
1
Running head: Membrane anchor of StREM1.3 Remorin 1
2
Correspondence: 3
Dr Sébastien MONGRAND, phD 4
Address: Laboratoire de Biogenèse Membranaire (LBM) 5
CNRS UMR 5200 / Université Bordeaux Segalen 6
146, rue Léo Saignat - Case 92 7
33076 Bordeaux Cédex 8
Email: [email protected] 9
Telephone: + 33 (0) 5 57 57 14 35 10
11
Journal research area: 12
Biochemical Processes and Macromolecular Structures 13
Cell Biology 14
Plant Physiology Preview. Published on August 1, 2012, as DOI:10.1104/pp.112.200519
Copyright 2012 by the American Society of Plant Biologists
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

2
Plasma membrane localization of StREM1.3 Remorin is mediated by 15
conformational changes in a novel C-terminal anchor and required for the 16
restriction of PVX movement 17
18
Artemis Perraki, Jean-Luc Cacas, Jean-Marc Crowet, Laurence Lins, Michel 19
Castroviejo, Sylvie German-Retana, Sébastien Mongrand1* and Sylvain Raffaele1 20
21
22
Laboratoire de Biogenèse Membranaire, (LBM) Unité Mixte de Recherche 5200 23
(UMR 5200) Centre National de la Recherche Scientifique (CNRS)-Université de 24
Bordeaux, 146 rue Léo Saignat, 33076 Bordeaux Cedex, France (SR, AP, JLC and 25
SM) 26
27
The Sainsbury Laboratory, John Innes Centre, Norwich NR4 7UH, UK (SR) 28
29
Centre de Biophysique Moléculaire Numérique, ULg, Gembloux Agro Bio Tech, 30
Passage des Déportés, 2, B-5030 Gembloux, Belgium (JMC and LL) 31
32
Laboratoire de Microbiologie Fondamentale et Pathogénicité Unité Mixte de 33
Recherche 5234 (UMR 5234) Centre National de la Recherche Scientifique (CNRS) 34
146 rue Léo Saignat 33076 Bordeaux Cedex, France (MC) 35
36 Equipe de Virologie, INRA and Université de Bordeaux, UMR 1332 Biologie du Fruit 37
et Pathologie, BP81, 33883 Villenave d'Ornon Cedex, France. (SGR) 38
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

3
Footnotes : 39
This work was supported by a Marie Curie IEF to S.R. (contract 255104). We 40
acknowledge the French ‘Agence Nationale pour la Recherche’ (ANR) for financial 41
support (contracts NT09_517917 PANACEA to S.M. and JLC). L.L. is senior 42
research assistant at the Belgian National Funds for Research (FNRS). LL and JMC 43
thank IAP project (iPROS) for financial support. 44
45
1 - SR and SM must be considered as co-last authors 46
47
SR Present address: Laboratoire des Interactions Plantes Micro-organismes, Chemin 48
de Borde Rouge BP 52627, 31326 Castanet Tolosan, France. 49
50
JLC Present address: Université de Bourgogne, UMR Plante-Microbe-Environnement 51
1088, Institut National de la Recherche Agronomique (INRA), 5184 CNRS, 21065 52
Dijon Cedex, France. 53
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

4
ABSTRACT 54
The formation of plasma membrane (PM) micro-domains plays a crucial role in 55
the regulation of membrane signalling and trafficking. Remorins are a plant-specific 56
family of proteins organized in six phylogenetic groups, and Remorins of the group 1 57
are among the few plant proteins known to specifically associate with membrane 58
rafts. As such, they are valuable to understand the molecular bases for PM lateral 59
organization in plants. However, little is known about the structural determinants 60
underlying group 1 Remorins specific association with membrane rafts. We used a 61
structure-function approach to identify a short C-terminal anchor (RemCA) 62
indispensable and sufficient for tight direct binding of Solanum tuberosum REMORIN 63
1.3 (StREM1.3) to the PM. RemCA switches from unordered to an alpha-helical 64
structure in a non-polar environment. Protein structure modelling indicates that 65
RemCA folds into a tight hairpin of amphipathic helices. Consistently, mutations 66
reducing RemCA amphipathy abolished StREM1.3 PM localization. Furthermore, 67
RemCA directly binds to biological membranes in vitro, shows higher affinity for 68
Detergent-Insoluble Membranes (DIM) lipids, and targets YFP to DIMs in vivo. 69
Mutations in RemCA resulting in cytoplasmic StREM1.3 localization abolish 70
StREM1.3 function in restricting potato virus X movement. The mechanisms 71
described here provide new insights on the control and function of lateral segregation 72
of plant PM. 73
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

5
INTRODUCTION 74
Protein-lipid interactions are increasingly recognised as key regulatory 75
processes for signal perception and cellular signalling cascades (Cho and Stahelin, 76
2005). During signal transduction and trafficking, a number of soluble proteins 77
dynamically associate with plasma membranes (PMs) to deliver their cargo and to 78
recruit pathway components to the sites of action (Seong et al., 2011). For such 79
proteins, membrane association can be critical for function (Porter and Koelle, 2010). 80
81
PM targeting of peripheral proteins is achieved through (i) binding to integral 82
membrane proteins, (ii) post-translational modifications, or (iii) directly by intrinsic 83
membrane anchor domains. Post-translational modifications function as auxiliary 84
modifications for transient or weak association of soluble proteins to the intracellular 85
face of the PM. In plants, these include N-myristoylation, S-palmitoylation, prenylation 86
by farnesyl or geranylgeranyl moieties, or attachment of glycosylphosphatidylinositol 87
(GPI) anchors (Thompson and Okuyama, 2000). GPI-anchors, for example, tightly 88
associate proteins to the extracellular face of PMs by interaction of the inositol head 89
group of the membrane lipid phosphatidylinositol with a glucosamine residue linked to 90
the C-terminal amino acid of the protein (Paulick and Bertozzi, 2008). As an 91
alternative mechanism, globular structures either recognize phospholipids in a 92
stereospecific manner or associate with membranes by their biophysical properties 93
(reviewed in Lemmon, 2008). Other proteins expose unstructured clusters of basic 94
and hydrophobic residues to mediate PM binding (McLaughlin et al., 2002; 95
McLaughlin and Murray, 2005). 96
97
Selective recognition of membrane compartments or domains by protein 98
anchors can be critical in triggering the appropriate downstream trafficking and 99
signalling events (for review: Gruenberg, 2003; De Matteis and Godi, 2004). 100
Membrane domain selectivity can be specified by the anchoring post-translational 101
modification or by a protein anchor domain. For instance proteins carrying GPI-102
anchors are over-represented in membrane rafts, indicating that addition of this lipid 103
anchor directs proteins to these micro-domains (Cordy et al., 2003; Kierszniowska et 104
al., 2009). Membrane rafts are enriched in highly-saturated long chain sphingolipids, 105
sterols and saturated phospholipids, creating tightly-packed domains, designated as 106
‘Liquid-ordered’. These lipids display a stronger affinity to saturated acyl chains as 107
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

6
found in GPI anchored and acylated proteins (Brown, 2006). The composition of 108
membrane rafts also prevents solubilisation by detergent at low temperature with 109
non-ionic detergent, and allows the partial purification of raft in so called Detergent-110
Insoluble Membrane (DIM) fractions, supposedly biochemical counterparts of 111
membrane rafts. Many signalling proteins are found in membrane rafts supporting the 112
hypothesis that they serve as key platforms for cellular signal transduction and cell-113
to-cell communication (Lingwood and Simons, 2010; Simon-Plas et al., 2011). For 114
example, in human cells, key soluble signalling components such as the Ser/Thr 115
kinase Akt (protein kinase B) are recruited to membrane rafts where they activate 116
signal transduction cascades (Lasserre et al., 2008). Nevertheless, few protein motifs 117
were described to contribute to raft targeting (Rossin et al., 2010) while the six amino 118
acid long raft target signal from human tyrosine phosphatase Src homology 2-119
containing phosphatase 1 (SHP-1) is the only known motif sufficient to anchor soluble 120
proteins specifically to domains of the intracellular face of the PM (Sankarshanan et 121
al., 2007). However, the anchoring mechanism itself remains to be unravelled. In 122
plants even though DIMs also exist (Mongrand et al., 2010) and functional PM 123
domains have been reported (Bhat et al., 2005), the molecular basis for specific 124
targeting and binding of proteins to membrane rafts has never been described. 125
126
Remorins form a diverse family of plant-specific proteins organized in six 127
distinct phylogenetic groups (Raffaele et al., 2007). Remorins from the group 1 have 128
been reported to localize to the PM despite their overall hydrophilic nature (Reymond 129
et al., 1996; Raffaele et al., 2007). Moreover, group 1 Remorins almost exclusively 130
associate to DIMs and localize to membrane micro-domains in a sterol-dependent 131
manner (Lefebvre et al., 2007; Kierszniowska et al., 2009; Raffaele et al., 2009). The 132
function of Remorins is mostly unknown, but we showed in a previous study that 133
StREM1.3 (Solanum tuberosum Remorin from group 1, homolog 3, initially described 134
in (Reymond et al., 1996) regulates cell-to-cell propagation of the potato virus X 135
(PVX) likely by directly interacting with the viral movement protein TGBp1 (Raffaele 136
et al., 2009). StREM1.3 localizes to the inner leaflet of PMs and along 137
plasmodesmata, bridges connecting neighbour cells essential for cell-to-cell 138
communication in plants (Maule, 2008). Other members of the Remorin family group 139
1 are likely involved in innate immune responses (Liu et al., 2009; Widjaja et al., 140
2009; Keinath et al., 2010). Remorins from group 2 are involved in the control of 141
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

7
infection by symbiotic bacteria at nodular infection threads and the peribacteroid 142
membrane (Lefebvre et al., 2010) These data suggest general roles for Remorins in 143
regulating signalling in plant-microbe interactions (Jarsch and Ott, 2011). 144
145
Elucidating the mechanisms driving StREM1.3 association with PM micro-146
domains therefore provides a unique opportunity for understanding the regulation 147
and function of membrane lateral segregation in plants. StREM1.3 does not contain 148
predictable transmembrane or membrane associated domains. The bases for its 149
association to PMs and selective targeting to DIMs are unknown. Here we identified a 150
novel membrane anchor domain required for StREM1.3 tight and direct association 151
with detergent insoluble fraction of the PM. We combined biophysics, in silico 152
analysis and directed mutagenesis to unravel the molecular bases of StREM1.3 153
membrane binding and its biological significance in the control of PVX propagation. 154
155
RESULTS 156
157
StREM1.3 is a strongly associated peripheral membrane protein 158
To identify structural domains potentially involved in PM localization of 159
StREM1.3, we performed in silico domain analysis and secondary structure 160
prediction. Proteins of the Remorin family typically contain a variable N-terminal 161
region (Marín and Ott, 2012) and a conserved C-terminal domain, termed Remorin_C 162
(PF03763), of approximately 120 amino acid (aa) residues that encompasses a 163
predicted coiled-coil domain. In StREM1.3, the Remorin_C domain extends from aa 164
85 to 195 with a coiled-coil domain predicted between aa 116 to 152 (Figure 1A). 165
StREM1.3 C-terminal region is predicted to be mostly alpha-helical composed of a 84 166
aa long coiled alpha helix ranging from position 85 to 169. This helix is predicted to 167
be interrupted at Glycine residue 171. A short (~12 aa) alpha helix is predicted 168
between residues 172 and 187. Finally the most C-terminal residues are predicted to 169
form a random coil except for residues 192 to 195 where a short alpha helix is 170
predicted. The N-terminal region of StREM1.3 contains the proline-rich Remorin_N 171
domain (PF03766) extending between aa 27 to 84 and is mostly unordered (Marín 172
and Ott, 2012). No recognizable transmembrane domains or any other membrane-173
targeting signals could be detected, indicating that StREM1.3 is a peripheral 174
membrane protein with an atypical membrane-binding domain. 175
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

8
To test the nature of Remorin association with biological membranes we 176
purified native microsomes and PMs from tobacco leaves and tested its affinity by 177
removal of extrinsic membrane proteins. In accordance with published data PM 178
stripping resulted in ~50% loss of total protein, representing peripheral proteins with 179
weak affinity to the PM (Santoni et al., 2003). Typical of peripheral membrane 180
proteins is Actin, which was almost completely removed from the PM after treatment 181
(Figure 1B). However, endogenous StREM1.3 remained almost entirely associated 182
with the PM, similar to true integral proteins such as the aquaporin PIP2 that was 183
used as an integral membrane protein control. Similar results were obtained when 184
purified PMs were washed under high ionic strength conditions, increasing EDTA 185
concentrations, and increasing pH for 30 minutes (Figure S1A). These results 186
suggest that the association of StREM1.3 with the membrane is not limited to 187
electrostatic interactions. 188
189
Remorin C-terminal Anchor (RemCA) is required and sufficient for plasma 190
membrane targeting in vivo. 191
A series of full length and truncated protein variants was generated and fused 192
to G/YFP to analyse the structural requirements for StREM1.3 PM localization. The 193
full-length protein localized to the PM (Figure 1C). This localization is consistent with 194
immunolocalization studies reported earlier (Raffaele et al., 2009; Lefebvre et al., 195
2010) suggesting that the fluorescent tag did not affect localization of the fusion 196
proteins. Deletion of the C-terminal residues (StREM1.31-170) resulted in an entire 197
loss of PM association indicating the presence of a PM anchor motif within these 198
residues (Figure 1C). We will refer to this 28 amino acid region as the “Remorin C-199
terminal Anchor” (RemCA). This hypothesis was confirmed by reciprocal experiments 200
where expression of soluble YFP tagged with RemCA led to clear YFP association 201
with the PM (Figure 1C). We designed a variety of different constructs to express the 202
fluorescently tagged RemCA but we were not able to prevent a partial cleavage of 203
the tag, resulting in a residual nucleo-cytoplasmic fluorescence in vivo. To confirm 204
that RemCA is sufficient to anchor soluble proteins to the PM, we performed 205
biochemical fractionation experiments on cells expressing StREM1.3 G/YFP fusion 206
constructs (Figure 1D). These experiments showed that the GFP:StREM1.3 is fully 207
microsomal whereas YFP:StREM1.31-170 is entirely soluble. The YFP:RemCA fusion 208
was recovered exclusively in the microsomal fraction, with only cleaved YFP in the 209
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

