Embed Size (px)
Citation preview

Plant Molecular Biology39: 1073–1078, 1999.© 1999Kluwer Academic Publishers. Printed in the Netherlands.
1073
Mini-review
Zinc-finger proteins: the classical zinc finger emerges in contemporaryplant science
Hiroshi TakatsujiLaboratory of Developmental Biology, National Institute of Agrobiological Resources, 2-1-2 Kannondai, Tsukuba,Ibaraki 305, Japan
Received 30 November 1998; accepted in revised form 15 December 1998
Key words:development, gametogenesis, stress response, target-sequence recognition, TFIIIA-type zinc finger,transcription factor
Abstract
TFIIIA-type zinc fingers have been found in a number of eucaryotic transcription factors as DNA-binding motifs.In plants, as many as 30 proteins have been reported that have either one, two, three or four zinc fingers. Plantzinc-finger proteins are characterized by long spacers of diverse lengths between adjacent fingers and a highlyconserved sequence, QALGGH, located within a putative DNA-contacting surface of each finger.In vitro DNA-binding experiments with two-fingered proteins of petunia have revealed that these proteins bind to target DNAsequences in a manner that is distinctive from that of their animal counterparts: (1) they specifically recognizethe spacing between two core sites in target DNA, (2) they have a unique base-determinant position. Regulatoryfunctions have been assigned to some of the TFIIIA-type zinc finger proteins inArabidopsis, petunia and chinesecabbage. SUPERMAN, AtZFP1, PetSPL3 and BcZFP1 have been implicated in the developmental regulation ofvarious floral and vegetative organs, presumably through the control of cell division and/or expansion in particularcell types. Several anther-specific zinc-finger proteins in petunia are presumed to be involved in the regulation ofgametogenesis in both reproductive and non-reproductive tissues of anther. STZ and ZPT2-2 are implicated in theresponse of plants to or tolerance for various stresses.
Introduction
The TFIIIA-type zinc finger (also referred to asCys2/His2 or classical zinc finger) is a sequence ofCX2−4FX5LX2HX3−5H in which two cysteines andtwo histidines tetrahedrally coordinate a zinc atom toform a compact finger structure containing aβ-hairpinand anα-helix. Structural studies have revealed thatthe α-helix makes contact with the major grooveof DNA (Choo and Klug, 1997). Recently, how-ever, some of the TFIIIA-type zinc fingers in certaintranscription factors have been implicated in protein-protein interaction, while the other fingers in the sametranscription factors interact with DNA (Mackay andCrossley, 1998). The first TFIIIA-type zinc finger inplants was found in a DNA-binding protein of petu-nia, ZPT2-1 (renamed from EPF1), that interacts with
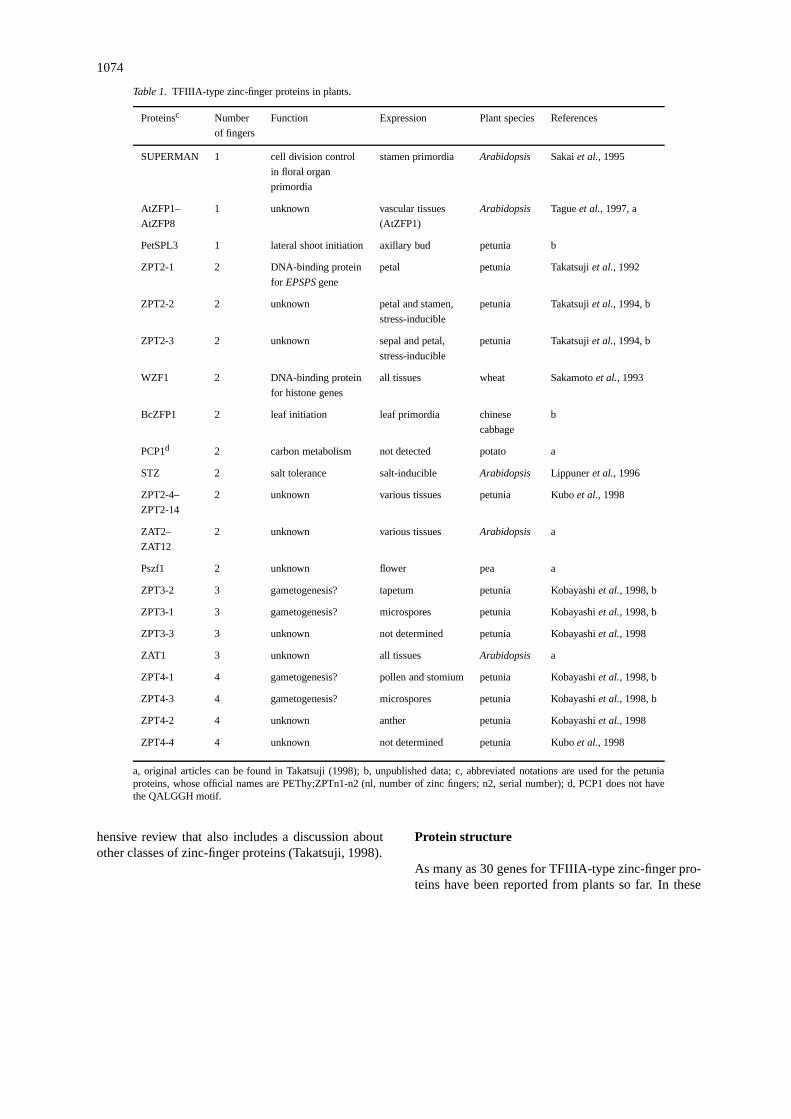
