Embed Size (px)
Citation preview

REVIEW
Carl Anderson á Francis L.W. Ratnieks
Worker allocation in insect societies: coordination of nectar foragersand nectar receivers in honey bee (Apis mellifera) colonies
Received: 15 December 1998 / Received in revised form: 6 March 1999 /Accepted: 12 March 1999
Abstract Nectar collection in the honey-bee is parti-tioned. Foragers collect nectar and take it to the nest,where they transfer it to receiver bees who then store itin cells. Because nectar is a ¯uctuating and unpredictableresource, changes in worker allocation are required tobalance the work capacities of foragers and receivers sothat the resource is exploited e�ciently. Honey beecolonies use a complex system of signals and otherfeedback mechanisms to coordinate the relative andtotal work capacities of the two groups of workers in-volved. We present a functional evaluation of each ofthe component mechanisms used by honey bees ± waggledance, tremble dance, stop signal, shaking signal andabandonment ± and analyse how their interplay leads togroup-level regulation. We contrast the actual regula-tory system of the honey bee with theory. The trembledance conforms to predicted best use of information,where the group in excess applies negative feedback toitself and positive feedback to the group in shortage, butthis is not true of the waggle dance. Reasons for this andother discrepancies are discussed. We also suggest rea-sons why honey bees use a combination of recruitmentplus abandonment and not switching between subtasks,which is another mechanism for balancing the workcapacities of foragers and receivers. We propose that thewaggle and tremble dances are the primary regulationmechanisms, and that the stop and shaking signals aresecondary mechanisms, which ®ne-tune the system.
Fine-tuning is needed because of the inherent unreli-ability of the cues, queueing delays, which foragers useto make recruitment decisions.
Key words Honey bee á Signals á Nectar foragers áReceiver bees á Worker allocation á Waggle dance áTremble dance á Shaking signal á Stop signal
Introduction
Insect societies are complex adaptive systems (Seeley1985, 1995, 1997; Bonabeau, in press) in which the in-dividuals form a highly coordinated and integrated unit.With little or no centralised control, many workers areable to work together and collectively tackle tasks farbeyond the abilities of any one individual (Bonabeauet al. 1997). The honey-bee, Apis mellifera, is the best-studied social insect and is an excellent model system forunderstanding the emergence of adaptive group-levelbehaviour. Much is known about honey bee group-levelcoordinated activities and the proximate mechanismsused by workers to achieve this. The key factor thatbinds individuals together as an adaptive unit is infor-mation transfer between individuals in the form of cuesand signals. A signal is taken to be a deliberate act ofcommunication that has been shaped by natural selec-tion (Lloyd 1983). An example of a signal is the honeybee waggle dance (von Frisch 1967). A cue is a structureor behaviour which conveys information, but only in-cidentally, and which has not been shaped by naturalselection. For instance, the time spent searching for anectar receiver or the carbon dioxide level in the nest(Seeley 1995, 1998) are both cues.
At least 17 signals and twice as many cues have beenidenti®ed in the honey-bee (Seeley 1998). Most honeybee signals are involved in either colony reproduction orforaging which are also two areas of colony life vital tocolony survival and likely to be subject to strong selec-tion. Four, possibly ®ve, signals, as well as numerous
Behav Ecol Sociobiol (1999) 46: 73±81 Ó Springer-Verlag 1999
C. Anderson1
Department of Genetics and EcologyUniversity of Aarhus, DK-8000 Aarhus C, Denmark
F.L.W. Ratnieks (&)Department of Animal and Plant SciencesUniversity of She�eld, She�eld, S10 2TN, UKe-mail: f.ratnieks@she�eld.ac.ukTel.: +44-114-2220070, Fax: +44-114-2760159
Present address:1Department of Zoology, Duke UniversityDurham, NC 27708-0325, USAe-mail: [email protected]

cues, regulate the number of workers involved in thenectar-harvesting process which involves both collectionfrom ¯owers and storage in cells. In the honey bee, nectarforaging is a task partitioned into two subtasks in whichone group of workers, the foragers, collect the materialand take it to the nest. However, instead of storing thenectar themselves, they transfer it to receiver bees whothen store it in cells. [Task partitioning is reviewed inJeanne (1986a) and Ratnieks and Anderson (1999).]Nectar availability and ease of collection can ¯uctuaterapidly and unpredictably, creating a need to alter thetotal foraging e�ort and balance the relative work ca-pacities of the forager and receiver groups. When thesework capacities are not matched, the workers are sub-optimally allocated and the nectar processing rate is lessthan it could be given the total number of workers in-volved (Anderson and Ratnieks, in press a, in press b;Ratnieks and Anderson, in press b). Honey bees havesophisticated mechanisms to regulate the number ofworkers in these two groups, both to recruit additionalworkers to exploit the ¯uctuating and ephemeral nectarresources (Seeley 1995) and to balance the relative workcapacities of foragers and receivers. These changes inworker allocation are mediated through four or ®vesignals: ``waggle dance'', ``tremble dance'', ``stop signal'',``shaking signal'' and, possibly, ``worker piping''.