9
soluble fraction. These results demonstrate that RemCA is required for StREM1.3 210
PM localization and sufficient to fully associate soluble proteins with the PM. 211
212
Non-polar environments trigger conformational changes in RemCA. 213
To analyse the structural properties of RemCA we obtained chemically 214
synthesized tag-free ultra pure peptides that were assessed using circular dichroism 215
(CD) spectroscopy. CD spectra were acquired in 100% 2,2,2-Trifluoroethanol (TFE), 216
a solvent mimicking biological membranes hydrophobicity (causing partial 217
desolvation of the peptide, a situation that occurs in membranes and during protein-218
protein and protein-lipid interactions), or in semi-hydrophobic solvent (50% TFE, 50% 219
Tris-HCl). CD spectra showed distinct minima at 208 and 222 nm, with a marked 220
maximum at 195 nm, consistent with RemCA forming an alpha helical structure 221
(Figure 2A). By contrast the spectrum of RemCA in aqueous solution (100% Tris-222
HCl) showed a reduced minimum at 208 nm and reduced concavity around 195 nm, 223
indicating that alpha helical contributions to the overall RemCA structure are reduced 224
under these conditions. We estimated RemCA protein secondary structure 225
composition from CD spectra (Figure 2B) and found that it undergoes transition from 226
15% alpha-helical content in aqueous solution to 50% in hydrophobic non-polar 227
solvents. Such estimations remain challenging and should thus be taken with 228
caution. Nevertheless, these data imply that the RemCA peptide is largely unordered 229
in aqueous solution and folds into a mostly alpha helical structure in hydrophobic 230
environments. 231
To gain further insights into the structural mechanisms underlying RemCA 232
binding to PMs, we performed homology 3D modelling of this peptide using the I-233
TASSER server. The predicted structure for RemCA corresponds to two alpha 234
helices arranged as a tight hairpin (Figure 2C, Supplemental File 1). This structure 235
is reminiscent of a recently published structure for Influenza hemagglutinin fusion 236
peptide (HAfp) (Lorieau et al., 2010) (Figure S2). The predicted structure has a high 237
helical content, therefore probably corresponding to the conformation adopted by 238
RemCA in hydrophobic environments. This hypothesis is consistent with the structure 239
of the HAfp which forms a tight hairpin of alpha helices in the presence of lipids 240
(Lorieau et al., 2010). The distribution of the relative hydrophobicity of amino-acids 241
reveal an accumulation of hydrophobic aa at the centre of the hairpin, with the side 242
chains of hydrophilic charged amino acids pointing outward (Figure 2C). The turn 243
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

10
connecting the two alpha helices is composed of the palindromic neutral sequence 244
Ala-Thr-Gly-Thr-Ala. 245
246
Amphipathy of RemCA alpha helices is required for Remorin anchoring to the 247
PM in vivo. 248
The predicted hydrophobic core at the centre of the RemCA hairpin and 249
solvent-exposed charged surfaces (Figure 2C) implies an asymmetry in the 250
distribution of hydrophobic and charged residues around the alpha-helices, a 251
property known as amphipathy. Amphipathic helices are crucial for transient 252
anchoring of some peripheral membrane proteins, notably viral fusion proteins (e.g 253
Lins and Brasseur, 2008; Drin and Antonny, 2009). We used the heliquest server to 254
calculate the difference, named hydrophobic moment (µH), in hydrophobicity 255
between the two faces of each alpha helix forming RemCA. The E172-A186 helix had a 256
µH of 0.153, similar to values obtained for viral fusion peptides (e.g 0.171 for the 257
Simian influenza virus -SIV- fusion peptide), whereas helical fragment P191-F198 258
had a µH of 0.661, with each helix organized with clearly distinct charged and 259
hydrophobic faces (Figure 3A). 260
To test the importance of alpha-helices amphipathy in the function of RemCA, 261
we designed a StREM1.3* variant harbouring a mutated RemCA* with reduced 262
amphipathy (Figure 3B, Supplemental File 2). The transient expression of a 263
YFP:StREM1.3* in tobacco leaves showed that the mutations completely abolished 264
membrane localization, leading to a fully cytoplasmic protein (Figure 3C). Loss of PM 265
association was confirmed biochemically by protein fractionation where the 266
YFP:StREM1.3* protein was exclusively detected in the soluble fraction, while 267
endogenous Remorin remained in the PM fraction (Figure 3D). 268
269
RemCA directly binds to lipid bilayers in vitro. 270
To test whether RemCA anchors StREM1.3 to the PM independently of post-271
translational modifications or association with other proteins, recombinant 272
6His:StREM1.3 (+), 6His:StREM1.31-170 (-, deletion of RemCA), and 6His:StREM1.3* 273
(*, mutated RemCA, see above) were purified from E. coli, and tested for binding to 274
liposomes mimicking plant PM lipid composition in vitro. Liposomes were separated 275
at the top of a sucrose gradient by centrifugation. The presence of StREM1.3 in the 276
liposome-bound or unbound fraction was assessed by Western Blot and lipids were 277
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

11
analysed by TLC (Figures 4A and S3) (Figure. StREM1.3 bound to liposomes in the 278
absence of any other proteins (Figure 4A). Although similar amounts of liposomes 279
were isolated for each sample (Figure S3B), binding was completely abolished in 280
StREM1.31-170 and StREM1.3* variants (Figure 4A). Furthermore, and in agreement 281
with our data obtained using purified PMs, treatment with denaturing agents did not 282
significantly reduce the amount of 6His:StREM1.3 bound to liposomes (Figure S1B). 283
These results demonstrate that RemCA confers direct membrane binding to 284
StREM1.3. Loss of membrane binding in StREM1.3* indicates that amphipathy plays 285
a crucial role in this process. 286
Insertion of a peptide into membranes results in a partial destabilization of 287
artificial bilayers and can thus be taken as a measure to discriminate between 288
physical insertion and outer association (Longo et al., 1997). To test whether RemCA 289
induces membrane destabilization, we performed lipid-mixing assays in which 290
application of purified peptides to a mixture of liposomes, labelled or not by the 291
lipophilic fluorescent dye octadecyl rhodamine chloride (R18), leads to a de-292
quenching of fluorescence. We confirmed the functionality of this assay by the 293
application of the well-known Simian immunodeficiency virus (SIV) fusion peptide 294
(Lorin et al., 2008) to liposomes (see materials and methods). By contrast liposome 295
treatment with TFE, used as a solvent for the peptides, did not result in lipid mixing. 296
Application of synthetic ultra pure RemCA peptides resulted in a ~40% de-quenching 297
of fluorescence relative to the values obtained with the SIV fusion peptide (Figure 298
4B) indicating destabilization of the membrane and insertion of the peptide in the 299
bilayer. 300
To verify that the increase in fluorescence was due to liposome destabilization rather 301
than liposome aggregation, we performed liposome leakage assays, in which 302
liposomes entrapping the fluorescent dye 1-hydroxypyrene-3,6,8-trisulfonate (HPTS) 303
and the corresponding cationic quencher α,α'-dipyridinium p-xylene dibromide (DPX) 304
were used. Application of fusion peptides leads to a de-quenching of fluorescence 305
due to physical separation of both components (Ellens et al., 1985). We used 306
liposomes composed of either negatively charged lipids or neutral lipids. As 307
expected, the use of the detergent TX100 entirely disrupted the liposomes, leading to 308
maximal fluorescence (Figure 4C). Application of the SIV fusion peptide induced 309
~90% leakage of negatively charged liposomes, and only ~9% leakage of neutral 310
liposomes, well above leakage induced by solvents used for the peptides (leakage 311
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

12
<2% by either DMSO or TFE for negatively charged and neutral liposomes 312
respectively). Application of synthetic RemCA peptide yielded in ~5% of leakage of 313
negatively charged liposomes, but did not cause any significant leakage of neutral 314
liposomes (Figure 4C). These experiments demonstrate that RemCA physically 315
inserts into membranes independently of other proteins, and predominantly binds 316
negatively charged lipids, suggesting a differential affinity for classes of lipids. 317
318
RemCA confers lipid-specific binding to StREM1.3 319
To test whether RemCA binds selectively to certain lipids, we probed 320
immobilized lipids with purified recombinant 6His:StREM1.3 and 6His:StREM1-170 in 321
lipid overlay assays. StREM1.3 showed differential affinity to 5 of the 7 lipids tested 322
(Figure 5A). Affinity of StREM1.3 was strong to PI(3,4)P2 phosphoinositol (3,4)-323
bisphosphate and sphingosine-1-phosphate (S1P), moderate to phosphatidic acid 324
(PA) and PI(3,5)P2, and weak to phosphatidylserine (PS). Binding to these 325
immobilized lipids was dependent on RemCA since it was almost completely 326
abolished in the StREM1.31-170 protein, with only a weak residual binding to PI(3,4)P2 327
remaining. Similar results were obtained with the purified recombinant 328
6His:StREM1.3* mutant (Figure S4A). These results indicate that RemCA has 329
affinity for minor anionic phospholipids and negatively charged lipids, and that 330
RemCA confers selectivity to the range of lipids bound in vitro. Interestingly, these 331
lipids are enriched in membrane rafts from plants (Vermeer et al., 2009; Furt et al., 332
2010) or animal cells (Hope and Pike, 1996; Pike et al., 2002), suggesting that 333
RemCA lipid-specificity contributes to driving the segregation of StREM1.3 into 334
membrane micro-domains. 335
To support this hypothesis, we first tested the binding of 6His:StREM1.3 to 336
liposomes with lipid composition mimicking either the PM or DIMs in vitro (see 337
materials and methods). Although the total amount of liposomes pulled down was 338
similar for all samples (Figure S4B), the relative quantification of 6His:StREM1.3 339
bound demonstrated a higher affinity for DIM-like liposomes compared to PM-like 340
liposomes (Figure 5B). 341
Finally, to test the specificity of RemCA-mediated binding in vivo, we extracted 342
DIMs from PM purified from tobacco plants expressing RemCA tagged with the 343
soluble YFP. As expected, ~80% of total proteins were excluded from the DIM 344
fraction (Figure 5C, S4C) (see also Raffaele et al., 2009) but the endogenous 345
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

13
tobacco Remorin proteins were almost exclusively found in DIM fractions (Figure 346
5C). Strikingly, the majority of YFP:RemCA proteins were also detected in DIMs 347
(Figure 5C). These results demonstrate that RemCA is sufficient to target soluble 348
proteins specifically to DIMs of plant PM. 349
350
StREM1.3 forms homo-trimers in vitro independently of RemCA 351
Previous work showed by direct electron microscopy observations and 352
chemical crosslinking with glutaraldehyde that group1 Remorins forms homo-353
polymeric filaments (Bariola et al., 2004). To test whether RemCA plays a role in the 354
assembly or stability of these higher-order polymers, we used exclusion column 355
chromatography to estimate the native molecular weight (MW) of 6His:StREM1.3 and 356
6His:StREM1.3* recombinant proteins. Chromatography revealed a peak of elution at 357
~1.25 mL (Figure S5A). Western blot analysis after denaturing SDS-PAGE of the 358
corresponding fractions confirmed that 6His:StREM1.3 eluted at this volume (Figure 359
S5B). Calibration of the gel filtration column with standards of known MW allowed to 360
estimate 6His:StREM1.3 native MW to ~120 kDa, suggesting that 6His:StREM1.3 361
forms meta-stable trimers in vitro (Figure 6A). Neither monomers nor dimers were 362
detected by gel filtration. Chromatography performed with 6His:StREM1.3* protein 363
showed similar elution pattern, indicating that a non-functional RemCA does not alter 364
the assembly of StREM1.3 into trimers. Interestingly, as discussed previously (Figure 365
S2), RemCA shows structural similarities with influenza virus hemagglutinin (HA) 366
fusion peptide. Influenza HA adopts a central trimeric alpha-helical coiled-coil 367
structure, and the formation of trimers of fusion peptide hairpins is critical for 368
membrane fusion (Cross et al., 2001). To get insights on the possible consequences 369
of StREM1.3 trimer formation on its membrane-binding properties, we used 370
homology modelling and geometry based docking to produce a 3D model of 371
StREM1.3 C-terminal domain (E87-F198) trimer (Figure 6B). This model suggests 372
that RemCAs are organized circularly, forming a hydrophobic core at the centre of 373
RemCA trimer of hairpins and exposing patches of positively charged residues all 374
around the C-terminus of the trimer. This conformation would create a strong 375
hydrophobic moment directed inward RemCA trimer, and likely increases the 376
strength and lipid-specificity of the membrane binding. 377
378
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

14
Mutations in RemCA* abolish StREM1.3 restriction of PVX cell to cell 379
movement 380
To test for the biological significance of StREM1.3 PM localization, we 381
exploited the previously reported role of StREM1.3 in restricting the cell-to-cell 382
propagation of the potato virus X (PVX) in tomato leaves (Raffaele et al., 2009). 383
Here, we transiently co-expressed in N. benthamiana leaves 35S:StREM1.3 or 384
35S:StREM1.3* with a highly diluted suspension of Agrobacterium tumefaciens 385
harbouring an infectious PVX:GFP clone, so that only isolated individual cells were 386
infected with the PVX:GFP. The accumulation of PVX:GFP in distinct infection foci 387
was visible 5 days post infection (dpi). StREM1.3 or StREM1.3* expression was 388
verified by Western blot (Figure S6). The effect of the expression of the StREM1.3 389
proteins was assessed by visualizing the virus infection foci and counting the number 390
of fluorescent cells of at least 30 infection foci by comparison with plants infiltrated 391
with A. tumefaciens carrying the PVX:GFP clone only. At 5 dpi, plants infiltrated with 392
A. tumefaciens carrying PVX:GFP alone showed infection foci covering 17 cells in 393
average (Figure 7A, B). As expected, the co-expression of StREM1.3 lead to a 394
significant decrease in PVX:GFP spreading, with foci covering 12 cells in average 395
(Figure 7A, B). By contrast, co-expression of StREM1.3* did not significantly alter 396
the size of the PVX:GFP infection foci (Figure 7A, B). StREM1.3 was shown to 397
directly interact with the TGBp1 viral movement protein during PVX infection 398
(Raffaele et al., 2009). To determine if the loss of activity in StREM1.3* was due a 399
loss of interaction with TGBp1, we tested the interaction of the StREM1.3* and 400
StREM1.31-170 mutants with TGBp1 in a Split Ubiquitin assay. In both case, a clear 401
interaction was observed, similar to the wild type StREM1.3 (Figure 7C). These 402
results clearly demonstrate that RemCA-mediated PM anchoring of StREM1.3 is 403
required for the function of this protein in the control of virus propagation and indicate 404
that this activity takes place at the PM. 405
406
DISCUSSION 407
In this paper we showed that StREM1.3 directly binds to the PM via a short C-408
terminal anchor domain (RemCA). This domain is unordered in aqueous solution and 409
spontaneously folds, probably into a tight hairpin of amphipathic alpha helices, in 410
non-polar environments. The amphipathy of RemCA helices is required for StREM1.3 411
PM localization. RemCA selectively binds to lipids enriched in DIMs and targets the 412
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

