Embed Size (px)
Citation preview

INTRODUCTION
The primordia of mesodermal tissues, such as visceral andsomatic mesoderm, dorsal vessel and fat body, becomedetermined as a result of progressive subdivisions of themesoderm along both the dorsoventral and anteroposterioraxes of the embryo (reviewed by Frasch and Nguyen, 1999;Frasch, 1999). The dorsoventral subdivision is a strictlyinductive process and leads to the formation of visceralmesoderm, the dorsal vessel, and specific somatic muscleprogenitors in the dorsal mesoderm. This induction requiresthe activity of ectodermal Dpp in conjunction with themesodermal NK homeodomain protein Tinman (Staehling-Hampton et al., 1994; Frasch, 1995; Xu et al., 1998). Whilethese dorsoventral activities act uniformly upon the entiretrunk mesoderm, the anteroposterior subdivisions of themesoderm occur within each parasegment in a repetitivefashion. These latter subdivisions involve both mesoderm-intrinsic and inductive patterning mechanisms and lead to ametameric arrangement of tissue primordia within the dorsalas well as the ventrolateral mesoderm (Dunin-Borkowski etal., 1995; Lawrence et al., 1995; Azpiazu et al., 1996). During
these processes, the mesoderm-intrinsic activities depend onpair-rule genes, such as even skipped(eve), whereas theinductive activities include Wingless (Wg) and Hedgehog(Hh), which are expressed in transverse stripes in theectoderm. Together, these findings have led to the view thatthe determination of individual mesodermal tissues ispromoted, or in some cases suppressed, in quadrants of cellsthat are positioned at the intersections of specific dorsoventraland anteroposterior cues.
The developmental events at the intersections between theDpp and Wg activity domains have been well studied. Inparticular, Wg has been shown to exert both positive andnegative effects in these domains. On the one hand, Wg signalsare instrumental for the induction of heart and dorsal muscleprogenitors at these intersections (Park et al., 1996; Wu et al.,1995; Azpiazu et al., 1996; Carmena et al., 1998); analogousWg activities also induce lateral and ventral muscle progenitors(Baylies et al., 1995). On the other hand, Wg signaling servesto suppress the induction of trunk visceral mesoderm at thesepositions, thereby restricting the formation of progenitors ofcircular midgut muscles to metamerically repeated cell clustersin dorsal areas between the striped domains of Wg activity
5497Development 127, 5497-5508 (2000)Printed in Great Britain © The Company of Biologists Limited 2000DEV7841
Inactivation of either the secreted protein Wingless (Wg)or the forkheaddomain transcription factor Sloppy Paired(Slp) has been shown to produce similar effects in thedeveloping Drosophilaembryo. In the ectoderm, both geneproducts are required for the formation of the segmentalportions marked by naked cuticle. In the mesoderm, Wgand Slp activities are crucial for the suppression of bagpipe(bap), and hence visceral mesoderm formation, and thepromotion of somatic muscle and heart formation withinthe anterior portion of each parasegment. In this report,we show that, during these developmental processes, wgand slp act in a common pathway in which slpserves as adirect target of Wg signals that mediates Wg effects in bothgerm layers. We present evidence that the induction ofslp by Wg involves binding of the Wg effector Pangolin(Drosophila Lef-1/TCF) to multiple binding sites within aWg-responsive enhancer that is located in 5′flankingregions of the slp1 gene. Based upon our genetic and
molecular analysis, we conclude that Wg signaling inducesstriped expression of Slp in the mesoderm. Mesodermal Slpis then sufficient to abrogate the induction of bagpipebyDpp/Tinman, which explains the periodic arrangement oftrunk visceral mesoderm primordia in wild type embryos.Conversely, mesodermal Slp is positively required,although not sufficient, for the specification of somaticmuscle and heart progenitors. We propose that Wg-induced slp provides striped mesodermal domains with thecompetence to respond to subsequent slp-independent Wgsignals that induce somatic muscle and heart progenitors.We also propose that in wg-expressing ectodermal cells, slpis an integral component in an autocrine feedback loop ofWg signaling.
Key words: wingless, sloppy paired, pangolin, LEF-1, dTCF,bagpipe, Mesoderm, Patterning, Myogenesis, Cardiogenesis,Signaling, Induction, Drosophila
SUMMARY
Wingless effects mesoderm patterning and ectoderm segmentation events
via induction of its downstream target sloppy paired
Hsiu-Hsiang Lee and Manfred Frasch*
Department of Biochemistry and Molecular Biology, Mount Sinai School of Medicine, One Gustave L. Levy Place, New York, NY10029, USA*Author for correspondence (e-mail: [email protected])
Accepted 3 October; published on WWW 14 November 2000

5498
(Azpiazu et al., 1996). An intriguing question is how Wgsignals achieve these opposing effects at the molecular level.
The homeobox genes even skipped(eve), slouch(slou), mshand the bHLH gene nautilus (nau) are examples of Wgdownstream genes that are positively regulated by Wgsignaling and important for normal somatic muscle and heartdevelopment (Wu et al., 1995; Baylies et al., 1995; D’Alessioand Frasch, 1996; Ranganayakulu et al., 1996; Keller et al.,1998; Knirr et al., 1999; Nose et al., 1998; Su et al., 1999).Conversely, the NK homeobox gene bagpipe(bap), which is afocus of the present study, is a primary example of a gene forthat is negatively regulated by Wg in the mesoderm. In thesegmental domains in which bap induction by the combinedactivities of Dpp and Tinman is not blocked by Wg, bapfunctions as a critical regulator that controls visceral mesodermdevelopment (Azpiazu and Frasch, 1993; Azpiazu et al., 1996).
As shown in our present study, the sloppy paired(slp) genesprovide a key to our understanding of the differential activitiesof Wg signals in the mesoderm. slp1 and slp2 (hereaftercollectively called slp) form a functionally redundant pair ofgenes, which were originally identified as segmentation genesthat encode forkheaddomain transcription factors (Nüsslein-Volhard et al., 1984; Grossniklaus et al., 1992; Cadigan et al.,1994b). More recently, Riechmann et al. (1997) have shownthat slp mutants display mesodermal phenotypes that arehighly reminiscent of wgphenotypes. Like wg mutantembryos, slp mutants show a loss of muscle and heartprogenitors, as well as derepression of bap and concomitantexpansion of the visceral mesoderm. These similaritiesindicated that wgand slpact in a common pathway duringmesoderm patterning. Based upon previous observations,which showed that slp is required for the maintenance ofectodermal wg expression (Cadigan et al., 1994a), Riechmannet al., proposed a model in which slp acts within the ectodermand upstream of wgduring the induction of heart and somaticmuscle progenitors. However, because of the stronger effectsof slp, as compared with wg, on bap, it was assumed that thenegative effects of slpinvolve a separate pathway, in which slpacts independently of wg.
We show herein that slp is downstream of wgand mediatesWg effects during both baprepression and somaticmuscle/heart induction. slpis expressed in a striped patternwithin the mesoderm that, after the disappearance of pair-rulegene expression, becomes strictly dependent on Wg signals.Moreover, our identification of functionally important bindingsites for the Wg effector Pangolin within a Wg responsiveenhancer element from the slplocus suggests that slpis a directtarget of the Wg signal transduction cascade. We further showthat the wgeffects on bap repression are exclusively mediatedby slp, whereas the wgeffects on somatic muscle and heartinduction requires an additional slp-independent pathway.Altogether, these observations integrate previous observationson the mesodermal functions of wg and slp,and provide acohesive picture of their roles in mesoderm patterning alongthe anteroposterior axis.