The main focus of this paper is the interaction ofthese signals, together with other non-signalling feed-back mechanisms such as patch abandonment, in theregulation of foraging. The following speci®c questionsare addressed. How exactly do these signals act andcause feedback on each other in order to integrate theforaging population into a ``group-level adaptive unit''(Seeley 1997)? What is the interplay between waggledancing and tremble dancing? Why do honey bees needstop and shaking signals? Additional questions focus onwhy the honey bee is organised the way it is rather thanin another manner. For example, why do honey-bees notneed a patch abandonment signal? How best shouldindividuals use signalling information? When signals areproduced, who should produce them: the group in excessor the group in shortage? Why do honey bees use re-cruitment and abandonment to balance work capacitiesrather than switching between subtasks?
Regulation mechanisms in nectar foraging
This section provides background information on the®ve signals plus one other feedback mechanism, aban-donment, used in regulating foraging. The emitter of thesignal is referred to as the ``sender'' and the intendedaudience or target of the signal as the ``recipient''. Be-cause full details of each signal are published elsewhere,we focus on the signal ``message'', i.e. the intended in-formation transmitted by the sender, its ``meaning'', i.e.the interpretation by the recipient, and its e�ects onworker allocation and activity.
Waggle dance
Probably the most well-known honey bee signal, andwhat is primarily referred to by the term ``dance lan-guage'', the waggle dance was deciphered by the pio-neering work of von Frisch (1967). It is a signal sent bynectar foragers and is directed at unemployed foragers(Seeley 1995), and possibly workers involved in othertasks [Fig. 1a(i)]. Waggle dances are also used to conveyinformation about the location of water, pollen andpotential nest sites (Seeley 1995). Waggle dances arenormally performed by a returning nectar forager whohas experienced a relatively short delay, typically lessthan 20 s (Seeley 1992, 1995, 1997), while searching for areceiver bee to whom she can transfer her nectar. Onecomponent of the dance, the direction of the waggle run,tells the dance followers the direction of the food sourcein relation to the azimuth of the sun. A second com-ponent, the duration of the waggle run, conveys thedistance (von Frisch 1967). Its message is: ``I have founda good food source worthy of greater exploitation. It islocated in direction X and distance Y '', while itsmeaning is: ``I should help exploit this patch''. Thewaggle dance, therefore, exerts positive feedback on thenumber of active foragers [Fig. 1a(ii); Seeley 1995].
Tremble dance
Returning foragers may also perform the tremble dance[Fig. 1b(i)]. In contrast to the waggle dance, this signal issent when a returning bee experiences a relatively longsearch delay, typically more than 50 s (Seeley 1992, 1995,1997). Its message is apparently: ``I have visited a richnectar source worthy of greater exploitation, but alreadywe have more nectar coming in than we can handle''(Seeley 1992), and has multiple meanings. For theworkers in the nest, its meaning is: ``I should switch tothe task of nectar processing'' (Seeley 1992), which has apositive feedback e�ect on the number of receivers.However, its meaning for other foragers is: ``I shouldrefrain from recruiting additional foragers to my nectarsource'' (Seeley 1992). Kirchner (1993) demonstratedthat playback of arti®cial tremble dance sounds reducedwaggle dance duration, or in some cases caused waggledancers to stop dancing immediately, and reduced therecruitment success of the dances. Thus, the trembledance can have a negative, or at least non-positive, e�ecton foragers in that it reduces forager recruitment (Seeley1992; Kirchner 1993 and references therein). But it has apositive e�ect on the number of receivers, recruitingmore workers to receiving (Seeley et al. 1996).
Stop signal
Originally termed a begging signal (Esch 1964), the stopsignal (Nieh 1993) comprises the sender emitting anaudible piping sound while head-butting the recipient,
74

who then stops moving or stops dancing. Althoughproduced by tremble dancers, dance followers andwaggle dancers, most stop signals (85%) are producedby tremble dancers ± with almost all of the remaining15% of stop signals sent by dance followers ± and are
mainly directed at waggle dancers, who are 44% of allrecipients [Fig. 1c(i); Nieh 1993]. Recipient waggledancers tend to stop waggle dancing and leave the dance¯oor. Thus, the stop signal is mainly sent by trembledancers, directed at waggle dancers, and inhibits addi-tional forager recruitment [Fig. 1c(ii)].
