
A STUDY OF UREASE ACTIVITY
IN THE RUMEN lUCROFLORA OF SHEEP
by
Graham Alfred Jones
A THESIS
Submitted to the Faculty of Graduate Studies and Research, McGill University, in partial fulfilment of the requirements
for the degree of Doctor of Philosophy
Department of Agricultural Bacteriology, Macdonald College of McGUl University, Que bec. April, 196.3

TABLE OF CONTENTS
Page
ACKNON!,EDGmŒN'IS •••••••••••••••••••••••••••••••••••••••••••••••••••• v
CLAIM OF CONTRIBUTION TO KNO\v.LEDGE •••.••••••••••••••••••••••••••••• vi
GENERAL INTRODUCTION •••••••••••••••••••••••••••••••••.•••••••••••••• 1
GEtJERAL LITERA ID RE. REVI.m'l. • • • • • • • • • • • • • • • • • • • • • ••••••••••••••••••••• 3
A. Function of the rlllll6n ••••••••••••••••••••••••••••••••••••••• 3 B. Nitrogen metabolism in the rumen •••••••••••••••••••••••••••• 4 C. The utUization of NPN feed supplements by ruminants •••••••• 9
PART I. THE ATT1!11PTED ISOLATION OF ANAEROBIC UREASB-PRODUCING BACTERIA FROM SHmP RUMEN CONTENTS
INTRODU CTl: ON • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 13
LITERATURE REVIEW •••••• ·······•·•·····•·•···•·•····••··••·•··••••· 14
A. HYdrolysis of urea in the rumen ••••••••••••••••••••••••••• 14 B. Enumeration of rumen bacteria ••••••••••••••••••••.•••••••• 16 C. Cultural detection of urease production by bacteria ••••••• 17 D. Urease-producing rumen bacteria ••••••••••••••••••••••••••• 20
l!AT:ER.IAIS AN'D .WTHOœ. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 24
A. Ex:p:er.imental aniJna.l.s. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 24 a) Sileep 2.. • • • • • • • • • • . • • • • • • • • • • • • • • • • • • • • • • • . • • • • • • . • • 24 b) Sheep 35 alld 37. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 24 c} Steers 1 and 2 ••••.•••••••••••.••••••••••••••••••••• 24
B. Collection of rumen contents •••••••••••••••••••••••••••••• 25 a) Slleep 2 ••••••••••••••••••••••••••••••••••••••••••••• 25 b) Sheep 35 and 37 •••••••••••••••·••••••••••••••••••••• 25 c) Steers 1 and 2 ••••••••••••••••·•·••••••••••••••••••• 25
c. Preparation or rumen fluid ••••••••••••••••••••••••••••••• 25 D. Fractionation of rumen :tluid ••••••••••••••••••••••••••••• 26
a) Yicrobial and supernatant fractions ••••••••••••••••• 26 b) Subfractionation of the microbial fraction •••••••••• 26 c) Separation of rumen protozoal fraction •••••••••••••• 28
E. Measurement of urease activity ••••••••••••••••••••••••••• 29 F. Bacterial viable counts •.•••••••••••••••••••••••••••••••• 30
a) Total viable counts ••••••••••••••••••••••••••••••••• 31 b) Counts of viable ureolytic organisms •••••••••••••••• 32

- ii -
G. Urease-producing bacteria from rumen fluid ••••••••••••••• 33 a) Isolation ••••••••••.•••••••••••••.••••••••••••••••••• 33
RESUL1S
A. B. c.
i) Isolation from buffered reinforced clostridial broth ..•••.•..•.•••.•.•.•.........•...•........
ii) Isolations from supplemented rumen supernatant 33
liquor medium •••••••••••••••••••••••••••••••••• 34 iii) Isolations of rumen staphylococci by enrichmant
in a brain heart infusion - NaCl medium •••••••• 37 b) Identification of isolated organisms •••••••••••••••• 38 c) Urease activity of ureolytic isolates ••••••••••••••• 38
.......................................................... Urease activity of rumen fluid and its component fractions Proportion of ureolytic bacteria in rumen fluid •••••••••• Attampts at the isolation and characterization of ureaaeproducing bacteria from rumen fluid ••••••••••••••••••••••
39
39 44
a) Isolation of bacteria from cultures in buffered reinforced clostridial medium ••••••••••••••••••••••• 46
i) Isola te 1 • • • • • • • . . • . . . • . . . • . • . • . . . • . . . • . . . • . • • . 47 ii) Isolate 2 •.•••................................. 49
iii) Urease activity of isolate 2 ••••••••••••••••••• 51 b) Isolation of bacteria from cultures in a supple-
mented rumen supernatant liquor medium •••••••••••••• 52
DISCUSSION ....................................................... 57
S~Y . . . • • • . . • . . . . . • . . . . . • . . . . . . . . . . . • . . • . . • . . . . • . • . . . • . . . . . . . • 61
PART II. FACTORS AFFECTING THE ACTIVITI OF RUMEN UREASE
INTRODUCTION ••••••••••••••••••••••••••••••••••••••••••••••••••••• 65
LITERAT1JRE REVImV' • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 67
A. Factors affecting the activity of jack-bean urease ••••••• 67 a) Urease stimulating agents ••••••••••••••••••••••••••• 67 b) Urease inhibiting agents •••••••••••••••••••••••••••• 69
B. Properties of urease produced by non-rumen bacteria •••••• 70 C. Properties of rumen urease ••••••••••••••••••••••••••••••• 71
MATERIAIS AND METHODS ............................................ 72
A. B.
Collection and preparation of rumen fluid •••••••••••••••• 72 Preparation of ~d rumen ureaae •••••••••••••••••••••••• 72
a) Whole oeil preparations of rumen microorganisme ••••• 72 b) Acetone-dried powders of rumen microorganisme ••••••• 72 c) Cell-free extracts of rumen microorganisme •••••••••• 73

c. D. E.
RESULTS
iii
Preparation of jaek-bean urease •.•....•......••••...••... Preparation of ashed rumen supernatant liquor •••••••••••• Measurement of urease aetivity •••••••••••••••••••••••••••
a) Routin.e method •.••••••.•...•.•...•.•••••.•...•••.••. b) Urease activity in the presence of added cations ••••
..........................................................
74 74 74 74 74
76
A. The identity of factors affeeting the aetivity of r\.lllen UNase • . • . . . . . . . . . . • • • . . • . . • • • . . . . . . . . . . . • . • . • • • . . . • • . . . • 76
a) Experimenta with intact rumen microorganisme •••••••• 76 b) Experimenta wi th an acetone-dried powder of rumen
microorganisms •••••••.••••••.•••••••.•••••••••.•.••• 88 e) Experiment with a cell-free extract of rumen micro-
orgards.ms • • • • • • . . . • • • • . . . . . . • . • . . . • . . . • . • . . • . • . . • . . . 90 B. Effect of di valent ions on the activity of jack-bean
urease • . . . . . . • • • . . . . • • . . . • . . . • . . . • . . . • . . . . . • . • . . . . . . . • . . . 92 C. Loealization of u:œase aetivi. ty in. rumen microbial cells.. 92
DISCUSSION ....................................................... 103
sm~Y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 108
BIBLIOGRA.PIII' • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 112
APPENDIX. REPORT OF A PRELlMINARY EXPERIME:NT ON THE FATE OF N15-LABELLED UREA IN THE: RUMEN OF A SHEEP
INTRODUCTION ••••••••••••••••••••••·••••••••••••••••••••••••••••• 129
LITERATl.J'IŒ REVIE:Vf • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 132
MATERIA.I.S AND l!ETHODS • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 136
A. Exper:iJnental a.nimaJ. ••••••••••••••••••••• , •• ~ .••••• , • • • • • 136 B. Nl5-urea •......•••.........•..•....•...•.......•........ 136 c. Collection and preparation of samples •••••••••.•••.•.•.• 137
a) Rumen contents ••••••.••••.••.•.•••••••••••••••••••• 137 b) Saliva ....•............••.........................• 137 c) Urine . . . • • . . . . . . . . . . . . . . • . . . . . . . . • • . . . • . . . . . . . • • . • • 138
D. Estimation of nitrogen in samples ••••••••••••••••••••••• 138 a) Ammonia in rumen supernatant liquor •••••••••••••••• 138 b) Estimation of total nitrogen ••••••••••••••••••••••• 139
i) Rumen supernatant liquor •••••••••••••••••••••• 140 ii) Rumen microbial fraction •••••••••••••••••••••• 140
iii) Sali va . • • . . . . . . . . . . . . . . . . • . . • • . . . • . • . • . . • • . • • • 14.0 iv) Urine . . • . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14.0
E. Nl5 Analyses •••••..•••••••.•.•.•••••••.•.•.•.•••.••••.•• 141 a) Oxidation of ammonia-nitrogen to N2 ••••••••••••.•.• 141 b) Determination of N14JN15 ratio •·••••••••••••••••••• 141

iv
RESULTS ......................................................... 143
A. Rumen fluid • . • • • • . . . • . • • . . . . . . • • • . • • • • . . . . . . . . . . • . • • • . • • 143 a) pH of rumen fluid •••••••••••••••••••••••••••••••••• 143 b) Ammonia content of rumen supernatant 1iquor • • • • • • • • 143 c) Total nitrogen content of rumen supernatant 1iquor
and its enrichment with N15 •••••••••••••••••••••••• 146 d) Total nitrogen content of the microbial fraction
of rumen fiuid and its enricbment with N15 • • • • • • • • • 149 B. Saliva: N15 enrichment •••••••••••••••••••••••••••••••••• 151 C. Urine: excretion of N15 in urine •·•••••••••••••••••••••• 153
DISCUSSION ·······················•·••·····•·······•••·•·•·•····• 156
S~RY • • • • • . . . . • . . • • . • • • • • . . . . . . • • . • . • . • . • • . • • • • . • • • . • • . • . • . • . • 159
BIBLIOORA.Pli'I' • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 161

The author is indebted to the following members of staff
of Macdonald College, and is pleased to express his gratitude to
them for the assistance they have afforded him during the course
of these investigations.
Dr. A. c. Blackwood, Professor, and Dr. R. A. MacLeod,
Associate Professor, Department of Agricultural Bacteriology,
for general guidance and for frequent and generous discussion
of the problems encountered.
Dr. L. E. Lloyd, Professor, Department of Animal Science,
for providing facilities for the maintenance of the sheep
used in the investigation, and for advice on the interpretation
of the resulte presented in the Appendix to this thesis.
Dr. R. Knowles, Assistant Professor, Department of Agri
cultural Bacteriology, for adviee and assistance with the
Nl5 analyses.
'lhe work was carried out under a grant from the Macdonald
Agricultural Research Fund.

CLAIM OF CONTRIBUTION TO KNCHLEDGE
Studies were made of some aspects of the urease activity of
strained rumen fluid from a sheep fed a diet in which urea satisfied
a proportion of the nitrogen requirement of the animal, and the assoc
iation ot rumen urease activity lfith the rumen bacteria was contirm.ed.
It is claimed that the following turther findings of these etudies
constitute a contribution to knowledge.
1) The baeteria responsible for 65% of rumen urease activity
were mainly the group of larger organisms, eamprised of several morpho
logical types, whieh sedimented at a relative centrifuga! foree of
1,200 x G.
2) Approximately 35% ot the total viable population of rumen
bacteria eonsisted of organisme which bad the capacity to produce
urease when cultured in a buffered reintoreed clostridial :medium.
3) A medium consisting of SO% rumen supernatant liquor,
supplemented wi th glucose, phytor~e and urea and incuba ted anaerobically,
provided enriehment cultures of rumen bacteria in which the urease
aetivity per unit concentration of cella after two successive transfera
was increased four-fold over that prevailing in rumen fluid.
4) The activity of preformed rumen urease was enhanced in
the presence of rumen supernatant liquor. This eftect was due to
the presence of an inorganic urease stimulating factor in rumen
supernatant liquor.

-vii-
5) Rumen urease activity was specifically enhanced in
the presence of the divalent cations :Mn++, Mg++, Ca++, sr++ and Ba++.
The activity was reduced in the presence of Na+, K+ and Co++, and the
enzyme was completely inhibited by Zn++, Cu++ and Fe+++.
6) Jack-bean urease was inhibited in the presence of con
centrations of divalent cations which brought about optimum stimulation
of rumen urease.
7) When intact cella of rumen microorganisme were washed
in 0.02M-maleate buffer there was a progressive reduction in the urease
activity of the cella as the number of washings increased. Up to tœ
third washing rumen supernatant liquor and solutions of Mn++, Mg++ and
Ca++ could restore the activity of the washed cella to the leval pre
vailing in unwashed cella. After the sixth and nin th washings the
activity of the cella could not be fully restored in this way.
8) Inorganic ions penetrated resting cella of rumen
ureolytic microorganisms at rates which varied with the ion involved.
The sequence of the penetration rates of rumen urease stimulating
ions into who le cells was , in decreasing order, :Mn++.> Mg++ ,:::::::..ca++;;::::...
Treatment of the cella with acetone modified the rates
at which these ions penetrated the cella. The sequence of the
penetration rates of the ions into acetone-treated cella was, in
decreasing order, Mg++= Ca++;::.:::--)ln++7Sr++:;::::..Ba++.
9) The urease activity of a cell-free extract of rumen
microorganisms was immediately enhanced when solutions of Mn++, .Mg++,

- viii -
and ca++ were added, and immediately reduced when solutions of co++
and K+ were added. When solutions of Sr++, Ba++ and Na+ were added,
however, there was a delay before the affects of these ions on the
urease activity of such an extract became apparent.
10) In addition to variations in cell penetration rate, the
different urease stimulating and inhibiting cations bad varying
absolute affects upon the intracellular urease enzyme of whole cells
of rumen microorganisme after they had penetrated the cella. The
sequences of stimulation and inhibition brought about in this way
were the sruœ as the sequences o:t penetration rates of the respective
groups of ions.

ŒNERAL INTRODUCTION
The ruminant animal, by virtue of the population of active
microorganisms whi.ch is supported in the rumen, can utilize a much
wider range of nitrogen containing substances for body maintenance
and production than can the monogastric animal. The rumen micro
organisme, chie fly the bacteria, synthesize microbial prote in of
high biological value from non-protem nitrogen sources in the rumen,
and this becomes available to the host animal by nonnal processes of
protein digestion in the abomasum. The animal is therefore not
entirely dependent upon dietary protein to satisfy its nitrogen re
quirements; indeed expansive dietary protein may be conserved by
feeding cheaper non-protein supplements to ruminants.
A non-protein nitrogen compound which has been widely used
as a prote in replacement is ure a, and a great deal of 110rk has been
devoted to defining the conditions under which this compound may be
most efficiently utilized by the rumen microfiora for protein synthesis.
This has primarily involved f'eeding trials and in vitro experimentation
wi th the mixed rumen micropopulation.
Dietary urea nitrogen becomes incorporated into body protein
by a process which initially involvea the conversion of urea to ammonia
by rumen urease; subseqœntl.y the ammonia is incorparated into micro
bial protein and this is then digested by the animal. The aymbiotic
rumen bacteria elaborate a highly active urease enzyme which catalyses

- 2 -
tœ hydrolysis of urea, but the mechanism by which this h;ydrolysis is
brought about bas received little study. The reason for this is that
it has not yet proved possible to isolate in pure culture organisme
canprising tm predominant ureolytic flora of the rumen.
In an attem.pt partly to remed;y this situation, investigations
were made of some aspects of the urease activity of rumen contents
from a sheep receiving a dietary urea supplement. Following experi
menta directed towards confirmation of previous work concerning the
source and location of rumen urease, soma attempts were made to isolate
representative ureolytic bacteria from rumen contents. These attempts
are described in Part I. No suitable organisns 11ere isolated, however,
and the mi.Jœd rumen microflora was therefore used as a source of enzy
me in a study of factors affecting rumen urease activity. This aspect
of the investigation is described in Part II.

GENUAL LITERATURE REVIEW
A. Function of the Rumen
The d.igesti va tract in herbivorous a.nimals is equipped
with a capacious organ in which fermentation of bulky, fibrous food
ca.n take place; in ruminants this is the complex stomach, espe cially
that part known as the rumen. The rumen has an important me chanica.l
action upon food material contained in it, and also provides rela.tivaly
constant conditions for the growth and physiological a.ctivity of a
large number of microorganisme, chie fly protozoa. and ba.cteria. It is
essentia.lly a. highly reducing, anaerobie, continuous fermentation
system under a gas phase of carbon dioxide, methane and hydrogen, a.t
a tempe rature of 39°0 and buffe red at a slightly a.cid pH. The dyna.mic
nature of the system is mainta.ined by the continuous inf'low of saliva,
the periodic intake of food and water, the passage of partia.lly f'erm
ented material to the om.asum and the absorption of' s ane soluble ferm
entation products by the rumen wall.
In t.he adul.t sheep the rumen may have a capacit.y of 20 - 25 1. ,
and the .fluid volume may excee d 5 1. Counts of rumen protozoa and
bacteria have averaged respectively 106 and lolO organisme per ml. of
fluid. The rumen is there.fore a site of considerable biochemica.l
activity. An important function of the rumen population is the ferm
entation of complex dieta.ry ca.rbohydrates, principally cellulose, with
the production of volatile fatty acids which are the major source of'

-4-
energy for the animal. Dietary protein degradation oceurs simultan
eousl.y with the incorporation of non-protein nitrogen into microbial
protein. Rumen microorganisms can also synthesit<e all components of
the vitamin B co.mple.x and vitamin K in sufficient quantities to meat
the entire demanda or the adult animal.
Rumen metabolism in its m.a.ny aspects has been comprehens
ively reviewd by Annison and lewis (1959) and Barnett and Reid (1961).
B. Nitrogen lletabolism in t:œ R.ulœn
The nitrogen requirements of non-ruminants are met by
absorption of the peptides and amino acids which result from the
degradation of digested protein in the stomach and small intestine,
a.rrl a specifie protein has a biological value determined largel.y by
the nature and availability of its amino acids. In the case of rumin
ants, however, the presence of a population of microorganisme in the
rumen markedly changes the relationship between tœ :tom of tœ nitro
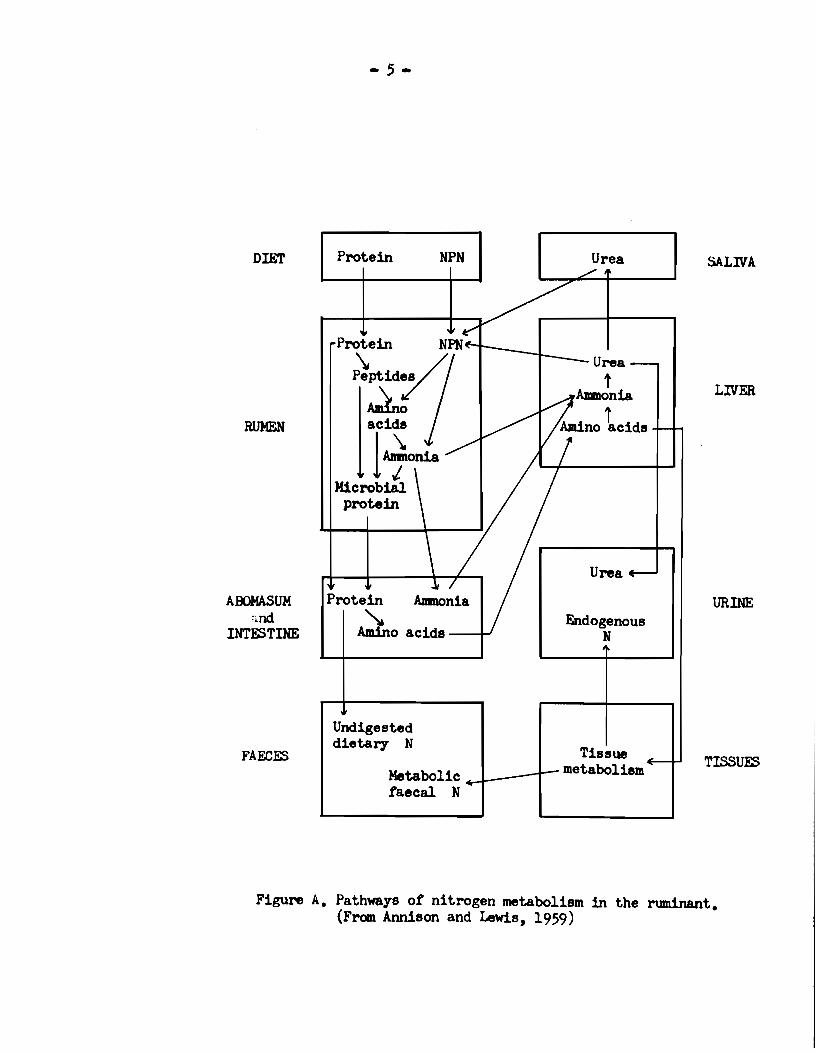
gan ingested orally and its availability to the animal. A schematic
representation of the present:cy known pathways of nitrogen metabollsm
in the ruminant is sho1m in Figure A.
On a natural forage diet, 80 - 90% of the total nitro gan
ingested by a ruminant may be in the form of protein, and the rema.inder
consista of such non-protein nitrogen (NPN) compounds as free amino
acids , peptides, nucleic a.cids, purine and pyrimidine bases, a.lkaloids,
urea, nitrates and ammonia (Chibnall, 1939; Lugg, 1949). lücrobial
D.IE'l'
RUMEN
A.oo.MASUM ;.nd
INTESTINE
FAECE.S
-5-
Prote in NPN
ac ids
1~ Microbial prote in
Protein Ammonia '\.
Am.ino acida --+J
Undigested dietary N
Metabolic . L
faecal N
Urea
Urea
Endogenous N
Tissue metabolism
SALIVA
LIVER
URINE
TISSUES
Figure A. Pathways of nitrogen metabolism in the ruminant. (From Annison and Lewis, 1959)

- 6 -
proteases in the rumen degrade a proportion of the ingested protein
(Pearson and Smith, 1943b) and tte amino acids form.ed are tten att
acked by deaminases and transaminases (Otagaki, Black, Goss and
Kleiber, 1955) of microbial origin. The activity of these enzymes
is such that ammonia is a major end-product of the degradation and
is generally the main component of the rumen NPN fraction (McDonald,
1948), but deamination of amino acids also resulta in the production
of energy-rich volatile fatty acids (Sirotnak, Doetsch, Brown and
Shaw, 1953) which supplement those form.ed during cellulose ferment
ation. The actual proportion of ingested protein which is degraded
in this way depends upon the nature of the protein and tha nature and
amount of carbohydrate in the diet (I.awis and McDonald, 1958).
The rumen microflore. can also attack a wide range of NPN
compounds entering the rumen. Individual amino acide (Sirotnak ,!:!! ~·,
1953), nucleic acids (Ellis and Pfander, 1958), purines and pyrimid
ines (Jurtshuk, Doetsch and Shaw, 1958), and urea (Pearson and Snith,
1943a) can ail be broken down to ammonia, carbon dioxide and volatile
fatty acids in various proportions, and nitrate may be reduced to
amm.onia (Iawis, 1951).
Diss:imilation of nitrogenous compounds in the rumen there
fore resulta largely in the production of ammonia, the concentration
of which may vary with time and diet from 5 - 120 mg. per 100 ml.
rumen liquor (Annison, Cha.lmers, Marshall and Synge, 1954). The
variations with time depend upon the rate of production of ammonia

- 7 -
relative to tha.t of its removal from the rumen solution by several
pathways.
The first of these pathwa.ys has been clearly demonstrated
by McDonald (1948) and Bouckaert and Oyaert (1952). A proportion of
the ammonia passes directly into the ruminal veina, which form part
of the hepatic portal system. This is probably achieved by simple
diffusion since the level of ammonia in portal blood is proportional
to that in tb! rwnen solution (Lewis, Hill and Annison, 1957). The
second pathway by which ammonia is removed from tb! rwnen involves
the passage of rumen ammonia to the abomasum. and sma.ll intestine
along with the normal flow of ingesta. Fran there it is absorbed
into the hepatic portal system. If the amount of anmonia absorbed
by the portal blood is not excessive all of it is converted to urea
in the liver. otherwise sone ammonia passes through the liver and
appears in the peripheral blood (Lewis !! al., 1957). Clinical
symptoms of tox:icity may be ma.nifested in the animal if the concent
ration of ammonia in the peripheral circulation increases to a crit
icaJ. vaJ.ue (Lewis, 1960), but. this is unlikely under nozmal conditions.
Urea formed in the li ver may be excreted in the urine, thus resulting
in a losa of ni trogen to the animal, re cycled to the rumen via the
saliva (MeDonald, 1948), or, of greater quantitative signiticance,
it may be secreted from the bloodstream through the rumen wall into
the rumen (Houpt, 1959).
The third pathway by which a.nmonia is removed from the

- 8 -
rumen is by incorporation into rumen microbial protein. There is a
considerable body of evidence available to show that this pathway is
of great significance to the nitrogen nutrition of the animal.
Zuntz (1891) was the first to suggest that conversion of
NPN to microbial protein might occur in the rumen and evidence for
this process was later provided by indirect methods auch as measure
ment of changes in the relative concentrations o:f rumen nitrogen
:fractions a:fter fee ding (Gray, Pilgrim and Waller, 1953), and demon
stration o:f amino acid synthesis in l"''l..œn contents (Loosli, Williams,
Thomas, Ferris and Maynard, 1949). More recently, Wamer (1961)
showed that during incubation o:f an in vitro system comprising rumen
liquor, a protein substrate and N15-ammonia, labelled nitrogen was
incorporated into bacterial protein.
Evidence :for the quantitative signi:ficance of protein
synthesis in the rumen lias obtained by Pearson and Smith (1943b).
These worlœrs incubated bovine rumen liquor and 0.3% starch in vitro
:for two hours in the presence o:f various NPN substrates and d.emonstr
ated protein synthesis to the extent of 8 mg. nitrogen per lOO g.
rumen liquor. I:f this rate .are ma.intained :for 24 hr. in !!!2, 72 g.
nitrogen would be incorporated. This would represent synthesis of
450 g. protein, which is su:f:ficient to satis:fy one-third o:f the total
protein require.ment o:f a cow yielding 2 - 3 gallons o:f milk per day.
Rumen microbial protein lias shown by McNaught, Owen, Henry
and Kon (1954) to have a. high digestibility and biological value for

- 9 -
rats and, in the absence of suitable methods for direct measurement,
it is assum.ed to have a similarly high value for the host animal, for
ruminants are independant of dietary sources of amino acids regarded
as essential for monogastric animals (Loosli et al., 1949). --Nitrogen metabolism in the rumen is thus characterized by
a dynamic equilibrium between the various ni trogenous components of
the rumen contents, with degradative, absorptive, and synthetic pro-
cesses occurring sim.ultaneously. The net result is that a considerable
proportion of the protein nitrogen and NPN entering the rumen event-
ually leaves as microbial protein, which is digested by animal prot
eases in the abomasum and small intestine (Pounden, Ferguson and Hibbs,
1950).
C. Utilization of NPN Feed Supplements by Ruminants
The conversion of rumen NPN to microbial protein was quickly
recognized as a process which could be exploited in ruminant feeding
practice to conserve relatively expansive dietary protein. Many NPN
compounds, auch as ammonium salta, urea, biuret and creatine, and
ammoniated by-products, auch as molasses and com cobs, have been
tested for their protein replacement value when fed as dietary supple-
ments. A major proportion of this work has been based upon feeding
trials, and these have clearly shown that auch supplements can support
weight gain and a positive nitrogen balance in the ruminant animal.
Because of its cheapness, high nitrogen content and commercial

- 10 -
ava.ilibility, ure a has recei ved a great deal of attention. Conditions
for its most efficient use in ruminant rations have been well defined
on the basis of the resulta of innumerable feeding experimenta, and
it is clear that urea ean supply a proportion of the nitrogen in an
otœ rwise nutritionall.y adequate diet required by daœstic ruminants
for maintenance, growth, fattening and milk production.
It is generally considered, however, tha.t some preformed
dietary protein is a neeessary constituent of tll."EErsupplemnted rations.
The proportion of the total ration nitrogen which should be represented
by protein is dependent on the nature of otœr canponants of the dist.
es:t:acially the carbohydrate (Belasco, 1956), and the nitrogen equllib
rium of the animal. Hamilton, Robinson and Johnson (1948) found tha.t
25% of tœ total nitrogen of the diet of lambs should consist of pre
formed protein when the protein equivalent wa.s not more than 12%.
The anima.ls then thrived when urea nitrogen constituted the remaining
75% of the total ration nitrogen. Harris and Mitehall (1941), on the
othar hand, ma.intained sheep in body and nitrogen equilibrium when
urea provided 9C1I> o~ the ni.trogen requi.re.œnt. At nitrogen equi.J.i.bri.um.
the biologicaJ. value of urea nitrogen was found to be 62 whereas that
of casein nitrogen was 79.
The pref'ormed protein requirement in urea-supple.mented
rations is generally accounted for in terms of a limitation in the
quantity of protein synthesized in the rumen of urea-fed a.nima.ls
which is imposed by the nature of the material ingested. Evidence

-ll-
has been obtained to shaw tha.t the limiting factor may be sulphur.
Thomas, Loosli, Williams and Maynard (1951) round that growing lambs
deficient in dietary su1phur did not utilize urea nitrogen; addition
ot inorganic sulphur to the diet, howewr, caused an increase in
lfeight and a change to positive nitrogen and sulphur balance in the
animals. The addition of Jœ thionine to a ration containing ure a as
a source or nitrogen has also been shown to increase the rate or gain
in weight and the nitrogen balance or lambs (Loosli and Harris, 1945).
Johanson, Yoir and Underwood (1949) have shown rumen bact
erial protein to be particularly rich in the sulphur-containing
amino acids, cystine and methionine, and prefonned dietary protein
may therefore represent a source or sulphur without which bacterial
protein synthesis in the rumen is inadequate to supply the nutritional
requirements or the animal.

PartI
The Attempted Isolation of Anaerobie Urease-produeing
Baeteria from the Rwœn Contents of a Sheep.

INTRODUCTION
An essential prerequisite to the detailed study of a given
enzyme is tbat it should be available in a puritied state. Urease
was first crystallized from jack-beans by Sumner in 1926, and since
tbat tim.e crystalline jack-bean urease bas been t:œ subject of a
fonn.idable am.ount of exper.1mentalwork. Despite the fact tbat the
rumen is a particularl.J rich source of bacterial urease, even a
crude purification of the enzyme from this source bas never been
attempted, because the first step in such a procedure, namely the
isolation in pure culture of symbiotic rumen bacteria which might
make a significant contribution to rumen urease activity, has not
yet been successfully accomplished.
In the present investigation, the bacterial origin of rumen
urease was first confinn.ed, and several attempts were then made to
isola te anaerobie urease -producing bacteria from the rumen in order
to provide a source of enzyme for subsequent studies on its purific
ation and properties.

