Embed Size (px)
Citation preview

Week 4: History / philosophy, distance methods
Genome 570
January, 2010
Week 4: History / philosophy, distance methods – p.1/73

Ernst Mayr and George Gaylord Simpson
Ernst Mayr (1905-2005) George Gaylord Simpson (1902-1984)
Week 4: History / philosophy, distance methods – p.2/73

A horse tree drawn by Simpson
Week 4: History / philosophy, distance methods – p.3/73

A phylogeny of the living Craniata
Mam
mal
s
Bird
s
Cro
codi
lians
Sna
kes,
Liz
ards
Tur
tles
Fro
gs
Sal
aman
ders
Lung
fish
Coe
laca
nth
Sha
rks,
Ray
s
Lam
prey
Hag
fish
Am
phio
xus
Mos
t fis
hes
Week 4: History / philosophy, distance methods – p.4/73

Vertebrates are a monophyletic group
Mam
mal
s
Bird
s
Cro
codi
lians
Sna
kes,
Liz
ards
Tur
tles
Fro
gs
Sal
aman
ders
Lung
fish
Coe
laca
nth
Sha
rks,
Ray
s
Lam
prey
Hag
fish
Am
phio
xus
The vertebratesM
ost f
ishe
s
Week 4: History / philosophy, distance methods – p.5/73
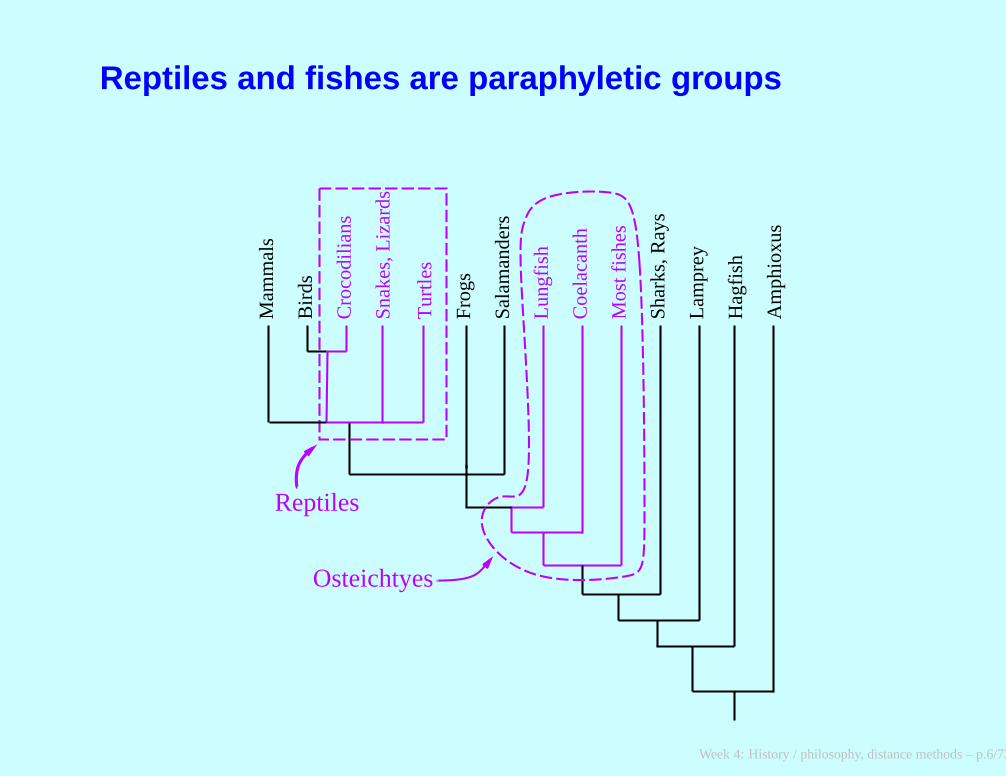
Reptiles and fishes are paraphyletic groups
Mam
mal
s
Bird
s
Cro
codi
lians
Sna
kes,
Liz
ards
Tur
tles
Fro
gs
Sal
aman
ders
Lung
fish
Coe
laca
nth
Sha
rks,
Ray
s
Lam
prey
Hag
fish
Am
phio
xus
Osteichtyes
Reptiles
Mos
t fis
hes
Week 4: History / philosophy, distance methods – p.6/73