MATERIALS AND METHODS
Drosophila strainsThe following Drosophilamutant lines were used in this study: wgcx4,
hh13C29 and slp∆34B. In slp∆34B both slp1 and slp2 are inactivated(Grossniklaus et al., 1992). The UAS and Gal4 lines used in this studyare UAS-wg2, UAS-hh1(Azpiazu et al., 1996), pUAST-D-en (Guillenet al., 1995), SL159.3F(UAS-slp1insertion on the third chromosome;a gift from G. Struhl, Columbia University), E22c-Gal4(Yoffe et al.,1995) for ectodermal expression and GALSG30(twi-Gal4 on the Xand 24B-Gal4on the third chromosome) or GALSG24(twi-Gal4 onX) (Brand and Perrimon, 1993; Greig and Akam, 1993) formesodermal expression. For UAS/GAL4 induced ectopic expression,embryos were collected at 29°C. For ectopic expression of bothslp1 and wg in the same embryos, UAS-wg2 and SL159.3Fwererecombined onto the same chromosome.
Embryo stainingAntibody staining was performed as described in Azpiazu et al.(1996), and double fluorescent staining was as described in Knirr etal. (1999). The following antibodies were used in this study:monoclonal mouse anti-En antibody (4D9, 1:4, Corey Goodman/Developmental Studies Hybridoma Bank, Univ. of Iowa), rabbit anti-Eve antibody (1:3000; Frasch et al., 1987), guinea pig anti-Slpantibody (1: 100; Kosman et al., 1998), monoclonal mouse anti-Wgantibody (4D4, 1:12; Steve Cohen/Developmental Studies HybridomaBank, Univ. of Iowa), monoclonal mouse anti-Fasciclin 3 antibody(7G10, 1:20, Corey Goodman/Developmental Studies HybridomaBank, Univ. of Iowa) rabbit anti-β-gal antibody (1:3000, Cappel) andrabbit anti-Bap antibody (1:250). The Bap antibody was raised againsta His fusion protein that was induced from a pQE-9 plasmid(Novagen), which contained a 600 bp PstI fragment from the bapcDNA 4 (Azpiazu and Frasch, 1993).
Construction of P-transformation plasmidsA series of constructs containing various slp1 upstream enhancerelements was generated by PCR using P1 clone DS00065 (BerkeleyDrosophila Genome Project) as template and different pairs of primersdesigned to contain either a BamHI or SalI site at their 5′ends. AllPCR fragments were cloned into BamHI/XhoI of pCaSpeRhs43β-galvectors in the native orientations relative to the basal promoters. Forthe slp5′-1 fragment (−981/−1905), primer slp0.9R GCGTCGACAT-GTGGATCTCTGGCAATCC and primer slp1.8 GCGGATCCTA-TCGCTCGATTGCCGTCTC were used; for slp5′-2 fragment(−981/−2553), primers slp0.9 and slp2.5 GCGGATCCAGGATC-TCGAATCGCTAATC were used; for slp5′-3 (−3340/−3980), primersslp3.3R GCGTCGACGAATGAATCCTGCTACTTGG and slp3.9GCGGATCCCTTTATAATTCGATCAACTC were used; for slp5′-4(−4660/−5396), primers slp4.5R GCGTCGACTCCCAGCTAGCT-CCCTTCA and slp5.3 GCGGATCCGCCCAAAGTCGGTCGAA-CAG were used.
For in vitro mutagenesis of the slp5′-2 fragment, slp5′-2 in pKS+was used as PCR template with different primer sets, which weredesigned to introduce new restriction sites replacing the Pan-bindingsites. For example, to mutate Pan A and Pan B in slp5′-2, EcoRIand HindIII sites were introduced with the PCR primersGCGAATTCCCCCAAAGGATGTTTGTCC and CCCAAGCTTGG-TCCGGACTTTTCGTTTG. The PCR products were digested withEcoRI and HindIII, then ligated with a double-strandedoligonucleotide, CCCAAGCTTTCCATTTCGGTAGCAAAGTAGA-GTCGGCGAGACTTCGAATTCGC, which spans the sequencesbetween Pan A and B, to generate a slp5′-2 fragment with mutationsin both Pan A and B. The following primers and oligonucleotides wereused to mutate additional Pan-binding sites in slp5′-2 with ananalogous strategy. The primers, ATAAGATCATATGTGCACAGTG-CCGCCGACA and CCCCATCGATTAGCAACAACACATGTTC-CA, and oligonucleotide CCATCGATCTGTTTTGTCAAGTTGTCT-AAGATCATATGCGCACAG, were used to mutate Pan E and F. Theprimers, CGGAATTCCTCTTACGACCAGAAACTAT and GCGAA-TTCGAATTATATATCCCTTCCTG, were used to mutate Pan I. Theprimers, GAAGATCTACACAAAAACCACTTGTACATTC and CA-
H.-H. Lee and M. Frasch

5499Slp effects Wg signals in mesoderm and ectoderm
TGCCATGGCACAATTGAACAAATATAC, and oligonucleotideCATGGCTACATGCCATAAAAACATAAAAGCA, were used tomutate Pan C and D. The mutated construct slp5′-2m5 contains fivemutated Pan binding sites (A, B, E, F and I), and construct slp5′-2m7contains seven mutated sites (A, B, C, D, E, F and I). All constructswere sequenced to confirm that only the intended mutations wereintroduced and cloned into pCaSperhs43β-gal.
DNA-binding assaysFootprinting assays were performed essentially as described in Yin etal. (1997) with the following minor modifications. The slp5′-2fragment was divided into three overlapping sub-fragments by PCR,which were cloned into pKS+ vectors. To generate single-end-labeledprobes, the DNA fragments were amplified by PCR with 5′-phosphorylated T3 or T7 primers, respectively, and an internalunphosphorylated primer. This DNA fragment was then single-endlabeled with γ32P-ATP by T4 DNA polynucleotide kinase, since the5′-end on the other side was blocked. The internal primers usedto make DNA fragments for probe labeling were slp1.3CCGGGATCCTGTATATTTGTTCAAT, slp1.3R GCGGGATCCG-TTAGAACAATTGAAC and slp2.5. Different amounts of GST-Pan(dTCF HMG domain; amino acids 271-366; Waltzer and Bienz, 1998)were added to the reaction mixture (50 mM KCl, 25 mM HepespH7.9, 6 mM MgCl2, 1 mM DTT, 10% glycerol, 1.5 µg poly-dIdC,0.25 mg/ml and 104 cpm of single end-labeled probe). Afterincubation on ice for 1 hour, 0.03 U DNaseI (Boehringer) was addedto a final concentration of 6×10−4 U/µl and incubated on ice fortwo minutes. The purified DNA fragments were separated on 6%polyacrylamide DNA sequencing gels.
For gel mobility shift assays, 50 ng of annealed oligonucleotideswere labeled with γ32P-ATP using T4 DNA polynucleotide kinase.Recombinant GST-Pan protein was added in a 10 µl volumecontaining 104 cpm probe, 0.5 µg poly (dI-dC), variable amountsof specific competitor DNA, 4% Ficoll, 20 mM Hepes (pH7.6),50 mM KCl, 1 mM DTT and 0.25 mg/ml BSA. After incubatingon ice for 30 minutes, the mixture was loaded on a 4%polyacrylamide:bisacrylamide (29:1) gel and run in 0.25×TBE bufferfor 2 hours. The following oligonucleotides were used as labeledprobes or specific competitor DNA: Pan I-wt ATATAATTTTCAAA-GGTCTTAC and Pan I-mt ATATAATTCGAATTCCTCTTAC.