Shaking signal
The shaking signal, also known as the dorso-ventralabdominal vibration (Milum 1955), is sent by foragers toa diverse set of recipients, i.e. workers involved in vari-ous tasks throughout the nest [Fig. 1d(i)]. Like the
Fig. 1 Information ¯ow and feedbacks for various regulatory signalsand mechanisms used in the nectar-foraging process in the honey-bee:waggle dance (a), tremble dance (b), stop signal (c), shaking signal (d),abandonment (e), a±e combined (f). (i) Information ¯ow from sendersto recipients (dashed arrows) and the ¯ow of workers that respond tothe signals (solid arrows). (ii) Feedbacks in task allocation that resultfrom the signals (+ excitatory, ) inhibitory). c Dance followers aretaken to be in the group of unemployed foragers. d Waggle dancerssometimes make shaking signals as well (Seeley et al. 1998)
75

stop-signal this is a ``one-to-one'' signal in which thesender physically shakes a single recipient (Allen 1959).Recipients tend to crawl onto the dance area (Schneideret al. 1986) or towards the entrance (Nieh 1998) wherethey are more likely to come into contact with othersignals, typically waggle dances, and may therefore berecruited as additional foragers. Evidence (Seeley 1995;Nieh 1998; Seeley et al. 1998) suggests that shakingsignals are used in conjunction with waggle dances toincrease the number of active foragers [Fig. 1d(ii)] whennectar availability starts to increase after a nectar dearthand thus additional foragers are required. The messageof the shaking signal appears to be ``prepare for greateractivity'' (Seeley et al. 1998).
Abandonment
Although not a signal, abandonment of a ¯ower patchby foragers is an important feedback mechanism inforaging regulation. In the honey bee, foragers work asmall patch or ¯owers (Ribbands 1953) and although thepro®tabilities of one forager's patch may decline or be-come unrewarding, this does not necessarily mean thatother patches are also unrewarding. As foragers are ef-fectively independent while outside the hive, a localmechanism enabling them to make their own decisionsto abandon a patch is required. In doing so they areavailable for recruitment to better patches (Fig. 1e).
Worker piping
The above four signals plus abandonment may not bethe only mechanisms used by honey bees. The possibilitythat there are more to discover can be appreciated byrealising that 5 of the 17 known signals have only beenelucidated within the last decade (Seeley 1998). Onecandidate signal is worker piping (Armbruster 1922).Little is known about this signal. It is almost certainly anaerial and substrate-mediated broadcast signal and ap-pears to be sent by workers involved in foraging, col-lection and use of water, and collection of pollen andperhaps nectar, but whose recipients, messages, andmeanings are unknown (Pratt et al. 1996). Pratt et al.(1996) report that in three cases in which the sender'sprevious activity was known, one was a pollen collector,one was a water collector and one was a water receiver.However, the possibility that it is also a signal sent bynectar foragers cannot be discounted, especially as theonly case in which more than one piper was observedwas during a day of high nectar intake (Pratt et al.1996).
A complex system of feedbacks
Figure 1f(ii) combines the feedbacks from the signalsand mechanisms described in the previous section.
Clearly, regulation of nectar foraging is complex.However, various feedback systems, some of which aremuch simpler than that used by the honey bee, could beformulated which produce the same global regulatoryproperties (e.g. Fig. 2). Why then do honey bees utilisetheir particular and complex system?
The interplay between waggle dancingand tremble dancing
The relationship between waggle and tremble dancing isrevealing. In a two-stage partitioned task (i.e. with twosubtasks) with direct transfer, such as nectar collectionand storage in the honey bee, queueing delays occur(Anderson and Ratnieks, in press a, in press b; Ratnieksand Anderson, in press b). These delays can increasetremendously when the work capacities of the twogroups are unmatched. Ratnieks and Anderson (in pressa) have shown that if queueing delays are used as anindicator of the relative work capacities of the twogroups, then the group in excess (that is those that ex-perience longer mean queueing delays) possess better-quality information than the group in shortage (whoexperience shorter delays). From this we would expectthe group in excess to apply negative feedback tothemselves and positive feedback to the group inshortage. This is true in the case of the tremble dance inwhich foragers indirectly apply negative feedback to therecruitment of foragers by inhibiting waggle dancing(Seeley 1992, 1995; Kirchner 1993 and references there-in; Nieh 1993) and positive feedback on the number ofreceivers. However, foragers also perform the waggledance (Seeley 1995) which recruits unemployed foragers.This is positive feedback from the group in shortage,and is not as expected from best use of the informationavailable from queueing delays (Ratnieks and Anderson1999).
Fig. 2 One of the simplest sets of feedbacks that would produce thesame global properties as the system used in the honey bee [Fig. 1f(ii)].In this example, the foragers perform a forager recruitment signal (i)and receivers perform a receiver recruitment signal (iii), and bothgroups may abandon their subtask (ii), in contrast to the situationfound in honey bees in which both forager and receiver recruitment ismediated solely through foragers. As in ®gure 1, dashed arrowsindicate the ¯ow of information from senders to recipients, and solidarrows, the ¯ow of workers that respond to the signals
76

When foragers are in shortage they recruit additionalforagers through the waggle dance. A priori, this is anobvious feedback solution: when in need recruit othersyourself. On this basis we might expect receivers to re-cruit additional receivers. However, this is not the case.It is foragers, and not receivers, who recruit the addi-tional receivers. Why do foragers have a greater regu-latory role than receivers, and in particular, why doforagers recruit new receivers? We suggest the reason isthat foragers have essential additional information notavailable to receivers such as the location of ¯owers andthe rate at which nectar can be collected. The only in-formation about the forage patch available to receiversis the nectar concentration, but even this informationmay be less than accurate because foragers may evapo-rate nectar by regurgitation whilst foraging (Heinrich1979). Honey bee colonies would ®nd it almost impos-sible to track the constantly changing locations of re-warding patches of ¯owers using shared information ifrecruitment of foragers was via receivers and the limitedinformation at their disposal.