Positions on classification as of about 1960Evolutionary systematics. George Gaylord Simpson and ErnstMayr led a movement that allowed non-monophyletic (paraphyletic)groups such as reptiles, on the assumption that groups could beseparated by real differences of rates of evolution (sometimes“grades” rather than “clades”).
Phylogenetic systematics. Willi Hennig advocated purelymonophyletic classification.
Phenetics. Sokal and Sneath advocated making a classificationwithout reference to evolution, using numerical clustering methods
Week 4: History / philosophy, distance methods – p.7/73

Peter Sneath in 1962 and Robert Sokal in 1964
Week 4: History / philosophy, distance methods – p.8/73

The first numerical phylogeny, by Sokal and Michener 1957
Week 4: History / philosophy, distance methods – p.9/73

Cavalli-Sforza and Edwards, 1963; Edwards, 1970
Week 4: History / philosophy, distance methods – p.10/73
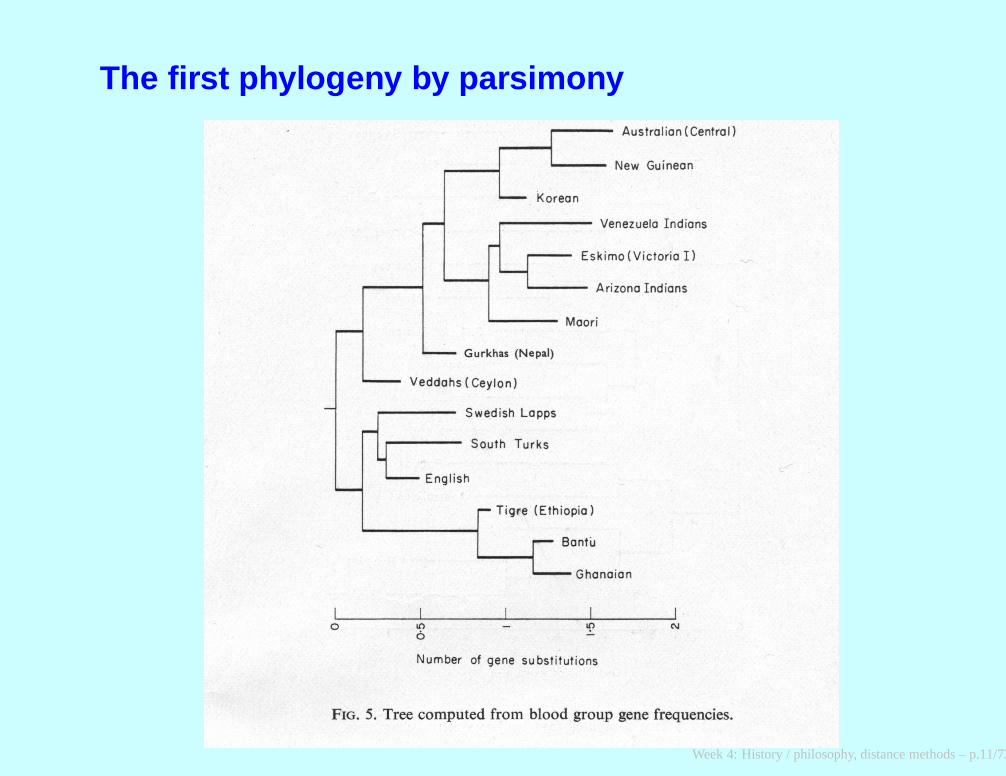
The first phylogeny by parsimony
Week 4: History / philosophy, distance methods – p.11/73
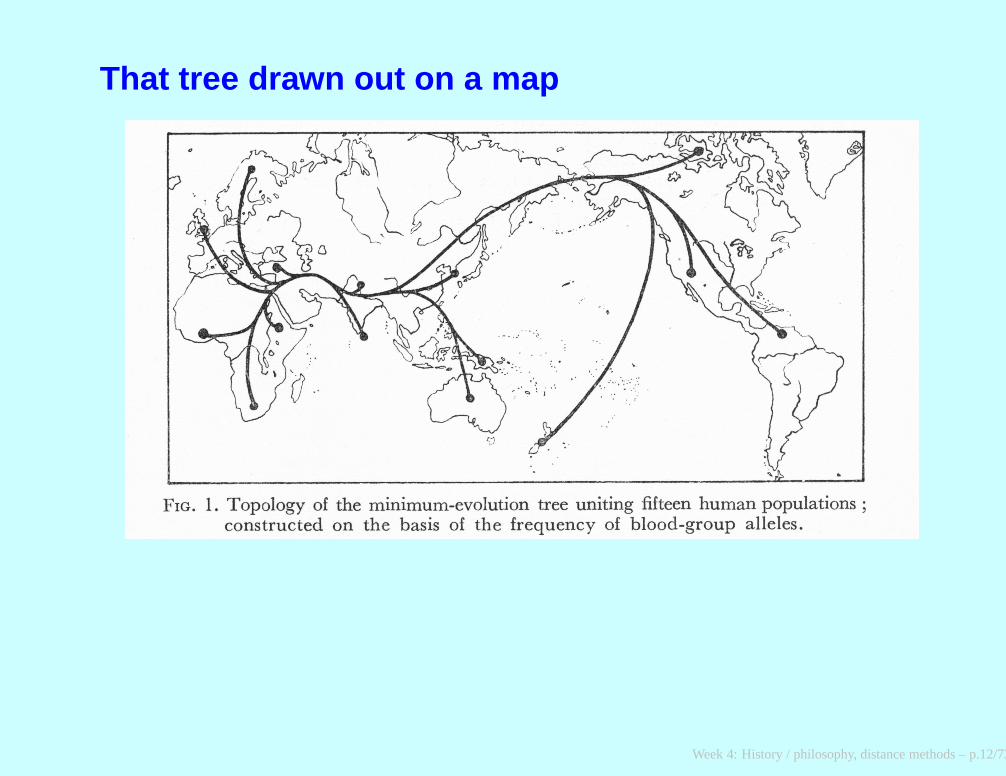
That tree drawn out on a map
Week 4: History / philosophy, distance methods – p.12/73