RESULTS
slp acts downstream of wg to repress bap andvisceral mesoderm formationwg and slp are required to abrogate bapexpression in asegmental fashion, thereby confining the primordia of trunkvisceral mesoderm to metameric quadrants of cells in thedorsal mesoderm (Azpiazu et al., 1996; Riechmann et al.,1997). These functional similarities and the overlappingdomains of wgand slpexpression (Cadigan et al., 1994a)suggest that the two genes are components of a commonregulatory pathway that leads to baprepression. Therefore, wetested the epistatic relationships between wg and slpin thispathway. In addition to the mesoderm, we also monitored the
Fig. 1. Repression of bapand suppression of trunk visceral mesoderm by mesodermal Slp. Confocal images are shown of stage 10 embryosstained with antibodies against Bap (green) and Slp (red), except for D,E, which show stage 12 embryos stained for fasciclin 3. (A-C,F) Mergedscans displaying Bap signals, i.e. mesodermal images. (G) Merged scans of exclusively ectodermal images. Broken lines indicate the bordersbetween the mesoderm (ms) and ectoderm (ec). (A) Wild-type embryo with mutually exclusive domains of Bap and Slp expression.(B) Embryo with ectopic mesodermal slpexpression (driven by SG30-GAL4). Bap expression is strongly reduced. (C) Ectopic mesodermal slpexpression (driven by twist-GAL4) in a slpmutant background. Mesodermal Slp is sufficient to repress bap. (D) Wild-type embryo stained forfasciclin 3, which marks the trunk visceral mesoderm (vm). (E) Embryo with ectopic mesodermal slpexpression as in C. Fasciclin 3 stainingreveals strong reduction of trunk visceral mesoderm. (F) Embryo with ectopic mesodermal enexpression (driven by SG30-GAL4).Disappearance of mesodermal Slp correlates with ectopic Bap expression. (G) Ectodermal view of same embryo as in F, showing normalectodermal Slp pattern.

5500
regulatory relationships between wgand slpin the ectodermduring the course of these experiments.
At the stage when bap is activated, wgis not expressed atdetectable levels in the mesoderm (Baker, 1987; van denHeuvel et al., 1989; H.-H. L. and M. F., unpublishedobservations). By contrast, confocal microscopy showed thatSlp was strongly expressed in the mesoderm during this period(Fig. 1A). Mesodermal expression of Slp occurred intransverse stripes, which are positioned exactly underneath theectodermal Slp stripes. Importantly, Slp and Bap wereexpressed in mutually exclusive domains in the dorsalmesoderm, which invokes the possibility that Slp is a repressorof bap. To test this possibility, we used the binary UAS/GAL4system to drive ectopic slpexpression. As shown in Fig. 1B,uniform expression of slpin the mesoderm resulted in analmost complete repression of bapexpression. A similar degreeof bap repression was observed when slpwas uniformlyexpressed in the mesoderm of slp mutant embryos, whichnormally display complete derepression of bap along theanteroposterior axis (Fig. 1C; Riechmann et al., 1997). Ectopicexpression of slpin the mesoderm prevented the formation oftrunk visceral mesoderm, presumably as a consequence of therepression of bap(Fig. 1E, compare with Fig. 1D). We alsotested the effects of a reciprocal experiment, in which slpexpression was eliminated only in the mesoderm. As shown inFig. 1F, ectopic mesodermal engrailed (en)removed theexpression of slpspecifically in the mesoderm, whileectodermal slp expression was unaffected (Fig. 1G). Aspredicted, bap was completely derepressed under theseconditions in the dorsal mesoderm (Fig. 1F). Taken together,we conclude that, in the normal situation, Slp acts in themesoderm to repress bapin a segmental fashion.
Although previous studies have indicated that, in theectoderm, slp is an upstream activator of wg(Cadigan et al.,1994a,b), the above data suggest that, in the mesoderm, slpactsdownstream or in parallel to wgto repress bap. In agreementwith this notion, the absence of wg activity resulted in thecomplete loss of slpin the mesoderm after stage 9 ofembryogenesis (Fig. 2A,B). In the ectoderm of wgmutants, slpexpression was also lost except for narrowly-striped areas inthe neuroectoderm (Fig. 2A,B; see also Bhat et al., 2000). Lossof slp expression in wgmutants was accompanied by anexpansion of bapexpression, which initiated in a segmentedpattern but then rapidly became continuous (Fig. 2A,B). Thenormal initiation of bapexpression could either be due to thepresence of Slp stripes at earlier stages, which are initiallyestablished by pair-rule genes, and/or due to the presence of astriped coactivator. In this respect, previous studies had shownthat hh has a positive effect on bapexpression and that ectopichh expression in the ectoderm of wgmutants yielded the sameextent of bapderepression as observed in slpmutants (Azpiazuet al., 1996, Riechmann et al., 1997). As shown in Fig. 2C,D,embryos with this particular genotype wg−, E22c>>hh, showedan equal loss of slp expression as embryos that were mutantfor wgalone. bapexpression initiated along the entire A/P axisof these embryos (Fig. 2C) and later occurred throughout thedorsal mesoderm (Fig. 2D). In a converse experiment, whichinvolved ectopic expression of wg in the ectoderm of hhmutants, an almost total absence of bapexpression wasobserved (Azpiazu et al., 1996). Notably, this genotype hh−,E22c>>wg, produces strong expansion of slp expression in the
mesoderm, particularly in the posterior segments (Fig. 2E).Based upon the data with ectopic slp (Fig. 1B), this expansionof slp could account for the absence of bap expression.However, the lack of significant levels of bap expression insome of the remaining gaps of slpexpression indicates that theabsence of hhactivity could also contribute to the loss of bapexpression in a slp-independent manner. In the ectoderm,ectopic expression of wgin the absence of hhproducesexpanded slp stripes as well, although the expansion is less
H.-H. Lee and M. Frasch
Fig. 2.Regulation of slpand bapexpression by wgand hh. Shownare embryos stained for Bap (green) and Slp (red). (A) Stage 9 wgmutant embryo. Striped Slp expression is fading and Bap expressioninitiates in a segmental pattern (v, ventral midline). (B) Stage 10 wgmutant embryo. Except for some neuroectodermal cells, Slpexpression has disappeared, while Bap expression has expanded.(C) Stage 9 wgmutant embryo with ectopic ectodermal hhexpression (driven by E22c-GAL4). Slp expression is fading, whileBap expression is initiating in a uniform pattern along the A/P axis.(D) Stage 10 embryo, genotype as in C. Except for someneuroectodermal cells, Slp expression has disappeared, while Bap isexpressed uniformly in the dorsal mesoderm. (E) Stage 10 hhmutantembryo with ectodermal ectopic expression of wg(mergedmesodermal scans). Expansion of mesodermal Slp is accompaniedby the loss of Bap expression. (F) Same embryo as in E, but mergedectodermal scans. Slp stripes in the ectoderm are expanded, althoughnot as strongly as in the mesoderm. ec, ectoderm; ms, mesoderm.

5501Slp effects Wg signals in mesoderm and ectoderm
pronounced when compared with the mesoderm (Fig. 2F). Ofnote, ectopic expression of wgin the ectoderm or mesodermof wild-type embryos does not cause a significant expansion ofslp expression (data not shown), which is consistent with themild effects of ectopic wgon bapexpression (Azpiazu et al.,1996). Together, these observations indicate that slp respondsto Wg in both the mesoderm and ectoderm, particularly in theabsence of Hh signaling. In addition, the effects of wg on bapcan be largely explained by its positive effects on theexpression of Slp, which in turn appears to serve as a repressorof bap.
