Embed Size (px)
Citation preview

RESEARCH ARTICLE
Zombie ant death grip due to hypercontractedmandibular musclesColleen A. Mangold1,2,*, Melissa J. Ishler2,3, Raquel G. Loreto2,3,4, Missy L. Hazen5 and David P. Hughes2,3,6
ABSTRACTThere are numerous examples of parasites that manipulate thebehavior of the hosts that they infect. One such host–pathogenrelationship occurs between the ‘zombie-ant fungus’Ophiocordycepsunilateralis sensu lato and its carpenter ant host. Infected ants climbto elevated locations and bite onto vegetation where they remainpermanently affixed well after death. Themandibular muscles, but notthe brain, of infected ants are extensively colonized by the fungus.Wesought to investigate the mechanisms by which O. unilateralis s.l.may be able to influence mandibular muscle contraction despitewidespread muscle damage. We found that infected muscles showevidence of hypercontraction. Despite the extensive colonization,both motor neurons and neuromuscular junctions appear to bemaintained. Infection results in sarcolemmal damage, but this is notspecific to the death grip. We found evidence of precise penetration ofmuscles by fungal structures and the presence of extracellularvesicle-like particles, both of which may contribute to mandibularhypercontraction.
KEY WORDS: Ophiocordyceps, Host–pathogen interactions,Fungus, Behavioral manipulation, Motor neuron, Extracellularvesicles
INTRODUCTIONParasites are capable of expressing their phenotypes in the bodiesof the hosts that they occupy. This is an example of an extendedphenotype (Dawkins, 1982). Specific viruseswithin the Baculoviridaefamily including Lymantria dispar nucleopolyhedrovirus (Hooveret al., 2011) andMamestra brassicae nucleopolyhedrovirus (Goulson,1997) infect L. dispar and M. brassicae larvae, respectively. Thistriggers distinct changes in behavior where infected larvae climb upto higher elevations on vegetation, die and liquify. This process ishypothesized to positively impact the transmission of infectiousprogeny virions to other hosts (Goulson, 1997; Heil, 2016). Precisemanipulation of host behavior is achieved via the coordinatedexpression of specific viral genes including ecdysteroid UDP-glucosyltransferase, protein tyrosine phosphatase, chitinase andcathepsin (Han et al., 2015; Hawtin et al., 1997; Hoover et al.,2011; Kamita et al., 2005; Katsuma et al., 2012). Another example is
the lancet liver fluke Dicrocoelium dendriticum, which infects anumber of different grazing animals, but requires transmission frommollusks to intermediate ant hosts for maturation prior totransmission to their ruminant definitive hosts (Manga-Gonzálezet al., 2001). Once infected, ants display a change in behavior wherethey climb to the tops of blades of grass and bite down during theearly dawn and evening hours (Botnevik et al., 2016; Manga-González et al., 2001). It has been suggested that D. dendriticumexpresses this extended phenotype via a direct influence on the brainsof their ant hosts (Botnevik et al., 2016; Carney, 1969; Krull andMapes, 1952; Martín-Vega et al., 2018; Moore, 1995).
One of the most well-studied microbial manipulators in insects isthe entomopathogenic fungus Ophiocordyceps unilateralis sensulato. Infection withO. unilateralis s.l. triggers a pronounced changein behavior where infected ants climb up vegetation and bite ontoleaf veins or twigs, where they ultimately die (Andersen et al., 2009;Hughes et al., 2011; Loreto et al., 2018). The fungus grows withinthe ant’s body, consuming all of its resources, extends a stalkfrom the ant’s head and then sporulates, dispersing infectious sporesfrom the stalk onto uninfected ants foraging along the forest floorbelow. The fungus is unable to grow or spread within the nest,making the manifestation of these behavioral changes in areasaround individual colonies essential for efficient transmission tonew hosts (Loreto et al., 2014). The ‘zombie-ant’ phenotype iscomplex and is composed of two distinct behaviors, (1) climbing upvegetation and (2) the ‘death grip’, where the ant bites onto a leafvein or twig prior to death. Previous studies examining infected hostmandibular muscle morphology show extensive muscle cell atrophyand colonization of the head capsule by fungi at the moment ofmanipulation (Fredericksen et al., 2017; Hughes et al., 2011).Moreover, within the host, cells of Ophiocordyceps kimflemingiae(=unilateralis; Araújo et al., 2018) act cooperatively and form aninterconnected community around individual mandibular musclecells in the infected ant. This causes the muscle fibers to becomewidely separated as the invading organism occupies up to roughly40% of the biomass of the ant host (Fredericksen et al., 2017).
Vertebrate muscle cells propagate signals from neuromuscularjunctions (NMJs) across the length of the muscle, resulting in fullmuscle contraction. In contrast, invertebrate muscle cells can beinnervated bymultiple motor neurons at a number of different pointsalong the length of the muscle cell (Edwards, 1959; Usherwood,1967). Contraction of ant mandibular closer muscle is controlled by10–12 motor neurons (Paul and Gronenberg, 2002). In thecockroach abdominal muscle, NMJs are spaced 4–30 µm apart,with each muscle fiber possessing 1–4 junctions (Edwards, 1959).Although not definitively quantified in ant mandibles, this suggeststhat ant mandibular muscle should have numerous contacts withmotor neurons.
Prior observations have demonstrated extensive atrophy andmuscle invasion at the time of the death grip behaviorin O. kimflemingiae-infected ants (Fredericksen et al., 2017;Hughes et al., 2011). These observations combined with ourunderstanding of the architecture of neuromuscular systems inReceived 30 January 2019; Accepted 3 June 2019
1Department of Biochemistry andMolecular Biology, Pennsylvania State University,University Park, PA 16802, USA. 2Center for Infectious Disease Dynamics,Huck Institutes of the Life Sciences, Pennsylvania State University, University Park,PA 16802, USA. 3Department of Entomology, College of Agricultural Sciences,Pennsylvania State University, University Park, PA 16802, USA. 4Unit of Geneticsand Genomics of Insect Vectors, Department of Parasites and Insect Vectors,Institute Pasteur, Paris 75015, France. 5Huck Institutes of the Life SciencesMicroscopy and Cytometry Facility, Pennsylvania State University, University Park,PA 16802, USA. 6Department of Biology, Eberly College of Science, PennsylvaniaState University, University Park, PA 16802, USA.