Camin in the 1970s, one of the Caminalcules
Week 4: History / philosophy, distance methods – p.13/73

J. S. Farris and Arnold Kluge in the 1980s
Week 4: History / philosophy, distance methods – p.14/73

Margaret Dayhoff, 1966; Walter Fitch, 1975
Week 4: History / philosophy, distance methods – p.15/73
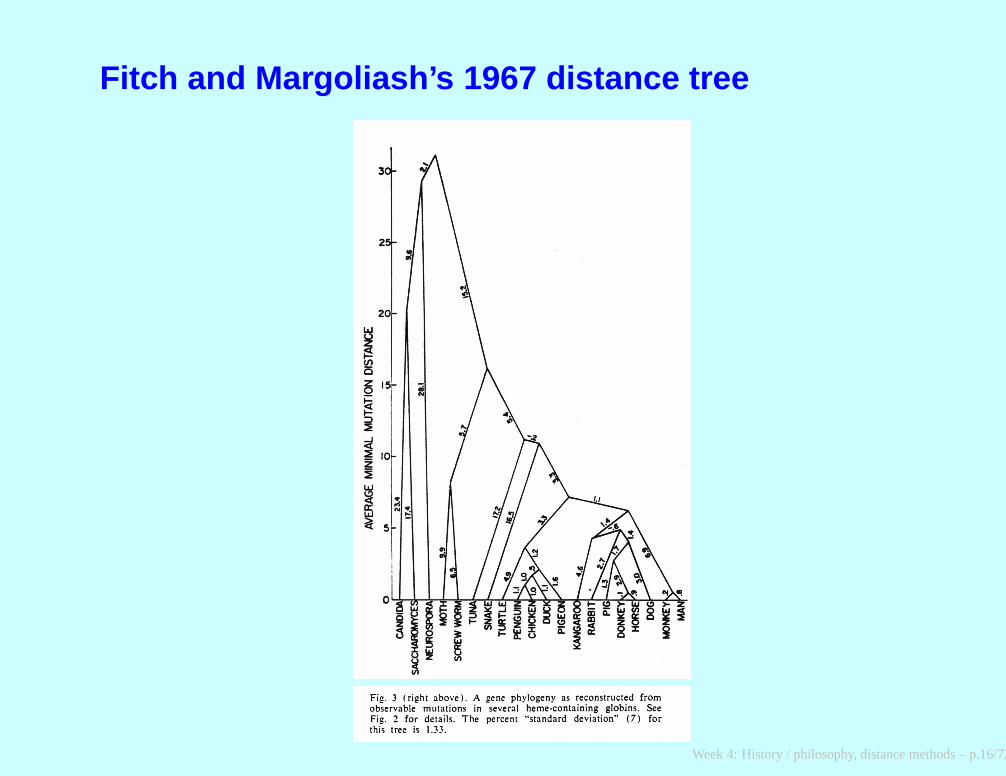
Fitch and Margoliash’s 1967 distance tree
Week 4: History / philosophy, distance methods – p.16/73

