N., Turbide, C., Afar, D., Bell, lr., Raymond,
M., Stanners, C. P. and Fuks, A. A mouse analogue of the hUll1an
carcinoembryonic antigen. Cancer Res. 49: 2017-2021, 1989.
27. Turbide, C., Rojas, M., Stanners, C.P. and Beauchemin, N.
A mouse carcinoernbryonic antigen gene family member ls d
calcium-dependent cell adhesion molecule. J. Biol. Chem. 266:
309-315, 1991.
28. von Heijne, G. A new method for predictlng signa l
sequence cleavage sites. Nucl. Acids Res. 14: 4683-4690, 1986.
29. Thompson, J.A., Mauch, E.M., Chen, F.C., Hinoda, Y.,
Schrewe, H., Berling, B., Barnert, S., von Kleist, S.,
Shively, J.E. and Zimmermann, W. Analysis of the size of thp
carcinoembryonic antigen (CEA) gene famlly: Isolation and
sequencing of N-terminal domain exons. Biochem. Blophys. Res.
Comm. 158: 996-1004, 1989.
30. Lin, S.H. and Guidotti, G. Cloning and expression of d
cDNA coding for a rat liver plasma membrane ecto-NfPùse: thE'
primary structure of the ecto-ATPase is similar to that of rh(~

'f 1
68
human biliary glycoprotein 1. J. Biol. Chem. 264: 14408-14414,
1989.
31. Wong, G.G., Witek, J.S., Temple, P.A., Wilkins, K.M.,
Leary, A.C., Luxenberg, D.P., Jones, S.S., Brown, E.L., Kay,
R.M., Orr, E.C., Shoernaker, C., Golde, D.W., Kaufman, R.J.,
Huwick, R.M., Wang, E.A. and Clark, S.C. Human M-CSF:
molecular cloning of the complementary DNA and purification of
the natural and recombinant proteins. Science 228: 810-814,
1985.
32. Ozawa, M. and Kemler, R. Correct proteolytic cleavage is
required for the cell adhesi ve function of uvomorulin. J. Cell
Biol. 111: 1645-1650, 1990.
33. Rojas, M., Fuks, A. and Stanners, C.P. Biliary
glycoprotein, a member of the immunoglobulin supergene family,
Eunctions in vitro as a Ca2+-dependent intercellular adhesion
molecule. Cell Growth and Diff. 1: 527-533, 1990.
34. Tingstrom, A., Blikstad, r., Aurivillius, M. and Obrink,
B. C-CAM (Cell-CAM 105) is an adhesive cell surface
glycoprotein with homophilic binding properties. J. Cell Sei.
96: 17-25, 1990.
35. Ocklind, C. and Obrink, B. Intercellular adhesion of rat

1 69
hepatocytes: Identi f iCdtion of a cell surface glycoprot ein
involved in the initial adhesion process. J. Biol. Chem. 257:
6788-6795, 1982.
36. Thompson, J. and Zimmermann, W. The carcinoembryonmic
antigen gene family: structure, expression dnd evolut ion.
Tumor Biol. 9: 63-83, 1988.
37. Rebstock, S., Lucas, K., Thompson, J .A. and zirnmermclnn, W.
cDNA and gene analyses imply a novel structure for èl rdt
carcinoernbryonic antigen-related protein. J. Biol. Chem. 261):
7872-7879, 1990.
38. Kuroki, M., Arakawa F., Matsuo Y., Oikawa, S. Nakazato H.
and Matsuoka Y. Three novel molecular forms of bj l id ry
glycoprotein deduced from cDNA clorles from a human J eukocyte
library. Biochem. Biophy. Res. Comm. 176: 578-585, 1991.
39. Cournoyer, D., Beauchemin, N., Boucher, D., Bench imol, S.,
Fuks, A. and Stanners, C.P. Transcription of genes of the
carcinoernbryonic antigen family in malignant and nonmal iqnant
human tissues. Cancer Res. 48: 3153-3157, 1988.
40. Zimmermann, W., Weber, B., Ortlieb, B., Rudert, F.,
Schempp, W., Fiebig, H.H., Shively, J.E., Von Kleist, S. and
Thompson, J .A. Chromo s oma 1 localization of the

70
carcinoembryonic antigen gene family and differential
expression in various tumors. Cancer Res. 48: 2550 -2554, 1988.
41. Neumaier, M., Zimmermann, W., Shively, L., Hinoda, Y.,
Riggs, A.D. and Shlvely, J.E. Characterization of a cDNA
clone for the nonspecific cross-reacting antigen (NCA) and a
comparlson of NCA and carcinoembryonic antigen.
Chem. 263: 3202-3207,1988.
J. Biol.
42. Tawaragl, Y., Oikawa, S., Matsuoka, Y., Kosaki, G. and
Nakazato, H. primary structure of nonspecific crossreacting
ant 1gen (NCA) , a member of the carcinoembryonic antigen (CEA)
gene f ami ly, deduced f rom cDNA sequence.
Res. Comm. 150: 89-96, 1988.
Biochem. Biophys.
43.0ikawa, S., Inuzuka, C., Kosaki, G. and Nakazato, H. Exon
intron organization of a gene for pregnancy-specific 1S1-
glycoprotein, a subfamily member of CEA family: implications
for i t s characteristic repetitive domains and C-terminal
sequences. Biochem. Biophys. Res. Conun. 156: 68-77, 1988.
44. Frohrndn, M. A., Dush, M.K. and Martin G.R. Rapid
production of full-length cDNAs from rare transcripts:
amplification using a sinyle gene-specifie oligonucleotide
primer. Proc. Natl. Acad. Sei. U.S.A. 85: 8998-9001, 1988.

l l
71
45. Nagafuchi, A. and Takeichi, M. Cell binding function of E
cadherin is regulated by the cytoplasmic domain. EMBO J. 7:
3679-3684, 1988.
46. Oikawa, S. , Inuzuka, C., Kuroki, M. , Arakawa, F. ,
Matsuoka, Y., Kosaki, G. and Nakazato, H. A specifie
heterotypic cell adhesion activity between members of
carcinoembryonic antigen farnily, W272 and NCA, is mediated hy
N-dornains. J. B10l. Chern. 266: 7995-8001, 1991.
47. Aurivillius, M., Hansen, O.C., Lazrek, M.B.S., Bock, E.
dnd Obrink, B. The celi adhesion moelcule Cell-CAM 105 is an
ecto-ATPase and a member of the inununogiobulin superfami1y.
FEBS Letters 264: 267-269, 1990.
48. Huang, J.Q., Turhide, C., Daniels, E., Jothy, S. and
Beauchemin, N. Spatioternporal expression of rnurine
carcinoembryonic antigen (CEA) gene family members during
rnouse embryogenesis. Development 110: 573-588, 1990.
49. Rutzky, L.P., Tom, B.H., and Kahan, B.D. Biologicai <lnd
antigenic analysis of hurnan colon cancer celi clones. Pro!].
Cancer Res. Ther. 9: 135-145, 1984.
50. Parker, B.A. and Stark, G.R. Regulation of simian virus 40
transcription: Sensitive analysis of the RNA specit:'!s present:.

72
ear ly in infections by virus or viral DNA. J. Virole 31: 360-
369, 1979.
51. Southern, P.J., and Berg, P. Transformation of mammalian
cells to antibiotic resistance with a bacterial gene under the
control of the SV40 early region promoter. J. Mol. Appli.
Gen. 1: 327-341, 1982.
52. Newton, C.R., Kalsheker, N., Graham, A., Powell, S.,
Gammack, A., Riley, J. and Markham, A.F. Diagnosis of (ll
antitrypsin deficiency by enzymatic amplification of human
genomic DNA and direct sequencing of polymerase chain reaction
products. Nucl. Aeid Res. 16: 8233-8243, 1988.
53. Sanger, F., Nicklen, S., and Coulson, A.R. DNA sequencing
with chain-terminating inhibitors. Proc. Natl. Acad. Sci.
U.S.A. 74: 5463-5467, 1977.
54. Devereux, J., Haeberli, P. and Smithies, O. A
comprehensive set of sequence analysis programs for the VAX.
Nucl. Aeids Res. 12: 387-395, 1984.
55. Feinberg, A.P. and Vogelstein, B. A technique for
radiolabeling DNA restriction endonuclease fragments to high
specifie activity. Anal. Bioehem. 132: 6-13, 1983.

l \
i
73
56. Cleveland, D.W., Lopata, M.A., MacDonald, R.J., Cowdn,
N.J., Rutter, vLJ. and Kirschner, M.W. Number dnd evolutonary
conservation of (l- and fS-tubulin and cytoplasmic fS- dnd y-
actin genes using specifie cloned cDNA probes. Cel! 20:
95-105, 1980.
57. Peterson, G.L. A simplification of the protein assay
method of Lowry et al. which is more generally applicable.
Anal. Biochem. 83: 346-356, 1977.
58. Lin, S. H. and Fain, J. N. Puri f icat ion of (Cal. -Mg;' ) -ATPdse
from rat liver plasma membranes. J. Biol. Chem. 259: 3016-
3020, 1984.
59. Laemmli, V.K. Cleavage of structural proteins during thl'
assembly of the head of bacteriophage T4. Nature 227: 680-685,
1970.
60. Towbin, H., Staehelin, T. and Gordon, J. Electrophoretic
transfer of proteins from polyacrylamide gels to
nitrocellulose sheets: procedure and sorne applications. Proc.
Natl. Acad. Sei. U.S.A. 76: 4350-4354, 1979.