*Author for correspondence ([email protected])
C.A.M., 0000-0001-7205-4540
1
© 2019. Published by The Company of Biologists Ltd | Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

insects raise different questions. For example, how can suchprecise manipulation of host behavior occur when the muscleis extensively atrophied? And how can motor neurons andNMJs be maintained when the muscles are so heavily invaded bythe fungal parasite? Answering these questions may provideimportant insight concerning the mechanisms utilized byO. unilateralis s.l. to trigger the death grip. Given the dense,interconnected networks formed by O. kimflemingiae aroundmuscle cells (Fredericksen et al., 2017) and the previouslydocumented muscle cell atrophy (Hughes et al., 2011), wehypothesized that motor neurons innervating the mandibularmuscle may become degraded as the fungal cells begin to crowdthe intermuscular space. The fungus could then act locally to triggermuscle contraction.To investigate overall muscle morphology and test for the
presence and integrity of motor neurons, we analyzed mandibularmuscle fromO. kimflemingiae-infected ants using scanning electronmicroscopy (SEM) to visualize gross muscle, motor neuronand fungus structure. We then utilized transmission electronmicroscopy (TEM) to assess the effects of infection on musclecytoarchitecture and test for the presence of nervous tissue.As controls, we used healthy ants and ants infected with thefungus Beauvaria bassiana, a generalist and non-manipulativeentomopathogenic fungus within the same order (Hypocreales) asO. kimflemingiae. Our data indicate that at the moment of biting,mandibular muscles of zombie ants demonstrate hypercontraction.Despite the abundance of fungus surrounding muscle cells,motor neuron innervation was maintained and NMJs werepresent. Sarcolemma breakdown was evident; however, this alsooccurred in the mandibular muscle of B. bassiana-infected ants.Further investigation identified potential alternative mechanismsunderlying muscle hypercontraction at the time of biting. Theseinclude the formation of interconnected fungal cell networks, directinsertion of hyphal tubes into muscle and the secretion ofextracellular vesicle-like particles. Together, the data presentedhere suggest that the zombie ant death grip phenotype may be aresult of O. kimflemingiae-specific secretion or direct introductionof modulators that mediate muscle contraction. Compounded withextensive damage to the sarcolemma, this may cause musclehypercontraction. These results provide an important step towardsunderstanding the mechanisms underlying the manifestation of thezombie ant death grip.
MATERIALS AND METHODSMaintenance of ant coloniesCamponotus castaneus (Latreille 1802) ants were collected inDonalds, SC, USA, and housed in plastic containers (Sistema,Mangere Auckland, New Zealand) within a temperature-controlledroom (25.8–23.2°C). The insectary was maintained on a 12 h:12 hlight:dark cycle with a relative humidity of roughly 79–99%. Waterand 10% sugar water were available ad libitum and replenishedevery 1–2 weeks. Biting platforms made of Plaster of Paris (DAP,Baltimore, MD, USA) and embedded with autoclaved toothpicks(Diamond, Jarden Home Brands, Fishers, IN, USA) were placedinside each cage to provide a substrate for climbing and bitingfollowing behavioral manipulation. Nests were made using a Plasterof Paris base with a red transcolor PVC film (TAP Plastics Inc.,Sacramento, CA, USA) over an empty pipette tip box lid (200 µl,VWR Zap, Radnor, PA, USA). Ants were randomly assigned to atreatment group (infected or control) and were individually paintedwith a paint marker (Edding 751, Ahrensburg, Germany) todistinguish group assignment.
Infections and behavioral characterizationOphiocordyceps kimflemingiae infectionManipulated ants were collected from two separate infections.For the first infection, O. kimflemingiae was harvested from thegaster (part of the thorax) of a recently deceased C. castaneus ant.Fungal tissue was incubated on a potato dextrose agar mediaplate for 10 months until growth reached roughly 1.5 cm indiameter. Inoculum was prepared as previously described(de Bekker et al., 2014). Roughly 0.375 cm2 of fungal tissuewas excised, homogenized in 500 µl Grace’s insect medium(Sigma-Aldrich, St Louis, MO, USA) and diluted 4-fold. Antswere injected with 1 µl of inoculum into the membrane locatedunderneath the forelegs using a laser-pulled 10 µl micropipette(Drummond Scientific Co., Broomall, PA, USA) and aspirator tube(Drummond Scientific Co.).
The second infection was executed as follows. Deceased antsbiting onto twigs were collected in Donalds, SC, USA. Followingsurface sterilization with 70% ethanol, the cuticle was removed andall internal tissue containing O. kimflemingiae was placed into asterile conical 1.5 ml tube (VWR) with two sterile 5/32 inch metalballs (Wheels Manufacturing Inc., Louisville, CO, USA) and 100 µlGrace’s Insect Media (Thermo Fisher Scientific, Hampton, NH,USA) supplemented with 10% fetal bovine serum (LifeTechnologies, Carlsbad, CA, USA). Fungal material was thenlysed for 1 min at 30 Hz using a Tissue Lyser II (Qiagen,Germantown, MD, USA), and briefly centrifuged using a tabletopmicrocentrifuge (Benchmark Scientific, Sayreville, NJ, USA) toremove cellular debris. The resulting supernatant was used as theinoculum; 1 μl of inoculum was then injected into C. castaneus antsas described above. Control ants (n=6) were injected with 1 µlGrace’s Insect Media supplemented with 10% fetal bovine serum.Infection was monitored for roughly 30 days with behavioralmanipulation occurring most often between days 15 and 30. Antswere identified as behaviorally manipulated if they were biting ontoa substrate, exhibited muscle spasms, did not respond to externalstimuli and did not detach from the substrate when touched (TEM,n=3; SEM, n=6). At the time of biting when the ants were still alive,they were immediately flash frozen or dissected for histology.
Beauvaria bassiana infectionPositive control ants were infected with B. bassiana, anentomopathogenic fungus that does not induce the death gripbehavior. Beauvaria bassiana conidia were provided by Dr NinaJenkins at Penn State University. Infectious conidia were suspendedin 0.05% Tween/phosphate-buffered saline, poured onto filterpaper, and placed in a 60 mm Petri dish. Camponotus castaneusants were then placed in the Petri dish and allowed to walk overinfectious spores for 24 h. Infected ants were then placed in a newcage. For B. bassiana-infected ants, there are no stereotypicalbehavioral phenotypes like those observed in O. kimflemingiae-infected ants that happen prior to host death, which occurs between3 and 5 days post-infection. Infected ants may exhibit sicknessbehaviors well before death, making it difficult to correctly identifyants that will die within the next few hours. As such, between days 3and 5 post-infection, infected ants were periodically monitored in1 h intervals to ensure collection of ants within 1 h of death. Antswere then flash frozen in liquid nitrogen until processing forhistology (control, n=3; infected, n=3).
