Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, June 1986, p. 1332-1342 Vol. 51, No. 60099-2240/86/061332-11$02.00/0Copyright X 1986, American Society for Microbiology
Microbial Transformation of Quinoline by a Pseudomonas sp.tONKAR P. SHUKLA
Division ofBiochemistry, Central Drug Research Institute, Lucknow-226001, India
Received 11 March 1985/Accepted 22 November 1985
A Pseudonas sp. Isolted from sewage by enricument culture on quinoline metabolized this substrate bya novel pathway invovig S-hydroxycoumarinb During early growth of the organism on quinoline, 2-kydroxyquhmline accua tas the intermediate; 8-hydroasycoumnrin accumulated as the major metaboliteon further incebaten. 2,S-Dlydroxyquiollne and 2,3-dihydroxyphenylprophinic acid were identified as theother intermediates. Iidbition of quinole metabolism by 1 mM sodium arsenite led to the accumulation ofpyruvate, whereas inhbiton by 5 atM arseiti resuled in the accumulation of 2-hydroxyquinoline as the majormetaboiNte and 2,-Ehydroxyquine as the minor metabolite. Coumarin was not utilized as a growthsubstrate by this bacteriin, but quindlinegrown cells converted it to 2-hydroxyphenylpropionic acid, whichwas not further metaholzed. Quinoline, 2-hydroxyqalnoline, 8-hydroxycoumarin, and 2,3-dihy-droxyphenylproplonke acid were rapidly oxidized by quinoline-adapted cells, whereas 2,8-dihy-diroxyquinoNne was oxidized very slowly. Quinoline catabolism in this Pseudomonas sp. is therefore initiated byhydroxylation(s) of the nolecule followed by cleavage of the pyridine ring to yield 8-hydroxycoumgrin, whichis further metabolized via 2,3-dihydroxyphenylpropioniic acid.
Quinoline and its derivatives occur widely in coal tar,bone oil, oil shale (1, 21), and plant alkaloids (18) ard serveas intermediates and solvents in the chemical industry (27).Quinoline derivatives have antimalarial, antiamoebic,anticoccidial, ard antifungal activities (43) and have alsobeen shown to display toxic, mutagenic, and carcinogenicactivities (27). Quinoline has been reported to serve as asource of nitrogen for oats and corn (15), probably owing tomineralization of nitrogen by microbial action (28). In recentyears, quinoline has been found to be rapidly degraded insewage (6), with a half-life of 2 h. A Moraxella sp. (17) anda Pseudomonas sp. (35) were isolated from soil by enrich-mnent culture and were found to utilize quinoline as the solesource of carbon, nitrogen, and energy. The Moraxella sp.accumulated 2-hydroxyquinoline, and the involvement of2,6-dihydroxyquinoline as intermediate was postulated ow-ing to its oxidation by the quinoline-adapted cells. Naphtha-lene-grown cells ofPseudomonas putida have been reportedto convert quinoline to o-aminophenyl-p-hydroxypropionicacid (22). Animal systems metabolize quinoline byhydroxylation and excretion of glucuronide and sulfate con-jugates (42), and further ring cleavage has not been reportedih animal systems. The degradation of the quinoline deriva-tive kynurenic acid, an intermediate in tryptophan metabo-lism, occurs by dihydroxylation and meta-cleavage of thebenzene ring (11, 38). The mode of degradation of quinolineitself in bacteria is still not known. The present paper reportsthe isolation of a Pseudomonas sp. which degrades quinolineand the characterization of 2-hydroxyquinoline, 2,8-dihydroxyquinoline, 8-hydroxycoumarin, and 2,3-dihy-droxyphenyl propionic acid as intermediates of quinolinecatabolism in this organism.
MATERIALS AND METHODSReagents and chemicals. Quinoline and 8-hydroxyquinoline
were products ofBDH, Poole, England; quinoline was furtherpurified by distillation. The following compounds weresynthesized by published methods: 2-hydroxyquinoline (7),
t Central Drug Research Institute communication no. 3680.
6-hydroxyquinoline (20), 1,8-dihydroxyquinoline (12), 8-hydroxycoumarin (8), phenylpropionic acid (40), meliloticacid, 2-hydroxyphenylpropionic acid (24), 2,3-dihydroxy-trans-cinnamic acid (5), and 2,3-dihydroxyphenylpropionicacid (4). Reagent-grade chemicals were used for thepreparation of media and other biochemical studies.
Cell-
(i) Extract with ethylacetate and (ii)acidify (phosphoricacid), saturate(NaCI), and extractwith ethyl acetate
Ethyl a
NaHCO3 extract
-free broth
cetate extract
5% aqueous NaHCO3
Ethyl acetate layer
Cool, acidify, andextract withethyl acetate
Na2CO3 extract
lO0o aqueousNa2CO3
Ethyl acetate extract
Acidify andextract withethyl acetate
ACIDS PHENOLIC COMPOUNDS
Ethyl acetate Acidextract QUIN
2-HYDROXYQUINOLINEFIG. 1. Fractionation bf quinoline metabolites.
0.1 NaqueousH2SO4
I extract[OLINE
1332
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
MICROBIAL TRANSFORMATION OF QUINOLINE 1333
0.
E
CN
tn4
0Cz0I-zm-umz0In
r-
Hr.
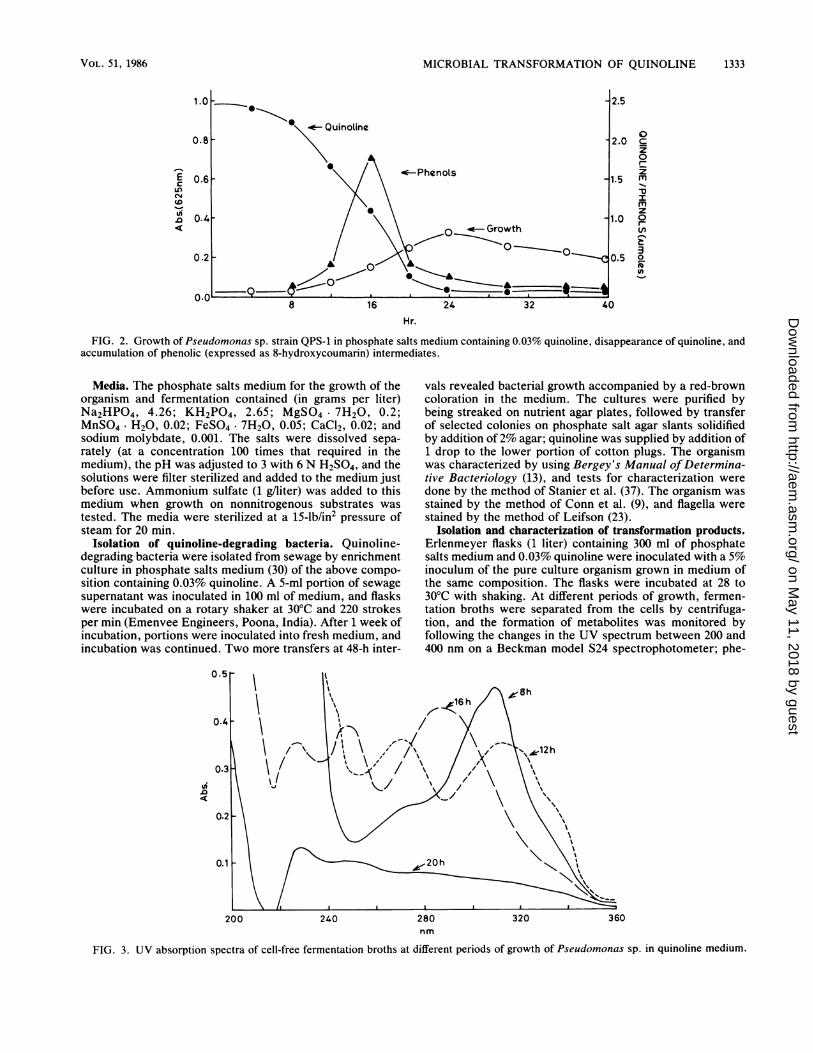
FIG. 2. Growth of Pseudomonas sp. strain QPS-1 in phosphate salts medium containing 0.03% quinoline, disappearance of quinoline, andaccumulation of phenolic (expressed as 8-hydroxycoumarin) intermediates.
