Embed Size (px)
Citation preview

DEPARTMENT OF PRIMARY INDUSTRIES
Wheat spindle streak mosaic virus
Diagnostic Manual

Wheat spindle streak mosaic virus Diagnostic Manual
2
Wheat spindle streak mosaic virus
Diagnostic Manual
Bonny Rowles-van Rijswijk
Dr James Cunnington
Dr Brendan Rodoni
Department of Primary Industries, Knoxfield
Private Mail Bag 15
Ferntree Gully Delivery Centre
Victoria 3156
Australia
Published by Primary Industries Research Victoria
Department of Primary Industries
Knoxfield
June 2005
© Copyright State of Victoria 2005
This publication is copyright. No part may be reproduced by any process except in
accordance with the provisions of the Copyright Act 1968.
Authorised by the Victorian Government, 1 Treasury Place, East Melbourne.
ISBN 1 74146 397 1

Wheat spindle streak mosaic virus Diagnostic Manual
3
This publication may be of assistance to you but the State of Victoria and its
employees do not guarantee that the publication is without flaw of any kind or is
wholly appropriate for your particular purposes and therefore disclaims all liability for
any error, loss or other consequence which may arise from you relying on any
information in this publication.
For more information about DPI visit the website at www.dpi.vic.gov.au
or call the Customer Service Centre on 136 186.

Wheat spindle streak mosaic virus Diagnostic Manual
4
Table of Contents
1. Introduction _________________________________________________ 6
2. Wheat spindle streak mosaic virus _______________________________ 7
2.1. Distribution and yield loss _______________________________________ 7
2.2. Wheat spindle streak mosaic virus description _______________________ 7
2.3. Host range of Wheat spindle streak mosaic virus ____________________ 10
2.4. Symptoms associated with Wheat spindle streak mosaic virus __________ 10
2.5. Diseases associated with causing symptoms similar to Wheat spindle streak
mosaic virus____________________________________________________ 15
2.5.1. Soil-borne wheat mosaic virus _______________________________________ 15
2.5.2. Wheat streak mosaic virus __________________________________________ 18
2.6. Management of Wheat spindle streak mosaic virus __________________ 19
3. Polymyxa graminis __________________________________________ 20
3.1. The impact of infection by the fungus, Polymyxa graminis _____________ 20
3.2. Description of Polymyxa graminis ________________________________ 21
3.3. Life cycle of Polymyxa graminis _________________________________ 24
3.3. Polymyxa graminis fungal isolation _______________________________ 30
3.4. Transmission of Wheat spindle streak mosaic virus by Polymyxa graminis _ 31
3.5. Management strategies for Polymyxa graminis ______________________ 33
4. Detection of Wheat spindle streak mosaic virus ____________________ 34
4.1. Diagnostic Flow-chart _________________________________________ 34
4.2. Sample collection ____________________________________________ 35
4.2.1. Plant ___________________________________________________________ 35
4.2.2. Soil ____________________________________________________________ 35
4.3. Total RNA extraction __________________________________________ 36
4.3.1. Equipment required _______________________________________________ 36
4.3.2. Reagents required ________________________________________________ 37
4.3.3. Method _________________________________________________________ 37
4.4. Detection of Wheat spindle streak mosaic virus in total RNA extracts using one
step RT-PCR ___________________________________________________ 39
4.4.1. Equipment required _______________________________________________ 39

Wheat spindle streak mosaic virus Diagnostic Manual
5
4.4.2. Reagents _______________________________________________________ 40
4.4.3. One-step RT-PCR detection of Wheat spindle streak mosaic virus___________ 42
4.5. DNA Sequencing of PCR Products _______________________________ 43
4.5.1. Equipment required _______________________________________________ 43
4.5.2. Reagents _______________________________________________________ 43
4.5.3. Method _________________________________________________________ 43
4.6. Detection of Wheat spindle streak mosaic virus with Enzyme-linked
immunosorbent assay (ELISA) _____________________________________ 44
4.6.1. Equipment_______________________________________________________ 44
4.6.2. Reagents _______________________________________________________ 44
4.6.3. Biological Reagents _______________________________________________ 45
4.6.4. Method _________________________________________________________ 45
5. Detection of Polymyxa graminis ________________________________ 47
5.1. Diagnostic Flow-chart _________________________________________ 47
5.2. Sample collection ____________________________________________ 48
5.2.1. Plant ___________________________________________________________ 48
5.2.2. Soil ____________________________________________________________ 48
5.3. Total DNA extraction __________________________________________ 49
5.3.1. Equipment required _______________________________________________ 49
5.3.2. Method _________________________________________________________ 49
5.4. Detection of Polymyxa graminis in total DNA extracts using PCR. _______ 50
5.4.1. Equipment required _______________________________________________ 50
5.4.2. Reagents _______________________________________________________ 51
5.4.3. Method _________________________________________________________ 52
5.5. DNA Sequencing of PCR Products _______________________________ 52
Acknowledgments _____________________________________________ 53
References __________________________________________________ 54
Related Articles _______________________________________________ 59
Appendix 1 – ELISA sample plans ________________________________ 61

Wheat spindle streak mosaic virus Diagnostic Manual
6
1. Introduction
The cost to the grains industry by incursions of exotic pests and pathogens has been
recognised by Plant Health Australia and the Department of Primary Industries,
Victoria. In preparation for potential incursions, it was decided to develop a
diagnostic capability for Wheat spindle streak mosaic virus (WSSMV), as this viral
disease has not been reported in Australia and poses a significant threat to the
grains industry. WSSMV outbreaks are favoured by cool climates (Cadle-Davidson
and Bergstrom, 2004), and it is feasible that this virus could establish under southern
Australian conditions.
WSSMV was first reported in 1957, in Ontario, Canada, and is now ranked amongst
the three most common viral diseases of wheat in this region (Tenuta and Johnson,
2003). WSSMV is transmitted by the soil-borne fungus, Polymyxa graminis
Ledingham (OMAF Staff, 2002). There is only one record of a “P. graminis-like”
fungus in Australia, that was lodged on 8 September 1959, in Murrumburrah, NSW,
on Poa annua L. (DAR 48987) (APPD, 2005). It must be noted that the identification
was not to subspecies level, and that this record has never been confirmed.
P. graminis is non-pathogenic (Kanyuka et al., 2003), but its the ability to acquire and
transmit a range of plant viruses via zoospores (swimming spores) that invade root
hairs and epidermal cells of young plants during periods of high soil moisture that
results in serious diseases in cereal crop species (Kanyuka et al., 2003; OMAF Staff,
2002). The fungus can remain in the soil for at least 8 years (OMAF Staff, 2002),
with viral particles protected from the environment within resting spores (Kanyuka et
al., 2003). The persistent, soil-borne nature of the WSSMV-vector P. graminis,
makes the use of virus-resistant crop varieties the only practical and environmentally
friendly means of control (Kanyuka et al., 2003).
As the virus and vector are so intrinsically linked, this diagnostic protocol will deal
with methods to detect both WSSMV and P. graminis. The sampling and detection
methods in this manual have not yet been trialed under Australian conditions. It is
highly recommended that the methods be validated prior to establishment as a
national standard.

Wheat spindle streak mosaic virus Diagnostic Manual
7
2. Wheat spindle streak mosaic virus
2.1. Distribution and yield loss
WSSMV in France, Germany, India, Italy, Japan, North America, and possibly China
(Brunt et al., 1996 onwards; Jones, 2004).
Losses from WSSMV are greatest when a sensitive wheat variety is grown, and
when fields have had several crops of winter wheat in the past 8-10 years, resulting
in the contamination of soil over the entire paddock with the fungal vector P. graminis
(Jones, 2004; OMAF Staff, 2002; Tenuta and Johnson, 2003). Wheat yield losses
are attributed to a reduction in the numbers of tillers produced (Bowden, 2005).
Losses are dependent on the wheat variety and the weather, and are difficult to
measure. A long cool spring prolongs the symptomatic period and increases losses,
as the optimum temperature for symptom development is 5°C-15°C (Bowden, 2005;
OMAF Staff, 2002; Tenuta and Johnson, 2003). Alternatively, in a warm spring,
losses may be minimal (Bowden, 2005). Data from North America shows the
variability in yield loss due to WSSMV infection; in 2004 soft white winter wheat
losses were reported to be 2-18% in Michigan state, and 7-59% in Canada. In
Pennsylvania and Georgia, losses were estimated to be 7-24% and 22-35%,
respectively (Bowden, 2005).
2.2. Wheat spindle streak mosaic virus description
WSSMV is a member of the genus Bymovirus, family Potyviridae. The virus consists
of single stranded positive sense RNA encapsulated by a protein coat (Brunt et al.,
1996 onwards). The long flexuous filamentous WSSMV particles (fig. 1) are 16 nm
wide, with no clear modal length. Measurements range from 300-2000 nm, with the
modal length probably around 700-1000 nm (Brunt et al., 1996 onwards; Jones,
2004; Lommel et al., 1986). Virions are found in the cytoplasm of leaves, roots,
mesophyll and vascular parenchyma of WSSMV-infected plants, and distinctive
pinwheel inclusion bodies are produced (Lommel et al., 1986). Infection by WSSMV
causes the chloroplasts and mitochondria to swell, resulting in disruption of these
organelles (Brunt et al., 1996).

Wheat spindle streak mosaic virus Diagnostic Manual
8
Figure 1. Electron micrograph of Wheat spindle streak mosaic virus. Par t icles t rapped
by an t iserum f rom in f ect ed leaves. Lengt h of bar [nm ]: 100 (Adam s, 2002).
WSSMV is transmitted by the obligate fungus P. graminis (Barr, 1979). The
subspecies of P. graminis that transmits WSSMV requires 3 hours at 15oC for
attachment and host penetration to occur, and WSSMV transmission has been
reported to occur within 16 days at temperatures ranging from 7.5 to 17.5oC (Cadle-
Davidson and Bergstrom, 2004). Environmental fluctuation through thermoperiodism
between 5 to 15oC (night-day) stimulates WSSMV transmission (Cadle-Davidson and
Bergstrom, 2004). The virus is not seed transmitted (Jones, 2004), nor is it
transmitted by grafting, pollen, or contact between plants (Brunt et al., 1996
onwards).
In an infected plant, Carrol et al. (1997) detected WSSMV in roots one month prior to
its detection in leaves, suggesting that virus moves slowly from roots to leaves.
WSSMV was detected in non-symptomatic leaves from two months after sowing,
through to crop senescence, and it was concluded that WSSMV is acquired and
spread by the vector during the majority of the crop cycle (Carrol et al., 1997).
There has been much confusion surrounding the relatedness of WSSMV to Wheat
yellow mosaic virus (WYMV), and how to distinguish the two viruses. Yellow mosaic
symptoms in wheat were first recorded in Japan, in 1927, and were attributed to
WYMV (Namba et al., 1998). In Canada similar symptoms in wheat were attributed
to WSSMV, and in the USA, India, France, Italy and China the causal agent has
been labelled either WYMV or WSSMV (Chen et al., 1999; Clover and Henry, 1999;

Wheat spindle streak mosaic virus Diagnostic Manual
9
Namba et al., 1998). A bymovirus occurring in rye, in Germany, has been described
as WSSMV or WYMV (Chen et al., 1999).
In 1979, Usugi and Saito drew the conclusion that WSSMV was a strain of WYMV
due to similarities in morphology, virus particle length, geographic distribution,
buoyant density, stability in sap, and because the two viruses had common antigens
in serological studies. However, there were slight differences in the symptoms
produced on infected wheat varieties, and not all serological antigens were common
to both viruses (Usugi and Saito, 1979). Hariri et al. (1996) also concluded that
WSSMV and WYMV were different strains of the same virus because of their strong
serological relationship. Carroll et al. (1995) developed a polyclonal antiserum that
reacted with bymovirus isolates of WSSMV, WYMV, and Barley yellow mosaic virus.
The International Committee on Taxonomy of Viruses website
(http://image.fs.uidaho.edu/vide/descr887.htm) continues to list WYMV as a strain of
WSSMV (Brunt et al., 1996 onwards).
The discrimination between WSSMV and WYMV is important as WSSMV has a
broader host range than WYMV, infecting Triticum aestivum and T. durum, as well as
Secale cereale. WSSMV infection has also been reported to breakdown field
resistance to Soil-borne wheat mosaic virus (SBWMV) in wheat (Clover and Henry,
1999; Lommel et al., 1986).
After generating sequence data from two fragments amplified from WYMV and
WSSMV (corresponding to RNA1 (7.6 kb) and RNA2 (3.6 kb)), Namba et al. (1998)
deduced that the two viruses shared only 77% amino acid sequence identity in their
coat proteins, and 74% nucleotide sequence identity in their 3’ non-coding regions.
This result had not been expected because of their close serological relationship, but
clearly suggests that WYMV and WSSMV are distinct virus species. This
discrimination was supported by sequencing of a 1.7 kb fragment representing the 3’-
terminal sequence of RNA1 of a Canadian WSSMV isolate, as this sequence shared
98.0% homology with the French WSSMV isolate used by Namba et al. (1998) (Lu et
al., 1998). Further analysis by Chen et al. (1999) was able to divide the isolates into
two distinct groups, i) WSSMV, for North American and European isolates, and ii)
WYMV for Asian isolates; based on differences in nucleotide between the groups of
over 30%. These findings were confirmed by Clover and Henry (1999), after analysis
of a further 23 viral sequences. As there was no serological method able to
discriminate between WSSMV and WYMV and no molecular diagnostic protocols

Wheat spindle streak mosaic virus Diagnostic Manual
10
available for either virus, Clover and Henry (1999) developed a WSSMV-specific RT-
PCR protocol. The WSSMV-specific RT-PCRs developed by Clover and Henry
(1999) and Gitton et al. (1999) form the basis of the diagnostic protocol
recommended for the detection of WSSMV (section 4).
2.3. Host range of Wheat spindle streak mosaic
virus
WSSMV infects wheat (T. aestivum, T. durum), which is used to maintain and
propagate the virus for experimentation, and rye. WSSMV does not infect barley
(Hordeum vulgare), nor any dicotyledonous species (Brunt et al., 1996 onwards;
Jones, 2004).
2.4. Symptoms associated with Wheat spindle
streak mosaic virus
WSSMV transmission occurs primarily during autumn and to a lesser extent in spring
(Carrol et al., 1997), with significant infections taking place during cool, wet autumn
periods. Often large areas of a field may be affected (Stromberg, 2002) (fig. 2 and
3), due to the uniform distribution of P. graminis, and WSSMV infection sites tend to
follow the lower, wetter areas of the field (Bowden, 2005).
Symptoms only appear in early spring, right after green-up (Bowden, 2005),
regardless of when plants have become infected (Carrol et al., 1997). However, the
earlier in the life cycle that the wheat is infected, the more severe the symptom
expression (Stromberg, 2002). The virus requires temperatures below 17oC for
symptom development (Lommel et al., 1986). The optimum temperature for
symptom development is 10oC (Smith, 2005), but symptoms will express between
9°C and 13°C air temperature (Stromberg, 2002), and 5 to 13oC soil temperature
(Smith, 2005). If soil temperatures exceed 15oC infected plants will fail to develop
mosaic symptoms, leaving little visual evidence of the disease (Carrol et al., 1997;
Smith, 2005), although symptoms may persist on older leaves (Bowden, 2005).
In favourable conditions during early spring, the symptoms of WSSMV appear as
yellow to light green streaks or dashes on a dark green background, in young leaves.