Dissections and fixation for electron microscopyFor samples analyzed by SEM (n=3–6 per group), mandibularmuscle still attached to cuticle was rapidly dissected and placed in
2
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

2.5% glutaraldehyde in 0.1 mol l−1 sodium phosphate bufferovernight at 4°C. Tissue was then washed 3 times with 0.1 mol l−1
sodium phosphate buffer for 5 min each, dehydrated with 25%, 50%,70%, 85% and 95% ethanol once each for 5 min, and then 100%ethanol 3 times. Dehydrated samples were then dried by critical pointdrying (Leica EM CPD300, Wetzlar, Germany). Images wereacquired using a Zeiss Sigma VP-FESEM (Thornwood, NY, USA).To assess average extracellular vesicle diameter, 100 particles pergroup were measured using Fiji (ImageJ).For samples analyzed by TEM (n=3 per group), mandibular
muscle was immediately removed, placed in modified Karnovsky’sfixative (2.5% paraformaldehyde, 1.5% glutaraldehyde, 0.1 mol l−1
sodium cacodylate buffer) and incubated overnight at 4°C. Tissuewas then washed 3 times in 0.1 mol l−1 sodium cacodylate buffer for5 min and fixed in 2% osmium tetroxide for 1 h protected fromlight. Samples were washed twice in 0.1 mol l−1 sodium cacodylatebuffer, once in Milli-Q water, and then stained for 1 h in 2%aqueous uranyl acetate protected from light. Samples weredehydrated in ethanol (50%, 70%, 85%, 90%, 95% and 100%once each for 5 min, and 3 times in 100% EM grade ethanol),washed 3 times in acetone, and then infiltrated with Spurrs asfollows: 50% acetone/50% Spurrs overnight, 25% acetone/75%Spurrs ∼8 h, 100% Spurrs overnight, 100% Spurrs ∼8 h, 100%Spurrs overnight, and then polymerized in fresh Spurrs overnight at60°C. Ultrathin sections (70 nm, Leica EM UC6 Ultramicrotome)were placed on grids, post-stained in 2% uranyl acetate/50%ethanol, washed in distilled water, and then stained in lead citrate.Grids were washed, allowed to dry, and then imaged on a JEOLJEM 1200 EXII (Peabody, MA, USA).
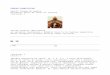
RESULTSInfected mandibular muscle demonstrates diffuse presenceof fungal cells and evidence of over-contractionPrevious research has shown the dense network of fungal cellsinfiltrating mandibular muscle in infected ants at the time of biting(Fredericksen et al., 2017). In support of these previous data, SEManalysis of uninfected (Fig. 1A) and infected (Fig. 1B,C)mandibular muscle demonstrated the diffuse presence of fungalcells on and around individual muscle fibers of manipulatedants. This phenotype was also seen in B. bassiana-infected ants,where fungus was observed throughout the mandibular musclespace (Fig. S1). Numerous muscle cells in O. kimflemingiae-infected ants demonstrated a unique morphology where z-linesappeared to be swollen and the sarcomeres shortened, giving agrooved-like appearance (Fig. 1B,C). Additionally, extensivedamage to the sarcolemma was evident (Fig. 1C, arrowheads).Closer inspection of these regions of sarcolemma breakdownrevealed what seem to be individual myofibrils beneath themembrane (Fig. 1D, arrow). Areas presumed to be the z-lines(Fig. 1D, arrowheads) demonstrated significant damage, whereinthe fibers appeared frayed and broken.
Given the distinct change in muscle cell appearance at the time ofthe death grip, we wanted to investigate the structural effectsof O. kimflemingiae infection on muscle cells, using TEM tovisualize intracellular muscle fiber ultrastructure. As muscle fromB. bassiana-infected ants more closely resembled control muscle,where z-line shortening was not as prevalent nor as severe, wedid not pursue further analysis of these samples by TEM.In O. kimflemingiae-infected muscle, we were able to observe
BA
C D
M
M
M
F
F
F
ZMN
MM
M
T
T
Fig. 1. Invasion of mandibular muscle space by Ophiocordyceps kimflemingiae – evidence for muscle over-contraction. Infected mandibularmuscle displays unique characteristics as visualized by scanning electronmicroscopy (SEM) (controls, n=3; infected, n=6). (A) In uninfected controls, mandibularmuscle cells demonstrate an even morphology with regularly distributed striations (z-lines). Motor neurons and tracheoles are abundant. Scale bar: 20 µm.M, muscle; MN, motor neuron; T, tracheole; Z, z-line. (B) Fungal cells completely invade the inter-muscle space and are in close contact with individual musclecells. Infected muscle cells demonstrate a unique morphology wherein the z-lines appear to be swollen and the sarcomeres shortened, giving the regularstriations a very pronounced appearance. Scale bar: 20 µm. M, muscle; F, fungus. (C) As a result of extensive damage to the sarcolemma, individual myofibrilsunderneath the membrane are exposed in infected muscle. Areas of sarcomere shortening (arrows: swollen, unexposed regions; arrowheads: exposedregions) are evident. Scale bar: 20 µm. (D) Magnification of the boxed region in C. Presumed z-lines are indicated by arrowheads while individual myofibrils areindicated by arrows. The z-lines display a frayed, damaged appearance. Scale bar: 1 µm.
3
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

distinct changes in z-line morphology at the time of biting. Thez-lines in control ants exhibited a connected, regular, linearmorphology (Fig. 2A). In contrast, infected muscle demonstratedchanges in z-line morphology where they were swollen (Fig. 2B),irregular and/or disengaged from neighboring z-lines (Fig. 2C),suggesting sarcomere disruption and shortening. Additionally, thepresence of numerous myelin-like structures was evident withininfected muscle (Fig. 2B, arrows).
Motor neurons and NMJs remain intact at the time of bitingWe next sought to examine the presence and integrity of motorneurons and NMJs within the mandibular muscle. It was more
difficult to locate motor neurons and NMJs in infected tissue thanin control tissue (Fig. 3A), but we were able to identify thesestructures and see that they remained intact (Fig. 3B). This was alsotrue in B. bassiana-infected muscle (Fig. S2). In some cases,O. kimflemingiae cell clusters were observed in direct contact withmotor neurons and near NMJs (Fig. 3B). Unfortunately, surfaceimaging did not allow for determination of the presence of motorneurons within fungal cell clusters. We were therefore unable toquantify exact differences in motor neurons between infectedmandibular muscle and healthy controls. Nonetheless, in bothcontrol and infected ants (Fig. 3C,D, respectively), nervous tissuewas present around and in contact with muscle cells.
BA C
N
M
M
S
S
M
M
M
S
S
M
Fig. 2. Infected mandibular muscle demonstrates evidence of damage to z-lines. Transmission electron microscopy (TEM) analysis of control (n=3) andO. kimflemingiae-infectedmuscle (n=3). (A) Control ants demonstrate the presence of healthy, regularly spaced sarcomeres (S) as evidenced by long, connectedparallel z-lines (arrowheads) through the muscle cell. Large mitochondria (M) are present throughout each muscle cell. Scale bar: 5 µm. (B,C) Infectedmuscle demonstrates extensive sarcomere disruption including z-line damage, either in a hypercontracted, swollen state (B, arrowheads) or with a broken,non-uniform appearance (C, arrowheads). The presence of multiple double-membraned bodies (B, arrows) is also evident. Scale bars: B, 5 µm; C, 1 µm.