Why do honey bees need a stop signal?
Due to ¯uctuations in nectar availability, the work ca-pacity of one of the two groups, foragers or receivers,will often exceed that of the other. To redress this im-balance, one would expect that only one type of workershould be recruited at one time. That is, the bees in acolony should either perform waggle dances or trembledances (or neither) but not both. However, these twodances can occur together (Nieh 1993; Seeley 1995). Oneprobable reason for this are the time lags that occurbetween information acquisition and dancing so thatinformation becomes out of date. A second reason is theunreliability inherent in the cues, search delays, whichforagers use to make their recruitment decisions (Rat-nieks and Anderson 1999, in press a). Unsurprisingly,therefore, there is a regulatory mechanism, the ``stopsignal'' (Nieh 1993), which directly inhibits forager re-cruitment. Most stop signals (85%; Nieh 1993) are sentby tremble dancers and directed at waggle dancers withthe e�ect that 16% of the waggle dancers immediatelystop dancing and leave the nest, presumably to continueforaging but without recruiting more foragers. The stopsignal is a negative feedback signal on forager recruit-ment, conveying the message ``receivers are currentlyoverloaded: stop recruiting more foragers''. Dance fol-lowers give most of the remaining signals. Given thatfollowers move around the dance area, on which bothdances occur (Seeley 1995), they are in a good positionto determine whether waggle and tremble dances arebeing made concurrently. Interestingly, Nieh (1993)found that tremble dancers, who were the largest groupto send stop signals, directed 34% of their signals towaggle dancers and 29% to tremble dancers. However,dance followers, the only other signi®cant group to sendstop signals, directed all of their signals to waggle
dancers. Is there some information available to trembledancers (active foragers) but not to dance followers(inactive workers) that can explain this di�erence in thetarget(s) of the signal sent by these two groups of stopsignal emitters?
Why do honey bees need the shaking signal?
The shaking signal appears to be emitted when nectaravailability increases after a period of low nectar forageractivity. In such a situation, most foragers will be un-employed and inactive. Clearly, it is in the interest of thecolony to recruit additional foragers rapidly to exploitthis new resource before conditions deteriorate. How-ever, there will not be many foragers to do the recruit-ing, because there is little forager activity. Also there willnot be many unemployed foragers in the dance area tobe recruited. Therefore, recruitment through waggledances alone is likely to be slow, leading to the need foran additional signal to complement forager recruitment:the shaking signal. After having been shaken, over halfof the recipients moved to the dance area where theycould be recruited to foraging (Schneider et al. 1986).There appears to be a strong link between the shakingsignal and the waggle dance. Seeley et al. (1998) foundthat bees that send shaking signals appear to follow aparticular pattern of behaviour. These workers, whichare generally the ®rst workers to leave the nest and in-spect potential forage patches, emit shaking signals atthe end of their early trips. On the next few trips theyemit a mix of both shaking signals and waggle dancesand then on subsequent trips tend to send only waggledances (see Fig. 3±6 of Seeley et al. 1998). Thus, theshaking signal, which conveys a general message thatforagers are required, appears to be linked to the waggledance which provides speci®c information as to whereforagers should work.
Why do honey bees not use a patchabandonment signal?
Why is there no known signal indicating poor patches inhoney bee foraging regulation? That is, why is aban-donment an individual decision made without any in-formation from others rather than the result of a speci®cpatch abandonment signal? Although the absence ofevidence for a patch abandonment signal does not meanthat no such signal exists (5 of the 17 known honey beesignals have only been elucidated within the last10 years), there are logical reasons why a patch aban-donment signal is not expected. Honey bee foraging is ahighly energetic activity (e.g. Wolf et al. 1989) and at®rst sight it would make sense to warn nest-mates thatan area is unpro®table. In fact, bumblebees use repellentscent marks for this purpose (Stout et al. 1998). In thecontext of a negative signal for a particular forage patch,consider three categories of foraging honey bees ±
77
unemployed foragers, foragers working other patches,and foragers working the same patch ± and whether thepatch abandonment signal would be useful to them. Theabsence of a negative recruitment signal will not reducethe number of unemployed foragers that are directed toa poor patch. This is because no unemployed foragerswill be directed to this patch anyway because recruit-ment of additional foragers is via positive recruitment togood patches. For foragers working other patches, theinformation is not relevant. And for foragers workingthe same patch, the information is redundant becausethey already have this information. In addition, if therewere a patch abandonment signal, the foragers wouldhave to spend time receiving and transmitting this signalin the nest.