Media. The phosphate salts medium for the growth of theorganism and fermentation contained (in grams per liter)Na2HPO4, 4.26; KH2PO4, 2.65; MgSO4 7H20, 0.2;MnSO4. H20, 0.02; FeSO4- 7H20, 0.05; CaCl2, 0.02; andsodium molybdate, 0.001. The salts were dissolved sepa-rately (at a concentration 100 times that required in themedium), the pH was adjusted to 3 with 6 N H2SO4, and thesolutions were filter sterilized and added to the medium justbefore use. Ammonium sulfate (1 glliter) was added to thismedium when growth on nonnitrogenous substrates wastested. The media were sterilized at a 15-lb/in2 pressure ofsteam for 20 min.
Isolation of quinoline-degrading bacteria. Quinoline-degrading bacteria were isolated from sewage by enrichmentculture in phosphate salts medium (30) of the above compo-sition containing 0.03% quinoline. A 5-ml portion of sewagesupernatant was inoculated in 100 ml of medium, and flaskswere incubated on a rotary shaker at 30°C and 220 strokesper min (Emenvee Engineers, Poona, India). After 1 week ofincubation, portions were inoculated into fresh medium, andincubation was continued. Two more transfers at 48-h inter-
0.5
0.4
0.3 \/\\'
vals revealed bacterial growth accompanied by a red-browncoloration in the medium. The cultures were purified bybeing streaked on nutrient agar plates, followed by transferof selected colonies on phosphate salt agar slants solidifiedby addition of2% agar; quinoline was supplied by addition of1 drop to the lower portion of cotton plugs. The organismwas characterized by using Bergey's Manual ofDetermina-tive Bacteriology (13), and tests for characterization weredone by the method of Stanier et al. (37). The organism wasstained by the method of Conn et al. (9), and flagella werestained by the method of Leifson (23).
Isolation and characterization of transformation products.Erlenmeyer flasks (1 liter) containing 300 ml of phosphatesalts medium and 0.03% quinoline were inoculated with a 5%inoculum of the pure culture organism grown in medium ofthe same composition. The flasks were incubated at 28 to30°C with shaking. At different periods of growth, fermen-tation broths were separated from the cells by centrifuga-tion, and the formation of metabolites was monitored byfollowing the changes in the UV spectrum between 200 and400 nm on a Beckman model S24 spectrophotometer; phe-
360
FIG. 3. UV absorption spectra of cell-free fermentation broths at different periods of growth of Pseudomonas sp. in quinoline medium.
200 240 280 320nm
VOL. 51, 1986
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
12.5 10.0 7.5 5.0 2.5
Time (min)
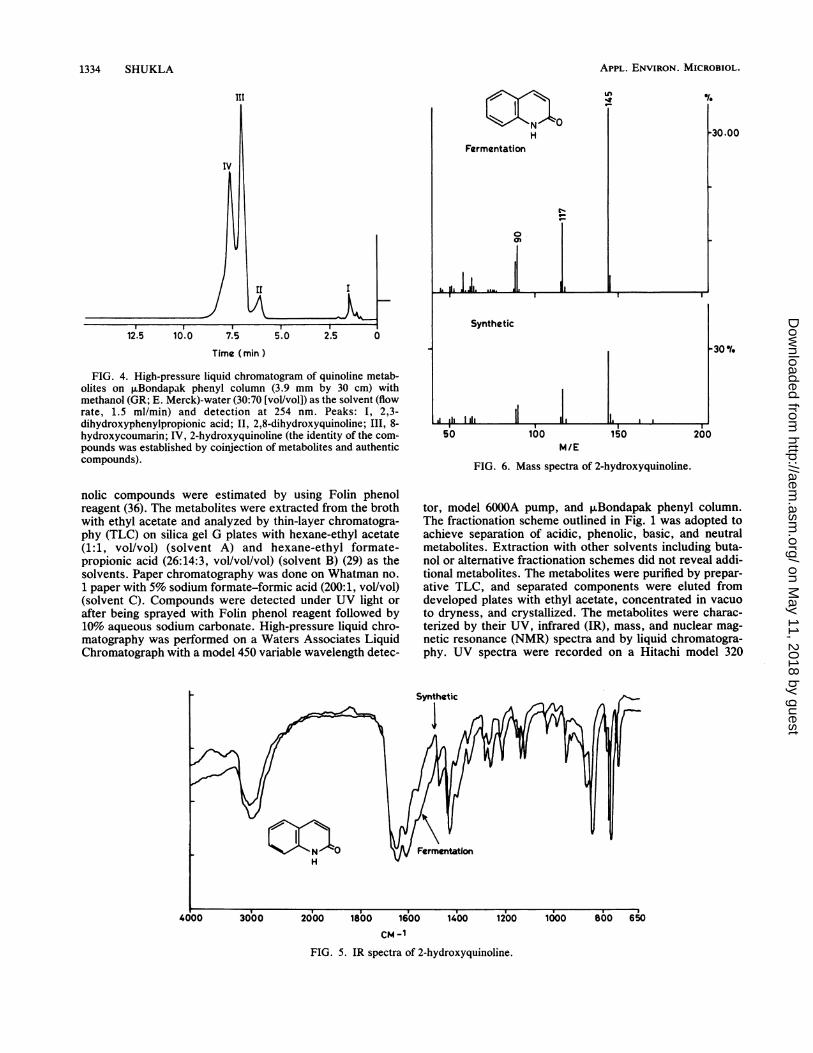
FIG. 4. High-pressure liquid chromatogram of quinoline metab-olites on ,uBondapak phenyl column (3.9 mm by 30 cm) withmethanol (GR; E. Merck)-water (30:70 [vol/vol]) as the solvent (flowrate, 1.5 ml/min) and detection at 254 nm. Peaks: I, 2,3-dihydroxyphenylpropionic acid; II, 2,8-dihydroxyquinoline; III, 8-hydroxycoumarin; IV, 2-hydroxyquinoline (the identity of the com-pounds was established by coinjection of metabolites and authenticcompounds).
nolic compounds were estimated by using Folin phenolreagent (36). The metabolites were extracted from the brothwith ethyl acetate and analyzed by thin-layer chromatogra-phy (TLC) on silica gel G plates with hexane-ethyl acetate(1:1, vol/vol) (solvent A) and hexane-ethyl formate-propionic acid (26:14:3, vol/vol/vol) (solvent B) (29) as thesolvents. Paper chromatography was done on Whatman no.1 paper with 5% sodium formate-formic acid (200:1, vol/vol)(solvent C). Compounds were detected under UV light orafter being sprayed with Folin phenol reagent followed by10% aqueous sodium carbonate. High-pressure liquid chro-matography was performed on a Waters Associates LiquidChromatograph with a model 450 variable wavelength detec-
30.00
-30 ¶.
M/E
FIG. 6. Mass spectra of 2-hydroxyquinoline.
tor, model 6000A pump, and ,uBondapak phenyl column.The fractionation scheme outlined in Fig. 1 was adopted toachieve separation of acidic, phenolic, basic, and neutralmetabolites. Extraction with other solvents including buta-nol or alternative fractionation schemes did not reveal addi-tional metabolites. The metabolites were purified by prepar-ative TLC, and separated components were eluted fromdeveloped plates with ethyl acetate, concentrated in vacuoto dryness, and crystallized. The metabolites were charac-terized by their UV, infrared (IR), mass, and nuclear mag-netic resonance (NMR) spectra and by liquid chromatogra-phy. UV spectra were recorded on a Hitachi model 320
CM -1
FIG. 5. IR spectra of 2-hydroxyquinoline.