74
LEGENDS TO FIGURBS
FIG. 1 Restriction Map and Topology of mmCGMla.
A: Restriction analyses: S'UT: 5' untranslated region, L:
leader, N: N-terminal domain, Al, Bl, A2: C2-type
immunoglobulin domain, Li: linker, TM: transmembrane domain,
CytS: p'ltative short-tail intracytoplasmic domain, 3'UT:
3'untranslated region.
B: DNA and predicted protein sequen~es of mmCGMla. Underlined
residues: N-linked consensus glycosylation sequences. C' :
cysteine residues thought to form intrachain disulphide bonds.
protein domains are separated by arrowheads placed above the
DNA sequence. This sequence has been deposited with GeneBank
and has received the accession nurnber X15351.
FIG.2: Homology of mmCGlIla and mmCGlIlb with rat ecto ATPase
and human BGP.
The amine dcids are represented using the one-letter code.
Dots indicate identical residues. Dashes indicate absence of
sequence or insert ions to maximize alignment. Underlined
residues are N-linked glycosylation consensus sequences. The
first set represents the leader (L) sequence. The next two
sets are the N-terminal (H) domain. The next five sets are
the Al, 81 and A2 domains. The last two sets are the
transmenmrane (TM) and intracytoplasmic (Cyt) domains.

1 75
FIG. 3 Expression of mmCGM1~-related transcripts in murine
tissues:
A: Hybridization with the clone 46 probe: 20 J..1g of tot al RNA
was separated on a formaldehyde-agarose gel as descibed in
Materials and Methods, trans ferred to Hybond-N membranes dnd
hybridized at 42°C in a 50% forn\1.mide solution with l X 10"
cpm/mlof 32P-labelled clone 46 insert. The filter was washed
at 65°C in a 0.1 SSC + 0.1 % SDS solution and exposed ta Kodak
XAR film.
B: Hybridization with a 8-actin probe: The filter used above
was rehybridized at 42°C in a solution containing 50%
formamide with a 32P-labelled chicken g-actin probe and washed
at 65°C in a solution of 0.1 SSC + 0.1% SDS.
C: Hybridization with several restriction fragments or an
oligonucleotide: 10 Jlg of colon RNA was separated on
formaldehyde-agarose gels as described ln Materials dnd
Methods and transferred to either Hybond N membrdnes (panels
A, B, D) or GeneScreen Plus membranes (panel C). These
Northern s t:rips were hybridi zed us ing candi t ions clescrihed.
panel A: an EcoRl-Dde1 fragment represent ing the 5' UT, panel
B: an EcoRl-BamHl fragment representing the 5' UT, the leader
sequence (L) and 2/3 of the N-terminal domain (N), panel C: a
19-mer oligonucleotide with little cross-reactivity with
mmCGMlb and ,-allel D: a 60 bp Acc1-EcoR1 fragment specifie to
the 3' UT. These blots were washed in a 0.1 SSC ~ 0 .1% SOS
solution at either 20°C (panel Cl, 42°C (panels A and D) or

76
FIG. ,,: protein Bxpression
A: Immunoprecipitation and Western Analyses: 300 J.lg of mouse
colon membrane proteins, 3 J.lg of purified p120 mouse colon
protein or 300 J.lg of proteins from transfectant cell lyzates
were irnmunoprecipi tated with ei ther a rabbi t anti -human CEA
antibody (lanes 1-5, 12), a normal rabbit serum (lanes 6 and
11) or a rabbit anti-rnouse p120 antibody (lanes 7-10). The
proteins were separated on a SDS-PAGE and irnmunoelectroblotted
as described in Materials and Methods. The membrane was
probed with an anti-rnouse p120 antibody and the proteins
revealed by incubation with 125r-labelled protein A as
described in Materials and Methods. Lanes 1, 6, 7, 11, 12:
Inouse colon membrane proteins. Lane 2: purified mouse colon
p120 protein. Lanes 3 and 8:
proteins.
proteins.
proteins.
Lanes 4 and 9 :
Lanes 5 and 10:
A4 transfectant cell lyzate
86 transfectant cell !yzate
S11 transfectant cel! !yzate
B: Immunofluorescence: 511 sense transfectant cells were
lifted from a dish with a PBS-citrate solution. The cells
were first incubated with a rabbit anti-mouse p120 antibody at
a 1/500 dilution, then with 6.4 Jlg of DTAF-conjugated goat
dnti-rabbit IgG (see Materials and Methods) and cytospun
(Cytospin 2, Shandon) at 500 rpm for 5 min. The cells were

1 77
analysed for fluorescence using a Leitz Orthoplan fluorescenc~
microscope. Arrowheads indicate presence of p120 protein dt
cell surface. Magnification X 630.
FIG. 5: Adhesion Assays
A: 2 X 10 6 transfectant cells bedring the nunCGMld cDNA in tilt")
sense (S11) or the antisense (A4) orientation were resuspenct(~d
in (l-MEM medium containing 0.8% fetai bovine serum ëmd
incubated at 37°C with spinning for 2 hrs. Al iquots were
retrieved at time intervals and single cells were counted.
Alternatively, 511 sense transfectant cells were inC'ubated
with 0.5 mg/ml of Fab fragments of the p120 antibouy (+Pab).
B: 2 X 106 cells of the cell lines described in b: werp
resuspended in Puck' s saI ine in the absence (Sll-Ca2+) or
presence (A4, Sll + 10 mMCa2+) of 10 mM Cal' and incubdted dt
37°C for 2 hrs.
C: 2 X 10 6 of either the S11 and the A4 rnrnCGMla t rdns fectclnt
celis or the human CEA tranfectant celis (CEA) were incubated
in (l-MEM medium containing 0.8% fetal bovine serum at 4°C for
2 hrs.
Results are expressed as the number of single cells remainin9
divided by the total number of cells.

f ,
Figure 1:
A SUT L N At BI A2 uTM CylS
B
mmCGM1. ' --' ! , , 1 j il Il
) 1 .. = = .. Eu ~ üi a aa. ~~ :f t! ~~ !Il
done 46
clone 23
u~~~~mm~~~~~~~~~~~m~rn 1 Il [ l
1 Il GCC TCA GCA CA T CTC CAC III. GGG CAli GTT CCt TG; G;A GGA CT A tTG CTC ACA Get TCA 4AS AHlHKliQY'''lililllTAS
L~N 173 CTT TTA Gee Tet TG& AGC eCT Get Act ACT GeT GAA GTt ACe ATT GAG GCT GTG CCG CCC 24 l lAS " S , A T T A E v T 1 E A Y P ,
m~~WGAA~~~mmmmm~~~~rn~mGGA 4. Q v. A [ D Il Il V l l l V Hill PLA l G
m~~~~mWGGA~ACTG~m~WGAAmGCArum~ 64AF AIIYK'~AIDKE IAA FV
mru~w~m~m~~~~m~~~~mmm~ e4'" SIIIILL..lliQAYSGA E Il YS
4U MT GGA TCC CTIO CTC TTC CM ATG ATC Act ATIi AAIi GAT ATG GGA lOTe TAC ACA eTA GAT 104 IL-i...-j l l F Q " 1 T Il K D Il G V Y T l 0
~Al mm~WGAA~mruwACTCAIi~~m~mw~~~m mllT DEIIY •• TQATVAF HYH'I
mru~wœ~mACA~~~ru~~m~w~~ru~ 144 l l t( , IL-L..l S L..L..S Il , V EGO D S v
m~mACCm~ru~~~ruw~m~mru~~~~ 164 S l T , D S y T D , D " l " Y l Il SR"
m~~~mruGAA~~~mAAlirum~~w~~mACT 184 G [ S l S [ , D • L K L S E G L.L..l l T
7U TTA CTC MT GTC ACIi AIiIi AAT ~ ACA Ga ccc TAT GTIi TGT GAA ACC CGIi MT CCA GTG 204 l l LL-I A L.Il.-l G , Y V C: E T A Il f' v
~B1 m~~~w~~rurn.m~mmmm~~ACCrum 224 5 V L..L....l 0 , F S Lili 1 Y li 'DT , 1
mm~m~wmmmwruGGGru~m~ru~mwGCA 244 1 S , 5 D 1 Y !. " , CS" L Il L 5 C. H A
m~~~rum~CAIi~m~rnm~~AAlimw~ru~ 264 A Sil' , A Q y f Il LIli E Il , " A S S
mCM~mmmœ~mACTACTw~_""ACCruACCmrnm 284Q E Lf 1 'IL..L...1T~liT YT,CiFY
~A2 m3~wrum~~m~~Act~mAAli~m~mrn~ru 304 " " 5 V T , l S • T T v It L..L.-I v lEP
3'UT
ü u <
112 3
172 ZJ
232 43
292 63
352 83
412 103
412 123
532 143
S92 163
652 183
712 203
712 2Z3
8JZ 243
892 21i3
1152 ZI3
la12 303
la72 323
1073 liTIi ACT CAli ccc TTC CTC CM liTt ACe ~ ACt ~ liTt III. GAA eTA ~ TCT GTG ACt 1132 lZ4 y T Q , F L Q v T lL..L...1 v Il E L D S V T 343
1U3 CTIi ACe Tee TTIi TCG AAT ~ ATT Ga '" IN. ATC ~ TGIi CTC TTt AAT AGe CAG AGT 1192 lU l T " l SilO l'A " 1 Q Il L F " S Q S 363
1191 CTT CAli eTC ACA GAIi ~ ATG ~ CTt ttc ~ ~ ~ ~ ATC ru ~ ATA ~ ceT 1252 364lQlTEAIITLSQL...IL..iI LAlO' 3e3
~m~AliliGAAW~~~mCAlimGAAmrum~m_mAlili 384 1 l A [ DA' [ y Q $. E 1 S " PYS y R
.... L. 1313 AIiIi AIiC AM: TCA ATC MG CTG GAt ATA ATA m ~ CCA ACA CM GGA Gee CTC TCA GAT 404 R S " S 1 II: L D 1 1 F D P T q G & l S 0 .,TM ~~~rn~~mmmGarnm~GGG~~rnm~~rn 424 li Il 1 A li 1 CV l , , V A li Y ALI A 10 L
_~ .vt 1433 ~ TAT TTC eTC TAT Tce AIlIi AAIi TCT Gee GIiA TCT liQ( Tet TTC TGA CM CTe Tee TAA 444 A Y Fl YSRItS"SCSF·
1312 403
1372 423
1432 443
1492 458
1493 CM GGA TCT CAC CAT ~ GeC TT; "" Act m GAA CTA T'T ~ CM AGT TG& ceT TGA 1552
1553 ACT GTG GeC ATA CAC TT' ceT TAIi CCT TTC ATIi TGe TG; GAT ~ AM TGT GTG eTA eCA 1612
1613 CAC CM GeT ~ AM AGT ATT eTT GAA ~ ACA CM CTG TGA AAT CCA GTA TG& GTe TeT 1672
1673 ACT ceT CM CAC TGe ACA GAA AIiA CAli ACT GGT CM TG& GTt CCA TGA GTC TAC TAC MG 1732
1733 AGT GTG TGT TGG MT TAT eTC TGe CCT 'TG GTT MT TTC TGG eTA TGA eTC CTA 1786