15
soluble YFP to DIMs in vivo. StREM1.3 forms trimers in vitro. This does not require a 413
functional RemCA and may increase the stability of PM anchoring in vivo. PM 414
anchoring is crucial for StREM1.3 function in restricting PVX movement since 415
mutations in RemCA abolishing PM binding also abolish StREM1.3 function towards 416
PVX. Molecular mechanisms controlled by raft-mediated signalling and invasion of 417
host cells via membrane micro-domains may be altered by directly targeting proteins 418
to these sites. Using RemCA as an anchor may thus provide the opportunity to 419
modulate raft-mediated signalling processes or alter host resistance towards 420
pathogenic microbes. 421
422
Molecular bases for the assembly of Remorin micro-domains at the PM: a 423
model. 424
Our results indicate that the folding of RemCA into amphipathic alpha helices, 425
probably organized as a tight hairpin, is required for StREM1.3 PM anchoring and 426
function. Our CD data showed that RemCA switches from unordered to alpha helical 427
folding in non-polar environments. RemCA folding could be triggered by proximity 428
with the hydrophobic vicinity of the PM (Figure 8A). Similar observations were made 429
for Fis1 Tail-anchor protein (Suzuki et al., 2003), the ArfGAP1 lipid sensor motif 430
(Bigay et al., 2005), and for the amphipathic membrane-binding helix of the epsin N-431
terminal homology (ENTH) family of clathrin adaptors (Ford et al., 2002). 432
Alternatively, the formation of StREM1.3 trimers would result in the formation of a 433
hydrophobic core, thereby creating an environment favourable to RemCA folding 434
(Figure 8B, C). Association of group 1 Remorins into higher-order complexes has 435
been reported in vitro (Bariola et al., 2004) and in vivo (Raffaele et al., 2009; 436
Lefebvre et al., 2010), but whether this is triggered by external signals remains 437
unknown. We propose that RemCA anchoring occurs as described for other lipid-438
binding proteins (Stahelin et al., 2003; Peter et al., 2004) in a two-step manner. First, 439
positively charged residues E172-A186 increase the affinity of RemCA for negatively 440
charged lipids of the inner leaflet of membrane rafts (Figure 8A, D). This destabilizes 441
the internal monolayer of the PM and allows RemCA to insert deeper in the 442
membrane (Figure 8B, E). The complete folding of RemCA results in the formation of 443
a tight hairpin of alpha helices defining a hydrophobic pocket at its core. As described 444
for Influenza HAfp (Han et al., 2001), formation of this hairpin could result in a deeper 445
insertion of RemCA explaining the intrinsic-like behaviour of Remorins (see Figure 446
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

16
1B). Considering that Influenza HAfp is non fusogenic if not properly kinked (Lai and 447
Tamm, 2007), the tight closure of RemCA hairpin of helices might be required for 448
stable anchoring in the PM. Finally, the possibility that the formation of StREM1.3 449
multimers occurs at the PM, eventually under control of external signals (Figure 8B, 450
E), would allow the aggregation of membrane rafts into higher order clusters, as 451
suggested during the response to pathogen (Raffaele et al., 2009). 452
453
Biophysical properties defining RemCA lipid-specificity 454
In this work, we have shown that RemCA alone is able to anchor soluble 455
hydrophilic proteins to the PM, such as G/YFP (Figure 1). We cannot exclude that 456
additional structural features of StREM1.3 protein modulate this activity, such as 457
post-translational modifications (e.g. phosphorylation), and the formation of 458
StREM1.3 multimers that would exacerbate local gradients of hydrophobicity and 459
charge. It is remarkable that RemCA preferentially binds to negatively-charged lipids 460
abundant in membrane rafts and target the soluble YFP to DIMs. A number of 461
globular domains is known to bind to the surface of the PM by specifically recognizing 462
certain lipids, particularly discriminating between different phospholipid types 463
(Lemmon, 2008; Kutateladze, 2010). A combination of steric recognition of specific 464
lipid polar head groups, electrostatic attraction to negatively charged lipids and 465
membrane destabilization-mediated penetration govern the function of these 466
domains. Some of those such as the Annexin, ENTH, ANTH (‘AP180 N-terminal 467
Homology’), BAR and PX (‘Phox homology’) domains adopt an alpha-helical fold. 468
Overall the hydrophobicity, the hydrophobic moment, the net charge and helix-469
breaking amino acids contribute to the membrane binding properties of amphipathic 470
helices (Drin and Antonny, 2009). In small GTPases, polybasic clusters provide PM 471
specificity by binding to negatively charged PI(4,5)P2 and PI(3,4,5)P3 of the PM (Heo 472
et al., 2006). The charge of the polar face of ArfGAP1 amphipathic helices has been 473
associated with the ability to perceive membrane curvature, with the introduction of 474
lysine residues at the polar-non polar interface reducing curvature sensitivity and 475
allowing binding to large liposomes (Drin et al., 2007). It is therefore tempting to 476
speculate that RemCA membrane binding could sense membrane curvature as a 477
mean to specifically target certain domains in the membrane. In addition, large 478
hydrophobic residues on both sides of the kink are required for setting the angle of 479
the hairpin structure and for function in Influenza HAfp (Lai and Tamm, 2007). The 480
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

17
L174, L180, I194 and F198 could play this role in RemCA. The overall conformation 481
of RemCA as well as individual residue side chains are likely to contribute to its 482
membrane binding ability, and further investigations will be required to dissect how 483
this activity is mediated. 484
485
Towards understanding the molecular bases of specific association with 486
membrane rafts 487
RemCA drives the soluble YFP to DIM PM fractions, and lipids bound by 488
RemCA are enriched in DIMs. Indeed, polyphosphoinositides and sphingolipids are 489
enriched in membrane rafts from plants (Vermeer et al., 2009; Furt et al., 2010) and 490
animal cells (Hope and Pike, 1996), the latter being also enriched in PS (Pike et al., 491
2002). The ability to specifically target DIMs might therefore be related to the 492
differential affinity of RemCA to certain negative lipids, as known for SNARE proteins 493
(Mima and Wickner, 2009). However, It should be noted that the presence of a 494
protein in DIMs is not sufficient to associate it with membrane rafts (Munro, 2003; 495
Simons and Gerl, 2010; Tanner et al., 2011). In the case of StREM1.3, we previously 496
reported that enrichment in DIMs correlate with clustering into microdomains of 70 497
nm in the PM detected by immunogold-labeling (Raffaele et al., 2009). Importantly, 498
both DIM enrichment and clustering at the PM were phytosterol-dependent 499
(Kierszniowska et al., 2009; Raffaele et al., 2009; Tanner et al., 2011). Since lipid-500
lipid interactions are sufficient to drive the segregation of liquid-ordered and liquid-501
disordered phases in model membranes (Dietrich et al., 2001; Baumgart et al., 2003), 502
it can be hypothesized that RemCA selective lipid binding properties could be 503
sufficient to restrict the localization of StREM1.3 to membrane rafts. 504
Nevertheless, it is reasonable to assume that in addition to RemCA other 505
domains or motifs may determine the organization of StREM1.3 into PM micro-506
domains. Indeed, the association of Remorins with DIMs is regulated quantitatively 507
by signals such as cold (Minami et al., 2009) and pathogen-associated molecular 508
patterns (PAMPs) (Keinath et al., 2010). The molecular bases of this regulation are 509
not fully understood but may likely involve protein-protein interactions or post 510
translational modifications. It is also not known whether the high-ordered organization 511
of Remorins into multimers is regulated by external signals. In any case, protein-512
protein interactions mediated by the coiled-coil or the N-terminal domain of Remorins 513
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

18
(Lefebvre et al., 2010; Tóth et al., 2012) likely modulate the localization of Remorins 514
in vivo. 515
516
517
Implications for the mechanisms underlying PVX cell-to-cell movement 518
In a previous study we reported that StREM1.3 restricts potato virus X (PVX) 519
cell-to-cell movement independently of viral replication and that StREM1.3 physically 520
associates in vivo and in vitro with the triple gene bloc (TGB) protein 1 of the virus 521
(Raffaele et al., 2009). Here we show that restriction of PVX movement is abolished 522
in StREM1.3* cytoplasmic mutant. We also show that deletion or mutation of RemCA 523
do not alter StREM1.3 interaction with PVX TGBp1 in a split ubiquitin assay, which is 524
consistent with evidences that RemCA would insert into the PM. This indicates that 525
RemCA is not directly involved in the interaction with TGBp1 but rather that 526
StREM1.3 –TGBp1 interaction takes place at the PM. The sub-cellular localization of 527
TGBp1 is complex and depends on the other TGB proteins, viral RNA, and on the 528
host plant species (Verchot-Lubicz et al., 2010). When expressed alone in N. 529
benthamiana, TGBp1 is mostly nuclear and cytoplasmic. However in the presence of 530
viral RNA, TGBp1 could traffic along the ER with TGBp2 and TGBp3 granules, and 531
reach plasmodesmata. Given that StREM1.3 is anchored on the cytosolic face on the 532
PM, it could associate with cytoplasmic TGBp1 at the periphery of the cell, explaining 533
why we could detect association when TGBp1 was expressed alone. Future work will 534
investigate the specificity of StREM1.3 association with TGBp1 and the impact of 535
viral RNA on this interaction. The recent report of the role of Arabidopsis 536
synaptotagmin 1 (SYT1) in the control of Begomovirus and Tobamovirus cell-to-cell 537
movement (Lewis and Lazarowitz, 2010) shed new light on the involvement host 538
membrane proteins in virus movement. SYT1 resides to the cortical endoplasmic 539
reticulum, endosomes, in DIMs at the PM and in PDs (Schapire et al., 2008; Minami 540
et al., 2009; Lewis and Lazarowitz, 2010; Yamazaki et al., 2010), and regulates 541
endocytosis and PM repair processes (Schapire et al., 2008; Yamazaki et al., 2008; 542
Lewis and Lazarowitz, 2010). These studies support a link between DIMs and PDs, 543
and provide evidence for a role of DIM proteins in cell-to-cell communication via PDs. 544
545
Considering the diversity of proteins in the Remorin family (Raffaele et al., 546
2007), there is probably a corresponding diversity of sub-cellular localizations and 547
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

19
functions to be discovered. Analysing the variability the RemCA region in relation with 548
Remorins sub-cellular location, the variability of the membrane lipid composition 549
according to the nature of membrane compartments and plant species, and specific 550
functions devoted to each protein in the Remorin family will provide valuable insights 551
in the mechanisms regulating lateral heterogeneity of plant PM. 552
553
MATERIALS AND METHODS 554
Molecular cloning and sequence analyses 555
StREM1.3 was cloned from Potato Solanum tuberosum cv. Desiree cDNAs by 556
standard Gateway techniques using the pDONR221 entry vector and primers 557
containing the recombination site sequences and the gene specific sequences 5’-558
ATGGCAGAATTGGAAGCT-3’ (forward) and 5’-TCAAAATATTCCAAGGAT-3’ 559
(reverse for N-terminal fusion), 5’-AAATATTCCAAGGAT-3’ (reverse for C-terminal 560
fusion), or 5’- TCAACGTTTAGCTTCAATCAT-3’ (reverse for YFP :StREM1-170). A 561
forward primer containing the gene specific sequence 5’-GGAGAAGATCTTCTC-3’ 562
was used to clone the RemCA constructs. StREM1.3* was generated by PCR 563
amplification using a reverse primer containing the sequence 5’- 564
TCAAAATATTACAGAGATTTTCTTTGGAGAAGTTACAGTGGAACGGACTTTTGC-565
3’. Mutated residues were selected at the C-terminus of RemCA to facilitate PCR-566
based mutations. The position of Glycines and the aromatic residue were shown to 567
be important in HA fusion peptide function (Cross et al., 2001), therefore G188V and 568
G196V were introduced to preserve small size while introducing hydrophobic amino-569
acids and Y184V to remove the aromatic residue. A186S and A190S were 570
introduced to preserve the tiny size of amino acids while removing hydrophobic 571
residues and L195S allowed reducing further the local hydrophobicity of RemCA 572
peptide. These mutations allowed reducing the hydrophobic moment from 0.182 to 573
0.112 for the whole RemCA peptide. These entry clones were recombined into the 574
pDEST17 (6His fusions), pH7YWG2, pH7WGY2 or pk7WGF2 (YFP/GFP fusions) 575
vectors. Secondary structure predictions were ran on SSPro 576
(http://download.igb.uci.edu/sspro4.html) and JPred 3 577
(http://www.compbio.dundee.ac.uk/www-jpred/) servers. Coiled-coil domains were 578
predicted using Marcoil1.0 server (http://www.isrec.isb-579
sib.ch/webmarcoil/webmarcoilC1.html).. Amphipathic helix analyses were performed 580
with heliquest server (http://heliquest.ipmc.cnrs.fr/) and 3D modelling via the I-581
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