1334 SHUKLA
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
MICROBIAL TRANSFORMATION OF QUINOLINE 1335
Fermentation
a
N
H
7
Synthetic
7ppm (6)
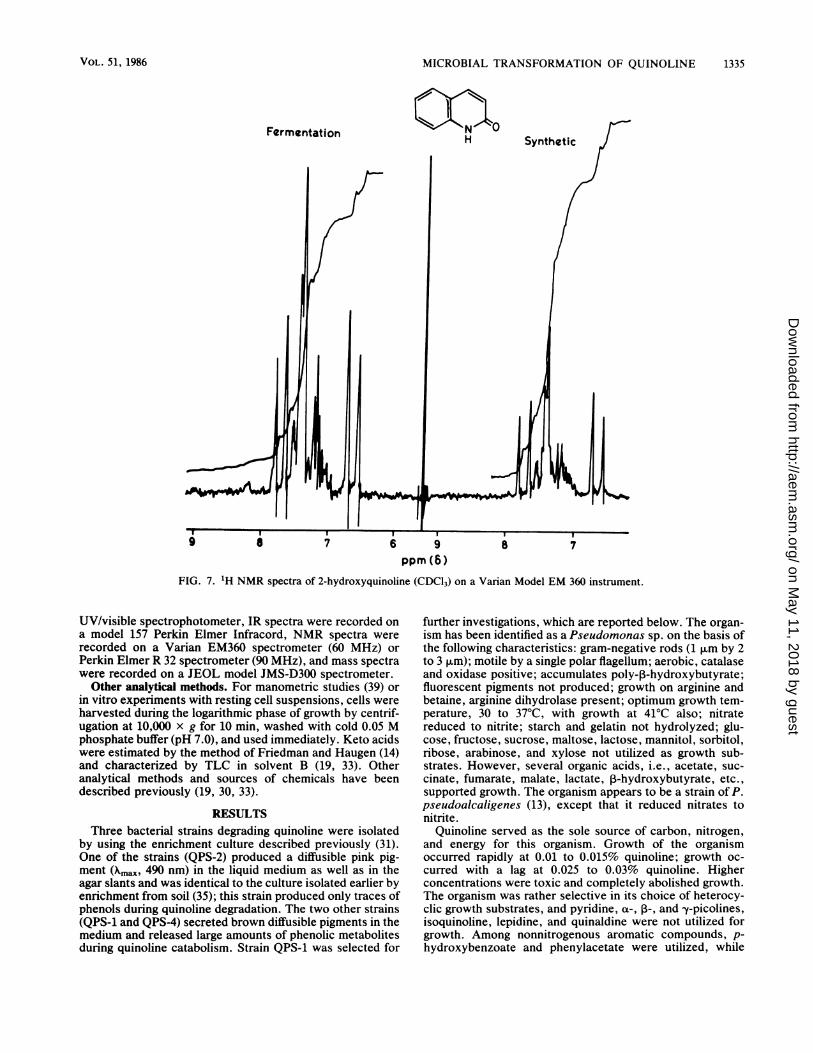
FIG. 7. 'H NMR spectra of 2-hydroxyquinoline (CDCl3) on a Varian Model EM 360 instrument.
UV/visible spectrophotometer, IR spectra were recorded ona model 157 Perkin Elmer Infracord, NMR spectra wererecorded on a Varian EM360 spectrometer (60 MHz) orPerkin Elmer R 32 spectrometer (90 MHz), and mass spectrawere recorded on a JEOL model JMS-D300 spectrometer.Other analytical methods. For manometric studies (39) or
in vitro experiments with resting cell suspensions, cells wereharvested during the logarithmic phase of growth by centrif-ugation at 10,000 x g for 10 min, washed with cold 0.05 Mphosphate buffer (pH 7.0), and used immediately. Keto acidswere estimated by the method of Friedman and Haugen (14)and characterized by TLC in solvent B (19, 33). Otheranalytical methods and sources of chemicals have beendescribed previously (19, 30, 33).
RESULTSThree bacterial strains degrading quinoline were isolated
by using the enrichment culture described previously (31).One of the strains (QPS-2) produced a diffusible pink pig-ment (Xm,[,,, 490 nm) in the liquid medium as well as in theagar slants and was identical to the culture isolated earlier byenrichment from soil (35); this strain produced only traces ofphenols during quinoline degradation. The two other strains(QPS-1 and QPS-4) secreted brown diffusible pigments in themedium and released large amounts of phenolic metabolitesduring quinoline catabolism. Strain QPS-1 was selected for
further investigations, which are reported below. The organ-ism has been identified as a Pseudomonas sp. on the basis ofthe following characteristics: gram-negative rods (1 ,um by 2to 3 p,m); motile by a single polar flagellum; aerobic, catalaseand oxidase positive; accumulates poly-,-hydroxybutyrate;fluorescent pigments not produced; growth on arginine andbetaine, arginine dihydrolase present; optimum growth tem-perature, 30 to 37°C, with growth at 41°C also; nitratereduced to nitrite; starch and gelatin not hydrolyzed; glu-cose, fructose, sucrose, maltose, lactose, mannitol, sorbitol,ribose, arabinose, and xylose not utilized as growth sub-strates. However, several organic acids, i.e., acetate, suc-cinate, fumarate, malate, lactate, j-hydroxybutyrate, etc.,supported growth. The organism appears to be a strain of P.pseudoalcaligenes (13), except that it reduced nitrates tonitrite.
Quinoline served as the sole source of carbon, nitrogen,and energy for this organism. Growth of the organismoccurred rapidly at 0.01 to 0.015% quinoline; growth oc-curred with a lag at 0.025 to 0.03% quinoline. Higherconcentrations were toxic and completely abolished growth.The organism was rather selective in its choice of heterocy-clic growth substrates, and pyridine, a-, P-, and -y-picolines,isoquinoline, lepidine, and quinaldine were not utilized forgrowth. Among nonnitrogenous aromatic compounds, p-hydroxybenzoate and phenylacetate were utilized, while
VOL. 51, 1986
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
1.61
1000*SYNTHETIC
ilii III
t0OH
I I
1000-
OH
FERMENTATION
r.0IIIn .. IIII .. .50
?00 240 280 320 360 400
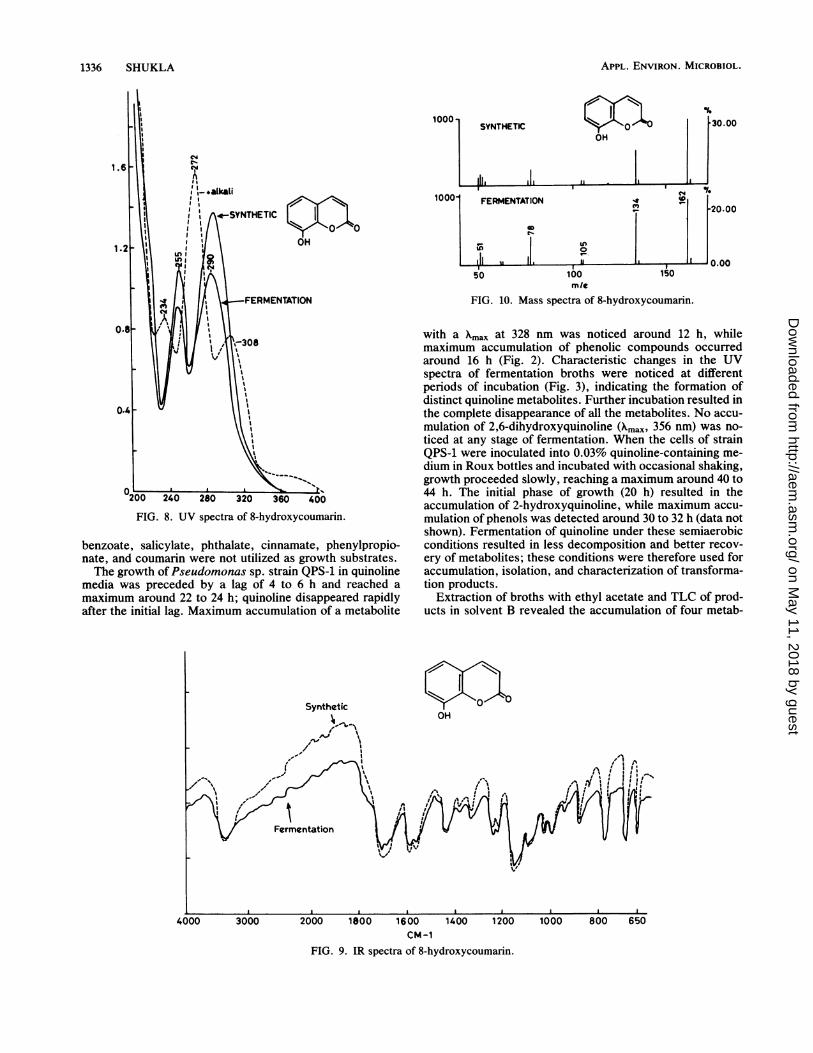
FIG. 8. UV spectra of 8-hydroxycoumarin.