the promoter region of the 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene (Takatsujiet al.,1992). Subsequently, WZF1 was identified in wheat asa DNA-binding protein that specifically interacts withaciselement of histone genes (Sakamotoet al., 1993).Thereafter, several proteins that contain the TFIIIA-type zinc finger have been reported from petunia,Arabidopsisand other plant species as summarized inTable 1. Although TFIIIA-type zinc-finger proteins inplants have not been characterized as extensively asother classes of plant transcription factors, zinc-fingerproteins have attracted more attention as importantfunctions are assigned to the members of this pro-tein family. In this article, I review recent progressin the functional characterization of TFIIIA-type zinc-finger proteins in plants including several unpublishedresults. Another recent article presents a more compre-
1074
Table 1. TFIIIA-type zinc-finger proteins in plants.
Proteinsc Number Function Expression Plant species References
of fingers
SUPERMAN 1 cell division control stamen primordia Arabidopsis Sakaiet al., 1995
in floral organ
primordia
AtZFP1– 1 unknown vascular tissues Arabidopsis Tagueet al., 1997, a
AtZFP8 (AtZFP1)
PetSPL3 1 lateral shoot initiation axillary bud petunia b
ZPT2-1 2 DNA-binding protein petal petunia Takatsujiet al., 1992
for EPSPSgene
ZPT2-2 2 unknown petal and stamen, petunia Takatsujiet al., 1994, b
stress-inducible
ZPT2-3 2 unknown sepal and petal, petunia Takatsujiet al., 1994, b
stress-inducible
WZF1 2 DNA-binding protein all tissues wheat Sakamotoet al., 1993
for histone genes
BcZFP1 2 leaf initiation leaf primordia chinese b
cabbage
PCP1d 2 carbon metabolism not detected potato a
STZ 2 salt tolerance salt-inducible Arabidopsis Lippuneret al., 1996
ZPT2-4– 2 unknown various tissues petunia Kuboet al., 1998
ZPT2-14
ZAT2– 2 unknown various tissues Arabidopsis a
ZAT12
Pszf1 2 unknown flower pea a
ZPT3-2 3 gametogenesis? tapetum petunia Kobayashiet al., 1998, b
ZPT3-1 3 gametogenesis? microspores petunia Kobayashiet al., 1998, b
ZPT3-3 3 unknown not determined petunia Kobayashiet al., 1998
ZAT1 3 unknown all tissues Arabidopsis a
ZPT4-1 4 gametogenesis? pollen and stomium petunia Kobayashiet al., 1998, b
ZPT4-3 4 gametogenesis? microspores petunia Kobayashiet al., 1998, b
ZPT4-2 4 unknown anther petunia Kobayashiet al., 1998
ZPT4-4 4 unknown not determined petunia Kuboet al., 1998
a, original articles can be found in Takatsuji (1998); b, unpublished data; c, abbreviated notations are used for the petuniaproteins, whose official names are PEThy;ZPTn1-n2 (nl, number of zinc fingers; n2, serial number); d, PCP1 does not havethe QALGGH motif.
hensive review that also includes a discussion aboutother classes of zinc-finger proteins (Takatsuji, 1998).