Wheat spindle streak mosaic virus Diagnostic Manual
11
The discontinuous streaks are usually 3 to 6.5 mm long, and oriented parallel to leaf
veins and taper at the ends to form chlorotic spindles (Bowden, 2005; Smith, 2005;
Tenuta and Johnson, 2003) (fig. 4 to 7). WSSMV is most easily diagnosed by
symptoms when the chlorotic spindle shaped lesions appear on the upper leaves of
the plant prior to heading (Smith, 2005).
During cool spring conditions, and as plants mature, the spindle-shaped lesions and
the chlorotic streaks on older leaves coalesce and become necrotic (Smith, 2005;
Stromberg, 2002). The older leaves look mottled with yellow and green blotches. If
temperatures remain cool, spindle shaped lesions will continue to develop throughout
spring in the upper new leaves (Smith, 2005). Warmer temperatures may prevent
symptom development on newly expanded leaves, giving the appearance that plants
have outgrown the disease, with symptoms confined to older, lower leaves (Smith,
2005).
WSSMV can also cause stunting, reduced tillering, smaller heads and less seed per
head than healthy plants, although the kernels produced by infected plants are not
shrivelled or light in weight (Bowden, 2005; OMAF Staff, 2002; Smith, 2005;
Stromberg, 2002; Tenuta and Johnson, 2003) (fig. 8 and 9).

Wheat spindle streak mosaic virus Diagnostic Manual
12
Figure 2. Infection by Wheat spindle streak mosaic virus is usually uniform in a wheat field,
because of the uniform distribution of the fungal vector, Polymyxa graminis (Source: http://www.uky.edu/Agriculture/IPM/scoutinfo/wheat/disease/wssm/wssm.htm).
Figure 3. Wheat field showing a large central patch of the crop affected by Wheat spindle streak mosaic virus (© Erik L. Stromberg, Department of Plant Pathology, Physiology, and
Weed Science, Virginia Tech).
Figure 4. Symptoms of Wheat spindle streak mosaic virus (WSSMV) infection typically
appear in early spring right after green-up. The symptoms of WSSMV are yellow to light green streaks or dashes on a dark green background. Dashes are usually 3 mm to 6.5 mm long.
The spindle-shaped dashes and streaks are oriented parallel to the leaf veins (©Robert L. Bowden, Kansas State University).

Wheat spindle streak mosaic virus Diagnostic Manual
13
Figure 5. Wheat spindle streak mosaic virus produces yellow streaks running parallel to the
leaf veins (Courtesy of Gary Bergstrom, © Queen's Printer for Ontario).
Figure 6. A wheat leaf with spindle streak symptoms associated with Wheat spindle streak
mosaic virus infection (© Erik L. Stromberg, Department of Plant Pathology, Physiology, and Weed Science, Virginia Tech).

Wheat spindle streak mosaic virus Diagnostic Manual
14
Figure 7. Symptoms of advanced Wheat spindle streak mosaic virus infection, showing well-
developed short light green to yellow streaks running parallel with the veins in leaves (Source: http://www.uky.edu/Agriculture/IPM/scoutinfo/wheat/disease/wssm/wssm.htm).
Figure 8. The effect of stunting caused by Wheat spindle streak mosaic virus (WSSMV)
infection is demonstrated through the use of WSSMV-resistant (left) and WSSMV-susceptible (right) wheat varieties (© Alabama Cooperative Extension System).
Figure 9. A wheat field with a cultivar susceptible to Wheat spindle streak mosaic virus growing on the left, and a resistant cultivar growing on the left, showing the reduction in
tillering due to virus infection (© Erik L. Stromberg, Department of Plant Pathology, Physiology, and Weed Science, Virginia Tech).

Wheat spindle streak mosaic virus Diagnostic Manual
15
2.5. Diseases associated with causing symptoms
similar to Wheat spindle streak mosaic virus
WYMV has not been included in this list, as symptoms caused by both WYMV and
WSSMV are very similar, and both infect wheat (Clover and Henry, 1999). These
viruses can only be accurately differentiated using molecular tests.
2.5.1. Soil-borne wheat mosaic virus
Soil-borne wheat mosaic virus (SBWMV) (synonym Wheat soil-borne mosaic virus)
and WSSMV have similar symptoms, life cycle and field distribution patterns, and
share the same fungal vector (Bowden, 2005; OMAF Staff, 2002; Tenuta and
Johnson, 2003). In certain cases, both SBWMV and WSSMV may be present in the
same field (OMAF Staff, 2002; Tenuta and Johnson, 2003), and the situation is
further confused by the fact that many plants are found to be infected with both
viruses (Bowden, 2005).
Although both viruses are vectored by the same fungus, the similarity between
WSSMV and SBWMV is only superficial, as the viruses are unrelated (Bowden,
2005; Linker et al., 1994; Smith, 2005). WSSMV belongs to the genus Bymovirus,
and SBWMV belongs to the genus Furovirus. Also, WSSMV infects wheat and rye,
whereas SBWMV can also infect barley (Linker et al., 1994).
As with WSSMV, symptoms of SBWMV infection often appear in low, wet areas, and
may cover all or most of a field due to the distribution of the fungal vector (fig. 10)
(Linker et al., 1994). Symptoms of WSSMV are usually visible prior to symptoms
associated with SBWMV, which typically appear in early spring right after green-up
(Bowden, 2005). The symptoms of SBWMV are most obvious in the early spring
months (Linker et al., 1994), and as with WSSMV, symptoms will not be expressed in
late spring when conditions are warmer (Linker et al., 1994; Smith, 2005).
The characteristic symptoms of WSSMV are the yellow-to-light green tapered streaks
running parallel to the leaf veins (OMAF Staff, 2002). In contrast, SBWMV typically
has a mosaic of green islands or blotches occurring on the leaves, ranging from mild
green to a prominent yellow that are longer than they are wide, against a yellow

Wheat spindle streak mosaic virus Diagnostic Manual
16
background (fig. 11 and 12) (Bowden, 2005; Linker et al., 1994; OMAF Staff, 2002;
Smith, 2005; Tenuta and Johnson, 2003). It must be noted that plants severely
infected with WSSMV resemble infection by WSBMV as they may develop mottled
leaves (Bowden, 2005), and that both WSSMV and SBWMV cause plant stunting
(fig. 13) (Linker et al., 1994; OMAF Staff, 2002).
Figure 10. Soil-borne wheat mosaic virus (SBWMV) field disease pattern (left), and patch of
SBWMV affected winter wheat (right) (Left; Source: http://www.plantpathology.tamu.edu/appel/vwheat3.htm, right; ©Department for Environment,
Food and Rural Affairs)
Figure 11. Soil-borne wheat mosaic virus (SBWMV) symptoms on wheat (left), and close-up
of SBWMV affected plants (©Department for Environment, Food and Rural Affairs)

Wheat spindle streak mosaic virus Diagnostic Manual
17
Figure 12. In each photograph, a healthy plant (left) is compared to a plant showing
symptoms of Soil-borne wheat mosaic virus infection (right).
Figure 13. Severe stunting of wheat caused by Soil-borne wheat mosaic virus
(© Ohio State University).

Wheat spindle streak mosaic virus Diagnostic Manual
18
2.5.2. Wheat streak mosaic virus
Wheat streak mosaic virus (WSMV) belongs to the genus Rymovirus (family
Potyviridae), and is transmitted by the wheat leaf curl mite (Aceria tulipae). This virus
produces symptoms that are similar to WSSMV infection, ie., stunted plants with
mottled green and yellow streaks extending the length of the leaf (fig. 14 and 15).
Plants can also exhibit rolling or curling of the youngest leaves due to damage by the
mite vector. Symptoms of WSMV typically appear in the spring (Stromberg, 2002).
Unlike WSSMV, WSMV field infection is not evenly distributed, infection is usually
located near the field edges, and infected plants continue to show symptoms
throughout the increasingly warm spring weather (Smith, 2005; Stromberg, 2002).
Figure 14. Streak mosaic pattern of Wheat streak mosaic virus (Left; © 2000 by the
University of Nebraska, right; http://www.oznet.ksu.edu/path-ext/Alerts/2002%20alerts/alert04.asp)
Figure 15. As the wheat crop develops, plants affected with Wheat streak mosaic virus are typically severely stunted with yellow mottled and streaked leaves. (© Erik L. Stromberg,
Department of Plant Pathology, Physiology, and Weed Science, Virginia Tech).

Wheat spindle streak mosaic virus Diagnostic Manual
19
2.6. Management of Wheat spindle streak mosaic
virus
There is no cure for a WSSMV-infected plant, and therefore preventative measures
are employed to reduce infection rates (Anon, 2003). Crop rotation as a
management tool to reduce the incidence of disease is only moderately effective
because the virus can remain infective in the soil for many years inside spores of the
fungal vector (Bowden, 2005; Linker et al., 1994; OMAF Staff, 2002; Tenuta and
Johnson, 2003). Manipulation of planting dates, irrigation schedules and paddock
drainage by avoiding exposure of crops to high risk low-lying wet areas (Bowden,
2005) could reduce virus incidence. Anecdotal evidence has shown fields that have
had liberal amounts of poultry and livestock manure added have a reduction in
WSSMV build-up (OMAF Staff, 2002; Tenuta and Johnson, 2003).
Chemical control is ineffective, as fungicides will not control the virus or the vector
(Anon, 2003) due to the longevity of the fungal resting spores (Campbell, 1996;
Kanyuka et al., 2003). The use of fumigants is not a practical or an affordable option
for broadacre farmers (Roget, 2004), and internationally, it is not an ecologically
acceptable form of disease control (Kanyuka et al., 2003).
The current thinking is that the only way to control WSSMV is to plant resistant
varieties (Anon, 2003). Presently there are no cultivars completely resistant to
WSSMV (Bowden, 2005), although many sources of resistance to WSSMV have
been identified in commercial wheat varieties and in wild Triticum species, with the
resistance trait being highly heritable (Kanyuka et al., 2003). Other wheat cultivars
such as Sierra, Mustang, and Thunderbird are highly susceptible and should be
avoided in high risk districts (Bowden, 2005).

Wheat spindle streak mosaic virus Diagnostic Manual
20
3. Polymyxa graminis
3.1. The impact of infection by the fungus, Polymyxa
graminis
Polymyxa graminis is a plasmodiophorid, a weakly pathogenic eukaryotic obligate
biotrophic parasite of plant roots with a worldwide distribution (Barr, 1979; Gitton et
al., 1999; Kanyuka et al., 2003; Ward et al., 2005), which includes New Zealand
(Jones, 2004). Although P. graminis is weakly pathogenic, its ability to vector cereal
crop diseases is what causes significant yield reductions (Kanyuka et al., 2003).
As previously stated, P. graminis is the vector for WSSMV. This fungus is also a
vector for the following viral diseases (Anon, 2005; Barr, 1979; Gitton et al., 1999;
Kanyuka et al., 2003; Morales et al., 1999; Ward et al., 2005):
- Aubian wheat mosaic virus
- Barley mild mosaic virus
- Barley yellow mosaic virus
- Chinese wheat mosaic virus
- Indian peanut clump virus (IPCV)
- Oat golden stripe virus
- Oat mosaic virus
- Peanut clump virus (PCV)
- Rice necrosis mosaic virus
- Rice stripe necrosis virus (RSNV)
- Soil-borne cereal mosaic virus
- Soil-borne wheat mosaic virus
- Sorghum chlorotic spot virus
- Wheat yellow mosaic virus
As the viruses are protected from the environment within the P. graminis resting
spores, which may remain dormant but viable for decades, the diseases are very
persistent (Kanyuka et al., 2003). After fungal spore germination, the viruses are
carried within the swimming zoospores to the plant host (Ward et al., 2005). The
host specificity of P. graminis is complex. For example, a West African isolate of P.
graminis that transmits RSNV is able to complete its lifecycle within rice roots. A
second West African isolate of P. graminis that is capable of transmitting PCV can
not complete its lifecycle within rice roots (Anon, 2005). It is therefore imperative to
understand the biology of local isolates of P. graminis and their ability to transmit
plant viruses.
Wheat spindle streak mosaic virus Diagnostic Manual
21
3.2. Description of Polymyxa graminis
Polymyxa graminis is an obligate root-infecting organism that was originally
described from wheat by Ledingham (1939). Barr (1979) examined roots of common
weeds for the presence of plasmodiophoraceous fungi, but was unable to identify a
natural reservoir of host species. The recorded hosts for P. graminis are listed in
Table 1.
Table 1: Recorded plant hosts of Polymyxa graminis.
Common name Botanical name Source
Barley Hordeum vulgare Barr, 1979; Rush, 2003
Couch grass, Quackgrass Agropyron repens Barr, 1979
Millet Panicum miliaceum Rush, 2003
Rye Secale cereale Barr, 1979; Rush, 2003
Sorghum Sorghum vulgare Rush, 2003
Wheat Triticum aestivum, T. durum Barr, 1979; Rush, 2003
Polymyxa graminis belongs to the order Plasmodiophorales, family
Plasmodiophoraceae (Kanyuka et al., 2003; Ward et al., 2005). In total, ten genera
are recognised in the Plasmodiophorales, all of which are intracellular parasites of
higher plants or fungi (Ward and Adams, 1998): Polymyxa, Spongospora,
Plasmodiophora, Ligniera, Membranosorus, Octomyxa, Sorodiscus, Sorosphaera,
Tetramyxa and Woronina (Kanyuka et al., 2003). Several species of Polymyxa,
Spongospora and Plasmodiophora genera, including P. graminis, are of significant
agronomic importance (Kanyuka et al., 2003).
There are several distinctive characteristics common amongst plasmodiophorids.
They are obligate intracellular parasites (Kanyuka et al., 2003; Rush, 2003), replicate
through cruciform nuclear division (Rush, 2003), they form multinucleated plasmodia
(Kanyuka et al., 2003; Rush, 2003), and their zoospores have two, anterior whiplash
flagella of unequal length (Kanyuka et al., 2003; Rush, 2003) (fig. 16). All produce
environmentally resistant, long-lived resting spores (cysts) that are often clustered
together to form a sporosorus (cystosorus) (Braselton, 1995; Rush, 2003). The
production of zoospores and long-lived sporosori are significant features in the
epidemiology of the diseases caused by plasmodiophorids, and the viruses they
vector (Rush, 2003).