benzoate, salicylate, phthalate, cinnamate, phenylpropio-nate, and coumarin were not utilized as growth substrates.The growth ofPseudomonas sp. strain QPS-1 in quinoline
media was preceded by a lag of 4 to 6 h and reached amaximum around 22 to 24 h; quinoline disappeared rapidlyafter the initial lag. Maximum accumulation of a metabolite
ILO _
100mle
[30.00
20.00
150
FIG. 10. Mass spectra of 8-hydroxycoumarin.
with a Xmax at 328 nm was noticed around 12 h, whilemaximum accumulation of phenolic compounds occurredaround 16 h (Fig. 2). Characteristic changes in the UVspectra of fermentation broths were noticed at differentperiods of incubation (Fig. 3), indicating the formation ofdistinct quinoline metabolites. Further incubation resulted inthe complete disappearance of all the metabolites. No accu-mulation of 2,6-dihydroxyquinoline (Xmax, 356 nm) was no-ticed at any stage of fermentation. When the cells of strainQPS-1 were inoculated into 0.03% quinoline-containing me-dium in Roux bottles and incubated with occasional shaking,growth proceeded slowly, reaching a maximum around 40 to44 h. The initial phase of growth (20 h) resulted in theaccumulation of 2-hydroxyquinoline, while maximum accu-mulation of phenols was detected around 30 to 32 h (data notshown). Fermentation of quinoline under these semiaerobicconditions resulted in less decomposition and better recov-ery of metabolites; these conditions were therefore used foraccumulation, isolation, and characterization of transforma-tion products.
Extraction of broths with ethyl acetate and TLC of prod-ucts in solvent B revealed the accumulation of four metab-
CM-1FIG. 9. IR spectra of 8-hydroxycoumarin.
Ili I .LL-
1336 SHUKLA
#a'".I
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
MICROBIAL TRANSFORMATION OF QUINOLINE 1337
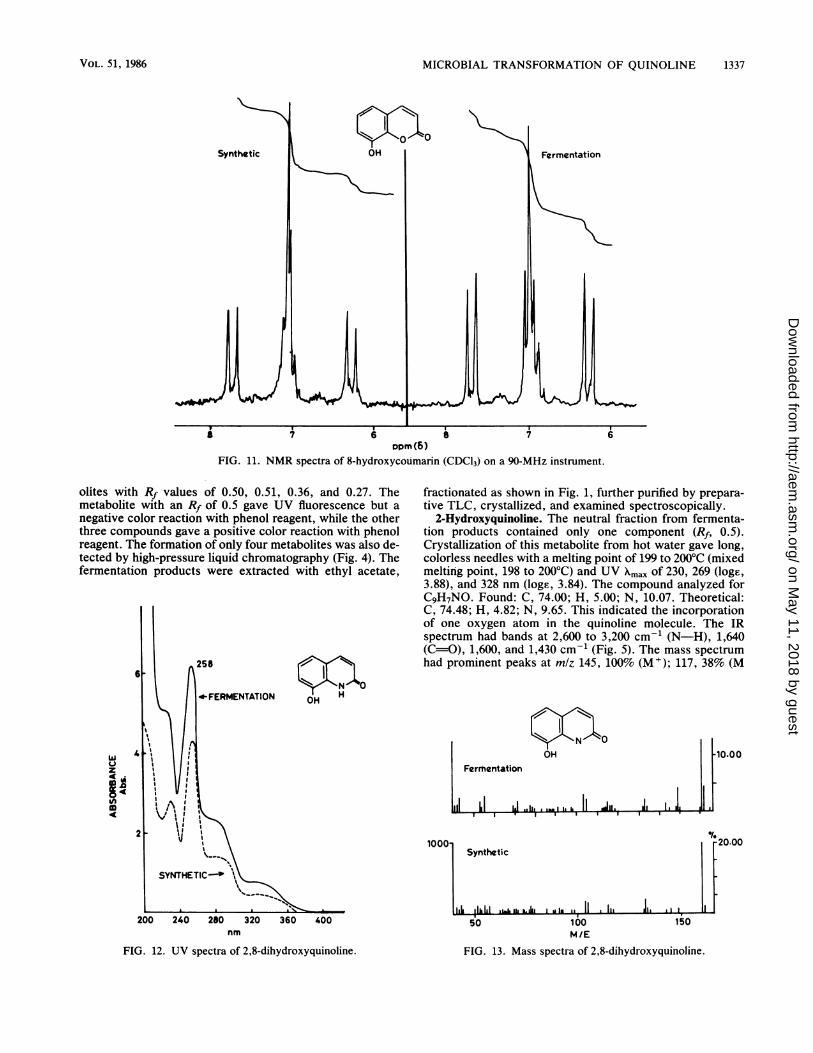
Dopm(6)FIG. 11. NMR spectra of 8-hydroxycoumarin (CDC13) on a 90-MHz instrument.
olites with Rf values of 0.50, 0.51, 0.36, and 0.27. Themetabolite with an Rf of 0.5 gave UV fluorescence but anegative color reaction with phenol reagent, while the otherthree compounds gave a positive color reaction with phenolreagent. The formation of only four metabolites was also de-tected by high-pressure liquid chromatography (Fig. 4). Thefermentation products were extracted with ethyl acetate,
z
In
258
fractionated as shown in Fig. 1, further purified by prepara-tive TLC, crystallized, and examined spectroscopically.
2-Hydroxyquinoline. The neutral fraction from fermenta-tion products contained only one component (Rf, 0.5).Crystallization of this metabolite from hot water gave long,colorless needles with a melting point of 199 to 200°C (mixedmelting point, 198 to 200'C) and UV Xmax of 230, 269 (loge,3.88), and 328 nm (loge, 3.84). The compound analyzed forCqH7NO. Found: C, 74.00; H, 5.00; N, 10.07. Theoretical:C, 74.48; H, 4.82; N, 9.65. This indicated the incorporationof one oxygen atom in the quinoline molecule. The IRspectrum had bands at 2,600 to 3,200 cm-' (N-H), 1,640(C=O), 1,600, and 1,430 cm-' (Fig. 5). The mass spectrumhad prominent peaks at m/z 145, 100% (M+); 117, 38% (M
HNOH
H
<N N 0
OH
Fermentation
1000-
240 280 320 360 400nm
FIG. 12. UV spectra of 2,8-dihydroxyquinoline.