20
TASSER server (http://zhanglab.ccmb.med.umich.edu/I-TASSER/) (Roy et al., 2010). 582
StREM1.3 C-terminal domain models were assembled into a trimer using the 583
SymmDock server (Schneidman-Duhovny et al., 2005). Models were rendered using 584
UCSF Chimera (Pettersen et al., 2004). 585
586
Plant transformation and fluorescence microscopy 587
Four week old tobacco plants (Nicotiana tabacum cv. Xanthi) were used for 588
Agrobacterium tumefaciens (strain GV3101)-mediated transient expression (Batoko 589
et al., 2000). Confocal imaging was performed two days after agroinfiltration using 590
Leica TCS SP2/5 confocal microscopes with 20x to 63x oil/water immersion 591
objectives. Laser and image settings were as described by (Raffaele et al., 2009). 592
593
Preparation of pure PM and Detergent Insoluble Membrane (DIM) 594
Microsomal fractions and PMs from tobacco leaves were purified described in 595
(Mongrand et al., 2004). For DIM preparation, TX100 (10 % w/v) was added to a ratio 596
detergent-to-PM proteins of 10 (at 1% w/v final concentration), and the PM 597
preparations were solubilized at 4°C for 30min. Treated PMs were brought to a final 598
concentration of 48 % sucrose (w/w), overlaid successively with layers of 40, 35 and 599
30 % sucrose in TBS buffer (w/w), and then centrifuged for 16 h at 200,000 g in 600
TST41 rotor (SORVALL). DIMs could be recovered above the 30-35 % layers as an 601
opaque band and this fraction was washed in 4 ml of TBS buffer to remove residual 602
sucrose. The protein concentration was determined with a BCA protein assay to 603
avoid TX100 interference, using bovine serum albumin as a protein standard. 604
605
Protein analysis, Western blots and protein purification 606
6-Histidine tagged constructs were expressed in E. coli BL21 DE3 cells. Cells were 607
lysed by ultasonication, and the crude extract was centrifuged at 10,000g for 15 min. 608
6-Histidine tagged proteins were mostly recovered in the soluble fraction. Soluble 609
proteins were then and purified using fast flow chelating sepharose resin (Amersham) 610
according to manufacturer’s instructions. Ultrapure RemCA peptides were obtained 611
by de novo peptide synthesis (GL Biochem Shanghai Ltd; www.glschina.com). 612
Western blot analyses were performed using either anti-REM (Raffaele et al., 2009), 613
anti-PMA (Lefebvre et al., 2004), anti-PIP (Santoni et al., 2003), anti-actin or 614
commercial anti-GFP (Millipore/Roche) antibodies. 615
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

21
616
Extrinsic PM proteins stripping 617
Stripping on PM preparations and microsomal fractions was performed as described 618
in (Santoni et al., 2003) with minor modifications. Membranes (200 μg) were 619
incubated in 40 ml/1 ml of 5 mM EDTA, 5 mM EGTA, 4 M urea and 5 mM Tris-HCl, 620
pH 9.5, for 5 min on ice before being centrifuged for 20 min at 100,000 g. The 621
subsequent pellet was suspended in 20 mM NaOH, and centrifuged at 100,000 g for 622
20 min. The membranes were then washed in 2 mM EDTA, 2 mM EGTA, 100 mM 623
NaCl and 5 mM Tris-HCl, pH 8, centrifuged at 100 000 g for 20 min, and finally 624
resuspended in loading buffer for SDS-PAGE and Western blot where samples were 625
loaded at equal volumes. The mean protein-purification yield was ~50%. A negative 626
control treated with TBS was added (mock), yielding 95% recovery. Total protein 627
amounts were quantified by Bradford or by image quantification of the Coomassie-628
stained gels. 629
630
Liposome and lipid binding assays 631
Liposomes mimicking the lipid content of tobacco PM were freshly prepared in TBS 632
buffer according to (Doms et al., 1985). For PM-like liposome, the lipid content was 633
phosphatidylethanolamine (PE)/ phosphatidylcholine (PC)/ phosphatidic acid (PA) / 634
glucosylcerebrosides (gluCER) / tobacco mix of free sterols / acyl steryl glucosides 635
(ASG)/ steryl glucosides (SG) at molar ratio 1/0.6/0.2/0.6/0.8/0.5/0.5 and for DIM-like 636
liposomes same lipids at molar ratio 1/0.6/0.2/1.2/2.5/3/1 (Mongrand et al., 2004; Furt 637
et al., 2010). Signals were quantified by the ImageJ software. Standard plant lipids 638
were purchased from SIGMA and MATREYA. 5 µg of purified 6His:StREM1.3, 639
6His:StREM1.31-170 or 6His:StREM1.3* was incubated for 30 min at 30°C with 5 mM 640
liposomes. The samples were further processed in 62% sucrose, and placed at the 641
bottom of a discontinuous sucrose gradient (35/30/15/5% w/w). After centrifugation, 642
liposomes floating at the 15-5% interface (top of the gradient) and fractions at the 643
bottom of the gradient were collected, proteins were precipitated with 15% 644
trichloracetic acid (TCA) and analysed by western blot. To control for liposome 645
recovery, lipids were extracted and analysed by Thin Layer Chromatography as 646
described previously (Mongrand et al., 2004). 647
648
Circular dichroism (CD) measurements 649
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

22
CD spectra were recorded on a Jasco J-815 CD spectrometer with 10-mm path 650
length quartz cuvettes. Ten scans were taken and automatically averaged in the 651
wavelength range from 190 to 250 nm. Peptide secondary structures were 652
determined using CDpro software package, which involved CDSSTR, SELCON3, 653
and CONTINLL methods (Adam et al., 2004; Zakharov et al., 2004). Percentages 654
were calculated by averaging the percentages provided by the three methods. The 655
peptides stock solutions used for the measurements were diluted in 1 mM Tris buffer 656
at pH 7.4, or in TFE. 657
658
Liposome leakage assays 659
Membrane perturbation and vesicle release can be measured by the assay of (Ellens 660
et al., 1985) based on the quenching of HPTS by DPX. HPTS and DPX were co-661
encapsulated in the aqueous phase of the same liposomes. When leakage of vesicle 662
content occurs, the quenching by DPX stops and subsequently the fluorescence of 663
HPTS increases. Small unilamellar vesicles (SUVs) were used in our experiments. 664
These vesicles were prepared from a solution of multilamellar vesicles (MLV) 665
obtained after hydration for 1 h at 37°C of dry lipid films. These films were mixtures 666
by weight of 26.6 % PC, 26.6 % SM, 26.6 % PE, and 20.2 % CHOL for neutral 667
liposomes and 30 % PC, 30 % PE, 2.5 % PI, 10 % PS, 5 % SM, and 22.5% CHOL for 668
negatively charged liposomes. Large unilamellar vesicles (LUVs) were prepared by 669
the extrusion technique of Mayer et al. The MLV suspension was submitted to five 670
successive cycles of freezing and thawing and thereafter extruded 10 times through 671
stacked polycarbonate filters (pore size, 0.08 μm), under a nitrogen pressure of 20 672
bars using an extruder (Lipex Biomembranes, Vancouver Canada). The 673
concentration of the liposome suspensions was determined by phosphorus analysis. 674
The peptides, dissolved in TFE or HFP/TFE, were added to a mixture of liposomes 675
with encapsulated HPTS and DPX. HPTS fluorescence was measured on a Perkin-676
Elmer LS- 50B fluorimeter (excitation, 360 nm; emission, 520 nm). Liposomes were 677
prepared as described above but were rehydrated with 1 ml of 12.5 mM HPTS (45 678
mM NaCl), 45 mM DPX (20 mM NaCl), and 10 mM Tris-HCl at pH 7.4 and passed 679
through a Sephadex G-75 column to removed un-encapsulated material. 680
681
Lipid mixing assays 682
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

23
The induction of vesicular lipid mixing by the different peptides was tested with 683
liposomes (SUVs) made of PC/PE/SM/chol at 26.6/26.6/26.6/20.2 molar ratio and 684
PC/PE/PI/PS/SM/CHOL at a molar ratio 30/30/2.1/10/5/22.5. Labelled liposomes 685
were obtained by incorporating R18 in the dry lipid film at a concentration of 6.3 % of 686
the total lipid weight. Labelled and unlabelled liposomes were mixed at a weight ratio 687
of 1:4 and a final concentration of 50 mM in 10mM Tris-HCl, 150 mM NaCl, 0.01% 688
EDTA, 1 mM NaN3 (pH 8). Mixing of liposome membranes was followed by 689
measuring the fluorescence increase of R18, a lipid soluble probe, occurring after the 690
fusion of labelled and unlabelled liposomes, as described in (Lins et al., 2002). 691
Incubation of labelled and unlabelled vesicles in buffer alone did not modify the 692
fluorescence intensity. Fluorescence was recorded at room temperature (excitation: 693
560 nm, emission: 590 nm) on an LS-50B PerkinElmer fluorimeter. 694
695
Size-exclusion chromatography 696
Size-exclusion chromatography was performed on an AKTApurifier apparatus (GE 697
Healthcare). 6His:StREM1.3 and 6his:StREM1.3* purified from E. Coli cultures 698
(1.2g/l) were centrifuged for 5 min at 20,000 g before being processed. The 699
molecular size of the proteins was analysed by chromatography on a Superdex 75 700
10/30 column (GE Healthcare) equilibrated with 20 mM of HEPEs, pH 7,4, 300mM 701
NaCl. Proteins (200 µl) were eluted with a flow rate of 0.2 ml/min and recorded by 702
continuously monitoring the absorbance at 280nm. The column was calibrated with 703
the standard proteins described in Fig.6. 704
705
Viral spreading and split ubiquitin assays 706
To assess spreading of PVX:GFP in N. benthamiana leaves Agrobacterium 707
tumefaciens strain GV3101 carrying the respective StREM1.3 constructs were 708
infiltrated at a final OD600= 0.2 together with the same strain carrying the plasmid 709
pGr208, which expresses the PVX:GFPcDNA, as well as the helper plasmid pSoup 710
(Peart et al., 2002) at final OD600 = 0.001. Spreading of PVX:GFP was visualized by 711
confocal laser-scanning microscopy at 5 days post infection and the number of 712
fluorescent cells of at least 30 PVX:GFP infection foci were counted. Protein extracts 713
from the infiltrated N. benthamiana leaves were subjected to cell fractionation and 714
Western Blot. The experiment was repeated 3 times with same results. Split ubiquitin 715
assays were performed as described in (Raffaele et al., 2009) with yeasts grown on 716
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

24
SD medium supplement with uracile (50 mg.L-1), leucine (380 mg.L-1), lysine (76 717
mg.L-1) and 5’fluoro-orotic acid (1 g.L-1). 718
719
ACKNOWLEDGEMENTS 720
We would like to thank Mohammed-Amine Belka, Christelle Flore and Jeanny 721
Laroche-Traineau their valuable contributions. We are very grateful to Claudia Popp 722
and Thomas Ott (LMU Munich) for inspiring discussions, shared data, contagious 723
enthusiasm, and a lot of help. We thank all other members of the labs of Sébastien 724
Mongrand (LBM Bordeaux) and Thomas Ott (LMU Munich) for fruitful discussions. 725
We acknowledge the platform Métabolome- Fluxome-Lipidome of Bordeaux 726
(http://www.biomemb.cnrs.fr/INDEX.html 727
https://www.bordeaux.inra.fr/umr619/RMN_index.htm) for contribution to lipid 728
analysis and the unit of plant imaging of Bordeaux Imaging Center (BIC) for confocal 729
imaging. This work has been funded by the French National Agency for Research, 730
ANR program (NT09_517917, ANR PANACEA to SM, JLC). 731
AUTHOR CONTRIBUTION 732
A.P., L.L., S.M. and S.R. designed research. A.P., JL.C., JM.C., S.M. and S.R. 733
performed research. JM.C., L.L. and S.G-R. contributed new reagents and analytic 734
tools. A.P., JM.C, S.M. and S.R. analysed data. S.M. and S.R. wrote the paper. All 735
authors read, edited and approved the manuscript. 736
737
CONFLICT OF INTEREST 738
The authors declare no conflict of interest. 739
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

25
REFERENCES 740
Adam B, Lins L, Stroobant V, Thomas A, Brasseur R (2004) Distribution of hydrophobic residues is 741 crucial for the fusogenic properties of the Ebola virus GP2 fusion peptide. J Virol 78: 2131-742 2136 743
Bariola P, Retelska D, Stasiak A, Kammerer R, Fleming A, Hijri M, Frank S, Farmer E (2004) 744 Remorins form a novel family of coiled coil-forming oligomeric and filamentous proteins 745 associated with apical, vascular and embryonic tissues in plants. Plant molecular biology 55: 746 579-594 747
Batoko H, Zheng HQ, Hawes C, Moore I (2000) A rab1 GTPase is required for transport between the 748 endoplasmic reticulum and golgi apparatus and for normal golgi movement in plants. Plant 749 Cell 12: 2201-2218 750
Baumgart T, Hess ST, Webb WW (2003) Imaging coexisting fluid domains in biomembrane models 751 coupling curvature and line tension. Nature 425: 821-824 752
Bhat RA, Miklis M, Schmelzer E, Schulze-Lefert P, Panstruga R (2005) Recruitment and 753 interaction dynamics of plant penetration resistance components in a plasma membrane 754 microdomain. Proceedings of the National Academy of Sciences of the United States of 755 America 102: 3135 756
Bigay J, Casella JF, Drin G, Mesmin B, Antonny B (2005) ArfGAP1 responds to membrane 757 curvature through the folding of a lipid packing sensor motif. Embo J 24: 2244-2253 758
Brown DA (2006) Lipid rafts, detergent-resistant membranes, and raft targeting signals. Physiology 759 21: 430 760
Cho W, Stahelin RV (2005) Membrane-protein interactions in cell signaling and membrane trafficking. 761 Annu. Rev. Biophys. Biomol. Struct. 34: 119-151 762
Cordy JM, Hussain I, Dingwall C, Hooper NM, Turner AJ (2003) Exclusively targeting beta-763 secretase to lipid rafts by GPI-anchor addition up-regulates-site processing of the amyloid 764 precursor protein. Proceedings of the National Academy of Sciences 100: 11735 765
Cross KJ, Wharton SA, Skehel JJ, Wiley DC, Steinhauer DA (2001) Studies on influenza 766 haemagglutinin fusion peptide mutants generated by reverse genetics. The EMBO journal 20: 767 4432-4442 768
De Matteis MA, Godi A (2004) PI-loting membrane traffic. Nature cell biology 6: 487-492 769
Dietrich C, Bagatolli L, Volovyk Z, Thompson N, Levi M, Jacobson K, Gratton E (2001) Lipid rafts 770 reconstituted in model membranes. Biophysical journal 80: 1417-1428 771
Doms RW, Helenius A, White J (1985) Membrane fusion activity of the influenza virus hemagglutinin. 772 The low pH-induced conformational change. J Biol Chem 260: 2973-2981 773
Drin G, Antonny B (2009) Amphipathic helices and membrane curvature. FEBS Lett 584: 1840-1847 774
Drin G, Casella JF, Gautier R, Boehmer T, Schwartz TU, Antonny B (2007) A general amphipathic 775 -helical motif for sensing membrane curvature. Nature 200: 7 776
Ellens H, Bentz J, Szoka FC (1985) H+- and Ca2+-induced fusion and destabilization of liposomes. 777 Biochemistry 24: 3099-3106 778
Ford MGJ, Mills IG, Peter BJ, Vallis Y, Praefcke GJK, Evans PR, McMahon HT (2002) Curvature 779 of clathrin-coated pits driven by epsin. Nature 419: 361-366 780
Furt F, Konig S, Bessoule JJ, Sargueil F, Zallot R, Stanislas T, Noirot E, Lherminier J, Simon-781 Plas F, Heilmann I, Mongrand S (2010) Polyphosphoinositides are enriched in plant 782 membrane rafts and form microdomains in the plasma membrane. Plant Physiol 152: 2173-783 2187 784
Gruenberg J (2003) Lipids in endocytic membrane transport and sorting. Current opinion in cell 785 biology 15: 382-388 786
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