As a final experiment to clarify the epistatic relationshipsbetween wg and slp with respect to bapregulation, weexpressed each of the two genes in the absence of the other. Asshown in Fig. 3B, uniform mesodermal expression of wg in slpmutant backgrounds did not result in any repression of bap(compare with Fig. 3A for an analogously stained wild-typeembryo; for bap-stained slpmutants, see Fig. 3R, Riechmannet al., 1997). By contrast, uniform mesodermal expression ofslp in wg mutants produces full repression of bap, similar tothe effects of ectopic slpin wild-type backgrounds (Fig. 3C,compare with Fig. 1B). Altogether, these data suggest that slpexpression is induced downstream of wg in both the ectodermand mesoderm. In the mesoderm, Slp in turn represses bapexpression in a segmental fashion.
wg and slp cooperate in the generation of heart andmuscle progenitorsSince the progenitors of pericardial cells, cardioblasts andsomatic muscles are largely derived from the segmental areasbetween the bapdomains, and their formation requires wgandslp as positive activities, we asked whether in this regulatorypathwayslp also acts strictly downstream of wg. In wild-typeembryos, eve-expressing progenitors of pericardial cells anddorsal muscles are located underneath the ectodermal wgstripes (Jagla et al., 1997; Carmena et al., 1998) and within thedomains of mesodermal slpexpression (Fig. 4A). In wgmutantembryos, these progenitors are not formed (Wu et al., 1995;Lawrence et al., 1995). Interestingly, while ectopic slpexpression in the mesoderm of wg mutants rescued baprepression (see Fig. 3C), it fails to rescue the formation of heartand dorsal muscle progenitors (Fig. 4B). Identical effects areobserved in slpmutants (Riechmann et al., 1997) and in slp
mutant embryos with ectopic mesodermal expression of wg(Fig. 4C); neither produces any eve-expressing heart andmuscle progenitors. The results of ectopic expression of slp orwg in wild-type backgrounds were also consistent with theseobservations. Although ectopic expression of slp in themesoderm of wild-type embryos caused complete repressionof bap (see Fig. 1B), it did not produce a significant increaseof eve-expressing cells under the same conditions (Fig. 4D).Similarly, ectopic expression of wgin the mesoderm (orectoderm) of wild-type embryos with the GAL4/UAS systemfailed to produce a major increase of eve-expressing cells (Fig.4E; see also Lawrence et al., 1995 and Park et al., 1996, whoreported slightly larger increases of evecells in their ectopicexpression experiments with UAS-wgand hs-slp). By contrast,when wg and slp were coexpressed in the mesoderm under thesame conditions as before, there was a dramatic increase ineve-expressing cells, which became distributed all along thedorsal margin of the mesoderm (Fig. 4F). Analogousexperiments have shown that the formation of slouch-expressing ventrolateral muscle progenitors and tin-expressingheart progenitors also requires the joint activities of wgand slp(data not shown). These combined data show that neither wgnor slp alone is sufficient to allow the formation of variousheart and muscle progenitors. Instead, it appears that wg andslp cooperate in parallel or consecutive pathways during theformation of these cell types.
We next asked whether one of these pathways involves theregulation of twistexpression by mesodermal Slp. Previousreports have indicated that wgand slpare required to generatesegmentally elevated levels of twistexpression during stage 11and that these higher levels of twistare necessary for normalsomatic muscle specification (Baylies and Bate, 1996,Riechmann et al., 1997). As shown in Fig. 4G,H, the stripes ofelevated Twist coincided with the mesodermal domains of Slpduring stage 11. Moreover, uniform expression of slp in themesoderm produced uniform expression of high Twist levels,even in the absence of Wg activity (Fig. 4I). Thus, it appearsthat one route of wgfunction in myogenesis involves theupregulation of twistby Wg-induced slp in the mesoderm.
Additional experiments confirm that, analogous to baprepression, the mesodermal component of slpis essential forsomatic muscle and heart specification. In these experiments,mesodermal slp expression was specifically removed by
Fig. 3.Epistatic relationships between wgand slpwith respect to bapregulation. (A) Stage 10 wild-type embryo stained for Bap (green) andWg (red) proteins. Confocal scans of both the ectodermal and mesodermal layers were merged. (B)slp mutant embryo with uniform Wgexpression driven by twi-GAL4 in the mesoderm (stained as in A). bap is uniformly expressed and not repressed by Wg. (C)wg mutant embryowith uniform Slp expression driven by twi-GAL4 in the mesoderm (stained for Bap/green and Slp/red). Slp represses bapas strongly as in thewild-type background.

5502
ectopically expressing enin the mesoderm of wild-typeembryos, as described above. Embryos of this genotype,SG30>>en showed a complete absence of eve-expressingmuscle and pericardial progenitors, and almost completeabsence of Mef2-labeled somatic mesoderm (Fig. 4J, compare
with Fig. 4M). The expression of nautilus in muscle founderswas also missing in embryos of this genotype (data not shown).In addition, tin-expressing heart progenitors failed to beformed (Fig. 4K, compare with Fig. 4N) and only a fewsomatic muscle fibers were present in late-stage embryos (Fig.
H.-H. Lee and M. Frasch
Fig. 4.Regulation of muscle andheart development by wg andmesodermalslp. (A) Stage 10wild-type embryo stained for Eve(green) and Slp (red). Eve-staineddorsal muscle and pericardialprogenitors are found within themesodermal Slp domains. High-magnification views (insets) shownuclear co-localization of Eve andSlp during stage 10. (B)wgmutant embryo with twi-GAL4driven slpexpression, stained as inA. Eve-expressing dorsalmesodermal cell types are notrescued by slp. (C)slpmutantembryo with twi-GAL4 driven wgexpression, stained for Eve (green)and Wg (red). Eve-expressingdorsal mesodermal cell types arenot rescued by wg. (D) Embryowith SG30-GAL4 drivenmesodermal expression of slp,stained as in A. (E) Embryo withSG30-GAL4 driven mesodermalexpression of wg, stained as in C.(F) Embryo with SG30-GAL4driven mesodermal expression ofslpand wg, stained as in A. Notethat the ectopic expression levelsof slpare lower than theendogenous levels. The number ofEve cells is strongly increased.(G,H) Stage 11 wild-type embryostained for Slp (red) and Twist(green). The stripes with highTwist levels coincide with themesodermal Slp stripes. (I) Stage11 wgmutant embryo with twi-GAL4 driven slpexpression,stained for Twist (green) and Slp(red, data not shown). Themesoderm displays uniformly highlevels of Twist. (J) Stage 12embryo with SG30-GAL4 drivenmesodermal expression of en,stained for Mef2 (brown) and Eve(purple). Pericardial and muscleprogenitors are missing and only afew somatic mesodermal cellsremain. (K) Stage 13 embryo withgenotype as in J, stained for Tinprotein. Tin-expressing heartprecursors are missing, whilesegmentally-stained visceralmesodermal cells are present. (L) Stage 17 embryo with genotype as in J, stained for myosin heavy chain. Only a few somatic muscle fibers and some unfused myoblasts areobserved. (M) Stage 12 wild-type control embryo stained as in J. (N) Stage 13 wild-type control embryo stained as in K. (O) Stage 17 wild-typecontrol embryo stained as in L. ec, ectoderm; gmc, ganglion mother cells; hp, heart progenitors; mp, muscle progenitors; ms, mesoderm; pc,pericardial progenitors; sm, somatic mesoderm/muscles; vm, trunk visceral mesoderm.

5503Slp effects Wg signals in mesoderm and ectoderm
4L, compare with Fig. 4O). In contrastto the loss of derivatives of themesodermal A (slp) domains (Azpiazuet al., 1996 Riechmann et al., 1997),derivatives of the P (eve) domains,including trunk visceral mesoderm (Fig.4J,M) and fat body (data not shown),were present and even expanded in theseembryos. In the aggregate, the evidencesuggests that Wg-induced slp activity inthe mesoderm is required but notsufficient to generate somatic muscleand heart progenitors. It appears that, inaddition to this route, wg is requiredindependently of slpfor the formation ofthese cell types.
A wg-dependent enhancerelement of slp contains essentialPangolin-binding sitesBecause previous genetic analysis hadimplicated 5′ flanking regions of theslp1genein the regulation of both slp1and slp2(Grossniklaus et al., 1992), we tested elements withinthis region for the presence of potential Wg responsiveenhancers (Fig. 5A). This analysis identified an element, slp5′-2, that activates reporter gene expression between stages 10 and11 in a transversely striped pattern, which is similar to the wg-dependent pattern of slpat these stages (Fig. 5B). A shorterversion, slp5′-1, produces an identical pattern but theexpression is much less robust (data not shown). As shown forendogenous slp, reporter gene expression with slp5′-2 occurredin register with Wg expression (Fig. 5C), and is observed inboth ectoderm and mesoderm (Fig. 7A). Moreover, the activityof this enhancer in striped domains is absolutely dependent onwg activity (Fig. 5E, compare with D). Together, the spatialand temporal expression, as well as its dependency on wg
activity suggest that slp5′-2 represents a Wg response elementthat mediates the activities of slpin both the mesoderm andectoderm.