Synthetic
1150
*III150100
M/E
FIG. 13. Mass spectra of 2,8-dihydroxyquinoline.
of-20.00
VOL. 51, 1986
.LI ,1111.1 .1-L I. k.bj I I
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
CM-1
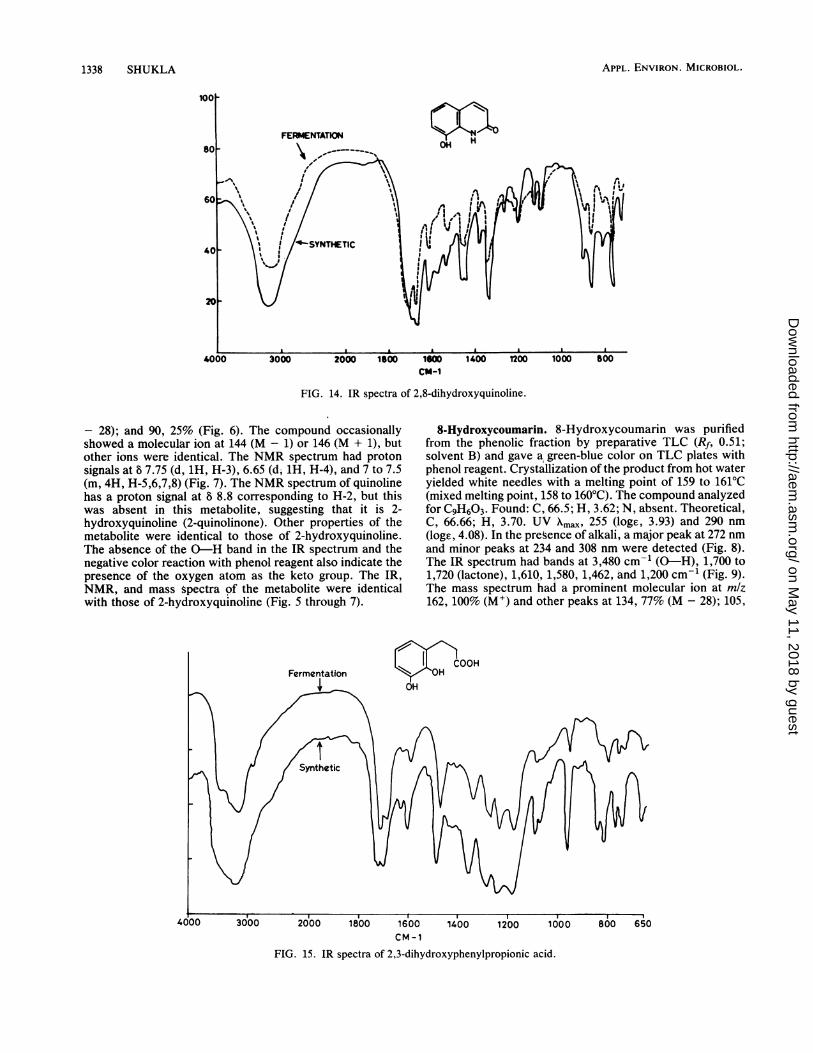
FIG. 14. IR spectra of 2,8-dihydroxyquinoline.
- 28); and 90, 25% (Fig. 6). The compound occasionallyshowed a molecular ion at 144 (M - 1) or 146 (M + 1), butother ions were identical. The NMR spectrum had protonsignals at 8 7.75 (d, 1H, H-3), 6.65 (d, 1H, H-4), and 7 to 7.5(m, 4H, H-5,6,7,8) (Fig. 7). The NMR spectrum of quinolinehas a proton signal at 8 8.8 corresponding to H-2, but thiswas absent in this metabolite, suggesting that it is 2-hydroxyquinoline (2-quinolinone). Other properties of themetabolite were identical to those of 2-hydroxyquinoline.The absence of the 0-H band in the IR spectrum and thenegative color reaction with phenol reagent also indicate thepresence of the oxygen atom as the keto group. The IR,NMR, and mass spectra of the metabolite were identicalwith those of 2-hydroxyquinoline (Fig. 5 through 7).
8-Hydroxycoumarin. 8-Hydroxycoumarin was purifiedfrom the phenolic fraction by preparative TLC (Rf, 0.51;solvent B) and gave a green-blue color on TLC plates withphenol reagent. Crystallization of the product from hot wateryielded white needles with a melting point of 159 to 161°C(mixed melting point, 158 to 160°C). The compound analyzedfor C9H603. Found: C, 66.5; H, 3.62; N, absent. Theoretical,C, 66.66; H, 3.70. UV nmax, 255 (loge, 3.93) and 290 nm(loge, 4.08). In the presence of alkali, a major peak at 272 nmand minor peaks at 234 and 308 nm were detected (Fig. 8).The IR spectrum had bands at 3,480 cm-' (0-H), 1,700 to1,720 (lactone), 1,610, 1,580, 1,462, and 1,200 cm-1 (Fig. 9).The mass spectrum had a prominent molecular ion at mlz162, 100% (M+) and other peaks at 134, 77% (M - 28); 105,
1600CM-1
FIG. 15. IR spectra of 2,3-dihydroxyphenylpropionic acid.
1338 SHUKLA
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
MICROBIAL TRANSFORMATION OF QUINOLINE 1339
N4N4
ICOOHOH
Fermentation
o," 04
N
1.* ImF.- -a-.1 0 Is
Synthetic
,,. I,a,1 ,ii,iwU iL iI, ,4
_ .__ ___ -_50 100 150
M/E200
1000-
-10.00
-10-00
250
1000o
0C1_2
COOHOH
0
Fermentation
I. JI_IsaIZ|*Is s fi *' "
Synthetic
l1.1LjLILL..lAII50 100 150
M/E200 240
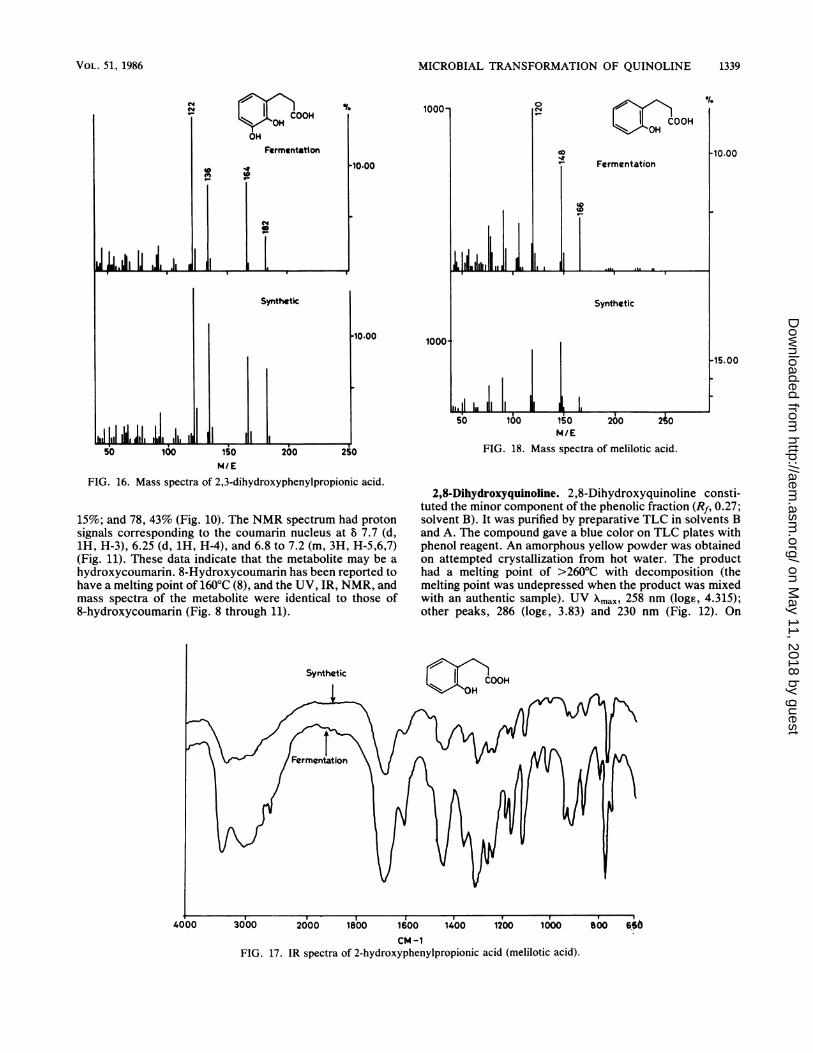
FIG. 18. Mass spectra of melilotic acid.
FIG. 16. Mass spectra of 2,3-dihydroxyphenylpropionic acid.