Protein structure
As many as 30 genes for TFIIIA-type zinc-finger pro-teins have been reported from plants so far. In these

1075
proteins, the number of zinc fingers ranges from oneto four. There are two prominent structural featuresof plant TFIIIA-type zinc-finger proteins which distin-guish them from their counterparts in other eucaryotes.One is the long spacers of diverse lengths betweenadjacent fingers. This is in contrast to TFIIIA-typezinc-finger proteins in animals that mostly have spac-ers of short and invariant length (seven amino acids) assummarized in Figure 1B. Another structural featureof the plant zinc-finger proteins is the high conser-vation of a unique sequence, QALGGH, within aputative DNA-contacting surface of each finger (Fig-ure 1A). Of all the TFIIIA-type zinc-finger proteinsreported so far, only one lacks this particular sequenceor a close relative.In vitro binding assays with mutag-enized proteins have demonstrated that this conservedsequence is critical for DNA-binding activity (Kuboet al., 1998). Interestingly, this sequence has so far notbeen found in the zinc fingers of any other organisms,suggesting that it may have evolved in association withplant-specific regulatory processes. All the proteinsreferred to in this article contain the zinc fingers in-cluding the QALGGH motif. The zinc fingers havingthis sequence motif will be referred to as the EPF typehereafter.
Recognition of target-DNA sequence
The structure of the EPF proteins suggests that theymay interact in unique manner with their target DNAwhich is distinct from that of the TFIIIA-type zinc-finger proteins in animals. Understanding the mecha-nism of this protein-DNA interaction would certainlybe beneficial to the molecular characterization of pro-teins. Our preliminary characterization of the DNA-binding activities of some EPF proteins of petuniahave revealed some unique protein-DNA interactionswhich are described below.
Significance of the diverse lengths of spacers betweenadjacent fingers
In most TFIIIA-type zinc-finger proteins in animals,multiple zinc fingers are present as tandem arraysseparated by short conserved sequences of invariantlength (7 amino acids). These proteins recognize con-tiguous sets of triplet DNA sequences, with eachtriplet being recognized by one finger. By contrast, atwo-fingered petunia EPF protein, ZPT2-2 (renamedfrom EPF2-5), with a spacer of 44 amino acids, binds
to two separate AGT core sites with each finger mak-ing contact with one core site (Takatsujiet al., 1994;Takatsuji and Matsumoto, 1996). This interaction issensitive to the spacing between the two core sites, theoptimum being 13 bp. Another two-fingered protein,ZPT2-1 (renamed from EPF1), which has a spacerof 61 amino acids, binds to the same core sites buttolerates a spacing of 13 to 16 bp. In petunia, 14 two-fingered proteins have been identified so far and thelengths of their spacers range from 19 to 65 aminoacids (Figure 1B) (Kuboet al., 1998). Given the differ-ent preference for the spacing of ZPT2-2 and ZPT2-1,it is tempting to speculate that these two-fingered pro-teins with such diverse lengths of spacers have rathervariable preferences for the spacings in the target DNAsequence. Thus, in multiple-fingered proteins, the di-versity of the length of spacers might contribute to thedifferent specificity for target DNA sequence of eachprotein. To test this hypothesis, we are conductingsimilar experiments with other members of the proteinfamily. A similar argument for three- and four-fingeredproteins has been discussed comprehensively by Kuboet al. (1998).