A second enhancer, slp5′-4, is activated only from late stage11 onwards and drives reporter gene expression in transversestripes that are restricted to the ventral ectoderm (Fig. 5F). Thispattern represents part of the late pattern of endogenous slpexpression, which also becomes restricted to the ventralectoderm after stage 11 (Grossniklaus et al., 1992). As seen inFig. 5G, the slp5′-4 dependent stripes only partially overlappedwith the Wg stripes and extended more anteriorly, thusimplying that they are shifted slightly anteriorly as comparedwith the slp5′-2-dependent stripes. This shift was also apparentin comparison with the En stripes (Fig. 5H, compare with D).
Fig. 5. Wg-dependent enhancer elementsfrom the slplocus. (A) Summary ofenhancer elements identified in 5′flankingregions of slp1. (B-E) Embryos carryingslp5′-2/lacZ and (F-I), embryos carryingslp5′-4/lacZ insertions. (B) Stage 11 embryostained for β-gal (dorsal view). (C) Stage 10embryo stained for β-gal (green) and Wg(red), which shows coinciding Wg proteinand lacZexpression domains. (D) Stage 11embryo stained for β-gal (green) and En(red). lacZexpression abuts the En stripesanteriorly. (E) Stage 11 wgmutant embryostained as in D. Both the β-gal and Enstripes are missing. (F) Stage 11 embryostained as in B (dorsal view). (G) Stage 11embryo stained as in C. The β-gal stripesoverlap with the anterior portions of eachWg stripe and extend into more anteriorareas with low levels of detected Wg. (H) Stage 11 embryo stained as in D. Thereis an approx. 1-cell-wide gap between the β-gal and En stripes. (I) Stage 11 wgmutantembryo stained as in H. Both the β-gal andEn stripes are missing.

5504
As with slp5′-2 and endogenous slp in the neuroectoderm, theactivity of slp5′-4 was also dependent on wg in the head andtrunk region of the embryo (Fig. 5I). We presume that thisenhancer mediates the slp-dependent effects of wgon row 4and 5 neuroblast specification (Bhat et al., 2000). A thirdenhancer element, slp5′-3, was active in the prospectiveembryonic head (between ca. 70-90% egg length) from lateblastoderm until germ-band elongation, which reflects thepattern of endogenous slpduring its earliest stage of expressionin blastoderm embryos (data not shown; Grossniklaus et al.,1992).
Because the enhancer slp5′-2 appeared to be directlyrelevant for the regulation of mesoderm patterning by wg,further functional analysis was devoted to this particularelement. Pangolin, a nuclear HMG family protein, has been
shown to mediate Wg signal transduction (Brunner et al., 1997;Riese et al., 1997; van de Wetering et al., 1997). To assesswhether Pangolin could bind to the slp5′-2 enhancer, DNaseIfootprinting analysis with bacterially expressed GST-fusionproteins encoding the HMG domain of Pangolin wasperformed. A total of nine protected binding sites weredetected within the slp5′-2 element (Fig. 6A-D, and data notshown). Alignment of the sequences spanning the protectedregions showed that five of these sites (sites A, B, E, F and I)conformed to the consensus binding sequence of Pan (dTCF),as determined by binding site selection, and to the canonicalbinding motif of mammalian TCF (Fig. 6F, top; van deWetering et al., 1993, 1997). While four other sites (sites C, D,G and H) displayed only weak similarities with the canonicalTCF binding sequence, all of them contain a sequence motif
H.-H. Lee and M. Frasch
Fig. 6. In vitro DNA-binding experiments with Pangolin and wild-type or mutated slp5′-2 sequences. (A-D) DNase I footprinting analysis withincreasing amounts of Pan protein or no Pan (lanes ‘C’) as indicated. Binding experiments with wild-type sequences are shown on the left andexperiments with mutated sequences (see asterisks) on the right. (E) Gel mobility shift assays with Pan and oligonucleotides spanning bindingsite I. While a 500-fold excess of wild-type (wt) oligonucleotides competes for binding, an equal excess of oligos with mutated binding sitesdoes not. (F) Sequence alignments of protected Pan-binding sites within slp5′-2. The numbering defines the 5′ most genomic nucleotidematching the slp1 cDNA sequence (GenBank Accession Number X66095) as +1. rc, reverse complement.
5505Slp effects Wg signals in mesoderm and ectoderm
that conforms to the consensus binding sequence of HMGdomain proteins (Fig. 6F, bottom; Churchill et al., 1995Clevers and van de Wetering, 1997). We also noted that sitesA, B, C, E and F appeared to bind Pangolin with a higheraffinity than the remaining four sites (Fig. 6 and data notshown).
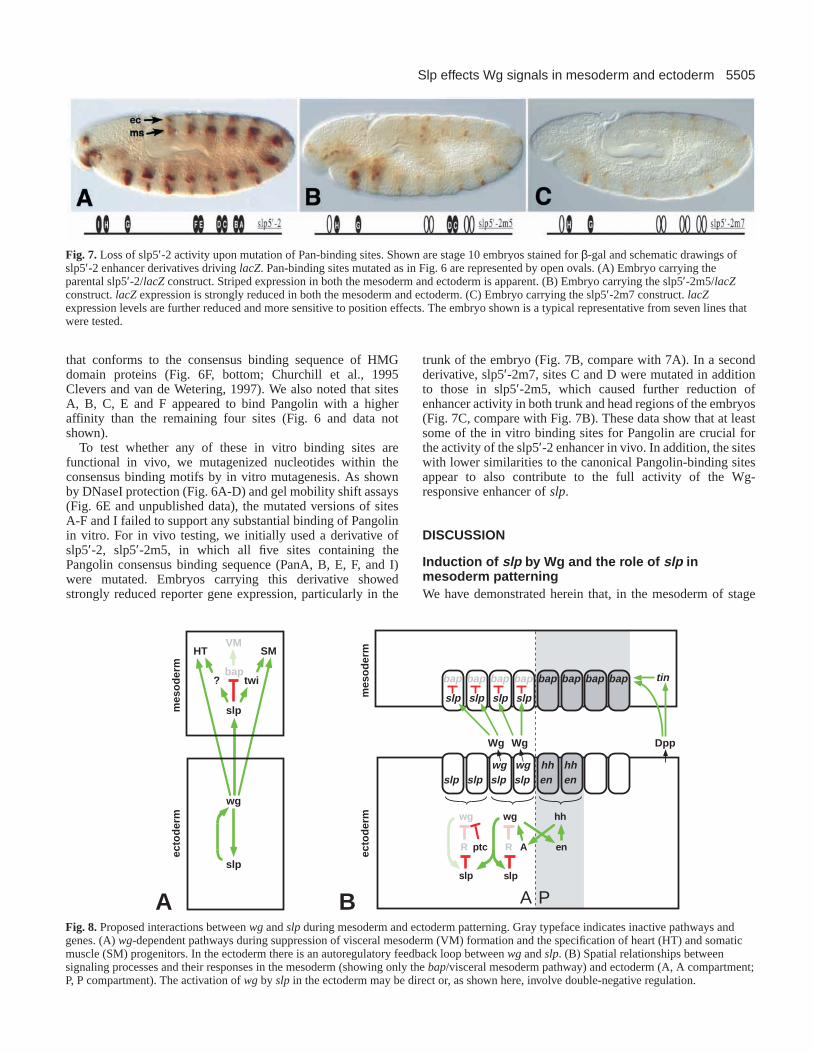
To test whether any of these in vitro binding sites arefunctional in vivo, we mutagenized nucleotides within theconsensus binding motifs by in vitro mutagenesis. As shownby DNaseI protection (Fig. 6A-D) and gel mobility shift assays(Fig. 6E and unpublished data), the mutated versions of sitesA-F and I failed to support any substantial binding of Pangolinin vitro. For in vivo testing, we initially used a derivative ofslp5′-2, slp5′-2m5, in which all five sites containing thePangolin consensus binding sequence (PanA, B, E, F, and I)were mutated. Embryos carrying this derivative showedstrongly reduced reporter gene expression, particularly in the
trunk of the embryo (Fig. 7B, compare with 7A). In a secondderivative, slp5′-2m7, sites C and D were mutated in additionto those in slp5′-2m5, which caused further reduction ofenhancer activity in both trunk and head regions of the embryos(Fig. 7C, compare with Fig. 7B). These data show that at leastsome of the in vitro binding sites for Pangolin are crucial forthe activity of the slp5′-2 enhancer in vivo. In addition, the siteswith lower similarities to the canonical Pangolin-binding sitesappear to also contribute to the full activity of the Wg-responsive enhancer of slp.