26
Han X, Bushweller JH, Cafiso DS, Tamm LK (2001) Membrane structure and fusion-triggering 787 conformational change of the fusion domain from influenza hemagglutinin. Nature Structural & 788 Molecular Biology 8: 715-720 789
Heo WD, Inoue T, Park WS, Kim ML, Park BO, Wandless TJ, Meyer T (2006) PI (3, 4, 5) P3 and PI 790 (4, 5) P2 lipids target proteins with polybasic clusters to the plasma membrane. Science 314: 791 1458 792
Hope HR, Pike LJ (1996) Phosphoinositides and phosphoinositide-utilizing enzymes in detergent-793 insoluble lipid domains. Mol Biol Cell 7: 843-851 794
Jarsch IK, Ott T (2011) Perspectives on Remorin Proteins, Membrane Rafts, and Their Role During 795 Plant-Microbe Interactions. Molecular Plant-Microbe Interactions 24: 7-12 796
Keinath NF, Kierszniowska S, Lorek J, Bourdais G, Kessler SA, Shimosato-Asano H, 797 Grossniklaus U, Schulze WX, Robatzek S, Panstruga R (2010) PAMP (pathogen-798 associated molecular pattern)-induced changes in plasma membrane compartmentalization 799 reveal novel components of plant immunity. Journal of Biological Chemistry 285: 39140 800
Kierszniowska S, Seiwert B, Schulze WX (2009) Definition of Arabidopsis sterol-rich membrane 801 microdomains by differential treatment with methyl-beta-cyclodextrin and quantitative 802 proteomics. Mol Cell Proteomics 8: 612-623 803
Kutateladze TG (2010) Translation of the phosphoinositide code by PI effectors. Nature chemical 804 biology 6: 507-513 805
Lai AL, Tamm LK (2007) Locking the kink in the influenza hemagglutinin fusion domain structure. 806 Journal of Biological Chemistry 282: 23946 807
Lasserre R, Guo XJ, Conchonaud F, Hamon Y, Hawchar O, Bernard AM, Soudja SMH, Lenne 808 PF, Rigneault H, Olive D (2008) Raft nanodomains contribute to Akt/PKB plasma membrane 809 recruitment and activation. Nature chemical biology 4: 538-547 810
Lefebvre B, Batoko H, Duby G, Boutry M (2004) Targeting of a Nicotiana plumbaginifolia H+ -811 ATPase to the plasma membrane is not by default and requires cytosolic structural 812 determinants. Plant Cell 16: 1772-1789 813
Lefebvre B, Furt F, Hartmann MA, Michaelson LV, Carde JP, Sargueil-Boiron F, Rossignol M, 814 Napier JA, Cullimore J, Bessoule JJ, Mongrand S (2007) Characterization of lipid rafts 815 from Medicago truncatula root plasma membranes: a proteomic study reveals the presence of 816 a raft-associated redox system. Plant Physiol 144: 402-418 817
Lefebvre B, Timmers T, Mbengue M, Moreau S, Herve C, Toth K, Bittencourt-Silvestre J, Klaus 818 D, Deslandes L, Godiard L, Murray JD, Udvardi MK, Raffaele S, Mongrand S, Cullimore 819 J, Gamas P, Niebel A, Ott T (2010) A remorin protein interacts with symbiotic receptors and 820 regulates bacterial infection. Proc Natl Acad Sci U S A 107: 2343-2348 821
Lemmon MA (2008) Membrane recognition by phospholipid-binding domains. Nature Reviews 822 Molecular Cell Biology 9: 99-111 823
Lewis JD, Lazarowitz SG (2010) Arabidopsis synaptotagmin SYTA regulates endocytosis and virus 824 movement protein cell-to-cell transport. Proceedings of the National Academy of Sciences 825 107: 2491-2496 826
Lingwood D, Simons K (2010) Lipid rafts as a membrane-organizing principle. Science 327: 46-50 827
Lins L, Brasseur R (2008) Tilted peptides: a structural motif involved in protein membrane insertion? 828 J Pept Sci 14: 416-422 829
Lins L, Flore C, Chapelle L, Talmud PJ, Thomas A, Brasseur R (2002) Lipid-interacting properties 830 of the N-terminal domain of human apolipoprotein C-III. Protein Eng 15: 513-520 831
Liu J, Elmore JM, Fuglsang AT, Palmgren MG, Staskawicz BJ, Coaker G (2009) RIN4 functions 832 with plasma membrane H+-ATPases to regulate stomatal apertures during pathogen attack. 833 PLoS biology 7: e1000139 834
Longo ML, Waring AJ, Hammer DA (1997) Interaction of the influenza hemagglutinin fusion peptide 835 with lipid bilayers: area expansion and permeation. Biophys J 73: 1430-1439 836
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

27
Lorieau JL, Louis JM, Bax A (2010) The complete influenza hemagglutinin fusion domain adopts a 837 tight helical hairpin arrangement at the lipid:water interface. Proc Natl Acad Sci U S A 107: 838 11341-11346 839
Lorin A, Lins L, Stroobant V, Brasseur R, Charloteaux B (2008) The minimal fusion peptide of 840 simian immunodeficiency virus corresponds to the 11 first residues of gp32. Journal of Peptide 841 Science 14: 423-428 842
Marín M, Ott T (2012) Phosphorylation of intrinsically disordered regions in remorin proteins. Front 843 Plant Sci. 3:86. 844
Maule AJ (2008) Plasmodesmata: structure, function and biogenesis. Current opinion in plant biology 845 11: 680-686 846
McLaughlin S, Murray D (2005) Plasma membrane phosphoinositide organization by protein 847 electrostatics. Nature 438: 605-611 848
McLaughlin S, Wang J, Gambhir A, Murray D (2002) PIP2 and proteins: interactions, organization, 849 and information flow. Annual review of biophysics and biomolecular structure 31: 151-175 850
Mima J, Wickner W (2009) Complex lipid requirements for SNARE- and SNARE chaperone-851 dependent membrane fusion. J Biol Chem 284: 27114-27122 852
Minami A, Fujiwara M, Furuto A, Fukao Y, Yamashita T, Kamo M, Kawamura Y, Uemura M (2009) 853 Alterations in detergent-resistant plasma membrane microdomains in Arabidopsis thaliana 854 during cold acclimation. Plant Cell Physiol 50: 341-359 855
Mongrand S, Morel J, Laroche J, Claverol S, Carde JP, Hartmann MA, Bonneu M, Simon-Plas F, 856 Lessire R, Bessoule JJ (2004) Lipid rafts in higher plant cells: purification and 857 characterization of Triton X-100-insoluble microdomains from tobacco plasma membrane. J 858 Biol Chem 279: 36277-36286 859
Mongrand S, Stanislas T, Bayer EMF, Lherminier J, Simon-Plas F (2010) Membrane rafts in plant 860 cells. Trends in plant science 15: 656-663 861
Munro S (2003) Lipid rafts: elusive or illusive? Cell 115: 377-388 862
Paulick MG, Bertozzi CR (2008) The Glycosylphosphatidylinositol Anchor: A Complex Membrane-863 Anchoring Structure for Proteins†. Biochemistry 47: 6991-7000 864
Peart JR, Lu R, Sadanandom A, Malcuit I, Moffett P, Brice DC, Schauser L, Jaggard DAW, Xiao 865 S, Coleman MJ (2002) Ubiquitin ligase-associated protein SGT1 is required for host and 866 nonhost disease resistance in plants. Proceedings of the National Academy of Sciences 99: 867 10865 868
Peter BJ, Kent HM, Mills IG, Vallis Y, Butler PJ, Evans PR, McMahon HT (2004) BAR domains as 869 sensors of membrane curvature: the amphiphysin BAR structure. Science 303: 495-499 870
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE (2004) 871 UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem 872 25: 1605-1612 873
Pike LJ, Han X, Chung KN, Gross RW (2002) Lipid rafts are enriched in arachidonic acid and 874 plasmenylethanolamine and their composition is independent of caveolin-1 expression: a 875 quantitative electrospray ionization/mass spectrometric analysis. Biochemistry 41: 2075-2088 876
Porter MY, Koelle MR (2010) RSBP-1 Is a Membrane-targeting Subunit Required by the G {alpha} q-877 specific But Not the G {alpha} o-specific R7 Regulator of G protein Signaling in Caenorhabditis 878 elegans. Molecular biology of the cell 21: 232 879
Raffaele S, Bayer E, Lafarge D, Cluzet S, German Retana S, Boubekeur T, Leborgne-Castel N, 880 Carde JP, Lherminier J, Noirot E, Satiat-Jeunemaitre B, Laroche-Traineau J, Moreau P, 881 Ott T, Maule AJ, Reymond P, Simon-Plas F, Farmer EE, Bessoule JJ, Mongrand S (2009) 882 Remorin, a solanaceae protein resident in membrane rafts and plasmodesmata, impairs 883 potato virus X movement. Plant Cell 21: 1541-1555 884
Raffaele S, Mongrand S, Gamas P, Niebel A, Ott T (2007) Genome-wide annotation of remorins, a 885 plant-specific protein family: evolutionary and functional perspectives. Plant Physiol 145: 593-886 600 887
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

28
Reymond P, Kunz B, Paul-Pletzer K, Grimm R, Eckerskorn C, Farmer EE (1996) Cloning of a 888 cDNA encoding a plasma membrane-associated, uronide binding phosphoprotein with 889 physical properties similar to viral movement proteins. Plant Cell 8: 2265-2276 890
Rossin A, Kral R, Lounnas N, Chakrabandhu K, Mailfert S, Marguet D, Hueber AO (2010) 891 Identification of a lysine-rich region of Fas as a raft nanodomain targeting signal necessary for 892 Fas-mediated cell death. Experimental cell research 316: 1513-1522 893
Roy A, Kucukural A, Zhang Y (2010) I-TASSER: a unified platform for automated protein structure 894 and function prediction. Nat Protoc 5: 725-738 895
Sankarshanan M, Ma Z, Iype T, Lorenz U (2007) Identification of a novel lipid raft-targeting motif in 896 Src homology 2-containing phosphatase 1. The Journal of Immunology 179: 483 897
Santoni V, Vinh J, Pflieger D, Sommerer N, Maurel C (2003) A proteomic study reveals novel 898 insights into the diversity of aquaporin forms expressed in the plasma membrane of plant 899 roots. Biochem J 373: 289-296 900
Schapire AL, Voigt B, Jasik J, Rosado A, Lopez-Cobollo R, Menzel D, Salinas J, Mancuso S, 901 Valpuesta V, Baluska F (2008) Arabidopsis synaptotagmin 1 is required for the maintenance 902 of plasma membrane integrity and cell viability. The Plant Cell 20: 3374-3388 903
Schneidman-Duhovny D, Inbar Y, Nussinov R, Wolfson HJ (2005) PatchDock and SymmDock: 904 servers for rigid and symmetric docking. Nucleic acids research 33: W363-W367 905
Seong J, Ouyang M, Kim T, Sun J, Wen PC, Lu S, Zhuo Y, Llewellyn NM, Schlaepfer DD, Guan 906 JL (2011) Detection of focal adhesion kinase activation at membrane microdomains by 907 fluorescence resonance energy transfer. Nature Communications 2: 406 908
Simon-Plas F, Perraki A, Bayer E, Gerbeau-Pissot P, Mongrand S (2011) An update on Plant 909 Membrane rafts. Current opinion in plant biology 14: 642-649 910
Simons K, Gerl MJ (2010) Revitalizing membrane rafts: new tools and insights. Nature Reviews 911 Molecular Cell Biology 11: 688-699 912
Stahelin RV, Long F, Peter BJ, Murray D, De Camilli P, McMahon HT, Cho W (2003) Contrasting 913 membrane interaction mechanisms of AP180 N-terminal homology (ANTH) and epsin N-914 terminal homology (ENTH) domains. Journal of Biological Chemistry 278: 28993 915
Suzuki M, Jeong SY, Karbowski M, Youle RJ, Tjandra N (2003) The solution structure of human 916 mitochondria fission protein Fis1 reveals a novel TPR-like helix bundle. J Mol Biol 334: 445-917 458 918
Tanner W, Malinsky J, Opekarová M (2011) In plant and animal cells, detergent-resistant 919 membranes do not define functional membrane rafts. The Plant Cell 23: 1191-1193 920
Thompson GA, Jr., Okuyama H (2000) Lipid-linked proteins of plants. Prog Lipid Res 39: 19-39 921
Tóth K, Stratil TF, Madsen EB, Ye J, Popp C, Antolín-Llovera M, Grossmann C, Jensen ON, 922 Schüßler A, Parniske M (2012) Functional Domain Analysis of the Remorin Protein 923 LjSYMREM1 in Lotus japonicus. PLoS One 7: e30817 924
Verchot-Lubicz J, Torrance L, Solovyev AG, Morozov SY, Jackson AO, Gilmer D (2010) Varied 925 movement strategies employed by triple gene block-encoding viruses. Molecular Plant-926 Microbe Interactions 23: 1231-1247 927
Vermeer JE, Thole JM, Goedhart J, Nielsen E, Munnik T, Gadella TW, Jr. (2009) Imaging 928 phosphatidylinositol 4-phosphate dynamics in living plant cells. Plant J 57: 356-372 929
Widjaja I, Naumann K, Roth U, Wolf N, Mackey D, Dangl JL, Scheel D, Lee J (2009) Combining 930 subproteome enrichment and Rubisco depletion enables identification of low abundance 931 proteins differentially regulated during plant defense. Proteomics 9: 138-147 932
Yamazaki T, Kawamura Y, Minami A, Uemura M (2008) Calcium-dependent freezing tolerance in 933 Arabidopsis involves membrane resealing via synaptotagmin SYT1. The Plant Cell 20: 3389-934 3404 935
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