Wheat spindle streak mosaic virus Diagnostic Manual
22
Figure 16. Scanning electron micrograph of a single biflagellate Polymyxa graminis zoospore,
showing the classic plasmodiophorid anterior whiplash flagella of unequal length (Kanyuka et al., 2003).
The genus Polymyxa, has two recognised species, P. graminis and P. betae. Both
species are morphologically indistinguishable (Barr, 1979; Legreve et al., 2002; Ward
and Adams, 1998), but were separated due to host range (Barr, 1979; Legreve et al.,
2002; Rush, 2003), and differences in cyst walls and zoosporangial plasmodia (Barr,
1979). P. graminis parasitises mostly monocotyledons, in the family Poaceae,
whereas P. betae favours dicotyledons within the Chenopodiaceae and related plant
families, such as Amaranthaceae, Portulaceae and Caryophyllaceae (Legreve et al.,
2002; Ward and Adams, 1998; Ward et al., 2005). The cyst walls in P. graminis are
fused, whereas in P. betae they are separated by a cementing substance, and the
zoosporangial plasmodia of P. graminis have longer discharge tubes than those of P.
betae (Barr, 1979). In recent times, ribosomal DNA ITS sequences have separated
P. graminis from P. betae, with isolates of P. graminis exhibiting a wider diversity in
ITS sequence when compared to P. betae (Rush, 2003).
The diversity within ITS sequences of P. graminis isolates may be a reflection of their
differences in temperature and host range. Indian isolates have a significantly higher
temperature optimum (27-30oC) than isolates from Europe and Canada (15-20oC,
Rush, 2003). A reduction in temperature to 15oC disables the Indian isolates, making
them incapable of vectoring PCV, although at that temperature the virus can replicate
and cause symptoms of infection (Rush, 2003).
The diversity in P. graminis isolates is reflected in host range (Legreve et al., 2002),
with tropical and subtropical isolates exhibiting a continuum in host specificity, as
opposed to an absolute, discrete host range favoured by isolates from temperate
regions (Rush, 2003). P. graminis normally infects graminaceous species, but
isolates from India are capable of infecting the dicotyledon, peanut (Arachis
hypogaea) (Rush, 2003). From an epidemiological standpoint, this distinction is

Wheat spindle streak mosaic virus Diagnostic Manual
23
important, as in vector-virus-host relationships, tropical isolates of P. graminis do not
need to complete their life-cycle within the virus susceptible plant (Rush, 2003). An
example is the P. graminis capable of transmitting PCV and IPCV. Both viruses
infect peanut, but peanut is a poor host for the fungal vector (Rush, 2003). Due to
the aggressiveness of this P. graminis isolate, its inability to complete its life cycle in
the peanut plant has little effect on disease severity of PCV and IPCV (Rush, 2003).
Legreve et al. (2002) proposed dividing P. graminis into five taxa based on the
combination of host range, temperature requirements, and genome characteristics,
as listed below.
1. P. graminis f. sp. temperata Legreve, Delfosse and Maraite
P. graminis on graminaceous species, particularly barley or wheat. P. graminis has
been detected once infecting sugar beet after heavy inoculation, favoured by
temperatures of 15-20oC, from temperate regions (Belgium, Canada, China, France,
Germany, UK), ITS1-5.8S gene-ITS2 sequence type Pg-I.
2. Polymyxa graminis f. sp. tepida Legreve, Delfosse and Maraite
P. graminis on graminaceous species, particularly barley, oat and wheat, favoured by
temperatures of 15-20oC, from temperate regions (Canada, UK) and ITS1-5.8S gene-
ITS2 sequence type Pg-II.
3. Polymyxa graminis f. sp. tropicalis Legreve, Delfosse and Maraite
P. graminis on graminaceous species, particularly sorghum, pearl millet and maize,
occasionally on wheat and barley, rarely on groundnut or sugar beet, favoured by
temperatures above 23oC, from tropical regions (India, Senegal), ITS1-5.8S gene-
ITS2 sequence type Pg-IIIa or b.
4. Polymyxa graminis f. sp. subtropicalis Legreve, Delfosse and Maraite
P. graminis on graminaceous species, particularly sorghum and pearl millet but also
wheat and barley, occasionally groundnut and sugar beet, favoured by temperatures
above 23oC, from sub-tropical regions (India, Pakistan) and ITS1-5.8S gene-ITS2
sequence type Pg-IVa or b.
5. Polymyxa graminis f. sp. colombiana Legreve, Delfosse and Maraite
P. graminis on rice from Colombia, ITS1-5.8S gene-ITS2 sequence type Pg-V.

Wheat spindle streak mosaic virus Diagnostic Manual
24
Of the five P. graminis taxa, two are known to occur in temperate regions (Ward et
al., 2005). The biological significance of the two temperate ribotypes is not clear, but
the trend found by Ward et al. (2005) was that ribotype I was isolated from barley,
and ribotype II from wheat and other cereals.
3.3. Life cycle of Polymyxa graminis
The lifecycle of P. graminis is separated into two phases, 1) the production of
zoospores, and 2) the production of resting spores, with each phase initiated by the
attachment and penetration of the plant host epidermal or root hair cells by
zoospores (Jones, 2004; Kanyuka et al., 2003). The zoospores form within
zoosporangia located within root hairs or epidermal cells (Kanyuka et al., 2003), and
the zoosporangia (resting spores) burst open releasing the short-lived, motile
zoospores into the soil around the roots (Kanyuka et al., 2003) (fig. 17 and 18).
Figure 17. Diagram of the lifecycle of Polymyxa graminis (Kanyuka et al., 2003)
The zoospores, with the aid of their two flagella, swim short distances through the
soil moisture film (moisture between soil particles) to infect roots (Jones, 2004). The

Wheat spindle streak mosaic virus Diagnostic Manual
25
fungus invades the root hairs of young susceptible plants in the autumn during
periods of high soil moisture (Bowden, 2005) (fig. 19). The zoospore contents
enlarge within the invaded host cell and undergo several cycles of synchronous
mitotic 'cruciform' nuclear divisions and reduction in nuclear size (Kanyuka et al.,
2003). The Polymyxa cytoplasm becomes an irregularly shaped multinucleate sac
(plasmodium), which develops into a zoosporangium (Ward et al., 2005), separated
from the host cytoplasm by a distinct cell wall (Kanyuka et al., 2003). Septa form
within the zoosporangium, dividing it into lobes that expand in volume followed by
several cycles of 'non-cruciform' mitotic nuclear divisions, and many form exit tubes
extending to the host cell wall (Kanyuka et al., 2003). The secondary zoospores are
cleaved apart, become rounded when they mature, and the septa between
zoosporangial segments disintegrate (Kanyuka et al., 2003), later developing into
resting spore clusters (Ward et al., 2005).
Figure 18. Life cycle of Polymyxa graminis: (a) mature sporangia with exit tubes passing through cells. Motile zoospores emerging formed from one sporangium; (b) biflagellate zoospores; (c) living amoeboid zoospores; 9d) amoeboid spores during period of active growth; (e) individual plasmodia (meronts); (f) same cell as d after the plasmodia have coalesced; (g) plasmodia just prior to cleavage into incipient cystosori; (h) cleavage of
plasmodia into polygonal resting spores (cystosori); (i) wheat cell completely filled with resting spores. (Adapted from Ledingham) (Integrated Pest Management Copyright © 2002
University of Illinios at Urbana-Champaign).

Wheat spindle streak mosaic virus Diagnostic Manual
26
Figure 19. Diagram of Polymyxa graminis zoospore encystment and penetration of root cells
(S: Stachel, R: Rohr: adhesium, N: nucleus, V: vacuole, L: lipid droplet) (Kanyuka et al., 2003).
The fungus forms dark clusters of resting spores in the cortical tissues within the
infected root (fig. 20 to 25). These spore balls are released into the soil when the
roots decay (Bowden, 2005), and the fungus survive as clusters of thick-walled
resting spores within the soil (Ward et al., 2005) for more than 15 years in the
absence of access to susceptible hosts (Jones, 2004). The cycle is restarted when
the resting spores germinate to each produce one biflagellate zoospore (Kanyuka et
al., 2003), which, in turn, will infect the root epidermal cells of a suitable plant host
(Ward et al., 2005), and behaving in the same manner as zoospores derived from
zoosporangia (Kanyuka et al., 2003).
Figure 20. Transmission electron micrograph of resting spores of Polymyxa graminis in root cell of Triticum monococcum (Source: http://oak.cats.ohiou.edu/~braselto/plasmos/poly.html)