l Figure 2:
IIUIICGMls L IIUIICGMlb
ECTO b.BGP
IIUIICGMl a IIUIICGM1b ECTO
N b.BGP
IIUIICGMl s IIUIICGMlb ECTO hBGP
IIUIICGHl a IIUIICGMlb ECTO
A1 hBGP
IIUIICGMl a IIUIICGMlb ECTO
81 hBGP
IIUIICGMla IIUIICGM1b EC'l'O
A
BGP
IIUIICGMla IIUIICGHlb EC'l'O hBGP
IlCGHla TMJIIIIICGMlb
EC'l'O hBGP
Cyt
JIIIIICGMl a IIUIICGM1b ECTO hBGP
EC'l'O BGP
MELASAHLHKGQVPWGGLLLTASLLASWSPA'l"l'A • ••••.•••••••.. v ••..•••••••••. P ••• • ••••• R. u .. 1 •• R ••••••••• T'YH •• L ••• • CHL •• P •• RVR ••• Q ••••••••• HF .HL ••••
EVTlEAVPPQVAEDNHVLLLVHNLPLALGAFAWYKGJa:'l:AIDKEIARFVPNSNH • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • .. • • PVSTNA. .. VIIQ • TGT..a.X Q •. VD •••• H.V.EKS •••• A •••• QEFQV.Y •••• T.LHP.S •••• YIRSD~ QL.T.SM.FH ••• GKE ••••••••• QQ.PGYS •••• BRVDGHRQ.VGYAIGTQQ
IICGQAYSGREIIYSH.G.SLLFQMITMlmMGVY'1'LDMTDENYRRTQATVRFHVH %'1' •• P.H •••• TV ............ l.RV. V •• T •••• lE ••••• F ••• 2 ..• 0 ••.• SK •• P .••••• T ............ F •• ~.ERA ••• SVF.QQFHPI.TS.O.R.Y ATP.I-.H •••• T •• P.Jw. •• l.~QL%.F •• LQVIICSDLVHEE .. GQ ••• y
PILLKPHl%SBKSHPVEGDDSVSLTCDSYTDPDNIHYLWSRHGESLSEGDRLKLSEG~
.A.O •• ~G~ •• M •• EPF ••• M.EP •• --~S ••••••••••••••. VTF .•• ~ PELP •• S. SS~ •••• DDA.Ar •• EPE.--<JD'rr ••• WI • Ima,.PVSP •• Q •• N . .&-A..L
L~PYVCET.RKPV~DPFSLHIIYGPDTPIISPSDIYLHPGSNLHLSC
• ••••• R. T .K. Y. E •• 1. ••• ATF 4..1..4' •• H .DV ••••• 1.. V ••• P ••••• Q •••• ~. • ••• S ••• .A.L.I. ••• B •• IQ ..... A~ ... VT • ....Jl:%. ......... T ....... T • YR ... A.a..a.!i •••
HAASHPPAQYFWLIHEKPHASSQELFIPIW:'1'lU:lSGTYTCFVHHSVTGLSR'l"l'VlQi.Il:V
• • D •••••••••••••• I.QT ••••••• S~ • ..a...a..L ••• 11. ••• ~ ............. ".A...I-L" F Y ••••••••• S •••• GTFQQ.T •••••• ~V~.S ••• HA~ ••• C~ ••• T.I.T
E---PVTQPFLQ~SV'1'LTCLSHDlGAHIQWLFHSQSLQLTERMTLSQHHSI Q--- ........ s .... "aI...I.I." ............................................................... ...LJP-.L ..
• ---••••• SI. l .............. G •••••• P .K. T. VSVR ••••••••••• D •••••• O~ .LSPV.AK.QlKASK •• TGDK.S.~.ST •• T.ISIR.P.KN ••• PSS ••• K •• G~
LRI~PIKREDAGEYQCEISHPVSVRRSHSlKLDII FDPTQG--GLSD •••••••••••.••.•••••••.• K •••..••••• • ••••••••••• D •••••••••• P • l • HP •••• v . P ••••• HS ••• E SIH-.V •••••• T.W •• VF •• I.~P.M.NVH YHALPQEN ••• P
GAIAGIVIGVVAGVALIACLAYPLY · ....................... . • •••••••• - •••••••• A •••••• •••••••••••• L ••• V •••• C .• H
SRKSGGSGSF
:::T::GSéHRoLTEHKPSTSSHNLGPSDDSP~DVSYSVLNFNAQQSKRPTSASSS -G. T .RASDQ ••••••••• V.H.TQDH.N.P ••• MHE. T •• T ••• E ••• PTQ •.••• P.
PT--ETVYSV'VKlŒ L.AT.II •• Z ••• Q
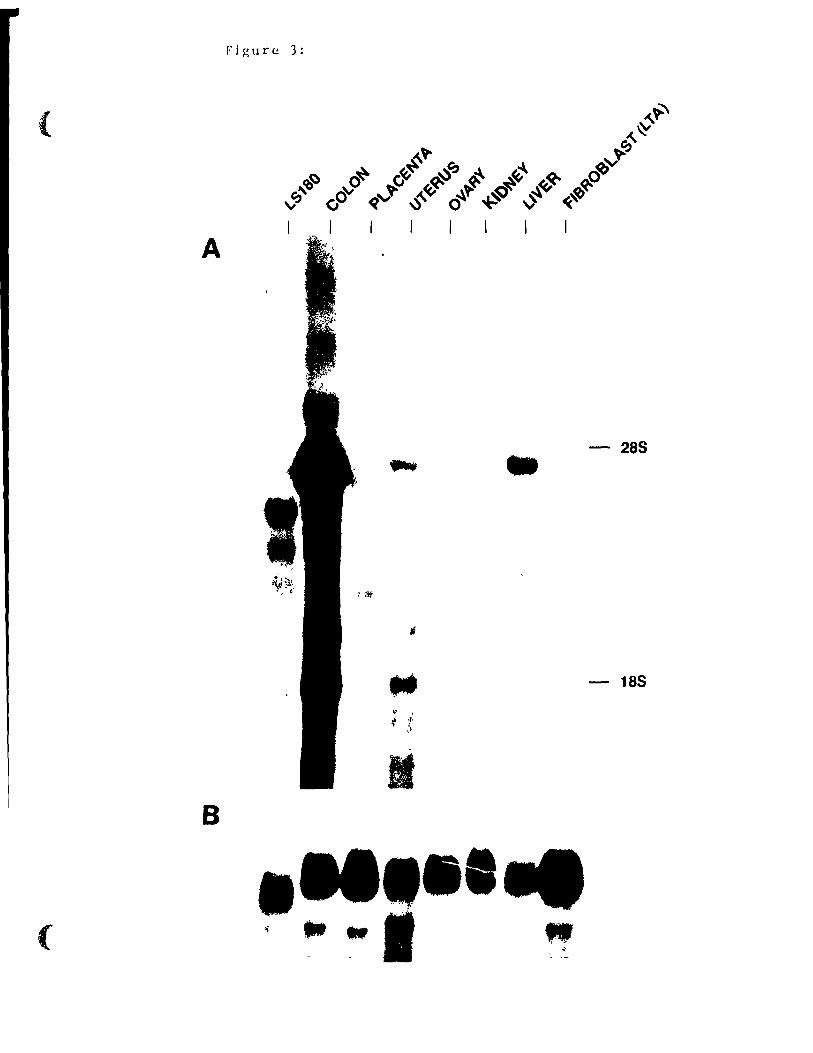
Figure 3:
{
A
- 28S •
- 18S
,
1 B
1 ( •• "