LITIRATDRE REVIEW
A. ff;vdrolYsis of Urea in the Rumen
Although the use of urea as a prote in replacement in
ruminant rations has been tœ subject of much experimental work,
relatively little is known of the mechanisms by which it is broken
dawn and utilized in the rumen. As alre ady indicated, however, ure a
also enters the rumen as an integral part of the physiological nitro
gan cycle of the ruminant animal. Thus the mechanism of urea break
dawn is of interest not only for an understanding of the .f'ate o.f'
dietary urea but also because the metabolisœ o.f' urea is a normal func
tion o.f' rumen contents even in animals on natural diets.
Wagner, Booth,Bohstedt and Hart (1941) fed urea to a hei.f'er
at the rate o.f' 1% of the ration dry matter and found that it had dis
appeared completely from the rumen in 1 hr. Since the levels o.f'
ammonia and protein increased during this period, it seemed probable
that the urea nitrogen had been converted to anmonia and to protein
nitrogen.
Pearson and Smith (1943a) concluded that the .f'irst stage
in urea utilization in the bovine rumen was hydrolysis of the urea
to ammonia by the enzyme urease. Incubation of strained llquor with
urea in vitro, with subsequent determination of the amm.onia released,
provided a means of measuring the urease activity of the liquor. It

- 15-
was shown that urease activity was manifested in the rwuen at all
times of the day to auch a degree that 100 g. of rwuen contents
could completely hydrolyze 100 mg. of urea in 1 br. Indeed substrate
concentration had little affect upon the rate of hydrolysis over the
range 66 - 714 mg. of urea per 100 g. of rumen contents, which indi
cated that conversion of all the urea likely to be fed to a ruminant
as a protein replacement would readily occur within l hr. of feeding.
This was confirmed by Rys, Gorski and Styczynski (1956) who fed a
sheep 16 g. of urea per day, together with non-green fodder, o-ver a
4 hr. period. Urea was detectable in the rumen contents at the end
of the feeding period but not l hr. t:œreafter.
Since the diet of the animal used by Pearson and Smith as
a source of rumen contenta contained only inaignificant amounts of
urease, and be cause urease is not secreted by the rumen mucosa or in
the saliva, they concluded that the enzyme was elaborated by the rumen
microorganisme. An attem.pt to prepare a sterile but enz;yma.tically
active filtrate of rumen liquor by Berkefeld filtration was unsuccess
tul., however, as was an attempt to separate the enzyme .f'rom bacteria
by law speed (1000 r.p.m.) centrifugation.
The reason for these failures becam.e clear from the work of
Gibbons and McCarthy (1957). These workers determined the urease
activity of fractions of rumen liquor crudely prepared by differentiai
centrifugation. They shoYJed the rumen bacteria to be largely reapons
ible for rumen urease activity, and the complete absence of activity

- 16 -
from the cell-f'ree rumen liquor indicated that the enzyme was entirely
intracellular.
Soejima, Sugawara and Shimura (1958) have postulated that
rumen protozoa can produce urease adaptively in a urea-containing
medium af'ter a 3 hr. lag period. In view of' the rapid rate of urea
hydrolysis by rumen bacteria, however, it seems unlikely that adaptive
protozoal urease could have any quantitative significance in ~·
B. Enumeration of Rumen Bacteria.
The enumeration of rumen bacteria has been generally
recognized as an important starting point in studies of the rumen
microbial population, since knowledge of the total number of organisns
present, or of the numbers in various physiological groups, provides
a useful method of def'ining the character of' the population (Warner,
1962).
Despite the comp1exity of the rumen environment, involving
as it does numerous and little understood relationships between
different groups o.f organisme, the enumeration o.f the rumen micro
population is achieved by methods which are essentially similar to
those used for counting organisms in simpler environments (Hobson,
1961). They f'all into two categories: direct counting of organisms
in counting chambers or in stained smears, and viable counts by
cultural means. Neither of these methods, however, is really adequate
for counting rumen bacteria. Although direct methods still suf'f'er

- 17 -
from the disadva.ntages that living bacterial cella are trequently
indistinguishable fran plant traS'Jil.ents and dead cella, sOJœ recent
modifications in the se œthods (Warner, 1962) have he1ped partially
to overcome auch ditticulties and direct counting of rumen micro
organisme appears to be gaining considerable tavour as a rapid and
reasonably accurate means of enumerating the rumen population.
Cultural methods, on the other hand, which depend upon the
ability ot indi vidual organisme to grow in artificial media, are
limited in accuracy by tœ ditticulties invo1ved in creating artitic
ial conditions suitab1e tor the growth of all of the various members
ot the rumen population. No medium with the necessary degree of
non-selsctirlty to pe:rmit growth of the many types of rumen bacteria
has yet been devised. Neverthe1ess, viable counts have been widely
used in etudies of the rumen microtlora, and Bryant (1959) has pre
sented a canprehensive review of the methods adopted by various
worlœrs, inc1uding a discussion of the canposition of the many media
used. The most valid method of assessing the ability of a given medium
to support the growth of the mi:xed rumen fi.ora. is by corre~a.tion of
the cultural counts it provides lr.i.th direct counts of bacteria in the
same samp1e of rumen tluid. Som.e workers, tor example Hungate (1947)
and Wilson and Briggs (1955), have obtained auch correlations, but
others, tor example Doetsch, Robinson am Shaw (1952) and Bauma.n and
Foster (1956), have considered media gi ving viable counts of 108 -
1010 organisme per g. of rumen contents to be satisfactory and have

- 18 -
not com.pared viable and dire ct counts.
Wilson and Briggs (1955) developed a medium and technique
suitable for speedy and large scala use in the determination of numbers
of viable rumen bacteria. They modified the rainforced clostridial
medium of Hirsch and Grinsted (1954) by the inclusion of a NaHC03-C02
buffer, and used it to enum.erate the bacteria in bovine rumen contents
by mea.ns of a dilution technique. Anaerobie conditions •re achieved
by satura ting the sterile medium. wi th carbon dioxide and maintained
by dispensing it, without exposure to the atmosphare, into tubes
containing sterile liquid paraffin. The initial ten-fold dilution
of rwœn contents was made in a buffared redueing solution, and
subsequent dilutions were made directly in the culture medium., a
simple procedure which helped to main tain anaerobiosis. Counts of
108 - lolO organisms per g. were regularly obtained in this way.
These correlated well w:i.th microscopie counts and were not improved
by enriehm.ent of the medium with rumen fluid.
C. Cultural Detection of Ureasa Production by Ba.cteria.
Species of the genus Proteus, and .lll.alll.bers of the paracolon
aerobacter and intemed:iate groups of baeteria ditfer from other Gram
negative enteric bacilli in that they produœ urease. On the basie
of a po si ti ve urease test, the se organisme may be eliminated from
turther study in schemas for the identification of intestinal
pathogens. Considerable attention has therefore been given to

- 19 -
methods :for the qualitative determination of urease production.
Nearly all the tests which hava been devised for urease
depend upon demonstration of the change in reaction of the test
medium when amm.onia is released from urea by the action of the
enzyme. Ruatigian and Stuart (1941) described a highly buffered
llquid medium., containing urea, which would support the growth of
Proteus spp. Urease-positive cultures attacked urea with the re
lease of amm.onia which caused the developœnt of alkaline conditions,
this change being deteetab1e by the use of a suitable indicator.
Christensen (1946) eonsidered this medium. to be too weak nutritionally
and too highly buffered to enable the detection of urease activity in
Gram-negative intestinal baeteria other than Proteus spp. Accord
ingly, he devised a more weakly buffered agar medium., containing urea,
peptone and dextrose. Cultures which produce ammonia from peptone or
. acid from dextrose, however, may interfere with tm detection of
urease activity on Christensen' s medium (Brisou, 1949) which is
therefore unsuitable for use with non-ente rie, weakly ureo1ytie
organisme.
Urease test media which do not involva growth of tbe
organism., and therefore need to contain no nutrients, are preferred
wbere non-enteric organisme are coneerœd. In this type of test a heav;y
suspension of the organism. is added to a buffered solution containing
urea, salta and indicator, but no carbon source. After incubation
for a few hours the indicator changes colour if urease is present,

- 20-
and sinee there is no other source of ammonia, alkali production must
be due to ureolysis. A further advantage of this type of test is
that sterility is not required.
D. Urease-produeing Rumen Bacteria.
Of the bacteria which are responsible for the production of
rumen urease very little is known, since species comprising the pre
dominating ureolytio microflora of the rumen have yet to be isolated.
With few exceptions ali tœ urease-producing rumen isolates so far
obtained have been facultative anaerobes developing only a weak
activity in artificial culture.
llann, :Masson and Oxford (1954) isolated 11 ureolytic strains
of eoagulase-negative Staphylococcus spp. from the rumen of a hay-fed
sheep 1 and Mann and Oxford (1955) isolated 8 similar strains from the
rumen of a 14-week old calf. :MacKay and Oxford (1954) obtained a
number of ooliform organisme from oalves and sheep at dilutions in
dicating populations of 104 - 106 organisme per g. of rumen contents.
13% of the 1solatesshowed a weak urease act1v:i.ty and 11ere 1d.ent1:r:isd
as Aerobacter aerogenes. A small Gram-negative rod with a stronger
a.otivity was isola.ted at dilutions of 1/104 from the rumens of two
ealves but this orga.niam was not identified.
Faculta.tively anaerobie rumen bacteria are relatively easy
to enumerate since tœy have non-exacting nutritiona.l requirements
and the ability to grow aerobioally. Generally, populations of the se

-21-
organisms in the rumen seldan exceed 106 - 10? cells p:.~r g. of rumen
contents (Bryant, 1959). Since the organisms isola.ted have had onl.y
a weak urease activity one is led to conclude that the facultative
anaerobes are not responsible for the bulk of rumen urease production.
Experimental evidence to support this conclusion was obtained by
Gibbons and :U:cCarthy (1957). These workers prepared a washed suspen
sion of the mixed rumen microfiora which contained 1 x lolO organisme
per ml. and showed urease activity. Of this population, facultative
bacteria 11ere shown by an aerobic plate count to comprise 0.04%, or
4 x 106 organisme per ml. Sixty-nine colonies were isolated from the
plates and one-third of these 11ere found to be urease-positive on
Christensen 1s medium. (Christensen, 1946). Washed cell suspensions
of three of the most strongly ureolytic strains were prepared at a
density (5 x 106 organisa per ml.) approxima. ting that of facultative
anaerobes in tm suspension of rumen organisms, but not one suspension
shawed a measurable urease activity. Neither was activity shown by
wasbed suspensions of Proteus Yulgaris and !J:. mirabilis at the same
density. In this conte.xt it may be noted that Huhtanen and Ga.ll
(1955) !ound 2 x 109 cella per ml. of Pr. mirabilis necessary to give
a urease activity equivalent to that of rumen fluid.
With respect to the low urease activity .manifested by
organisme so far isolated from the rum.en, Gibbons and his associa tes
(Gibbons am :U:cCarthy, 1957; Gibbons and Doetsch, 1959) have pointed
out that an organism show.ing only a weak activity when grown on

-22-
artiticial media may well produce a higber specifie activity under
conditions existing in the rumen. The totality of evidence suggests,
however, that the predominating urease-producing bacteria in rumen
contents are obligately anaerobie and have yet to be isolated in pure
culture, althougb several species of this type producing small amounts
of urease have been obtained. Gall, Stark and Loosli (1947), tor
example, isolated a number of strictly anaerobie Gram-positive rode
vith urease activity tram rumen contents of cattle and sheep at dil~
tions of l/1olO - l/1011• In pure culture, however, where initiall7
the sole nitrogen source was urea at a concentration of o.o4%, gener
ally leas than one-fifth of the urea wa.s hJ'drolyzed in 48 hr.
The aim of a recent stud)r b7 Gibbons and Doetsch (1959) was
the isolation of obligately anaerobie urea-hydrolyzing bacterie. trom
bovine rumen contents. Using a 1% rumen tluid medium., the authors
succeeded in isolating a G~positive rod resembling Lactobacillus
bifidus, which at the maximum urease activity obtained in artifieial
culture h7drolyzed 36.8 J.LM ot urea per mg. of eell-nitrogen per hr.
Enzym.e production was JU.rkedl.y intluenced by the type of medium. used
for cultivating the organism. In a casein hJ'drolyzate medium, a chemi
eally defined amino acid medium and a medium containing autoclaved rumen
fiuid, al1 of which were high in organic nitrogen, more urease was
produced as the level of nitrogenous eompounds was deereased. In
the reinforced clostridialmedium ot Hirsch and Grinsted (1954), on
the other hand, lese urease was produced as the amount of organic

-23-
nitrogen was decreased.. •zym.e production was impaired. in the
presence of urea 1 ammonium sulpha te and 1118llg&nese 1 al.though iron
was able to overcom.e the inhibitory properties of the latter.
Inhibition of enzym.e production by Mnft at a concentration of
o.ool% (w/v) MnS04.H2o was noted but the ion had no effect on the
activity of prefo~ed urease. The authors could not estiψte the
contribution 'Which this ,&. bifidus stre.in might make to rumen urease
activity .3::!! vivo, but beca.use it was present to the extent of only
about 105 cells per g. of rumen contents it wa.s not considered to be
the organism. mainly responsible for rumen urease activity.
No other report concerning the isolation in pure culture
of ureo~ic rumen baeteria was found; apparently organism.s eom
prising the predominating urease-producing microflora of the rumen
still await isolation.

MATERIAIS AND METHODS
A. Experimental Animal s.
Five rum1nant animals, three sheep and two steers, fed
different diets, were used as sol:lrces of' runan contents.
a) Sheep 2
The a.nimal from which rurœn contents were routinely coll
ected was an adult Cheviot eft, which weighed 60 kg. and was equipped
with a rumen fistula. The animal was kept in a metabollsm crate and
fed a ration consisting of 1400 g. of chopped timothy or brome-grass
bay, 175 g. of ground bar ley and 21 g. of urea {fertilizer grade).
Tb3 concentrate and urea were mixed with the hay at the time of :read
ing and the .mixture was readily consumed by the animal. It was offered
daily at 8 a.m. and remained accessible throughout the following 24 hr.
period. Water was available ,24 llbitUfll.
b) Sheep 35 and 37
These two non-:f'istul.ated animals were kept in metabolism
crates am fed a d.iet of deh.ydrated alfalfa. meal containing 1%
vegetable oil and ethoxyquin, a vitamin A stabilizer.
e) Steers l ani 2
These fistulated Holstein anim.als, which were tethered in
loo se boxes with access to water ani mineral licks, were maintained
on a diet of alfalfa hay.

- 25 -
B. Collection of Rumen Contents.
a) Sheep 2
Following initial installation of the animal in the meta
bolisn crate, a two week period of adaption to confinement and to the
ration was allowed to elapse before the first collection of rumen
contents was made. Sam.ples of rumen contents were withdrawn through
the fistula into a Buchner tlask by means of an S mm. Nalgon tube,
under a negative pressure of 500 mm. mercury.
b) Sheep 35 and 37
Sam.ples of rumen contents were withdrawn in the way
described above except that the tube was introduced into the rumen
via the mouth and oesophagus.
c) Ste ers 1 and 2
Solid material was withdralfll from the rumen by hand through
the fistula, and then compressed to express the liquid portion.
C. Preparation of Rumen Fluid.
Sam.ple s of :rumen cœtents were returned to the laboratory
in completely filled bottles. These œre imlœrsed in ice-water when
measurements of urease activity were to be made, and wrapped in cotton
wool in an insulated bucket, previously wa:med to 37°C, when rumen
microorganisns were to be cultured. In the laboratory the material
was routinely strained through four layera of cheesecloth before use
to remove larger food particles. The resulting "filtrateu, referred

-26-
to as "rumen fiuid", wa.s very rich in microorganisms and was used
throughout the present experimenta.
D. Fraetionation of Rumen Fluid.
a) Microbial and supernatant tractions
Aliquots of 25 ml. of rumen fiuid •re centri!uged at
13,200 x G for 10 min. at 10°C. The clsar brown supernatant liquor
was decanted and tl:e cella were then washed by suspending them in
o.021l-phosphate or maleate butter at pH6.8, centrituging the suspen
sion and again decanting tl:e supernatant liquid. This procedure was
generally repeated twice. The microbial fraction was finally resuspend
ded in butter and tl:e suspension diluted to 25 ml.
b) Subfractionation of tœ microbial fraction
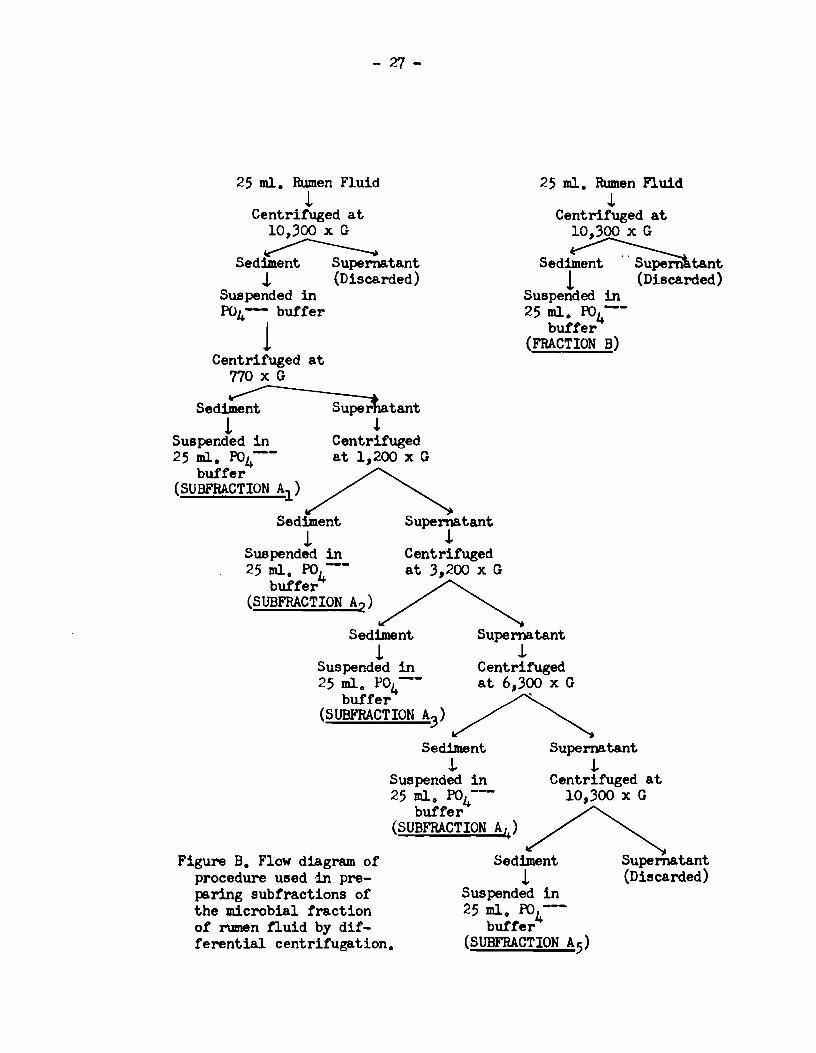
This was carried out by the differentia! centrifugation
procedure shown in the flow diagram, Figure B. Two 25 ml. volumes of
rumen fiuid "nere centrituged at 10,300 x G for 10 min. and the sedi
ments resuspended in ma.leate butter. One suspension was then centri
tuged repeatedl.y at progressi-wly increasing speeds for 10 min. periods;
atter each centrifugation the supernatant liquid was decanted and re
centrituged at the next higher speed. The centrifuge speeds were
arbitrarily chosen and were equivalent to 770 x G, 1,200 x G, 3,200 x G,
6,300 x Gand 10,300 x G, respectively. The sediments obtaiœd at
each centrifugation were each made up to 25 ml. with ma.lsate butter.
Six suspensions resulted from the application of this procedure. Each
was examined by phase contrast microscopy to determine the predominant
- 27-
25 ml. Rumen Fluid l,
25 ml. Rumen Fluid J.
Centrifuged at 10,300 x G
Centrituged at
~ 10,300 x G
Sed~tant l (Disearded)
Suspended in
Sediment Supernatant ! (Disearded)
Suspended in P04- butter
1 25 ml. P04--
butter
Centri.fuged at 770 x G
Sed~tant ! !
Suspended in Centrituged 25 ml. P04 -- at 11 200 x G
butter (SUBFRACTION A;:a_)
Sediment .L
Suspended in 25 ml. P04-
butfer (SUBFRACTION A2)
Supernatant J.
Centrituged at 31 200 x G
(FRACTION B)
Sediment Supernatant .L l.
Suspended in Centrituged 25 mlo Po4-- at 61300 x G
butter ~ (SUBFRACTioti A3) / ~
Sediment Supernatant l. i
Suspended in Centrituged at 25 ml. P04 -- 10,300 x G
butter (SUBFRACTION At•)
Figure B. Flow diagram of' procedure used ,in preparing subtractions of' the microbial traction of rumen tluid by dif'terential centrifugation.
Sediment L
Suspended in 25 ml. P04-
butf'er (SUBFRACTION A5)
Supematant (Discarded)

- 2S-
types of organism.s present.
c) Separation of rumen protozoal fraction
The protozoa were initially removed from rumen .tluid by
the sedimentation technique of Masson and Oxford (1951).
A sample of rumen contents was retumed to the laborator;y
in an insulated container pre-warmed to 37°C. Glassware and solutions
were similarly pre-warmed, and all manipulations ware carried out in
a room mainta.ined at this temperature. 500 ml. of rumen fluid, to
which was added 0.05% (w/v) each of glucose and maltose (Sugden, 1953),
was allo11ed to stand in a separatory tunnel for là hr. The protozoa
which had sedimented during this time were then drawn off and re susp
ended in the bu:f'fer solution of Abou Akkada and Howard (1960). This
contained: K2HP04 0.63%, KH;!04 0.50%, NaHCO., 0.10%, NaCl 0.06%,
Mgso4 o.ol%, CaCl2 0.009%, Na~ 0.02%, and chloramphenicol 5 pg. par
ml. The pH of the bu:f'fer was ad.justed to 6.9. The protozoal suspen-
sion in butter was replaced in the tunnel. This was allowed to stand
for a further li hrs. and tm sediment of protozoa was again drawn off.
This procedure was repeated t'Wice and tbs greyish-white protozoal
fraction was finally suspended in 500 ml. of rumen supernatant liquor
to give a cell density approximating that of the original rumen :f'luid.
The purity of the suspension was checked microscopically in a hanging
drop preparation.
The rumen .tluid fran which the protozoa had been removed was
centrif'uged at 75 x G for 2 min. to remove residual protozoa, and the

- 29 -
bacteria-rich supernatant, which is ref'erred to as protozoa-.t'ree
rumen .t'luid, was decanted and retained.
1. Measurement of' Ure a se Activi ty.
The activity of rumen urease preparations was routinely
measured at 37°C by the manometric method of Huhta.nen am Gall (1955).
In this procedure , the hydrolysis is carried out in convent
ional Warburg vessels under an atmosphere of co2 which is in equilib
rium wi th C~ in the aqueous phase. Ure a is hydrolyzed by urease to
amm.onia and co2 according to the equation:
CO(NH2}2 + H2) ____. 2NH3 + C02
For ewry mole of' C02 produced, therefore, 2 moles of NH3 are released,
and this NH3 is retained in the slightly acid aqueous phase. Since
twice as muàlalkali as acid is released, C02 is absorbed from the
atm.osphere by the aqueous phase to maintain equilibrium conditions.
The volume of co2 absorbed, which is measured in terms of changes in
pressure of the gas phase in the conventional mannar, is proportional
to the amount ot urea hydrolyzed.
The vessels contained 3 ml. of the urease preparation,
suspended in either rumen supernatant liquor or in O.OaL-maleate
buffer at pH 6.8, in the main compartment and 0.2 ml. of 0.3M-u.rea
solution in the side-arm. With manometer tapa and vent plugs open,
the vessels were gassed with 100% co2 and shaken for 10 min. to
achieve temperature equilibration. At the end of this time the tapa

-30-
and vent plugs were simultaneously closed, leaving an atm.osphere of
C~ inside the vessels. Shaking was continued for a further 5 min.
to allow equilibration of the gas with the liquid phase in the
vessels; the substrate was then tipped in and .manom.eter readings
begun. The shaking amplitude was 4 cm.. and the frequency 90
oscillations per min.
Determinations were carried out at least in duplicata and
endogenous controls of the urease preparation in the absence of ure&
were included for each determination. Readings 11ere generall;y taken
every 15 min. for a 1 hr. period, but when complete hydrolysis of
substrate was required, incubation was continued until C02 absorption
ceased before a final reading was taken. The com.pl.eteness of hydro-
lysis was them checked by Nessleriza.tion of the rel.eased NH3• This
procedure pe:rmitted determination of the volume of co2
absorbed per
micran.ole of NH3 rel.eased during the hydrolysis of a known am.ount of
urea, ani this relationship was then used in the calculation of
indices of urease activity for suspensions of rumen urease in sub-
sequent. e.x:periments. The urease activit.y index, <mH3
, was expressed
in conventional manner in te:rms of the we ight of ammonia released
during urea hydrolysis rather than in terms of the amount of urea
hydrolyzed. ~ • Jlg• of NH3 released per ml. of suspension per hr.
F. Bacterial Viable Counts.
Samples of rumen contents •re taken from Sheep 2 ten hours
after feeding. All glassware and equipœrrt. used for collection and
preparation of the sam.ples was previously sterilized by autoclaving.

-31-
a) Total viable counte
100 ml. of strained .fluid, containing a drop of sterils
Dow Anti.toam A, was agitated for 30 sec. in a sterile Waring blendor
jar to disrupt chains and clumps of organisme. During agitation a
stream of sterile C02 was blown into tm jar to minimize aeration of
the .fluid. The initial ten-fold dilution wa.s prepared in the butfered
reducing solution (dilution fluid A) of Wilson and Briggs (1955)
containing: Na~ 0.02%, NaHC0:3 0.25%, and Tween-80 0.05% {v/v). The
pH of this solution was adjusted to 6.8. Subsequent ten-fold dilutions
wre prepared in a solution (dilution fiuid B) which bad the follow.ing
composition: yeast extract (Difco) 0.3%, beef extract (Difco) 1.0%,
peptone (Difco) 1.0%, soluble starch o.l%. dextrose 0.5%, NaCl 0.5%,
cysteine hydrochloride 0.05%, citric acid 0.175%, Na2HP04.7H20 1.8%,
and Tween-80 0.05% (v/v). The fluid was filtered and the pH adjusted
to 6.8 before autoclaving. The culture medium bad the sam.e composit
ion as dilution fiuid B except that Tween-80 was omitted and 0.05% {w/v)
agar was includad. It th us had a similar composition to the "modi.tied
reinforced clostridial œdium" of Wilson and Briggs (1955) • the
NaHOO, - 002 buffer system used by these workers being replaced in
the present case by Na2HP04 and citric acid. This modification in
buffer composition was made because in prelim.inary experimenta it
was found that following gassing of the medium with co2 a precipitate
developed during incubation which was ditficult to distinguish from
bacterial growth. The madium is subsequently referred to as "buffered
reinforced clostridial broth" (BRCB) •

- 32-
Immediataly on removal from the autoclave, the medium was
aseptically dispensed into tubes containing 2 ml. of sterile liquid
paraffin to provide an anaerobie seal. No further anaerobie pre
cautions were taken. Tubed medium was hsld at 37°C for 24 hr.
bef'ore use and any tubes showing growth were discarded.
Five 1 ml. aliquots of' each dilution of' rumen f'luid from
1/102 to l/1ol2 were inoculated into 9 ml. of' BRCB, which was thsn
incubated a.t 37°C for a. minimum of 5 days and examined thereaf'ter
every 48 hr. untll no further initiation of growth occurred. The
numbers of tubes showing growth at the three signif'icant highest
dilutions (Prescott_ Winslow and McGrady, 1946) 111ere noted and used
to calculate the most probable number of viable bacteria par ml. of
strained rumen fiuid from the MPN tables of Hoskins (1934).
b) Counts of viable ureolytic organisme
All growing cultures were examined by phase contrast
microscopy to determine the predominating types of' organisme present
and wre then centrifuged. Tœ supernatant liquid wa.s decanted and
the cella 11ere washed by resuspending them in 0.02M-maleate buffer
at pH 6.8, and centrif'uging the suspension. Again ths supematant
liquid was decanted and the cella were f'inally suspended in 1 ml. of'
m.aleate buffer. To this suspension was then added 1 ml. of a solution
of 10% urea and 0.0001% phenol red in 0.9% NaCl, neutralized to an
orange eolour with dllute acetie acid. Finally 1 ml. of paraffin
was added and the tubes were incubated at 37°C. Urea hydrolysis was

-33-
shawn by a change in colour of the indicator to a deep cerise. Tubes
wre exam:ined trequently and those showing a positive result in 48 hr.
were considered to have contained ureolytic bacteria, although soma
times positive resulta were obtained in as little as 2 hr. A 24 hr.
culture of Proteus vulgaris, grown in 10 ml. of nutrient broth and
prepared in the way described, needed 6 hr. to give a positive result.
The llPH tables of Hosldns (1934) were used to calculate
the most probable number of viable ureolytic bacteria in tœ strained
rumen .fluid fran the number of cultures prepared from the three con
secutive highest dilutions of rumen .fluid which showed urea hydrolysis;
the calculations were based on a total of five tubes per dilution even
though less than ti ve may have initially shown growth.
G. Urease-producing Bacteria traœ Rumen Fluid.
a) Isolation
i) Isolations fran buffered reinforced clostridial broth
Urea-hydrolyzing bacteria were isolated from cultures used
!or enumeration o! -the ureol.ytic rum.en f'J.ora. Trana!ers made f'rom
cultures subsequently .t'ound to contain ureolytic bacteria. were
streaked on plates containing buttered reintorced clostridial broth
solidified by the addition of 1.45% agar, and the rest were discarded.
This solid medium is subsequently reterred to a.s bu.t'fered reintorced
clostridial agar (BRCA). The plates were incubated .t'or 24 hr. at
37°C in Brewer anaerobie jars under an atmosphere of N2 and co2
(95%: 5%), and colonies were then picked at random onto BRCA slopes.

-34-
After incubation at 37°C, the isolates 1ere checked for morphologieal
purity and for response to oxygen. All grew as well aerobically as
anaerobically and therefore no attempt was made to exclude atmospheric
o::xygen in subsequent work. Transfera were made to slopes of urea
agar (Difco) and organisms producing an alkaline reaction within 12 hr.
0 at 37 C were selected for further study. Cultures of the se organisms
were maintained at 10°C on slopes of tryptone glucose extract agar
(Difco) supplemented with 0.2% yeast extract.
ii) Isolations from supplemented rumen supernatant
liquor medium.
Efforts were made to devise a medium. suitable for prepa.ring
enriclment cultures of rumen ureolytic bacteria. For this purpose a
series of media were devised, each of which eontained 80% rumen
supernatant liquor, 0.5% NaH<X3 and 0.05% cysteine hydrochloride,
together with various canbinations of the following nutrients at
concentrations of o. 5% each: glucose, eellobiose, sodium acetate,
phytone, easamino acids and urea. In addition, to one medium was
added the mineral solution of Hu.ngate (1947) modified to provide in
the medium the folloldng final concentrations: NaCl 0.036%, (NH4),~4 0.012%, K2HP04 0.02%, KH2Po4 0.012%, CaC12 0.006%, llgS04 0.006%,
KnS04 0.0001%, FeCl3 0.0001%, CoCl2 0.0001%, and (NH4)6 M~024 0.0001%.
Appropria te eombinations or solutions of glucose, cellobiose,
sodium. acetate , phytone , easamino acids, minerals and cysteine hydro
chlorida in a volume or 2 ml. or water were added to screw-capped test

-35-
tubes "Wbich 11ere them autoclaved at 121°C for 15 min. Eight ml. of
fresh rumen liquor, f'ollowed by solutions of urea and NaHCù:3 which
had been sterilized by membrane filtration, ll3re then added. The
final volume of medium. in each case was approximately li ml.
Each tube was inoculated with two drops of f'resh rumen f'luid
and the free space above the liquid was flushed with sterile co2.
When the caps were tightened and the tubes shaken' the co2 absorbed
gaw a final pH in each medium of' appro:ximately 6.8. The cultures
were incubated at 37°0 for 48 hr. and then examined for growth as
evidenced by the developnent of turbidity. A number between 0 and 4
was assigned to each culture on the basis of a visual estimation of
tm amount of' growth present. Those cultures showing growth were
tested f'or the presence of ureolytic organism.s by suspending the cella
in the urease test solution described in section F(b) above. The
cell suspensions were observed at half-hourly intervals and the time
taken for the development of an alkaline reaction, as demonstrated by
a colour change in tm indicator, was recorded.
From the resulta of this study two rœdia were selected and
further tested for their ability to produce enrichment cultures of
rumen ureolytic bacteria. The media 'Were prepared in lOO ml. volumes
and each was inoculated with 1 ml. of strained rumen fluid. Tm
microbial fraction from 25 ml. of the rumen fiuid used for inoculation
of the media was washed by centrifuging, decanting the rumen super
natant liquor, resuspending the cella in 0.034-maleate butter at pH 6.8,

- 36-
centrif'uging the suspension, and again decanting the aupematant
liquid. The celle were finally suspended in maleate bu.ffer and the
suspension diluted to 25 ml. ldth bu.ffer. The optical density of a
1/10 dilution of this suspension was measured in a Coleman Junior
co1orimeter at 590 Illf• The instrument sh011ed maximum sensitivity
with the greenish cell suspension at this wave1ength. The urease
acti vi ties of the who1e rumen fiuid and of the cell suspension in
bu.ffer were then measured manometrically. After incubation of the
inoculated œdia at 37°C for 48 hr. the follawing procedure was used
with each medium. One ml. was transferred to 100 ml. of fresh medium.
Celle were then harvested from 25 ml. of the residual medium by
centri.fuging and l'Jere washed once by decanting the supematant 1iquid,
resuspending tœ cells in 0.0211-ma.1eate buffer, centri.fuging the
suspension and again decanting the supernatant liquid. The cella
were then suspended in maleate buffer and the suspension diluted
with buffer unti1 a 1/10 dilution of the suspension had an cptical
density at 590 IllJl equal to that of the di1uted suspension of cella
from rumen f1uid. In this way the density of the cells in the various
suspensions wa.s standardized. The urease activity of the cell suspen
sion was then œasured. By the repeated use of this procedure, the
urease activity developed in the enrichment media during tl:e incubation
of successive transfera of rumen organisme was compared to the activity
of the original rumen fiuid.
When a medium was f'ound which supported the d.eve1opnent of
enhanced urease activity, it was used for the a.ttempted isolation of

- 37 -
rumen ureolytic ba.cteria. For this purpose a culture of rumen micro
organisme enriched 'llith respect to urea.se a.ctivity, wa.s examined
microscopica.lly and then serially diluted in sterile O.OaL-m.a.lea.te
buffer, a.t pH 6.8, conta.ining 0.05% cysteine hydrochloride. Bottles
of dilution fluid were nushed with sterile COz and shaken, and 1 ml.
volumes in duplica.te of ea.ch dilution from l/10 to l/lolO -were pla.ted
with the enrichment medium., "Which wa.s solidified with 1.5% agar. The
plates 'Were incuba.ted in Brewer anaerobie jars a.t 37°C for 48 hr. under
an atmosphere of COz and Hz· After incubation, colonies were picked
from the plates into 3 ml. volumes of the enrichment medium.. These
were la.ter transferred to agar slopes of the same medium., and the
liquid cultures "Were tested for the presence of urea.se-producing
bacteria in the manner described in section F(b) above.
iii) Isolations of rumen staphylococci by enricbment in
a brain heart infusion - Na.Cl medium.
Rumen sta.phylococci were isola.ted from the rumen contents of
sheep Z by a modification of the staphylococcus enrichment method of
Wil.son, Poter and I.ew:i.s (~959). One ml.. ot aseptically prepared rumen
f~uid was a.dded to 30 ml. of brain heart infusion (Difco) containing
7.5% NaCl. The suspension wa.s shaken for 9 hr. in a wa.terbath at
37°0 and streak inoculations were tban made on plates of sta.phylococcus
medium no. 110 (Difco). Atter incubation for 36 hr. a.t 37°C, colonies
were picked into brain heart infusion medium. and subsequently
characterized.