Thomas Jukes and Charles Cantor, middle, in the 1990s
Week 4: History / philosophy, distance methods – p.17/73

Jerzy Neyman
Week 4: History / philosophy, distance methods – p.18/73

Willi Hennig (in 1972) and Walter Zimmermann (in 1959)
Willi Hennig (1913-1976) Walter Zimmermann (1890-1980)
Week 4: History / philosophy, distance methods – p.19/73

Hennig and conflict of characters
W. Hennig (1966) says that in the case of homoplasy,
“it becomes necessary to recheck the interpretation of [the]characters"
He also says (1966, p. 121)
“the more certainly characters interpreted as apomorphous(not characters in general) are present in a number of differentspecies, the better founded is the assumption that thesespecies form a morphological group."
Farris, Kluge, and Eckardt (1970) argue that this should be translated as:
“the more characters certainly interpretable as apomorphous..."
Week 4: History / philosophy, distance methods – p.20/73

Did Hennig invent parsimony?
Farris (1983, p. 8):
I shall use the term in the sense I have already mentioned:most parsimonious genealogical hypotheses are those thatminimize requirements for ad hoc hypotheses of homoplasy. Ifminimizing ad hoc hypotheses is not the only connotation of“parsimony" in general useage, it is scarcely novel. BothHennig (1966) and Wiley (1975) have advanced ideas closelyrelated to my useage. Hennig defends phylogenetic analysison the grounds of his auxiliary principle, which states thathomology should be presumed in the absence of evidence tothe contrary. This amounts to the precept that homoplasyought not be postulated beyond necessity, that is to sayparsimony.
Week 4: History / philosophy, distance methods – p.21/73

Hennig’s auxiliary principle
Hennig discusses the case in which “only one character can certainly orwith reasonable probability be interpreted as apomorphous."Hennig (1966, p. 121):
In such cases it is impossible to decide whether the commoncharacter is indeed synapomorphous or is to be interpreted asparallelism, homoiology, or even as convergence. I havetherefore called it an “auxiliary principle" that the presence ofapomorphous characters in different species “is alwaysreason for suspecting kinship [i.e. that the species belong to amonophyletic group], and that their origin by convergenceshould not be assumed a priori" (Hennig 1953). This wasbased on the conviction that “phylogenetic systematics wouldlose all the ground on which it stands" if the presence ofapomorphous characters in different species were consideredfirst of all as convergences (or parallelisms), with proof to thecontrary required in each case.
Week 4: History / philosophy, distance methods – p.22/73

Farris and Kluge on Hennig and parsimony
Unfortunately, AIV is not sufficiently detailed to allow us toselect a unique criterion for choosing a most preferable tree.We know that trees on which the monophyletic groups sharemany steps are preferable to trees on which this is not so. ButAIV deals only with single monophyletic groups and does nottell us how to evaluate a tree consisting of severalmonophyletic groups. One widely used criterion – parsimony –could be used to select trees. This would be in accord withAIV, since on a most parsimonious tree OTUs [tips] that sharemany states (this is not the same as the OTUs’ being describedby many of the same states) are generally placed together. Wemight argue that the parsimony criterion selects a tree most inaccord with AIV by “averaging" in some sense the preferabilityof all the monophyletic groups of the tree. Other criteria,however, may also agree with AIV.
Farris, Eckardt and Kluge, 1970
Week 4: History / philosophy, distance methods – p.23/73

Philosophical frameworks: hypothetico-deductive
Gaffney (1979, pp. 98-99)
In any case, in a hypothetico-deductive system, parsimony isnot merely a methodological convention, it is a direct corollaryof the falsification criterion for hypotheses (Popper, 1968a, pp.144-145). When we accept the hypothetico-deductive systemas a basis for phylogeny reconstruction, we try to test a seriesof phylogenetic hypotheses in the manner indicated above. Ifall three of the three possible three-taxon statements arefalsified at least once, the least-rejected hypothesis remainsas the preferred one, not because of an arbitrarymethodological rule, but because it best meets our criterion oftestability. In order to accept an hypothesis that has beensuccessfully falsified one or more times, we must adopt an adhoc hypothesis for each falsification .... Therefore, in a systemthat seeks to maximize vulnerability to criticism, the addition ofad hoc hypotheses must be kept to a minimum to meet thiscriterion.
Week 4: History / philosophy, distance methods – p.24/73

more from Gaffney
Gaffney (1979)
“the use of derived character distributions as articulated byHennig (1966) appears to fit the hypothetico-deductive modelbest."
Gaffney (1979, p. 98)
“it seems to me that parsimony, or Ockham’s razor, isequivalent to ‘logic’ or ‘reason’ because any method that doesnot follow the above principle would be incompatible with anykind of predictive or causal system."
Week 4: History / philosophy, distance methods – p.25/73