C
Figure 3:
S'UT L mmCGM1. : f----t 1
A
B
A
28S .. 18 S-
N Al 1 1 ,...
I,... 0 Eu c cau Ï co<!
C *
B
- .
91 A2 LiTM CytS 3'UT ri
1 1 ri
.... ........ ,... .... c- CT C'_ u (JI
~ ~/E u
Cf) <{
0
C 0
•

1 f f ••
-,.
A
kO 200-
Figure 4:
1 2 3 4 5 6 7 8 9 10 11 12
116 --'_ .• •
97 -
67 -
B
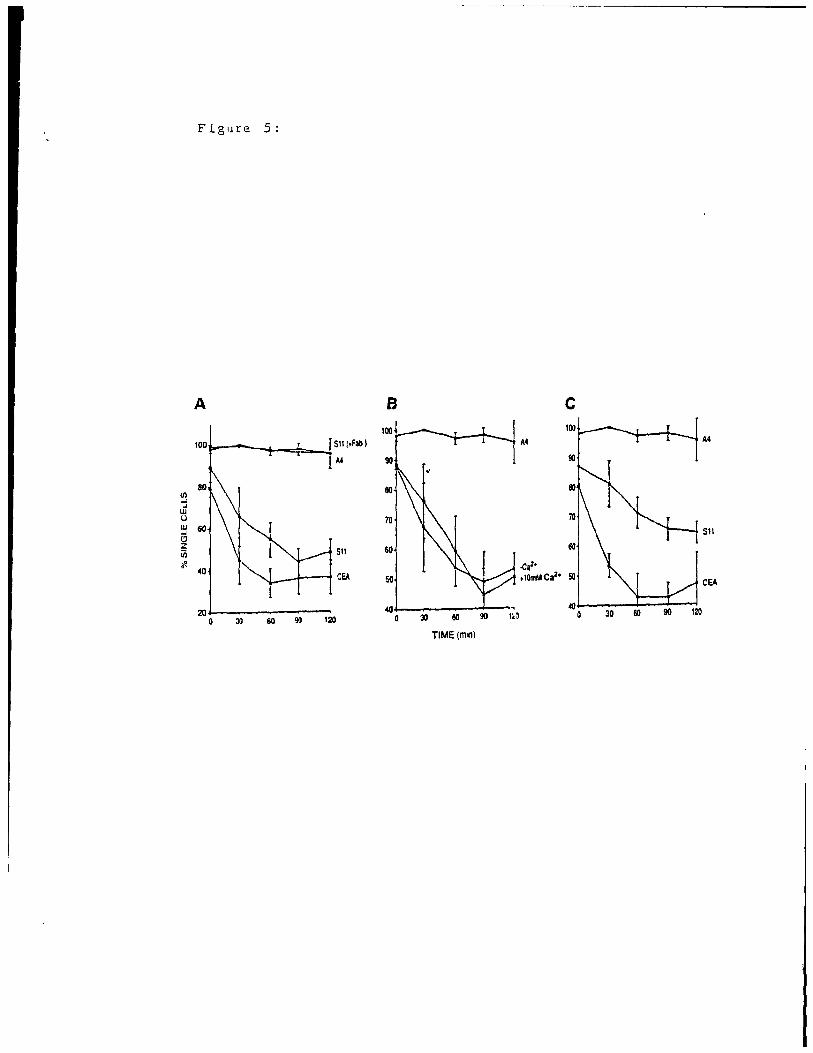
Figure 5:
A B C
100
100 A4 A4
A4
ffl ao
.... ~
W u 70 70 w 60 .... Sl1 Cl Z üi Sil 60 60
;ft .(;a2• 40 CEA 50 .'0mMCaZ, 50
CEA
20 40 ~
0 JO 60 90 120 0 J) 60 90 1<0 30 60 90 120
TIME(mln)

DISCUSSION AND CONCLUSION

78
DISCUSSION
Structure and Alternative Splicing:
When the existence of mouse analogs of hurnan CEA was
first reported in 1989, cDNAs encoding four different members
of the murine CEA family were identified (Beauchemin et al.,
1989) . This dissertation describes mainly the
characterization and function of mmCGMla. The previous
chapter reports two findings; first, that the mmCGMla mRNA is
generated by alternative splicing of one gene, and second,
that transfectant cells expressing mmCGM1a protein on their
cell surface exhibit calcium- and temperature-independent
adhesion in vitro (McCuaig et al., 1992).
The regulation of gene expression is often accomplished
by controlling the initintion of transcription, and therefore,
the abundance of transcripts present in a cella This
initiation involves cis-acting elements and trans-acting
factors which may activate or down-regulate transcription.
Post-transcriptional control mechanisms may also regulate the
level of protein product produced from one gene. One way to
!::"8yulate the amount of active protein produced, is to generate
proteins with similar or diverse functions by alternative
splicing of a single precursor mRNA.
Every mature rnRNA is generated by removing introns and
splicing together exons of a precursor rnRNA. In sorne pre-

1 79
mRNAs, the splicing pattern is constitutive while in others,
it is regulated. However, aIl splicing is performed by the
spliceosome, a multisubunit cornplex containing smdll nuclear
ribonucleotide particles (snRNPs) (Maniatls and Reed, 1987;
Sharp, 1987; Guthrie, 1991). U2AF is one of the early
binding factors in spliceosorne assembly and appear"s t 0 bind to
the introns of constitutively spliced sites more efficiently
than to those of regulated splice sites. Binding affinity is
higher with increased pyrimidine content and increélsing
polypyrimidine tract length. The relative binding affinities
of U2AF offers an explanation for the mechanism of splicing
regulation (Mullen et al., 1991; Smith et al., 1989).
There are several active genes identified where
alternative splicing of the primary transcript regulates the
activity of the protein product. For example, one splice
variant of erbAa is modified in the carboxy-terminal domain,
therefore preventing ligand binding (Koeing et al. 1 1989).
The different isoforms of erbAn exhibit a tissue-speci f ic
distribution and therefore different tissues have different
responsiveness to thyroid hormone. Fibroblast growth factor
(FGF) receptors may also exhibit differential splicing of the
prirnary transcript. There are four different FGF receptors
(FGFR1, FGFR2, FGFR3, and FGFR4) encoded by four distinct
genes. Recently, a second splice variant of FGFR1 has been
isolated; it is expressed mainly in the skin and has a fifty
fold lower affinity for basic FGF (Werner et al. 1 1992).

"
'.
80
In other systems, alternative splicing may remove part of
the activation domain, rendering them incapable of activation.
For example, mTFE3, a transcriptional activator that binds to
the enhancer of the immunoglobulin heavy chain gene, can exist
in either a Jong form or a truncated, inactive form. The
relative amounts of these two mRNAs varies in different
tissues, allowing regulation of transcription factor activity
(Roman et al., 1991). Another example is the cyclic AMP
responsive element modulator (CREM) which plays a role ln
mammalian spermatogenesis. In premeiotic germ cells, CREM is
expressed at low amounts in the antagonist form, lacking the
acti vating domain. Subsequently, in later stages of
spermatogenesis, alternative splicing generates an activator
form, CREMr (Foulkes et al., 1992).
Human biliary glycoprotein also undergoes extensive
alternative splicing (Barnet t et al., 1989); however, the
functional significance of this remains unknown. To date,
there have been at least eleven different splice variants
identlfied at the cDNA level (Barnett, personal
communication). The mmCGM proteins, by sequence homology, are
the mouse homologs of human BGP. The gene encoding mouse CEA
related family members exhibits alternative splicing patterns
similar to those seen for human BGP.
Through the screening of a mouse colon cDNA library, many
different transcripts were isolated dnd sequenced. Sequence
comparisons revealed llunCGMla-like transcripts with certain