Wheat spindle streak mosaic virus Diagnostic Manual
27
Figure 21. Clusters of Polymyxa graminis resting spores inside root cells of wheat (X 205)
(Source: http://www.bsu.edu/classes/ruch/msa/barr.html).
Wheat spindle streak mosaic virus Diagnostic Manual
28
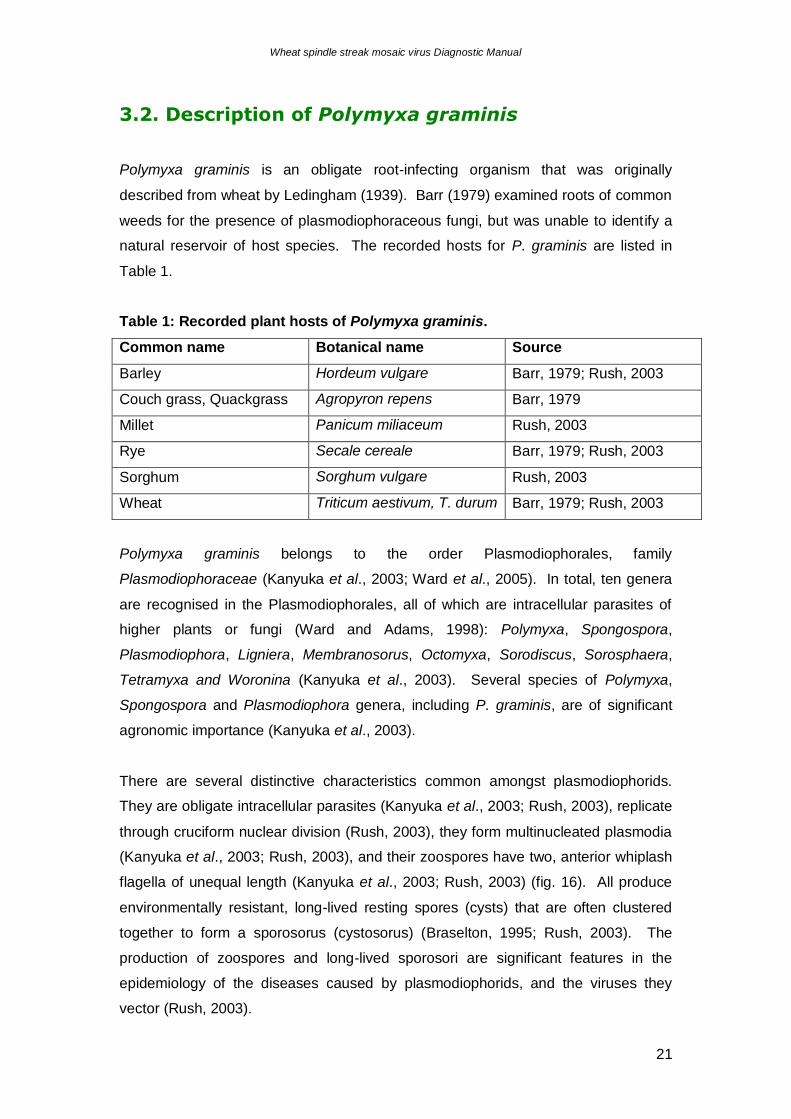
Figure 22. Grass roots cleared of protoplasm to reveal the presence of clusters of small
resting spores of Polymyxa graminis. Top: low-power view of a root. Bottom: two clusters of resting spores in a single root cortical cell
(Source: http://helios.bto.ed.ac.uk/bto/microbes/chytrid.htm).
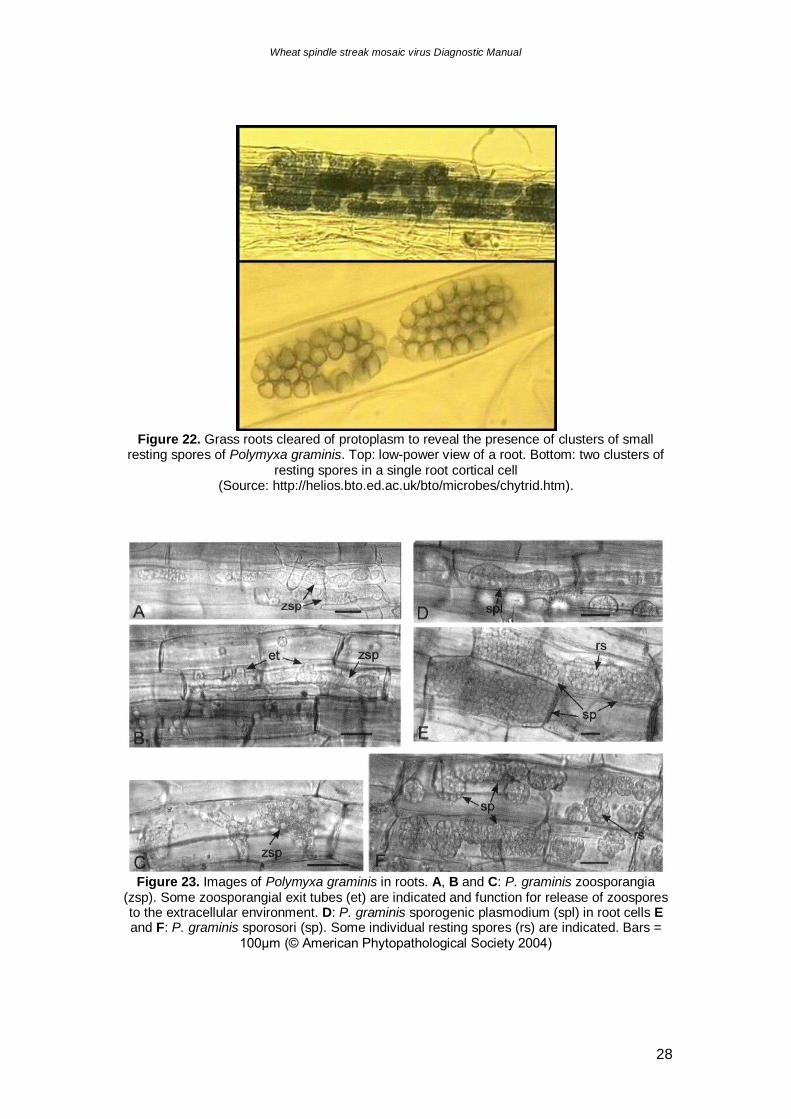
Figure 23. Images of Polymyxa graminis in roots. A, B and C: P. graminis zoosporangia
(zsp). Some zoosporangial exit tubes (et) are indicated and function for release of zoospores to the extracellular environment. D: P. graminis sporogenic plasmodium (spl) in root cells E and F: P. graminis sporosori (sp). Some individual resting spores (rs) are indicated. Bars =
100μm (© American Phytopathological Society 2004)
Wheat spindle streak mosaic virus Diagnostic Manual
29
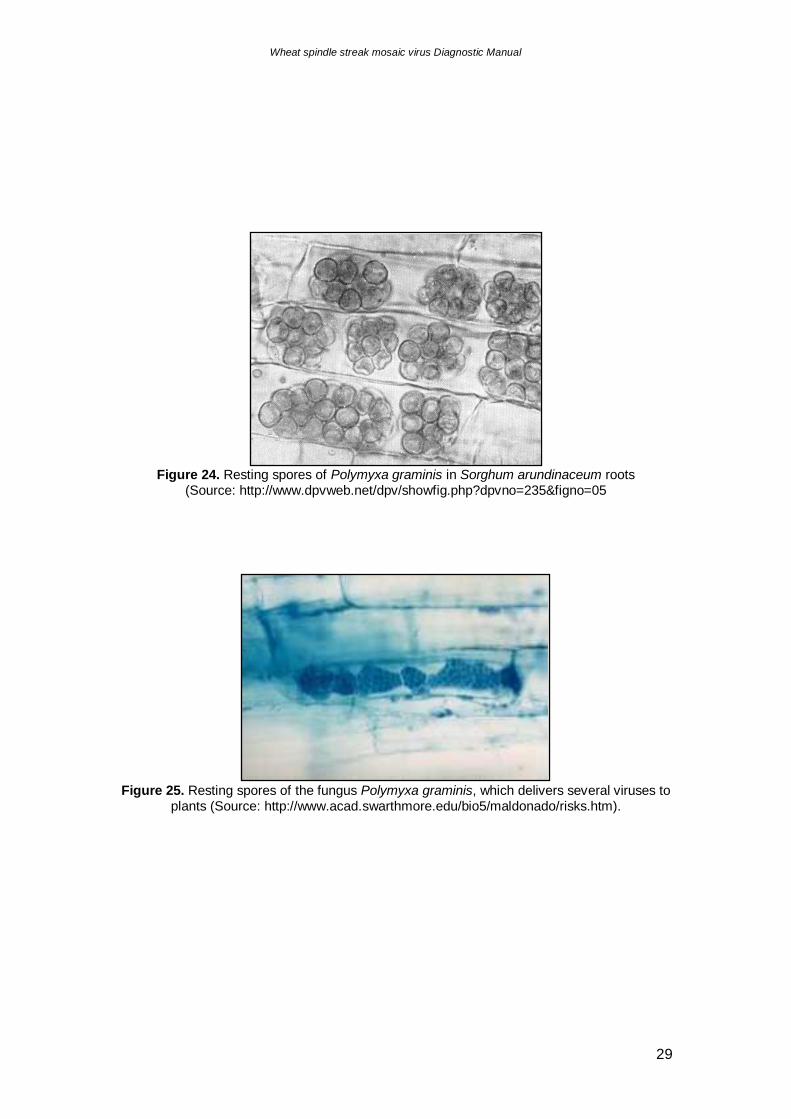
Figure 24. Resting spores of Polymyxa graminis in Sorghum arundinaceum roots
(Source: http://www.dpvweb.net/dpv/showfig.php?dpvno=235&figno=05
Figure 25. Resting spores of the fungus Polymyxa graminis, which delivers several viruses to
plants (Source: http://www.acad.swarthmore.edu/bio5/maldonado/risks.htm).

Wheat spindle streak mosaic virus Diagnostic Manual
30
3.3. Polymyxa graminis fungal isolation
Polymyxa graminis is an obligate biotroph and can therefore only be maintained in
the roots of host plants (Kanyuka et al., 2003; Subr et al., 2002). As P. graminis can
not be grown in culture, all purified preparations could potentially be contaminated
with soil microorganisms and plant debris (Subr et al., 2002). It is generally difficult
to obtain good quality P. graminis DNA for molecular studies that is free from the
contaminating DNA of the host plant or other organisms (Ward et al. 1994),
especially when samples are collected from the field (Kanyuka et al., 2003).
Adams et al. (1986), Mutasa et al. (1993), Subr et al. (2002) and Ward et al. (1994)
have isolated P. graminis spores for molecular biological experiments by propagating
the fungus on host plant roots in semi-sterile sand cultures. Subr et al. (2002) used
subtractive hybridisation to further isolate P. graminis-specific DNA from plant roots.
However, even in experimental glasshouse conditions, P. graminis multiplication is
slow, taking 34 weeks to produce zoospores and approximately 23 months to
produce resting spores (Kanyuka et al., 2003).
Wheat spindle streak mosaic virus Diagnostic Manual
31
3.4. Transmission of Wheat spindle streak mosaic
virus by Polymyxa graminis
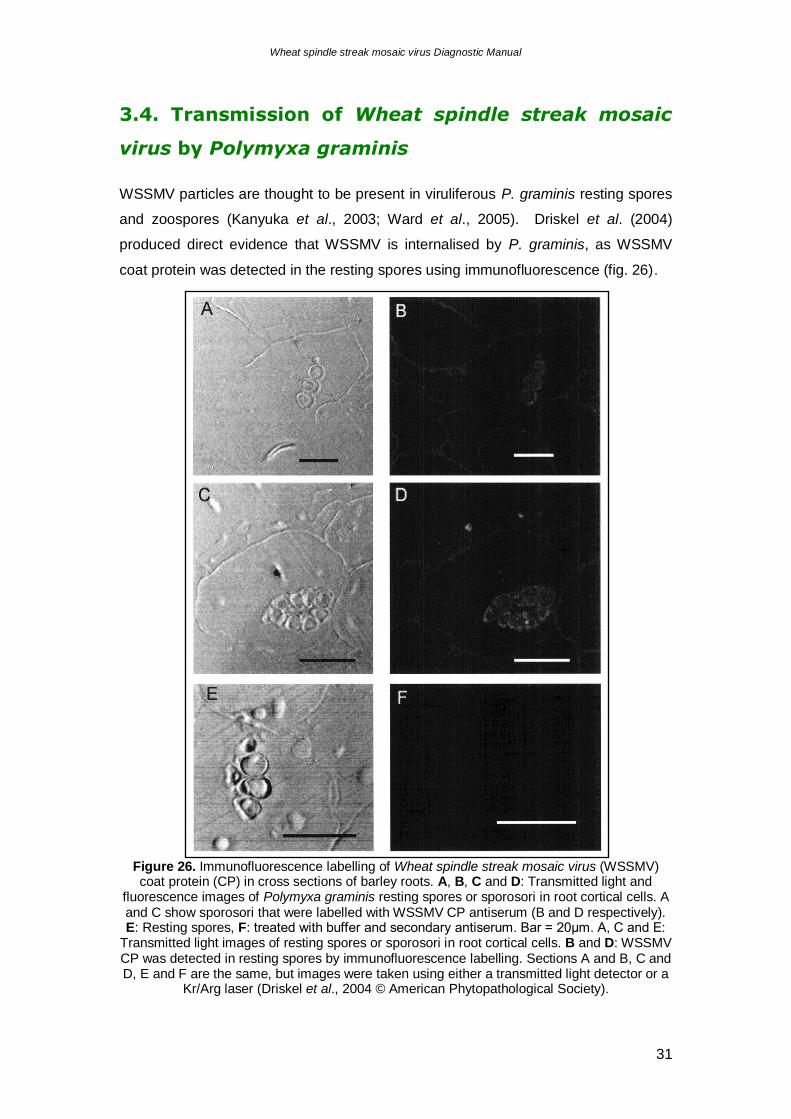
WSSMV particles are thought to be present in viruliferous P. graminis resting spores
and zoospores (Kanyuka et al., 2003; Ward et al., 2005). Driskel et al. (2004)
produced direct evidence that WSSMV is internalised by P. graminis, as WSSMV
coat protein was detected in the resting spores using immunofluorescence (fig. 26).
Figure 26. Immunofluorescence labelling of Wheat spindle streak mosaic virus (WSSMV) coat protein (CP) in cross sections of barley roots. A, B, C and D: Transmitted light and
fluorescence images of Polymyxa graminis resting spores or sporosori in root cortical cells. A and C show sporosori that were labelled with WSSMV CP antiserum (B and D respectively). E: Resting spores, F: treated with buffer and secondary antiserum. Bar = 20μm. A, C and E:
Transmitted light images of resting spores or sporosori in root cortical cells. B and D: WSSMV CP was detected in resting spores by immunofluorescence labelling. Sections A and B, C and D, E and F are the same, but images were taken using either a transmitted light detector or a
Kr/Arg laser (Driskel et al., 2004 © American Phytopathological Society).

Wheat spindle streak mosaic virus Diagnostic Manual
32
WSSMV cannot be removed from zoospores by washing, or inactivated by
application of antiserum. P. graminis resting spores remain viruliferous after
treatments with diluted NaOH and HCl (Kanyuka et al., 2003). The precise
mechanism of virus uptake and transfer is unknown (Rush, 2003), and it is also not
known whether WSSMV is able to replicate within P. graminis, although indirect
evidence suggests that it does not (Kanyuka et al., 2003). Zoospores released from
viruliferous P. graminis isolates grown in virus-resistant host plants no longer contain
WSSMV, or loose the ability to transmit the virus (Kanyuka et al., 2003).
Polymyxa graminis acquires WSSMV when it multiplies inside virus-infected plant
cells as the virus is incorporated in the resulting plasmodium (Rush, 2003). If this
plasmodium develops into a zoosporangium, the secondary zoospores will be
viruliferous, and if it develops into a sporosorus, the virus particles will survive inside
the resting spores for years (Rush, 2003). When the plant host cell dies and
deteriorates, the infected resting spores are released into the surrounding soil, and
upon germination, will release motile viruliferous primary zoospores in search of a
plant host (Rush, 2003) (fig. 17).
Motile P. graminis zoospores are released from resting spores or zoosporangia, and
infects the plant via root hairs or epidermal cells (Rush, 2003). If the zoospore is
viruliferous, virus particles are introduced into the plant cytoplasm soon after contents
of the zoospore are injected into the cell (Rush, 2003). The virus replicates and
initiates disease, becoming systemic, causing symptoms in leaves, and affecting
plant growth and yield (Rush, 2003; Ward et al., 2005).