- 38-
b) Identification of isolated organisme
Praliminary classification of the isolates into thair
respective genera was made from tm rasults of tests recommended by
Skerman (1959). Subsequent species identification was made according
to Bergey (1957).
c) Urease activity of uraolytic isolates
Each organiam wa.s inoculated on to tryptone glucose 0.2%
yeast extract agar contained in Roux bottles. The inoculated bottles
were incubated at 37°C for 48 hr. The cella were then washed off the
agar with O.OaL-maleate buffer, pH 6.8, and the suspension of cells
was centrifu.ged. The supernatant liquid wa.s decanted and the cells
were wa.shed once by resuspending them in buffer, centrifu.ging the
suspension and again decanting tm supernatant liquid. The cells
from two Roux bottles were finally suspended in about 8 ml. of buffer,
and 3 ml. volumes of this suspension were used for manom.etric measura
ments in duplicata of the urease activity of tm cella. The number of
organisms in the suspension wa.s detennined by a standard count using
sterile 0.9% NaCl. as the dil.uting fl.uid and tryptone gl.ucose 0.2%
yeast extract agar as ths plating medium. Plates were incubated at
37°C for 48 hr.

RESULTS
A. Urease Activity of Rumen Fluid and its Compg!!!nt Fractions.
The urease activities of samples of whole rumen tluid fran
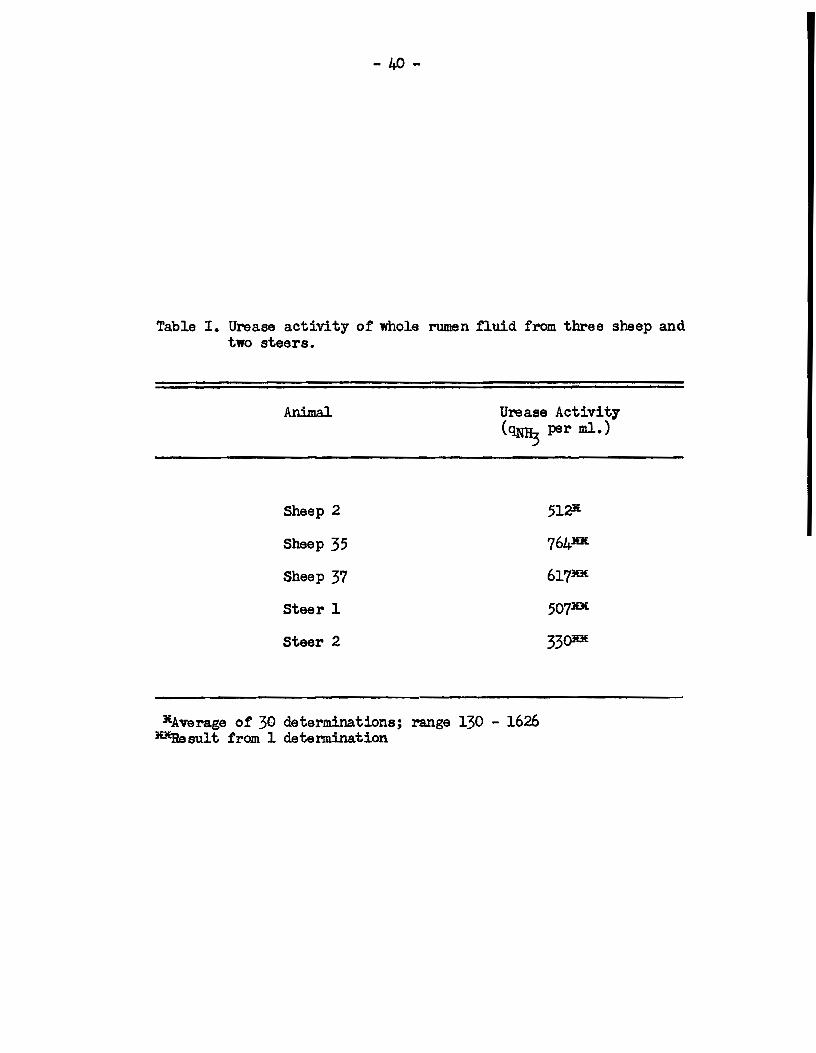
t:œ five experimental anima.ls used in this study are shown in Table I.
The range of activities found with sheep 2 represented a variation in
the amount of urea hydrolyzable to ammonia from 25 - 310 mg. per 100 ml.
of rumen fluid per hr., and therefore indicated a geœrally high leval.
of rumen urease activity. The average activity found with this urea
fed animal was sim.ilar to the levels of activity found in single det
em.ina tions wi th the othar four anima.ls, which were fed more natural
diets. It is evident from the resulta with sheep 2, however, that a
wide range of urease activities may be found from time to tim.e.
Clearly, therefore, a single determination of rumen urease activity
may not be indicative of the generally prevailing urea-hydrolyzing
capacity of rumen fluid from an individual animal.
Ali the experimenta subsequently reported in this section
"Were carried out with rumen fluid samples from sheep 2.
Table II shows the urease acti viti.es of the component
fractions of wbole rumen fluid, together with those of suspensions
of the hay and grain fed to the animal. The urease activity of the
whole fluid was almost quantitatively recovered in the protozoa-free
rumen tluid, tha.t is, in the bacterial fraction. (This fraction was
represented by whole tluid from which the protozoa had been removed
-40-
Table I. Urease activity of whole rumen fiuid from three sheep and two steers.
Sheep 2
Sheep 35
Sheep 37
Steer 1
Steer 2
Urease Activity (~ per ml.)
507lDE
33QlDf.
lEAverage of 30 determinations; range 130 - 1626 lilEResult from 1 determination

-41-
Table II. Urease activity of tractions or rumen tluid.
Fraction
Protozoal fraction
Protozoa-free rumen fiuid (bacteriai traction)
Rumen supernatant liquor
Dietary grain suspension
Dietary hay suspension
Whole rumen fiuid
Urease Activity (qNH_3 par ml.)
4
189
0
0
0
193

- 42-
by a sedimentation technique and subsequent centritugatio~ The
occurrence in the protozoal fraction of a small proportion of the
urease activity of 'Whole rwnen fluid wa.s probably due to the presence
in the suspension of small numbers of ureolytic bacteria. The absence
of ureolytic activity from the suspensions of hay and grain elim.inates
the presence of these components in the bacterial fraction as a poss
ible source of urease, and sinca no animal enzymes are sacreted by
the rumen epithelium (Dukes. 1955), the resulta show that the product
ion of urease in whole rumen fiuid is attributable entirely to the
rumen bacteria. Sinee there was no activity in the relatively cell
free rwnen supernatant liquor, i t is furthermore evident tha t the
enzyme activity assoeiated wi th the bacteria wasex clusively intra
cellula.r.
In order to find out whether urease activity was assoc:ia.ted
with a particular group of morphological types of rumen bacter:ia.,
subfractions of the rumen microbial population wera preparad by
differential centrifugation. Suspensions of these subfractions in
buffer wsre round to have the compositions and urease activities
shown in Table Ill. The resulta show that the bacteria responsible
for about 65% of rumen urease aetivity 'Wera the larger organisme
which were thrown down when tl:e suspension of the entire population
was eentrifuged at 1,200 x G. When occurring in clum.ps or chains,
many of these organisme sedimented at 770 x G. The subfraction
sedimenting at 3,200 x G, which contained many types of smaJJer
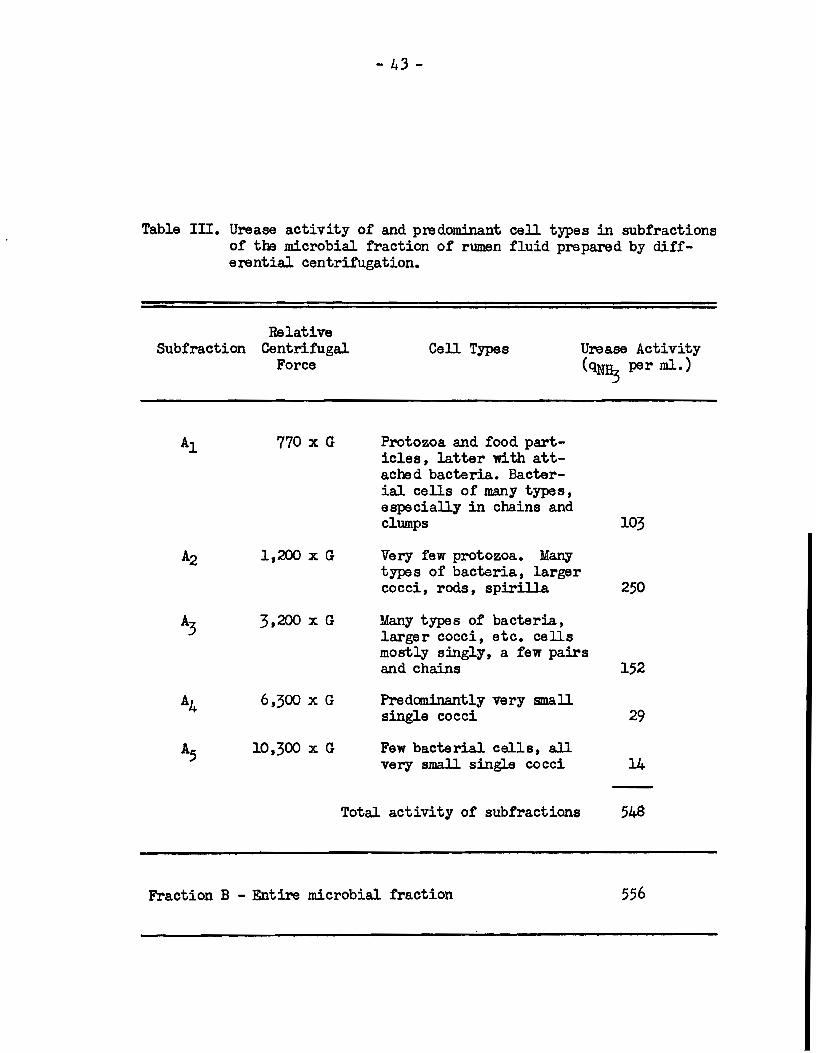
- 4:3-
Table Ill. Urease activity of and predomjnant cali types in subfractions of the microbial fraction of rumen fluid prepared by differentia! centrifugation.
Relative Subfraction Centrifuga!
Force
Al 770 x G
A.2 1,200 x G
A.3 3,200 x G
A4 6,300 x G
A5 10,300 x G
Cell Types
Protozoa and food part-icles, latter with att-achad bacteria. Bacter-ial cella of many types, especially in chains and clum.ps
Very few protozoa. Many
Urease Activity {qN~ per ml.)
103
type a of bacteria, larger cocci, roda, apirilla 250
Many types of bacteria, larger cocci, etc. cella most ].y singly, a few pairs and chaina 152
Predaninantly very small single cocci 29
Few ba.cterial cells, ali very sma.ll single cocci 14
-Total activity of subfractions 548
Fraction B - Entire microbial fraction 556

- 44-
bacteria, bad associated with it about 2!7% of the total urease
activity of the rumen microbial population. -Onl.y a com.paratively
low activity was associated with the very small cocci which were
obtained upon centrifuging the suspension above 3,200 x G.
These resulta suggest that rumen urease activity was
associated with a relatively limited number of types of rumen bacteri.a,
which appeared to comprise the larger organisms :round in rumen nuid.
B. Proportion of Ureolrtic Bacteria in Rumen Fluid.
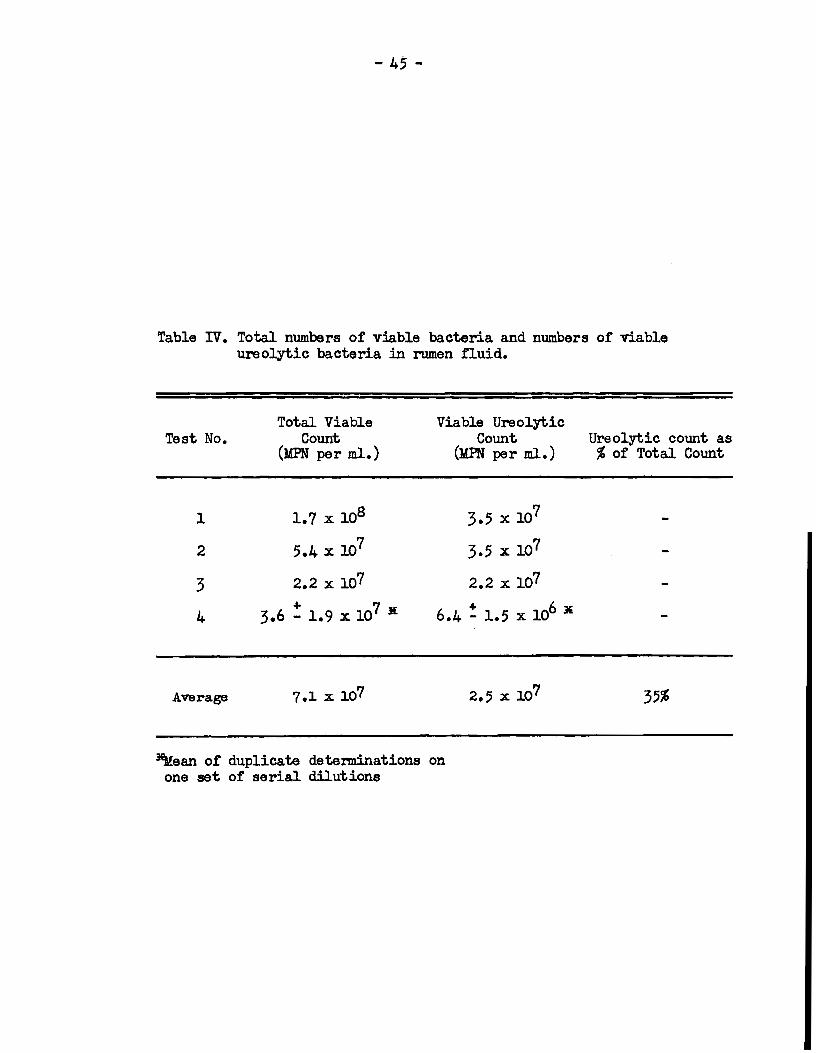
Evidence was obtained in the exper:iment described above that
organisms representing a l:imited group of rumen bacteria were mainly
responsible for rumen urease production. A review of the lite rature,
however, failed to reveal any estimate of the proportion of the total
numbers of rumen bacteria represented by urease-producing organisme.
An attempt was therefore made to estimate the proportion of ureolytic
bacteria in the rumen fluid of the urea-fed sheep. Using the method
of most probable numbers, the populations of viable bacteria and of
viable ureolyt.ic bacteria in rumen f'luid w:n-e determined on !our
different occasi0118 in a 10-week period. The results, which are
presented in Table IV, show tha.t the average proportion of rumen
bacteria :round to produce urease was 35%. Microscopie examination
of the cultures used in these determinations showed that non-m.otile,
Gram-negative cocci, arranged singly, in pairs and in short cha.ins,
were invariably present in predominating numbers in cultures with
- 4$-
Table IV. Total numbers of viable bacteria. and numbers of viable ureolytic bacteria in rumen fluid.
Test No.
1
2
3
4
Average
Total Viable Count
(MPN per ml.)
1.7 x 108
5.4 x 107
2.2 x 107 + 7 lE 3.6 - 1.9 x 10
Viable Ureolytic Count Ureolytic count as
(llPN per ml.) % of Total Count
3.5 x 107
3.5 x 107
2.2 x 107 + 63' 6.4 - 1.5 x 10
JI!.Mean of duplicata determinations on one set of serial dilutions

- 46-
urease activity. These resulte support the conclusion that the high
urease activity of rumen fluid from the urea-fed sheep was due to
the presence of large numbers of bacteria of relatively few types.
C. Attempts at the Isolation and Characterization of Urease-producing
Bacteria from Rumen Fluid.
Atte.m.pts were made to isolate in pure culture organisme
representing the predominating urease-producing flora in tœ rumen
fluid of the urea-fed animal, sheep 2.
a) Isolation of bacteria from cultures in buffered reinforced
clostridial medium
First attempts at isolations of ureolytic rumen bacteria
were made using the cultures in buffered :reinforced clostridial broth
employed for the enumeration of these organisme. The cultures were
streaked on buffe red reinforced clostridial agar. Of a total of Z7
colonies picked at random fran tbese plates, 13 consisted of organisme
which produced an alkaline reaction on urea agar within 12 hr. When
t.hese isol.at.es were transf'erred to tryptone gl.ucose 0.2% yeast e.xtract
brotht two types of growth developed. In tœ case of one, the bacter
ia, which were found to be short, Gram-negative rods, produced a
heavy mucoid pellicle and a light turbidity. In tm other, a unifozm
be avy turbidity was produced by smal1, Gram-negative cocci which re
sembled in morphology and arrangement the organisme sean in the BRCB
cultures. One organism of each type was selected at randan,

- 47-
characterized ani identitied. 'l'hase organisms were re.ferred to
respectively as isolates 1 and 2.
i) Isolate 1
This organiam, wt4ch was isolated from a rumen .fluid dilution
of lJ109 , was identitied as Pseudom.onas aeruginosa on the basis o.f the
.follow:i.ng characters.
Cella rod-shaped, 0.8 by 1.5 to 3.0 microns, arranged singly and in
pairs. Motile at 37°C, with one polar .flagellum, non-motile at
42°0. Encapsulated. Non-spore-forming. Gram-negati"Ve.
Agar colonies: circular, ill-defined edge, .flat, smooth, translucent,
green water-soluble pigmentation.
Agar alope: modera.te growth, spreading, glistening, butyrous, agar
greened.
Nutrient broth: pellicle, strong even turbidity, abundant .flocculent
sediment, green especially at 20°0.
Gelatin stab: rapid hydrolysis.
Li'bnus milk: alkaline curd, rapid peptonization, litmua reduced.
Indole not produced.
Nitrate reduced to nitrite and nitrogen.
Starch not hydrolyzed.
Acetoin not produced.
Methyl red negative.
Hydrogen sulphide not produced.
Tributyrin hydrolyzed.

- 48-
No fermentation of arabinose , rhamnose , xylose , glucose , fructose,
galactose, lactose, mannose, sucrose, maltose, trehalose , cello
biose, saccharose, raffinose, melezitose, glycerol, adonitol,
mannitol, sorbitol, dulcitol or salicin.
No hydrolysis of aesculin, inulin or cellulose.
Catalase produced.
Urease produced.
Citrate utilized as sole source of carbon.
Blood haemolyzed.
Aerobic.
The organisn showed no growth within 72 hr. when inoculated
on to tryptone glucose 0.2% yeast ext.ract agar in a spray dish anaerobie
culture, neither did it grow in a broth culture of tha same medium which
was made anaerobie by tbe addition of a paraffin oil seal before auto
claving. It wa.s therefore concluded to be a strict aerobe. Since it
seemed unlikely that such an organian could maintain a population of
the order of 109 cells per ml. of rumen fluid undar the conditions of
low redox potential ex:tsting in the rumen, it was thought probable
that the organism. wa.s a contaminant which bas been introduced into tbe
primary culture. Support for this conclusion was provided by the
failure of an attempt, made after a lapse of eight weeks, to reisolate
Ps. aeruginosa from the rt1D.en contents of sheep 2, using tœ sam.e
procedure as bef ore. The isola te the re fore received no furtmr
attention.

- 49-
ii) Isolate 2
This organism., which was isolated from a rumen f'luid
dilution of 1/108 , was identifiad as a coagulase-negative member of'
the ~nus Staphylococcus on the basis of the following characters.
Cella spherical, 1.0 to 1.6 microns in diameter, arranged singly,
in pairs, in chaine of up to 6 ce ils and in irre gular clumps.
Non-motile. Non-iodophilic. Gram-positive, becoming negative.
Agar colonies: circular, raised, en tire , smooth, opaque , white
becoming golden.
Agar slope: abundant, filifonn, glistening, butyrous, white growth.
Nutrient broth: strong even turbidity, abundant granular sediment.
Potato: weak golden pigmentation.
Litmus mil.k: slow acid curd, litmus reduced.
Indole not produoed.
Nitrate reduced to nitrite.
Starch not hydrolyzed.
Acetoin not produced.
Methyl red positive.
Hydrogen sulphide not produced.
Tributyrin hydrolyzed.
Acid from glucose, fructose, galactose, lactose, maltose, mannose,
sucrose, glycerol and mannitol. No fermentation of arabinose,
rhamnose, xylose, trehalose, cellobiose, raffinose, melezitose,
adonitol, sorbitol, dulcitol or salicin.

-50-
No hydrolysis of aesculin, inulin or cellulose.
Catal.ase produced.
Urease produced.
Citrate not used as sole source of carbon.
Anunonium. sulphate not used as sole source of nitrogen.
Methylene blue reduced.
Coagulase not produced.
Serum not dige sted.
Blood not haemolyzad.
Facultatively anaerobie.
Tha organism essent:ially satisfied the oonditions suggested
by Gall and Huhtanen (1951) for consideration as a true rumen organism.
It was, for exa.mple, facultati vely anaerobie, it occurred in rumen
fluid in num.bers exceeding 106 per ml. and brought about reactions,
such as the fermentation of soluble carbohydrates, known to occur in
the rumen. The organism. was tberefore considered to be a normal
canponent of the rumen microflora.
Attempts were made to reisolate it from the rumen contents
of sheep 2, eight months a:rter the original isolation. For this
purpose, a prima.ry enricbnent of the staphylococci in a sa.mple of
rumen fluid was made in a brain heart infusion - NaCl medium, and
!ive rumen staphylococci were subsequently isolated. Although none
was identical to isolate 2, all of them were very similar to it and
generally differed only with respect to the range of soluble

- 51 -
carbohydrates which were fermented. This failure to reisolate the
organism, however, was not considered to provide evidence against
tm conclusion that isolate 2 was a true rumen organism. Thus it
was further investigated to assess the possible contribution of the
organism to rumen urease activity.
iii) Urease activi ty of iaolate 2
Isolate 2 was grown on tryptone glucose 0.2% yeast e.x.tract
agar and the cells suspended in ma.leate buffer. The suspension wa.s
shown by a standard plate count to contain 1. 9 x 1011 cella per ml. ,
and its urease activity index found to be 19. Thus, the urease
activity of the organism was very law. The activity produced by an
organism when growing on an artificial medium, however, may not give
a valid indication of the activity it would produce when grown und.er
the environmental conditions prevailing in the rumen (Gibbons and
Doetsch, 1959).
In the following section a medium wbich provided enricbnent
cultures of rumen ureolytic bacteria is described. It consisted of
so~ rumen supernatant liquor supplemented with glucose, phytone and
urea (medium 17, Table V). In view of the hii?P urease activity
expressed by .mi:œd cultures of rumen ureolytic bacteria when grown
in this medium, it was thought probable that it .might support enhanced
urease production by isolate 2. Batches of the medium -were therefore
inoculated with the organism, and after incubation the cella were
harvested and suspended in maleate buffer. The suspension contaiœd

- 52 -
approx:imately 2 x lolO cella per ml. When its urease activity was
determined manometrically, however, it was found to have no œasurable
activity whatsoever.
It was therefore concluded from these experimenta that the
Staphylococcus sp. isolated from the rumen contents of sheep 2,
aJ.though weakly ureolytic when grown on artificial media, did not make
a significant contribution to rumen urease activity ~ !!!2·
b) Isolation of bacteria from cultures in a suppleœnted rumen
supernatant liquor medium.
Owing to the lack of auccess experienced in the attempted
isolation of rumen bacteria with a high specifie urease activity by . the method so far described, a second approach to the problem was
made. This involved the development of a medium which would provide
enrichment cultures of rumen ureolytic bacteria, and pennit subsequent
isolation of organisme fran these cultures.
For this purpose, various supplements were added to a basal
medium eontaining 90% rumen supernatant l.iquor, and the rel.ative
abilities of twenty-four such media to support growth and urease
production by rumen bacteria in mixed culture 11ere determined. The
re sul ts are shown in Table V.
Growth developed to varying extents in ail media except
medium 1, which contained no supplements, but the greatest development
of urease activity occurred in those media which were supplemented
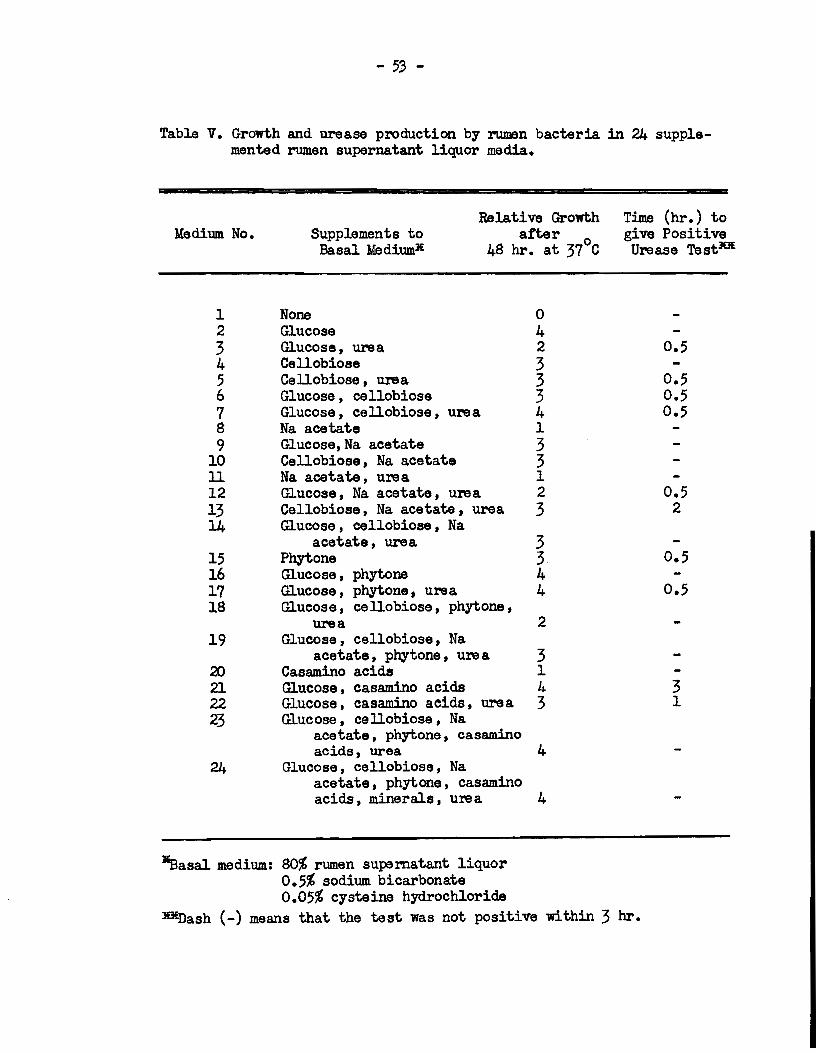
-53 -
Table V. Growth and urease production by rumen bacteria in 24 supplemented rumen supernatant liquor media.
Medium No.
1 2 3 4 5 6 7 8 9
10 ll 12 13 14
15 16 17 18
19
20 21 22 23
24
Supplements to Basal ll.ediumlE
Relative Growth a:tter
0 48 hr. at 37 C
None 0 Glucose 4 Glucose, ure a 2 Cellobiose 3 Cellobiose , ure a 3 Glucose, cellobiose 3 Glucose, cellobiose, ure a 4 Na acetate 1 Glucose,Na acetate 3 Cellobiose, Na acetate 3 Na acetate, ure a 1 Glucose, Na acetate, ure a 2 Cellobiose, Na acetate , urea 3 Glucose , cellobiose , Na
acetate, ure a 3 Phytone 3 Glucose, phytone 4 Glucose, phytone, ure a 4 Glucose, cellobiose, phytone,
ure a 2 Glucose , cellobiose, Na
acetate, phytone, ure a 3 Casam.ino acids 1 Glucose , casamino acids 4 Glucose, casamino acids, ure a 3 Glucose, cellobiose, Na
acetate , phytone, casamino acids, urea 4
Glucose, cellobiose, Na acetate, phytone, casamino acids, .m.inerals, ure a 4
X.Sasal medium: 80% rumen supamatant liquor 0. 5% sodium bicarbonate o.o;% cysteine hydrochloride
Time (hr.) to give Positive Urease TestlŒ
o.; o.; o.; o.;
o.; 2
o.; o.;
3 1
lBEDash (-) means that the test was not positive within 3 hr.

-54-
with urea and either glucose or cellobiose. The presence of sodium
acetate, casamino acids or miœrals did not appear to be related to
enzyme production.
Two media, 5 and 17, 11ere se le cted from among those which
supported production of urease to the greatest extent and these were
then compared for their ability in this respect in another experiment.
The media were prepared and inoculated in lOO ml. volumes. After
incubation the cella from the two cultures ware suspendsd in buffer
and the volumes of th:l suspensions were adjusted until both had the
same optical density. It was then assumed that they centaine d approx
imately equal numbers of cella per ml. The urease activities of the
suspensions were measured manometrically, and the activity of the
suspension of cella from medium 17 (qN~ • 1403) was fotmd to be some
lfha.t greater than that from medium 5 (qlffi_3 = 1316). Medium 17 was
therefore chosen for further study, and tested for its abUity to
provids enrichment cultures of rumen ureolytic bacteria.
Figure 1 shows that successive transferring of rumen
organisms through medium 17 resulted in a considerable increase in
urease activity per unit concentration of cella over that of the
rumen fiuid from which the first culture was inoculated. The
enhanced urease activity of the cella was only maintained to the
second transf'er, but it was concluded that this œdium was capable
of providing the enrichment cultures required.
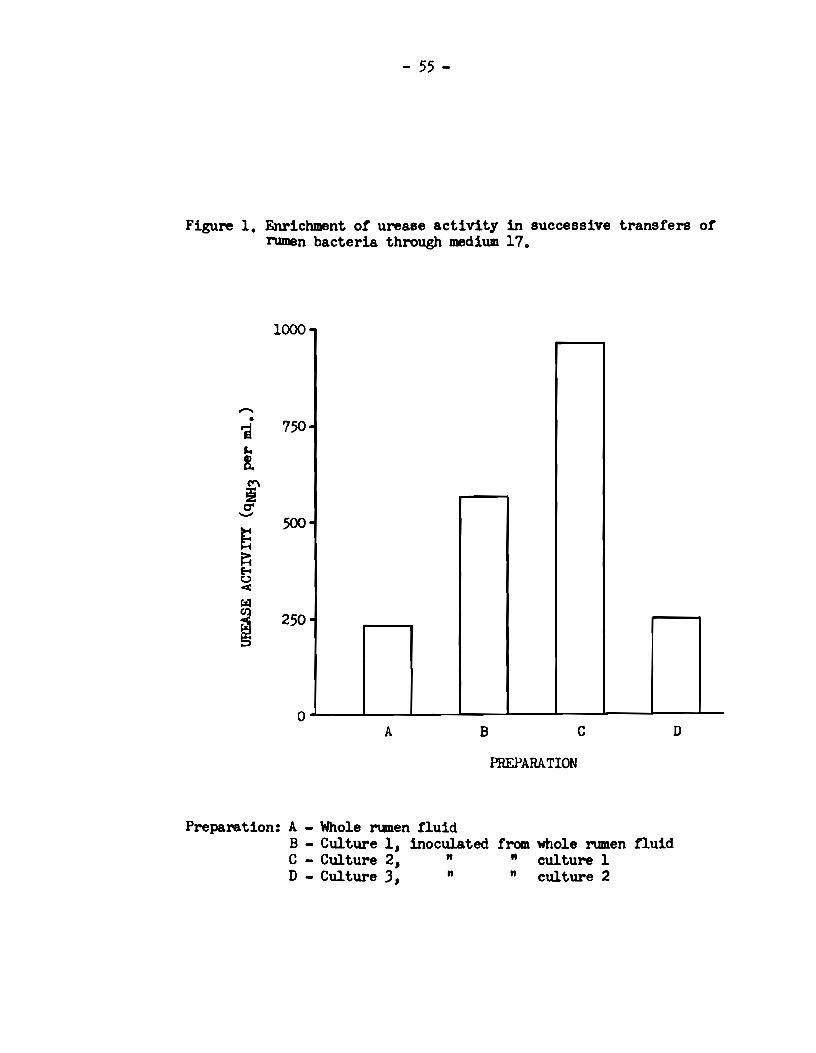
- 55 -
Figure 1. Enrichment or urease activity in successive transfera ot rumen bacteria through medium 17o
1000·
-• 750 i . ~
!. ("'\
~ -E
500 . i=i E-1 tJ <
1 250 .
A B c D
PREPARATION
Preparation: A - Who1e rumen nuid B - Culture 1, inoculated traa who1e rumen fiuid C - Culture 2, " " culture 1 D - Culture 3, " " culture 2

- 56 -
An attem.pt was therefore made to isolate rumen ureolytic
bacteria fran an enriched culture. For this purpose, a series of
enrichment cultures of rt'lœn bacteria in md.ium. 17 was prepared and,
by successive transferring, the urease activity per unit concentration
of cells was increased to a level 3t times greater than that of the
whole rumen fiuid fran which the inoculum. was obtained. Microscopie
examination of the final culture in the series showed it to contain
several morphologi.cal types of ba.cteria. An aliquot of the culture
was serially diluted and plated, and alter anaerobie incubation 57
colonies were picked from the plates. The organism.s isolated 'Were
che cked for purity and for the ability to produce urease when growing
in pure culture in medium 17. Altbough the medium was prepared and
incubated in the same way as for the enrichment cultures, however,
not one of the isolated organisms produced a detectable amount of
urease.