Figure A:
L N1 A1a B1a A2a lITMCyt • ..
rnn-CGM1a 1 m·8*:œ§X§X«§}W Il • L N1 A1a 81a A2a li TM Cyt
rnn-CGM1d 1 t§.j«$$}§$$§8§!§$-@ Il
L N1 A2a UTM~yt
rnn-CGM1d iJ$$$--;,i§=t;:;2;g.:.:&g@ "
• L N1 A2a UTM cyr
rnrrC3M1g rra&Y:§§$§$!.$$2·m Il
L N2 A2b UTMCyt rnrrCGM1b 1 '~~:%%~~%~:%~~%%%~~~~%1~' Il •
L N2 A1b B1b A2b UTMCyt
rnrrCGM1e 1 I"""",,,.~,,,,,,,,,,.,,l Il •
L N2 A1b B1b A2b UTM cyr
mmCGM1f 1 1""""""""""",1 Il
L N2 A2b UTM Cyt
rnrrCGM1h f"""""""""",.,,;,j Il
* indicates one base pair change

'f
."
81
domains included or deleted. Sorne cDNAs were missing the
entire region encoàing t.he Al and B1 internal domains.
Transcripts containing an extra 60 base pairs after the region
coding for the transrnembrane domain were also identified.
This insertion into the mRNA results in a reading frame shift
during t ranslat ion, extending the cytoplasmic domain from 10
amino acids to 73 amino acids. As weIl, nunCGMla transcripts
possessing the 3' untranslated sequence of mmCGMlb (another
family member which has been previously reported) were
isolated. Data generated from our group and from Kay Holmes'
group suggest that there are, in fact, two different 5'
untranslated regions and two different 3' untranslated regions
possible for t.he nunCGMla transcripts, where the second 3'UT
i solated contains three di f ferent pOlyadenylation si tes.
In arder to cOllfirm the expression of these different
splice variants in mouse colon, analyses were performed using
polymerase chain reaction (PCR) techniques. A first strand
cDNA was synthesized by reverse transcription of total mouse
colon RNA. The reaction was initiated elther with a primer in
the 3'UT reported for mmCGM1b or with a poly-dT adaptor.
AmpliflC"ation of specifie cDNAs was achieved in a thermocycler
using VENT" polymerase (a heat stable DNA polymerase which is
less error-prone than the Taq DNA polymerase, Neuner et al.,
1990) and different pair combinat ions of oligonucleotides
specifie to different domains of the rnrnCGM1a or the mmCGMlb
cDNA . The amplified products were cloned and sequenced,

1 8:2
revealing transcripts with eight different codlng sequences;
four encoded the N-terminal domain sequence of nm1CGMla (N 1)
and its corresponding Ala, Bla, and A2a domdins, while th\;'
other four encoded the N-terminai domain sequence of nunCGM 1 b
(N2, which is 87% homologous to N1) and i ts correspondi nq
internaI domains (A1b, Blb, and A2b, which are 99% homolgous
to Ala, B1a, and A2b, respectively). The longest mdture mRNA
species (mmCGM1d or mmCGMlf) encode an N-terminal, Al, B1, dmi
A2 domains, a transmembrane, and a 73 amine acid cytopldsmiC'
domaine Other splice variants (mmCGMlg or mmCGMlh), have the
Al and B1 coding reg~ons spliced out. The transcriptn
designated mmCGM1a and mmCGHle have 60 bp deleted, result ing
in a mRNA encoding a 10 ami no acid cytoplasmic domaine The
minimum coding sequences (mmCGM1b and mmCGMlc) have the
regions encoding the Al and B1 domains as weIL as 60 bp uf t hf~
cytoplasmic domain removed. These cDNAs are represented in
Figure A. As weIL as alternatively spliced transcripts with
different coding regions, the 3' untranslated regions may he
different, thus affecting mRNA stabliity (Wilson and Treisman,
1988). PCR analyses by Ray Holmes' group demonstrated thêÜ
the longest 3' untranslated region is more abundant in the
liver while in the colon the shortest 3' untranslated sequence
is more prominent (Deiffenbach, personal communication).
The relevance of this alternative splicing and usage of
different polyadenylation sites is unknown; however, we dre
presently investigating if the relative amounts of eé1ch

83
species are tissue-specifie by RNAse proter:tion and by PCR
dna lyses. Tissue-specifici ty or
would suggest that each proteln
developmental regulation
product generated by
alternatlvely spliced rnRNAs has a different function necessary
in a spatially OI" temporally controlled fashion.
Adhesion Molecule:
Olle function for human BGP and the mouse CEA-related
EdITuly members has been elucidated. BGP, mrnCGMla, and
mmCGMlb, when expressed on the cell surface of transfectant
cells, mediated the formation of cell aggregates. BGP (Rojas
et al., 1990) and mmCGM1b (Turbide et al., 1991), however,
required calcium and physiological temperatures to function as
adhesl0n molecules while rnrr .. :GMla did not \4 '~r:uaig et al.,
1992). As prp.viously described, rnrnCGM1a and rnrnCGM1b possess
different N-terminal domains sharing 87% homology, and
rrunCGMla, unl ike rnrnCGMl b, expresses the Al and B1 domains. The
region conferring calcium dependency may lie in one of these
domains. This calcium dependency may also be an effect of the
cellular background or of the amount of protein expressed on
the cell surface; both rnrnCGMla and mmCGMlb were transfected
into mouse fibroblasts (rnrnCGMla into NIH/3T3, rnmCGMlb into LTA
cells) dnd recent data on human BGP adhesion properLies
suggests that high BGP expression on the cell surface of
transfectant cells abolishes the calcium dependency of
ddhesion (M. Rojas, unpublished data) .

84
Role in Tumor Formaticn:
Specifie cell adhesion governs morphogenesis in embryonu'
development and maintains tissue integri ty ln adult hoo<1.
Altered recognition and adhesion properties of cells mdy also
be important in varioùs disease processes such as i nVdS ion clnt!
metastasis of cancer cells. The adhesion molecule E-cadherln
has been shown to behave as an invasion suppressor. Tumor
cells expressing high levels of E-cadherin are less metastdt le
than those with low expression (Frixen et al., 199 ~) .
Transfecting E-cadherin into highly invasive eplthel id 1 t umOl
cell lines decreased invasiveness; however, the InvdsivenesG
could be restored by Lreatment with anti-E-cadherin dntibodl.efj
(Vleminckx et al., 1991). CEA expression lS also altered in
tumors, but unlike E-cadherin, the higher the eXpreS:ë-31orl, the'
more tumorigenic the tumor is (Jessup et al., 1988).
Malignant tumors ir1 mice also have al tered level s of CEA
like family members. Normal adult mouse colon produces large
quantities of CEA-like proteins; however, compdring the
expression of rnrnCGM transcripts in pr imary colon t umor fj to
that in adj acent normal mucosa, showed decreased express] on HI
tumors (N. Beauchemin, unpublished data). Furthermor~,
Northern analyses on tumor cell lines revealed thdt four mour;e
colon carcinoma cell lines (CT26B, CT36B, CT51G, dnd C'r51P)
had no detectable CEA-like transcripts and a mouse rer;td 1
tumor cell line, CMT93, expressed a 4.0 kb transcrlpt dt 10-20
times lower levels than normal colon (N. Beauchemin,

'.
85
unpubllshed data). In mouse mammary tumors, the opposite
pattern occurs. Normal mouse mammary tissue does not express
CEA related family members, however, chemically-induced or
rnurine mammary tumor virus-induced mammary tumors express high
levels of these transcripts (Robbins et al., 1991). The
s 19ni f icance of the change in expression of CEA-related family
members during turnor formation is nol known. In colon
cdrclnomas, lts change in expression pattern is suggestive of
é.l tumor suppressor role, while in mammary tumors, mmCGM
prott~ins appear to dct in tumor formation. An explanation
for this discrepancy may reside in the tissue of origin;
mmCGM protel.ns may have different functions in different
t issue types. As weIl, the manner in which the tumors were
induced may be important; tumor markers are specifie to
di Eferent cell types as weIl as to the virus or chemical used
ta induce tumor formation (Shuster et al., 1980).
Homologues in The Rat:
Other functions hypothesized for the mouse CEA related
fdmi ly members are based on observations found in the rat.
Identlcal proteins which have been independently isolated as
ppJ20 (Rees-Jones et al., 1985), HA4 (Hubbard et al., 1985),
('e11-Cell Adhesion Molecule 105 (Cell-CAM 105) (Ocklind and
Obrink, 1982), or rat hepatocyte ecto-ATPase (Lin and
Guidotti, 1989), are the rat homologues of human BGP and of
mousE' CEA gene fami ly members. The purification of each of

j 8t,
these proteins was achieved by exploit ing a function tn
ident; fy a protein. They wel. e lc1ter found ta be ident iCd 1
proteins by comparing expression pat terns, protel.n dE'<)raclat lOt1
patterns, antibody crossreacti vit y , and/or amino dcid or DNA
sequence (Margolis et al., 1988 and 1990; Lin et al., 1991).
The mouse CEA gene family memLers were isoJated uSlng d
!:everse stategy i a cDNA library was screened wi th cl probp
from the coding sequence of human CEA. Once cONAs wer p
isolated, the corresponding proteins were expr0ssed in tissue
culture permitting large scale production of the proteln of
interest, and allowing possible funct ions to be assiqned. The
rat homologues gave sorne indication of the factors to be
evaluated.
The first function to be assigned to mmCGM protei r1f;
represents that of an adhesion function which is in accorddnce
with the described function of the rat proteins. Cell-CAM 105
was originally purified from rat hepatocytes by looking for d
protein which neutralized the inhibition of ceU aggregdt ion
caused by multispecific antlbodies directed against the cel]
surface (Ocklind and Obrink, 1982). By then incorpordtinq
ceU-CAM 105 into liposomes, these liposomes could bind to
hepatocytes in suspension independent of calcium ions (Obri nk
et al., 1988).
Tc greater understand the functional slgnificance of j tr;
adhesion properties, the tissue dis tribut l.on and cell u] ar
locations of cell-CAM 105 were examined (Odin and Ohrink,