Based-determinant position
According to the studies with the TFIIIA-type zinc-finger proteins in animals, three positions (base-determinant positions) in theα-helical region of thezinc finger participate in major interactions with se-quences in target DNA. These positions are hypervari-able among different zinc fingers thereby conferringvariable specificities for triplets in target sequence.Combinations of zinc fingers with the specificities fordifferent triplet sequences generate a huge diversity oftarget sequences. In the EPF-type zinc fingers, twopositions within the highly conserved QALGGH se-quence (underlined) correspond to two of the threebase-determinant positions of the animal zinc fingers.If the same positions are used for base determinationin plant zinc fingers then the repertoire of target se-quences would be greatly limited for the plant proteinsdue to their high conservation. (Moreover, the con-served glycine at position 6 has no side chain to inter-act with bases in DNA, see Figure 1A). These circum-stances led us to look for other base-determinant posi-tions by domain swapping and amino acid substitutionexperiments using recombinant proteins for ZPT2-2 and ZPT2-3 (Takatsuji, 1996). These experimentshave indicated that position 10 is a base-determinantposition in the EPF-type zinc finger (Figure 1A). Po-

1076
Figure 1. Structure of TFIIIA-type zinc finger proteins in plants. A. EPF-type zinc-finger consensus sequence. Positions of amino acids arenumbered relative to the first putative helical residue (+1). Putativeα-helical region is shadowed and the conserved QALGGH sequence boxed.The three positions corresponding to the base-determinant positions in animal zinc-finger proteins are shown by white letters. One of thebase-determinant positions in EPF-type zinc finger is circled. B. Schematic representations of protein structure. Zinc finger regions are shaded.Shown below each schematic structure are the numbers of amino acids between adjacent zinc fingers.
sition 10 lies outside the QALGGH sequence and isdifferent from any of the three positions in animalzinc fingers. This position, however, is still withinthe putativeα-helical region. Further characterizationof protein-DNA interaction including the search forother base-determinant positions is in progress usingapproaches such as nuclear magnetic resonance.
Developmental regulation
Several proteins that play important regulatory rolesin development are TFIIIA-type zinc-finger proteins.SUPERMANmutants ofArabidopsisproduce extrastamens at the cost of normal carpel development. Be-fore its cloning, the function ofSUPERMANgene wasassumed to be suppression of the expression of B classorgan identity genes in the fourth whorl because ec-topic expression of the B class genes in this whorl,in combination with C class genes, would lead to thegeneration of stamens, consistent with the mutant phe-notype. TheSUPERMANgene was isolated by map-based cloning and found to encode a single TFIIIA-type zinc finger containing the conserved QALGGHmotif (Sakai et al., 1995). Expression analysis re-vealed this gene to be under the control of B classgenes, just the opposite from what was originallypresumed.In situ hybridization studies demonstratedthatSUPERMANtranscripts were present in the sub-
domain of the third-whorl primordia adjacent to thefourth whorl. A revised model based on the expres-sion analyses in wild-type and mutant plants was thenproposed suggesting thatSUPERMANacts to represscell division in the third-whorl primordia by defininga boundary between the third- and the fourth-whorlcells. According to this model, generation of extra sta-mens in place of carpels in theSUPERMANmutant isdue to the alteration in the boundary resulting in ex-cessive cell divisions in the third-whorl meristem andin suppressed proliferation of cells in the fourth whorl.
AtZFP1 is one of the eight known proteins ofArabidopsiswhich contain single TFIIIA-type zincfingers including the QALGGH motif (Tagueet al.,1997). Fusions of the AtZFP1 promoter to the GUSreporter gene revealed that AtZFP1 is first expressedat the apical region of seedlings. Later, it is expressedat higher levels in vascular and surrounding tissuesof petioles and in leaf veins. Interestingly, whentransgenicArabidopsislines overexpressing AtZFP1were generated using tissue-culture-based transforma-tion, transformed calli remained green and healthy butfailed to regenerate shoots. Tague and coworkers spec-ulated that it might be due to tissue-specific repressionof cell proliferation in callus by the ectopic expres-sion of AtZFP1 by analogy to the proposed functionof SUPERMAN.