DISCUSSION
Induction of slp by Wg and the role of slp inmesoderm patterningWe have demonstrated herein that, in the mesoderm of stage
Fig. 7.Loss of slp5′-2 activity upon mutation of Pan-binding sites. Shown are stage 10 embryos stained for β-gal and schematic drawings ofslp5′-2 enhancer derivatives driving lacZ. Pan-binding sites mutated as in Fig. 6 are represented by open ovals. (A) Embryo carrying theparental slp5′-2/lacZconstruct. Striped expression in both the mesoderm and ectoderm is apparent. (B) Embryo carrying the slp5′-2m5/lacZconstruct. lacZexpression is strongly reduced in both the mesoderm and ectoderm. (C) Embryo carrying the slp5′-2m7 construct. lacZexpression levels are further reduced and more sensitive to position effects. The embryo shown is a typical representative from seven lines thatwere tested.
slp slp slp slp
bap bap bap bapbapbapbapbap
slp slp slp slpwg wg
Wg Wg
tin
Dpp
enhh hh
en
PA
hh
en
slp
wg
R
wg
ptc
slp
AR
wg
slp
baptwi
slp
VMSMHT
?
mes
od
erm
ecto
der
m
mes
od
erm
ecto
der
m
A BFig. 8.Proposed interactions between wgand slpduring mesoderm and ectoderm patterning. Gray typeface indicates inactive pathways andgenes. (A)wg-dependent pathways during suppression of visceral mesoderm (VM) formation and the specification of heart (HT) and somaticmuscle (SM) progenitors. In the ectoderm there is an autoregulatory feedback loop between wgand slp. (B) Spatial relationships betweensignaling processes and their responses in the mesoderm (showing only the bap/visceral mesoderm pathway) and ectoderm (A, A compartment;P, P compartment). The activation of wgby slpin the ectoderm may be direct or, as shown here, involve double-negative regulation.

5506
10-11 embryos, the slp gene is apparently a direct downstreamtarget of the Wg signaling cascade. We have also shown thatslp acts within the mesoderm to regulate the development ofvisceral mesoderm, somatic muscle and heart progenitors.These findings provide an alternative explanation for the closesimilarities between the wgand slp phenotypes in themesoderm, which were previously thought to be due mainly tothe loss of ectodermal wgexpression in slpmutants (Park etal., 1996; Riechmann et al., 1997; see Cadigan et al., 1994a).Analogous epistatic relationships between wg and slpduringspecification of certain neuroblasts have been recentlydescribed by Bhat et al. (2000).
During earlier stages, slpexpression is established andmaintained in the mesoderm (and ectoderm) by the pair-rulegenes, whose activity declines between stages 8 and 9 ofembryogenesis. The similarities of the mesodermal wgandslp phenotypes suggest that slp exerts its major functions inthe mesoderm during its wg-dependent phase of expression.However, the stronger effects of mutations of slp, whencompared with wg, on bapderepression indicate that the pair-rule dependent phase of slp expression, which is unaffectedin wg mutants, also provides some slpactivity for mesodermpatterning. In addition, it may be that the induction of slpexpression by Wg requires pre-existing Slp protein as acoregulator, which would imply that pair-rule gene regulationis required although not sufficient for mesodermal slpfunction. Hence, slpcould be a candidate for one of the genes,‘N’, which have been proposed to be activated by pair-rulegenes in striped domains within the mesodermal germ layerand to provide the mesoderm with a segmental prepattern(Azpiazu et al., 1996; see also Fig. 4 by Riechmann et al.,1997).
A model of how this newly-defined pathway may controlbap expression and trunk visceral mesoderm development isshown in Fig. 8A,B. In its simplest version, synergisticactivities of Dpp and Tinman would be sufficient for bapactivation in the entire dorsal mesoderm of the embryonictrunk. However, segmentally induced slp would abrogate theactivities of Dpp/Tinman in a periodic fashion and thus restrictbap, and consequently visceral mesoderm specification, tometameric domains within the dorsal mesoderm. Slp may actas a repressor by direct binding to regulatory elements of thebap gene during this process. This proposed mechanism isconsistent with our findings that a functional bap enhancerelement contains an in vitro binding site for Slp (H.-H. L. andM. F., unpublished observations). In addition, we have noticedconserved sequence motifs in the N-terminal portions of bothSlp1 and Slp2 that are related to previously defined repressiondomains from the Engrailed protein (Slp1, KSSFSINSILP;Slp2, KSNFSIDAIL A; underlining indicates identity with En)(Smith and Jaynes, 1996).
While wg regulates visceral mesoderm development in anegative manner and strictly through slp, its role in the positiveregulation of somatic muscle and heart development is morecomplex. We have shown that induction of slp by Wg in themesoderm is required, but not sufficient, for mediating thesewgactivities. This could be due to a two-step process, in whichWg-induced slp provides the cells in the mesodermal slpdomains with the competence to respond to subsequent Wgsignals. During the second step, Wg signals could then requireSlp protein, or a Slp-dependent factor such as Twist (Fig. 8A;
Baylies and Bate, 1996; Riechmann et al., 1997), as acoregulator to induce the expression of genes such as eve, slou,nau or mef2 in heart and somatic muscle progenitors,respectively. These latter pathways seem to imply that Slp canact not only as a transcriptional repressor, but also as anactivator and that perhaps different promoter-specific cofactorsswitch its activity from a repressor to an activator. However,we cannot exclude the possibility that Slp is strictly repressingand acts by negatively regulating repressors of twist and othersomatic muscle- and heart-specification genes.
As indicated in Fig. 8B, wg is expressed in two cell-widestripes in the ectoderm at the relevant stage, but the diffusibleWg protein is able to induce slpin approximately four cell-wide stripes in both germ layers. Similar to the situation in theectoderm, Wg activity in the mesoderm is polarized, since itdoes not induce slp across the parasegment boundary. In part,this restriction could be due to an additional requirement forSlp protein, which is limited to domains that are initiallydefined by pair-rule gene activities, as discussed above.Moreover, Hh signaling from the ectodermal P compartmentsappears to limit the range of Wg signaling in the mesoderm(Fig. 2G,H; Azpiazu et al., 1996). As recently shown bySanson et al. (1999), a similar Hh-dependent block of Wgsignaling in the ectoderm involves a reduction of the transportand/or stability of the Wg protein. Alternatively, Hh signalingor its targets could interfere with Wg signal transduction inthese cells.