29
Yamazaki T, Takata N, Uemura M, Kawamura Y (2010) Arabidopsis synaptotagmin SYT1, a type I 936 signal-anchor protein, requires tandem C2 domains for delivery to the plasma membrane. 937 Journal of Biological Chemistry 285: 23165 938
Zakharov SD, Kotova EA, Antonenko YN, Cramer WA (2004) On the role of lipid in colicin pore 939 formation. Biochim Biophys Acta 1666: 239-249 940
941
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

30
FIGURE LEGENDS 942
943 Figure 1. A C-terminal anchor peptide (RemCA) from StREM1.3 Remorin is 944
required and sufficient for strong plasma membrane (PM) anchoring in vivo. (A) 945
Predicted secondary structure and protein domains in StREM1.3. Numbers indicate 946
amino acid position. (B) Stripping of peripheral PM proteins is not sufficient to wash 947
out StREM1.3. Purified PMs from tobacco leaves were washed according to Santoni 948
et al. 2003 (Stripped) or treated with TBS (Mock). In average, 50% (n=3) of the PM-949
associated proteins, such as Actin, were stripped whereas the quantity of Remorin 950
protein associated to the PM was unaffected, as for the Plasma membrane Intrinsic 951
Protein 2 (PIP2) aquaporin intrinsic PM protein. (C) Subcellular localization of YFP-952
tagged StREM1.3, truncated StREM1.3 comprising amino-acids 1 to 170, and 953
truncated StREM1.3 comprising amino acids 171 to 198 (RemCA peptide only) 954
transiently expressed in tobacco epidermal cells. Scale bars indicate 20 µm. (D) Cell 955
fractionation of the constructs shown in C. Crude protein extracts (CE) from tobacco 956
leaf cells expressing each construct were fractionated into soluble (sol.) and 957
microsomal (µ) compartments by centrifugation (15µg of proteins per lane). In spite of 958
various attempts to solve this problem, we were not able to prevent partial cleavage 959
of the YFP:RemCA construct, resulting in a free YFP signal as shown. 960
961
Figure 2. Non-polar environments trigger conformational changes in the 962
RemCA peptide. (A) Circular dichroism spectra of synthetic ultrapure RemCA 963
peptide in various environments. The spectral signatures of alpha helices (at 195, 964
208 and 222nm) are indicated by arrows. Spectra were recorded in aqueous (100% 965
Tris-HCl), semi-hydrophobic (50% TFE) and hydrophobic (100% TFE) environments. 966
(B) Estimated distribution of secondary folds in RemCA peptide in the different 967
environments assayed. (C) Sequence and predicted structure of the RemCA peptide. 968
Homology modeling predicts a tight hairpin of alpha helices for RemCA, consistent 969
with the circular dischroism spectra collected in hydrophobic environments. The 970
hydrophobicity of residues (Kyte-Doolitle scale) is indicated by a color-code. TFE, 971
2,2,2 trifluoroethanol. 972
973
Figure 3. Amphipathy of RemCA alpha-helices plays a critical role in anchoring 974
StREM1.3 Remorin to the plasma membrane. (A) A helical wheel projection of 975
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

31
RemCA as a hairpin of α-helices showing amphipathy of each helix. Overall helix 976
hydrophobicity (H) and hydrophobic moment (µH) are indicated in the middle of the 977
helices, with an arrow showing the direction and intensity of the hydrophobic 978
moments. (B) Ribbon and surface models of the RemCA and mutated RemCA* 979
peptides with hydrophobicity (Kyte-Doolitle scale) shown by a color-code. The 980
surface model of RemCA shows the predicted hydrophobic core in red, and charged 981
surfaces in blue. Substitutions introduced in the RemCA* mutant aimed at reducing 982
the amphipathy of the α-helices by altering the distribution of hydrophobic residues 983
(Y184V, A190S, and L195S) and positively charged residues (A186S, G188V and 984
G196V). (C) Maximum projection of confocal scans through N. benthamiana 985
epidermal cells expressing YFP:StREM1.3* mutant, with plastids shown in red. Scale 986
bar indicates 20 µm. (D) Cell fractionation analysis of cells expressing GFP tagged 987
StREM1.3 with native (+) and mutated (*) RemCA sequence, confirming that 988
mutations introduced in StREM1.3* abolish PM localization. 989
990
Figure 4. RemCA confers intrinsic lipid binding to StREM1.3 Remorin. (A) 991
Affinity purified 6His:StREM1.3 (WT) binds to protein-free liposomes in vitro in a 992
RemCA-dependent manner. Liposome binding is abolished in 6His:StREM1.3 993
recombinant protein lacking RemCA (∆CA) or harbouring a mutated RemCA (*). The 994
soluble (Sol.) and liposome-bound (Bound) fractions recovered after centrifugation 995
are shown. Lipid mixing assays (B) and liposome leakage assays (C) show that 996
RemCA ultrapure peptides insert into liposome membranes containing negatively 997
charged lipids, as indicated by an increase in fluorescence de-quenching. By 998
contrast, liposome leakage assays (C) showed that RemCA is not able to insert into 999
neutral liposome membranes. SIV, Simian influenza virus fusion peptide; TFE, 1000
trifluoroethanol; TX100, Triton X-100. Solvent used in (C) was dimethyl sulfoxide for 1001
negatively charged liposomes and TFE for neutral liposomes. Error bars show 1002
standard deviation of 6-8 independent measurements. 1003
1004
Figure 5. RemCA targets soluble proteins to the PM in a lipid-specific manner. 1005
(A) Lipid overlay assays performed with 6His tagged StREM1.3 and StREM1.31-170 1006
show a differential affinity of StREM1.3 for various classes of lipids, dependent on 1007
RemCA. The right panel shows Western Blot of the protein input material illustrating 1008
equal loading for StREM1.3 and StREM1.31-170. PA, phosphatidic acid; PIP3, 1009
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

32
phosphoinositol 3,4,5, triphosphate; PIP2, Phosphoinositol bisphosphate, the position 1010
of phosphate groups is indicated below; S1P, sphingosine 1-phosphate. (B) 1011
6His:StREM1.3 shows higher affinity for liposomes with a composition mimicking 1012
detergent insoluble membranes (DIM) compared to plasma membrane (PM - 1013
described in materials and methods). Histograms show quantifications and standard 1014
error based on Western blot signals for three independent experiments. Differential 1015
affinity is significant with p<0.05 in a Student t-test. (C) RemCA-tagged GFP 1016
transiently expressed in tobacco leaves segregates into the DIM fractions of the PM, 1017
similar to the endogenous Remorin. Total proteins were quantified by bicinchoninic 1018
acid assay (BCA) relative to total proteins in purified PM (error bars show standard 1019
deviation from three independent replicates). µ, microsomal fraction; DSM, detergent 1020
soluble membranes. 1021
1022
Figure 6. StREM1.3 Remorin forms homo-trimers independently of RemCA in 1023
vitro. (A) Exclusion column chromatography of 6His:StREM1.3, 6His:StREM1.3* and 1024
elution standards allows to estimate the native MW of 6His:StREM1.3 to ~120 kDA, 1025
suggesting that it assembles into trimers, and shows that mutations introduced in 1026
6His:StREM1.3* do not alter the ability to form trimers. (B) A trimer model of the C-1027
terminal domain of StREM1.3. Views from the C-terminal end of the trimer with 1028
residues colored according to hydrophobicity (Kyte-Doolitle scale) reveal the 1029
formation of a hydrophobic core (in red) at the centre of the trimer, with positive 1030
surface patches surrounding the RemCA region, in blue. Standards used in A: 1, blue 1031
dextran (2MDa); 2, Thyroglobuline (670kDa); 3, Catalase (232kDa); 4, Ovalbumine 1032
(43kDa); 5, Chymotrypsinogene (25kDa); 6, soybean Trypsin inhibitor (21.5kDa); 7, 1033
Ribonuclease (13.7kDa); 8, Cytochrome C (12.3kDa). 1034
1035
Figure 7. RemCA-mediated plasma membrane binding is required for StREM1.3 1036
restriction of PVX cell to cell propagation. (A) Expression of PVX:GFP alone and 1037
in combination with StREM1.3 or the mutated StREM1.3* variant unable to bind to 1038
the plasma membrane. Images of infection foci were taken at 5 days post infection 1039
(dpi) of N. benthamiana leaves. Scale bars indicate 150µm. (B) Quantification of PVX 1040
cell to cell spreading showing the average number of cells in the infection foci at 5 1041
dpi. For each treatment, 90 pictures were analysed, with error bars showing standard 1042
deviation. Significance was assessed by Dunn’s multiple comparison test (***, 1043
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

33
p<0.001; n=3). (C) Split ubiquitin assays showing that StREM1.31-170 and StREM1.3* 1044
interact with PVX TGBp1 movement protein similar to the wild type StREM1.3. 1045
1046
Figure 8. Model for the molecular mechanisms leading to the assembly of 1047
StREM1.3 micro-domains at the plasma membrane (PM). Conformational 1048
changes in RemCA, leading to folding into amphipathic alpha helices, are required to 1049
anchor StREM1.3 to the PM. These conformational changes are triggered by non-1050
polar environments, such as the vicinity of the PM (A). Helix E172-A186 folds 1051
exposing clusters of positively charged residues (+). These residues establish 1052
electrostatic interactions with the negatively charged lipids of the inner leaflet of 1053
membrane rafts, destabilizing the PM and allowing insertion of RemCA in the bilayer 1054
(B). Hydrophobicity triggers RemCA hairpin formation, creating an hydrophobic 1055
pocket (small red star) that deepens and strengthen insertion in the membrane. 1056
Alternatively, an external signal may trigger the formation of StREM1.3 trimers (C). 1057
The hydrophobic core of the trimer (large yellow star) favours folding of helix E172-1058
A186 of RemCA (D), leading to PM anchoring as previously described. The formation 1059
of trimers, either prior to or after PM anchoring, is likely to increase the strength of 1060
the lipid binding, may promote membrane domain aggregation, lead to membrane 1061
deformation or mediate the assembly of protein complexes at the PM via Remorin 1062
protein-protein interaction domains (E). 1063
1064
SUPPLEMENTAL DATA 1065
1066
Figure S1. Membrane washing experiments show that StREM1.3 strongly binds 1067
to purified plasma membrane and liposomes. (A) Anti-Remorin Western blot on 1068
tobacco leaf purified plasma membranes incubated for 30 min in water, in Tris-HCl 1069
buffer saline (TBS) containing NaCl, urea, TBS of increasing pH and TBS containing 1070
Ethylene diamine tetra acetic acid (EDTA). (B) Anti-Remorin Western blot on 1071
liposomes supplemented with 1 µg of purified 6His:StREM1.3 (input). These samples 1072
were incubated in TBS in H2O or in TBS containing 1 M NaCl or 6 M Urea for 30 min 1073
at 4°C, and centrifuged. 1074
1075
Figure S2. Sequence and structure comparison between StREM1.3 RemCA and 1076
Influenza hemagglutinin fusion peptide (HAfp). StREM1.3 homology model (in 1077
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

34
blue) was manually superimposed with HAfp structure (in yellow, Lorieau et al., 1078
2010). Kyte-Doolittle scale is used to illustrate the hydrophobicity of residues along 1079
the alignment and in one of the structure views. 1080
1081
Figure S3. Controls for the production of purified recombinant proteins and 1082
liposome binding assays. (A) 6His-tagged StREM1.3 constructs were expressed in 1083
BL21 DE3 E. Coli cells, with induction controlled by addition of isopropyl β-D-1-1084
thiogalactopyranoside (IPTG - top left panel). Red arrowheads indicate the size 1085
expected for StREM1.3 constructs. Cells were lysed by ultrasonication (CE) and 1086
centrifuged. StREM1.3 variants were mostly recovered in the supernatant (Sup.) but 1087
not the pellet (Pel.). Soluble proteins were then processed using fast flow chelating 1088
sepharose resin with immidazole concentrations as indicated on the figure (mM). (B) 1089
To control the recovery of the liposomes, the lipids of half of the liposomes 1090
supplemented with StREM1.3, StREM1.31-170 or StREM1.3* were extracted and 1091
analysed by Thin Layer Chromatography as described in Mongrand et al., 2004. 1092
Lipids were identified by comparison with commercial standards. ASG, acyl steryl 1093
glucoside; DIM, detergent insoluble membranes; gluCER, glucosyl ceramide; PA, 1094
phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidylethanolamine; SG, 1095
Steryl glucoside. 1096
1097
Figure S4. Controls for lipid-specific binding of StREM1.3. (A) Lipid overlay 1098
assay comparing phospholipid binding properties of recombinant 6His:StREM1.3 and 1099
6His:StREM1.3*. Total protein used in the assay is shown in the top panel. (B) To 1100
control for the recivery of the liposomes in the liposome binding assays, the lipids of 1101
half the liposomes mimicking PM or DIM lipid composition were extracted and 1102
analyzed by Thin Layer Chromatography as described in Mongrand et al. 2004. 1103
Lipids were identified by comparison with commercial standards. ASG, acyl steryl 1104
glucoside; DIM, detergent insoluble membranes; gluCER, glucosyl ceramide; PA, 1105
phosphatidic acid; PC, phosphatidylcholine; PE, phosphatidylethanolamine; SG, 1106
Steryl glucoside. (C) Segregation of the PMA proton pump and Remorin along a 1107
sucrose gradient showing distribution of these proteins between Detergent Insoluble 1108
Membranes (DIM, fraction 1) and Detergent Soluble Membranes (DSM, fractions 2 to 1109
4). 1110
1111
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