.> , 87
1987; Odin et al., 1988). It is expressed late in
development and does not reach the level seen in mature liver
until three weeks after birth (Odin et al., 1986) In
regenerating li ver, a transient decrease in the amount of
Cell-CAM 105 occurs (Odin and Obrink, 1986) and l.n rat
hepatocellular carcinornas, it is absent or significantly
chemically modified (Hixson and McEntire, 1989). Since cell-
CAM 105 is expressed at high levels only in normal adult rat
liver, it appears to play a role in the maintenance of tissue
architecture. In development, liver regeneration, or
hepatocellular carcinornas where cell-CAM 105 protein is
decreased or not expressed, the hepatocytes are not as tightly
bound to each other, allowing changes in the tissue
architecture to occur.
Cell-CAM 105 appears to function in the regeneration of
rat liver, however this protein is also expressed on a variety
of other tissue types such as the bile canaliculi of the
liver, epithelia of the stomach, t.he small intestine, the
large intestine, the tubules of the kidney, vessel endothelia,
plat.elets, and granulocytes (Odin and Obrin~, 1987; Odin et
al., 1988). The apparent molecular weight of the protein
varies dmong different tissues, however, it is not completely
known whether this is a result of alternative splicing or of
differentidl glycosylation. It was also noted that ceJI-CAM
105 was mainly expressed on epithelial structures but showed
varying cell surface distribution. It can be present in cell-

1 88
cell borders between adjacent cells, on the membranes of
densely packed microvilli, or ln intracellular sites,
depending on the tissue and cell-type (Odin et dl., 1988).
This variation between tissues suggests that Cell-CAM 105 Indy
function in a different manner in different tissues with SOIne
involvement of membrane-membrane binding in each case. For
example, ce11-CAM 105 on platelets could be involved in
platelet aggregationi unactivated platelets contdin eell-CAM
105 intracellularly, however, when indueed to aggregate in
vi tro, it becomes heavily exposed on the surface (Odin et al.,
1988) .
The mouse CEA-related proteins are also expressed in a
variety of tissues, including colon, placenta, uterus, ovary,
kidney, gall bladder, small intestine, and li ver in adult miee
(McCuaig et al., 1992) 1 and meninges, eart i lage and bone 1
blood vesse! walls, placenta, dermis, muscle layers of the
stomach and intestine, and bronchioles in mouse embryos (Huang
et al., 1990). Northern blot analysis dernonstrated that
expression of the major 1.8, 4.1, and 4.6 kb transcripts was
most abudant in the colon. It could also be noted thàt a
number of other transcripts were present 1 sorne being tissue
specifie. The major transcripts seen in al! of the expressing
tissues may have a universal role, while the tissue-specifie
transcripts may have unique functions.

'.
89
Blastocyst Implantation:
Another functional hypothe3is to be investigated in rats
is the involvement of cell-CAM 105 in early stage blastocyst
implantation. The expression of cell-CAM 105 at the apical
surface of rat uterine epithelium is controlled by ovarian
steroid hormones (Svalander et al., 1987). At the tirne of
blastocyst implantation, cell-CAM 105 is absent in the lurninal
epithelial cells but present in the glandular epithelial cells
of the uterus. Pre-implantation blastocysts exhibit a
transient expression on the trophectoderrn (Svalander et al.,
1987). At the time of implantation, a polarity with respect
to staining for Cell-CAM 105 occurs. It is lost or rnasked
from the surface of the mural tropnoblast cells which are the
first to adhere to the uterine luminal epitheliurn during the
onset of implantation and invasion of the uterine stroma. The
downregulation of cell-CAM 105 in both the luminal epithelium
of the uterus and in the trophectoderm of the blastocyst may
be linked to the acquisition of trophoblast invasiveness
(Svalander et al., 1987). Mouse CEA-related transcripts are
expressed in the uterus but whether or not this has an ef fect
on blastocyst implantation has yet to be examined.
Signal Transduction:
Phosphorylation is an important mechanism in protein
function regulation. The addition of phosphate may serve to
either up-regulate or down-regulate the activity of a protein;

1
------ - --------------
90
for sorne proteins, the phosphoprotein 1S the act ive form,
while for others, it is the inactive form. There are two
types of protein kinases; those that phosphorylate substrates
on serine or threonine and those that phosphorylat e on
tyrosine residues.
Human BGP (Afar et al., 1992) as weIl as the rat pp120
(Rees-Jones and Taylor, 1985) have been found to be
phosphorylated on intracellular tyrosine residues. The
protein which phosphorylates human BGP has not yet been
identified; however, pp120 is an endogenous substrate for the
insulin receptor (Rees-Jones and Taylor, 1985). Stimulating
H-35 hepatoma cells with insulin showed a ten fold increase ln
insulin receptor phosphorylat~on and a two fold incredse ln
phosphorylation of pp120; however, stimulation with epidermal
growth factor (EGF) or with insulin-like growth factor (lGF)
had no effect (perctti et al., 1987). In a cell free system,
however,
insulin,
where the corresponding
epidermal growth factor,
receptors are present,
and insulin-llke growth
factor 1 stimulate the phosphorylation of tyrosine residues of
pp120 (Phillips et al., 1987; Fanciulli et al., 1989). The
receptors for these factors have an endogenous tyrosine kinase
activity which acts on other cellular proteins and results in
activation of a signal transduction cascade. Once the signal
has reached the nucleus, the cell can respond appropriately.
Although phosphorylation assays have not yet been done on
the mouse CEA-like proteins, the splice variants with 73 amine

91
acid cytoplasmic domains contain two tyrosines which may be
substrates for tyrosine kinases, as weIl as a consensus
sequence for cAMP dependent serine phosphorylation. Because
of the homology between the mouse CGM proteins and the rat
cell-CAM 105, it is postulated that sorne of the rnrnCCGM
proteins are endogenous substates of the insulin receptor
tyrosine kinase, or of other growth factor receptors. To
examine this postulation further, cells expressing the
respective receptors will be transfected with cDNAs encoding
mmCGM proteins. These cells will then be stimulated with
either insulin, EGF, or IGF, and the change in phosphorylation
status of the mmCGM proteins will be determined.
Recent studies provide evidence of an ectoprotein kinase
activity on the surface of several types of cells, including
neutrophils (Skubitz et al., 1991; Dusenbery et al., 1988).
The role of this ectoprotein kinase activity in cell function
is unknown; however, the identification and characterization
of its physiologie substrates should aid in the understanding
of the role of this activity. A 180 kDa phosphoprotein on the
surface of human neutrophils which is one of the maJor
endogenous substrates for the ectoprotein kinase has been
isolated (Skubitz et al., 1992). In resting cells there is
only a small amount of the 180 kDa protein on the cell surface
This surface expression is augmented by stimulation by the
chemotactic peptide FMLP, the calcium ionophore A23187, and
12 -o-tetrade-conoyl-phorbol-13 -acetate. Since this protein is

1
--~-----------------------------
92
cross-reactive with human CEA, NCA, and BGP, it hels been
postulated to be a 180 kDa form of the clelssical NCA on human
neutrophils.
Therefore,
status of the
as well as evaluating the phosphorylat ion
cytoplasmic domain of mouse CEA-related
proteins, phosphorylation on the external doma:n should aiso
be stu,:1ied. The addition of a phosphate group to tyrosine
residues in the extracellular domain of the protein mdy affect
the stren~th of adhesion mediated by these molecules. It is
the protein backbone and not the carbohydrate residues which
is responsible for the adhesion properties of CEA gene-family
members (Zhou, personnal communication); however, the effect
of phosphorylation on intracellular or extracellular domains
is not known.
Calmodulin Binding protein:
Recently it has been
calmodulin binding protein
function of calmodulin
shown that Cell-CAM 105
(Obrink et al., 1988).
is to recognize changes
is a
The
in
intercellular concentration of the second messenger, Ca1t, and
to transmit this information to proteins inside the cell.
Calmodulin binds calcium, and as a result, undergoes
conformational changes which increase its affinity for target
proteins by two or more orders of magnitude (Cohen and Klee,
1988) . Calmodulin confers Ca2'-dependence to a variety of
intercellular processes, incl uding energy and biosynthet ic