Autoregulatory loop between wg and slp in the Acompartments of the ectodermPrevious studies have shown that, in the ectodermal Acompartments, wg expression requires slpactivity (Cadigan etal., 1994a), as well as autocrine Wg signaling (Hooper, 1994;Yoffe et al., 1995). Our present data show that both of theseprocesses are in fact part of the same feedback loop, in whichWg maintains slpexpression and Slp is required to maintainwgexpression. The activation of wgby slpcould involve directbinding of Slp to wg enhancer elements. However, because ofthe presumed activity of Slp as a transcriptional repressor (seeabove) and observations that it can downregulate severalsegmentation genes (Cadigan et al., 1994a) as well as bap, analternative mode of action would be that slp represses anegative regulator of wg, as indicated in Fig. 8B (bottom).Because en is expanded in slp mutants (Cadigan et al., 1994a)and its encoded protein has properties of a transcriptionalrepressor (Smith and Jaynes, 1996, and this study), En may bea candidate for ‘R’ in this proposed double-negative regulationof wg (Fig. 8)
The feedback loop between wg and slpis active in only thetwo cells anterior to the parasegment border, which receive Hhsignals. By contrast, in their more anterior neighbors, it isrendered inactive by Ptc activity, which prevents wgexpression(Cadigan et al., 1994a), although slp can still be induced byparacrine Wg signals. Based upon our current understandingof the functions of the nuclear Hh effector Ci (Aza-Blanc andKornberg, 1999), it appears likely that an activating form of Ci(CiAct) is involved in the Hh-dependent promotion of thisfeedback loop and a repressing form (CiR) in its Ptc-dependentinterruption. Because a Hh/Ptc-responsive wgenhancer lacksany recognizable Ci-binding motifs (Lessing and Nusse, 1998),it is plausible that CiAct and CiR do not act directly upon wg
H.-H. Lee and M. Frasch

5507Slp effects Wg signals in mesoderm and ectoderm
but perhaps on yet undefined component(s) that are needed forwg activation (‘A’, Fig. 8) and repression.
Mechanistic aspects of slp regulationBased upon the breakpoints and phenotypes of several largedeletions in the slplocus, Grossniklaus et al. (1992) haveconcluded that the 5′flanking regions contain essentialregulatory elements that are shared between the slp1and slp2genes. Indeed, our present reporter gene analysis has identifiedthree separate enhancer elements within approx. 5.5 kb ofupstream sequences, each of which reproduces one particularaspect of endogenous slp expression. The two distal elementsdrive head-specific expression during blastoderm stages andstriped expression in the ventral ectoderm following stage 11,respectively. The third, more proximal element is active in boththe mesoderm and ectoderm between stages 10 and 11,coinciding with the period when mesodermal genes such asbap and eveare activated. Its spatial and temporal profile, aswell its wgdependency, are fully consistent with the notion thatthis enhancer is targeted by the Wg signaling cascade duringmesoderm induction and autocrine signaling in the ectoderm.Because it is not active during the period when the pair-rulegenes are expressed and is turned on in every parasegment,without displaying any transient pair-rule pattern, this enhancerappears to be exclusively responsive to Wg in terms of itsspatial regulation. Hence, there must be separate pair-ruleenhancer(s), which were not uncovered by our dissection. Theobservation that this Wg response element becomes rapidlyinactive at late stage 11, although wgcontinues to be expressed,indicates that it requires at least one additional, temporallyrestricted regulator in conjunction with the Wg signal.
Ubx is one of the few genes known to be a direct target ofWingless, although there are indications that enmay be anotherexample (Riese et al., 1997; Florence et al., 1997). An enhancerelement of Ubx, which is activated by Wg after the formationof the midgut visceral mesoderm, contains at least twofunctionally important Pan-binding sites (Yu et al., 1998). Inour present study, we have determined that a Wg-responsiveenhancer of slpcontains nine in vitro Pan-binding sites. We donot know whether all of these sites are functional because wehave not yet performed in vivo tests of mutations of individualsites or various combinations of them. However, our currentdata from two rounds of mutagenesis show that some of themare indeed functional in vivo. Our analysis also indicates thatthe activities of different functional binding sites are additiveand therefore partially redundant.
Although most of the known wg downstream genes areupregulated by Wg signals, there are several examples ofnegatively regulated genes, in addition to bap. For instance, theUbx midgut enhancer is downregulated by high levels of Wgsignaling through a mechanism that involves Smad-bindingsites. It has been speculated that Wg signals may induce thetranscription of an unknown repressor that competes withMad/Medea binding to these sites (Yu et al., 1998). Bycontrast, the repression of shavenbaby(ovo – FlyBase) in theepidermis of late stage embryos by Wg signaling (Payre et al.,1999) has been proposed to involve the direct binding ofhypothetical Pangolin/Armadillo/corepressor complexes toovo regulatory elements (Bejsovec, 1999). Our present studyprovides strong evidence for an indirect mode of baprepression by Wg that is more comparable to the one proposed
for Ubx. It appears that during this pathway, Wg signals inducethe transcription of slp, whose encoded protein productprobably acts as a repressor that interferes with Dpp/Tinmanmediated activation of baptranscription.
We thank Gary Struhl for generously providing the UAS-slp lineused in this study. We also thank Dave Kosman, John Reinitz, MariannBienz and the Developmental Studies Hybridoma Bank (supported byNICHD and University of Iowa) for antibodies and plasmids, andHanh Nguyen for comments on the manuscript. Confocal laserscanning microscopy was performed at the MSSM-MicroscopyCenter, supported with funding from NIH (1 S10 RR0 9145-01) andNSF (DBI-9724504). We acknowledge the funding of this research byNIH (HD 30832).
REFERENCES
Aza-Blanc, P. and Kornberg, T. B.(1999). Ci: a complex transducer of theHedgehog signal. Trends Genet.15, 458-462.
Azpiazu, N. and Frasch, M.(1993). tinmanand bagpipe: two homeoboxgenes that determine cell fates in the dorsal mesoderm of Drosophila. GenesDev.7, 1325-1340.
Azpiazu, N., Lawrence, P., Vincent, J.-P. and Frasch, M.(1996).Segmentation and specification of the Drosophila mesoderm. Genes Dev.10, 3183-3194.
Baker, N. E.(1987). Molecular cloning of sequences from wingless, a segmentpolarity gene in Drosophila: the spatial distribution of a transcript inembryos. EMBO J.6, 1765-1773.
Baylies, M., Martinez Arias, A. and Bate, M.(1995). winglessis requiredfor the formation of a subset of muscle founder cells during Drosophilaembryogenesis. Development121, 3829-3837.
Baylies, M. K. and Bate, M.(1996). twist: a myogenic switch in Drosophila.Science272, 1481-1484.
Bejsovec, A.(1999). Wnt signalling shows its versatility. Curr. Biol. 9, R684-R687.
Bhat, K. M., van Beers, E. H. and Bhat, P.(2000). sloppy pairedacts as thedownstream target of Wingless in the Drosophila CNS and interactionbetween sloppy paired and gooseberryinhibits sloppy pairedduringneurogenesis. Development127, 655-665.
Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a meansof altering cell fates and generating dominant phenotypes. Development 118,401-415.
Brunner, E., Peter, O., Schweizer, L. and Basler, K.(1997). pangolinencodes a Lef-1 homologue that acts downstream of Armadillo to transducethe Wingless signal in Drosophila. Nature385, 829-833.
Cadigan, K., Grossniklaus, U. and Gehring, W. (1994a). Localizedexpression of sloppy pairedprotein maintains the polarity of Drosophilaparasegments. Development120, 899-913.
Cadigan, K. M., Grossniklaus, U. and Gehring, W. J. (1994b).Functional redundancy: the respective roles of the two sloppy pairedgenes in Drosophilasegmentation. Proc. Natl. Acad. Sci. USA91, 6324-6328.
Carmena, A., Gisselbrecht, S., Harrison, J., Jimenez, F. and Michelson, A.(1998). Combinatorial signaling codes for the progressive determination ofcell fates in the Drosophilaembryonic mesoderm. Genes Dev.15, 3910-3922.
Churchill, M. E., Jones, D. N., Glaser, T., Hefner, H., Searles, M. A. andTravers, A. A. (1995). HMG-D is an architecture-specific protein thatpreferentially binds to DNA containing the dinucleotide TG. EMBO J.14,1264-1275.