35
Figure S5. StREM1.3 Remorin forms trimers in vitro. (A) Exclusion column 1112
chromatogram obtained for 6His:StREM1.3. (B) Fractions A8, A9, B4, B5, C3 and C4 1113
were analysed by SDS-PAGE (denaturing conditions) followed by anti-Remorin 1114
western blot showing that StREM1.3 elutes in fractions B4 and B5. 1115
1116
Figure S6. Western Blot controls for the expression of StREM1.3 and 1117
StREM1.3* in PVX:GFP propagation experiments. Protein extracts (10 µg) from 1118
infected tobacco leaves with PVX:GFP alone or co-expressing StREM1.3 or 1119
StREM1.3* were used for Western Blot analysis. 1120
1121
Supplemental File 1. Protein Databank file containing atom coordinates for the 1122
predicted StREM1.3 structure. 1123
1124
Supplemental File 2. Protein Databank file containing atom coordinates for the 1125
predicted StREM1.3* structure. 1126
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
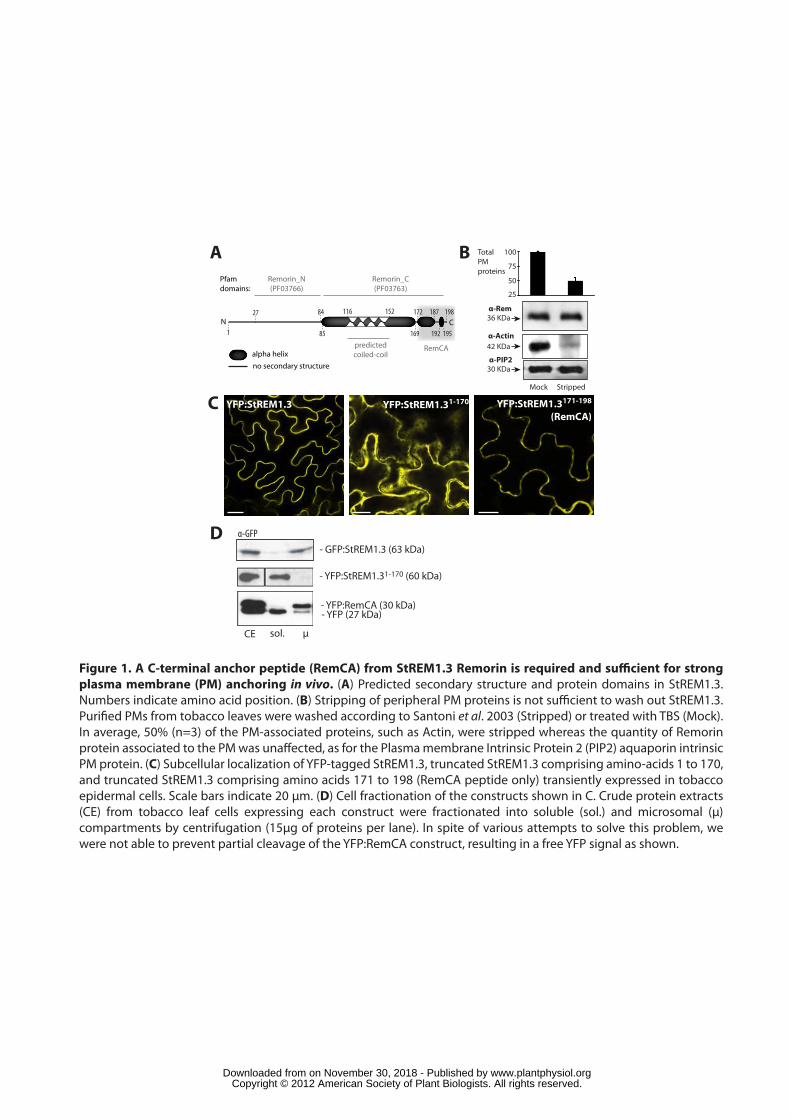
Figure 1. A C-terminal anchor peptide (RemCA) from StREM1.3 Remorin is required and sufficient for strong plasma membrane (PM) anchoring in vivo. (A) Predicted secondary structure and protein domains in StREM1.3. Numbers indicate amino acid position. (B) Stripping of peripheral PM proteins is not sufficient to wash out StREM1.3. Purified PMs from tobacco leaves were washed according to Santoni et al. 2003 (Stripped) or treated with TBS (Mock). In average, 50% (n=3) of the PM-associated proteins, such as Actin, were stripped whereas the quantity of Remorin protein associated to the PM was unaffected, as for the Plasma membrane Intrinsic Protein 2 (PIP2) aquaporin intrinsic PM protein. (C) Subcellular localization of YFP-tagged StREM1.3, truncated StREM1.3 comprising amino-acids 1 to 170, and truncated StREM1.3 comprising amino acids 171 to 198 (RemCA peptide only) transiently expressed in tobacco epidermal cells. Scale bars indicate 20 μm. (D) Cell fractionation of the constructs shown in C. Crude protein extracts (CE) from tobacco leaf cells expressing each construct were fractionated into soluble (sol.) and microsomal (μ) compartments by centrifugation (15μg of proteins per lane). In spite of various attempts to solve this problem, we were not able to prevent partial cleavage of the YFP:RemCA construct, resulting in a free YFP signal as shown.
YFP:StREM1.3 YFP:StREM1.31-170 YFP:StREM1.3171-198
(RemCA)
A B
C
D
100
75
50
25
α-Rem36 KDa
Total PMproteins
StrippedMock
α-Actin42 KDa
α-PIP230 KDa
- GFP:StREM1.3 (63 kDa)
CE sol. μ
α-GFP
- YFP:StREM1.31-170 (60 kDa)
- YFP:RemCA (30 kDa)- YFP (27 kDa)
1
198
85 169
172 187
192 195
alpha helixno secondary structure
116 152
predictedcoiled-coil
Remorin_N(PF03766)
Remorin_C(PF03763)
27 84
RemCA
Pfamdomains:
N C
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
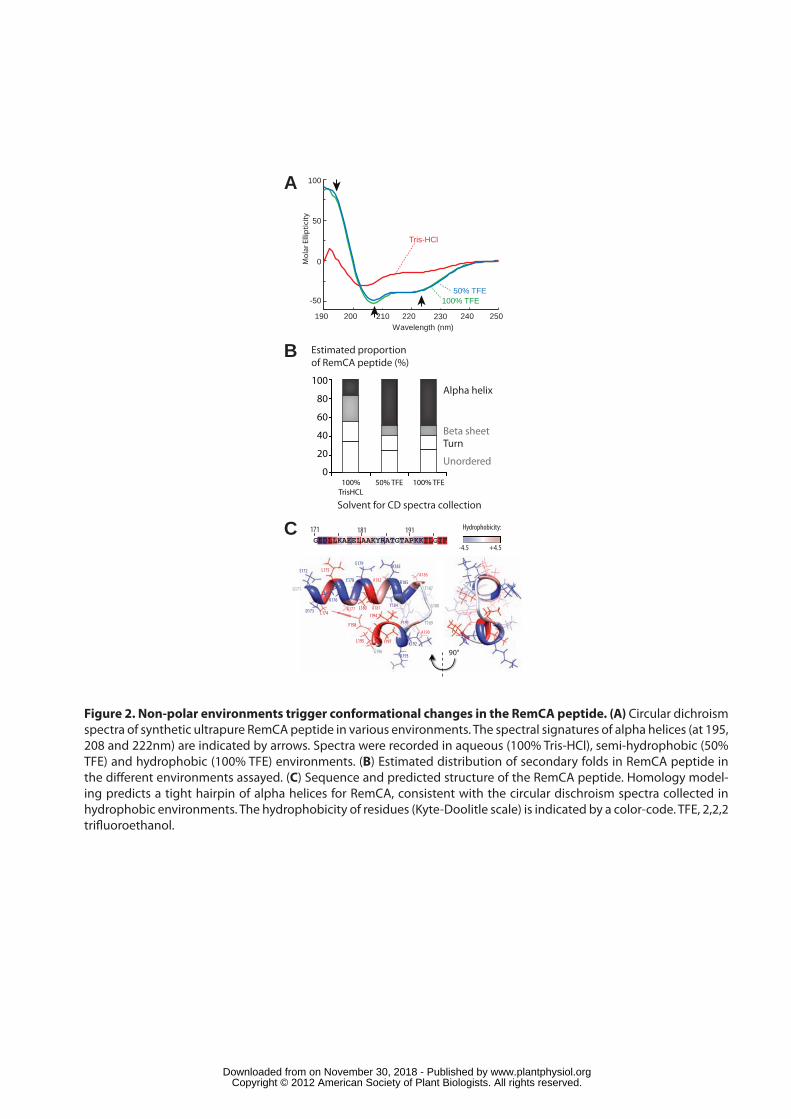
Figure 2. Non-polar environments trigger conformational changes in the RemCA peptide. (A) Circular dichroism spectra of synthetic ultrapure RemCA peptide in various environments. The spectral signatures of alpha helices (at 195, 208 and 222nm) are indicated by arrows. Spectra were recorded in aqueous (100% Tris-HCl), semi-hydrophobic (50% TFE) and hydrophobic (100% TFE) environments. (B) Estimated distribution of secondary folds in RemCA peptide in the di�erent environments assayed. (C) Sequence and predicted structure of the RemCA peptide. Homology model-ing predicts a tight hairpin of alpha helices for RemCA, consistent with the circular dischroism spectra collected in hydrophobic environments. The hydrophobicity of residues (Kyte-Doolitle scale) is indicated by a color-code. TFE, 2,2,2 tri�uoroethanol.
GEDLLKAEELAAKYRATGTAPKKILGIF
Wavelength (nm)190 250200 220 240
100
-50
0
50
Mol
ar E
llipt
icity
B
A
171
G171
210 230
C Hydrophobicity:
-4.5 +4.5
181 191
0
20
40
60
80
100
Estimated proportion of RemCA peptide (%)
100%TrisHCL
50% TFE 100% TFE
Solvent for CD spectra collection
Unordered
TurnBeta sheet
Alpha helix
E172
D173 L174
L175
K176
A177
E178
E179
L180 A181
A182
K183
Y184
R185A186
T187
G188
T189
A190
P191
K192
K193
I194
L195
G196
I197
F198
90°
Tris-HCl
100% TFE50% TFE
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
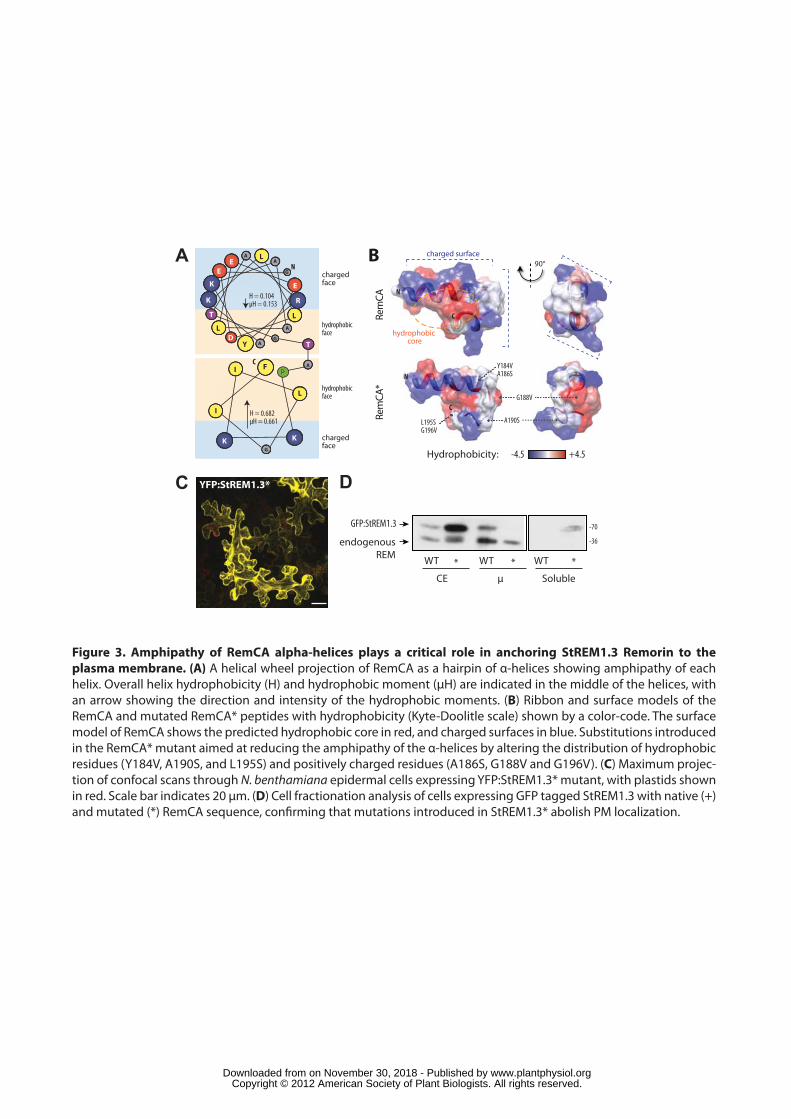
S
YFP:StREM1.3* D
GFP:StREM1.3
endogenousREM WT * WT * WT *
CE μ Soluble
A
C
hydrophobic core
charged surface
N
C
90°
-36
-70
BGE
LD
L
L
K
A
E
E
A
A
K
Y
R
A
T
GT
APP
KK
I
L
G
I
N
C
Hydrophobicity: -4.5 +4.5
chargedface
chargedface
hydrophobicface
hydrophobicface
H = 0.104μH = 0.153
H = 0.682μH = 0.661
F
Rem
CARe
mCA
*
L195SG196V
A190S
G188V
Y184VA186SN
C
Figure 3. Amphipathy of RemCA alpha-helices plays a critical role in anchoring StREM1.3 Remorin to the plasma membrane. (A) A helical wheel projection of RemCA as a hairpin of α-helices showing amphipathy of each helix. Overall helix hydrophobicity (H) and hydrophobic moment (μH) are indicated in the middle of the helices, with an arrow showing the direction and intensity of the hydrophobic moments. (B) Ribbon and surface models of the RemCA and mutated RemCA* peptides with hydrophobicity (Kyte-Doolitle scale) shown by a color-code. The surface model of RemCA shows the predicted hydrophobic core in red, and charged surfaces in blue. Substitutions introduced in the RemCA* mutant aimed at reducing the amphipathy of the α-helices by altering the distribution of hydrophobic residues (Y184V, A190S, and L195S) and positively charged residues (A186S, G188V and G196V). (C) Maximum projec-tion of confocal scans through N. benthamiana epidermal cells expressing YFP:StREM1.3* mutant, with plastids shown in red. Scale bar indicates 20 μm. (D) Cell fractionation analysis of cells expressing GFP tagged StREM1.3 with native (+) and mutated (*) RemCA sequence, confirming that mutations introduced in StREM1.3* abolish PM localization.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