A
C D
B
M
M
Fig. 3. Motor neurons and neuromuscularjunctions are maintained in infectedmandibular muscle at the time of biting. Incontrol ants (A), numerous motor neurons(arrows) and neuromuscular junctions (NMJs,arrowheads) are evident along the length ofindividual muscle cells (SEM image). Scalebar: 2 µm. (B) Motor neurons (arrow) and NMJs(arrowhead) in infected mandibular muscle aremaintained (SEM image). In some instances,these structures are in close contact with fungalcell bodies. Scale bar: 2 µm. In bothcontrol (C) and infected (D) ants, nervoustissue is present near and/or in contact withmuscle, as identified by TEM (nervous tissue,arrows; muscle cells, arrowheads).M, mitochondria. Scale bars: C, 500 nm; D,1 µm. Controls, n=3; infected, n=6.
4
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

Muscle cells exhibit diffuse degradation of sarcolemmaassociated with fungal infectionOne unanticipated finding was that in theO. kimflemingiae-infectedmandibular muscle samples examined, we observed grossdegradation of muscle fiber sarcolemma (Figs 1C,D and 4A).In most cases, these sections of sarcolemma breakdown werelocated directly beside clusters of fungal cells. Similar toO. kimflemingiae infection, B. bassiana infection also triggeredsarcolemma degradation (Fig. 4B). Some small patches ofsarcolemma destruction were observed in control tissue. However,these sections were most often localized near excision points of thedissection and are more than likely an artifact of the dissection itself.Degraded sarcolemma was much more prevalent in infected muscleand was localized to internal muscle fibers as well, suggesting theseobservations are not artifacts.
Ophiocordyceps kimflemingiae forms extensiveconnections with neighboring fungal cellsUsing SEM, we were also able to observe the previouslydocumented collective behavior of fungal cells: anastomosis tubesconnecting individuals, resulting in the formation of fungalnetworks around host muscle cells (Fredericksen et al., 2017)(Fig. 5, arrows). Individual fungal cells can form multipleconnections along the length of the entire cell body (Fig. 5A).Additionally, we were able to observe the formation of theseconnection points (Fig. 5A, arrowheads).Consistent with previous literature (Fredericksen et al., 2017), we
were able to observe numerous hyphal bodies in direct contact withant muscle (Fig. S3), as well as hyphal body invasion of musclecells (Fig. 6). In these cases, invading hyphal bodies were connectedto others via extensive anastomosis bridge networks. Additionally,we identified the presence of numerous extracellular vesicle-likeparticles associated with fungal cells within the mandibular musclespace (Fig. 7, arrows). Extracellular vesicle-like particles were alsoobserved in B. bassiana-infected muscle (Fig. S4). Extracellularvesicle-like particles identified within O. kimflemingiae-infected
muscle were small (averaging 463 nm in diameter), spherical,similar in size and often found in small (Fig. 7B) or large (Fig. 7D)clusters. In contrast, vesicles found within B. bassiana-infectedtissue appeared larger (averaging 1.6 µm in diameter), less sphericaland were randomly dispersed (Fig. S4).
DISCUSSIONManipulation of host behavior by O. unilateralis s.l. is a complexphenomenon that is manifested by the stereotypical ‘deathgrip’ where infected ants bite into vegetation prior to death(Andersen et al., 2009; Hughes et al., 2011). Studying how thisphenotype arises is of significant interest and will yield importantinsight into pathogen–host co-evolution as well as potentialmechanisms underlying fungal pathogenesis. In this study, wesought to understand the interactions between host muscle tissueand fungal cells.
Prior studies examining the interactions between entomopathogenicfungi and their hosts have demonstrated that in general, despite thewidespread distribution of fungal cells around host muscle,destruction and physical invasion are delayed until after death (Elyaet al., 2018; Funk et al., 1993; Gryganskyi et al., 2017). Thispotentially allows the infected hosts to continue moving around andconsuming nutrients until the time of death (Gryganskyi et al., 2017).InOphiocordyceps-infected ants, our lab has shown the developmentof an extensive, interconnected, fungal community around individualmuscle fibers in host mandibles, suggesting that fungal cells maywork cooperatively to consume host nutrients and induce the bitingbehavior seen in infected ants (Fredericksen et al., 2017). In thepresent study, we built on these previous findings to provide a moreglobal view of fungal–host interactions in this model system (seeTable 1 for a list of similarities and differences between the twostudies and fungal pathogens used). We confirmed that hostmandibular muscle is heavily populated by O. kimflemingiae cells(Fig. 1B). In addition, we observed numerous structural abnormalitiesin infected host cells, the most striking of which was z-line swellingand sarcomere shortening (Fig. 1B–D).
A B Fig. 4. Damage to the sarcolemma is aconsequence of fungal infection and notspecifically associated with the death grip.Damage to muscle cell sarcolemma is evidentin both O. kimflemingiae- (A) and Beauvariabassiana- (B) infected muscle (arrows) (SEMimages). Scale bars: A, 10 µm; B, 20 µm.Ophiocordyceps kimflemingiae-infected ants:controls, n=3; infected, n=6; B. bassiana-infected ants: controls, n=3; infected, n=3.
A B Fig. 5. Ophiocordyceps kimflemingiaefungal cells demonstrate collectivebehavior via anastomosis tube formation.(A,B) SEM reveals that, within infected muscle,individual O. kimflemingiae cells formextensive networks with each other, connectedvia the formation of anastomosis tubes(arrows). In many instances, numerousanastomosis tubes form along the length ofone fungal cell, and formation of new tubes(A, arrowhead) is evident. Scale bars: 10 µm.Controls, n=3; infected, n=6.
5
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

In order to investigate this phenotype further, we utilized TEM tocharacterize changes to muscle cell ultrastructure at the time ofbiting in O. kimflemingiae-infected ants. Areas of z-line swelling,shortening (Fig. 2B) and disruption (Fig. 2C) were evident ininfected muscle. Swelling and disruption of the z-line as well assarcolemmal corrugation have been observed in human skeletalmuscle following eccentric exercise, where the muscleconcomitantly lengthens as it contracts (Lauritzen et al., 2009),and is indicative of muscle hypercontraction. Similar phenotypesare also observed in insect striated muscle following intensivecontraction. In adult female locusts, Locusta migratoria,sarcolemma corrugation and z-line disruption are evident insupercontracted ovipositor muscle in preoviposition females(Jorgensen and Rice, 1983). Additionally, corrugation of thesarcolemma and the formation of myelin-like bodies (Fig. 2B) areabundant in O. kimflemingiae-infected muscle. The appearance ofmyelin-like structures occurs in rat heart muscle followingadministration of adrenalin (Kleimenova and Belen’kii, 1975),
suggesting that these bodies form in response to hypercontraction.Together, the data presented in the present study detailing z-lineswelling, sarcomere shortening and disruption, sarcolemmablebbing and the formation of myelin-like bodies in infectedmuscle (Fig. 2) support the hypothesis that mandibular muscle at thetime of biting is in a state of hypercontraction. This phenomenonhad not been previously reported in other studies examiningentomopathogenic fungus–insect interactions.