Clevers, H. and van de Wetering, M.(1997). TCF/LEF factor earn theirwings. Trends Genet.13, 485-489.
D’Alessio, M. and Frasch, M. (1996). mshmay play a conserved role indorsoventral patterning of the neuroectoderm and mesoderm. Mech. Dev.58,217-231.
Dunin-Borkowski, O., Brown, N. and Bate, M. (1995). Anterior-posteriorsubdivision and the diversification of the mesoderm in Drosophila.Development 121, 4183-4193.
Florence, B., Guichet, A., Ephrussi, A. and Laughon, A.(1997). Ftz-F1 isa cofactor in Ftz activation of the Drosophila engrailedgene. Development124, 839-847.

5508
Frasch, M. (1995). Induction of visceral and cardiac mesoderm by ectodermalDpp in the early Drosophilaembryo. Nature374, 464-467.
Frasch, M. (1999). Intersecting signalling and transcriptional pathways inDrosophilaheart specification. Semin. Cell Dev. Biol.10, 61-71.
Frasch, M. and Nguyen, H. T. (1999). Genetic control of mesodermpatterning and differentiation during Drosophilaembryogenesis. Adv. Dev.Biochem.5, 1-47.
Frasch, M., Hoey, T., Rushlow, C., Doyle, H. J. and Levine, M. (1987).Characterization and localization of the even-skippedprotein of Drosophila.EMBO J. 6, 749-759.
Greig, S. and Akam, M. (1993). Homeotic genes autonomously specifyone aspect of pattern in the Drosophilamesoderm. Nature 362, 630-632.
Grossniklaus, U., Pearson, R. and Gehring, W.(1992). The Drosophilasloppy pairedlocus encodes two proteins involved in segmentation thatshow homology to mammalian transcription factors. Genes Dev.6, 1030-1051.
Guillen, I., Mullor, J. L., Capdevila, J., Sanchez-Herrero, E., Morata, G.and Guerrero, I. (1995). The function of engrailedand the specification ofDrosophilawing pattern. Development121, 3447-3456.
Hooper, J. (1994). Distinct pathways for autocrine and paracrine Winglesssignalling in Drosophilaembryos. Nature372, 461-464.
Jagla, K., Frasch, M., Jagla, T., Dretzen, G., Bellard, F. and Bellard, M.(1997). ladybird, a new component of the cardiogenic pathway inDrosophila required for diversification of heart precursors. Development124, 3471-3479.
Keller, C. A., Grill, M. A. and Abmayr, S. M. (1998). A role for nautilusinthe differentiation of muscle precursors. Dev. Biol.202, 157-171.
Knirr, S., Azpiazu, N. and Frasch, M.(1999). The role of the NK-homeoboxgene slouch(S59) in somatic muscle patterning. Development126, 4525-4535.
Kosman, D., Small, S. and Reinitz, J. (1998). Rapid preparation of a panelof polyclonal antibodies to Drosophilasegmentation proteins. Dev. GenesEvol. 208, 290-294.
Lawrence, P., Bodmer, R. and Vincent, J.(1995). Segmental patterning ofheart precursors in Drosophila. Development121, 4303-4308.
Lessing, D. and Nusse, R.(1998). Expression of winglessin the Drosophilaembryo: a conserved cis-acting element lacking conserved Ci-binding sitesis required for patched-mediated repression. Development125, 1469-1476.
Nose, A., Isshiki, T. and Takeichi, M.(1998). Regional specification ofmuscle progenitors in Drosophila: the role of the mshhomeobox gene.Development125, 215-223.
Nüsslein-Volhard, C., Wieschaus, E. and Kluding, H.(1984). Mutationsaffecting the pattern of the larval cuticle in Drosophila melanogasterI.Zygotic loci on the second chromosome. Roux’s Arch. Dev. Biol.193, 267-282.
Park, M., Wu, X., Golden, K., Axelrod, J. D. and Bodmer, R.(1996). TheWingless signaling pathway is directly involved in Drosophila heartdevelopment. Dev. Biol.177, 104-116.
Payre, F., Vincent, A. and Carreno, S.(1999). ovo/svb integrates Wingless
and DER pathways to control epidermis differentiation. Nature 400, 271-275.
Ranganayakulu, G., Schulz, R. A. and Olson, E. N.(1996). winglesssignaling induces nautilus expression in the ventral mesoderm of theDrosophilaembryo. Dev. Biol.176, 143-148.
Riechmann, V., Irion, U., Wilson, R., Grosskortenhaus, A. and Leptin, M.(1997). Control of cell fates and segmentation in the Drosophilamesoderm.Development124, 2915-2922.
Riese, J., Yu, X., Munnerlyn, A., Eresh, S., Hsu, S., Grosschedl, R. andBienz, M. (1997). LEF-1, a nuclear factor coordinating signaling inputsfrom winglessand decapentaplegic. Cell 88, 777-787.
Sanson, B., Alexandre, C., Fascetti, N. and Vincent, J. P.(1999). engrailedand hedgehogmake the range of Wingless asymmetric in Drosophilaembryos. Cell 98, 207-216.
Smith, S. T. and Jaynes, J. B.(1996). A conserved region of engrailed, sharedamong all en-, gsc-, Nk1-, Nk2- and msh-classhomeoproteins, mediatesactive transcriptional repression in vivo. Development122, 3141-3150.
Staehling-Hampton, K., Hoffmann, F. M., Baylies, M. K., Rushton, E. andBate, M. (1994). dppinduces mesodermal gene expression in Drosophila.Nature372, 783-786.
Su, M., Fujioka, M., Goto, T. and Bodmer, R.(1999). The Drosophilahomeobox genes zfh-1 and even-skippedare required for cardiac-specificdifferentiation of a numb-dependent lineage decision. Development126,3241-3251.
van de Wetering, M., Cavallo, R., Dooijes, D., van Beest, M., van Es, J.,Loureiro, J., Ypma, A., Hursh, D., Jones, T., Bejsovec, A. et al.(1997).Armadillo coactivates transcription driven by the product of the Drosophilasegment polarity gene dTCF. Cell 88, 789-799.
van de Wetering, M., Oosterwegel, M., van Norren, K. and Clevers, H.(1993). Sox-4, an Sry-like HMG box protein, is a transcriptional activatorin lymphocytes. EMBO J.12, 3847-3854.
van den Heuvel, M., Nusse, R., Johnston, P. and Lawrence, P.(1989).Distribution of the winglessgene product in Drosophilaembryos; a proteininvolved in cell-cell communication. Cell59, 739-749.
Waltzer, L. and Bienz, M.(1998). DrosophilaCBP represses the transcriptionfactor TCF to antagonize Wingless signalling. Nature395, 521-525.
Wu, X., Golden, K. and Bodmer, R. (1995). Heart development inDrosophilarequires the segment polarity gene wingless. Dev. Biol.169, 619-628.
Xu, X., Yin, Z., Hudson, J., Ferguson, E. and Frasch, M.(1998). Smadproteins act in combination with synergistic and antagonistic regulators totarget Dpp responses to the Drosophilamesoderm. Genes Dev.12, 2354-2370.
Yin, Z., Xu, X.-L. and Frasch, M. (1997). Regulation of the Twist target genetinman by modular cis-regulatory elements during early mesodermdevelopment. Development 124, 4971-4982.
Yoffe, K. B., Manoukian, A. S., Wilder, E. L., Brand, A. H. and Perrimon,N. (1995). Evidence for engrailed-independent winglessautoregulation inDrosophila. Dev. Biol.170, 636-650.
Yu, X., Riese, J., Eresh, S. and Bienz, M. (1998). Transcriptional repressiondue to high levels of Wingless signalling. EMBO J.17, 7021-7032.
H.-H. Lee and M. Frasch





