We have identified several genes for single-fingered proteins from petunia (Nakagawaet al., un-

1077
published results). These proteins can be dividedinto two groups based on the relative similarity oftheir zinc-finger sequences to either SUPERMAN orAtZFPs. Recently, one of these genes encoding a zincfinger more similar to that of SUPERMAN (tentativelydesignated asPetSPL3) was found to induce vigorousbranching in transgenic plants when its cDNA is over-expressed under the control of the cauliflower mosaicvirus (CaMV) 35S promoter (Nakagawaet al., un-published results). These transformants produce first-and second-order branches at almost every leaf axil,whereas wild-type plants form only a few first-orderbranches and rarely initiate second-order branchingunder normal conditions. Promoter activity of thePetSPL3upstream sequences linked to the GUS re-porter gene has been detected at the base of axillarybuds. Upon decapitation of a shoot apex,PetSPL3expression rapidly disappears after the outgrowth ofaxillary buds has initiated. These results suggest thatthe gene is involved in the initiation of growth in ax-illary buds and plays a key role in the regulation ofapical dominance in response to hormone-mediatedenvironmental cues.
BcZFP1 is a two-fingered protein ofBrassicacampestris. Lee and coworkers have reported thatthe overexpression of BcZFP1 in transgenic tobaccounder the control of the CaMV 35S promoter re-tards the growth of the transformants and subsequentlytransforms the shoot apical meristem into a leaf pri-mordium, resulting in the termination of the mainshoot (J.S. Lee, personal communication).BcZFP1promoter-driven GUS expression was detected at thebase of leaf primordia initiating from the shoot apicalmeristem. GUS expression was not present in the cen-tral zone of the shoot apical meristem but was confinedto the cells underneath the outer three layers of thecells in the peripheral zone, with its timing concurrentwith the expansion of cells in the peripheral zone priorto developing into leaf primordia. Lee and coworkerspropose that BcZFP1 might provide positional infor-mation by producing factors that direct the cells in theperipheral zone to undergo a precise sequence of ge-netically programmed steps leading to leaf initiationfrom the shoot apical meristem.
Tissue- and development-specific zinc-fingerproteins expressed in anther
During pollen development, a series of geneticallyprogrammed events occur both in reproductive and
non-reproductive anther cells. The genetic control ofthese developmental events is presumed to be largelymediated by unknown transcription factors. In an at-tempt to identify such transcription factors, we haveobtained cDNAs for seven zinc-finger transcriptionfactors of the EPF-type containing either two, three,or four zinc fingers that express specifically or prefer-entially in anther (Kobayashiet al., 1998). The sevengenes are activated transiently at different develop-mental stages and, as a whole, appear to be expressedsequentially from meiosis through dehiscence, as ifthey form a regulatory cascade. Our recent resultsfrom a more detailed analysis using promoter-GUSfusion genes demonstrate that four of these genes aredifferent in their tissue-specific expression patterns(Kapoor et al., unpublished results).ZPT3-2 is ex-pressed specifically in the tapetal tissue that supportsthe development of male gametophytes by providingnutrients and lytic enzymes, and subsequently under-goes programmed cell death. TheZPT3-2expressionlasts from the tetrad stage through the degeneration oftapetal layer.ZPT4-3andZPT3-1are both expressedspecifically in the microspores at overlapping timesduring the uninucleate stage.ZPT2-6 is expressedin mature pollen grains and in stomium cells. Thestomium cells constitute a dehiscence zone that dis-integrates to release mature pollen grains by causinga split in the anther wall. Coincidentally,ZPT2-6 isexpressed transiently just prior to dehiscence. Thesetissue- and developmental-stage-specific expressionpatterns of the zinc-finger genes suggest their involve-ment in the regulation of developmental events duringgametogenesis. For more direct functional character-ization of respective transcription factors, transgenicand transposon-induced gene knockout experimentsare under way.
Involvement in stress-related regulation
Some of the TFIIIA-type zinc-finger proteins havebeen implicated in the regulation of responses to ortolerance for various stresses. STZ was identified dur-ing screening ofArabidopsisgenes that complementthe salt-sensitive phenotype of a yeast mutant whichis deficient in the phosphoprotein phosphatase, cal-cineurin (Lippuneret al., 1996). The expression of theSTZgene increases with salt treatment of plants. Takentogether, it suggests that STZ is likely to be involvedin the salt tolerance ofArabidopsis. The gene for petu-nia ZPT2-2 was initially identified as being expressed

1078
specifically in petals and stamens of petunia (Takat-suji et al., 1994). Using a luciferase (LUC) reportergene, the promoter ofZPT2-2was found to be respon-sive to various stresses and plant hormones in all planttissues whereas it was constitutively expressed in thetwo floral organs (van der Krolet al., manuscript inpreparation). The activity ofZPT2-2::LUCin leaf tis-sue is highly induced upon wounding, cold, UV-B anddesiccation treatments. In petals, it increases dramati-cally upon pollination from a basal level and declinesconcomitantly with petal wilting. The upregulation oftheZPT2-2::LUCactivity can be blocked by inhibitorsof ethylene perception. These expression patterns sug-gest that ZPT2-2 may play a role in ethylene hormonalresponses induced by stresses.