The motor neurons and NMJs of manipulated ants appear to bepresent and structurally intact (Fig. 3). This is true despite extensivecolonization of fungal cells around individual muscle fibers and thewide separation of muscle cells evident at the time of biting.Although we observed fewer motor neurons and NMJs, this may bedue to the large number of fungal cells present in the mandibularmuscle space. Alternatively, motor neurons may degenerate overthe course of infection, yielding a smaller number at the time ofbiting. The host nervous system is suggested to be important forbehavioral manipulation (Adamo, 2013). In fact, the central nervous
A B
C
Fig. 6. Ophiocordyceps kimflemingiaeinserts hyphal projections into hostmuscle. In some instances, physical invasionof muscle cells by fungal hyphae is evident(A; boxed regions are shown at highermagnification in B and C; SEM images).Interconnected networks of fungal cells(A, arrows) are able to access host muscle viadirect insertion of one fungal cell from thenetwork (B,C). Scale bars: A, 2 µm; B, 1 µm;C, 200 nm. Controls, n=3; infected, n=6.
B
D
A
C
Fig. 7. Ophiocordyceps kimflemingiaewithin the mandibles demonstrateassociation with extracellular vesicle-likeparticles. Numerous fungal cells possessextracellular vesicle-like particles (arrows)attached to their cell bodies (A,C; boxedregions are shown at higher magnification inB and D, respectively; SEM images). While inmost cases, these vesicles appear in smallclusters (B), in some instances, large massesare evident (D). Scale bars: A, 2 µm; B, 1 µm;C, 10 µm; D, 2 µm. Controls, n=3; infected,n=6.
6
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

system of the manipulated ants appears to be physically intact at themoment of the death grip (Fredericksen et al., 2017), and it isarguable that motor neurons are also preserved at this time.Therefore, our difficulty in finding motor neurons and NMJs may bea consequence of clusters of fungal cells that were physicallyobstructing our view (Fig. 1B).When assessing the morphology of infected mandibular muscle,
we observed overt damage to the sarcolemma localized throughoutthe mandibular muscle space in all samples infected withO. kimflemingiae (Fig. 4A). We hypothesized that perhaps thisdiffuse degradation of the sarcolemma may underlie aberrant musclecontraction observed in manipulated ants. However, we observedsimilar regions of sarcolemma degradation in themandibular musclesof B. bassiana-infected ants (Fig. 4B). Therefore, muscle membranedamage is not necessarily specific to infection with O. kimflemingiaeor the sole mechanism underlying initiation of the death grip.Nonetheless, sarcolemma degradation may still play a role ininitiating the death grip associated with manipulation.The sarcolemma is responsible for maintaining proper ionic
balance and resting membrane potential (Hopkins, 2006). Oncestimulated by excitatory motor neurons, the membrane potentialdepolarizes, releasing calcium stores from the sarcoplasmicreticulum, which triggers muscle contraction (Iwamoto, 2011;Josephson, 2009). Disruption of the sarcolemma can cause aberrantcalcium influx, resulting in alterations in muscle contraction andultimately muscle fiber degradation, as occurs in Duchenne musculardystrophy (Petrof, 2002). Degradation of the sarcolemma is alsothought to contribute to muscle hypercontraction in response tonotexin, a toxin in Australian tiger snake (Notechis scutatus scutatus)venom (Dixon and Harris, 1996). It is unlikely that the sarcolemmadamage directly causes the hypercontraction of mandibular musclesobserved during the death grip because it is also abundant on non-manipulated ants infected with B. bassiana. However, sarcolemmabreakdown could facilitate the direct introduction of fungal secretedfactors to trigger the observed hypercontraction.At the time of biting in O. kimflemingiae-infected ants, fungal
genes associated with ergot alkaloid biosynthesis are increasedin expression, suggesting these compounds may play a role inbehavioralmanipulation (deBekker et al., 2015).Claviceps purpurea,a fungus closely related to Ophiocordyceps, secretes ergotalkaloids, compounds known to trigger alterations in serotonergicneurotransmission, resulting in convulsions and hallucinations in
mammals (Eadie, 2003). Additionally, the entomopathogenic fungusMetarhizium anisopliae releases destruxins when infecting tobaccohornworm (Manduca sexta) larvae. These destruxins cause rapiddepolarization of host muscle membrane, potentially via modulationof calcium channels, thereby triggering sustained muscle contractionand paralysis, followedby flaccid paralysis (Samuels et al., 1988). TheO. kimflemingiae genome encodes 36 putative enterotoxins (deBekker et al., 2017). Importantly, transcription of one putativeenterotoxin ortholog is up-regulated ∼3000-fold at the moment ofbiting, with transcription decreasing ∼200-fold following the deathgrip (de Bekker et al., 2015, 2017). This gene is encoded in numerousOphiocordyceps species that are able to manipulate ant behavior (deBekker et al., 2017). These data suggest that enterotoxins may playimportant roles in the manifestation of the death grip. However, thesetoxins still need to be isolated from O. kimflemingiae and fullycharacterized. Further studyof these fungus-derived toxins is requiredto understand their potential involvement in disease pathogenesis andmanifestation of the death grip.
We observed tight association of fungal cells to host sarcolemma(Fig. S3), invasion of host muscle by fungal hyphae (Fig. 6), and thepresence of fungus-associated extracellular vesicle-like particles(Fig. 7). Physical invasion of host muscle by O. kimflemingiae mayprovide a mechanism by which the fungus can extract nutrients fromhost tissue and feed the network of fungal cells via anastomosis tubeconnections. This is supported by data demonstrating invasion ofFrankliniella occidentalis thorax muscle by Verticillium lecaniipost-mortem. Areas of tissue clearing are evident around hyphal tipswithin muscle tissue, indicating enzymatic degradation (Schreiteret al., 1994). A similar process may be utilized by O. kimflemingae,where invading hyphae may digest local muscle tissue, andnutrients can then be shuttled through the interconnected networkof fungal cells.