DISCUSSION
Tœ generally high leval of rumen urease activity round
in tœ present stud.y is in agreement wi.th the findings of previous
work (Pearson and Smith, 1943a; Huhtanen and Gall, 1955). The
variations in urease activity from time to time observed with rumen
fluid from sheep 2 were probably due to the affects of several factors.
For exarnple, changes in tœ rate of formation of the enzyme by micro
bial synthe sis relative to its rate of removal from the rumen in the
normal flow of rumen oontents, and the dilution of rumen contents
with water, would both be expected to influence tœ observed urease
activity of the rumen fluid. Despite fluctuations in the leval of
rumen urease activity, however, there seems to be some active urease
present in the rumen at ali times. This is of two-fold significance
with respect to the nitrogen metabolism of the animal. First, it
resulta in the unique ability of ruminants to utilize dietary urea
for the indirect synthesis of animal protein. Second, since urease
converts urea entering the rtmen from the bloodstream to am.monia,
and since a proportion of this ammonia may then be incorporated into
microbial protein, the presence of urease in the rumen enables the
retention of some nitrogen which would otherwise be lost to the
animal by renal excretion.
The results of this stud.y have confirmed the findings of
Gibbons and McCarthy (1957) with respect to the production of urease
by the rumen bacteria, and have extended these f'indings by providing

- 58 -
infom.ation concerning the proportion of bacteria which produced
urease in the rllllen of a urea-fed sheep. The total viable counts
ot rumen bacteria obtained were geœrally lower than the counts
reported by many previous workers, which have indicated a mean
bacterial population of 109 - 1()10 organisms per ml. of l"W'Den fluid
(Annison and lewis, 1959). Severa! factors may have been responsible
for the low counts obtained in the present case. First, it has been
shawn, for example, that severa! species ot rumen bacteria require
C02 for growth (Johns, 1951; Wright, 1960). The metabolism or the
bacteria present in the buffered reinforced clostridial broth cult
ures would probably result in the production of 002 during incubation,
but since the medium contained no added 002 tœ possibility exista
that the · 002 tension in the medium was not adequate for the growth of
SCllle types of rumen bacteria. Second, the technique used to prepare
the cultures was probably not sufficiently controlled to provide
completely anaerobie conditions in tœ inoculated medium. The
presence of oxygen in the cultures may therefore have exarted a
selective action upon the facultative anaerobes present in the
original inoculum. by inhibiting the growth of obligate anaerobes.
Support for this possibility is provided by the resulta of micro
scopie examination of the cultures. These showed that the predom
inating type of organism occurring in these cultures was similar in
cell morphology ani arrangement to tm organism subsequently isolated
and round to be a facultatively anaerobie Staphzlococcus sp.

- 59 -
Notwithstanding these limitations, it may be concluded
from the studies on enumeration of l!"WWIen ureolytic organism.s that
the rumen fiuid from sheep 2 contained a high proportion of urease
producing bacteria. The resulta of this study apply only to this
sheep 2, however, and it might be expected that in an animal main
tained under a different feeding regime the num.bers and proportion
of urease-producing rumen bacteria would be different from those
f'ound with the urea-fed animal. The high level of urease activity
which is generally characteristic of rumen contents suggests,
nevertheless, that large numbers of urease-producing bacteria are
always present in the rumen contents of nonnal anima.ls.
Despite the deomonstration of' a large number of ureolytic
bacteria in the rumen nuid of sheep 2, it was possible to isolate
only one organism. which showed urease activity i::a vitro. This was
a facultatively anaerobie organism. which could not be considered to
produce significant amounts of urease in the rumen. Several other
workers (lla.eKay and Oxford, 1954; Huhtanen and Gall, 1955; Gibbons
and MeCarthy, 1957) have similarly concluded tbat facultative anaerobes,
a.lthough producing urease when grown on la.boratory media, make an
insignificant contribution to urease activity i::a vivo.
The attempts made to isola.te pure cultures of a.nerobic
urease-producing organisms having a high specifie activi ty were
unsuccessful. This is again in accord with previous attempts by
other workers (lla.cKay and Oxford, 1954; Blackbum and Hobson, 1962).

-60-
The only anaerobie urease-producing rumen species so far reported
to have been isolated was a Gram-positive rod resembling Lactobacillus
bifidus, which was obtained from the bovme rumen by Gibbons and
Doetsch (1959). Although this organisrn produeed a relatively high
urease activity on suitable laboratory media, it was not eonsidered
to be representative of the predominating ureolytic nora of the
rumen.
Anaerobie, ureolytic rumen ba ete ria produeing significant
amounts of urease, therefore, still await isolation. The f'ailures
which have been experienced in attempts to isolate such organisme
are probably attributable to the inability of artifieial cultures
to provid.e conditions whieh adequately sim.ulate those prevailing in
the rumen. In the opinion of' Hobson (1962), the problem is one of
devising primary isolation media which will satisf'y the nutritional
requirements of the organisme, but it is also œcessary, from the
point of view of recognising the organisme when isolated, that media
used for growing them be capable of supporting the production of
urease in quantities comparable to those they produce in the rumen.
The developnent in the present study of a medium which would provid.e
enric:tment cultures of rumen ureolytic bacteria may have made a
contribution to the ultimate solution of' these problems.

SUMMARY
A study was made of the urease activity of strained rumen
fluid from a sheep fed a diet in which urea satisfied a proportion
of the nitrogen requirement of the animal.
The urease activity of the rumen fluid Yaried from time to
time within a range which represented a variation in the amount of
urea hydrol.yzable to ammonia frcm 25 - 310 mg. par 100 ml. of rumen
f1uid per hr.
Ueasurement of the urease activitiea of the compoœnt
fractions of rumen f1uid showed that the capacity to hydrolyze urea
was associated entirely with the rumen bacteria, a finding which
confirma the resulta of previous workers with respect to the source
of rumen urease.
When the total microbial population of rumen f1uid was
divided into arbitrary subfractions by differentia! centrifugation,
it was found that the bacteria responsible for 65% of rumen urease
activity were mainly the group of larger organisms, comprised of
severa1 morphologiea1 types, which sed:imented at a relative cent
rifuga! force of 1,200 x G.
Approximately 35% of the total population of bacteria in
the rumen fluid of the urea-fed shaep had the capacity to produce
urease when cul tured in a buffe red reinforced cloatridial medium.

- 62 -
Attempts to isolate representative anaerobie urease-pro
ducing bacteria from cultures of rumen microrganisms in this medium,
however, were unsuccess.ful. One facultatively anaerobie Staphylococcus
sp. was isolated, but since it had only a low urease aetivity, even
wben grown in medium in which rumen microorganisns grew and produeed
large quantities of urease, it was coneluded tha. t this organism was
unlikely to be of significanee in rumen urease production.
A medium which would provide enrichment cultures of rumen
ureolytic baeteria was devised. It contained 80% rumen supernatant
liquor supplemented with glucose, phytone and urea. Two successive
trans fers of rumen organisns through this medium provided a cul ture
in which the urease activity per unit concentration of cells was
approximataly four times that of the rumen fluid from which the first
culture was inoculated.
An attem.pt was made to isolate fran such an enriched
culture strictly anaerobie bacteria rapresenting the predaminating
ureolytic species of the rumen, but this was unsuccessful.
It was concluded that although rumen fiuid contains large
numbers of urease-producing bacteria of a relatively limited range
of types, insu.fficient is known of the requiremen:ts of these organisms
for growth to allow of the ir isolation in pure culture.


Part II
Factors Affecting the Activity of Rumen Urease

INTRODUCTION
Early observations of urease activity were made during
studias of the aumoniacal fer:rœntation of urea-containing materia.ls
such as urine; auch fer.mentation is brougbt about by urease of
bacterial origin. Recognition of the jaok-bea.n, Canavalla ensifonnis,
as a partioularly rich source of the enzyme urease (Mateer and Marshall,
1916), however, and the deve1opnent by Sumner (1926) of a simple
method for its crystallization, subsequently caused attention to be
directed very 1argely towards this urease. Since that time the
properties of crystalline jack-bean urease have been widely studied,
and, apart from demonstrations of its presence in many mcroorganisms,
interest in urease from other sources bas not been great. Indeed, it
was not until 1954 that a serious attempt was made to prepare bacteria1
urease in a purified for.m.
In 1943, Pearson and Smith showed that rumen fiuid invariab-
1y has a highly deve1oped capa city to hydrolyze ure a, and this is
now known to be attributa.bl.e to urea.se of ba.cterial origin. It
would therefore be of considerable interest to compare the proJ:erties
of rumen urease with those of urease from other sources, especially
since the accum.ula.ted evidence suggests th.a.t urease-producing rumen
bacteria are obligate1y anaerobie whereas virtually a11 non-rumen
bacteria. so far reported to produce urease be1ong to aerobic or
facultative1y anaerobie genera. Only cursory attempts have been

- 66 -
made to study the properties of rumen urease, ho"Wever, beeause the
organisms implieated in its production have as yet proved impossible
to isola te in pure cult ure. Sorne work has been done using the
mixed rumen flora as a source of enzyme, but at present our know
ledge of the meehanisms involved in rumen ureolysis is very limited.
In the present investigation a study was made of soma
factors affeeting the aetivity o:f rumen urease in vitro. The mixed
rumen mierobial :fraction :from fresh rumen contents was used as a
source o:f urease sinee the attempts deseribed in Part I to isolate
anaerobie urease-produeing rumen baeteria proved unsueeess:ful.

LITERATURB REVIEW
A. Factors Aff'ecting the Activity of Jaek-bean Urease.
The kinetics of urea hydrolysis by jack-bean urease have
been widely studied with respect to the influence upon its activity
of such factors as temperature, pH and relative concentrations of
substrate and enzyme. It is clear that the eff'ects of these factors
are complex and inter-related. In addition, tœ eff'ects of se veral
agents which inhibit, prote ct or stimulate urease have received
considerable attention (Varner, 196o).
a) Urease stimulating agents
According to Sumner, there is no coenzyme for urease
(Sumner and Kirk, 1932) and no stimulation of the enzyme unless there
bas previously been some degree of reversible inhibition (Sumner, 1951).
Agents with an apparent stimulating ef'fect are believed to act in
either of two ways. First, substances such as proteins, am.ino acids
and gum arabie bind heavy metals pre sent as contaminants in the buffer
system and thereby protect the active sulpydryl groups of the enzyme
from the inhibiting effects of the metal ions (Pinter, Taskovska and
Karas, 1954). Second, reducing agents such as H~ and glutathione,
whieh reactivate the enzyme after mild oxidation (Sumner and Myrback,
1930; Argenziano and Giani, 1946) apparently reduce oxidized and
therefore inactive sulphydryl groups. The stimulation of soybean
urease by the oxidation-red uction vitamins nicotinamide and

- 68-
ribofla.vin (Jacobsohn and Cruz, 1945) is another example of this
type of activation.
Sane substances, however, have been :round to e:xsrt a
stimulating affect on plant urease which camot be explained in
thes.e tenns. Wall and Laidler (1953), for example, have shown that
glycine, DL-alanine and L-tyrosine speci:tically stimula.te urease.
Their resulta suggested that two separate affects were operative.
One was a general, non-competitive stimulation which was independant
of substrate concentration, and the other a reduction in substrate
inhibition {Laidler and Hoare, 1949) which predominated at urea
concentrations above O.l5K.
Shibata (1958) showed a stimulating affect of alkaline
earth metals on urease activity, in the order Mg++_?Ca+"/Ba++_.?Sr++.
The original report of this work has not been seen, however, and the
source of urease used by the author is not known.
Talœda (1960) observed that low concentrations of the
antibiotice viomycin and penicillin stimulated urease but that very
high concentrations inhibited it. He sugge.sted that the effects
observed involved combinat ion between enzyme and antibiotic, and that
when the antibiotic accumulated on the urease molecule in excessive
amounts the approach of the substrate molecule was prevented and
inhibition therefore occur.red.

- 69 -
b) Ure ase inhibi ting agents
A num.ber of agents are known to inhibit urease specifically
and the inbibiting affects of heavy metals upon urease activity have
been lmown for many years (e.g., Jacoby, 1933). More recently,
evidence has been obtained which suggests the .mechanism of inhibition.
Shaw (1954) for exam.ple, has shown that the sequence of to:x:icity of
specifie metal ions for urease is: Ag+I"'.Hg••?Cu++:::>cd++7Co++7Ni•• 7
}ln•+. These metals appear to inaotivate the enzyme by com.bining with
it at the active site and those ions whioh have the greatest affinity
for negatively charged sulphur are the most to:x:ic. Shaw and Raval
(1961) have suggested the reaction to be analagous to the formation
of metal sulphides, and indeed the to:x:ioity sequence was found to
correlate perfectly with the relative insolubilities of the metal
sulphides and with the relative stabilities of the metal complexes
so formed (Shaw, 1961).
Na• and K+ have an inhibiting action on urease which is
dependent on pH. Thus, Fasman and Niema.nn (1951) have shown a
specifie inhibii;ion of urease by these ions at pH 7.0, whereas
Kistiakowsky and Shaw (1953) round that at pH 8.9 urease activity
was influenced by the ionie strength of the buffer solution rather
than by the type of ion (Na+ or K+) present.
Quastel (1933) showed an inhibition of urease by quinone
which could be prevented by ~ and cysteine, and Mystkowsld (1928)
shœed a pœerful inhibition of the enzyme by fluoride ions.

- 70-
Talœda. (1960) .round that urease activity wa.s reduced in
the presence of the antibiotics colimycin, chloromycetin, terramycin
and dihydrostreptomycin, and explained these affects in te:nns of
competition between antibiotic and substrate for attachment at the
active site of the enzyme.
B. Prwrties of Urease Produced by Non-rumen Bacteria.
Although some 200 species of non-rumen bacteria have been
reported to pr<Xluce urease, etudies of the properties of the enzl7JII.e
have been surprisingly llmited both in num.ber and in spe cies of
organism. used. Whole cella of !J:. vulgaris were used by Sizer (1941)
as a source of enzyme; La.rson and Kallio (1954) used partially
purified Bacillus pasteurii urease, and Lister (1956) used cell-free
extracts of Corynebacterium renale. Com.parisons between the resulta
of tœse workers are made difficult by the differences in conditions
of temperature, pH and buffer composition used in the measurement of
kinetic parameters, but apparently an essential similarity exista
between plant and bacterial ure ases, the kinetics of enzymes from
these different sources differing only in minor respects.
From e:xperiments on its susceptibility to sulphydryl group
inhibitors, Yall and Green (1952) suggested that the urease produced
by Micrococcus pyogenes var. aureus was of two types, depending upon
the cultural conditions under which the organiem was grown. Urease
produced in tœ presence of urea was inhibited by furacin, p-mereuri
benzoate and trivalent arsenical compounds, but Tfhen urea was omitted

- 71-
fran the growth medium the enzyme produced was not inhibited either
by sulphydry1 inhibitors or by furacin. The authors postulated a
shift in the fom of enzyme from. su1phydry1 to non-sulphyd.ry1 to
account for this difference in affinity of the enzyme for su1phydry1
inhibitors.
C. Properties of Rumen Urease.
The report of Pearson and Smith (1943&) representa the
only published investigation of the propertjs s of rumen urease.
These workers used strained rumen fluid fran a urea-fed steer as a
source of .m.ixed rumen urease and were the re fore dealing wi th urease
contained intracellular1y within intact rumen bacteria. They
determined the effects of changes in temperature, pH and substrate
concentration, and of som.e inhibiting agents, upon the ability of
strained rumen f1uid to hydrolyse urea.
The optimum. temperature for activity of the enzyme was
49°C, sorne 10°C above the nonnal body temperature of the ruminant
animal. The optimum. pH lay between 7 and 9. Pearson and Smith
determined no kinetic parameters of rumen urease, such as activation
energy or Michaelis constant, to characterize the enzyme, but con
c1uded from the resulta of their experimenta that rumen urease was
very similar in constitution and properties to jack-bean urease.

JlATERIALS AND METHOUS
A. Collection and Preparation of Rumen F1uid.
The experimental anima1s used in this study and tl»
methods of collection and preparation of rumen contents from them
•re de scribed in Part I.
B. Preparation of Mixed Rumen Urease.
a) Who1e ce11 preparations of rumen microorganism.s
The m.i:md rumen microfiora was removed from 25 ml. aliquots
of rumen f1uid by centrifuging them at 13,200 x Gin a refrigerated
centrifuge at 10°C and decanting the rumen supernatant liquor. The
microbia1 fraction was then washed by suspending it in 0.0214-phosphate
or maleate buf'fer, at pH 6.8, centrifuging the suspension, and again
decanting the supernatant 1iquid. This procedure was routine1y
repeated twice and the microbia1 fraction was finally suspended in
the appropriate buffer and the suspension di1uted to 25 ml. with
buf'fer.
b) Acetone-dried powders of rumen microorganisme
Protozoa and larger food partic1es were removed from 600 -
000 ml. of rumen f1uid by centrifugation at 75 x G for 2 min. in a
refrigerated centrifUge at 10°C. The bacteria-rich supernatant
liquid was decanted and centrifuged at 13,200 x G for 10 min. at
10°C and tœ cella washed three times, as described above, in

- 73-
0.021l-maleate buffer. A.tter the final washing the cella were suspend
ad in about 100 ml. of cold distilled water and 10 volumes of acetone
0 at 10 C were immediately added. The suspension was well stirred and
then filtered with suction using Whatma.n no. 2 filter paper. The cell
precipitate was washed with cold acetone until the washings 111ere no
lon~r green in colour. Air was drawn through the precipitated cells
on the filter to remove residual acetone. The precipitate was allowed
to stand for 24 hr. over CaC12 in a dessicator at 10°0 and then ground
in a mortar to a fine powder. The powders, if .round to possess app
reciable urease activity, 111ere stored in sealad brown botties in a
dessicator at 10°0.
c) Cell-free extracts of rumen microorganisms
The mixed flora fran lOO ml. of rumen fluid was washed
three times in buffer as described above and suspended in 10 - 15 ml.
of buffer. The cells were disrupted by sonic oscillation usjng an
MSB-Kulla.rd ultra-sonic disintegrator. The cell suspension in a
beaker was placed in a larger beaker containing crushed ice and
water. The cells were aubjected to sonic oscillation for four suce-
essi ve 10 min. periods. At tb3 end of each period 5 min. were allowed
to permit the suspension to cool. After microscopie exa.mination,
which revealed a high degree of cell rupture, the suspension was
centrifuged at 105,400 x G in a Spinco ultracentri.tuge for 30 min.
The clear supematant liquid was decanted, assayed rapidly to deter-
mine its approx.ima.te urease activi.ty and then, if necessary, diluted
with cold buffer for quantitative studies.

- 74 -
C. Preparation of Jack-bean Urease.
Twenty-five mg. of jack-bean urease (Harleco) were suspended
in 25 ml. of 0.02M-maleate bu:tfer at 10°C with shaking. The suspen
sion was filtered in the refrigerator and used without delay to avoid
decomposition. In experments where the activity of jack-bean urease
was to be determined, 0.5 ml. of the filtrate (equivalent to 0.5 mg.
of powder), diluted to 3.0 ml. with buffer, was used in the main
compartment of each Warburg vessel.
D. Preparation of Ashed Rumen Supematant Liquor.
A 25 ml. volume of f'resh rumen supernatant liquor wa.s
evaporated to 2 - 3 ml. and transferred to a platinum crucible. It
was then charred and ashed for 12 hr. in a muffle-furnace at 470°C.
The ash was taken up in the water, f'iltered, evaporated to dryness,
and tm residue dissolved in 25 ml. of 0.031-maleate buff'er.
E. Measurement of Urease Activity.
a.) Routine method
The activities of tm va.rious preparations used in this
investigation were measured mananetrically as described in Part I.
b) Urease activity in tm presence of added cations
When the effects of individual cations upon rumen urease
activity were to be m.easured, urease preparations were suspended in
0.02M~ea.te buffer containing an appropriate concentration of ion.

- 75 -
Ions were generally used in the form of the chlorida salt of the
corre sponding metal.
Where measuremants of the tirne required for initiation of
an affect of tm ions on urease activity were to be made, hovever,
the procedure used was as follows. Double side-arm Warburg vessels
were used and 2. 7 ml. of m-ease preparation in m.aleate buffer were
added to the main canpartment. To one side-arm was added 0.2 ml.
of 0.3M-urea, and to the other 0.3 ml. of a solution of the test ion
in buffer at a concentration of 128 mll. In control vessels tœ ion
solution was replaced by addition of 0.3 ml. of bu.ffer to the main
compartment. A.fter equilibration of the vessels, the substrate
solution was added to the main compartment and two sets of m.anom.eter
readings were taken at 15 min. intervals. The ion solutions were
then added as quickly as possible and a third set of readings
immediately taken. Readings were continued at 15 min. intervals for
a further li - 2 hr.

RESULTS
A. The Identitz of Factors Affecting the Activity of R'llm'3n Urease.
a) Experimenta with intact rumen microorganisns
The microbial fraction from a sample of rumen fluid from
sheep 2 was separated by centrifugation and resuspended in various
media, and the urease activities of the suspensions were determined.
The resulta are presented in Table VI. They show that when the cella
were resuspended in phosphate or maleate buffer the urease activity
of tm suspensions was appreciably lesa than that of the uncentrifuged
rumen fluid. When the cella were resuspended in the supernatant
liquor removed from the cella when the rumen f'luid was centrifuged,
however, tm activity of the re sul ting suspension was grea ter than
that of' the suspensions in buffer. This suggested that there waa a
factor in the rumen supernatant liquor capable of st:imulating tm
urease activity of the microbial cella. To determine whether this
factor was organic or inorganic in nature , a sample of rumen super
natant liquor was ashed and the inorganic residue dis sol vad in
malea.te bu.t'.fer. The resulta in Table VI show that when the rumen
microbial fraction was resuspendsd in this solution the urease
activity of the suspension was even greater tha.n that of an equiv
alent amount of uncentrif'uged rumen fluid. It thua appeared that
rumen supernatant liquor contained an inorganic factor capable of
st:imulating the urease activity of the rumen microbial fraction.

- 77-
Table VI. Ef'fect of various suspending media on the urease activity of rumenmicroorganisms.
Preparation
Uncentrifuged rumen f'luid
Microorganisme in 0.0~-phosphate buffer
lt
"
"
If o.02M-maleate buffer
" RSLi[
" maleate buffer plus RSL ash
:KRumen supernatant liquor
Urease Activity (~ per ml.)
3
1472 1: 23
936 t: s 882 t: 12
lJ.44. l: 4
2D47 t: 45

- 78-
The affect of repeatedly washing the rumen microbial
fraction on the capacity of rumen supernatant liquor to enhance t.œ
urease acti vit y of the washed cella was then tested. The procedure
uaed to obtain suspensions of washed cella was as follows. Aliquote
of rumen fluid were centrif'uged and the rumen supernatant liquor wa.s
decanted. Two portions of the microbial fraction were then resuspend
ed respectively in rumen supernatant liquor and in 0.02M:-maleate buffer
at pH 6.8 for tests of urease activity. The remaining portions of the
microbial fraction wre suspended in buffer and the suspensions were
centrif'uged. The supernatant liquid was again decanted and the cella
were resuspended in buffer. This washing procedure was repeated a
sufficient number of t.imes to provide suspensions of cella for test
ing which bad been washed 3, 6 and 9 times, respectiveiy. Af'ter each
of these numbers of washings the urease activity of the cella was
compared when suspended in rumen supematant liquor and in maleate
buffer. The resulta in Figure 2 show that the urease activity of the
suspensions in buffer was in avery case lower than that of equivalent
amounts of the cella suspended in rumen supematant liquor. There
was, moreover, a progressive decrease in urease activity of the cells
resuspended in buf'fer as the number of washings increased. Up to the
third washing the activity of the cella could be completely restored
to the leval prevailing in unwashed cella by resuspending t:œ was~ed
celle in rumen supernatant liquor. Af'ter the si:x:t.h and ninth washings
the urease activity of the celle could not be so restored, suggesting
that seme irreversible losa of enzyme activity occurred on prolonged
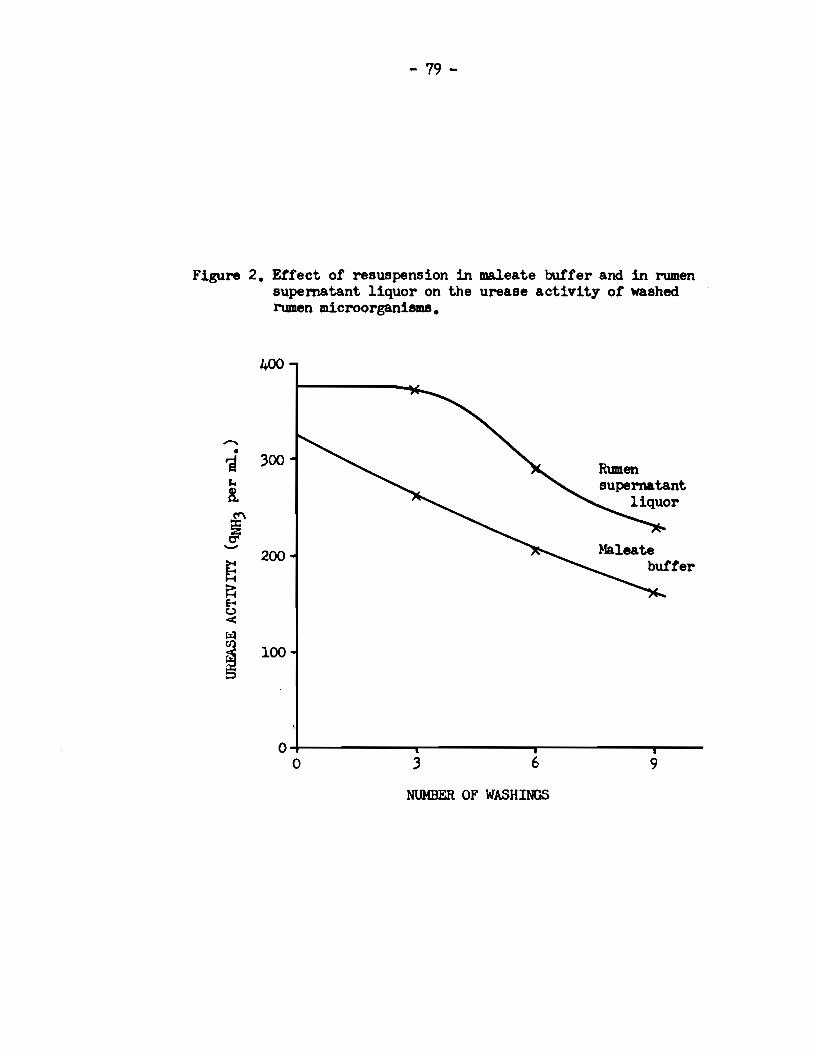
- 79 -
Figure 2. Effect of resuspension in maleate butter and in rumen supernatant liquor on the urease activity of vashed rumen microorganisma.
400
-• 'if 300 Rumen s.. supernatant 8. liquor
f cr - Maleate
~ 200
butter
~ E-t 0 <(
rz1 Cl)
100 ~
01-----------~----------~----------~----0 6 9
NUMBER OF WASHINGS

-80-
washing. On the basis of the se resulta, rumen microorganis:ns l'lB re
routinely prepared for subsequent experimenta described in this
section by washing them three times in .maleate buffer.
In order to identify the inorganic component of rumen
supernatant liquor which was responsible for the stimulation of
urease acti vit y, the capaci ty of combinat ions of related cations to
stimulate the activity of intact cella was determined. The resulta
are presented in Table VII. They show that an appreciable stimulation
of urease was brought about by the group of di valent ions, Ca ... +, Mg"" ... ,
Sr ... + and Ba++. In order to determine the affects on rumen urease of
the individual ions in the various combinations, each was tested over
a range of concentrations. The resulta are presented in Figure 3.
They show that each of the alkaline earth metal ions had a capacity
to sti.mulate the urease activity of the whole cella. Moreover, Jln++
had an even grea ter stimula ting affect than Mg+ ... or Ca....._. The affect
of Zn++, Cu ...... and Fe+"" at all the concentrations tested, although
not sholfll in Figure 3, was to inhibit urease activity completely.
When the concentrations at which the alkaline earth ions
and Jin++ exerted half-.maxi.mum stimulation of urease activity are
eompared, it is sean that in ali cases the concentration was 2 - 8 Jill of ion per ml. of suspension. Since a number of different ions had
a capacity to sti.mulate the mierobial urease, it was necessary to
establish whether or not the stimulation was due to a non-specifie
ionie strength affect. For this purpose washed cella were suspended

- 81 -
Tabla VII. Capacity of cambinations of related cations to stimulate tœ urease activity of washed rumen microorganisns.
Cations Added!lt
None
Group A: Na4-, K4-
Group B: Ca++, Mg++, sr++, Ba++
Group C: Zn++, Cd++, cu++' )(n++
Group D: Fe++, Fe+++-, Co++
lEconcentration of each cation: 4 Ji1! per ml. Cella suspended in 0.0211-maleaté buffer
Urease Activity (cum3 per ml.)
367 :!: 10
353 ± 4
552 :!: 2
0
0
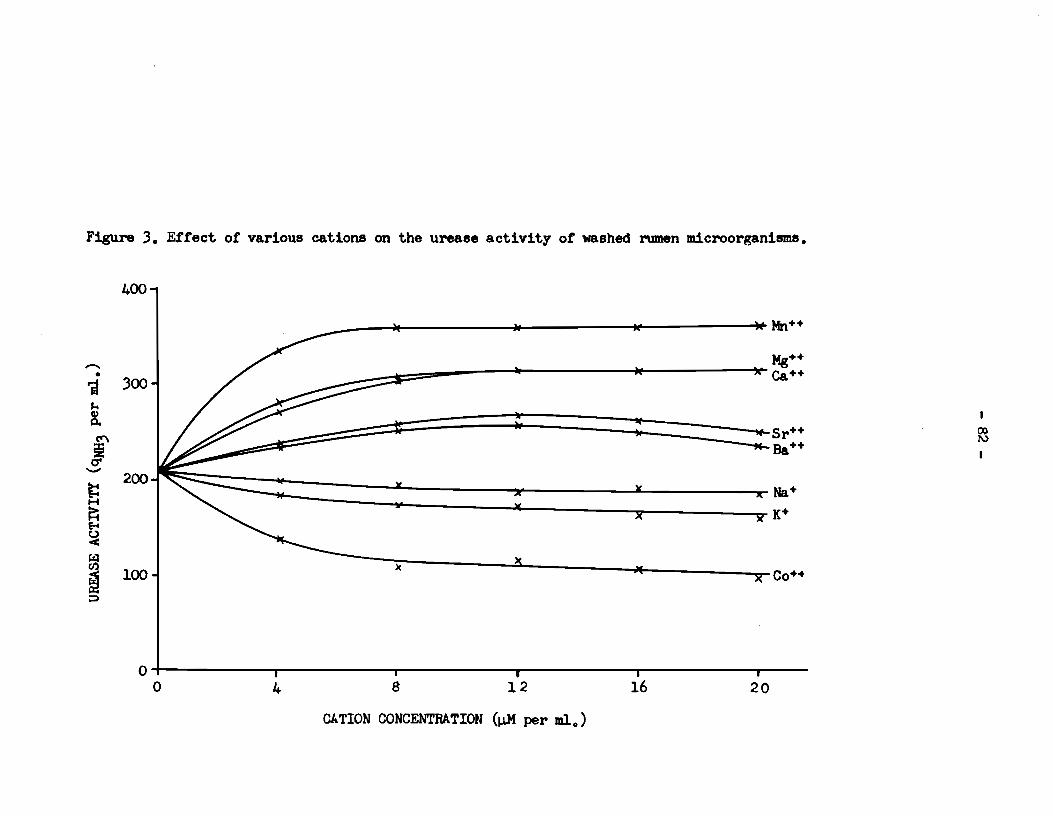
Figure 3. Effect ot various cations on the ureaae activity of washed rumen microorganima.
400
1 Mn++
- 300 1 / Kg++
• ca•• '"d J.. Q) D. ... ~ 1 )j
)l' ::sr++ ~
~ Ba++
1 -E
200
1 ~ 1(1 : :· : Na+
~ o K+ E--4 ~
1 100~ )( Co++
:::>
o~--------~~---------r----------~----------r---------~-----0 4 8 12 16 20
CATION CONCENTRATION {IJ.M per ml.)