15%; and 78, 43% (Fig. 10). The NMR spectrum had protonsignals corresponding to the coumarin nucleus at 8 7.7 (d,1H, H-3), 6.25 (d, 1H, H-4), and 6.8 to 7.2 (m, 3H, H-5,6,7)(Fig. 11). These data indicate that the metabolite may be a
hydroxycoumarin. 8-Hydroxycoumarin has been reported tohave a melting point of 160°C (8), and the UV, IR, NMR, andmass spectra of the metabolite were identical to those of8-hydroxycoumarin (Fig. 8 through 11).
2,8-Dihydroxyquinoline. 2,8-Dihydroxyquinoline consti-tuted the minor component of the phenolic fraction (Rf, 0.27;solvent B). It was purified by preparative TLC in solvents Band A. The compound gave a blue color on TLC plates withphenol reagent. An amorphous yellow powder was obtainedon attempted crystallization from hot water. The producthad a melting point of >260'C with decomposition (themelting point was undepressed when the product was mixedwith an authentic sample). UV Xmax, 258 nm (loge, 4.315);other peaks, 286 (loge, 3.83) and 230 nm (Fig. 12). On
CM-1FIG. 17. IR spectra of 2-hydroxyphenylpropionic acid (melilotic acid).
*1.
10.00
-15.00
VOL. 51, 1986
I I I lie III I I d t.LIA li I .11 I
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
80H -C
400
2X3-DHPPA
200 2,8-DHQ
Endo.
30 60 90
Time (min)
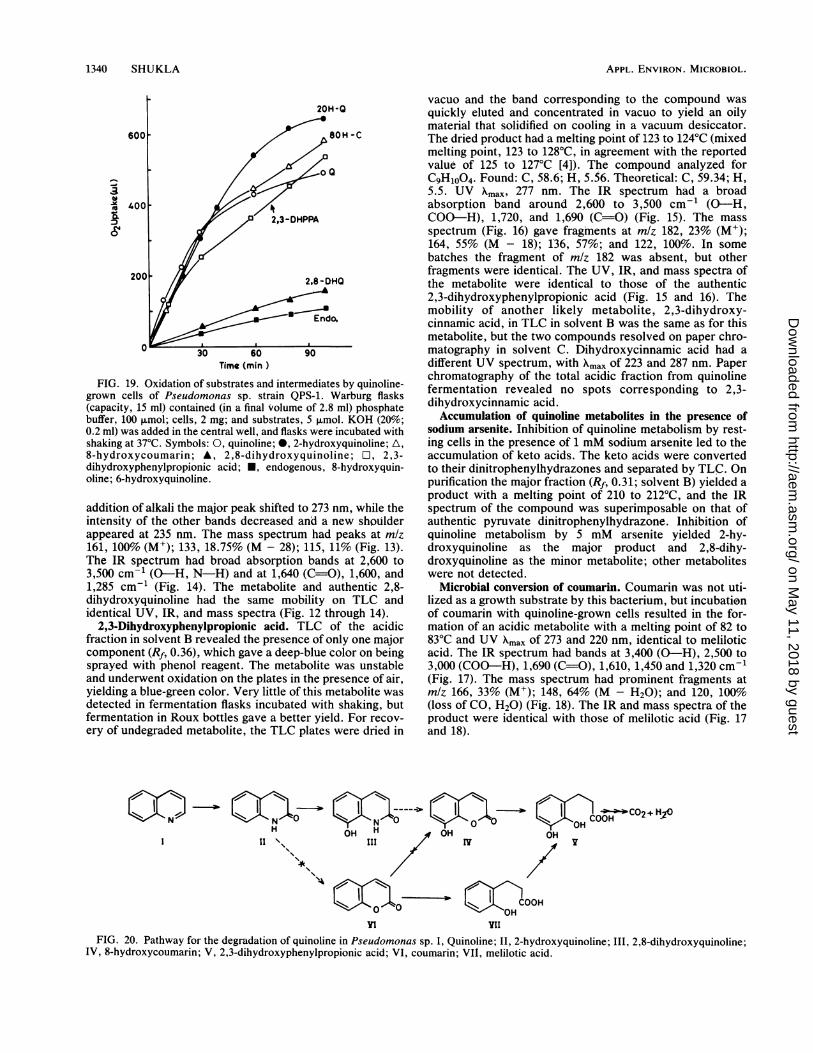
FIG. 19. Oxidation of substrates and intermediates by quinoline-grown cells of Pseudomonas sp. strain QPS-1. Warburg flasks(capacity, 15 ml) contained (in a final volume of 2.8 ml) phosphatebuffer, 100 p.mol; cells, 2 mg; and substrates, 5 ,umol. KOH (20%;0.2 ml) was added in the central well, and flasks were incubated withshaking at 37°C. Symbols: 0, quinoline; *, 2-hydroxyquinoline; A,8-hydroxycoumarin; A, 2,8-dihydroxyquinoline; O1, 2,3-dihydroxyphenylpropionic acid; *, endogenous, 8-hydroxyquin-oline; 6-hydroxyquinoline.
addition of alkali the major peak shifted to 273 nm, while theintensity of the other bands decreased and a new shoulderappeared at 235 nm. The mass spectrum had peaks at mlz161, 100% (M+); 133, 18.75% (M - 28); 115, 11% (Fig. 13).The IR spectrum had broad absorption bands at 2,600 to3,500 cm-' (0-H, N-H) and at 1,640 (C=O), 1,600, and1,285 cm-' (Fig. 14). The metabolite and authentic 2,8-dihydroxyquinoline had the same mobility on TLC andidentical UV, IR, and mass spectra (Fig. 12 through 14).
2,3-Dihydroxyphenylpropionic acid. TLC of the acidicfraction in solvent B revealed the presence of only one majorcomponent (Rf, 0.36), which gave a deep-blue color on beingsprayed with phenol reagent. The metabolite was unstableand underwent oxidation on the plates in the presence of air,yielding a blue-green color. Very little of this metabolite wasdetected in fermentation flasks incubated with shaking, butfermentation in Roux bottles gave a better yield. For recov-ery of undegraded metabolite, the TLC plates were dried in
vacuo and the band corresponding to the compound wasquickly eluted and concentrated in vacuo to yield an oilymaterial that solidified on cooling in a vacuum desiccator.The dried product had a melting point of 123 to 124°C (mixedmelting point, 123 to 128°C, in agreement with the reportedvalue of 125 to 127°C [4]). The compound analyzed forC9H1004. Found: C, 58.6; H, 5.56. Theoretical: C, 59.34; H,5.5. UV Xmax, 277 nm. The IR spectrum had a broadabsorption band around 2,600 to 3,500 cm-' (0-H,COO-H), 1,720, and 1,690 (C=O) (Fig. 15). The massspectrum (Fig. 16) gave fragments at m/z 182, 23% (M+);164, 55% (M - 18); 136, 57%; and 122, 100%. In somebatches the fragment of m/z 182 was absent, but otherfragments were identical. The UV, IR, and mass spectra ofthe metabolite were identical to those of the authentic2,3-dihydroxyphenylpropionic acid (Fig. 15 and 16). Themobility of another likely metabolite, 2,3-dihydroxy-cinnamic acid, in TLC in solvent B was the same as for thismetabolite, but the two compounds resolved on paper chro-matography in solvent C. Dihydroxycinnamic acid had adifferent UV spectrum, with Xmax of 223 and 287 nm. Paperchromatography of the total acidic fraction from quinolinefermentation revealed no spots corresponding to 2,3-dihydroxycinnamic acid.Accumulation of quinoline metabolites in the presence of
sodium arsenite. Inhibition of quinoline metabolism by rest-ing cells in the presence of 1 mM sodium arsenite led to theaccumulation of keto acids. The keto acids were convertedto their dinitrophenylhydrazones and separated by TLC. Onpurification the major fraction (Rf, 0.31; solvent B) yielded aproduct with a melting point of 210 to 212°C, and the IRspectrum of the compound was superimposable on that ofauthentic pyruvate dinitrophenylhydrazone. Inhibition ofquinoline metabolism by 5 mM arsenite yielded 2-hy-droxyquinoline as the major product and 2,8-dihy-droxyquinoline as the minor metabolite; other metaboliteswere not detected.