Further research is required to determine whether the identifiedextracellular vesicle-like particles are derived from host cells inresponse to infection, or are fungus derived. At least eight speciesof fungi have been shown to release extracellular vesicles,including the human pathogenic fungi Cryptococcus neoformans,Histoplasma capsulatum, Paracoccidiodes brasiliensis andCandida albicans (Albuquerque et al., 2008; Gehrmann et al.,2011; Oliveira et al., 2013; Rodrigues et al., 2007, 2011; Vallejoet al., 2011; Vargas et al., 2015; Zamith-Miranda et al., 2018). Therelease of extracellular vesicles may play important roles inmodulating the interactions between fungi and their hosts.Specifically, they may contribute to fungal pathogenesis andvirulence (Huang et al., 2012; Oliveira et al., 2010, 2013;Panepinto et al., 2009; Rodrigues et al., 2008, 2011; Zamith-Miranda et al., 2018). These findings combined with the datapresented here suggest that perhaps O. kimflemingiae secretesputative enterotoxins packaged within extracellular vesicles in asynchronous fashion that triggers local mandibular musclehypercontraction. We observed similar invasion by B. bassiana aswell as evidence of extracellular vesicle-like particles inB. bassiana-infected mandibular muscle (Fig. S4). Further testingneeds to be done to determine whether B. bassiana- andO. kimflemingiae-derived extracellular vesicles contain similar orunique cargo. These data would provide valuable informationdetailing potential mechanisms of distinct pathogenesis strategies.
Unraveling the mechanisms by which a microbe is able tomanipulate host behavior will aid in elucidating the fundamentalmechanisms underlying animal behavior as well as detailmechanisms of fungal pathogenesis and virulence. In the case ofzombie ants, many questions remain unanswered. In the present
Table 1. Phenotypes associated with fungal infection in Beauvariabassiana- and Ophiocordyceps kimflemingiae-infected Camponotuscastaneus ants
Phenotype B. bassiana O. kimflemingiae
Evidence of fungalinfection aroundmandibular muscle
+* +*
Mandibular musclehypercontraction
− +
Presence ofneuromuscularjunctions
+ +
Sarcolemma breakdown + +Fungal invasion ofmuscle
+* +*
Fungal cell appearance Elongatedinterconnectedhyphae*
Short hyphal bodiesconnected by conidialanastomosis tubes*
Extracellular vesicles + +
*Observed in both the present study and Fredericksen et al. (2017).
7
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

study, we sought to investigate how despite the extensive fungalcolonization and atrophy of mandibular muscles, manifestation ofthe death grip still occurs. We were able to show that ratherthan exhibiting overt atrophy, muscle cells are in a state ofhypercontraction and motor neuron innervation is maintained.Additionally, individual O. kimflemingiae cells form a collectivenetwork around muscle cells, can physically invade the muscleand possess numerous extracellular vesicle-like particles on theirsurface. Taken together, we hypothesize that potential mechanismsutilized by O. kimflemingiae to trigger synchronized mandibularhypercontraction may include sarcolemma degradation and directaction on host muscle by secreted factors. The results presented hereprovide a strong foundation from which to begin investigating thecontribution of these different proposed mechanisms to musclehypercontraction and the manifestation of the zombie ant death grip.
AcknowledgementsWewould like to thank the faculty and staff members of the Huck Microscopy Facilityat Penn State University including John Cantolina and Dr Greg Ning for their supportand guidance during all the imaging experiments. We would also like to thank DrsVivian Budnik andWulfila Gronenberg for their invaluable insight on, and assistancewith data interpretation.
Competing interestsThe authors declare no competing or financial interests.
Author contributionsConceptualization: C.A.M.; Methodology: C.A.M., M.J.I., R.G.L., M.L.H.; Formalanalysis: C.A.M., D.P.H.; Investigation: C.A.M., M.J.I., R.G.L., M.L.H.; Data curation:C.A.M.; Writing - original draft: C.A.M., M.J.I.; Writing - review & editing: C.A.M.,M.J.I., R.G.L., D.P.H.; Visualization: C.A.M.; Supervision: D.P.H.; Projectadministration: D.P.H.; Funding acquisition: D.P.H.
FundingThis work was supported in part by the American Heart Association(16POST29920001, C.A.M.), the National Science Foundation (grant IOS-1558062to D.P.H.) and the National Institutes of Health (grant R01 GM116927-02 to D.P.H.).Deposited in PMC for release after 12 months.
Supplementary informationSupplementary information available online athttp://jeb.biologists.org/lookup/doi/10.1242/jeb.200683.supplemental
ReferencesAdamo, S. A. (2013). Parasites: evolution’s neurobiologists. J. Exp. Biol. 216, 3-10.doi:10.1242/jeb.073601
Albuquerque, P. C., Nakayasu, E. S., Rodrigues, M. L., Frases, S., Casadevall,A., Zancope-Oliveira, R. M., Almeida, I. C. and Nosanchuk, J. D. (2008).Vesicular transport in Histoplasma capsulatum: an effective mechanism for trans-cell wall transfer of proteins and lipids in ascomycetes. Cell. Microbiol. 10,1695-1710. doi:10.1111/j.1462-5822.2008.01160.x
Andersen, S. B., Gerritsma, S., Yusah, K. M., Mayntz, D., Hywel-Jones, N. L.,Billen, J., Boomsma, J. J. and Hughes, D. P. (2009). The life of a dead ant: theexpression of an adaptive extended phenotype. Am. Nat. 174, 424-433. doi:10.1086/603640
Araujo, J. P. M., Evans, H. C., Kepler, R. and Hughes, D. P. (2018). Zombie-antfungi across continents: 15 new species and new combinations withinOphiocordyceps. I. Myrmecophilous hirsutelloid species. Stud. Mycol. 90,119-160. doi:10.1016/j.simyco.2017.12.002
Botnevik, C. F., Malagocka, J., Jensen, A. B. and Fredensborg, B. L. (2016).Relative effects of temperature, light, and humidity on clinging behavior ofmetacercariae-infected ants. J. Parasitol. 102, 495-500. doi:10.1645/16-53
Carney, W. P. (1969). Behavioral and morphological changes in carpenter antsharboring dicrocoeliid metacercariae. Am. Midl. Nat. 82, 605-611. doi:10.2307/2423801
Dawkins, R. (1982). The Extended Phenotype: the Gene as the Unit of Selection.Oxford [Oxfordshire]; San Francisco: Freeman.