93
metabolism, cell motility, exocytosis, and skeletal assembly,
by directly interacting wi th specific proteins. Rabbit
skeletal muscle myosin light chain kinase (MLCK) calmodulin
binding domain has been studied to determine the essential
residues for calmodulin binding. Calmodulin binding domains
have the following features: clusters of basic residues,
clusters of hydrophobic residues adjacent to the basic
residues, and the ability to form a-helices. These features
are aIl contained within sequences of less than 25 residues
(Cohen and Klee, 1988).
The ability of calmodulin to bind to the intracellular
domains of the mouse CEA-related proteins will be assessed.
A variety of approaches will be taken. First,
immunoprecipitating with an anti-mouse CEA-related protein
antibody on transfected cells, and determining what proteins
co-precipitatei second, running colon extracts over a
calmodulin-agarose column and assessing whether or not mouse
CEA-l ike proteins bind i third, attaching mouse CEA-like
proteins to a column matrix and then running a variety of
proteins through the columns, determining which ones are
retained.
The intracellular domain of the mouse CEA related
proteins bearing short intracytoplasmic tails will most likely
not interact with any cytoplasmic proteinsi the cytoplasmic
domain is very short, only 10 amino acids, and calmodulin
binding domains this short have yet to be identified. The

1 94
amine acid sequence of the cytoplasmic domain of the 10ng
tailed proteins does not show any striking homology to
calmodulin binding domains, however, the structure dnd not the
absolute sequence is important for calmodulin binding.
virus Receptor:
A wide variety of proteins belong to the immunog1obulin
superfamily. They are expressed on many different cell types
and perform diverse functions although they are aIl involved
to sorne extent in cell recognition. Immunoglobulins make up
a large portion of the proteins expressed on cell surfaces,
allowing them to be exploited as virus receptors. In humans,
the envelope protein of HIV binds to the CD4 molecule (Maddon
et al., 1986), rhinovirus binds to lCAM-1, and polivirus uses
another immunoglobulin as a receptor (Mendelsohn et dl., 1989;
Greve et al., 1989, Staunton et al., 1989).
Virus receptors have also been identified in other
species. Mouse hepatitis virus (MHV) , a coronavirus, is often
used to study viral infections in vivo (Wege et al., 1982)
since coronaviruses are also common pathogens among humans
causing respiratory il] ness. One MHV-resistant strain of
mouse (SJL/J) does not possess the virus receptor identified
as a 110-120 kDa glycoprotein on the plasma membranes of
hepatoeytes and intestinal epithelial cells of MHV-susceptible
BALB/e mice (Williams et al., 1990). Injecting resistant mice
with a tissue extract from susceptible mice induced the

95
recipients to synthesize antibodies against the MHV receptor.
These antibodies were used to purify the receptor protein
(Williams et al., 1991), and allow the cDNA encoding it to be
cloned. By sequence homoJogy, the MHV receptor was identified
as the mmcm-Ha protein presently described (Dveksler et al.,
1991). To determine the domain(s) important for bindi~g of
MHV to its receptor, many of the splice variants described
here have been sent to Dr. Kay Holmes and the ability of these
molecules to confer susceptibility to viral infection on
transfectant hurnan and hamster cells will be examined. To
date, no specifie domain has been found to be essential for
infection.
Resistant SJL/J mice, by Western analysis, do not produce
intact mmCGM1a protein. They do, however, produce a protein
of different molecular weight which crossreacts with an anti-
human CEA antibody (Williams et al., 1990). Phenotypically,
this mouse strain appears to be normal up to adulthood; at
this point, they then develop tumors. Perhaps these rnice give
sorne indication of the importance of mouse CEA-like proteins
in the maintenance of tissue architecture. The related
prat ein expressed in SJL/ J mice is enough to allow development
to proceed normally, however, it is unable to rnaintain the
tissue architecture throughout adulthood.
By genomic analysis and by RNAse protection assays,
further studies will be done to identify the mutation in the
mmCGMl locus. By determining which CEA crossreactive protein

1 96
is expressed in SJL/J rnice, perhaps we can assign different
functions to the different proteins generated by alternative
splicing; sorne rnay play an important role in development 1
whiJe others have a tumor suppressor role.
Conclusion:
Several CEA-related proteins have been identified in the
mouse. The transcripts encoding these different proteins are
generated by alternative splicing of one precursor messenger
RNA. The tissue distribution and temporal regulation of these
different farnily members will have to be established in order
to understand the significance of this extensive alternative
splicing.
There are many possible functions for these proteins
which will have to be investigated, as described above. One
function which has already been established is that of an
adhesion molecule. The expression pat tern during mouse
development as well as the change in expression upon tumor
formation supports the hypothesis that mouse CEA-related
proteins mediate cell adhesion in vivo. However, the fact
that at least eight different related proteins exist in the
mouse, allows speculation that there is more than one function
for these proteins. Further functional studies on the mouse
CEA-related proteins, as weIl as regulatory studies on the
gene encoding them are in progress.

97
References
Afar, D.E.H. , Stanners, C.P. and Bell, J. Tyrosine
phosphorylation of biliary glycoprotein, a cell adhesion
molecule related to carcinoembryonic antigen. Biochemica et
Biophysica Acta (in press) .
Barnett, T., Tretschner, A., Austen, D., Goebel, S., Hart, J.,
Elting, J. and Kamarck, M. Carcinoembryonic antigens
alternative splicing accounts for the multiple rnRNAs that code
for novel members of the carcinoembryonic antigen family. J.
Celle Biol. 108: 267-276, 1989.
Beauchemin, N., Turbide, C., Afar, D., Bell, J., Raymond, M.,
Stanners, C.P. and Fuks, A. A mouse analogue of the human
carcinoembryonic antigen. Cancer R~s. 49: 2017-2021, 1989.
Cohen, P., and Klee, L.B. Calmodulin, vol. 5: molecular
aspects of cellular regulation. Amsterdam: Elsevier, 1988.
Dusenbery, K., Mendiola, J., and Skubitz, K. Evidence for
ecto-protein kinase activity on the surface of human
neutrophils. Biochem. Biophys. Res. Commun. 153: 7, 1988.
Dveksler, G., Pensiero, M., Cardellichio, C., Williams, R.,
,( Jiang, G-S., Holmes, K. and Dieffenbach, C. Cloning of the

1 mouse hepatitis
and hamster cell
98
virus (MHV) receptor: expression ln humc1n
lines confers susceptibility to MH'I. ,1.
Virol. 65: 6881-6891, 1991.
Fanciulli, M., Paggi, M., Mancini, A., Del Carb, C., Floridi,
A., Taylor, S. and Perotti, N. pp120: a COIlUTIon endogenous
substrate for insulin and IGF-1 receptor-associated tyrosine
kinase activity in the highi.y malignant AS-30D rat hepatoma
cells. Biochem. Biophys. Res. Commun. 160: 168-173, 1989.
Foulkes, N.S., Mellstrom, B., Benvsiglio, E. and Sassone
Coursi, Developmental switch of CREM function during
sperrnatogenesis: from antagonist to activator. Nature 355: 80-
84, 1992.
Frixon, U., Behrens, J., Sacha, M., Eterle, G., Voss, B.,
Warda, A., Lochner, D., and Birchmeier, W. E-cadherin mediated
cell-cell adhesion prevents invasiveness of human carcinomd
cells. J. Cell Biol. 113: 173-185, 1991.
Greve, J.M., Davis, G., Meyer, A.M., Fort~, C.P., Vast, S.C.,
Marlor, C.W., Kamarck, M.E. and McClelland, A. The major human
rhinovirus receptor is ICAM-1. Cell, 56: 839-847, 1989.
Guthrie, C. Messenger RNA splicing in yeast: clues to why thé
spliceosome is a ribonucleoprotein. SciencE' 253: 157 -] 63,

" "
: 99
1991.
Hlxon, D. and McEntire, K. Detection of an altered form of
cell-CAM 105 on rat trdnsplantable and primary hepatocellular
carcinomas. Cancer Res. 49: 6788-6794, 1989.
Huang, .LQ., Turbide, C., Daniels, E., Jothy, S., Beauchemin,
N. Spatiotemporal expression of murine carcinoembryonic
antigen (CEA) gene family rnembers during mouse embryogenesis.
Development 110: 573-588, 1990.
Hubbard, A.L. , Bartles, J.R. and Braiterman, L.T.
Identification of rat hepatocyte plasma membrane proteins
I.lsing monoclonal antibodies. J. Cell Biol. 100: 1115-1125,
1985 .
• Jessup, J.M., Giavazzi, R. , Campbell, O., Cleary, R. ,
Morikawa, K. and Fidler, I.J. Growth potential of human
colorectal carcinomas in nude mice: association with the
preoperative serum concentration of carcinoembryonic antigen
in patients. Cancer Res. 48: 1689-1692, 1988.
Keoing, R.J., Lazar, M.A., Hodin, R.A., Brent, G.A., Larsen,
P.R., Chin, W.W. and Moore, 0.0. Inhibition of thyroid hormone
action by a non-hormone binding c-erbA protein generated by
alternative mRNA splicing. Nature 337: 659-661, 1989.