Wheat spindle streak mosaic virus Diagnostic Manual
33
3.5. Management strategies for Polymyxa graminis
Polymyxa graminis is a ubiquitous soil inhabitant wherever susceptible crops are
produced (Rush, 2003). Once the fungus is introduced to new areas of cereal
production, eradication is difficult as the dormant resting spores are able to persist for
many years in soil in the absence of a plant host (Jones, 2004). Therefore, strict
farm hygiene must be implemented to prevent spread, and techniques such as
minimum or zero tillage can be utilised to reduce soil movement via cultivation
(Jones, 2004).
This fungus survives in a variety of soil types that vary greatly in pH, texture, and
structure (Rush, 2003). For example, P. graminis can function as a vector in soil pH
values 4 to 8 with minimal effect on its ability to vector Rice stripe necrosis virus
(Rush, 2003). Infection of plant roots by P. graminis is favoured by high soil moisture
conditions, so well drained soils typically reduce infection (Rush, 2003). However,
heavy soils with poor structure and compaction layers, which slow drainage and
retard root growth, enhance root infection (Rush, 2003).
Due to the longevity and hardiness of P. graminis resting spores, host resistance to
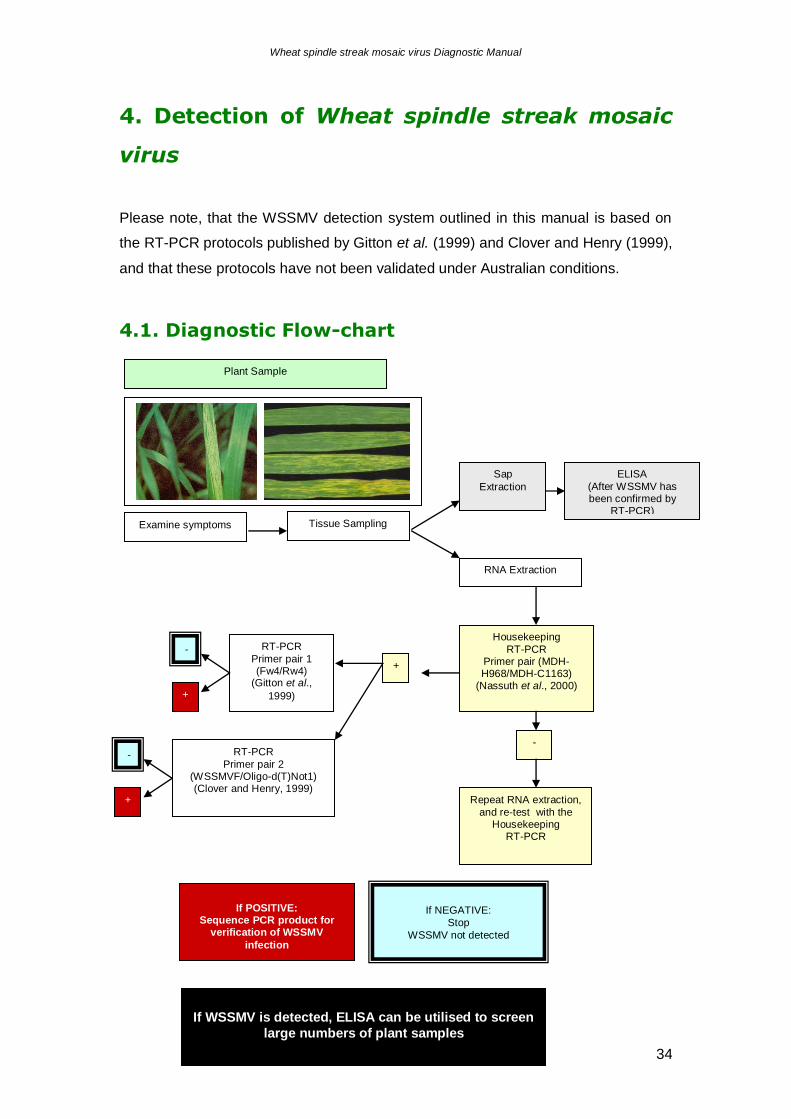
the fungus by cereal cultivars would be a very desirable trait. Unfortunately, at
present there are no sources of natural P. graminis resistance. The European barley
cultivars tested so far have been highly susceptible to P. graminis and there were
some differences between wheat cultivars in their degree of susceptibility (Ward et
al., 2005).
Wheat spindle streak mosaic virus Diagnostic Manual
34
4. Detection of Wheat spindle streak mosaic
virus
Please note, that the WSSMV detection system outlined in this manual is based on
the RT-PCR protocols published by Gitton et al. (1999) and Clover and Henry (1999),
and that these protocols have not been validated under Australian conditions.
4.1. Diagnostic Flow-chart
If POSITIVE: Sequence PCR product for
verification of WSSMV
infection
If NEGATIVE:
Stop
WSSMV not detected
RNA Extraction
Housekeeping
RT-PCR Primer pair (MDH-H968/MDH-C1163)
(Nassuth et al., 2000)
+
RT-PCR
Primer pair 2 (WSSMVF/Oligo-d(T)Not1) (Clover and Henry, 1999)
RT-PCR
Primer pair 1 (Fw4/Rw4)
(Gitton et al.,
1999)
Examine symptoms Tissue Sampling
-
Repeat RNA extraction,
and re-test with the Housekeeping
RT-PCR
+
-
+
-
Plant Sample
If WSSMV is detected, ELISA can be utilised to screen
large numbers of plant samples
Sap
Extraction
ELISA
(After WSSMV has been confirmed by
RT-PCR)

Wheat spindle streak mosaic virus Diagnostic Manual
35
4.2. Sample collection
4.2.1. Plant
Record plant symptoms. Collect leaves and roots from plants suspected of WSSMV
infection. Wash roots clean of soil under fast running tap water. Store plant material
at 4oC or at –20oC (Gitton et al., 1999; Kingsnorth et al., 2003).
4.2.2. Soil
Collect soil samples in a “W” pattern across the field. Take a 1-cup sample of soil
every ten metres. Soil samples can be pooled. To increase the likelihood of
detecting virulent P. graminis, include soil samples collected from the wettest area of
the paddock.
To recover WSSMV from the soil samples, rear T. aestivum seedlings in the suspect-
virus-infected soil (Clover and Henry, 1999). Incubate two-week old seedlings of T.
aestivum in a slurry of 40 g of infected soil at 20oC for 10-14 days. After incubation,
transplant into sterile sand to give a 1:9 dilution of infected soil:sand. Grow the
plants for a further 3 weeks at 20oC, then decrease the temperature to 10oC. Record
plant symptoms. Take leaf samples monthly. Store plant material at 4oC or at –
20oC.
Currently, there are no soil extraction protocols for extracting WSSMV or P. graminis
directly from soil.

Wheat spindle streak mosaic virus Diagnostic Manual
36
4.3. Total RNA extraction
Wear disposable gloves and a lab coat at all times. Have an autoclave bag ready to
dispose of all plant material, tips, tubes, gloves, and paper towel that has come into
contact with any suspect plant material. A footbath containing disinfectant located at
the doorway of the laboratory must be used when exiting the lab.
4.3.1. Equipment required
1. 2-20 L, 20-200 L, and 200-1000 L pipettes and sterile tips
2. Autoclave
3. Autoclave bags
4. Balance (at least 2 decimal places)
5. Disposable gloves
6. Microcentrifuge
7. Sterile microcentrifuge tubes
8. Paper towel
9. RNeasy® Plant Mini Kit (QiagenTM) (Kingsnorth et al., 2003)
10. Sharps container
11. Sterile scalpel blades and scalpel blade handle
12. Waterbath or heatblock set at 70oC
13. Weighboats
ALSO OR OR
1a. 1b. 1c.
Autoclaved mortar Qiagen Tissue Lyser Homex tissue macerater
and pestle (QiagenTM) Homex bags
Fume hood 2 ml snap-lock tubes (Bioreba AG / BioSys)
Sterile sand Liquid Nitrogen Plastic disposable
Stainless steel beads pasteur pipettes

Wheat spindle streak mosaic virus Diagnostic Manual
37
4.3.2. Reagents required
1. MacKenzie buffer (MacKenzie et al., 1997)
Chemical Amount Final Concentration
Guanidine thiocyanate (CH5N3·CHNS) 23.64 g 4 M
3M Sodium acetate (C2H3NaO2) 3.33 ml 0.2 M
0.5M EDTA (C10H16N2O8) 2.5 ml 25 mM
PVP-40 (Polyvinylpyrrolidone) 1.25 g 2.5% (w/v)
Add sterile distilled water to final volume of 50 ml
Store at room temperature
Please note, fresh MacKenzie buffer should be prepared every 3-6 months
2. β-mercaptoethanol (C2H6OS)
3. 20% N-Lauroylsarcosine solution (w/v)
4. 100% Ethanol
4.3.3. Method
The RNA extraction method is based on that described by MacKenzie et al. (1997).
All steps are carried out at room temperature and is as follows:
1a. If using a mortar and pestle to homogenise samples:
1a-1. Determine the number of samples and label plastic tubes accordingly.
1a-2. Use new clean gloves and scalpel blades for each sample.
1a-3. Cut each new sample on fresh paper towel on the bench.
1a-4. Weigh out 400 mg of plant sample.
1a-5. Place sample in mortar.
1a-6. Add 1980 l of MacKenzie buffer.
1a-7. Add 20 l of β-mercaptoethanol in the fumehood.
1a-8. Homogenise in fume hood.
1a-9. Pipette 1.0 ml of the mixture into a labelled microcentrifuge tube (you
may need to cut the end of the pipette tip if the slurry is too thick).
1a-10. Continue to step 2.

Wheat spindle streak mosaic virus Diagnostic Manual
38
1b. If using the QiagenTM Tissue Lyser:
1b-1. Determine the number of samples and label the 2 ml snap-lock tubes
accordingly.
1b-2. Use new clean gloves and scalpel blades for each sample.
1b-3. Cut each new sample on fresh paper towel on the bench.
1b-4. Weigh out 100 mg of plant sample and place sample in the
appropriate tube.
1b-5. Add 990 l of MacKenzie buffer.
1b-6. Add 10 l of β-mercaptoethanol in the fumehood.
1b-7. Close tubes.
1b-8. Place tubes in the Adaptor Set, in the QiagenTM Tissue Lyser. Grind
for 1 min at 30 Hz.
1b-9. Rotate tubes within the Adaptor Set, so that tubes in the centre are
moved to the outside.
1b-10. Continue to step 2.
1c. If using the Homex tissue macerater:
1c-1. Determine the number of samples and label plastic tubes accordingly.
1c-2. Use new clean gloves and scalpel blades for each sample.
1c-3. Cut each new sample on fresh paper towel on the bench.
1c-4. Weigh out 200 mg of plant sample.
1c-5. Place sample in Homex bag.
1c-6. Add 1980 l of MacKenzie buffer.
1c-7. Add 20 l of β-mercaptoethanol in the fumehood.
1c-8. Macerate tissue with the Homex.
1c-9. With a plastic disposable pasteur pipette, transfer 1.0 ml of the mixture
into a labelled microcentrifuge tube.
1c-10. Continue to step 2.
2. Carefully read the RNeasy Mini Handbook.
3. Add 100 l of 20% Sarkosyl to each tube and mix.
4. Incubate tubes at 70oC for 10 minutes.
5. Spin tubes in microcentrifuge for 1 minute at 13,000 rpm.
6. Continue with step 4 of the “RNeasy® Plant Mini Protocol for Isolation of Total
RNA from Plant Cells and Tissues and Filamentous Fungi” on page 75 of the
RNeasy Plant Mini Handbook and follow as per manufacturer's instructions.