Hypothetico-deductivists on falsification
Eldredge and Cracraft (1980, p. 69) are careful to point out that
“Falsified" implies that the hypotheses are proven false, butthis is not the meaning we (or other phylogenetic systematists)wish to convey. It may be that the preferred hypothesis willitself be “rejected" by some synapomorphies.
Wiley (1981, p. 111):
In other words, we have no external criterion to say that aparticular conflicting character is actually an invalid test.Therefore, saying that it is an invalid test simply because it isunparsimonious is a statement that is, itself, an ad hocstatement. With no external criterion, we are forced to useparsimony to minimize the total number of ad hoc hypotheses(Popper, 1968a: 145). The result is that the mostparsimonious of the various alternates is the most highlycorroborated and therefore preferred over the lessparsimonious alternates.
Week 4: History / philosophy, distance methods – p.26/73

Farris on hypothetico-deductivism
Farris (1983, p. 8):
Wiley [(1975)] discusses parsimony in a Popperian context,characterizing most parsimonious genealogies as those thatare least falsified on available evidence. In his treatment,contradictory character distributions provide putative falsifiersof genealogies. As I shall discuss below, any such falsifierengenders a requirement for an ad hoc hypothesis ofhomoplasy to defend the genealogy. Wiley’s concept is thenequivalent to mine.
Week 4: History / philosophy, distance methods – p.27/73

Philosophical frameworks: Logical-parsimony
Beatty and Fink (1979):
We can account for the necessity of parsimony (or some suchconsideration) because evidence considerations alone are notsufficient. But we have no philosophical or logical argumentwith which to justify the use of parsimony considerations – anot surprising result, since this issue has remained aphilosophical dilemma for hundreds of years.
Week 4: History / philosophy, distance methods – p.28/73

Kluge and Wolf on logical parsimony
Kluge and Wolf (1993, p. 196):
Finally, we might imagine that some of the popularity of theaforementioned methodological strategies and resamplingtechniques, and assumption of independence in the context oftaxonomic congruence and the cardinal rule of Brooks andMcLennan (1991), derives from the belief that phylogeneticinference is hypothetico-deductive (e.g. Nelson and Platnick,1984: 143-144), or at least that it should be. Even the uses towhich some might put cladograms, such as “testing"adaptation (Coddington, 1988), are presented ashypothetico-deductive. But this ignores an alternative, thatcladistics, and its uses, may be an abductive enterprise(Sober, 1988). We suggest that the limits of phylogeneticsystematics will be clarified considerably when cladistsunderstand how their knowledge claims are made (Rieppel,1988; Panchen, 1992).
Week 4: History / philosophy, distance methods – p.29/73

Elliot Sober on falsificationSober (1988, p. 126):
Popper’s philosophy of science is very little help here,because he has little to say about weak falsification. Popper,after all is a hypothetico-deductivist. For him, observationalclaims are deductive consequences of the hypothesis undertest .... Deductivism excludes the possibility of probabilistictesting. A theory that assigns probabilities to various possibleobservational outcomes cannot be strongly falsified by theoccurrence of any of them. This, I suggest, is the situation weconfront in testing phylogenetic hypotheses. (AB)C is logicallyconsistent with all possible character distributions (polarized ornot), and the same is true of A(BC). [Emphasis in the original]
Week 4: History / philosophy, distance methods – p.30/73

Philosophical foundations: Logical probability?
Popper’s corroboration formula
C(h, e, b) =p(e, hb) − p(e, b)
p(e, hb) − p(eh, b) + p(e, b)
whereb = background knowledgeh = hypothesise = evidence (= d, data?)
C(h, e, b) =Prob (d|h) − Prob (d)
Prob (d|h) − Prob (d&h) + Prob (d)
Week 4: History / philosophy, distance methods – p.31/73