1 100
Lin, S-H., Guidotti, G. Cloning and expression of a cDNA
coding for a rat liver plasma membrane ecto-A'l'Pase: the
prirnary structure of the ecto-ATPase is simildl: to that of the
human biliary glycoprotein. J. Biol. Chern. 264: 14408-14414,
1989.
Lin, S-H., Culic, O., Flanagan, D., Hixon, D. Irnmunochernical
characterization of two isoforrns of rat liver ecto-ATPase that
show irnmunological and structural identi ty wi th a glycoprotein
cell-adhesion molecule with Mr 105 000 Biochem. J. 278: 155-
161, 1991.
Maddon, P.J., Dalgleish, A.G., McDougal, J.S., Clapharn, P.R.,
Weiss, R.A. and Axel, R. The T4 gene en,~odes the AlDS virus
receptor and is expressed in the immune system and the brain.
Cell 47: 333-348, 1986.
Maniatis T. and Reed, R. The role of smal1 nuclear
ribonucleoprotein particles in pre-mRNA splicing. Nature 325:
673-678, 1987.
Margolis, R. ,
Identification
hepatocellular
Taylor 1 S., Serninara 1 D. and Hubbard, A.
of pp120, an endogenous substrate for the
insulin receptor tyrosine kinase, dS an
integral membrane glycoprotein of the bile canalicu lar doma j n.
Proc. Natl. Acad. Sei. U.S.A. 85: 7256-7259, 1988.

r
101
Margolis, R., Schell, M., Taylor, S. and Hubbard, A.
Hepatocyte plasma membrane ecto-ATPase (pp12 0 /HA4) is a
substrate for tyrosine kinase activity of the insulin
receptor. Biochem. Biophys. Res. Commun. 166: 562-566, 1990.
Mendelsohn, C.L., Wirruner, E. and Racaniello, V.R. Cellular
receptor for poliovirus molecular cloning, nucleotide
sequence, and expression of a new member of the immunoglobulin
superfamily. Cell, 56: 855-865, 1989.
McCuaig K., Turbide, C., and Beauchemin, N. rnrnCGMla: a mouse
carcinoembryonic antigen gene family member generated by
alternative splicing, functions as an adhesion molecule. Cell
Growth Diff. 3: 165-174, 1992.
Mullen, M. P., Smith, C. w. S., Patton, J.G. and Nadel-Ginard, B.
a-Tropomyosin mutually exclusive exon selection: competition
between branchpolnt/polypyrimidine tracts determines default
exon choice.
Genes Dev. 5: 642-655, 1991.
Neuner, A., Jannasch, H.W., Belkin, S. and Stetter, K.O. A
new species of extremely thermophillic marine archaebacteria.
Arch. Microbiol. 153: 205-207, 1990.
Obrink, B. Epithelial cell adhesion molecules. Exp. Cell Res.

1 102
1163: 1-21, 1986.
Obrink, B., Blikstad, M., Hansson, M., Odin, P., Svalander, P.
and Tingstrom, A. Cell-CAM 105 - an adhesive cell surface
glycoprotein. Protoplasma 145: 182-187, 1988.
Ocklind, C. and Obrink, B. Intercellular adhesion of rat
hepatocytes. Identification of a cell surface glycoprotein
involved in the initial adhesion proces~. J. Biol. Chem. 257:
6788-6795, 1982.
Odin, P. and Obrink, B. Dynamic expression of the cell
adhesion molecule cell-CAM 105 in fetai and regenerating rat
liver. Exp. Cell Res. 164: 103-114, 1986.
Odin, P. and Obrink, B. Quantitative deterrnination of the
organ distribution of the cell adhesion molecule cell-CAM 105
by radioimmunoassay. Exp. Cell Res. 171: 1-15, 1987.
Odin, P., Asplund, M., Busch,
Irnrnunocytochernical localization of
C. and
cell-CAM
Obrink, B.
105 ln rat
tissues. Appearance in epithelia, platelets and granulocytes.
J. Histochem. Cytochern. 36: 729-739, 1988.
Perotti, N., Accili, D., Marcus-Sarnuels, B., Rees-.Jones, R.,
and Taylor, S. Insulin stimulates phosphorylation of a 120 kDa

(
103
glycoprotein substrate (pp120) for the receptor-associated
protein kinase in intact H-35 hepatoma eells Proc. Natl. Aead.
Sei. U.S.A. 84: 3137-3140, 1987.
Phillips, S., Perotti, N. and Taylor, S. Rat liver membranes
eontain a 120 kDé. glyeoprotein which serves as a substrate for
the tyrosine kinases of the receptors [or insulin and
epiderrnal growth factor. FEBS 212: 141-144, 1987.
Rees-Jones, R. and Taylor, S. pp120 in an endogenous substrate
for the insulin receptor-associated tyrosine kinase J. Biol.
Chern. 260: 4461-4467, 1985.
Robbins, J., Robbins, P., Kozak, C., and Callahan, R. The
mouse biliary glycoprotein gene (Bgp): partial nueleotide
sequence, expression, and ehromosomal assignment. Genoroics 10:
583-587, 1991.
Rojas, M., Fuks, A. and Stanners, C.P. Biliary glycoprotein,
d member of the immunoglobulin supergene faroily functions in
vitro as a Ca2t -dependent intercellular adhesion molecule. Cell
Growth niff. 11: 527-533, 1990.
Roman, C., Cohn, L. and Calame, K. A dominant forro of
transcription activator mTFE3 created by differential
splicing. Science 254: 94-97, 1991.

1
. ,
104
Sharp, P.A. Splicing of messenger RNA precursors. Science,
235, 766-771, 1987.
Shuster, J.D., Thompson, M.P., Fuks, A. and Gold, P.
Immunologie approaches to diagnosis of malignancies. Prog.
Exp. Tumor Res. 25: 89-139, 1980.
Skubitz, K.M., Ehresmann, D.D. and Ducker, T.P.
Characterization of human neutrophil ecto-protein kinase
activity released by kinase substrates. J. Immunol. 147: 638,
1991.
Skubitz, K., Ducker, T. and Goueli, S. CD66 monoclonal
antibodies recognize a phosphotyrosine-containing protein
bearing a carcinoembryonic antigen cross-reacting dntigen on
the surface of human neutrophils. J. Immunol. 148: 852-860,
1992.
Smith, C.W.J., Porro, E., Patton, J.G. and Nadel-Ginard, B.
Scanning from an independently specified branch point defines
the 3' splice site of mammalian introns. Nature 342: 243-247,
1989.
Staunton, D.E., Merluzzi, V.J., Rothlein, R., Barton, R.,
Martin, S.D. and Springer, T.A. Cell adhesion molecule ICAM-l,
is the major surface receptor for rhinoviruses. Cell 56: 849-

105
853, 1989.
Svalander, P., Odin, P., Nilsson, B.a., and Obrink, B.
Trophectoderm surface expression of the cell adhesion
molecule cell-CAM 105 on rat blastocysts. Development 100:
653-660, 1987.
Turbide, C., Rojas, M., Stanners, C.P. and Beauchemin, N. A
'llouse carcinoembryonic antigen gene family member is a
calcium-dependent cell adhesion molecule. J. Biol. Chem. 266:
309-315, 1991.
Vleminckx, K., Vakaet, L., Mareel, M., Fiers, W. and Van Roy,
F. Genetic manipulation of E-cadherin expression by epi thelial
tumor cells reveals an invasion suppressor role. Cell 66: 107-
119, 1991.
Wege, H., Siddel, S. and terMeulen, V. The biology and
pathogenesis of coronaviruses Curr. Top. Microbiol. Immunol.
99, 165-200, 1982.
Werner, S., Duan, D-S. 1 deVries, C., Pet ers , K., Johnson, D.,
and Williams, L. DifferentiaI splicing in the extracellular
region of fibroblast growth factor receptor 1 generates
receptor variants with different ligand-binding
specificities. Mol. Celle Biol. 12: 82-88, 1992.

1
"
106
Williams, R., Jiang, G-S., and Holmes, K. Receptor for mouse
hepatitis virus is a member of the carcinoembryonic antigen
family of glycoproteins. Proc. Natl. Acad. Sei. U.S.A.
88:5533-5536, 1991.
Williams, R., Jiang, G-S., Snyder, S., Frana, M. and Holmes,
K. Purification of the 110-kilodalton glycoprotein receptor
for mouse hepatitis virus (MHV) -A59 from mouse liver and
identification of a non-functional, homologous protein in MHV
resistant SJL/J mice. J. Virol. 64: 3817-3823, 1990.
Wilson, T. and Treisman, R. Removal of poly (A) and consequent
degradation of c-fos rnRNA facilitated by 3' AU-rich sequences.
Nature 336: 396-299, 1988.
Recommended

