Conclusion and future prospects
Several TFIIIA-type zinc-finger proteins in plantshave been implicated in various important regulatoryprocesses, viz. morphogenesis, gametogenesis, andstress responses. These investigations have providedclues as to how the proteins work. In the future, thesearch for target genes of zinc-finger transcription fac-tors will be one of the most important directions ofresearch. The rules of protein-DNA interaction re-vealed by virtue of petunia proteins will probably helpsuch investigations.
Acknowledgements
I am grateful to Drs Sanjay Kapoor, Hajime Sakaiand Takashi Aoyama for critical reading of the man-uscript. This work was supported by the Enhance-ment of Center of Excellence, Special CoordinationFunds for Promoting Science and Technology fromthe Science and Technology Agency of Japan and by aPROBRAIN grant from the Bio-Oriented TechnologyResearch Advancement Institution (BRAIN) of Japan.
References
Choo, Y. and Klug, A. 1997. Physical basis of a protein-DNArecognition code. Curr. Opin. Struct. Biol. 7: 117–125.
Kobayashi, A., Sakamoto, A., Kubo, K., Rybka, Z., Kanno, Y.and Takatsuji, H. 1998. Seven zinc-finger transcription factorsare expressed sequentially during the development of anthers inpetunia. Plant J. 13: 571–576.
Kubo, K., Sakamoto, A., Kobayashi, A., Rybka, Z., Kanno, Y.,Nakagawa, H., Nishino, T., and Takatsuji, H. 1998. Cys2/His2zinc-finger protein family of petunia: evolution and generalmechanism of target-sequence recognition. Nucl. Acids Res. 26:608–616.
Lippuner, V., Cyert, M.S. and Gasser, C.S. 1996. Two classes ofplant cDNA clones differentially complement yeast calcineurinmutants and increase salt tolerance of wild-type yeast. J. Biol.Chem. 271: 12859–12866.
Mackay, J.P. and Crossley, M. 1998. Zinc fingers are stickingtogether. Trends Biochem. Sci. 23: 1–4.
Sakai, H., Medrano, L.J. and Meyerowitz, E.M. 1995. Role of SU-PERMAN in maintainingArabidopsisfloral whorl boundaries.Nature 378: 199–203.
Sakamoto, A., Minami, M., Huh, G.H. and Iwabuchi, M. 1993.The putative zinc-finger protein WZF1 interacts with a cis-actingelement of wheat histone genes. Eur. J. Biochem. 217: 1049–1056.
Tague, B.W., Gallant, P. and Goodman, H.M. 1997. Expressionanalysis of anArabidopsisC2H2 zinc finger protein gene. PlantMol. Biol. 32: 785–796.
Takatsuji, H. 1996. A single amino acid determines the specificityfor the target sequence of two zinc-finger proteins in plants.Biochem. Biophys. Res. Commun. 224: 219–223.
Takatsuji, H. 1998. Zinc-finger transcription factors in plants. Cell.Mol. Life Sci. 54: 582–596.
Takatsuji, H. and Matsumoto, T. 1996. Target-sequence recognitionby separate-type Cys2 /His2 zinc finger proteins in plants. J. Biol.Chem. 271: 23368–23373.
Takatsuji, H., Mori, M., Benfey, P.N., Ren, L. and Cua, N.-H. 1992.Characterization of a zinc finger DNA-binding protein expressedspecifically inPetuniapetals and seedlings. EMBO J. 11: 241–249.
Takatsuji, H., Nakamura, N. and Katsumoto, Y. 1994. A new fam-ily of zinc finger proteins inPetunia: structure, DNA sequencerecognition, and floral organ-specific expression. Plant Cell 6:947–958.





