Wheat spindle streak mosaic virus Diagnostic Manual
39
4.4. Detection of Wheat spindle streak mosaic virus
in total RNA extracts using one step RT-PCR
For the reliable detection of WSSMV, total RNA extracts are subjected to three RT-
PCR tests, as outlined below, with primer sequences and annealing temperatures
listed in Table 2.
1. Primer pair Fw4 (F) / Rw4 (R). This primer pair amplifies a 457 bp region of the
RNA1 component of the WSSMV genome, from bases 730 to 1186 on the
reference WSSMV nucleotide sequence X73883. Please note that this primer
pair does not amplify isolates from Okalahoma or China, but that it will faintly
detect Barley yellow mosaic virus (Gitton et al., 1999).
2. Primer pair WSSMVF (F) / Oligo-d(T)Not1 (R). This primer pair amplifies a
region on the WSSMV coat protein gene on RNA1, generating a 982 bp fragment
(Clover and Henry, 1999).
3. House-keeping gene (Primer pair MDH-H968 (F) / MDH-C1163 (R). The MDH-
H968/MDH-C1163 primer pair are designed to amplify a 196 bp region of the
plant mRNA encoding malate dehydrogenase (MDH) gene (Nassuth et al., 2000).
This gene is highly conserved among plants and therefore RT-PCR amplification
of the MDH mRNA is used as an internal RT-PCR control to, a) determine the
quality of the RNA extract, and b) determine whether the RNA extract contains
inhibitors that will interfere with the activity of the reverse transcriptase and Taq
DNA polymerase enzymes. This RT-PCR is particularly important when
confirming the absence of WSSMV in the test sample.
4.4.1. Equipment required
1. 0-2 l, 2-20 l, 20-200 l, and 200-1000 l pipettes and sterile tips
2. 0.2 or 0.5 ml sterile PCR tubes
3. Bulb spinner or centrifuge
4. Disposable gloves
5. Freezer
6. Gel electrophoresis tanks, rigs and racks
7. DNA Molecular Weight markers
8. Ice
9. Leather gardening gloves
Wheat spindle streak mosaic virus Diagnostic Manual
40
10. Sterile microcentrifuge tubes to store reagents
11. Microwave
12. Power pack
13. Thermocycler
14. UV transilluminator with camera
4.4.2. Reagents
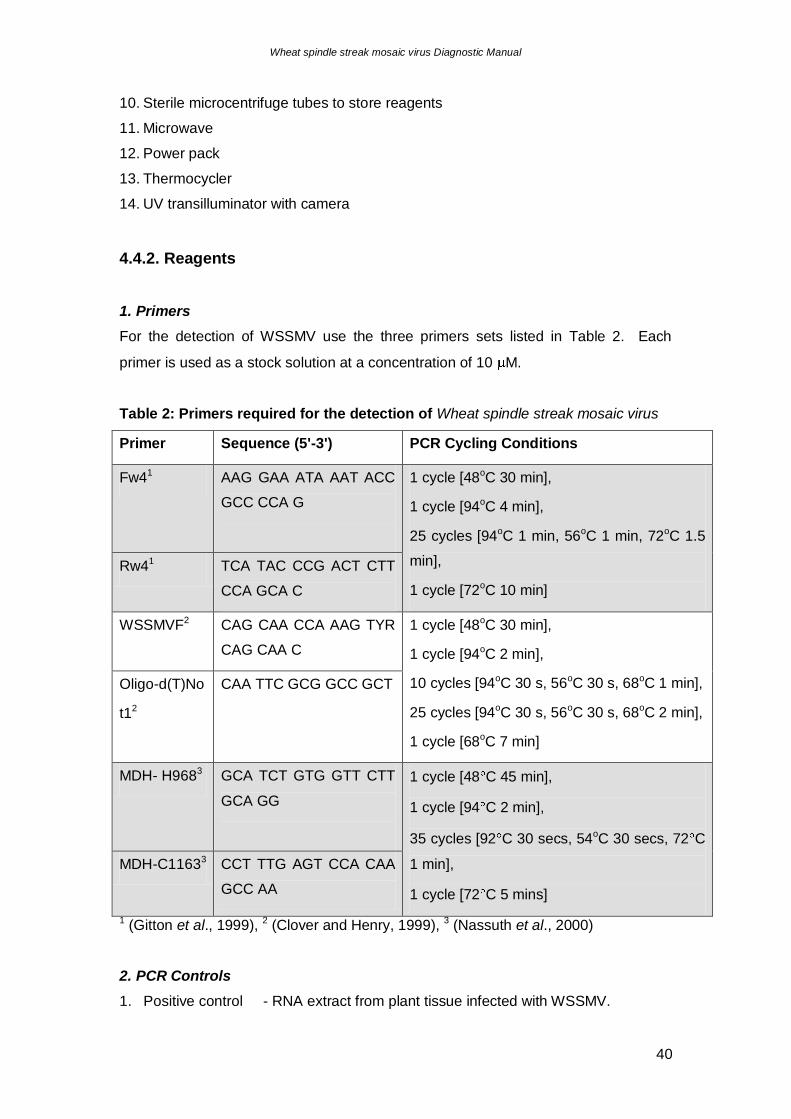
1. Primers
For the detection of WSSMV use the three primers sets listed in Table 2. Each
primer is used as a stock solution at a concentration of 10 M.
Table 2: Primers required for the detection of Wheat spindle streak mosaic virus
Primer Sequence (5'-3') PCR Cycling Conditions
Fw41 AAG GAA ATA AAT ACC
GCC CCA G
1 cycle [48oC 30 min],
1 cycle [94oC 4 min],
25 cycles [94oC 1 min, 56oC 1 min, 72oC 1.5
min],
1 cycle [72oC 10 min]
Rw41 TCA TAC CCG ACT CTT
CCA GCA C
WSSMVF2 CAG CAA CCA AAG TYR
CAG CAA C
1 cycle [48oC 30 min],
1 cycle [94oC 2 min],
10 cycles [94oC 30 s, 56oC 30 s, 68oC 1 min],
25 cycles [94oC 30 s, 56oC 30 s, 68oC 2 min],
1 cycle [68oC 7 min]
Oligo-d(T)No
t12
CAA TTC GCG GCC GCT
MDH- H9683 GCA TCT GTG GTT CTT
GCA GG
1 cycle [48 C 45 min],
1 cycle [94 C 2 min],
35 cycles [92 C 30 secs, 54oC 30 secs, 72 C
1 min],
1 cycle [72 C 5 mins]
MDH-C11633 CCT TTG AGT CCA CAA
GCC AA
1 (Gitton et al., 1999), 2 (Clover and Henry, 1999), 3 (Nassuth et al., 2000)
2. PCR Controls
1. Positive control - RNA extract from plant tissue infected with WSSMV.

Wheat spindle streak mosaic virus Diagnostic Manual
41
- Alternatively a “plasmid control” that has the target WSSMV
sequence cloned into the plasmid
2. Negative plant control - RNA extract from uninfected plant tissue of the same
species as that used for the positive control.
3. Negative buffer control - an aliquot of the RT-PCR “Master Mix” without template.
4. The house keeping RT-PCR, using primers MDH-H968/MDH-C1163, reduces the
risk of false negative results. The generation of a band confirms the presence of
RNA in the extract, and that the RNA extract does not contain inhibitors. Failure
to produce an amplicon of expected size (196 bp) indicates that either dilution of
the RNA extract or re-extraction of RNA from the sample is required.
3. RT-PCR reagents
1. One-step RT-PCR kit (Invitrogen® SuperScriptTM One-Step RT-PCR with
Platinum® Taq, Catalogue No. 12574-026, is recommended)
2. Nuclease-free water
4. 5x TBE Buffer
Per 1 litre
Tris (C4H11NO3) 54 g
Boric acid (H3BO3) 27.5 g
0.5M EDTA ([CH2.N(CH2.COOH).CH2COONa]2.2H2O) pH 8.0 20 ml
Store at room temperature.
5. 1% Agarose gel with ethidium bromide
Use a 1% DNA grade agarose gel made with 0.5x TBE solution, and stained with
0.03 g/ml Ethidium bromide.
6. 1x TE Buffer
Per 100 ml
1 M Tris-HCl (pH 8.0) 1 ml
0.5 M EDTA 200 μl
Adjust pH to 8.0± 0.2. Store at room temperature.
7. 6x loading dye
Per 100 ml

Wheat spindle streak mosaic virus Diagnostic Manual
42
1 x TE 10 ml
Glycerol (Sigma 200-289-5) 50 ml
Bromophenol blue (Sigma 263-653-2) trace (0.2%)
Store at room temperature.
4.4.3. One-step RT-PCR detection of Wheat spindle streak mosaic virus
This method is to be repeated for each set of the three primer pairs listed in Table 2.
Use one-step RT-PCR reagents as specified by the manufacturer. Some volumes
outlined below may vary depending on the buffer and enzyme concentrations
specified by the manufacturer. Ensure that the final volume of the RT-PCR reaction
is 25 µl by altering the volume of nuclease-free water accordingly.
1. Label sterile PCR tubes
2. Prepare "Master Mix" on ice in a sterile microcentrifuge tube.
The “Master Mix” usually contains buffer, forward and reverse primers, RT/Taq
and nuclease-free water.
Prepare the “Master Mix” according to the RT/Taq manufacturer’s
recommendations.
Ensure that the final volume for each reaction is 24 l.
Add 24 l of “Master Mix” to each PCR tube.
3. Add 1 l of each template (total RNA extract) to each corresponding PCR tube.
4. Cycle the tubes using the RT-PCR conditions listed in Table 2.
5. At completion of the RT-PCR, mix 10 l of each reaction with 2 l of 6x gel
loading dye, and load samples onto a 2% agarose gel with ethidium bromide.
6. Electrophorese in 0.5 x TBE at 100V for 45 minutes or until the bromophenol blue
front has migrated half way down the length of the gel.
7. Visualise and photograph gel on UV transilluminator.

Wheat spindle streak mosaic virus Diagnostic Manual
43
4.5. DNA Sequencing of PCR Products
4.5.1. Equipment required
1. 0-2 l, 2-20 l, 20-200 l, and 200-1000 l pipettes and tips
2. 0.2 or 0.5 ml PCR tubes
3. 1.5 or 2 ml centrifuge tubes to store reagents
4. Bulb spinner or centrifuge
5. Freezer
6. Ice machine
7. Latex gloves
8. PC with Internet access
9. QIAQuick® PCR Purification Kit (QiagenTM)
10. Thermocycler
11. UV illuminator
4.5.2. Reagents
1. ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kits (Applied
Biosystems (www.appliedbiosystems.com))
2. Forward and Reverse primers (As per section 4.4.2).
3. Sterile dH2O.
4.5.3. Method
PCR products are cleaned using the QIAquick Spin kit (Qiagen) as per
manufacturer’s instructions. The purified PCR products are prepared for sequencing
with ABI Big Dye (Roche), as per the manufacturer’s instructions. Sequencing is out-
sourced to Monash University, Melbourne or similar provider. The DNA sequences
are compared against sequences on the GenBank database using the program
BlastN (Altschul et al., 1997), to confirm if the positive PCR product sequence is from
WSSMV.

Wheat spindle streak mosaic virus Diagnostic Manual
44
4.6. Detection of Wheat spindle streak mosaic virus
with Enzyme-linked immunosorbent assay (ELISA)
4.6.1. Equipment
1. Wheat spindle streak mosaic virus coating antibody (Agdia; CAB 43001/0500)
(Store at 4oC)
2. Wheat spindle streak mosaic virus enzyme conjugated antibody (Agdia, ECA
43001/0500) (Store at 4oC)
3. Balance
4. Homex tissue macerater (Bioreba AG / BioSys)
5. Homex bags (Bioreba AG / BioSys)
6. Multiskan MS micro-titre plate reader (Labsystems)
7. Micro-titre plate (Nalgene)
8. Incubator
9. Fridge
10. 20-200 l pipette tips
11. Plastic sample cups
12. Plastic zip-lock bags
4.6.2. Reagents
Make all reagents with distilled water only.
1. Coating buffer
Per 1 litre
Sodium carbonate (anhydrous) 1.59 g
Sodium bicarbonate 2.93 g
Sodium azide 0.2 g
Adjust pH to 9.6. Store at 4oC.
2. ELISA extraction buffer
Per 1 litre
Sodium sulfite (anhydrous) 1.3 g
Polyvinylpyrrolidine (PVP) MW 24-40,000 MW 20.0g
Sodium azide 0.2 g

Wheat spindle streak mosaic virus Diagnostic Manual
45
Powdered egg (chicken) albumin, Grade II 2.0 g
Tween-20 20.0 g
Adjust pH to 7.4. Store at 4oC.
3. PBST buffer (Wash buffer)
Per 1 litre
Sodium chloride 8.0 g
Sodium phosphate, dibasic (anhydrous) 1.15 g
Potassium phosphate, monobasic (anhydrous) 0.2 g
Potassium chloride 0.2 g
Tween-20 0.5 g
Adjust pH to 7.4. Store at 4oC.
4. ECI buffer
Per 1 litre
Bovine serum albumin (BSA) 2.0 g
Polyvinylpyrrolidine (PVP) MW 24-40,000 MW 20.0g
Sodium azide 0.2 g
Adjust pH to 7.4. Store at 4oC.
5. Substrate visualising buffer
Dissolve 5 mg P-nitrophenyl phosphate in 5 ml substrate buffer (9.7 % (v/v)
Dethanolamine, pH 9.8)
4.6.3. Biological Reagents
Due to quarantine restrictions, there is no positive control available for WSSMV.
For a negative control use tissue from a non-infected plant of the same species as
that being tested.
4.6.4. Method
1. Dilute the concentrated WSSMV coating antibody into coating buffer at the
dilution recommended by the manufacturer. Make 1 ml per 8 wells. Use
immediately.
2. Add 100 μl of prepared coating buffer to each well. Incubate for 4 hours at 37oC.
(see Appendix 1 for layout with or without using the outer wells).