- 8.3 -
in buffer solutions adjusted to an ionie strength of 0.0611: (the
ionie strength of the optimum Mg++ concentration) using various salta.
The resulta in Figure 4 show that Na+ and K+ salta had little or no
eapacity to stimulate the urease activity of the washed eell suspensions.
It is thus evident that the stimulation of the urease aetivity of
intact cella by the alkaline earth metal ions and :u:n++ was not due to
an ionie strength effeet.
Of the di valent ions shown to stim.ulate the urease activity
of intact rumen mieroorganisns, Kg++ and Ca++ were more likely than
lin"'"", Sr++ or Ba+• to be tm active ions present in runen supematant
liquor. :u:n++' however, frequently plays a sim.ilar rele to that of
Mg++ and Ca+"' in the activation of enzyme systems (Di.xon and Webb,
19.58). The capaeity of solutions of :Mg++, Ca++ and :u:n++ to replace
rumen supernatant liquor in stim.ulating the urease activity of intact
cella was therefore tested. The eapacity of :Mg++ in this respect was
tested with washed cella from sheep 35 and 37 and steer 2; the
resulta are shown in Figure 5. Cella from. sheep 2 were used to
compare tœ affects of rumen super na tant liquor with Jln++, Mg++ and
Ca++; the resulta are presented in Figure 6. The resulta in Figure
5 clearly show that the addition of Mg++ to a buffer solution raised
the urease activity of washed cella suspended in the solution. In
the presence of tm concentration of Yg++ giving m.a.x:imum stimulation
of the enzyme, the aetivity was raised to the leval obtained with a
suspension of cella in rumen superna.tant liquor. Although the con
centrations of Mg++ in buffer need.ed to achieve this leval of ac ti vity
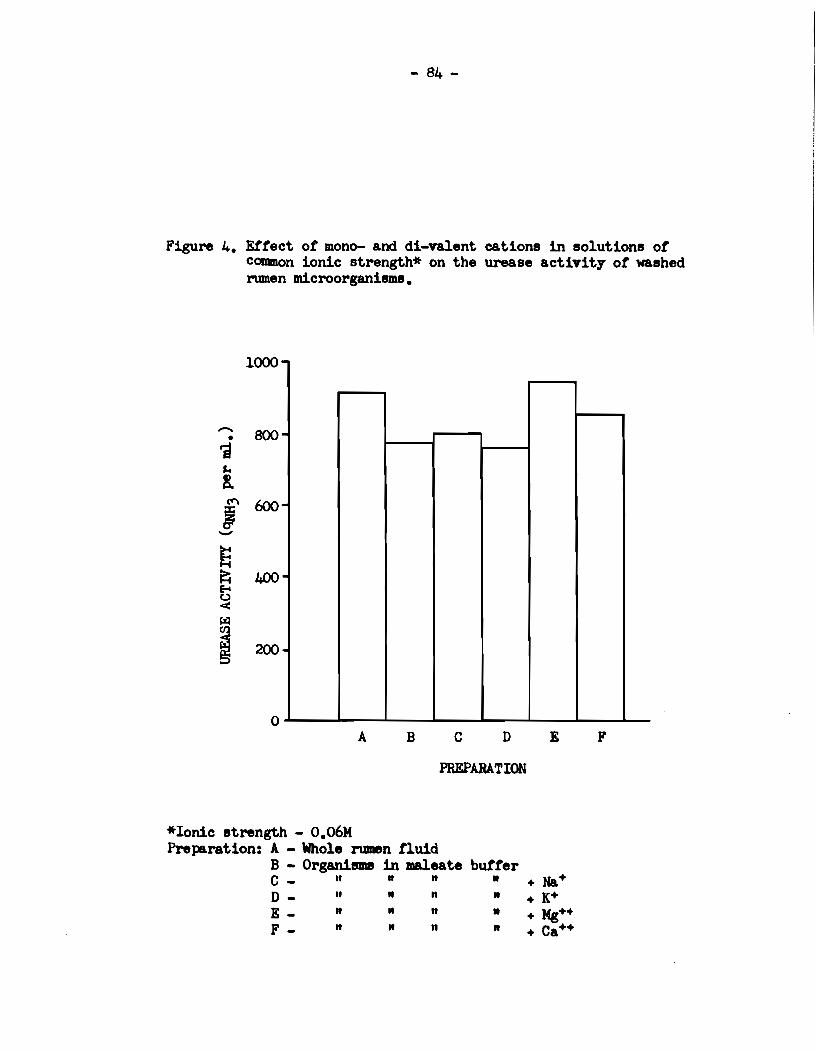
- 84 -
Figure 4. Ettect ot mono- and di-valent cations in solutions ot common ionie strength* on the urease activity ot washed rumen microorganisme.
1000·
- BOO . • i "" !. C""\ 600 -j -E ~ 400 . ~ < [;~';
~ 200 .
0~---~-----~--~------~--~----~----L--A B c D E
PREPARATION
*Ionie strength - o.06M Pre.r:aration: A - Whole rumen nuid
B - Organ181118 in mal.eate butter c- " " " • D- " " n •
E- " " n •
F- " " n •
+Na+ + K+ +Mg++ + ca++
F

- 85 -
differed slightly from animal to animal, the general affects
observed were the same with cella from ali three animals. It was
particularly noted that these effects did not differ significantly
from those found with Mg++ in the case of sheep 2 (Figure 6). It
was therefore concluded that the capacity of Mg++ to stimulate the
activity of rumen urease was not influenced by the presence of urea
in the diet of sheep 2.
The resulta in Figure 6 show that the effect of Ca++ on
the urease activity of washed cella from sheep 2 was almost identical
to that of Mg++. Between the affects of Mn++ and Mg++, however, an
appreciable quantitative difference was observed. Thus, in the
presence of 1 pM of Mn++ per ml. of buffer, the urease activity of
the washed cella was the same as that gi ven by a suspension of similar
cells in rumen supernatant liquor. In the presence of 12 - 20 JiM of
Mn++ per ml. the activity was raised to a level 56% above that given
by the cell suspension in rumen supernatant liquor and 41% above that
of whole rumen fluid. Thus it was concluded that di valent ions,
probably Mn++, Mg++ and Ca++, were the factors present in rumen super
na tant liquor which were responsible for stimulation of the urease
activity of intact rumen bacteria.
It was not possible to draw a conclusion regarding the way
in whichureas.e stimulation was brought about by these ions, however,
Since the experimenta had been carried out using intact rumen micro
organisns in which the enzyme was entirely intracellular, the affects
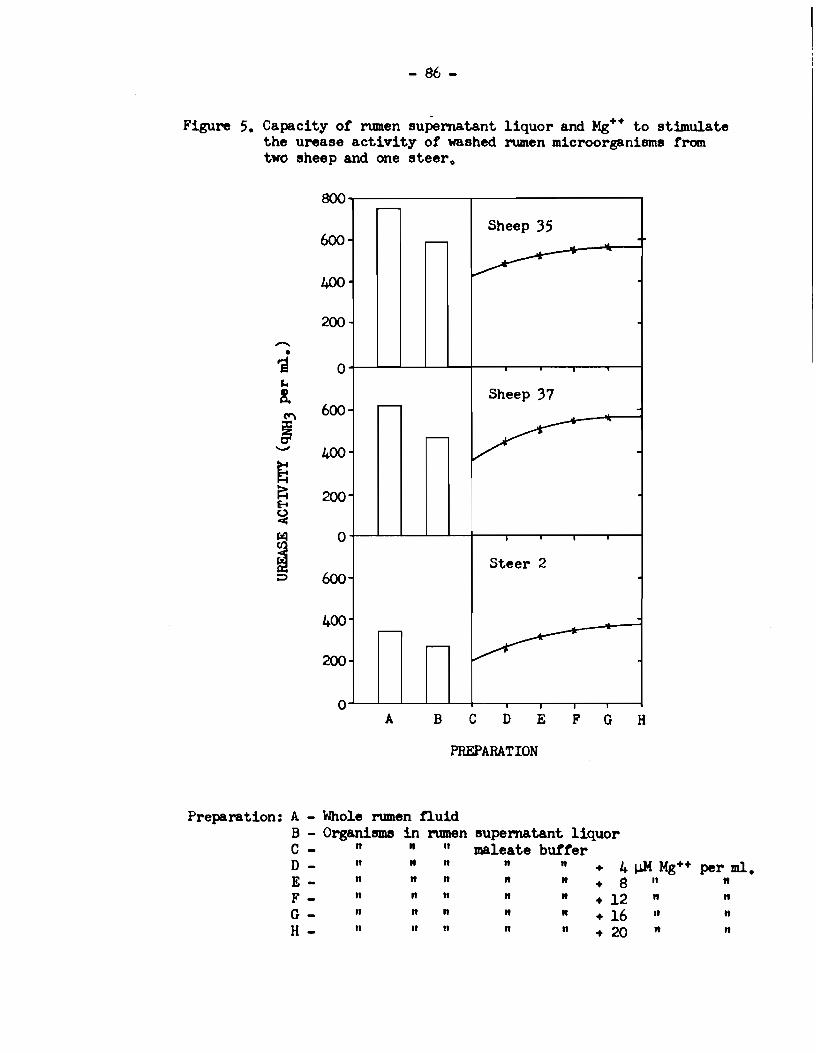
-86-
Figure 5. Capacity or rumen su}iematant liquor and Mg++ to stimulate the urease aetivity of washed rumen mieroor8&nisms tram two sheep and one steer.
aoo r--
600 . Sheep 35 r--
400 ~
200
-• 'i1 0 ,..
Sheep 37 r--
!. 600 ('t'\
r--~ . j - 400
~ ~ 200 E-t
~
Steer 2
P:l 0
~ 600 :::>
400 r--
r--200
0 • A B C D E F G H
PREPARATION
Preparation: A - Whole rumen tl.uid B - Organisms in rumen supernatant liquor c - " " " ma1eate butter D - " n " " n + 4 ~ Mg++ per ml. E- n tt n n " + 8 u " F _ n n tt n 11 + 12 n n G _ n n n tt tt + 16 11 tt
H- tt 11 n n n +20 n n
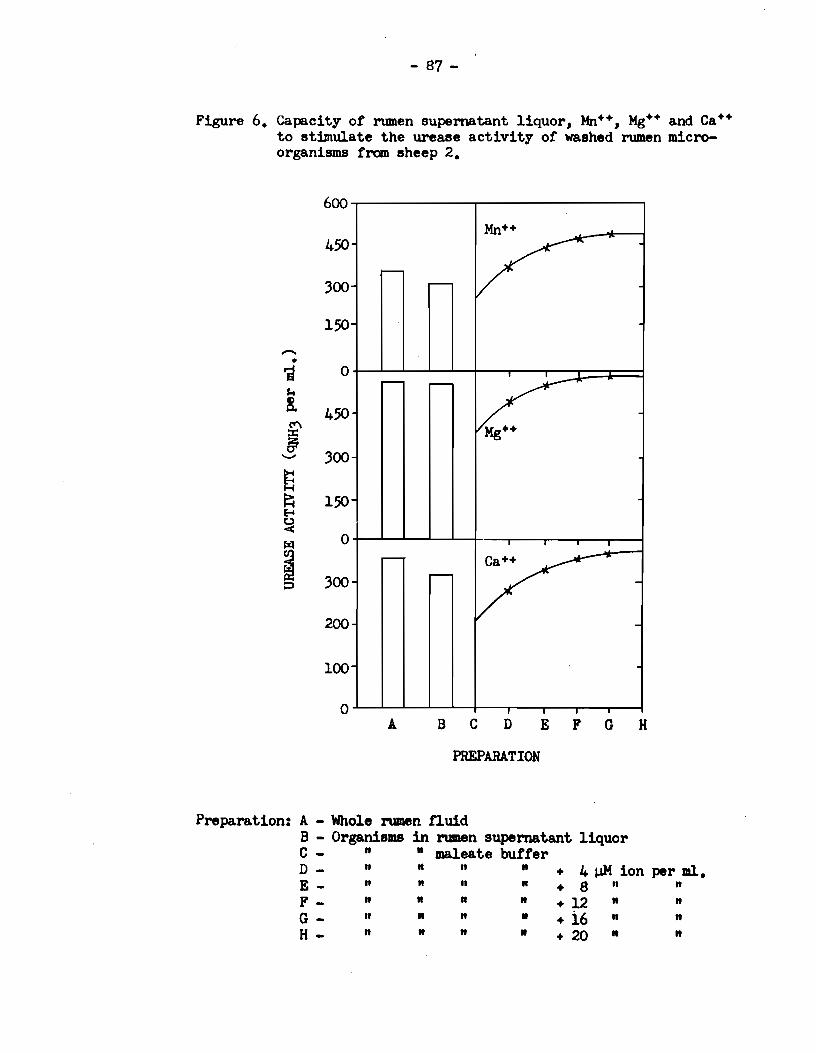
- 87-
Figure 6. Capacity or rumen supematant liquor, Mn++, Mg++ and ca++ to stimulate the urease activity or washed rumen microorganisme from sheep 2.
600
450
300
150
-• i 0 $.e
!. 450 ~ g;
300 -E ~ 150 f-4 ~ !';il 0
1 300 p
200
100
0 A B c D E F G H
PREPARATION
Preparation: A - Who1e rumen fiuid B - OrganisiiiB in rumen supematant 11quor C - " • maleate butter D - " " 11
• + 4 J,J.M ion per ml. E- n n 11 11 + 8 11 n
F- n " n • +12 • " G - n • " 11 + i6 " " H- " 11
" • +20 " "

- 88 -
observed could have been due either to the influence of the ions on
the permeability of the cells to urea or to an absolute effect of
the ions on the intraeellular enzyme. In order to de.monstrate which
of these affects was in fact involved, a series of experimenta was
carried out using as sources of urease preparations in which the
permeability properties of the intact cells were respectively modi
fied and eliminated, nam.ely, by preparing an acetone-dried powder
and a cell-free extract of the mixed flora of rumen microorganisme.
b) Ex:pe riments wi th an acetone -dried powder of rumen micro
organisns
An aeetone-dried powder was prepa.red from washed rumen
microorganisns and determinations were made of the affects on its
urease activity of suspending the powder in va.rioua media. The
resulta presented in Table VIII show that tm urease activity of the
powder was influenced in a mannar similar to that of the intact cella.
Thus, when the powder was suspended in rumen supernatant liquor or in
a solution in buffer of the ash of rumen supernatant liquor the
urease activity of the powder was appreciably greater than 'When it
was suspendad in buffer alone, indicating that urease activity was
enhanced by an :i,norganic factor present in rumen supernatant liquor.
The combinations of cations previously deseribed were te sted .for
their affects on the enzyme. It is evident from the resulta in
Table VIII that the group of alkaline earth metal ions again enhanced
urease aetivity. It was concluded there.fore that the ureaae aotivity
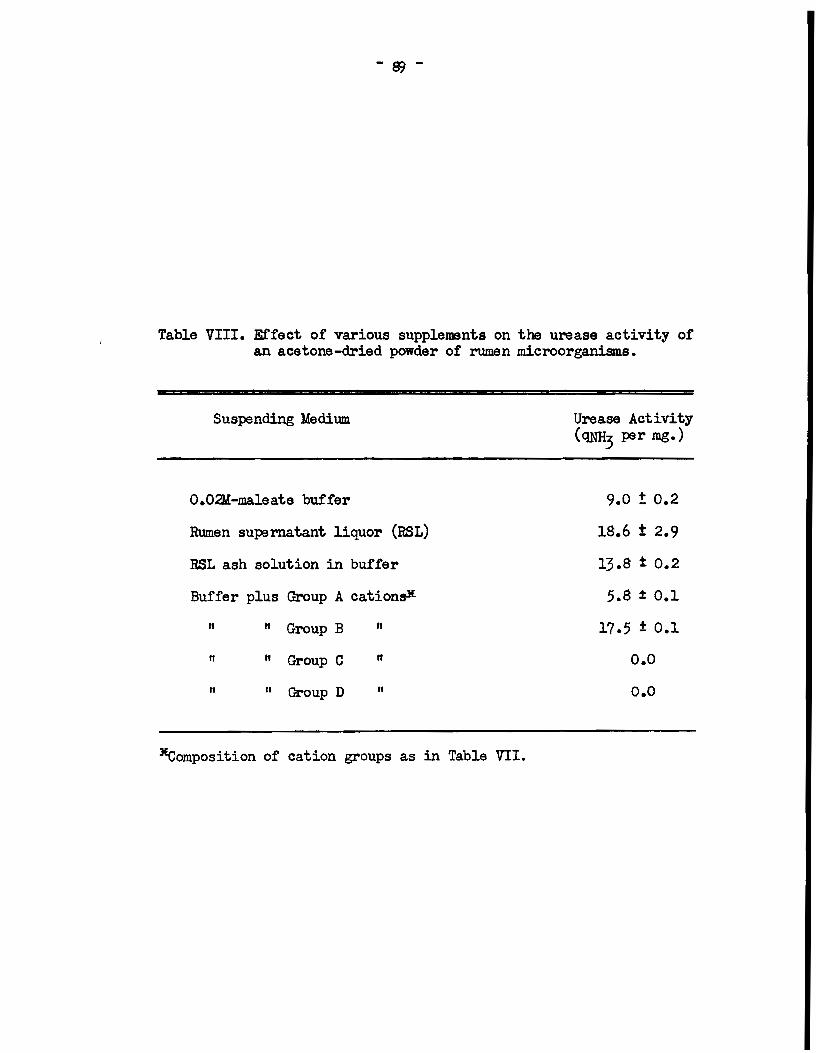
- fJJ -
Table VIII. Etfact of various supplements on the urease activity of an acetone-dried powder of rumen .microorganism.s.
Suspending Medium
o.02M-maleata buffer
Rumen supernatant liquor (RSL)
RSL ash solution in buffer
Buffer plus Group A cationsx
Il " Group B tl
11 " Group C
tt 11 Group D Il
lEcomposition of cation groups as in Table VII.
Ureasa Activity ( <lliH3 per mg.)
9.0 ! 0.2
18.6 t 2.9
13.8 t 0.2
5.8 :!: 0.1
17.5 :t 0.1
o.o
o.o

- 90-
of an acetone-dried preparation of rumen microorganisms wa.s stimul
ated by divalent cations.
c) Ex:periment with a cell-free extract of rumen microorganism.s
Since studies have shown that treating cells with acetone
modifies their permeabllity properties (Gunsalus, 1955), the affect
of inorganic ions on the urease activity of whole cells would appear
not to be an affect on membrane permeability. The re is no information,
however, on the extent to which acetone treatment modifies the pe:nnea
bllity pro}:erties of rumen microorganisme. To establish beyond doubt
whether the e:f'fects of' the ions on the 11\hole cella were due to affects
on the urease enzyme or enzymes, cell-free extracts of the organisms
were prepared. This was done by subjecting a suspension of washed
rumen microorganisme in maleate buffer to sonic oscillation. The
preparation was centrifuged free of whole cells and debris. The
affects of rumen supernatant liquor and various concentrations of
Mg++ on the urease activity of the extract 118re then determined. The
resulta, which are presented in Figure 7, show that the enzyme
activity was strongly stimulated by both rumen supernatant liquor and
Mg++. The resulta obtained with this cell-free extract of rumen
microorganisme therefore resembled closely those obtained with whole
cella. It was concluded that divalent ions exerted an absolute
stimulating affect upon the urease enzyme and that the affects
observed with the whole cells were not due primarily to the influence
of the ions on the permeability of the cell membrane.
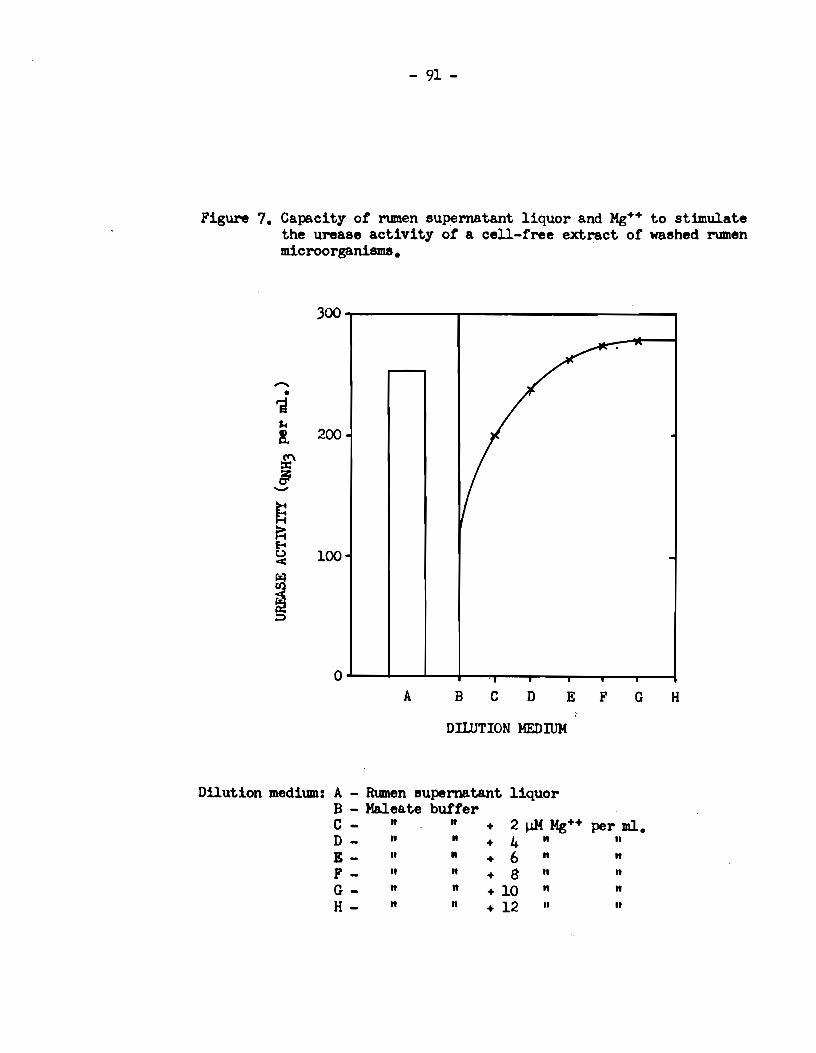
- 91 -
Figure 7. Capacitr of nunen su~rnatant 1iquor and Mg++ to stimula te the urease activitr of a cell-free extract of washed rumen microorganisme.
-• Et ... 8. 200
r;: §t -~ ~ f-4 ~ 100
1 :::::>
A B c D E F G H
DIWTION MEDD.JM
Dilution medium: A - Rumen supernatant 11quor B - Maleate butter c- " n + 2 IJ.M Mg++ per ml. D - lt lt + 4 n tt
B- " .. + 6 lt " F- tt n + 8 " n
G- n If + 10 n " H- n n + 12 Il n

- 92 -
B. Effect of Divalent Ions on the Activity of Jack-bean Urease.
In order to determine the affects of divalent ions on the
activity of jack-bean urease a solution of the enzyme in maleate
buffer was prepared and its activity in the presence of five ions
at concentrations of 12 uM per ml. of solution was measured. The
results are shown in Figure S. It is clear from these results that
the affects of the ions on jack-bean urease ware very different from
their affects on rumen urease. Thus, whereas the five ions ail
st.imulated rumen urease to varying extents, they all inhibited jack
bean urease, the sequence of inhibition being Ca++~sr++~;7Mg++ ~
Mn++/Ba++.
C. Localization of Urease Activity in Rumen Microbial Cells.
The results so far reported indicate that the stimulation
of the urease activity of intact rumen microorganisms by inorganic
ions was due prim.arily to the affects of the ions on the urease enzyme.
This suggested either that the urease enzyme was located on the cell
surface o:f these organisns or that the cell membrane of the intact
cells presented little or no obstacle to the passage of the ions into
the cell. In an effort to distinguish between the se two possibilities,
the following experimenta were perfonned.
When inorganic ions were added to suspensions of washed
rumen microorganisms it was observed that there was a t.ime lag before
the ions exerted their stimulating or inhibiting affects. This is
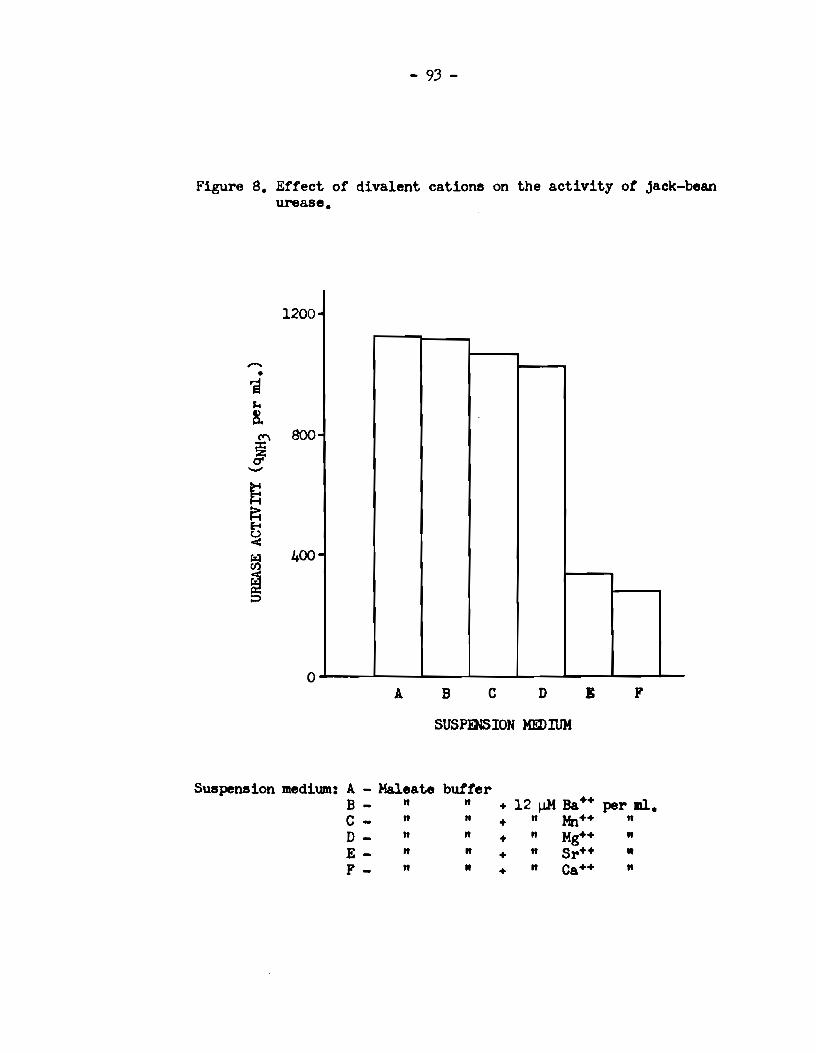
- 9:3 -
Figure s. Effect of diva1ent cations on the activity of jack-bean urease.
1200·
-• Et -.. !. "" 800 . J -E i:i E-4 c <
1 400 .
:::>
0._--~----~--~----._--~----~---------
A B c D B F
SUSPF.XSION Ml!DllJM
Suspension medium: A - Maleate butter B - " " + 12 ~ Ba++ C _ 1t 11 + n Mn++
D - " " + " Mg++ E - " " + " sr++ F - " " + " ca++
per ml. " lt .. "

- 94 -
shawn in Figure 9 with various ions capable of stimulating urease
activity. The shortest lag occurred with Mn++, the ion producing
the greatest stimulation. The longest time lag occurred with sr++.
Once the stimulation of enzyme activity had begun, the reaction
proceeded at a constant rate which varied wi th the stimula ting ion
tested. Similarzy, inhibiting ions showed a time lag bef ore inhib
ition could be observed, Figure 10. The time lags observed before
the initiation of stimulation or inhibition could beat be explained
if one assumed that the urease enzyme was intracellular and that the
different ions took varying lengths of time to cross the membrane and
reach a concentration at which they could e:xert an effect in tbs cells.
If' this were so, one could conclude that the rate of penetration of
the urease stimulating ion decreased in the order of increase in the
lag period, namezy, Mn++ ?Mg++ :;::::=--ca++ 7 Ba.++> sr++, and tho se of
the inhibiting ions in the ord.er Co++/ K+ >Na+.
If' tm time lags observed were indeed due to barriere to
the read.y penetration of the ions to tm site of an intracellular
enzy.m.e, the tim.e l.ags shoul.d be modi.fi.ed by factors which woul.d be
expected to modify the permeability of the membranes. For this
reason the time talœn by various ions to initiate stimulating or
inhibiting affects on rumen urease activity were determined using an
acetone powd.er of the rumen microbial cells. The resulta in Figures
11 and 12 show that even with the acetone powder there was a lag
before the effects of the stimulating and inhibiting ions expressed
themselves. This suggests that if the time lags were due to
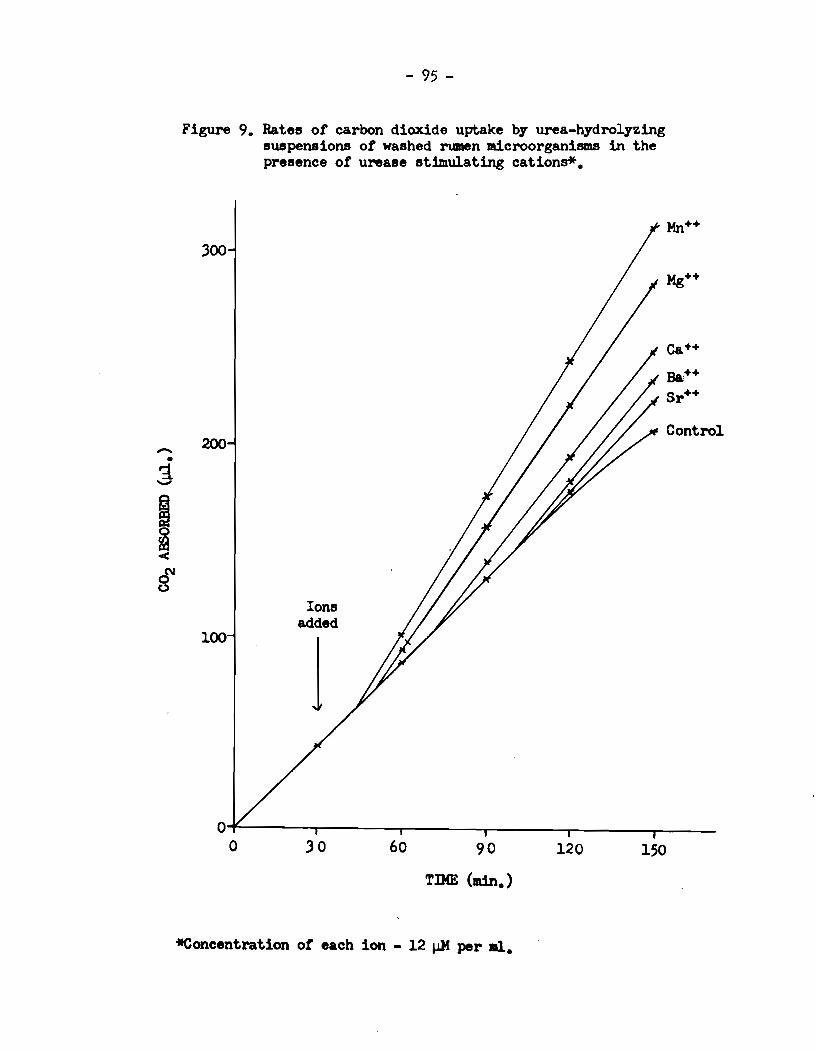
-• '1 -1 < ON 0
- 95 -
Figure 9. Rates ot carbon dioxide upt,ake by urea-hydrolyzing suspensions ot washed rumen m.icroorganisms in the presence ot urease stimula ting cations*.
300
200
Ions ad.ded
100
l
Mn++
Mg++
ca++
:sa++ sr++
Control
0~----~------,-----~,-----.-------~---0 30 60 90
TIME (min.)
*Concentration ot each ion - 12 IJ.M per al.
120 150
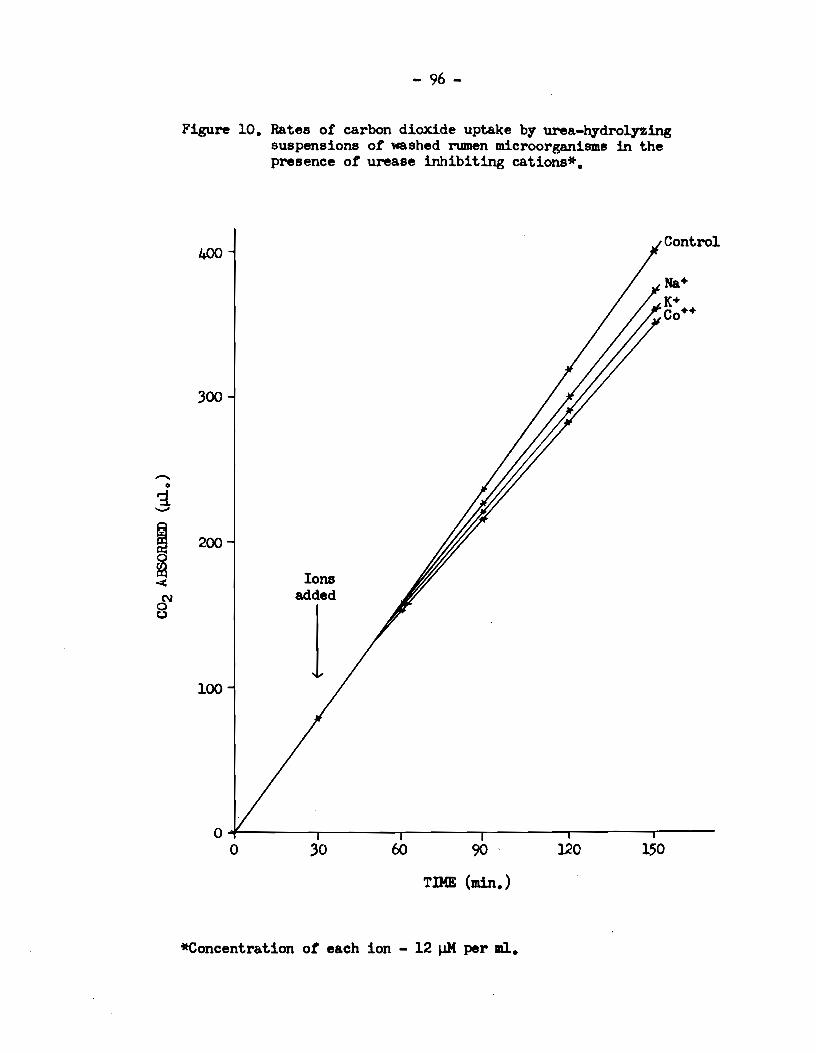
-• 3. -1 fe < N
0 0
- 96 -
Figure 10. Ra tes of carbon dioxide uptake by urea-hydrolyzing suspensions of washed rumen microorganisme in the presence of urease inhibiting cations*.
Control
300
200
Ions added
1 100
0~------~------~----~.-------.-------r-----o 30 60 90'
TlME (min.)
*Concentration of each ion - 12 ~ per ml.
120 150