Why do honey bees not switch between subtasks?
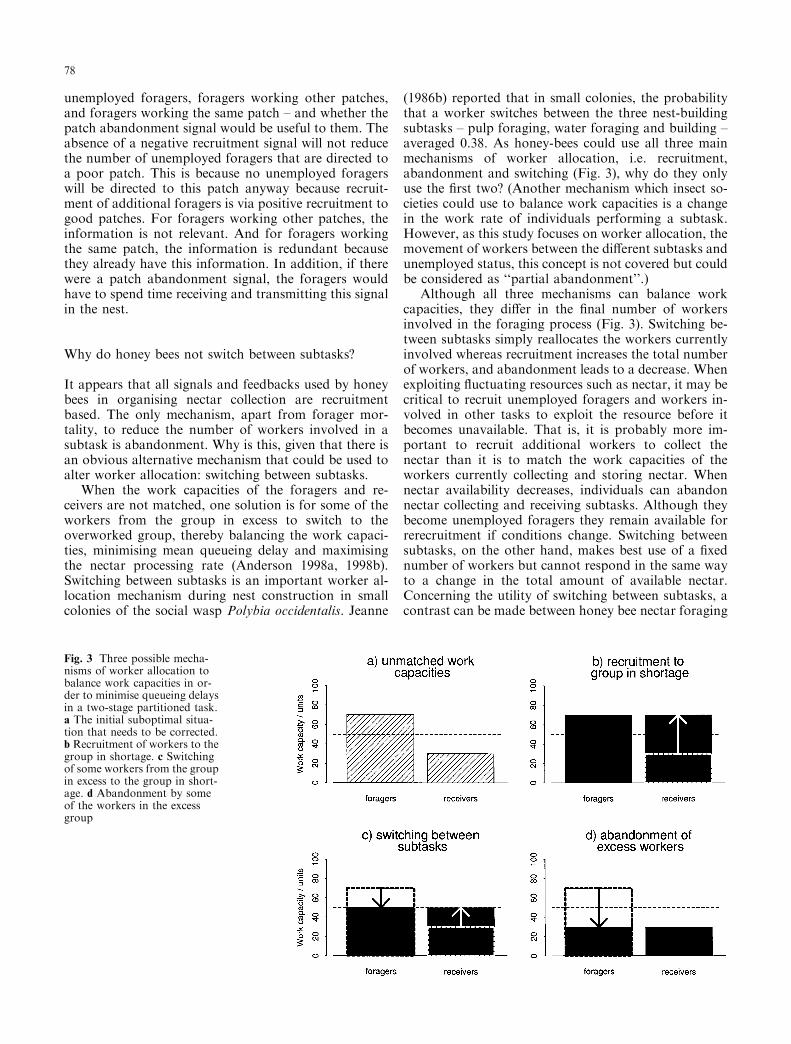
It appears that all signals and feedbacks used by honeybees in organising nectar collection are recruitmentbased. The only mechanism, apart from forager mor-tality, to reduce the number of workers involved in asubtask is abandonment. Why is this, given that there isan obvious alternative mechanism that could be used toalter worker allocation: switching between subtasks.
When the work capacities of the foragers and re-ceivers are not matched, one solution is for some of theworkers from the group in excess to switch to theoverworked group, thereby balancing the work capaci-ties, minimising mean queueing delay and maximisingthe nectar processing rate (Anderson 1998a, 1998b).Switching between subtasks is an important worker al-location mechanism during nest construction in smallcolonies of the social wasp Polybia occidentalis. Jeanne
(1986b) reported that in small colonies, the probabilitythat a worker switches between the three nest-buildingsubtasks ± pulp foraging, water foraging and building ±averaged 0.38. As honey-bees could use all three mainmechanisms of worker allocation, i.e. recruitment,abandonment and switching (Fig. 3), why do they onlyuse the ®rst two? (Another mechanism which insect so-cieties could use to balance work capacities is a changein the work rate of individuals performing a subtask.However, as this study focuses on worker allocation, themovement of workers between the di�erent subtasks andunemployed status, this concept is not covered but couldbe considered as ``partial abandonment''.)