de Bekker, C., Quevillon, L. E., Smith, P. B., Fleming, K. R., Ghosh, D.,Patterson, A. D. and Hughes, D. P. (2014). Species-specific ant brainmanipulation by a specialized fungal parasite. BMC Evol. Biol. 14, 166. doi:10.1186/s12862-014-0166-3
de Bekker, C., Ohm, R. A., Loreto, R. G., Sebastian, A., Albert, I., Merrow, M.,Brachmann, A. and Hughes, D. P. (2015). Gene expression during zombie antbiting behavior reflects the complexity underlying fungal parasitic behavioralmanipulation. BMC Genomics 16, 620. doi:10.1186/s12864-015-1812-x
de Bekker, C., Ohm, R. A., Evans, H. C., Brachmann, A. and Hughes, D. P.(2017). Ant-infecting Ophiocordyceps genomes reveal a high diversity of potentialbehavioral manipulation genes and a possible major role for enterotoxins. Sci.Rep. 7, 12508. doi:10.1038/s41598-017-12863-w
Dixon, R. W. and Harris, J. B. (1996). Myotoxic activity of the toxic phospholipase,notexin, from the venom of the Australian tiger snake. J. Neuropathol. Exp. Neurol.55, 1230-1237. doi:10.1097/00005072-199612000-00006
Eadie, M. J. (2003). Convulsive ergotism: epidemics of the serotonin syndrome?Lancet Neurol. Lond. 2, 429-434. doi:10.1016/S1474-4422(03)00439-3
Edwards, G. A. (1959). The fine structure of a multiterminal innervation of an insectmuscle. J. Biophys. Biochem. Cytol. 5, 241-244. doi:10.1083/jcb.5.2.241
Elya, C., Lok, T. C., Spencer, Q. E., McCausland, H., Martinez, C. C. and Eisen,M. (2018). Robust manipulation of the behavior of Drosophila melanogaster by afungal pathogen in the laboratory. eLife 7, e34414. doi:10.7554/eLife.34414
Fredericksen, M. A., Zhang, Y., Hazen, M. L., Loreto, R. G., Mangold, C. A.,Chen, D. Z. and Hughes, D. P. (2017). Three-dimensional visualization and adeep-learning model reveal complex fungal parasite networks in behaviorallymanipulated ants. Proc. Natl. Acad. Sci. USA 201711673. doi:10.1073/pnas.1711673114
Funk, C. J., Ramoska, W. A. and Bechtel, D. B. (1993). Histopathology ofentomophaga grylli pathotype 2 infections in melanoplus differentialis.J. Invertebr. Pathol. 61, 196-202. doi:10.1006/jipa.1993.1035
Gehrmann, U., Qazi, K. R., Johansson, C., Hultenby, K., Karlsson, M.,Lundeberg, L., Gabrielsson, S. and Scheynius, A. (2011). Nanovesicles frommalassezia sympodialis and host exosomes induce cytokine responses–novelmechanisms for host-microbe interactions in atopic eczema. PLoS ONE 6,e21480. doi:10.1371/journal.pone.0021480
Goulson, D. (1997). Wipfelkrankheit: modification of host behaviour duringbaculoviral infection. Oecologia 109, 219-228. doi:10.1007/s004420050076
Gryganskyi, A. P., Mullens, B. A., Gajdeczka, M. T., Rehner, S. A., Vilgalys, R.and Hajek, A. E. (2017). Hijacked: Co-option of host behavior byentomophthoralean fungi. PLoS Pathog. 13, e1006274. doi:10.1371/journal.ppat.1006274
Han, Y., van Houte, S., Drees, G. F., van Oers, M. M. and Ros, V. I. D. (2015).Parasitic manipulation of host behaviour: baculovirus SeMNPV EGT facilitatestree-top disease in spodoptera exigua larvae by extending the time to death.Insects 6, 716. doi:10.3390/insects6030716
Hawtin, R. E., Zarkowska, T., Arnold, K., Thomas, C. J., Gooday, G. W., King,L. A., Kuzio, J. A. and Possee, R. D. (1997). Liquefaction of autographacalifornica nucleopolyhedrovirus-infected insects is dependent on the integrity ofvirus-encoded chitinase and cathepsin genes. Virology 238, 243-253. doi:10.1006/viro.1997.8816
Heil, M. (2016). Host manipulation by parasites: cases, patterns, and remainingdoubts. Front. Ecol. Evol. 4. doi:10.3389/fevo.2016.00080
Hoover, K., Grove, M., Gardner, M., Hughes, D. P., McNeil, J. and Slavicek, J.(2011). A gene for an extended phenotype.Science 333, 1401-1401. doi:10.1126/science.1209199
Hopkins, P. M. (2006). Skeletal muscle physiology. Contin. Educ. Anaesth. Crit.Care Pain 6, 1-6. doi:10.1093/bjaceaccp/mki062
Huang, S.-H., Wu, C.-H., Chang, Y. C., Kwon-Chung, K. J., Brown, R. J. andJong, A. (2012). Cryptococcus neoformans-derived microvesicles enhance thepathogenesis of fungal brain infection. PLoSONE 7, e48570. doi:10.1371/journal.pone.0048570
Hughes, D. P., Andersen, S. B., Hywel-Jones, N. L., Himaman, W., Billen, J. andBoomsma, J. J. (2011). Behavioral mechanisms andmorphological symptoms ofzombie ants dying from fungal infection. BMC Ecol. 11, 13. doi:10.1186/1472-6785-11-13
Iwamoto, H. (2011). Structure, function and evolution of insect flight muscle.Biophys. 7, 21-28. doi:10.2142/biophysics.7.21
Jorgensen, W. K. and Rice, M. J. (1983). Superextension and supercontraction inlocust ovipositor muscles. J. Insect Physiol. 29, 437-448. doi:10.1016/0022-1910(83)90072-0
Josephson, R. (2009). Chapter 175-muscle system. In Encyclopedia of Insects,2nd edn (ed. V. H. Resh and R. T. Carde), pp. 675-680. San Diego: AcademicPress.
Kamita, S. G., Nagasaka, K., Chua, J. W., Shimada, T., Mita, K., Kobayashi, M.,Maeda, S. and Hammock, B. D. (2005). A baculovirus-encoded protein tyrosinephosphatase gene induces enhanced locomotory activity in a lepidopteran host.Proc. Natl. Acad. Sci. USA 102, 2584. doi:10.1073/pnas.0409457102
Katsuma, S., Koyano, Y., Kang, W. K., Kokusho, R., Kamita, S. G. and Shimada,T. (2012). The baculovirus uses a captured host phosphatase to induce enhancedlocomotory activity in host caterpillars. PLoS Pathog. 8, e1002644. doi:10.1371/journal.ppat.1002644
Kleimenova, N. N. and Belen’kii, E. E. (1975). Myelin-like structures in heartmuscle cells after administration of adrenalin. Bull. Exp. Biol. Med. 80, 1376-1379.doi:10.1007/BF00835193
8
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology

Krull, W. H. and Mapes, C. R. (1952). Studies on the biology of Dicrocoeliumdendriticum (Rudolphi, 1819) looss, 1899 (Trematoda: Dicrocoeliidae), includingits relation to the intermediate host, Cionella lubrica (Muller). VII. The secondintermediate host of Dicrocoelium dendriticum. Cornell Vet. 42, 603-604.