Microbial conversion of coumarin. Coumarin was not uti-lized as a growth substrate by this bacterium, but incubationof coumarin with quinoline-grown cells resulted in the for-mation of an acidic metabolite with a melting point of 82 to83°C and UV Xmax of 273 and 220 nm, identical to meliloticacid. The IR spectrum had bands at 3,400 (0-H), 2,500 to3,000 (COO-H), 1,690 (C=O), 1,610, 1,450 and 1,320 cm-1(Fig. 17). The mass spectrum had prominent fragments atmlz 166, 33% (M+); 148, 64% (M - H20); and 120, 100%(loss of CO, H20) (Fig. 18). The IR and mass spectra of theproduct were identical with those of melilotic acid (Fig. 17and 18).
H11 \\
'+
NOH H OH
III IY
OH COOHO2+ )
OHv
X0in
COOOHOH
ViI
FIG. 20. Pathway for the degradation of quinoline in Pseudomonas sp. I, Quinoline; II, 2-hydroxyquinoline; III, 2,8-dihydroxyquinoline;IV, 8-hydroxycoumarin; V, 2,3-dihydroxyphenylpropionic acid; VI, coumarin; VII, melilotic acid.
N
1340 SHUKLA
_ _
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from

MICROBIAL TRANSFORMATION OF QUINOLINE 1341
Oxidation of substrates and intermediates. Quinoline wasrapidly oxidized by quinoline-adapted cells of Pseudomonassp., but succinate-grown cells failed to oxidize quinoline.2-Hydroxyquinoline, 8-hydroxycoumarin, and 2,3-dihy-droxyphenylpropionic acid were also rapidly oxidized byquinoline-adapted cells; 2,8-dihydroxyquinoline was oxi-dized very slowly (Fig. 19). 8-Hydroxyquinoline and 6-hydroxyquinoline were not oxidized.
Incubation of quinoline, 2-hydroxyquinoline, and 8-hydroxycoumarin with quinoline-grown cells resulted in thecomplete disappearance of these substrates; 2,8-dihy-droxyquinoline is insoluble and yielded a suspension whenadded to the incubation mixtures. Incomplete recovery(75%) of 2,8-dihydroxyquinoline was obtained at the end ofthe incubation.
DISCUSSIONBenzene derivatives are generally degraded via introduc-
tion of two hydroxyl groups either ortho or para to eachother, followed by oxidative cleavage of the resulting diol toyield cis,cis-muconic acid or muconic semialdehyde deriva-tives as the intermediates (10, 16). Hydroxypyridines andpyridine carboxylic acid are also degraded via di- ortrihydroxy pyridines as intermediates (32). Pyridine, a-picoline, N-methylisonicotinate, and isonicotinate use alter-native pathways in which the ring is probably reduced to thedi- or tetrahydropyridine state followed by oxidative cleav-age to acylic intermediates which, on hydrolysis, generatesuccinic semialdehyde as a key intermediate (30, 32, 34, 41);glutaric semialdehyde has been detected as the intermediatein Nocardia sp. strain Z1, suggesting that hydrolytic cleav-age of the partially reduced pyridine ring has occurred (41).The pyridine ring differs from the benzene ring in having alone pair of electrons on the nitrogen, which causes the ringto assume a positive charge, making it less susceptible toelectrophilic attack (3). Pyridine is thus used as the solventfor the oxidation of other organic compounds with CrO3 (26).The pyridine ring of the quinoline molecule is also lesssusceptible to oxidative cleavage (25). The quinoline ring inkynurenic acid has been shown to be degraded via the7,8-diol, suggesting the preferential cleavage of the benzenering (11, 38). However, the mechanism of cleavage of thering in quinoline per se in bacteria is not known. Grant andAl-Najiar (17) isolated and characterized 2-hydroxyquinolineas the first metabolic product of quinoline in Moraxella sp.They implicated 2,6-dihydroxyquinoline as an intermediateon the basis of its oxidation by quinoline-adapted cells.Degradation of kynurine (4-hydroxyquinoline) in a Pseu-domonas sp. was reported to proceed via 4,7,8-tri-hydroxyquinoline, and a putative pink cleavage product waspostulated to arise from meta cleavage of the triol (2).Degradation of quinoline by a Pseudomonas sp. (QPS-2) anda Nocardia sp. proceed via 2-hydroxyquinoline and 2,6-dihydroxyquinoline (31). However the details of this path-way are still not known.The results presented in this paper indicate the existence
of an alternative pathway for quinoline catabolism in mi-crobes (Fig. 20); 2-hydroxyquinoline is the first intermediateof this pathway also. However, 2,8-dihydroxyquinoline,8-hydroxycoumarin, and 2,3-dihydroxyphenylpropionic acidhave been characterized as the other intermediates of thispathway. 2-Hydroxyquinoline and 2,8-dihydroxyquinolineaccumulated as the exclusive products when quinoline wasmetabolized in the presence of 5 mM arsenite. Quinoline-grown cells rapidly oxidized 2-hydroxyquinoline, 8-hy-droxycoumarin, and 2,3-dihydroxyphenylpropionic acid,
supporting their role as intermediates; 2,8-dihydroxy-quinoline was oxidized very slowly with overall consump-tion of only 0.5 mol of oxygen per mol of substrate. It alsodid not disappear significantly when incubated with restingcells for 24 h or more, suggesting that it may not be a trueintermediate; no formation of 8-hydroxycoumarin was de-tected under these incubation conditions. 2,8-Dihydroxy-quinoline is, however, highly insoluble and exists in suspen-sion when added to the cells. The solubility and permeabilityof the compound may be limiting factor(s) in demonstratingits role as an intermediate of quinoline catabolism.8-Hydroxycoumarin is the intermediate that has been
isolated and characterized rigorously by spectroscopic meth-ods as well as comparison with chemically synthesizedsubstrate. A metabolite with the spectroscopic characteris-tics of 8-hydroxycoumarin also accumulated in broths duringfermentation (16 h) (Fig. 3). It can be argued that the trueintermediate of quinoline catabolism may be cis-3,4-dihydroxycinnamic acid which may rapidly cyclize underacidic conditions of processing of broths and extraction withethyl acetate. However, extraction of broths without acidi-fication and concentration without application of heat alsoyielded 8-hydroxycoumarin. Incubation of 8-hydroxy-coumarin with resting cells led to its complete disappear-ance, while the parent compound, coumarin, was not de-graded beyond melilotic acid. The reactions of quinolinemetabolism seem to be very specific, and hydroxylation ofcoumarin to 8-hydroxyderivative does not occur in thisbacterium. Small quantities of 2,3-dihydroxyphenyl-propionic acid were detected during aerobic fermentation ofquinoline, but larger amounts accumulated in semi-aerobicfermentation in Roux bottles. This compound was oxidizedby quinoline-adapted cells. Closely related 2,3-dihy-droxycinnamic acid was not detected spectroscopically andwas not oxidized by the quinoline-adapted cells ruling itsparticipation. The present studies therefore establish 2-hydroxyquinoline, 8-hydroxycoumarin, and 2,3-dihydroxy-phenylpropionic acid as intermediates of quinoline catabo-lism in this organism.
LITERATURE CITED1. Albert, A. 1959. Heterocyclic chemistry, p. 105. The Athlone
Press of the University of London, London.2. Al-Najjar, T. R., R. J. Grout, and D. J. W. Grant. 1976.
Degradation of 4-hydroxyquinoline (kynurine) by a soilPseudomonad. Microbios Lett. 1:157-163.
3. Barnes, R. A. 1960. Properties and reactions of pyridine and itshydrogenated derivatives, p. 1-97. In E. Klingsberg (ed.),Pyridine and its derivatives, part I. Interscience Publishers Inc.,New York.
4. Blakley, E. R., and F. J. Simpson. 1964. The microbial metab-olism of cinnamic acid. Can. J. Microbiol. 10:175-185.
5. Bohme, H., and T. Severin. 1957. Optical studies on coumarins.II. Ultraviolet absorption of monohydroxycoumarins and deriv-atives. Arch. Pharm. (Weinheim) 290:405-412.