Wheat spindle streak mosaic virus Diagnostic Manual
46
3. Homogenise plant tissue: for negative controls and each sample, homogenise 1
gram of leaf or root tissue 10 ml extraction buffer.
4. After incubation of coating antibody, empty wells into a sink. Fill wells to
overflowing with PBST. Quickly empty. Repeat 4 to 8 times. Final rinse with
distilled water.
5. After washing, hold the plate upside down and tap firmly on a folded paper towel
to dry the wells.
6. To each micro-titre plate load 100 μl each of negative and buffer controls in
duplicate wells.
7. Add 100 μl of each sample homogenate into duplicate wells. Ensure that you
have a record of sample location (well grid reference) for each sample (Appendix
1).
8. Incubate plate overnight at 4oC.
9. Ten minutes prior to washing make enzyme antibody conjugate. Dilute the
concentrated WSSMV enzyme antibody conjugate into ECI buffer at the dilution
recommended by the manufacturer. Make 1 ml per 8 wells. Mix thoroughly.
10. After incubation of plant sap extract, empty wells into a sink with a quick flipping
motion. Fill wells to overflowing with PBST buffer. Quickly empty. Repeat 4 to 8
times and include several brief distilled water washes.
11. After washing, hold the plate upside down and tap firmly on a folded paper towel
to dry the wells.
12. Add 100 μl of prepared enzyme antibody conjugate per well.
13. Incubate plate at 37oC for 4 hours.
14. Wash plate three times with PBST buffer and three times with distilled water.
15. After washing, hold the plate upside down and tap firmly on a folded paper towel
to dry the wells.
16. Add 100 μl of substrate visualising buffer to each well. Incubate at room
temperature for 30 to 60 minutes.
17. Measure the micro-titre plate with a plate reader (Labsystems) at an optical
density of 405 nm.
18. Samples are considered positive if the OD readings are 2.5 times the (corrected)
healthy control OD reading.
Wheat spindle streak mosaic virus Diagnostic Manual
47
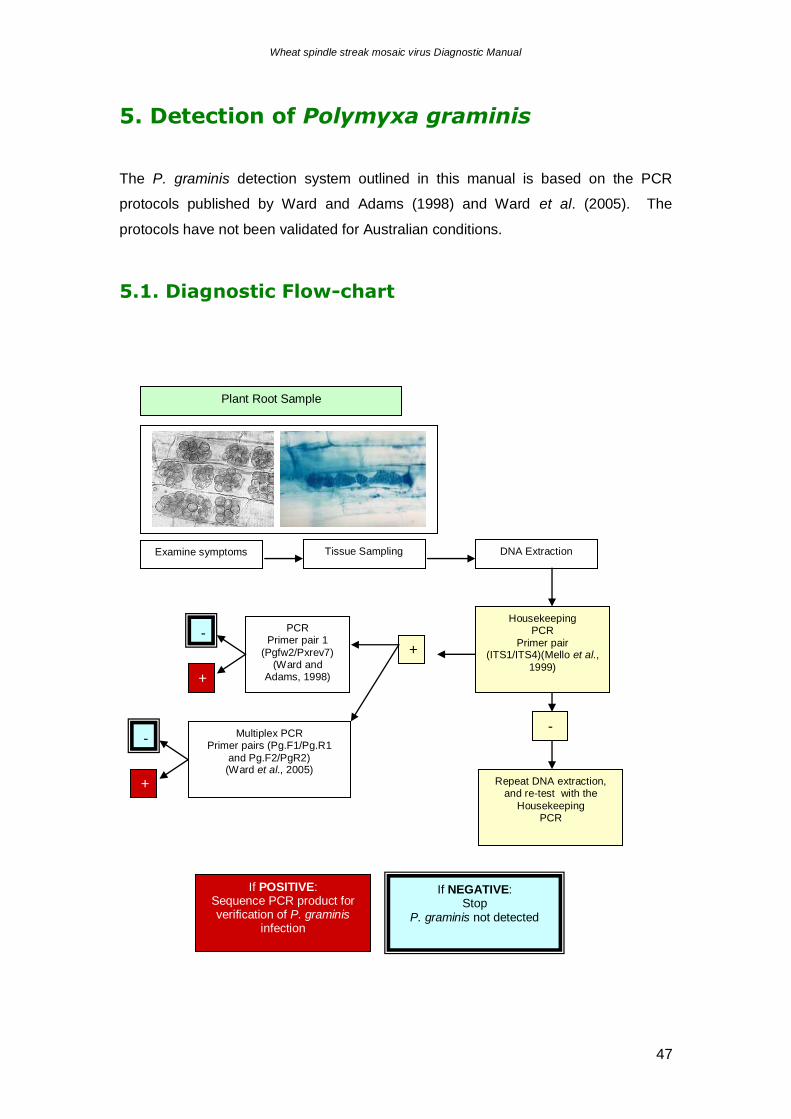
5. Detection of Polymyxa graminis
The P. graminis detection system outlined in this manual is based on the PCR
protocols published by Ward and Adams (1998) and Ward et al. (2005). The
protocols have not been validated for Australian conditions.
5.1. Diagnostic Flow-chart
If POSITIVE: Sequence PCR product for verification of P. graminis
infection
If NEGATIVE: Stop
P. graminis not detected
DNA Extraction
Housekeeping PCR
Primer pair (ITS1/ITS4)(Mello et al.,
1999)
+
Multiplex PCR Primer pairs (Pg.F1/Pg.R1
and Pg.F2/PgR2) (Ward et al., 2005)
PCR Primer pair 1
(Pgfw2/Pxrev7) (Ward and
Adams, 1998)
Examine symptoms Tissue Sampling
-
Repeat DNA extraction, and re-test with the
Housekeeping PCR
+
-
+
-
Plant Root Sample

Wheat spindle streak mosaic virus Diagnostic Manual
48
5.2. Sample collection
5.2.1. Plant
Collect plants suspected of P. graminis infection and wash roots clean of soil under
fast running tap water. Allow to air dry, and store in a cool dry place (Barr, 1979).
5.2.2. Soil
Collect soil samples in a “W” pattern across the field. Take a 1-cup sample of soil
every ten metres. Soil samples can be pooled. To increase the likelihood of
detecting P. graminis, include soil samples collected from the wettest area of the
paddock.
To recover P. graminis from the soil samples, grow T. aestivum seedlings in the
suspect fungus-infected soil (Clover and Henry, 1999). Incubate two-week old
seedlings of T. aestivum in a slurry of 40 g of infected soil at 20oC for 10-14 days.
After incubation, transplant into sterile sand to give a 1:9 dilution of infected soil:sand.
Grow the plants for a further 3 weeks at 20oC, then decrease the temperature to
10oC. Grow the plants for a further 9-12 months. Check roots periodically for P.
graminis infection. Treat collected roots as per section 5.2.1.

Wheat spindle streak mosaic virus Diagnostic Manual
49
5.3. Total DNA extraction
5.3.1. Equipment required
5. 2-20 l, 20-200 l, and 200-1000 l pipettes and sterile tips
6. Autoclave
7. Autoclave bags
8. Balance (at least 2 decimal places)
9. Disposable gloves
10. Microcentrifuge
11. Sterile microcentrifuge tubes
12. Paper towel
13. DNeasy® Plant Mini Kit (QiagenTM)
14. Sharps container
15. Sterile scalpel blades
16. Waterbath or heatblock
17. Weighboats
5.3.2. Method
Follow the protocol of the DNeasy® Plant Mini Kit (QiagenTM), as per the
manufacturer’s instructions.

Wheat spindle streak mosaic virus Diagnostic Manual
50
5.4. Detection of Polymyxa graminis in total DNA
extracts using PCR.
For the reliable detection of P. graminis, total DNA extracts are subjected to three
PCR tests, as outlined below, with primer sequences and PCR protocols listed in
Table 2.
1. Primer pair Pgfwd2 (F) / Pxrev7 (R). This primer pair detects isolates of P.
graminis, with the exception of isolates originating from India, generating a band
of 280 bp (P. graminis type I), and 320 bp (type II) (Ward and Adams, 1998).
2. Multiplex with all Pg primer pairs: Pg.F1 and Pg.F2 (F) / Pg.R1 and Pg.R2
(R). The forward primers are in ITS1 region, and the reverse primers in the ITS2
region. Primer pair Pg.F1/Pg.R1 amplifies a 292 bp fragment only from P.
graminis ribotype I, and primer pair Pg.F2/Pg.R2 a 430 bp fragment only from
ribotype II (Ward et al., 2005).
3. House-keeping gene (Primer pair ITS1 (F) / ITS4 (R). The ITS1/ITS4 primer
pair is designed to amplify a fragment between 500-1000 bp from the nuclear
ribosomal ITS region (Mello et al., 1999). This region is highly conserved
amongst fungi. Consequently, PCR amplification of the ITS region is used as an
internal PCR control to a) determine the quality of the DNA extract, and b)
determine whether the DNA extract contains inhibitors that will interfere with the
activity of the Taq DNA polymerase enzyme. This PCR is particularly important
when confirming the absence of P. graminis in the test sample.
5.4.1. Equipment required
1. 0-2 L, 2-20 l, 20-200 l, and 200-1000 l pipettes and tips
2. 0.2 or 0.5 ml PCR tubes and 1.5 or 2 ml centrifuge tubes
3. Bulb spinner or centrifuge
4. Freezer
5. Gel tanks, rigs, racks and power pack
6. Ice machine
7. Latex gloves
8. Microwave
9. Thermocycler
10. UV transilluminator with camera
Wheat spindle streak mosaic virus Diagnostic Manual
51
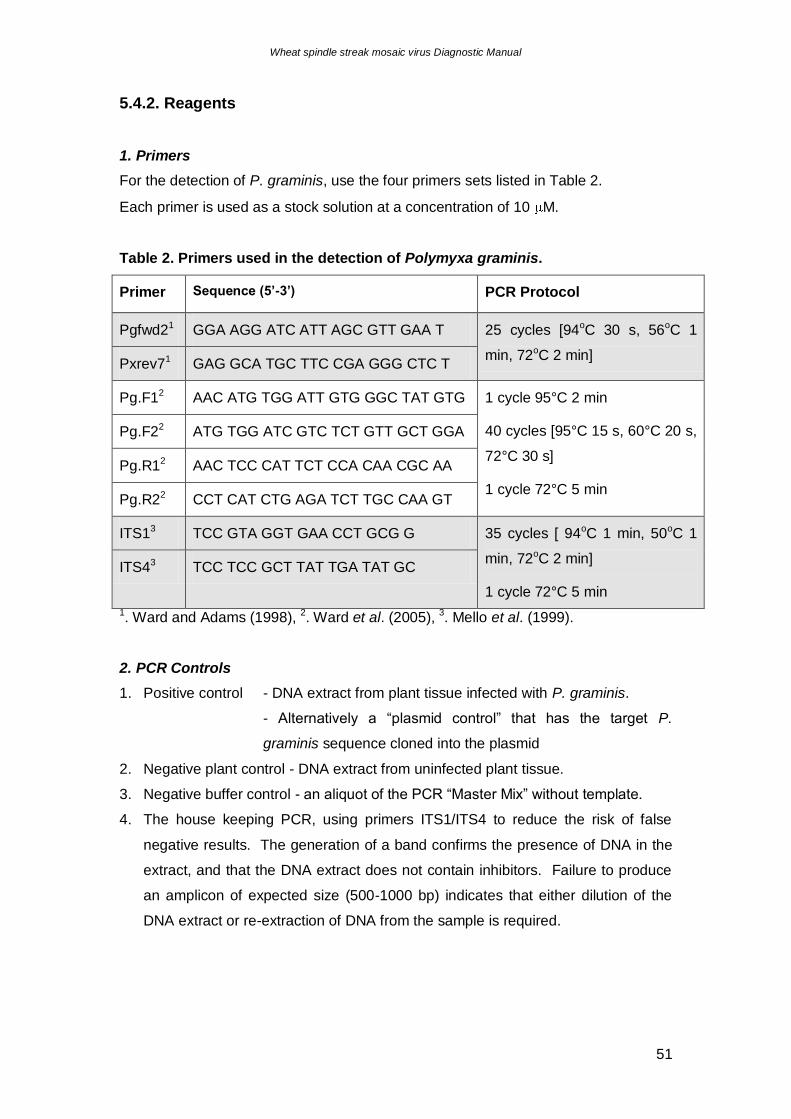
5.4.2. Reagents
1. Primers
For the detection of P. graminis, use the four primers sets listed in Table 2.
Each primer is used as a stock solution at a concentration of 10 M.
Table 2. Primers used in the detection of Polymyxa graminis.
Primer Sequence (5’-3’) PCR Protocol
Pgfwd21 GGA AGG ATC ATT AGC GTT GAA T 25 cycles [94oC 30 s, 56oC 1
min, 72oC 2 min] Pxrev71 GAG GCA TGC TTC CGA GGG CTC T
Pg.F12 AAC ATG TGG ATT GTG GGC TAT GTG 1 cycle 95°C 2 min
40 cycles [95°C 15 s, 60°C 20 s,
72°C 30 s]
1 cycle 72°C 5 min
Pg.F22 ATG TGG ATC GTC TCT GTT GCT GGA
Pg.R12 AAC TCC CAT TCT CCA CAA CGC AA
Pg.R22 CCT CAT CTG AGA TCT TGC CAA GT
ITS13 TCC GTA GGT GAA CCT GCG G 35 cycles [ 94oC 1 min, 50oC 1
min, 72oC 2 min]
1 cycle 72°C 5 min
ITS43 TCC TCC GCT TAT TGA TAT GC
1. Ward and Adams (1998), 2. Ward et al. (2005), 3. Mello et al. (1999).
2. PCR Controls
1. Positive control - DNA extract from plant tissue infected with P. graminis.
- Alternatively a “plasmid control” that has the target P.
graminis sequence cloned into the plasmid
2. Negative plant control - DNA extract from uninfected plant tissue.
3. Negative buffer control - an aliquot of the PCR “Master Mix” without template.
4. The house keeping PCR, using primers ITS1/ITS4 to reduce the risk of false
negative results. The generation of a band confirms the presence of DNA in the
extract, and that the DNA extract does not contain inhibitors. Failure to produce
an amplicon of expected size (500-1000 bp) indicates that either dilution of the
DNA extract or re-extraction of DNA from the sample is required.

Wheat spindle streak mosaic virus Diagnostic Manual
52
3. PCR reagents
- Sterile dH2O
- 1 mM dNTPs
- 10 x concentration buffer
- 25 mM MgCl2
- Taq DNA polymerase
Use all reagents at concentrations recommended by the manufacturer of the Taq
DNA polymerase.
Please refer to section 4.4.2 for the following;
- 5x TBE buffer
- 1x TE solution
- 1% Agarose gel with ethidium bromide
- 6x loading dye
5.4.3. Method
8. Label sterile 0.2 ml centrifuge tubes
9. Prepare PCR “Master Mix” in sterile 1 ml microcentrifuge tube
10. Add 2 l sdH2O to the negative control tube, 2 l test template to each tube, and
DNA extracted from plants suspected to be infected with P. graminis into positive
control tube.
11. Place tubes in thermocycler, and use PCR conditions as listed in Table 2.
12. At completion of PCR, mix 10 l each PCR sample with 5 l loading dye
13. Load samples onto a 1% agarose gel containing ethidium bromide
14. Electrophorese in 0.5 X TBE at 100V
15. Visualise and photograph gel on UV transilluminator.
5.5. DNA Sequencing of PCR Products
As per section 4.5.

Wheat spindle streak mosaic virus Diagnostic Manual
53
Acknowledgments
Plant Health Australia, for providing the funding for this project as part of their
National Diagnostic Protocols Initiative.
The Department of Primary Industries, Victoria, for allocating resources to this
project.
Dr Joanne Luck and Dr Fiona Constable for giving their time to review this document.