Although all three mechanisms can balance workcapacities, they di�er in the ®nal number of workersinvolved in the foraging process (Fig. 3). Switching be-tween subtasks simply reallocates the workers currentlyinvolved whereas recruitment increases the total numberof workers, and abandonment leads to a decrease. Whenexploiting ¯uctuating resources such as nectar, it may becritical to recruit unemployed foragers and workers in-volved in other tasks to exploit the resource before itbecomes unavailable. That is, it is probably more im-portant to recruit additional workers to collect thenectar than it is to match the work capacities of theworkers currently collecting and storing nectar. Whennectar availability decreases, individuals can abandonnectar collecting and receiving subtasks. Although theybecome unemployed foragers they remain available forrerecruitment if conditions change. Switching betweensubtasks, on the other hand, makes best use of a ®xednumber of workers but cannot respond in the same wayto a change in the total amount of available nectar.Concerning the utility of switching between subtasks, acontrast can be made between honey bee nectar foraging
Fig. 3 Three possible mecha-nisms of worker allocation tobalance work capacities in or-der to minimise queueing delaysin a two-stage partitioned task.a The initial suboptimal situa-tion that needs to be corrected.b Recruitment of workers to thegroup in shortage. c Switchingof some workers from the groupin excess to the group in short-age. d Abandonment by someof the workers in the excessgroup
78

and nest construction in Polybia wasps. In Polybia, thematerials collected are wood and water which are notexpected to ¯uctuate as they are superabundant ± that is,freely available for the taking. In addition, Polybia hasfar fewer workers than the honey-bee and may not haveexcess workers to allocate to these tasks. In this way,nest construction in Polybia is far more suited to subtaskswitching than is nectar collection in the honey bee.
There are at least four additional reasons why re-cruitment plus abandonment might be favoured oversubtask switching in the honey-bee. First, subtaskswitching causes workers to switch between roles. Ifthere is any ``performance e�ciency'' e�ect (Jeanne1986a) and workers improve their ability to perform atask through repetition and learning, this increasedability will be lost when workers switch between the twosubtasks. This is probably not the case with abandon-ment to unemployed status: skilled workers can wait inreserve until required and so the colony su�ers a smallerloss of skills. However, with simple tasks or where per-formance e�ciency is not a factor, recruitment plusabandonment is e�ectively equivalent to switching be-tween subtasks when recruitment rates and abandon-ment rates are comparable. That is, excess workers whoabandon their subtask and are then recruited to theother group, the group in shortage, are e�ectivelyswitching between subtasks and the total number ofactive workers is unchanged. Second, switching betweensubtasks will reduce what is termed the ``demographicadvantage'' (Jeanne 1986a). Foraging is generally themost risky subtask performed by a worker during herlifetime. If she performs this as the last task, she maxi-mises her lifetime work output. Switching between sub-tasks would cause some young bees to switch to foragingand die early. Although there would be no cost if a livingforager switched back to receiving, the costs arise fromworkers that have already died. [Visscher and Dukas(1997) showed that honey-bee foragers are subject to aconstant probability of death per trip.] In addition, ifswitching between foraging and receiving is used tobalance relative work capacities of the two groups, for-agers are unlikely to be able to switch from foraging toreceiving and prolong their life-span and thus their workcontribution to the colony. This is because the necessaryrate of switching from receiving to foraging required tocounteract the depletion of foragers would be muchhigher than from foraging to receiving (Anderson 1998a,1998b). Third, switching between subtasks in the honey-bee would introduce some time lags between being re-cruited and becoming available for the new subtask. Arecruited receiver may ®rst have to store any nectar inher crop before leaving as a forager, or a forager mayhave to store her load of nectar before she can startreceiving depending on when recruitment takes place. Inboth cases, the ability of a colony to track a changingenvironment rapidly will be reduced (Anderson 1998a,1998b). Fourth, switching between subtasks would causesome foragers, who work outside the nest and are moreprone to encounter parasites, to switch to an internal
subtask thus possibly facilitating the spread of disease[see Schmid-Hempel's (1998) ``conveyor belt model''].Moreover, task partitioning in itself increases the amountof interaction between workers (Ratnieks and Anderson1999). Perhaps, and more importantly, the interactionswill be increased between older and potentially diseasedand contagious foragers with low life expectancy and in-nest workers with higher life expectancy. However, theimportance of this e�ect is currently unknown.
Discussion
This paper has presented a functional evaluation of theproximate regulatory mechanisms employed by thehoney bee in the nectar collection and storage process.Regulation involves at least four signals ± the waggledance (von Frisch 1967), the tremble dance (Seeley1992), the stop signal (Nieh 1993) and the shaking signal(Allen 1959) ± and possibly a ®fth, worker piping (Prattet al. 1996), as well as one other mechanism, patchabandonment (Seeley 1995). In theory, abandonment incombination with just two signals should be su�cient:one signal to recruit additional foragers and another torecruit additional receivers (Fig. 2). These three mecha-nisms could deal with all worker allocation require-ments, such as the recruitment of additional workersduring a nectar ¯ow, matching of work capacities tomaximise e�ciency of those workers currently em-ployed, and a reduction in colony activity during anectar dearth. Why are at least two other signals used bythe honey bee? One probable reason is the shortcomingsand uncertainty in the information workers use to makerecruitment decisions. This is suggested by Ratnieks andAnderson (in press a) who show that queueing delays,which are the cues honey bees use to modulate waggleand tremble dances, are inherently unreliable indicatorsof the relative work capacities of foragers and receivers.The mere fact that waggle dances and tremble dancesoccur together, necessitating a mechanism such as thestop signal, demonstrates some of the problems of de-centralised control.