6. Bohonos, J., T. W. Chow, and R. J. Spanggord. 1977. Someobservations on biodegradation of pollutants in aquatic systems.Jpn. J. Antibiot. 30(Suppl):275-285.
7. Chichibabin, A. E. 1924. Preparation of hydroxyderivatives ofpyridine, quinoline and their homologs by a new method. J.Russ. Phys. Chem. Soc. 55:7-18.
8. Cingolani, E. 1954. Benzopyrones, coumarins derived fromo-vanillin. Gazz. Chim. Ital. 84:843-852.
9. Conn, H. J., J. W. Bartholomew, and M. W. Jennison. 1957.Staining methods, p. 10-36. In H. J. Conn and M. J. Pelczar, Jr.(ed.), Manual of microbiological methods. Society of AmericanMicrobiologists. McGraw Hill Book Co., New York.
10. Dagley, S. 1971. Catabolism of aromatic compounds by micro-
VOL. 51, 1986
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
organism. Adv. Microb. Physiol. 6:1-46.11. Dagley, S., and P. A. Johnson. 1963. Microbial oxidation of
kynurenic, xanthurenic and picolinic acids. Biochim. Biophys.Acta 78:577-587.
12. Diamant, J. 1895. On the direct introduction of hydroxyl groupsin oxyquinoline. Monatsh. Chem. 16:760-762.
13. Doudoroff, M., and J. J. Palleroni. 1974. Pseudomonas, p.217-243. In. R. E. Buchanan and N. E. Gibbons (ed.), Bergey'smanual of determinative bacteriology, 8th ed. The Williams &Wilkins Co., Baltimore.
14. Friedman, T. E., and G. E. Haugen. 1943. Pyruvic acid. II. Thedetermination of ketoacids in blood and urine. J. Biol. Chem.147:415-422.
15. Funchess, M. J. 1917. The nitrification of pyridine, quinoline,guanidine carbonate etc. in soils, p. 65-82. In Alabama Agricul-tural Experiment Station Bulletin no. 196 (Technical Bulletinno. 3).
16. Gibson, D. T., and V. Subramanian. 1984. Microbial degrada-tion of aromatic hydrocarbons, p. 181-252. In D. T. Gibson(ed.), Microbial degradation of aromatic compounds. MarcelDekker, Inc., New York.
17. Grant, D. J. W., and T. R. Al-Najjar. 1976. Degradation ofquinoline by a soil bacterium. Microbios 15:177-189.
18. Grunden, M. J. 1979. Quinoline alkaloids related to anthranilicacid, p. 105-198. In R. H. F. Manske and R. Rodrigo (ed.), Thealkaloids, vol. 17. Academic Press, Inc., New York.
19. Gupta, R. C., and 0. P. Shukla. 1979. Microbial transformationof isonicotinic acid hydrazide and isonicotinic acid by Sarcinasp. J. Biosci. 1:223-234.
20. Hargreaves, G. W. 1936. Quinone formation in the thalleoquinreaction: a new preparation of quinoline-o-quinone. J. Am.Pharm. Assoc. 25:975-976.
21. Katritzky, A. R., and J. N. Lagowski. 1960. Heterocycliccompounds, p. 29. John Wiley & Sons, Inc., New York.
22. Kucher, R. V., A. Turovskii, N. V. Dzumedzei, and A. G.Shevchenko. 1980. Microbiological transformation of quinolineby Pseudomonas putida bacteria. Mikrobiol. Zh. 42:284-287.
23. Leifson, E. 1960. Atlas of bacterial flagellation, p. 1-7. Aca-demic Press, Inc., New York.
24. Levy, C. C., and G. D. Weinstein. 1964. The metabolism ofcoumarin by a microorganism. II. The reduction of o-coumaricacid to melilotic acid. Biochemistry 3:1944-1947.
25. Paquette, L. A. 1968. Principles of modem heterocyclic chem-istry, p. 222 and 273. W. A. Benjamin, Inc., Reading, Mass.
26. Poos, G. I., G. E. Arth, R. E. Beyler, and L. E. Sarett. 1953.Approaches to total synthesis of adrenal steroids. V. 4b-methyl7-ethylenedioxy-1,2,3,4,4a,4b,5,6,7,8,10,10a-dodecahydro-phenanthrene-4-O1-1-one and related tricyclic derivatives. J.
Am. Chem. Soc. 75:422-429.27. Reinhardt, C. F., and M. R. Britteli. 1981. Heterocyclic and
miscellaneous nitrogen compounds, p. 2761-2763. In G. D.Clayton and F. E. Clayton (ed.), Patty's industrial hygiene andtoxicology, vol. IIA. John Wiley & Sons, Inc., New York.
28. Robbins, W. J. 1917. The cause of disappearance of coumarin,vanillin, pyridine and quinoline in the soil, p. 49-64. In AlabamaAgricultural Experiment Station Bulletin no. 195 (TechnicalBulletin no. 2).
29. Ronkainen, P. 1967. Chromatographic identification of carbonylcompounds. VI. TLC resolution of mixtures of keto acid2,4-dinitrophenylhydrazones. J. Chromatogr. 28:263-276.
30. Shukla, 0. P. 1974. Microbial decomposition of a-picoline.Indian J. Biochem. Biophys. 11:192-200.
31. Shukla, 0. P. 1984. Microbial transformation and biodegrada-tion of quinoline and isoquinoline, p. 323. Seventh InternationalBiotechnology Symposium, New Delhi, India.
32. Shukla, 0. P. 1984. Microbial transformation of pyridine deriv-atives. J. Sci. Ind. Res. 43:98-116.
33. Shukla, 0. P., and S. M. Kaul. 1973. Microbial transformationof a-picolinate. Indian J. Biochem. Biophys. 10:176-178.
34. Shukla, 0. P., and S. M. Kaul. 1974. A constitutive pyridine-degrading system in Corynebacterium sp. Indian J. Biochem.Biophys. 11:201-207.
35. Shukla, 0. P., and S. M. Kaul. 1981. Microbial metabolism ofquinoline and quinoline derivatives. Indian J. Biochem. Bio-phys. 18(Suppl):88.
36. Spies, J. R. 1957. Colorimetric procedure for amino acids.Methods Enzymol. 3:467-468.
37. Stanier, R. Y., N. J. Palleroni, and M. Doudoroff. 1966. Theaerobic pseudomonads: a taxonomic study. J. Gen. Microbiol.43:159-271.
38. Taniuchi, H., and 0. Hayaishi. 1963. Studies on the metabolismof kynurenic acid. III. Enzymatic formation of 7,8-dihy-droxykynurenic acid from kynurenic acid. J. Biol. Chem.238:283-293.
39. Umbreit, W. W., R. H. Burris, and J. F. Stauffer. 1964.Manometric techniques, p. 43. Burgess Publishing Co., Minne-apolis.
40. Vogel, A. I. 1966. A textbook of practical organic chemistry, p.768-769. John Wiley & Sons, Inc., New York.
41. Watson, G. K., and R. B. Cain. 1975. Microbial metabolism ofpyridine ring: metabolic pathways for pyridine degradation insoil bacteria. Biochem. J. 146:157-172.
42. Williams, R. T. 1959. Detoxication mechanisms, 2nd ed., p.639-645. Chapman and Hall, Ltd., London.
43. Wolff, M. E. 1979. Burger's medicinal chemistry, 4th ed. JohnWiley & Sons, Inc., New York.
1342 SHUKLA
on May 11, 2018 by guest
http://aem.asm
.org/D
ownloaded from














![Synthesis of highly functionalized benzo[h]quinoline and ...shodhganga.inflibnet.ac.in/bitstream/10603/39020/17/17...quinoline ) (16 ) and tetracyclic quinoline (3-(epimin omethano)](https://img.pdfslide.us/doc/110x75/606a70077d4f6141007ad728/synthesis-of-highly-functionalized-benzohquinoline-and-quinoline-16.jpg)