- 97 -
variations in the rates of penetration of the ions the acetone
treated cella still reta.ined soma of the permeability properties
of the whole cella. The relative lengths of the time lags with
the various ions showed differences from those obta.ined with whole
cella, however, suggesting tha.t the permeability properties of the
acetone-treated cells bad in fact been modified. Cell penetration
by J.fn++, for exa.mple, was slower with the acetone-treated cells tha.n
with the whole cells, whereas the absence of a time la.g in the case
of Co++ suggested that the acetone-treated cella were freely perm
eable to this ion. The order of increasing time lag for the stimul
ating ions using the acetone powder wa.s Mg++ = Ca++c:::::..Mn++L. sr++<
Ba+•. That of tœ inhibiting ions was co••.c:::::.K+; Na+ exerted no
apparent effect on urease activity within 2 br. There were aga.in
differences between the affects of the various ions after the initial
lag period. The sequence of these affects for the stimulating ions,
in decreasing order of magnitude, was :Mg++/Ca++,?Mn•+:::::>sr+•:;::::..Ba++,
which is similar to the order in which they appeared to penetrate the
acetone-treated eells. It was different, however, from the sequence
previously found for the absolute affects of these ions on the urease
activity of whole cells.
To establish if the time lags observed were in fact due to
permeability affects, the action of tœ ions on a cell-free extract
of rumen microorganisms was studied. The resulta obtained, Figure 13,
show tha.t Mn,.,.., :Mg++ and Ca,.,.. all stimula.ted urease activity immediately
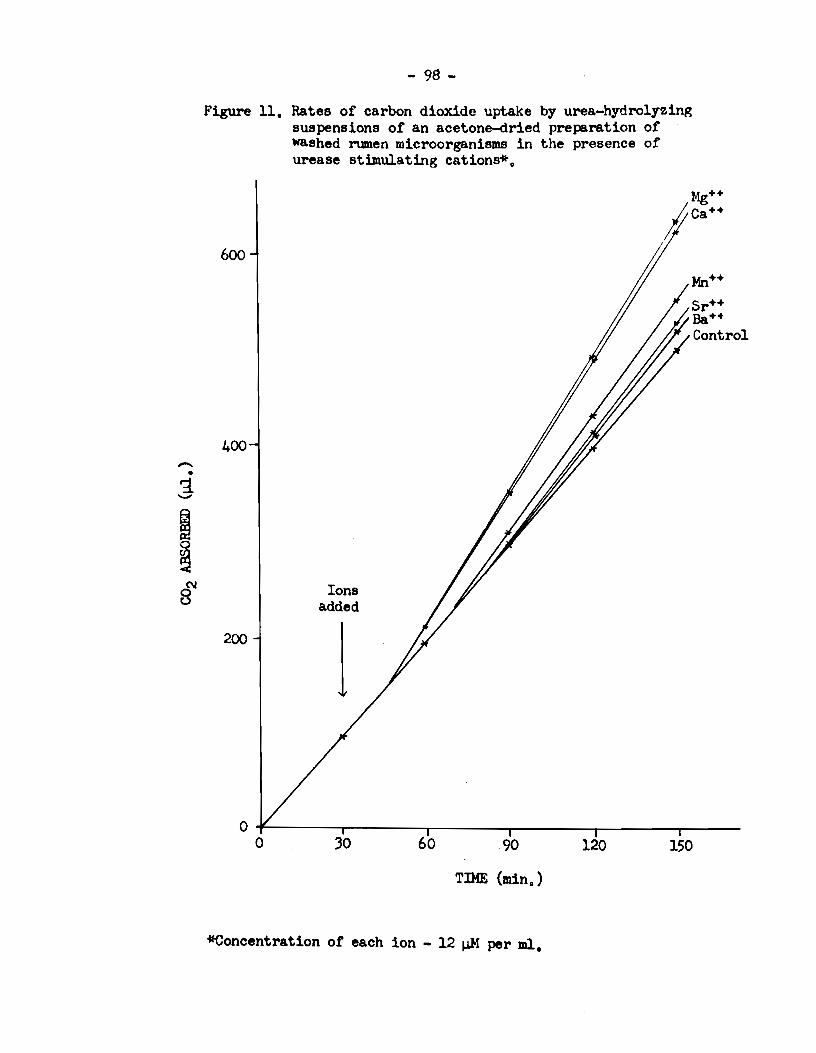
-• '1 -1 83 « 8N
- 98-
Figure 11. Rates of carbon dioxide uptake by urea-hydro1yzing suspensions of an acetone-dried preparation of w.ashed rumen microorganisms in the presence of urease stimulating cations*o
600
400
Ions added
200
1
Mg++
jca++
Mn++
sr++ Ba·· Control
0 ~------r-------r-------r-------r-------r-----0 30 60 90
TIME (mino)
*Concentration ot each ion - 12 ~ per mi.
120 150
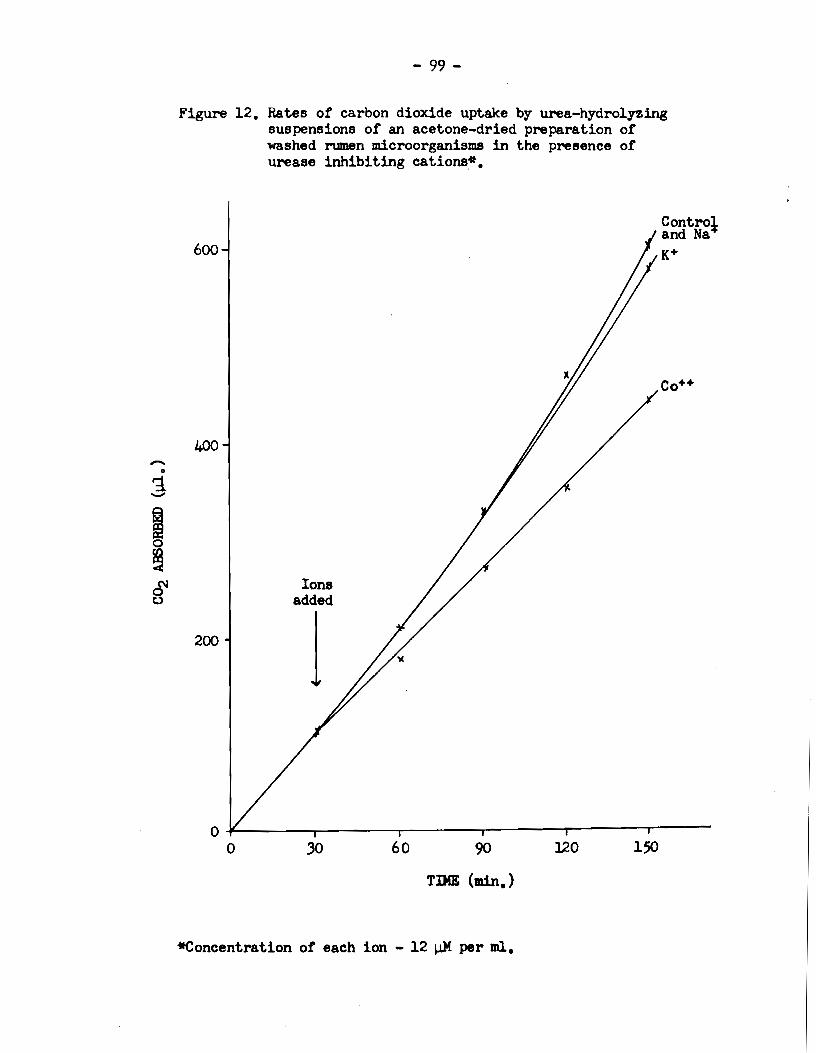
-0
3. -1 ~ ~ 0
- 99 -
Figure 12. Rates ot carbon dioxide upta.ke by urea-hydro1yzing suspensions ot an acetone-dried preparation ot washed rumen microorganisms in the presence ot urease inhibiting cations*.
600
co++
400
Ions added
200 1
0~----~------~----~------~----~-----0 30 60 90 120 150
TIME (min.)
*Concentration ot each ion - 12 j.1M per ml.
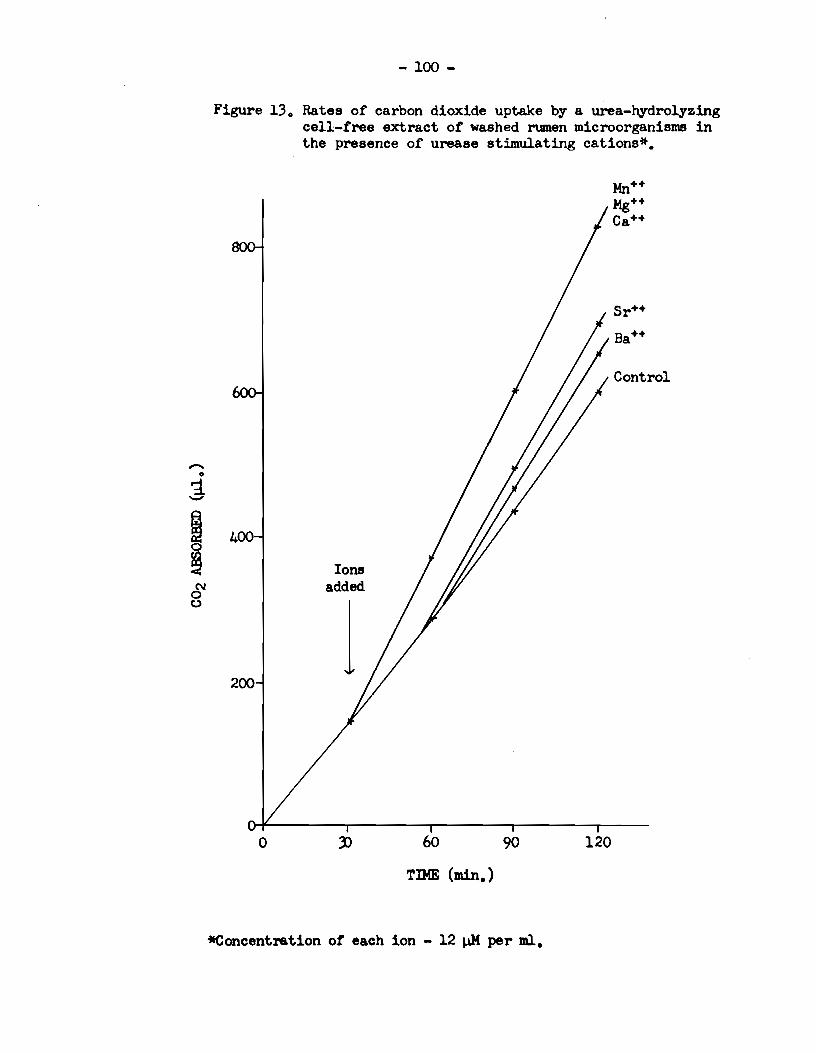
-• '1 -1 ~ N
0 t,)
-lOO-
Figure 13. Rates of carbon dioxide uptake by a urea-hydrolyzing cell-free extract of washed rumen microorganisms in the presence of urease stimulating cations*.
400
Ions added
1 200
60 90
TlME (min.)
*Concentration of each ion - 12 ~ per ml.
120
Mn++ Mg++ ca++
sr++
Ba++
Control
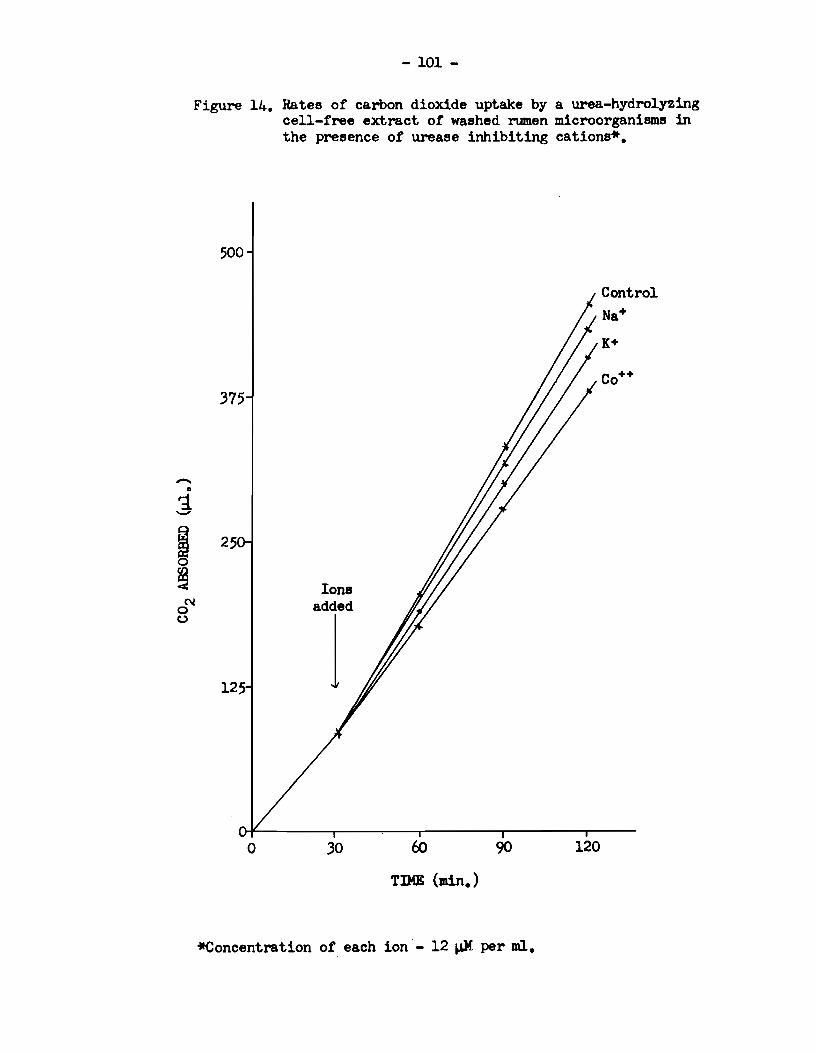
-0
'3. ..._,
1 ~
C\1 0 0
- 101 -
Figure 14. Rates of carbon dioxide uptake by a urea-hydrol)'fiing cell-free extract of washed rumen microorganisme in the presence of urease inhibiting cations*.
500
Control Na+
K+
co•• 375
25
Ions added
12 l
0 30 f:IJ 120
TDΠ(min.)
*Concentration of each ion - 12 ~ per ml.

- 102-
upon addition of the ions to the urea-hydrolyzing extract.sr+-+- and
Ba+-+, on the otber hand, both stim.ulated the enzyme only a:tter a
lag period. In the case of inhibiting ions, Figure 14, Co++ and K+
both inhibited the enzyme immediately upon addition to the enzyme
system, while Nat"was effective only a:tter a 1ag period.

DISCUSSION
The resulta of this study have clearly shown that the rumen
solution contains inorganic components which have the capacity to
stimulate intracellular rumen urease, and evidence has been presented
that the alkaline earth metal ions and Un++ also have this capacity.
Of these ions, Ca"'+ and llgu are known to have important
fun ct ions in ruminant metabolism (Duncan, 1958; Rook and Storry, 1962).
Analyses of rumen liquor from sheep on various diets (Gerton, 1951)
have shown that the rumen solution may contain concentrations of Ca++
and Mg++ equivalent respectively to 2.5 - 5.0 JiM and 4.0 - 5.0 )fM per
ml. The significance and function of Un++ in the rumen, on the other
hand, is but little understood (Barnett and Reid, 1961) and a search
of the literature has failed to reveal a reference to the normal
concentration of soluble Mn++ in the rumen solution. From experimenta
on the requirements of ruminants for dietary Un++, which have generally
been found to be lesa than 50 p.p.m. of the ration (Bentley and Phillips,
1951; Embry, Ga.stler, Radabaugh and OJ.son, J.95S), it may be deduced,
however, that the rumen concentration of soluble Mn++ is considerably
lower than that of either ca++ or :Mg++. Neither sr++ nor Ba++ is
known normally to have any significance in rumen metabolism. From
a consideration of the concentrations of divalent ions found to
stimulate rumen urease in vitro, it seems reasonable to conclude
that Ca++ and Mg++ were probably the components of the rumen solution
which were primarily responsible for its stimulatory affects on the

- 104-
enzyme.
A search of the literature revealed one other reference to
the stimulation of urease by alkaline earth metal ions. This wa.s a
report by Shibata (1958). The original paper was not seetJ,however,
and neitmr the source of the urease used by the author nor the
concentrations of the cations found to be effective is lmown. The
present investigation has shovm. that the affect of these ions on
jack-bean urease at the concentration tested was one of inhibition
rather than stimulation. Jack-bean urease was also found to be
inhibited by Mn++, and this is in agreement with the findings of
Shaw (1954). This au thor has postulated that the mechanism of
urease inhibition by several cations, including Jln"''t', involves
a.ttachment of the ion to the sulphydryl groups of the enzyme, since
ali the cations involved f'orm insoluble sulphides.
Clearly a .mechanism of f'undamentally different character
must be involved in the case of' Jln++ and rumen urease , for here an
appreciable increase in the activity of the enzyme resulted. Wha.t
this mechanism might be, however, cannot a.t present be suggested,
nor can a mechanism by which the other di valent cations may stimu1-
a.te rumen urease be postulated. Rumen urea.se wa.s found to be
inhibited by Co'"+, Na++ and K+ and therefore in this respect resem
bled jack-bean urease which is also inhibited by these ions (Wall and
Laidler, 1953; Shaw, 1954).
The reduction in urease activity which wa.s found in tm
present study to accompany the washing of intact rumen microorganisms

- 105 -
with buffer indicated a reduction in the intracellular concentration
of di valent cations. Furthermore, the suspension of resting intact
cells in solutions of divalent ions resulted in uptake of the ions
by the cells, together with a subsequent enhancement of urease
activity. These observations show, firstzy, that the ions were not
bound to any appreciable extent intracellularly, and secondly, that
the ions were qui te readily diffusible into the cella in the re sting
state. This situation shows some differences from that found to
e.xist in other whole cell systems. Tsuyuki and Maclsod (1951), for
example, round that extracellular Mg+-+ and )ln++, which ions are
essential cofactors for several enzymes involved in glycolysis, did
not affect the rate of glycolysis by resting cell suspensions of
Lactobacillus arabinosus grown in a Mzl++-deficient medium, although
:Mn++ was required for growth by the organism. The authors interpreted
their resulta as indicating that Mn++ was bound intracellularly and
that during washing of the cella the concentration of the ion within
the cella remained sufficiently high so that further addition of
divalent cations to a suspension of the cella bad no sti.mulating
effect on metabolism.
A dependance of divalent cation absorption on metabolic
activity was shown for baker 1s yeast in isotopie studies by Rothstein
and his associates (Rothstein and Hayes, 1956; Rothstein, Hayes,
Jennings and Hooper, 1958; Jennings, Hooper and Rothstein, 1958).
If resting cella of the organism were placed in solutions containing
Mn++, Mg++, ca++ or sr++, a rapid and reversible binding of the ion
at specifie sites on the cell membrane oecurred to a li.mited extent.

- 106-
Beyond this surface binding thare was no absorption of tm ion, nor
did divalent cations leak out of tm cytoplasm, showing that thase
ions could not penetrate tm cell membrane. If phosphate and glucose
11ere added to the suspension, however, in addition to the surface
binding soma di valent ion was irreversibly absorbed by tm ce lis.
The sequence of absorption of divalent cations by metabolizing cells
of the yeast was Mg·f..f· /Mrl'""">Ca++:;::>SrH. In the present study it
was found that Mn,++ penetrated tm resting cella of rumen bacteria
more rapidly than :Mg++, but otherwise the seque nee was similar.
It is concluded from this investigation, therefore, that
rumen ureolytic bacteria differ from soma other biological systems
both with respect to pe:nneability of the cella to cations, and with
respect to the susceptibility of their urease to stimulation by
dival.ent cations. It is of interest to note, when considering tm
per.meability of the cella to cations and the affects of these cations
on urease acti vity, that the time lags which occurred be fore J4n++,
:Mg+"" and Ca+.,. stimulated the urease activity of intact cella, were
attributable entirely to the time taken for these ions to penetrate
the ce lis, whereas in the case of sr++ and Ba++ this was not so.
Even when these latter ions were in apparently free contact with the
enzyme, i.e. , in the case of the cell-free extract of rumen micro
organisme, stimulation did not occur imrnediately. This suggests
that an active complex between ion and enzyme was not imrnediately
formed.

- 107 -
Rumen bacteria, as a.lready mentioned, no:rma.lly exist in a
medium, na.mely the rumen solution, whieh conta.ins signi.ficant concen
trations of soluble Ca++ and :Mg++. These concentrations are contin
ua.lly ma.inta.ined by solubilization of constituants of the anima.l8
diet. The rumen solution also contains relatively large a.mounts of
Na1' and K"'. For example, Garton {1951) found each ion to be present
in tha rumen solution of sheep to the extent of 100 - 200 mg. per
100 nù.. Ail these ions have been shown in the present study to be
absorbed by resting cells of urease-producing rumen organisme and
subsequently to influence the urease activity of the organisme. If
a simila.r absorption occurs in the case of actively metabolizing
cella, it may be postula.ted that tha activity ma.nifested by urea.se
producing organisme in !!!.2 is the resultant of opposing stimulating
and inhibiting affects on the intracellular enzyme brought about by
ions which are present in the rumen solution.
A detailed study of the bacteria predomina.ntly responsible
for rumen urease production must await the isolation in pure culture
of the organisme concemed. This, as indicated in Part I, is a matter
which presents considerable problems. Fresh attempts to isolate such
organisme, however, would appear to be of potential value, for the
present investigation has shown that a study of their physiologica.l
properties may prove to be a matter of considerable interest from.
the viewpoint of comparative biochemistry.

SUMMAR.Y
A study was made of soma factors affecting the activity
of rumen urease, uaing preparations of .mi::xed rumen microorganisme
as a source of enzyme •
When whole cells o:f rumen microorganisms were washed with
maleate buf:fer there was a reduction in the urease activity o:f the
cells. Resuspension o:f the washed cells in rumen supernatant liquor,
however, brought about an enhancement of the ir urease activity. This
e:f:fect was due to the pre sance of an inorganic rumen urease stjmulat
ing factor in rumen superna.tant liquor.
When whole cella were washed in bu:f:fer there was a pro
gressive reduction in the urease activity of the cella as the number
of washings increased. Up to the third washing the activity o:f tœ
cells could be restored to the level prevailing in unwashed cells
by resuspending the washed cells in rumen supernatant liquor. A:fter
the sixth and ninth washings, on the other hand, the urease activity
of the cella could not be fully restored in this way.
Five divalent cations also had the capacity speci:fically
to stimulate the urease activity of washed whole cells of rumen
microorganisme. The degree of urease stimulation brought about by
these ions decreased in the sequence Mn,++~g++7Ca++:;:>sr++/Ba++.
The activity of the cella was reduced in the presence of Na+, K+

- 109-
and Co+t-, and tm enzyme was completely inhibited by Zn++, eu++ and
Fe+++.
Solutions of Mg++ had the capacity to replace rumen super
natant liquor in the restoration of urease aetivity to washed whole
ce ils of run.en microorganisme from a urea-fed sheep and fran two
sheep and a steer not receiving dietary urea. Solutions of Ca++ had
a similar capacity to Mg++ in restoring the activity of washed cells
from the urea-fed animal. The affect of solutions of Mn++, however,
was to raise the activity of washed cells from this animal to a level
approximately 40% greater than that prevailing in unwashed cells.
The urease activity of an acetone-dried powder of washed
rumen microorganisme was stimulated by inorganic factors present in
rumen supematant liquor and also by the group of divalent cations
Mg++, Ca++, sr++ and Ba++.
The urease activity of a cell-free extract of washed
rumen microorganisma was stimulated by rumen supernatant liquor. of
Solutions Mg..,.., could replace rumen supernatant liquor in stimulating
the activi ty of the ext.ract.
It was concluded that di valent ions, probably Un++, Mgt-t-
and Ca++, were the factors present in rumen supernatant liquor which
were responsible for the stimulation of rumen urease. Moreoever, the
stimulating effect of these ions on the urease activity of whole cella
was due primarily to the affects of the ions on the urease enzyme and
not to their effects on permeability of the cells to urea.

- 110-
The activity of jack-bean urease was reduced in the presence
of all divalent ions found to stimulate the activity of rumen urease.
The degree of inhibition of the enzyme brought about by these ions
in decreasing order was Ca""""> Sr"""".>Mg""""?M'n""""?Ba""""· Apparently,
therefore, the re exista a funda.mental difference between rumen urease
and jack-bean urease with respect to their response to divalent cations.
When inorganic ions were added individually to actively urea
hydrolyzing suspensions of washed rumen microorganiams, there were time
lags before the ions exerted their st:imulating or inhibiting effects.
The time lags with urease stimulating ions increased in the order
Mn""+~ :Mg++<: Ca+"< Ba++<:::: Sr"""", and those wi.th the urease inhibiting
ions in the order Co""~ K+<.. Na .......
Time lags were also observed when inorganic ions were added
to urea-hydrolyzing suspensions of an acetone-dried powder of washed
rumen microorganisms. In this case the lags increased in the orders
MgH = Ca++<Mn""+< Sr++.C:::::::. Ba++ and Co++< K+ with the stimula ting
and inhibiting ions respectively.
The re l'lere no time lags, however, be .tore the initiation of
effects on urease activity when Mn+ .. , Mg....,, Ca++, Co++ or K+ "Were
added to a urea-hydrolyzing cell-free extract of washed rumen micro
organisms, although in the case of sr++, Ba++ and Na+ such time lage
were observed.
The tim.e lags observed with the whole cella were explained

- lll-
in tenns of the varying lengths of time taken by different ions to
cross the cell membrane and reach a concentration at which they
could exert an affect on the intracellular urease enzyme. Modifi
cation of the perm.eability properties of the cells by treatment with
acetone also modified the rates of penetration of the ions. The tim.e
lags observed when sr++, Ba++ and Na"' were added to the cell-free
extract suggested that there was a delay in the formation of an
active complex between the ions and the enzyme.
The absolute affects of the different ions upon the urease
enzyme varied with the different preparations of rumen urease studied,
in a mannar generally sim.ilar to that of variations in the cell
penetration rates of the ions.
The differences which exist between rumen ureolytic bacteria
and sana other biological systems, both with respect to the pe:rmeability
of cella to cations and the susceptibility of urease to stimulation
by di valent cations, were discussed with reference to the rumen envir
onment.

BIBLIOGRAPHY
Abou Akkada, A.R., and Howard, B.H.
1960 The biochemistry of rumen protozoa. 3. The carbohydrate metabolism of Entodinium. Biocham. J., 76: 445-451.
Amdson, E.F., Chalmers, M.I., Marshall, S.B.M., and Synge, R.L.M.
1954 Ruminal anmonia formation in relation to the protein requirement of sheep. n. Ruminal amnonia formation with various diets. J. Agr. Sei., 44: 'Z/0-2:73.
Annison, E.F., and Iswis, D.
1959 Metabolism in the Rumen. Metheun andco:-;-London.
Argenziano, G., and Giani, E.
1946 Boll. Soc. Ital. Biol. Sperim., 22: 123-124. Cited in Chem. Abstr., 21: 21453(1947).
Barnett, A.J.G., and Reid, R.L.
1961 Reactions,!!! the Rumen. Edward Arnold Ltd., London.
Bauman, H.E., and Foster, E.M.
1956 Characteristics of organisms isolated from the rumen of cows fed high and low roughage rations. J. Bacteriol., 71: 333-33S.
Belasco, I.J.
1956 The role of carbohydrates in urea utilization, cellulose digestion and fatty acid metabolism. J. Animal Sei., 15: 496-508.

- 113 -
Bentley, O.G., and Phillips, P.H.
1951 The effect of law manganese rations upon dairy cattle. J. Dairy Sei., 34: 396-403.
Bergey 1 s Manual of Determinative Bacteriology
1957 7th ed. Ed. by Breed, R.s., Murray, E.G.D., and Smith, N.R. Williams and Wilkins Co., Baltimore.
Blackburn, T.H., and Hobson, P.N.
1962 Further studies on the isolation of proteolytic baeteria from the sheep rumen. J. Gen. Microbiol., 29: 69-89.
Bouckaert, J.H., and Oyaert, W.
1952 Fate of some nitrogen eanpounds in the rumen. Zooteehnia, 1: 21-29.
Brisou, J.
1949 Bull. Soc. Chim. Biol., 31: 1656-1658. Cited in Chem. Abstr., 44: 7927(1950).
Bryant, M.P.
1959 Bacterial species of the rumen. Bacteriol. Rev. , 23: 125-153.
Chibnall, A. C.
1939 Protein Metabolism in tœ Plant. Yale Univ. Press, New Haven.

- ll4-
Christensen, W.B.
1946 Urea decomposition as a means of differentiating Proteus and paraco1on cultures from each otœr and from Salmonella and Shigella types. J. Bacteriol., 52: 461-466.
Dixon, M., and Webb, E.C.
1958 Enzy.mes. Longmans, Green and Co., London.
Doetsch, R.N., Robinson, R.Q., and Shaw, J.C.
1952 Techniques employed in cultural investigations of the bacterio1ogy of bovine rumen contents. J. Animal. Sei., li: 536-544 ..
Dukes, H.H.
1955 ~ PhYsiologY gf Domestic Animals. 7th ed. Camstock Pub1ishing Associates, Ithaea.
Duncan, D.L.
1958 The interpretation of etudies of calcium and phosphorus balance in ruminants. Nutr. Abstr. Rev., 28: 695-715.
El.l.is, W.C., and Ptander, W.H.
1958 In vitro rumen microbial protein and nuc1eic acid metabolism .. J. Animal Sei., 17: ll92.
Embry, L.B., Gastler, G.F., Radabough, D. V., and Olson, O.E.
1958 Manganese requirements of grow:i.ng and fattening catt1e. J. Animal Sei., 17: 1204.

- 115 -
Fasm.an, G.D. , and Niemann, C.
1951 A reinvestigation of the kinetics of the urease-catalyzed hydrolysis of urea. I. The ac ti vit y of urease in the pre sen ce of sodium and potassium phosphate. J. Am. Cham. Soc., 73: 1646-1650.
Gall, L.s., and Huhtanen, C.N.
1951 Criteria for judging a tru.e rumen organisn and a description of five rumen bacteria. J. Dairy Sei., 34: 353-362.
Gall, L.S., Stark, C.N., and Loosli, J.K.
1947 The isolation and preli.minary stud.y of soma physiological characteristics of the predominating flora fran the rumen of cattle ani sheep. J. Dairy Sei., 30: 891-899.
Garton, G.A.
1951 Observations on the distribution of inorganie phosphorus, soluble calcium and soluble magnesium in the stomach of the sheep. J. Exptl. Biol., 28: 358-368.
Gibbons, R.J., and Doetsch, R.N.
1959 Physiological s~udy of an obligately anaerobie ureol~ic bacterium. J. Bacteriol., 77: 417-428.
Gibbons, R.J., and McCarthy, R.D.
1957 Obligately anaerobie urea-hydrolyzing bacteria in the bovine rumen. Maryland Agr. Expt1. Sta., :Mise. Pub1. No. 291: 12-16.