Lauritzen, F., Paulsen, G., Raastad, T., Bergersen, L. H. and Owe, S. G. (2009).Gross ultrastructural changes and necrotic fiber segments in elbow flexor musclesafter maximal voluntary eccentric action in humans. J. Appl. Physiol. (1985) 107,1923-1934. doi:10.1152/japplphysiol.00148.2009
Loreto, R. G., Elliot, S. L., Freitas, M. L. R., Pereira, T. M. and Hughes, D. P.(2014). Long-term disease dynamics for a specialized parasite of ant societies:a field study. PLoS ONE 9, e103516. doi:10.1371/journal.pone.0103516
Loreto, R. G., Araujo, J. P. M., Kepler, R. M., Fleming, K. R., Moreau, C. S. andHughes, D. P. (2018). Evidence for convergent evolution of host parasiticmanipulation in response to environmental conditions. Evolution 72, 2144-2155.doi:10.1111/evo.13489
Manga-Gonzalez, M. Y., Gonzalez-Lanza, C., Cabanas, E. and Campo, R.(2001). Contributions to and review of dicrocoeliosis, with special reference to theintermediate hosts of Dicrocoelium dendriticum. Parasitology 123, 91-114. doi:10.1017/S0031182001008204
Martın-Vega, D., Garbout, A., Ahmed, F., Wicklein, M., Goater, C. P., Colwell,D. D. and Hall, M. J. R. (2018). 3D virtual histology at the host/parasite interface:visualisation of the master manipulator, Dicrocoelium dendriticum, in the brain ofits ant host. Sci. Rep. 8, 8587. doi:10.1038/s41598-018-26977-2
Moore, J. (1995). The behavior of parasitized animals.Bioscience 45, 89-96. doi:10.2307/1312610
Oliveira, D. L., Freire-de-Lima, C. G., Nosanchuk, J. D., Casadevall, A.,Rodrigues, M. L. and Nimrichter, L. (2010). Extracellular vesicles fromCryptococcus neoformans modulate macrophage functions. Infect. Immun. 78,1601-1609. doi:10.1128/IAI.01171-09
Oliveira, D. L., Rizzo, J., Joffe, L. S., Godinho, R. M. C. and Rodrigues, M. L.(2013). Where do they come from and where do they go: candidates for regulatingextracellular vesicle formation in fungi. Int. J. Mol. Sci. 14, 9581-9603. doi:10.3390/ijms14059581
Panepinto, J., Komperda, K., Frases, S., Park, Y.-D., Djordjevic, J. T.,Casadevall, A. and Williamson, P. R. (2009). Sec6-dependent sorting offungal extracellular exosomes and laccase of Cryptococcus neoformans. Mol.Microbiol. 71, 1165-1176. doi:10.1111/j.1365-2958.2008.06588.x
Paul, J. andGronenberg,W. (2002). Motor control of the mandible closer muscle inants. J. Insect Physiol. 48, 255-267. doi:10.1016/S0022-1910(01)00171-8
Petrof, B. J. (2002). Molecular pathophysiology of myofiber injury in deficiencies ofthe dystrophin-glycoprotein complex. Am. J. Phys. Med. Rehabil. 81, S162-S174.doi:10.1097/00002060-200211001-00017
Rodrigues, M. L., Nimrichter, L., Oliveira, D. L., Frases, S., Miranda, K.,Zaragoza, O., Alvarez, M., Nakouzi, A., Feldmesser, M. and Casadevall, A.(2007). Vesicular polysaccharide export in Cryptococcus neoformans is aeukaryotic solution to the problem of fungal trans-cell wall transport. Eukaryot.Cell 6, 48-59. doi:10.1128/EC.00318-06
Rodrigues, M. L., Nakayasu, E. S., Oliveira, D. L., Nimrichter, L., Nosanchuk,J. D., Almeida, I. C. and Casadevall, A. (2008). Extracellular vesicles producedby Cryptococcus neoformans contain protein components associated withvirulence. Eukaryot. Cell 7, 58-67. doi:10.1128/EC.00370-07
Rodrigues, M. L., Nosanchuk, J. D., Schrank, A., Vainstein, M. H., Casadevall,A. and Nimrichter, L. (2011). Vesicular transport systems in fungi. FutureMicrobiol. 6, 1371-1381. doi:10.2217/fmb.11.112
Samuels, R. I., Reynolds, S. E. and Charnley, A. K. (1988). Calcium channelactivation of insect muscle by destruxins, insecticidal compounds produced by theentomopathogenic fungus Metarhizium anisopliae.Comp. Biochem. Physiol. PartC Comp. Pharmacol. 90, 403-412. doi:10.1016/0742-8413(88)90018-7
Schreiter, G., Butt, T. M., Beckett, A., Vestergaard, S. and Moritz, G. (1994).Invasion and development of Verticillium lecanii in the western flower thrips,Frankliniella occidentalis. Mycol. Res. 98, 1025-1034. doi:10.1016/S0953-7562(09)80429-2
Usherwood, P. N. (1967). Insect neuromuscular mechanisms. Am. Zool. 7,553-582. doi:10.1093/icb/7.3.553
Vallejo, M. C., Matsuo, A. L., Ganiko, L., Medeiros, L. C. S., Miranda, K., Silva,L. S., Freymuller-Haapalainen, E., Sinigaglia-Coimbra, R., Almeida, I. C. andPuccia, R. (2011). The pathogenic fungus paracoccidioides brasiliensis exportsextracellular vesicles containing highly immunogenic α-Galactosyl epitopes.Eukaryot. Cell 10, 343-351. doi:10.1128/EC.00227-10
Vargas, G., Rocha, J. D. B., Oliveira, D. L., Albuquerque, P. C., Frases, S.,Santos, S. S., Nosanchuk, J. D., Gomes, A. M. O., Medeiros, L. C. A. S.,Miranda, K. et al. (2015). Compositional and immunobiological analyses ofextracellular vesicles released by Candida albicans. Cell. Microbiol. 17, 389-407.doi:10.1111/cmi.12374
Zamith-Miranda, D., Nimrichter, L., Rodrigues, M. L. and Nosanchuk, J. D.(2018). Fungal extracellular vesicles: modulating host–pathogen interactions byboth the fungus and the host. Microbes Infect. 20, 501-504. doi:10.1016/j.micinf.2018.01.011
9
RESEARCH ARTICLE Journal of Experimental Biology (2019) 222, jeb200683. doi:10.1242/jeb.200683
Journal
ofEx
perim
entalB
iology