Figure 4. RemCA confers intrinsic lipid binding to StREM1.3 Remorin. (A) A�nity puri�ed 6His:StREM1.3 (WT) binds to protein-free liposomes in vitro in a RemCA-dependent manner. Liposome binding is abolished in 6His:StREM1.3 recombinant protein lacking RemCA (∆CA) or harbouring a mutated RemCA (*). The soluble (Sol.) and liposome-bound (Bound) fractions recovered after centrifugation are shown. Lipid mixing assays (B) and liposome leakage assays (C) show that RemCA ultrapure peptides insert into liposome membranes containing negatively charged lipids, as indicated by an increase in �uorescence de-quenching. By contrast, liposome leakage assays (C) showed that RemCA is not able to insert into neutral liposome membranes. SIV, Simian in�uenza virus fusion peptide; TFE, tri�uoroethanol; TX100, Triton X-100. Solvent used in (C) was dimethyl sulfoxide for negatively charged liposomes and TFE for neutral liposomes. Error bars show standard deviation of 6-8 independent measurements.
A
-40
α-Re
m
B
TX100
100
10
1
100
75
0 SIV
Rem
CA
TFE
50
25
Relat
ive �u
ores
cenc
ede
-que
nchin
g
C
SIV Solvent RemCA
Relat
ive �u
ores
cenc
ede
-que
nchin
g
negatively chargedliposomes
neutral liposomes
WT*∆CAInput
BoundSol. BoundSol. BoundSol.
-37
∆CA * WT
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

Figure 5. RemCA targets soluble proteins to the PM in a lipid-speci�c manner. (A) Lipid overlay assays performed with 6His tagged StREM1.3 and StREM1.31-170 show a di�erential a�nity of StREM1.3 for various classes of lipids, dependent on RemCA. The right panel shows Western Blot of the protein input material illustrating equal loading for StREM1.3 and StREM1.31-170. PA, phosphatidic acid; PIP3, phosphoinositol 3,4,5, triphosphate; PIP2, Phosphoinositol bisphosphate, the position of phosphate groups is indicated below; S1P, sphingosine 1-phosphate. (B) 6His:StREM1.3 shows higher a�nity for liposomes with a composition mimicking detergent insoluble membranes (DIM) compared to plasma membrane (PM - described in materials and methods). Histograms show quanti�cations and standard error based on Western blot signals for three independent experiments. Di�erential a�nity is signi�cant with p<0.05 in a Student t-test. (C) RemCA-tagged GFP transiently expressed in tobacco leaves segregates into the DIM fractions of the PM, similar to the endogenous Remorin. Total proteins were quanti�ed by bicinchoninic acid assay (BCA) relative to total proteins in puri�ed PM (error bars show standard deviation from three independent replicates). µ, microsomal fraction; DSM, detergent soluble membranes.
BLiposomes + 6his:StREM1.3Input
PM-likeliposomes
DIM-likeliposomes
µ PM DSM DIM
α-GFP
α-REM
30
20
10
0
% o
f inp
ut b
ound
PM-like DIM-like
endogenousREM
RemCA:GFP
C
-40
-36
-30
Blank PS PA PIP3
PIP2
S1P3,54,5 3,4
StREM1.3
StREM1.31-170
A
Input
StREM1.3
StREM1.31-170
-40-37
**
0
50
100% of totalproteins in
PM
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
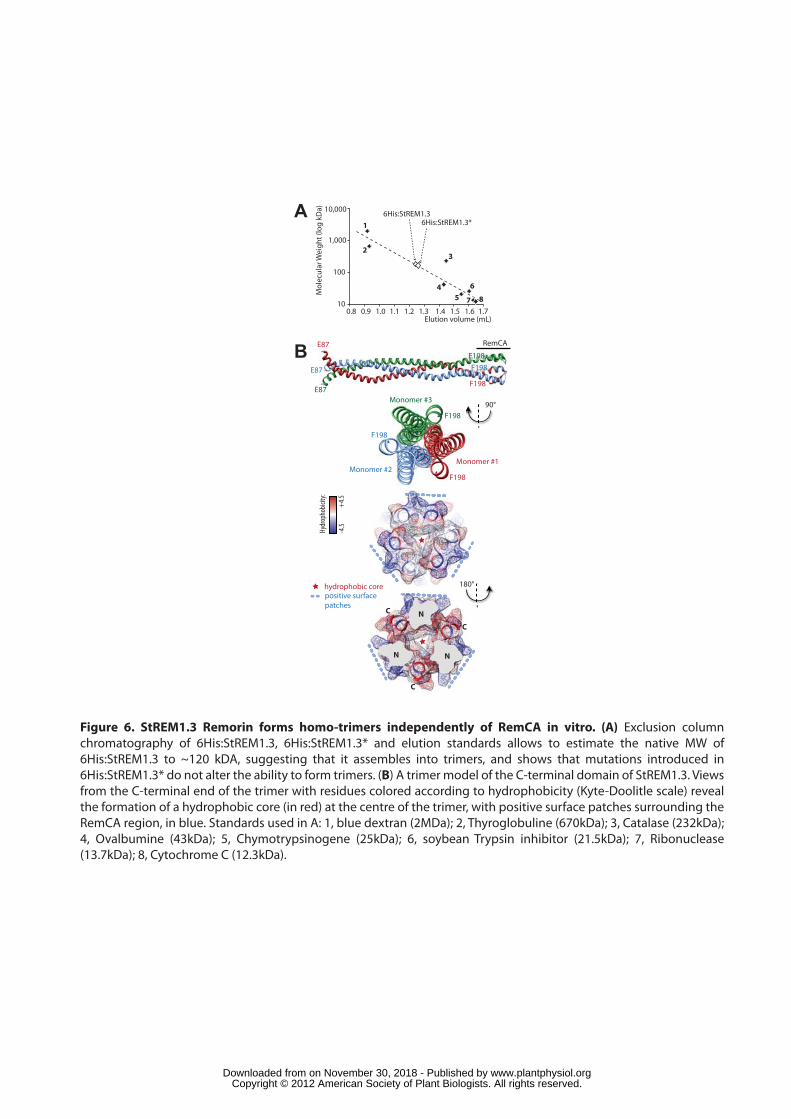
Figure 6. StREM1.3 Remorin forms homo-trimers independently of RemCA in vitro. (A) Exclusion column chromatography of 6His:StREM1.3, 6His:StREM1.3* and elution standards allows to estimate the native MW of 6His:StREM1.3 to ~120 kDA, suggesting that it assembles into trimers, and shows that mutations introduced in 6His:StREM1.3* do not alter the ability to form trimers. (B) A trimer model of the C-terminal domain of StREM1.3. Views from the C-terminal end of the trimer with residues colored according to hydrophobicity (Kyte-Doolitle scale) reveal the formation of a hydrophobic core (in red) at the centre of the trimer, with positive surface patches surrounding the RemCA region, in blue. Standards used in A: 1, blue dextran (2MDa); 2, Thyroglobuline (670kDa); 3, Catalase (232kDa); 4, Ovalbumine (43kDa); 5, Chymotrypsinogene (25kDa); 6, soybean Trypsin inhibitor (21.5kDa); 7, Ribonuclease (13.7kDa); 8, Cytochrome C (12.3kDa).
N
N N
C
C
C
B E87
E87
E87F198
RemCA
90°F198
F198
F198
Monomer #3
Monomer #1Monomer #2
180°
Hydrophobicity:
-4.5
+4.5
hydrophobic corepositive surfacepatches
A
F198F198
0.8 0.9 1.010
100
1,000
10,000
Elution volume (mL)M
olec
ular
Wei
ght (
log
kDa)
1.1 1.2 1.3 1.4 1.5 1.6 1.7
1
23
45
6
7 8
6His:StREM1.36His:StREM1.3*
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
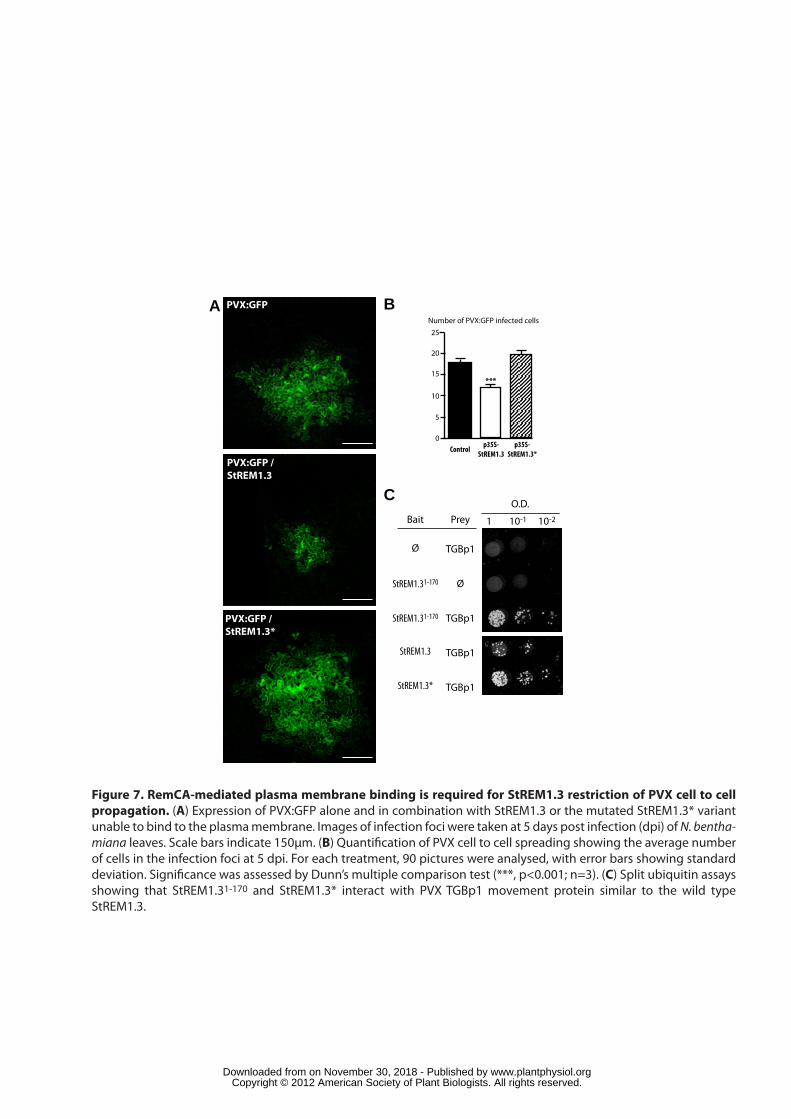
Number of PVX:GFP infected cells
0
5
10
15
20
25
***
Control p35S-StREM1.3
p35S-StREM1.3*
B
Figure 7. RemCA-mediated plasma membrane binding is required for StREM1.3 restriction of PVX cell to cell propagation. (A) Expression of PVX:GFP alone and in combination with StREM1.3 or the mutated StREM1.3* variant unable to bind to the plasma membrane. Images of infection foci were taken at 5 days post infection (dpi) of N. bentha-miana leaves. Scale bars indicate 150µm. (B) Quanti�cation of PVX cell to cell spreading showing the average number of cells in the infection foci at 5 dpi. For each treatment, 90 pictures were analysed, with error bars showing standard deviation. Signi�cance was assessed by Dunn’s multiple comparison test (***, p<0.001; n=3). (C) Split ubiquitin assays showing that StREM1.31-170 and StREM1.3* interact with PVX TGBp1 movement protein similar to the wild type StREM1.3.
StREM1.3
StREM1.3*
O.D.
1 10-1 10-2
Ø TGBp1
ØStREM1.31-170
StREM1.31-170 TGBp1
Bait Prey
TGBp1
TGBp1
A PVX:GFP
PVX:GFP /StREM1.3
PVX:GFP /StREM1.3*
C
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
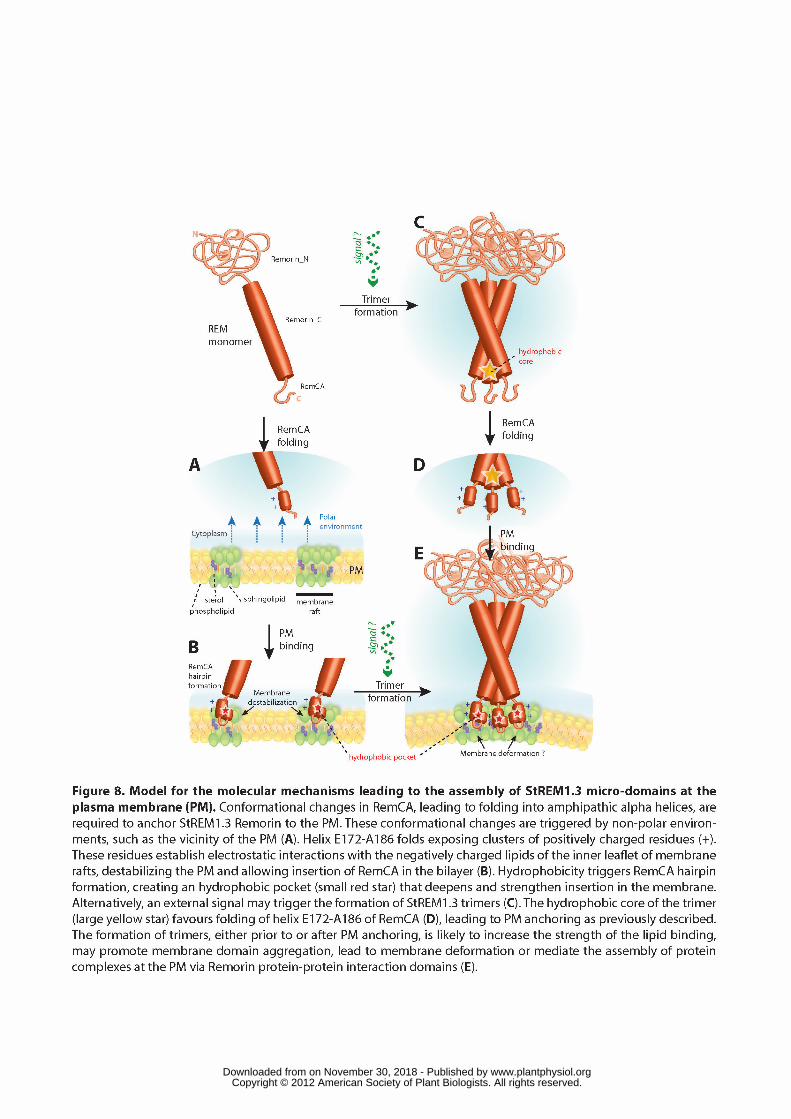
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
Recommended

