- 116-
Gray, F.V., Pilgrim, A.F., and Weiler, R.A.
1953 Conversion of plant nitrogen to microbial nitrogen in the rumen of the sheep. Nature, 172: 347-348.
Gunsalus, I.e.
1955 Extraction of enzymes from microorganisns. In Colowick, S.P., and Caplan, N.O. Methods .!n, Enzymologz, Vol. I, Academie Press Ine., New York.
Hamilton, T.S., Robinson, W.B., and Johnson, B.C.
1948 Furthar eomparison of the utilization of nitrogen of urea with that of some feed proteins by sheep. J. Animal Sei., 7: 26-34.
Harris, L.E., and Mitchell, H.H.
1941 The value of urea in the synthesis of protein in the pauneh of the ruminant. I. In maintenance. J. Nutr., 22: 167-182.
Hirseh, A., and Grinsted, E.
1954 Methods for the growth and enumeration of anaerobie spore -formsrs from chee se , with observations on the effect of nisîn. J. Dairy Res., 21: 101-110.
Hobson, P.N.
1961 Technique of eounting rumen microorganisns. In lewis, D., Ed. Digestive P}lysiologz and Nutrition of the Ruminant. Butterwortb;,:Lëndon.
Hobson, P.N.
1962 Personal communication.

- 117-
Hoskins, J.K.
1934 Most probable numbers for evaluation of coli-aerogenes tests by fementation tube method. Public Health Rept. (U.S.), 49: 393.
Houpt, T.R.
1959 Utilization of blood urea in ruminants. Am. J. Physiol., 197: 115-120.
Huhtanen, C.N., and Gall, L.s.
1955 Manametric estimation of rumen urease. J. Bacteriol., 69: 102-103.
Hungata, R.E.
1947 Studies on cellulose fer.mentation. III. The culture and isolation of cellulose-decomposing bacteria from the rumen of cat tle. J. Bacteriol., 53: 631-645.
Jacobsohn, K.P., and Cruz, J.M.
1945 Arch. Port. Sei. Biol., 7: 73-75. Cited in Biol. Abstr., 23: 20447(1949).
Jacoby, M.
1933 Biochem. z., 259: 211-222. Cited in Biol. Abstr., S: 420(1934).
Jennings, D.H., Hooper, D.C., and Rothstein, A.
195S The participation of phosphata in the for.mation of a carrier for the transport of magnesium and manganese ions into yeast cells. J. Gan. Physiol., 41: 1019-1026.

- 118-
Johanson, R., Moir, R.J., and Underwood, E.J.
1949 Sulphur-containing amino-acids in the rumen bacteria of sheep. Nature, 163: 101.
Johns, A.T.
1951 The mechanisms of propionic acid formation by Veil1onella gazogenes. J. Gen. Microbio1., 5: 326-336.
Jurtshuk, P., Doetsch, R.N., and Shaw, J.c.
1958 Anaerobie purine dissimilation by washed suspensions of bovine rumen bacteria. J. Dairy Sei., 41: 190-202.
Kistiakowsky, G.B., and Shaw, W.H.R.
1953 Ureo1ytic activity of urease at pH S.9. J. Am. Chem. Soc., 75: 2751-2754.
Laidler, K.J., and Hoare, J.P.
1949 The mo1ecular kinetics of the urea-urease system.. I. The kinetic 1aws. J. Am. Cham. Soc., 71: 2699-2702.
Larson, A.D., and Kallio, R.E.
1954 Purification and properties of bacterial urease. J. Bacterio1., 6S: 67~71.
lewis, D.
1951 The metabolism of nitrate and nitrite in the sheep. 2. Hydrogen donators in nitrate reduction by rumen microorganisns ia vitro. Biochem. J., 49: 149-153.

- 119-
Lewis, D.
1960 Ammonia toxieity in the ruminant. J. Agr. Sei., 55: lll-117.
Lewis, D., Hill, K.J., and Annison, E.F.
1957 Studies on the portal blood of sheep. 1. AbsorPtion of ammonia from the rumen of sheep. Bioehem. J., 66: 587-592.
lewis, D., and MeDonald, I.W.
1958 The inter-relationships of individual proteins and earbohydrates during fermentation in t:œ rumen of the sheep. I. The fermentation of casein in t:œ presence of starch or other earbohydrate materials. J. Agr. Sei., 51: 108-ll8.
Lister, A.J.
1956 The kineties of urease aetivity in Corypebaeterium renale. J. Gen. Microbiol., 14: 478-484.
Loosli, J. K., and Harris, L.E.
1945 Methionine inereases tœ value of urea for lambs. J. Animal Sei., 4: 435-437.
Loosli, J.K., Williams, H.H., Thomas, W.E. Ferris, F .H., and Maynard, L.A.
1949 Synthesis of amino aeids in the rumen. Science, llO: 144-145.
Lugg, J.W.H.
1949 Plant proteins. Advan. Protein Chem., 5: 229-304.

- 120-
MacKay, E.S.M., and Oxford, A.E.
1954 Soma facultatively anaerobie Gram-negative rods from the runen of the calf and t:œ sheep. J. Gen. Microbio1., 11: 472-476.
Mann, s.o., Masson, F.M., and Oxford, A.E.,
1954 Facultative anaerobie bacteria from the sheep's rumen. J. Gen. Microbiol., 10: 142-149.
Mam, s.o., and Oxford, A.E.
1955 Relationships between viable saccharolytic bacteria in rumen and abomasum of the young calf and kid. J. Gen. Microbio1., 12: 140-146.
Masson, F.M., and Oxford, A.E.
1951 The action of the ciliates of the sheep's rumen upon various water-solub1e carbohydrates, including polysaccharides. J •• Gen. Microbiol., 5: 664-672.
Mateer, J.G., and Marshall, E.K.
1916 The urease content of certain beans, with special reference to the jack-bean. J. Biol. Cham., 25: 297-305.
McDonald, I.w. 1948 The absorption of ammonia from the
rumen of the sheep. Biochem. J., 42: 584-587.
McNaught, M.L., Owen, E.C., Henry, K.M., and Kon, S.K.
1954 The uti1ization of non-protein nitrogen in the bovine rumen. S. The nutritive value of the proteins of preparations of dried rumen bacteria, rumen protozoa and brewer's yeast for rats. Biochem. J., 56: 151-156.

- 121-
M'ystkowski, E.M.
1928 Acta. Biol. Exptl., Varsovie, 2: 211-224.. Cited in Chem. Abstr., 23: 2192(1929).
Otagaki, K.K., Black, A.L., Goss, H., ani Kleiber, M.
1955 In vitro studies with rumen microorganisms using carbon-14.-labeled casein, glutamic acid, leucine and carbonate. Agr. Food Chem., 3: 94.8-951.
Pearson, R.M., and Smith, J.A.B.
1943a The utilization of urea in the bovine rumen. 2. The conversion of urea to a.mrnonia. Biochem. J., 37: 148-153.
Pearson, R.M., and Smith, J.A.B.
1943b The utilization of urea in the bovine rumen. 3. The synthesis and breakdom of protein in the rumen ingesta. Biochem. J., 37: 153-164.
Pinter, T. , Taskovska, D. , and Karas , V.
1954 Biochem. z., 325: 239-245. Cited in Chem. Abstr., 48: 7089(1954).
Pounden, W.D., Fergus on, L.C., and Hibbs, J .W.
1950 The digestion of rumen microorganisms by the host animals. J. Dair,y Sei., 33: 565-572.
Prescott, s.e., Winslow, C.-E.A., and McCrad)!.l M,H~,
1946 Water Bacteriology. 6th ed. Wiley and Sons,
New York.

- 122 -
Qua.stel, J.H.
1933 The action of polyhydric phenols on urease; the influence of thiol compounds. Biochem. J., 27: ll16-1122.
Rook, J.A.F., and Storry, J.E.
1962 Magnesium in tm nutrition of farm. anim.a.ls. Nutr. Abstr. Rev., 32: 1055-1077.
Rothstein, A., and Hayes, A.
1956 The relationship of the cell surface to metabolism. XIII. The cation-binding properties of the yeast cell surface. Arch. Biochem. Biophys., 63: 87-99
Rothstein, A., Hayes, A., Jennings, D.H., and Hooper, D.C.
1958 The active transport of magnesium and manganese ions into the yeast cell. J. Gen. Physiol., 41: 585-594.
Rustigian, R., and Stuart, C.A.
1941 Decomposition of urea by Proteus. Proc. Soc. Exptl. Biol. Med., 47: 108-112.
Rys, R. , Gor ski, L. , and Styczynski, H.
1956 Roczniki Nauk Rolniczych: Ser. B, 70: 573-575. Cited in Nutr. Abstr. Rev., 70: 573(1958).
Shaw, W.H.R.
1954 The inhibition of urease by various metal ions. J. Am. Chem. Soc., 76: 2160-2163.

- 123-
Shaw, W.H.R.
1961 Cation toxicity and the stability of the transition~tal complexes. Nature, 192: 754-755.
Shaw, W.H.R., and Raval, D.N.
1961 The inhibition of urease by metal ions at pH 8.9. J. Am. Chem. Soc. , 83: 3184-3187.
Shibata, K.
1958 Kyoto Furitsu Ika Daigaku Zasshi, 64: 355-364. Cited in Cham. Abstr., 54: 24969(1960).
Sirotnak, F.M., Doetsch, R.N., Brown, R.E., and Shaw, J .c.
1953 Am1no acid metabolism of bovine rumen bacteria. J. Dairy Sei., 36: lll7-1123.
Sizer, I.w.
1941 Temperature activation of the urease-urea system using urease of Proteus vulgaris. J. Bacteriol., 41: 511-527.
Skerman, V .B.D.
1959 A Guide to the Identification of the Genera ofBaCteria. -Williams and Wilkins Co. , Baltimore.
Soejima, M., Sugawara, K., and Shimura, K.
1958 Nippon Nogeikagaku Kaishi, 32: 917-921. Cited in Chem. Sbstr., 53: 18229(1959).
Sugden, B.
1953 The cultivation and metabolism of oligotrich protozoa from the sheep's rumen. J. Gen. Microbiol., 9: 44-53.

- 124..;.
Sumner, J .B.
1926 The isolation and crystallization of the enzyme urease. J. Biol. Cham., 69: 435-441.
Sumner, J .B.
1951 Urease. In Sumner, J.B., and Myrback, K., ed. The Enz;ymes. Vol. I, Ft. 2. Academie Press Inc., New York.
Sumner, J.B., and Kirk, J.s.
1932 Is there a co-enzyme for urease? Biochem. J., 26: 551-554.
Sumner, J .B., and Myrback, K.
1950 Hoppe-Seyler' s Zeitschr. Physiol., 189: 218-228. Cited in Biol. Abstr., 8: 17550(1934).
Takeda., N.
1960 Kyoto Furitsu Ika Daigaku Za.sshi, 67: 759-768. Cited in Cham. Abstr., 55: 1753(1961).
Thomas, W.E., Loosli, J .K., Williams, H.H. and Maynard, L.A.
1951 The uti1ization of inorganic sulfates and urea nitrogen by lambs. J. Nutr., 43: 515-523.
Tsuyuki, H., and Macleod, R.A.
1951 Ion antagonisms affecting glycolysis by bac te rial suspensions. J. Biol. Cham., 190: 711-719.

- 125 -
Varner, J .E.
1960 Urease. In Boyer, P.D., Lardy, H., and Myrbaok, K. The Enz:wnes. Vol. 4. 2nd ed. Academie Press Ino., New York.
Wall, M.C., and Laidler, K.J.
1953 The moleoular kinetios of the urea-urease system. VI. The activation of urease by amino aoids. Arch. Bioohem. Biophys., 43: 312-31S.
Warner, A.C.I.
1961 Personal communication.
Warner, A.C.I.
1962 Enumeration of rumen microorganisms. J. Gen. Miorobiol., 2S: 119-12S.
Wagner, M.I., Booth, A.N., Bohstedt, G., and Hart, E.B.
1941 Preliminary observations on ohemioal changes of rumen ingesta with and without u:rea. J. Dairy Sei., 24: 51-56.
Wilson, E., Foter, M.J., and Lewis, K.H.
1959 A rapid test for deteoting Stapbylooooous aureus in food. Appl. Miorobiol., 7: 22-26.
Wilson, M.K., and Briggs, C.A.E.
1955 The normal flora of the bovine rumen. II. Quantitative baoteriologioal studies. J. Appl. Baoterio1., lB: 294-306.

-126 -
\V'right, D .E.
1960 The metabolism of carbon dioxide by Streptococcus bovis. J. Gen. Microbiol., 22: 713-725.
Yall, I., and Green, M.N.
1952 Sulphydryl variation in bacterial enzymes in relationship to chemotherapy with tm nitro-furans. Proc. Soc. Exptl. Biol. Mad., 79: 306-311.
Zuntz, N.
1891 Pfluegers Arch. Ges. Physiol., 49: 483. Cited by Annison and Lewis (1959).


APPENDIX
Re port of a Preliminary E:xpe riment on the Fa te of
Nl5-labelled Urea in the Rumen of a Sheep

INTRODUCTION
When urea entera the rumen it is rapidly hydrolyzed to
ammonia by rumen bacterial urease. The ammonia produced is then
removed from the rumen solution by absorption into the portal blood
stream, by passage to the abomasum and by incorporation into micro-
bial protein. Broadly speaking, only th at incorporated into the
rumen microorganisms representa nitrogen of use to the animal since
most of the remaining ammonia is excreted as urea-nitrogen by the
kidneys, although soma may be recycled to the rtunen. :Many factors
affect the efficiency with which dietary non-protein nitrogen is
incorporated into microbial protein (Annison and lewis, 1959), among
the most important of these factors being the nature of other comp
onents of the diet.
A survey of the literature indicated that no estimate has
been made of the proportions of ammonia-nitrogen released fran urea
in the rumen which follow the various pathways of removal fran the
rumen solution. It was therefore decided to attempt to assess the
quantitative importance of these pathways in a sheep, using Nl5 as a
marker of dietary urea-nitrogen. The efficiency of utilization of
the urea-Nl5 was to be estimated under a number of experimental
conditions, including the administration of various carbohydrate
energy sources simultaneously with the labelled urea.
It was intended initially that too completion of this
study would be the primary objective of this Ph.D. research pro-

- 130 -
gram. It be came evident during the preJ jmj nary stages of the study,
hawever, that adequate arrangerœnts could not be made to obtain all
the Nl5 analyses required when necessary. For this reason the pro
jected study was abandoned and the program on rumen urease reported
in this thesis was substituted in its stead.
When the original investigation was planned, each experiment
was intended to involve the collection fran the animal of samples
of rumen fluid, saliva and urine over a period of 12 hr. following
the initial introduction of the urea into th3 rumen. The microbial
fraction was to be separated from the rtuiJ.an fluid samples, and total
nitrogen and Nl5 enricèment determinations were to be made on samples
from ali sources. In order to be able to put the re sul ts of tha
determinations made on the microbial fraction of rumen fiuid on an
absolute quantitative basis, measurements of tha liquid volume of
the rumen were to be made during the course of the experimental
period by the method of Hydén (1961), using polyethyleneglycol as a
marker.
As part of the original program, a preliminary experim.ent
was carried out in which labelled urea was introduced into the rumen
of a fistulated sheep which had been fasted during the previous 2 hr.
Determinations of rumen liquid volume and collections of samples from
the animal were made as planned. This prelirninary experiment was
completed and is described in this Appendix. Owing to its prelimin
ary nature, however, it contains a number of sources of error which

- 131-
it might have proved possible to reduce or eliminate had further
experimenta been performad. One of the most serious omissions is
that of the data obtained for changes in the rumen liquid volume
during the course of the experiment; when these data were studied
they did not seem sufficiently reliable to be used to convert the
resulta obtained from the nitrogen analyses of the rumen fluid sam.ples
to a "total rumen fluid volume" basis. The experiment therefore pro
vided lesa information conceming the fate of urea nitrogen added to
the rumen than had been anticipated.
It was, nevertheless, considered desirable to report the
resulta of this preliminary experiment since they might prove to be
of value in suggesting areas in which modifications in technique
would be necessary in the event that tre investigation were continued.

LITERATURE REVIEW
Watson, Davidson and Kennedy (1949) fed Nl5_labelled urea
to sheep over a period of 4 days prier to slaughter. They then
found an N15 excess present in the proteins of the liver, kidneys
and blood of the animals.
An experim.ent in which Nl5_labelled ammonium nitrate
(N1 5H4N"3) was added to tm ration of milking cows was described by
Virtanen and Land (1959). Within 1 hr. an Nl5-excess was detectable
in the milk, and 17 and 25% respectively of tre administered Nl5 was
recovered from the milk during the first 10 and 17 days respectively,
of the experiment. It was shown that the Nl5 was weil distributed
throughout the amine acids of the casein.
These exp:~riments showed that dietary NPN becam.es incorpor
ated into the body proteine of ruminants and the intermediate steps
in this process have now been broadly established. Sane of the
experimental evidence to support the various mechanisms involved was
described in the General Literature Review.
Several studies, which have already been referred to, have
shown that urea is rapidly hydrolyzed to ammonia as a result of the
high urease activity of the rumen bacteria. The major pathway of
significance to tm nitrogen nutrition of the animal which is sub
sequently followed by the released ammonia is that of its incorporation

- 133 -
into rumen microbial protein. N15-labelled NPN has been used by
several workers to confirm the occurrence of protein synthesis in
the rumen. Sugawara, Soejima and Sagisaka (1958), for example, fed
Nl5-labelled urea to a goat, and collected and fractionated rumen
contents from it at 1, 4 and 7.5 hr. intervals after administration.
The leval of ammonia in the rumen supernatant liquor was maximum in
the 1 hr. sa.mple, and an Nl5 excess was found in both the bacterial
and protozoal fractions from ail sa.mples. Warner (1961) demonstrated
that in an in vitro system comprised of sheep rumen liquor, a protein
substrats and Nl5_ammonia, the Nl5 was taken up into the proteins of
the bacteria and, to a lasser extent, of the protozoa. When the total
microbial nitrogen was fractionated into its components, Warner found
the N15 in almost equal concentration in all the amino acids tested,
including lysine and the dibasic amino acids, and in the amide
nitrogen.
The microbial protein into which rumen ammonia is incorpor
ated subsequently passes to the abomasum and small intestine where it
is digested by proteases of animal origin and from where the degrad
ation products are absorbed. Waller, Gray and Pilgrim (1958), using
diaminopimelic acid as a marker of bacterial protein, showed 63 - 82%
of the total nitrogen in the rumen to be microbial nitrogen, 11 - 27%
to be plant nitrogen and 5 - 10% soluble nitrogen. These results
give an indication of the extent of conversion of rumen nitrogen
components to microbial protein, and of the importance of this process

·134-
to tha nitrogen nutrition of the animal.
Sana of the amn.onia-nitrogen released from urea in the rumen
is largely lost to the animal by excretion as exogenous urinary nitro
gen, and nitrogen retention is primarily governed by the efficiency
with which the ammonia is incorporated into m.icrobial protein. This
in turn depends on such factors as the amount of NPN present in tha
anima] 's feed, the rate of release of ammonia from the NPN (or protein)
source and the availability of carbohydrate source. Oyaert and Bouckaert
(1960), using a sheep equipped with an omasal canula, found a highly
significant negative correlation between levels of rumen aamonia and
tœ proportion of this ammonia which was recoverable at the omasal
abomasal orifice. ApparentJ..y, wh3re the level of rumen ammonia was
high, a large proportion was absorbed directly fran the rumen.
Belasco (1954) :round that am.monia was released more slowly
!!! vitro from the amides of the monocarbo:x:ylic ac ids than from ure a,
bence these compounds gave rise to lower concentrations of ammonia
and promoted greater rates of nitrogen utilization by the rumen
microorganisms than did urea. Hendrick (1960) , in !!! vitro e:xperi
ments, found that am.monia was released from ammonium succinate
sufficiently slowly that S5% of the nitrogen released was incorporated
into the protein of rumen microorganisme whereas the e.xtent of incor
poration of an equivalent amount of urea-nitrogen was only 46%.
,!a ~ studies by severa! workers have demonstrated tha
importance of the type and concentration of carbohydrate available to

- 135 -
the rumen microflora in relation to the efficiency of urea utilization.
Protein synthesis from urea was shown to be stimulated in the presence
of soluble sugars (We gner, Booth, Bohstedt and Hart, 1940; Arias,
Burroughs, Gerlaugh and Bethke, 1951) and the special value of starch
as an energy substrate in this respect was ahown by Mills, Booth,
Bohstedt and Hart (1942). Belasco (1956) obtained 22% urea utilization
in the presence of starch, and found that the addition of cellulose to
the system increased utilization to 61%.
Despite the fact that urea has been widely used as a protein
replacement in ruminant rations, very little information is available
concerning the efficiency of its utilization in vivo. Evidence of a ---relatively efficient utilization under suitable conditions haa been
obtained from in vitro studies, but it is difficult to translate the
resulta of these to an !g_ vivo situation. No report was found in
the literature of any attempt to estimate the quantitative importance
of the varioua pathways which may be follollled by ammonia released from
dietary urea in the rumen.

KATERIALS AND JŒTHOOO
A. ExPerimental Animal.
Tœ animal used in this study was sheep 2, the diet and
management of which were described in Part I. It was fed the usual
ration at S a.m. on the day of the experiment and at 12 noon both feed
and water were withdravm. The animal was fasted for 2 hr. prior to
commencement of the experiment and during the succeeding 12 hr.
15 B. N -urea.
N15-urea (Isomet Corp., CO(Nl5H2)2) containing 98.7 atoms%
Nl5 was used as a source of labelled urea-nitrogen. The amount
administered to the animal was 0.50 g., calculated to contain 15.98 mg.
atoms of Nl5. It was dissolved in a solution of 10 g. of polyethylene
glycol (PEG; M.W. • 4000) in 500 ml. of water at 39°0, and at the
beginning of the experiment this solution was introduced into the rumen
of the animal through the fistula. The flask which had contained the
solution was rinsed with 100 m.1. o:! water and the rinsings were a1so
poured into the rumen. The PEG was added as part of the procedure
involved in carrying out the determinations of rumen fluid volume.
It was dis sol ved in 500 ml. of water in order to achieve a rapid
distribution of PEG in the rumen solution (Hydén, 1961). Further
additions of PEG solutions to the rumen 'Were made during the course
of tœ experim.ent as follows: 5 g. of PEG in 300 ml. of water at
each of the 4th and Sth hours. At the 7th hour 1000 ml. of water

- 137-
were added to the rumen because at that time it was found difficult
to obtain rumen fluid samples of adequate volume to enable the necessary
analyses to be carried out.
C. Collection and Preparation of Samples.
Immediately before addition of the N15-urea solution to the
rumen, and at i-hr. intervals during the ensuing 12 hr. period, sam.ples
of rumen contents, saliva and urine were taken from the animal. The
procedures de scribed below were carried out in the barn and the
resulting preparations were stored at -17°0 for later analysis.
a) Rumen contents
Rumen contents were collected and strained as described in
Part I. A 55 ml. volume of the strained rumen f'luid was retained and
any excess was immediately returned to the rumen. The pH of the fiuid
was measured with a Beckman modal N pH meter. The fiuid was then
centrif'uged at 11,500 x G for 10 min. at room temperature. The rumen
supernatant liquor was decanted and retained, an:i the cella were re
suspended in 55 ml. ot 0.9% NaCl solution.
b) Saliva
At each sampling time 2 - 3 ml. saliva wre collected by
inaerting a curved metal blowpipe tube, eonnected via a small test
tube to a vacuum pump, into the animal's mouth. The end of the
blowpipe was wetted, dipped into crystalline NaCl, and inserted into
the mouth of the animal. Saliva was withdrawn under slight vacuum.

- 138-
The samples were obtained with little dii'ficulty since the presence in
the mouth of salt, coupled with chewing by the animal on the blowpipe,
appeared to encourage salivation.
c) Urine
The metabolism crate in which the sheep was maintained was
equipped for the collection of' urine. The animal conveniently urinated
at the time each sample of rumen contents was taken. At hali'-hourly
intervals, therei'ore, a volume of urine was obtained which was taken
to represent the entire volume excreted by the kidneys during the
previous 1 hr. On three occasions during the 12 hr. period of' the
experiment, however, no urina.tion occurred and no collection could be
made. Following the tennination of the experimental period collections
of urine were made at arbitrary intervals during a 90-hr. period. The
volume of' each urine sample colle cted was measured.
D. Estimation of Nitrogen in Samples.
a) Ammonia in rumen supernatant liquor
The a.mmonia content o:f the rumen supernatant l.iquor sampl.es
was estimated by a method invol.ving Fol.in aeration of the ammonia
into standard acid.
A 10 ml. volume of' rumen supernatant liquor was made alkaline
by the addition of 5 ml. of' saturated K2C~ solution. A drop of' Dow
Antii'oam A was added and the anmonia was aerated in a Van Slyke-Cullen
urea train (Van Slyke and Cullen, 1914) for 1 hr. into standard HZ304.

- 139 -
The excess acid was then titrated against standard alkali.
b) Estimation ot total nitrogsn
The total nitrogen content of rumen supernatant liquor,
microbial fraction, saliva and urine samples was determined in duplicata
by the semi-micro-Kjeldahl method. The general procedure used was as
follows. A suitable volume (3 or 5 ml.) of the thawed sample was
transferred to a 30 ml. Kjeldahl flask and 0.5 ~· of K~04, 0.2 ~·
of HgO and 1 Hengar Se granule were added. This was followed by the
addition of 3 ml. of conc. H2so4• The mixture was heated gsntly on
a digestion rack until frothing ceased; tœ temperature was then
increased until the mixture boiled vigoroualy and digestion was
continued for 8 hr. The flask and its contents were then cooled and
5 ml. ot water were added. The diluted digest was tranaferred to a
micro-Kjeldahl distillation apparatus (Scientitic Glass Apparatua Co.,
Cat. No. JM-4250) and made alkaline by tœ addition of 2) ml. of a
aolut ion containing 50% NaOH and 5% Na~ 'iJ3. 5H~ ( w/v). Tba a.mm.onia
was distilled into standard acid and the exeess acid titrated against
standard a.lkal.i.
One ot the duplicate distillates was aciditied by the addition
of 1 ml. ot 0.2N-H2904 to prevent losa of ammonia. It was filtered
and then evaporated to dryness over a steam bath. The residue was
taken up in 2 ml. ot acidified distilled water and stored at roan
temperature to await N15 analysis.

- 140 -
The initial procedures f'ollowed with tœ va.rious samplas
"Were:
i) Rumen superna.tant liquor
Five ml. volumes of' supernatant were used directly for
digestion.
ii) Rumen microbial fraction
Af'ter thawing, suspensions of the microbial fraction in
0.9% NaCl were centrifuged. The supernatant liquid was decanted,
the cells resuspended in 0.9% NaCl and the suspension diluted to
40 ml. A 5 ml. volume of each suspension was used for digestion.
iii) Saliva
Saliva samples vere f'iltered to remove food particles and
mucus. The entire volume of each filtrate, which was not measured
but which was genera.lly less than 2 ml., was used for a single
detenn.ina.tion of total nitrogen content. The amount of nitrogen
subsequently found to be pre sent in each sample was of the order
0.5 - 1.0 mg. and since this was insufficient for Nl5 analysis, 1 ml.
of an (NH4)2so4 solution conta.ining 1.5 mg. of nitrogen per ml. was
a.dded to each neutral distillate bef'ore evaporation.
iv) Urine
Urine samples -were filtered and a 3 ml. volume of the
filtrats wa.s used t'or digestion.

- 141-
E. N15 Analyses.
a) Oxidation of runmonia-nitrogen to N2.
The 1abelled amnorrla-nitrogen, which was present in each
sample as (NHJ)2so4 was oxidized to N2 with alkal.ine NaOBr. This
reagent was prepared by the method of Sprinson and Rittenberg (1949).
50 ml. of Br2 was slowly added with 'Vigorous stirring to 150 ml. of
40% NaOH (w/v) in an ice bath. After the addition of a further
150 ml. of NaOH solution, the reagent was allowed to stand in a
polythene container in the refrigerator for several days. Before use
it wa.s decanted and centrifuged to remove precipitated NaBr.
The procedure used in the oxidations was a modification of
the method of Rittenberg (San Pietro, 1957), and has been described
by Laishley (1961). The labelled ammonium sulpha.te samp1e in 2 - 3
ml. of water was added to one bulb of the gas conversion a.ssembly and
5 ml. of alkaline NaOBr, diluted with an equa.l volume of water, was
added to the other. The contents of the bulbs were frozen by immersion
in 1iquid air and the assemb1y was evacua.ted to a pressure equivalent
to 10 p Hg. The contents of the two bulbs were them thawed and mixed
in vacuo; a violent reaction occurred and N2 gas wa.s libera.ted.
b) Determination of N14JN15 ratio
After o:xidation was complete, the gas conversion a.ssembly
was attached to the manifold of a Consolidated-Nier direct reading
mass spectraneter, and its contents were again frozen by immersion

- 142-
in liquid air. The gas sample was then introduced into the spectro
meter and the N14fN15 ratio in the sample was measured. From this
ratio, the atome % excess Nl5 in the sample was calculated. The
standard nitrogen gas sample used for comparison contained 0.366
atom% N15.
In the case of the saliva samples, corrections 'Were made
in the calculationa of enrichment for the dilution effect of the
added unlabelled (NH4)2so4
•