Wheat spindle streak mosaic virus Diagnostic Manual
54
References
Adams MJ (2002) Bymovirus Potyviridae In: Tidona CA and Darai G (Eds.) The
Springer Index of Viruses. Springer-Verlag Berlin Heidelberg New York, 832.
Adams MJ, Swaby AG and Macfarlane I (1986) The susceptibility of barley cultivars
to barley yellow mosaic virus (BaYMV) and its fungal vector, Polymyxa graminis.
Annals of Applied Biology, 109: 561-572.
Altschul SF, Madden TL, Scäffer AA, Zhang J, Zhang Z, Miller W and Lipman DJ
(1997) Gapped BLAST and PSI-BLAST: a new generation of protein database
search programs. Nucleic Acids Research, 25: 3389-3402.
Anon (2003) Wheat spindle streak mosaic virus. Kentucky Wheat IPM.
http://www.uky.edu/Agriculture/IPM/scoutinfo/wheat/disease/wssm/wssm.htm
Anon (2005) Towards sustainability of groundnut and cereal production in West
Africa: management of peanut clump virus. International Crops Research Institute for
the Semi-Arid Tropics, http://www.icrisat.org/gt3/DelSpproj.html.
APPD (2005) Australian Plant Pest Database (2005).
http://www.planthealthaustralia.com.au/APPD/
Barr DJS (1979) Morphology and host range of Polymyxa graminis, Polymyxa betae,
and Ligniera pilorum from Ontario and some other areas. Canadian Journal of Plant
Pathology, 1: 85-94.
Bowden RL (2005) Wheat spindle streak mosaic. http://www.oznet.ksu.edu/path-
ext/factSheets/wheat/wheat%20Spindle%20Streak%20Mosaic%20Virus.asp
Braselton (1995) In: Kanyuka K, Ward E and Adams MJ (2003) Polymyxa graminis
and the cereal viruses it transmits: a research challenge. Molecular Plant Pathology,
4: 393-406.

Wheat spindle streak mosaic virus Diagnostic Manual
55
Brunt AA, Crabtree K, Dallwitz MJ, Gibbs AJ, Watson L and Zurcher EJ (eds.) (1996
onwards). Wheat spindle streak mosaic bymovirus, In: Plant Viruses Online:
Descriptions and Lists from the VIDE Database. Version: 20 th August 1996.
http://biology.anu.edu.au/Groups/MES/vide/
Cadle-Davidson L and Bergstrom GC (2004) The effects of postplanting environment
on the incidence of soilborne viral diseases. Phytopathology, 95: 527-534.
Campbell RN (1996) Fungal transmission of plant viruses. Annual Review of
Phytopathology, 34: 87-108.
Carroll JE, Bergstrom GC and Gray SM (1997) Dynamics of wheat spindle streak
mosaic bymovirus in winter wheat. European Journal of Plant Pathology, 103: 313-
321.
Carroll JE, Gray SM and Bergstrom GC (1995) Use of antiserum to a New York
isolate of Wheat spindle streak mosaic virus to detect related bymoviruses from
North America, Europe, and Asia. Plant Disease, 79: 346-353.
Chen J, Sohn A, Chen JP, Lei J, Cheng Y, Schulze S, Steinbiss HH, Antoniw JF and
Adams MJ (1999) Molecular comparisons amongst wheat bymovirus isolates from
Asia, North America and Europe. Plant Pathology, 48: 642-647.
Clover G and Henry C (1999) Detection and discrimination of wheat spindle streak
mosaic virus and wheat yellow mosaic virus using multiplex RT-PCR. European
Journal of Plant Pathology, 105: 891-896.
Driskel BA, Doss P, Littlefield LJ, Walker NR and Verchot-Lubicz J (2004) Soilborne
wheat mosaic virus movement protein and RNA and Wheat spindle streak mosaic
virus coat protein accumulate inside resting spores of their vector, Polymyxa
graminis. Molecular Plant-Microbe Interactions, 17: 739-748.
Gitton F, Diao A, Ducrot O, Antoniw JF, Adams MJ and Maraite H (1999) A two-step
multiplex RT-PCR method for simultaneous detection of soil-borne wheat mosaic
virus and wheat spindle streak mosaic virus from France. Plant Pathology, 48: 635-
641.

Wheat spindle streak mosaic virus Diagnostic Manual
56
Hariri D, Delaunay T, Gomes L, Filleur S, Plovie C and Lapierre H (1996)
Comparison and differentiation of Wheat yellow mosaic virus (WYMV), Wheat spindle
streak mosaic virus (WSSMV) and Barley yellow mosaic virus (BaYMV) isolates
using WYMV monoclinal antibodies. European Journal of Plant Pathology, 102: 283-
292.
Jones R (2004) A national diagnostic protocol for soil-borne viruses of wheat.
Department of Agriculture, State of Western Australia, 1-5.
Kanyuka K, Ward E and Adams MJ (2003) Polymyxa graminis and the cereal viruses
it transmits: a research challenge. Molecular Plant Pathology, 4: 393-406.
Kingsnorth CS, Kingsnorth AJ, Lyons PA, Chwarszczynska DM and Asher MJC
(2003) Real-time analysis of Polymyxa betae GST expression in infected sugar beet.
Molecular Plant Pathology, 4: 171-176.
Ledingham GA (1939) Studies on Polymyxa graminis, n. gen. n. sp., a
plasmodiophoraceous root parasite of wheat. Canadian Journal of Research, 17: 38-
51.
Legreve A, Delfosse P and Maraite H (2002) Phylogenetic analysis of Polymyxa
species based on nuclear 5.8S and internal transcribed spacers ribosomal DNA
sequences. Mycological Research, 106: 138-147.
Linker HM, van Duyn JW, Bambara S, Bailey JE, Lewis WM, Jarrett RE, Kidd KA,
Leath S and Zarnstorff ME (1994) Scouting Small Grains in North Carolina.
http://ipm.ncsu.edu/grain/smgrain521.html, page 9.
Lommel SA, Willis WG and Kendall TL (1986) Identification of wheat spindle steak
mosaic virus and its role in a new disease of winter wheat in Kansas. Plant Disease,
70: 964-968.
Lu X, Kashiwazaki S, Tamura M and Namba S (1998) The 3’ terminal sequence of
RNA1 of Wheat spindle streak mosaic virus Canadian isolate (WSSMV-C). European
Journal of Plant Pathology, 104: 765-768.

Wheat spindle streak mosaic virus Diagnostic Manual
57
MacKenzie DJ, McLean MA, Mukerji S, Green M (1997) Improved RNA extraction
from woody plants for the detection of viral pathogens by reverse transcription -
polymerase chain reaction. Plant Disease 81, 222-226.
Mello A, Garnero L and Bonfante P (1999) Specific PCR-primers as a reliable tool for
the detection of white truffles in mycorrhizal roots. New Phytologist 141, 511-516.
Morales FJ, Ward E, Castano M, Arroyave JA, Lozano I and Adams MJ (1999)
Emergence and partial characterization of rice stripe necrosis virus and its fungus
vector in South America. European Journal of Plant Pathology, 105: 643-650.
Namba S, Kashiwazaki S, Lu X, Tamura M and Tsuchizaki T (1998) Complete
nucleotide sequence of wheat yellow mosaic bymovirus genomic RNAs. Archives of
Virology, 143: 631-643.
Nassuth A, Pollari E, Helmeczy K, Stewart S and KoFalvi SA (2000) Improved RNA
extraction and one-tube RT-PCR assay for simultaneous detection of control plant
RNA plus several viruses in plant extracts. Journal of Virological Methods 90:37-49.
OMAF Staff (2002) Cereals: Soil-borne wheat mosaic virus (SBWMV) Wheat spindle
streak mosaic virus (WSSMV).
http://www.gov.on.ca/OMAFRA/english/crops/pub811/6mosaic.htm
Roget D (2004) Fumigation reveals the soil's real potential.
http://www.grdc.com.au/growers/cd/south/southern_region04044.htm
Rush CM (2003) Ecology and epidemiology of Benyviruses and plasmodiophorid
vectors. Annual Review of Phytopathology, 41: 567-592.
Smith KL (2005) Wheat yellow mosaic. Fact Sheet AC-3-96, Ohio State University
Extension http://ohioline.osu.edu/ac-fact/0003.html
Stromberg EL (2002) Wheat spindle streak.
http://www.ppws.vt.edu/stromberg/smallgrain/biology/wsstreak.html

Wheat spindle streak mosaic virus Diagnostic Manual
58
Subr ZW, Kastirr U and Kuhne T (2002) Subtractive cloning of DNA from Polymyxa
graminis - an obligate parasitic plasmodiophorid. Journal of Phytopathology, 150:
564-568
Tenuta A and Johnson P (2003) Wheat Virus Symptoms Evident!
http://www.gov.on.ca/OMAFRA/english/crops/field/news/croppest/2003/08cpo03a1.htm
Usugi T and Saito Y (1979) Relationship between wheat yellow mosaic virus and
wheat spindle streak mosaic virus. Annals of Phytopathology Society Japan, 45: 397-
400.
Ward E, Adams MJ, Mutasa ES, Collier CR, and Asher MJC (1994) Characterization
of Polymyxa species by restriction analysis of PCR-amplified ribosomal DNA. Plant
Pathology, 43: 872-877.
Ward E and Adams MJ (1998) Analysis of ribosomal DNA sequences of Polymyxa
species and related fungi and the development of genus- and species- specific PCR
primers. Mycological Research, 102: 965-974.
Ward E, Kanyuka K, Motteram J, Kornyukhin D and Adams MJ (2005) The use of
conventional and quantitative real-time PCR assays for Polymyxa graminis to
examine host plant resistance, inoculum levels and intraspecific variation. New
Physiologist, 165: 875-885.

Wheat spindle streak mosaic virus Diagnostic Manual
59
Related Articles
Delfosse P, Reddy AS, Legreve A, Thirumala Devi K, Abdurahman MD, Maraite H
and Reddy DVR (2000) Serological methods for detection of Polymyxa graminis, an
obligate root parasite and vector of plant viruses. Phytopathology, 90: 537-545.
Driskel BA, Hunger RM, Payton ME and Verchot-Lubicz J (2002) Response of hard
red winter wheat to Soilborne wheat mosaic virus using novel inoculation methods.
Phytopathology, 92: 347-354.
Fraaije BA, Lovell DJ, Rohel EA and Hollomon DW (1999) Rapid detection and
diagnosis of Septoria tritici epidemics in wheat using a polymerase chain
reaction/PicoGreen assay. Journal of Applied Microbiology , 86: 701-708
Haufler (1996) In: Clover G and Henry C (1999) Detection and discrimination of
wheat spindle streak mosaic virus and wheat yellow mosaic virus using multiplex RT-
PCR. European Journal of Plant Pathology, 105: 891-896.
Kanyuka K, Lovell DJ, Mitrofanova OP, Hammond-Kosack K and Adams MJ (2004)
A controlled environment test for resistance to Soil-borne cereal mosaic virus
(SBCMV) and its use to determine the mode of inheritance of resistance in wheat cv.
Cadenza and for screening Triticum monococcum genotypes for sources of SBCMV
resistance. Plant Pathology, 53: 154-160
Khan AA, Bergstrom GC, Nelson JC and Sorrells ME (2000) Identification of RFLP
markers for resistance to wheat spindle streak mosaic bymovirus (WSSMV) disease.
Genome, 43: 477-482.
May KJ and Ristaino JB (2004) Identity of the mtDNA haplotype(s) of Phytophthora
infestans in historical specimens from the Irish Potato Famine. Mycological
Research, 108: 471-479.
Meunier A, Schmit JF, Stas A, Kutluk N and Bragard C (2003) Multiplex reverse
transcription-PCR for simultaneous detection of Beet necrotic yellow vein virus, Beet
soilborne virus, and Beet virus Q and their vector Polymyxa betae KESKIN on sugar
beet. Applied and Environmental microbiology, 69: 2356-2360.

Wheat spindle streak mosaic virus Diagnostic Manual
60
Mutasa ES, Chwarszczynska DM, Adams MJ, Ward E and Asher MJC (1995)
Development of PCR for the detection of Polymyxa betae in sugar beet roots and its
application in field studies. Physiological and Molecular Plant Pathology, 47: 303-
313.
Mutasa ES, Ward E, Adams MJ, Collier CR, Chwarszczynska DM and Asher MJC
(1993) A sensitive DNA probe for the detection of Polymyxa betae in sugar beet
roots. Physiological and Molecular Plant Pathology, 43: 379-390.
Ristaino JB, Groves CT and Parra GR (2001) PCR amplification of the Irish potato
famine pathogen from historic specimens. Nature, 411: 695-697.
Zagula KR, Barbara DJ, Fulbright DW and Lister RM (1990) Evaluation of three
ELISA methods as alternatives to ISEM for detection of the Wheat spindle streak
mosaic strain of Wheat yellow mosaic virus. Plant Disease, 74: 974-978.

Wheat spindle streak mosaic virus Diagnostic Manual
61
Appendix 1 – ELISA sample plans
1-45 Samples
1 2 3 4 5 6 7 8 9 10 11 12
A ● 2 6 10 14 18 22 26 30 34 38 42
B ● 2 6 10 14 18 22 26 30 34 38 42
C ● 3 7 11 15 19 23 27 31 35 39 43
D ● 3 7 11 15 19 23 27 31 35 39 43
E ● 4 8 12 16 20 24 28 32 36 40 44
F ● 4 8 12 16 20 24 28 32 36 40 44
G 1 5 9 13 17 21 25 29 33 37 41 45
H 1 5 9 13 17 21 25 29 33 37 41 45
● Positive control ● Buffer control ● Healthy control
1- 27 Samples
1 2 3 4 5 6 7 8 9 10 11 12
A - - - - - - - - - - - -
B - ● 1 4 7 10 13 16 19 22 25 -
C - ● 1 4 7 10 13 16 19 22 25 -
D - ● 2 5 8 11 14 17 20 23 26 -
E - ● 2 5 8 11 14 17 20 23 26 -
F - ● 3 6 9 12 15 18 21 24 27 -
G - ● 3 6 9 12 15 18 21 24 27 -
H - - - - - - - - - - - -
● Positive control ● Buffer control ● Healthy control





