We have proposed various reasons why honey-beesmight use a combination of recruitment and abandon-ment rather than switching between subtasks. Duringnest construction in small colonies of P. occidentalis,workers readily switch between the three buildinggroups (Jeanne 1986b). However, in this situation, allthree subtasks, pulp and water collection, and building,are external subtasks and thus transmission of diseaseinto the colony is unlikely to be a�ected by subtaskswitching. Furthermore, switching between water andpulp foraging is unlikely to involve any loss of perfor-mance e�ciency as both subtasks involve collecting amaterial that is likely to be readily available and in ex-cess. Similarly, there is no loss in the demographic ad-vantage as both are foraging subtasks. Interestingly,switching between subtasks in Polybia, and probably in
79

all species in which it occurs, does conform to the pre-dicted best use of information. Workers experiencinglong delays (which are more reliable sources of infor-mation than the short delays experienced by the group inshortage; Ratnieks and Anderson 1999) when trying too�oad their material are the ones who switch. Theyapply negative feedback to themselves and positivefeedback to the group in shortage, which is the best useof information.
We suggest that the two primary signals in the reg-ulation of nectar foraging are the waggle and trembledances and that these are complemented by two addi-tional secondary signals, the stop signal and the shakingsignal. Waggle and tremble dances are both broadcast(``one-to-many'') signals, although they only operate onquite a local scale, whereas both the stop signal and theshaking signal are clearly ``one-to-one'' signals. Wesuggest that the two broadcast signals, the waggle danceand the tremble dance, have evolved to recruit addi-tional workers quickly. Although quick acting, they areinherently imprecise because of the unreliability in thecue used, the queuing delay experienced by the dancer(Anderson 1998b; Ratnieks and Anderson, in press a), inmaking the decision whether or not to dance. Thus thesetwo signals produce a rapid coarse-tuning of the system.The stop signal and the shaking signal may be morereliable, in that they are based on a greater amount ofinformation and therefore less likely to be made underthe wrong colony conditions. For example, the shakingsignal is made by foragers during changing colonyconditions, something they can detect from their ownexperience and other cues from within the nest. Simi-larly, the cue that workers use when deciding to send thestop signal ± the co-occurrence of waggle and trembledancing ± is also reliable. Moreover, both of the majorgroups of stop signal senders, the tremble dancers anddance followers, are likely to encounter the two dances ifthese are occurring simultaneously. Whereas an indi-vidual waggle dancer performs her dance on a smallportion of comb and is unlikely to gain information onwhat other dancers are doing, a tremble dancer's signaltakes her over a much larger portion of comb (Seeley1992, 1995), and dance followers are free to move acrossthe whole dance area. It seems logical that the role of thestop and shaking signals in ®ne-tuning the system hasevolved after coarse tuning. Whereas the stop signaloperates on waggle dances and its present function hasclearly evolved at a later stage, it is not clear when theshaking signal evolved. It was possibly the ®rst signalthat honey-bees used to regulate foraging (as a way ofincreasing total foraging e�ort) but could also haveevolved later as ®ne-tuning.
Honey-bees still hold tantalising secrets concerningtheir colony organisation. Nectar foragers have a greaterrole than would be expected in that they recruit bothadditional foragers and receivers. What is it about for-agers' ``contextual information'', that is, their own ex-perience in addition to the information from other cuesand signals from that particular trip [stretching Seeley's
(1998) de®nition to include signal senders as well as re-cipients], which makes them more reliable recruitersthan are the nectar receivers? Certainly they know thelocations and qualities of various ¯ower patches, but dothey possess other valuable information? Are there anyadditional signals used in regulating foraging such aspiping? Our analysis does not discount this possibilitybut the honey-bee already has more signals than thetheoretical minimum needed. We predict that if more arediscovered, than they will be used to ®ne-tune the systemdue to limitations in information quality which preventthe system from otherwise being precisely optimised. Itis clear that the honey bee has a complex system ofregulating nectar collection. But, as we have demon-strated in this review, it is also subtle and sophisticated.
Acknowledgements C.A. was supported by a postdoctoral fellow-ship from the research network ``Social Evolution'' of the Uni-versities of Aarhus, Firenze, Keele, She�eld, Uppsala, WuÈ rzburgand the ETH ZuÈ rich, ®nanced by the European Commission viathe Training and Mobility of Researchers (TMR) programme.