RESULTS
In this preliminary experiment, 0.5 g. of Nl5_u.rea con
taining 15.98 .mg. atoms of N15 was introduced into the rumen of a
fasting sheep. Changes in the pH of the rumen solution and in the
concentrations, and levels of N15 enrichment, of various nitrogen
components in tœ rumen fluid, the saliva and the urine of the
animal were then followed during the succeeding 12 hr. period.
A. Rumen F1uid.
a) pH of rumen fiuid
Changes in the pH of the rumen f1uid of the animal following
administration of the N15-u.rea are shown in Appendix Figure 1. These
chan~s did not ref'1ect the presence in the rumen of the u.rea or of tœ
amnonia to which it would be rapidly converted by hydrolysis. On the
contrary, during tœ first 7 - 8 hr. of the experim.ent, the pH of the
rumen fluid gradually decreased. This affect was probab1y caused by
the production of volatile fatty acids by fermentation of food material
present in the rumen at the commencement of the experiment. The sub
sequent absorption of these fatty acids may then explain the fact that
the pH of the fluid rose again to its initial level.
b) A.mmonia content of rumen supernatant liquor
Changes in the ammonia content of rumen supernatant llquor
during the experimental period are shown in Appendix Figure 2. These
represent variations in the arnmonia content of the rwnen solution.
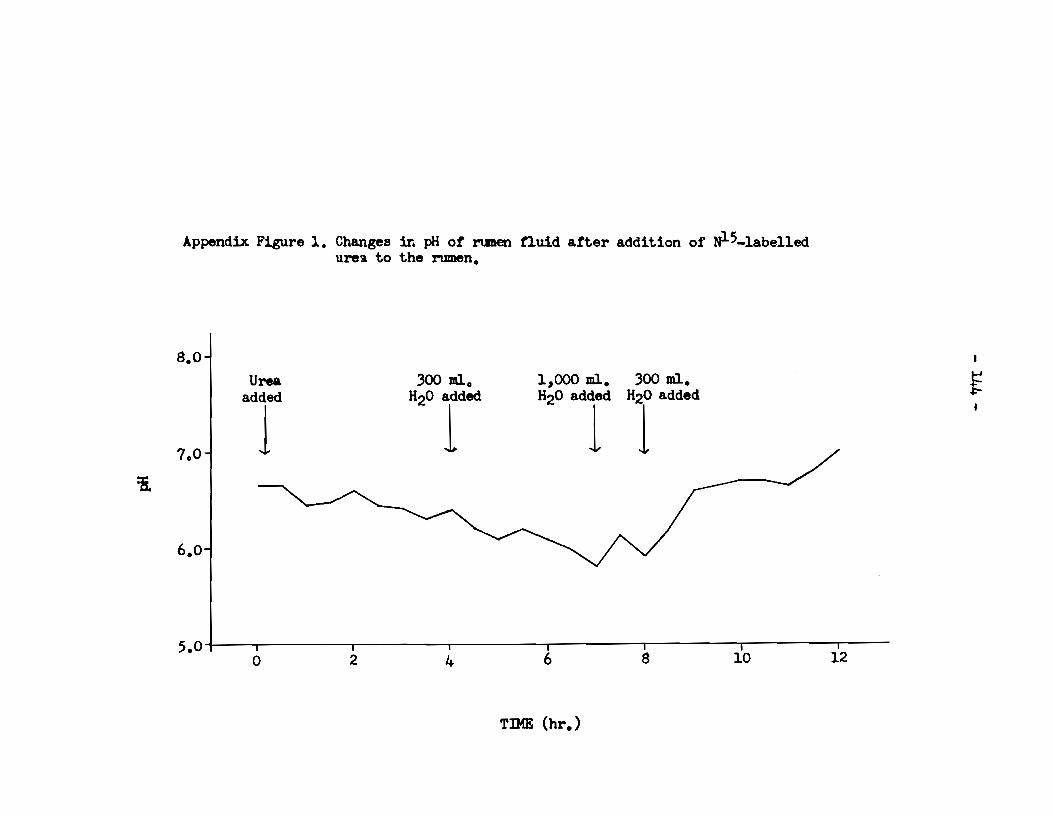
=a
Appendix Figure 1. Changes ir. pH of rumen fiuid arter addition of ~5-1abe11ed urea to the rumen.
s.o 1
Urea 300ml. 1,000 ml. 300ml. added H20 red H20 adr H20 added
7 .o-1 l 1
6.0
5.0 6 8 10 12 0 2 4
TIME (hr.)
f;
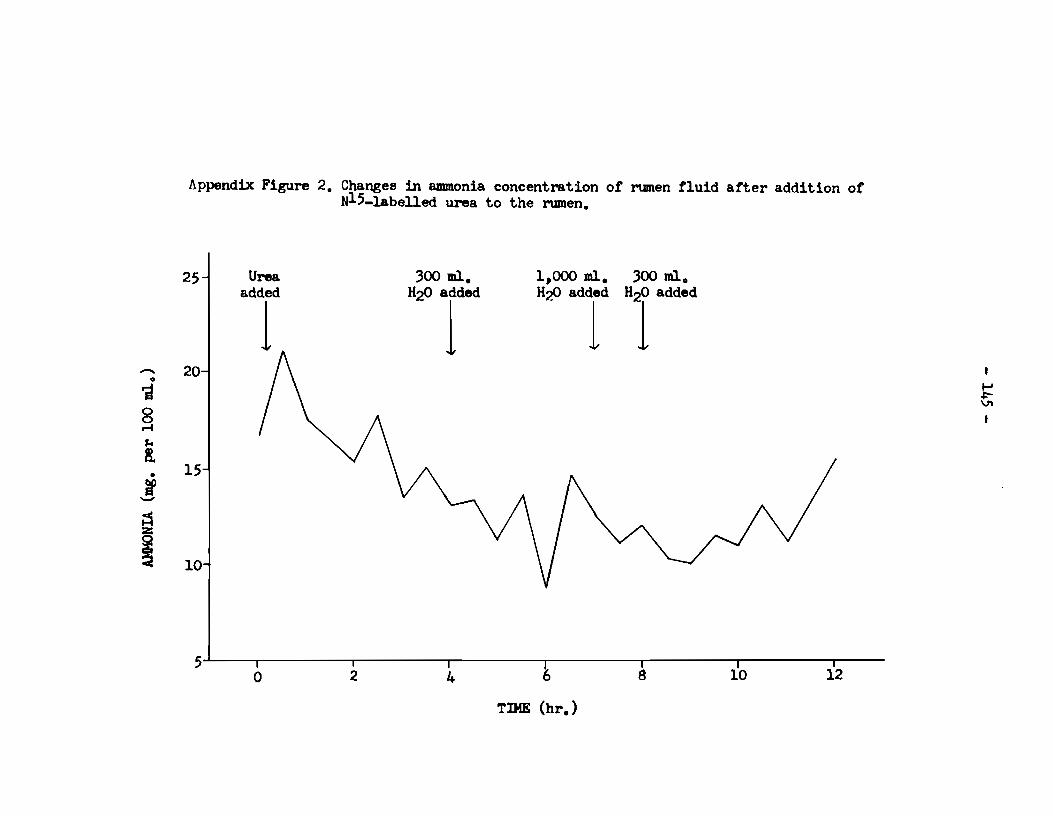
-0
~ 8 ,..... ,.. !. • r -~ :z:
1
Appendix Figure 2. Changes in ammonia concentration ot rumen f'1uid af'ter addition ot N15-labelled urea to the rumen.
25-l Urea 300ml. 1,000 ml. 300 ml. added H~r~ H20 added H20 added
lA - 1 l
1 20
1 1 \
15
10
5 6 8 w u 0 2 4
TIME (hr.)
1-' ~ VI

- 146-
The resulta show that the rumen ammonia concentration fluctuated
considerably throughout the experimental period. Wlthin the first
i hr. tba runen ammonia concentration rose to a level 25% greater
than its initial level, showing that the urea added to the rumen was
rapidly hydrolyzect · to ammonia. Thereafter, despite the fluctuations,
there was a trend towards a lower concentration during the following
7 - 8 hr., and then towards an increased concentration again during
the next 4 hr. period, by the end of which the concentration had
regained its initial levaL.
c) Total nitrogen content of rumen supernatant liquor and
its enricèment with Nl5.
The total nitrogen content of the rumen supernatant liquor
during the experimental period is shown in Appendix Figure 3. The
resulta represent the concentration of soluble nitrogen found in the
rumen fluid at each sampling time. The figure also shows the levels
of enrichment of this nitrogen fraction with Nl5. The degree of
enrichment is e:xpressed as atoms % excess N15 ovar the normal abundance
of 0.366 atom.s %. The results show that despite the fluctuations in
ammonia concentration of the rumen fluid which occurred during the
experimental period, and which were noted above, the soluble nitrogen
concentration remained relatively constant. There was a rapid decrease
following the 7th hour, when 1000 ml. of water Viere added to the rumen,
but apart from this the decrease in concentration during tha first 8 hr.
l'ms relatively small. It was followed by a slight increase in
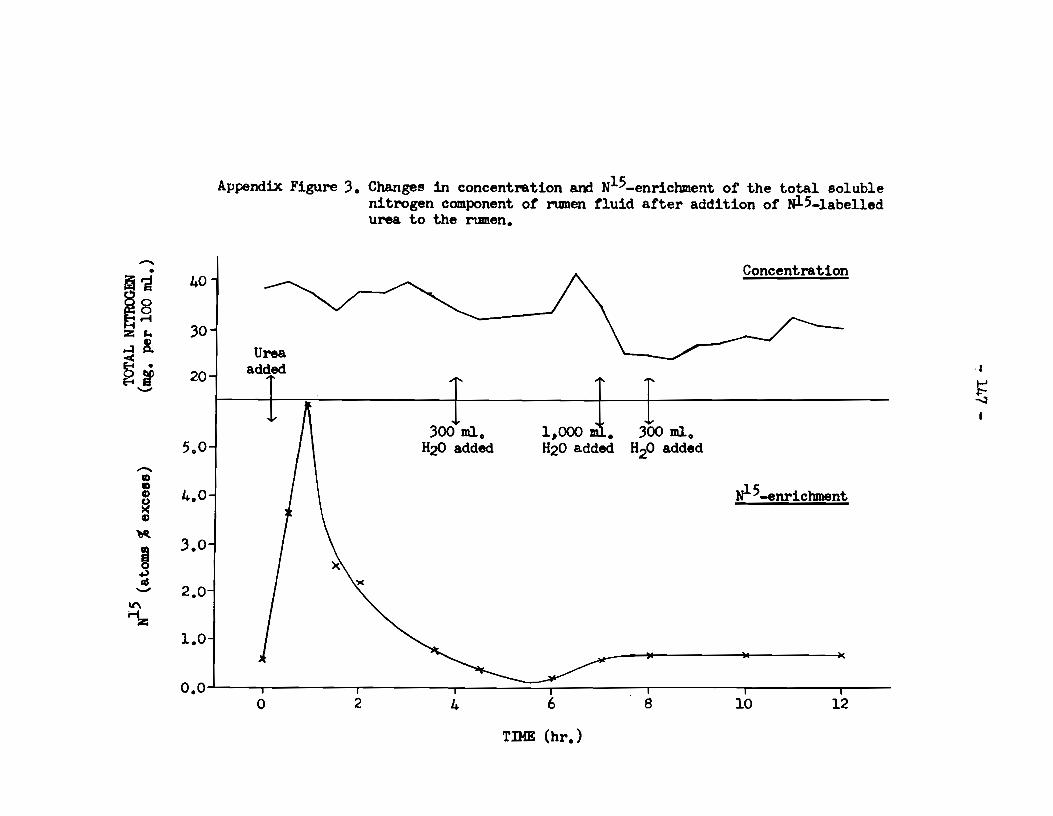
-•
~~ 40
:Z:J.. ~8. g . e-.r -
-Il) Il)
~ 'bit
J ~ 2.0
U'\
~ 1.0
Appendix Figure 3. Changes in concentration and Nl5_enrichment of the total soluble nitrogen camponent ot rumen tluid after addition ot Nl5-labelled urea to the rumen.
Urea added
300ml. H2Ûadded
1,000 ml. 300 ml. H20 added H2D added
Concentration
r5-enriehment
0.0~---.-------.--------r---~==~-------.-------.~------~---0 2 4 6
TIHE (hr.)
B 10 12
·1
~ 1

- 14$ -
concentration during the final 4 hrs. of the experimental period.
The level of enrichment of this nitrogen fraction rose very
sharply to 6 a toms % excess Nl5 within the first ! hr. after adminis
tration of the labelled urea. Thereafter tœ enrichm.ent decreased,
at first sharply and then more slawly. The initial sharp rise may be
attributed to the addition of a relatively large amount of Nl5-urea
to the rumen and its subsequent solution in the rumen fluid. The
rapid fall in enrichment indicates either tha.t the N15 -a.mmonia formed
by hydrolysis of the urea wa.s selecti vely absorbed fran the rumen
solution or that th9 added Nl5 wa.s rapidly diluted by unlabelled
nitrogen, which might have entered the rumen directly or indirectly
from the bloodstream. The former is probably the more likely poss
ibility, as evidencErl by the previously noted fall in tm ammonia
concentration in the rumen solution, which coincided with the decrease
in enrichment. Between the 5th and ?th hours of the experimental
period there was an increase in the enrichment of soluble nitrogen
with r5. This was probably caused by a sudden increase in tm amount
of Nl5 entering the rumen by way of the sali va or the rumen wall, since
there was a small increase in the total nitrogen content of the rumen
supernatant liquor during the sa.rœ period. A relatively steady state
in the leval of enrichment of soluble rumen nitrogen with Nl5 appeared
to be reached at the 7th hour. This indicates that Nl5 entering the
rumen was then balanced by that being removed.

- 149-
d) Total nitrogen content of the microbial fraction
of rumen fluid and its enricbnent with r5 Appendix Figure 4 shows tha total amount of microbial nitrogen
present in the rumen fluid samples collected during the experimental
period. It also shows the levels of enrichrnent of microbial nitrogen
with Nl5. Since the samples of the microbial fraction upon which
the determinations were made had been washed with saline during prep
aration, it was thought unlikely that there was a significant contamination
of the fractions with soluble nitrogen from the rumen fluid, and it was
therefore concluded that the nitrogen found in the samplas represented
incorporated nitrogen. However, the microbial fraction was frozen for
storage and later thawed and this may have resulted in sane degree of
cell lysis. Seme microbial nitrogen may therefore have been lost
bef ore the analyses were made. Apart from de cre ases whieh were brought
about by the addition of water to the rumen, the leval of microbial
nitrogen in the rumen fluid rose gradually throughout the experimental
period. This could be attributed either to the occurrence of micro
bial synthesis or to the absorption of water from tha rumen.
The lsv'els of enrichm.ent of microbial nitrogen with N15
indieate that a significant degree of incorporation of Nl5 into the
microbial nitrogen occurred, at least during tha first 8 hr. after
administration of the urea,. and thus they support the conclusion that
the utilization of ure a by the ruminant animal involves, at least in
part, the incorporation of urea-nitrogen into rumen microbial nitrogen.
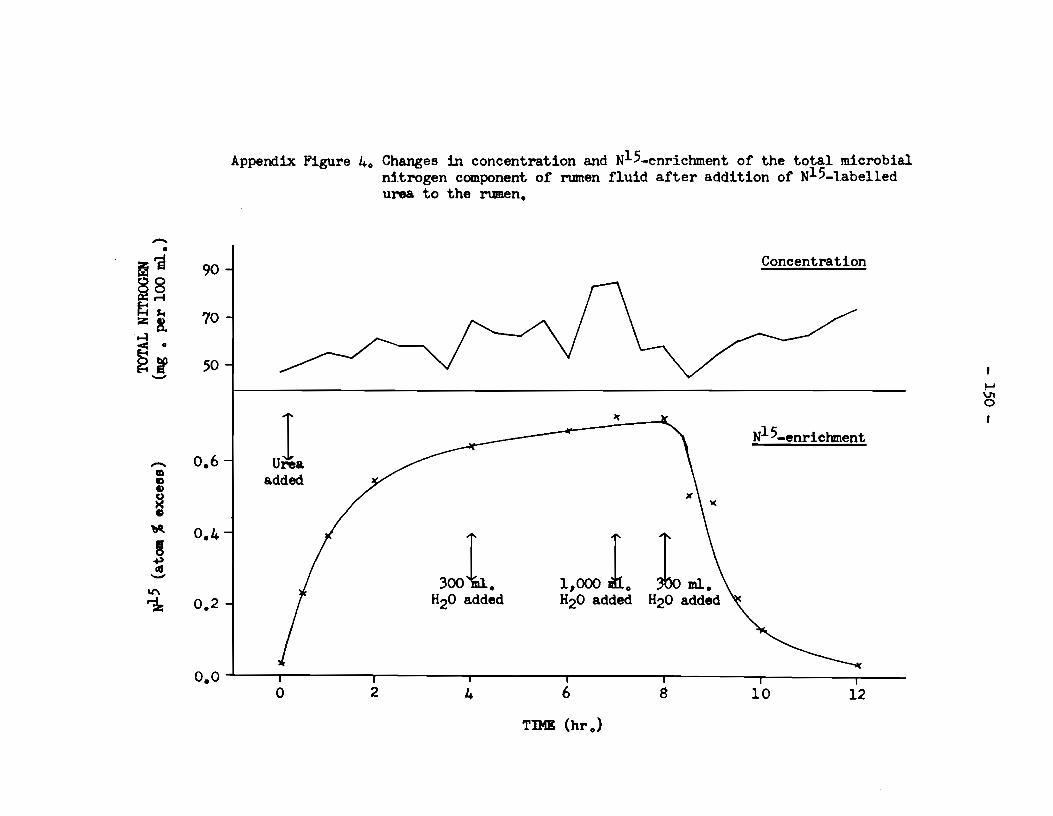
-• i:i'if ~§ ~~ :z:s. ·~
~ . ~r ._,
-ID .... .,
= 'tslt.
8 +) cd -
1.1'\
~
Appendix Figure 4. Changes in concentration and Nl5-enrichment of the total microbial nitrogen eom.ponent of rumen fluid after addition of Nl5-l.abelled urea to the rumen.
90 1 Concentration
70
50
ul loC
N15-enrichment
o.6 Î added
0.4] 1 300l. 1,000 l. lml.
0.2 H20 added H20 added H20 added
0.0~----r---------,----------r----------r----------r---------,----------~----
0 2 4 6
TlME (hr.)
8 10 12
!-' V1 0
'

- 151 -
The rate of incorporation of Nl5 was initially quite rapid
until at the 8th hour the leval of enriehment of rumen mierobial
nitrogen was O. 75 atom % excess Nl5. The level of enrieh:nent then
feil dramatically to 0.1 atom % exœss Nl5 within 2 hr. This is hard
to explain. There was no si.multaneous increase in the enrichment of
the soluble rumen nitrogen fraction, however, and the possibility that
tœ incorporated N15 was released from the rumen microorganisms by
exchange or by cali lysis may therefore be eli.minated. Since the
reduction in enrichment immediately followed the addition of 1000 ml.
of water the rumen, it is possible that this water rapidly flowad
from the rumen to the omasum. and carried with it a considerable
proportion of the rumen microbial fraction. However, in the absence of
rapid synthesis of new material in the rumen from nitrogen of low
enrichment, this would not result in a decrease in the Nl5 /Nl4 ratio
in microbial nitrogen. There was on the other hand, no si.multaneous
reduction in the level of microbial nitrogen in the rumen fluid.
No satisfactor.v explanation for the sudden drop in the level of
enrichment of the microbial nitrogen fraction with ~5 can therefore
be advaneed.
B. Saliva: Nl5 enrichment.
Changes in tœ leval of enrichment with Nl5 which were
found in the sam.ples of saliva collected during the experimental
period are shown in Appendix Figure 5. A significant degree of
enrichment of the saliva-nitrogen occurred within the first à hr.
after addition of the labelled urea to the rumen, but the peak leval
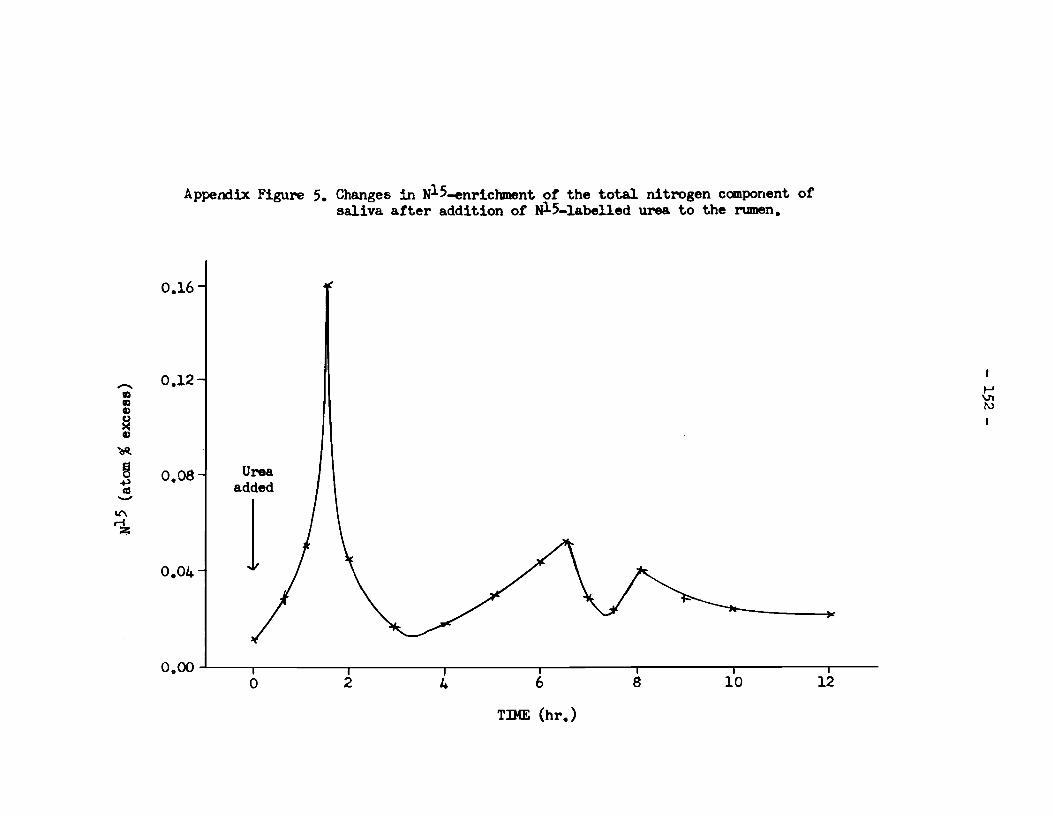
-ID ID Q)
~ Q)
~
e ~ cd -
"' ,.-1 ~
Appendix Figure 5. Changes in N15-enrichment of the total nitrogen canponent of saliva after addition of Nl5-1abelled urea to the rumen.
0.16
O.l2l Il
o.œl Urea added
1 o.o4....J
0.00~----.---------~r----------.-----------.----------,-----------.----------,------
0 2 4 6
TIME (hr.)
8 10 12
1
..... V'l 1\)
1

- 15.3 -
of enrichnent was not attained for a furthar 1 hr. It was thus
reached à hr. after the peak level of enrichment found in t:œ soluble
nitrogen fraction of rumen fiuid. Following the peak level tœre
was a rapid reduction in the enrichment of saliva-nitrogen with Nl5,
but a significant excess was present throughout t:œ experim~ntal
period. This indicates that N15 was continually entering tœ rumen
in saliva and tharefore, by implication, also entering and leaving
the rumen by diffusion through the rumen wall; tha resulta therefore
confirm that seme nitrogen absorbed from the rumen is recycled to that
organ xia the saliva.
C. Urine: Excretion of Nl5 in Urine.
At each sampling time a m.easured volume of urine was
collected fran the animal and the total nitrogen content of the
sample was then determined. Subsequently the Nl5-enrichment of
this nitrogen component was measured. It was therefore possible to
calculate the total amount of N15 lost to tœ animal by excretion in
the urine at each sampling time. The resulta of these calculations
from samples obtained during the 12 hr. experimental period are shown
in Appendix Figure 6, where the cumulative quantity of Nl5 excreted
is expressed as a percentage of the Nl5 originally added to the rumen
as urea. The resulta show that there was an almost linear increase
wi th time in the total amount of N15 excreted by the animal between
the lst and 12th hours of the experiment. By the 12th hour after
administration of the urea, 16% of the added N15 had been excreted.
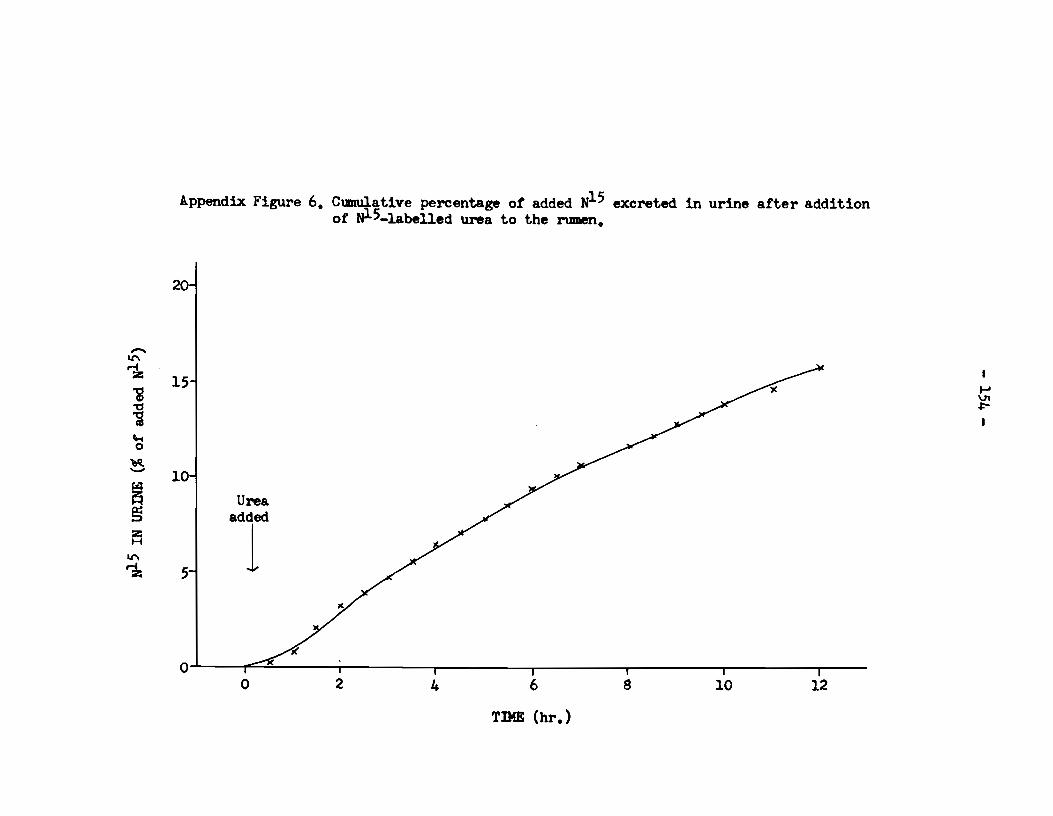
-Il'\ ~ i ~
al '-t 0
~ -~ § :z: H
Il'\
~
Appendix Figure 6. Cum.u,J.ative percentage of added W.5 exereted in urine after addition ot uJ-5-labelled urea to the rumen.
21
15
10
J Urea.
added
1
0 '"' 0 2 4 6
TIME (hr.)
s
~-
10 12
.......
"' ~ 1

- 155 -
S:imilar calculations were made on samples of urine
collected at the 30th, 78th and 102nd hours after administration
of the urea. The proportions of tœ added Nl5 which had been
excreted by these times were, respectively, 26%, 52% and 62%. By
the 102nd hour the degree of enrichment of urine-nitrogen with r5
had decreased to 0.03 atom % excess, suggesting that at this time
there was an almost insignificant excretion of N15.

DISCUSSION
The Prelimina.ry nature of tœ e.xperiment described, and the
reasons for some deficiencies in the resulta obtained from it, were
discussed in the Introduction. The chief factor limiting the value of
the resulta was the absence of reliable data for the changes in the
fluid volume of the rumen during the course of tœ e:xperiment which
could be used to convert the resulta obtained from the nitrogen analyses
to a "total rumen fluid volume" basis. Apparently the polyethylene glycol
solutions added to tha rumen did not become adequately mixed with the
rumen fluid, am subsequently measurements of the mG concentration in
rumen fluid samples gave an incorrect indication of the PEG distribution
volume and therefore also of the volume of liquid pre sent in the rumen.
For this reason, the resulta of thase determinations were discarded
and no attempt was made to put the resulta of the Nl5 analyses on an
absolute quantitative basie. Only the most general conclusions can
thus be drawn from tha resulta presented.
Within a very short time following the addition of labelled
ure a to the rumen of the sheep, all the nitrogen components analysed
were found to be enriched with N15 and furthermore, significant levels
of r5 enrichment occurred in them for at !east the succeeding 12 hr.
period. The enrichm.ent with Nl5 was found in the nitrogen of the rumen
microorganisme, and the sali va and urine of the animal. The re sul ts
therefore provide evidence in support of tha existence of some of the
pathways of nitrogen metabolism in the ruminant animal which have been

- 157 -
elucidated by various workers and which were illustrated in Figure A.
As already mentioned, the most important factor in the
utilization of dietary NPN by ruminants is the efficiency with which
it is incorp::>rated into microbial protein. Although resulta of the
present investigation clearly show that some labelled urea-nitrogen
was incorporated into the rumen microorganisms, they do not indicate
whether this occurred during synthesis of new bacterial protein or
whether the incorporation re sulted fran exchange between extracellular
Nl5 and intracellular nitrogen in the absence of synthesis. Nor do
the resulta indicate the proportion of added Nl5 which was utilized in
this way. An indication of the efficiency wi th 'Which tha animal utilized
added Ure!Hlitrogen can be obtained, however, from a consideration of
the amount of N15 excreted in tha urine. When 0.5 gm. urea was added
to the rumen the proportion of urea-nitrogen retained by the animal
within 12 hr. was 84%. By the 102nd hour after administration of the
urea, whan tha animal was again eating and drinking no:rmally, 62% of
the added N15 had been excreted in the urine. With respect to urine-
nitrogen excretion it is not possible to distinguish that of exogenous
origin although it is only this which representa a loss, through
inefficient utilization, of dietary nitrogen. Same nitrogen may also
be lost in the feces through inefficient absorption from the alimentary
tract, but no account was taken of fecal nitrogen in the present
investigation.
The figures obtained for urinary labelled nitrogen excretion
would undoubted.ly be subject to considerable variation if changes were

- 158 -
made in the experimental conditions. Nitrogen retention m.ight be
increased, for e.xampla, if a carbohydrate source such as glucose had
been added to the rumen at the same time as the urea, since this would
probably resulta in a greater incorporation of nitrogen into m.icrobial
protein. Retention might be reduced if a larger am.ount of urea were
added, since the rapid release of larger auantities of ammonia into
the rumen solution would result in more extensive absorption and
greater exogenous nitrogen excretion (Oyaert and Bouckaert, 1960).
This preliminary experiment has provided resulta which,
though requiring corroboration, would serve as a valuable starting
point for further, more extensive investigations on the fate of
Nl5_labelled urea in the rumen of the sbeep.

SlJIB[ARY
A preliminary experiment was carried out on the fate of
N15-labelled urea in the rumen of a sheep. Nl5-labelled urea was
introduced into the rumen of a fasting sheep, and changes in the pH
of the rumen solution and in the concentrations, and levels of ~5
enricbnent, of various nitrogen components in the rumen fluid, the
saliva and the urine of the animal -were followed during the succeed
ing 12 hr. period. Determinations of rumen volume were made during
the course of the experiment using polyethyleneglycol as a marker,
but the resulta were not reliable and were not reported.
The pH of tm rumen solution decreased gradually during the
first 7 - 8 hr. of the experiment and then increased again. The
ammonia concentration of the rumen solution showed considerable
fluctuations, but tended also to decrease to the 8th hour and then to
increase. The soluble nitrogen concentration of rumen fluid remained
relatively constant throughout the experimental period, but the level
of m.icrobial nitrogen gradually increasad.
Within i hr. following addition of the labelled urea to
the rumen and for the remainder of the experimental period, all the
nitrogen components analysed -were found to be enriched with r 5•
Generally the level of enrichment rose sharply within the first
i - 1 hr. and then decreased more slowly during the remainder of the
period. The leval of enrichment of m.icrobial nitrogen, ho-wever, rose
to o. 75 atom % excess Nl5 by the 8th hour after administration of the

- 160-
urea and than .feil dramatically to 0.1 atom % excess within 2 hr.
O.f t:œ total amount o.f Nl5 added to the rumen, 16% was
excreted in the urine o.f the animal by the 12th hour a.fter adminis
tration, and by the 102nd hour 62% had been excreted.
The resulta of this preliminary experiment provided
evidence in support o.f soma of the known pathways of nitrogen
metabolism. in the ruminant animal, and would be of value in planning
a more extensive investigation on the fate of urea-nitrogen in the
rumen.

BIBLIOGRAPHY
Annison, E.F. , and Lewis, D.
1959 Metabolisn in the Rumen. Metheun and. Co :-;-London.
Arias, C., Burroughs, w., Gerlaugh, P., and Bethke, R.M.
1951 The influence of different amounts and sources of energy upon in vitro urea utilization by rumen microorganisms. J. Animal Sei., 10: 683-692.
Belasco, I.J.
1954 New nitrogen feed compounds for ruminantsa laboratory evaluation. J. Animal Sei., 13: 601-610.
Belasco, I.J.
1956 The role of carbohydrates in urea utilization, cellulose digestion and fatty acid metabolism. J. Animal Sei. , 15: 496-508.
Hendrick, H.
1960 z. Tierphysiol. Tierernaehr. Futtermittelk., 15: 218-227. Cited in Cham. Abstr., 55: 4705(1961).
Hydén, S.
1961 Determination of the am.ount of fluid in the reticule-rumen of the sheep and its rate of passage to the omasum. Kungl. La.ntbrukshogskolans Ann., 27: 51-79.
La.ishley, E.J.
1961 Non-symbiotic nitrogen fixation studies in woodland soils. M.Sc. Thesis. McGill University.

- 162 -
Mills, R.C., Booth, A.N., Bohstedt, G., and Hart, E.B.
1942 The utilization of urea by ruminants as influenced by the presence of starch in the ration. J. Dairy Sei., 25: 925-929.
Oyaert, W., and Bouckaert, J.H.
1960 Zentr. Veterinaermed., 7: 929-935. Cited in Cham. Abstr., 55: 8557(1961).
San Pietro, A.
1957 The measurement of stable isotopes. In Oo1owick, S.P., and Cap1an, N.O. Methods ~ Enzymo1ogY. Vol. IV. Academie Press, Inc., New York.
Sprinson, D.B., and Rittenberg, D.
1949 The rate of utilization of ammonia for protein synthesis. J. Biol. Chem., 180: 707-714.
Sugawara, K., Soejim.a, M., and Sagisaka, s.
1958 Nippon Nogeikagaku Kaisha, 32: 921-924. Cited in Chem. Abstr., 53: 18230(1959).
Van S1yke, D.D., and Cullen, G.E.
1914 A permanent preparation of urease, and its use in the determination of urea. J. Biol. Cham., 19: 211-228.
Virtanen, A.I., and Land, H.
1959 Acta. Agral. Fennica, no. 94: 7-12. Cited in Chem. Abstr., 54: 9017(1960).
Warner, A.C .I.
1961 Personal communication.

- 163 -
Watson, C.J., Davidson, W.M., and Kennedy, J.W.
1949 The nutritive value of nitrogenous compounds for ruminants. II. The formation of body protein from ure a labelled with the isotope Nl5. Sei. Agri., 29: 185-188.
Wegner, M.I., Booth, A.N., Bohstedt, G., and Hart, E.B.
1940 The nin vitro 11 conversion of inorganic nitro~n to protein by microorganisms from the cow's rumen. J. Dairy Sei., 23: 1123-1129.
Weiler, R.A., Gray, F. V., and Pilgrim, A.F.
1958 The conversion of plant nitro~n to microbial nitrogen in the rumen of the sheep. Brit. J. Nutr., 12: 421-429.
Recommended

