References
Allen DM (1959) The ``shaking'' of worker honeybees by otherworkers. Anim Behav 7:233±240
Anderson C (1998a) Simulation of the feedbacks and regulation ofrecruitment dancing in honey bees. Adv Complex Syst 1:267±282
Anderson C (1998b) The organisation of foraging in insect soci-eties. PhD thesis, School of Mathematics, University of Shef-®eld
Anderson C, Ratnieks FLW (in press a) Task partitioning in for-aging: general principles, examples, e�ciency and informationreliability of queueing delays. In: Detrain C, Deneubourg JL,Pasteels JM (eds.) Information processing in social insects.BirkhaÈ user, Berlin
Anderson C., Ratnieks FLW (in press b) Task partitioning in insectsocieties. I. E�ect of colony size on queueing delay and colonyergonomic e�ciency. Am Nat
Armbruster L (1922) UÈ ber BienentoÈ ne, Bienensprache undBienengehoÈ r. Arch Bienenkd 4:221±259
Bonabeau E (in press) Social insect colonies as complex adaptivesystems. Ecosystems
Bonabeau E, Theraulaz G, Deneubourg JL, Aron S, Camazine S(1997) Self-organization in social insects. Trends Ecol Evol12:188±193
Esch H (1964) BeitraÈ ge zum Problem der Entfernungsweisung inden SchwaÈ nzeltaÈ nzen der Honigbiene. Z Vergl Physiol 48:534±546
Frisch K von (1967) The dance language and orientation of bees.Harvard University Press, Cambridge, Mass
Heinrich, B (1979) Keeping a cool head: honeybee thermoregula-tion. Science 205:1269±1271
Jeanne RL (1986a) The evolution of the organization of work insocial insects. Monit Zool Ital 20:119±133
Jeanne RL (1986b) The organization of work in Polybia occiden-talis: costs and bene®ts of specialization in a social wasp. BehavEcol Sociobiol 19:333±341
Kirchner WH (1993) Vibrational signals in the tremble dance of thehoney bee, Apis mellifera. Behav Ecol Sociobiol 33:169±172
Lloyd JE (1983) Bioluminescence and communication in insects.Annu Rev Entomol 28:131±160
Milum VG (1955) Honey bee communication. Am Bee J 95:97±104Nieh JC (1993) The stop signal of honey bees: reconsidering its
message. Behav Ecol Sociobiol 33:51±56
80

Nieh JC (1998) The honey bee shaking signal: function and designof a modulatory communication signal. Behav Ecol Sociobiol42:23±36
Pratt SC, KuÈ hnholz S, Seeley TD, WeidenmuÈ ller A (1996) Workerpiping associated with foraging in undisturbed queenrightcolonies of honey bees. Apidologie 27:13±20
Ratnieks FLW, Anderson C (1999) Task partitioning in social in-sects. Insectes Soc 46:95±108
Ratnieks FLW, Anderson C (in press a) Task partitioning in insectsocieties. II. Use of queueing delay information in recruitment.Am Nat
Ratnieks FLW, Anderson C (1999) Task partitioning in insectsocieties natural history, ergonomics, and information. ActesColloq Insectes Soc 12:1±10
Ribbands R (1953) The behaviour and social life of honeybees. BeeResearch Association, London
Schmid-Hempel P (1998) Parasites in social insects. PrincetonUniversity Press, Princeton, NJ
Schneider SS, Stamps JA, Gary NE (1986) The vibrational dance ofthe honey bee. I. Communication regulating the foraging ontwo time scales. Anim Behav 34:377±385
Seeley TD (1985) Honeybee ecology: a study of adaptation in sociallife. Princeton University Press, Princeton, NJ
Seeley TD (1992) The tremble dance of the honey bee: message andmeanings. Behav Ecol Sociobiol 31:375±383
Seeley TD (1995) The wisdom of the hive: the social physiology ofhoney bee colonies. Harvard University Press, Cambridge,Mass
Seeley TD (1997) Honey bee colonies are group level adaptiveunits. Am Nat 150:22±41
Seeley TD (1998) Thoughts on information and integration inhoney bee colonies. Apidologie 29:67±80
Seeley TD, KuÈ hnholz S, WeidenmuÈ ller A (1996) The honey bee'sdance stimulates additional bees to function as nectar receivers.Behav Ecol Sociobiol 39:419±427
Seeley TD, WeidenmuÈ ller A, KuÈ hnholz S (1998) The shaking signalof the honey bee informs workers to prepare for greater activity.Ethology 104:10±26
Stout JC, Coulson D, Allen JA (1998). Repellent scent-marking of¯owers by a guild of foraging bumblebees (Bombus spp.). BehavEcol Sociobiol 43:317±326
Visscher PK, Dukas R (1997) Survivorship of foraging honey bees.Insectes Soc 44:1±5
Wolf TJ, Schmid-Hempel P, Ellington CP, Stevenson RD (1989)Physiological correlates of foraging e�orts in honey-bees: oxy-gen consumption and nectar load. Behav Ecol Sociobiol 3:417±424
Communicated by R.F.A. Moritz
81





