Criticisms of statistical inferenceFarris (1983, p.17):
The statistical approach to phylogenetic inference was wrongfrom the start, for it rests on the idea that to study phylogenyat all, one must first know in great detail how evolution hasproceeded.
Kluge (1997a)
“As an aside, the fact that the study of phylogeny is concernedwith the discovery of historical singularities means thatcalculus probability and standard (Neyman-Pearson) statisticscannot apply to that historical science ...."
Week 4: History / philosophy, distance methods – p.32/73

Positions on classification nowadays
Phylogenetic systematics. Willi Hennig advocated purelymonophyletic classification. Now the (strongly) dominant approach.
Evolutionary systematics. Has almost faded away. Its adherentswere reluctant to make it algorithmic.
Phenetics. Although Sokal and Sneath strongly influenced the fieldof numerical clustering, their approach to biological classificationhas few adherents.IDMVM One person (me) takes the view that It Doesn’t Matter VeryMuch, as we use the phylogeny, and, given that, we never use theclassification system. This is widely regarded as a marginalcrackpot view [“A bizarre thumb in the eye to systematists” –Michael Sanderson].
Week 4: History / philosophy, distance methods – p.33/73
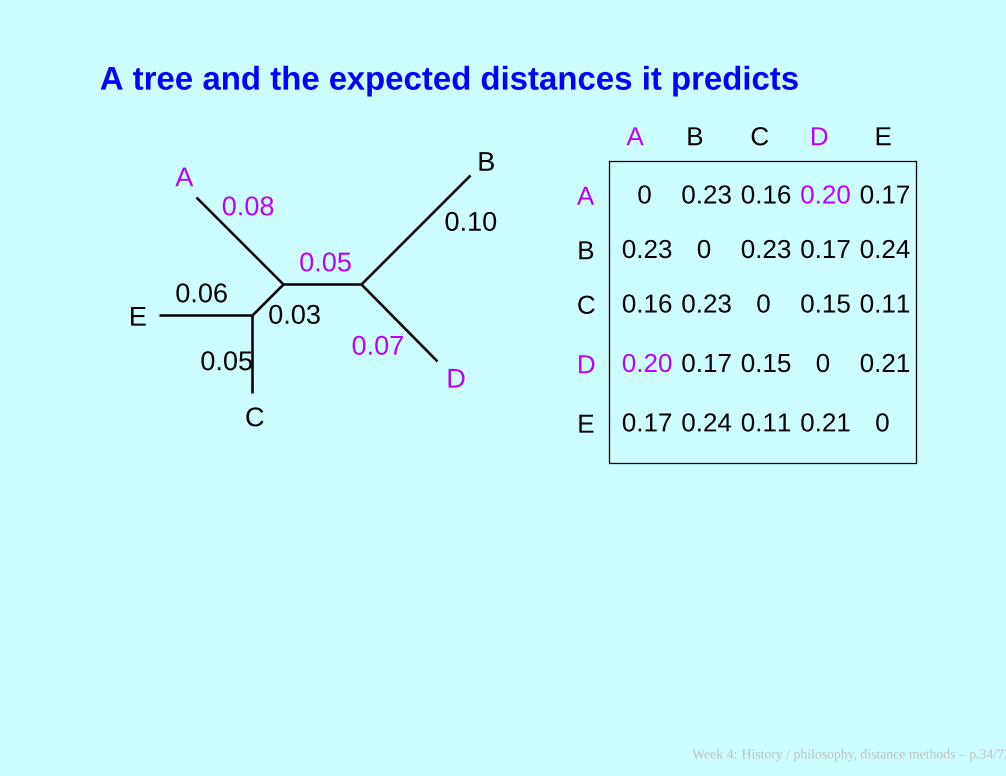
A tree and the expected distances it predicts
A B C D E
A
B
C
D
E
0
0
0
0
0
0.23 0.16 0.20 0.17
0.23 0.17 0.24
0.15 0.11
0.21
0.23
0.16
0.20
0.17
0.23
0.17
0.24
0.15
0.11 0.21
0.10
0.07
0.05
0.08
0.030.06
0.05
A B
CD
E
Week 4: History / philosophy, distance methods – p.34/73

A tree and a set of two-species trees
0.10
0.07
0.05
0.08
0.030.06
0.05
A B
CD
E
A
CD
E
B
The two-species trees correspond to the pairwise distances observedbetween the pairs of species.
Week 4: History / philosophy, distance methods – p.35/73
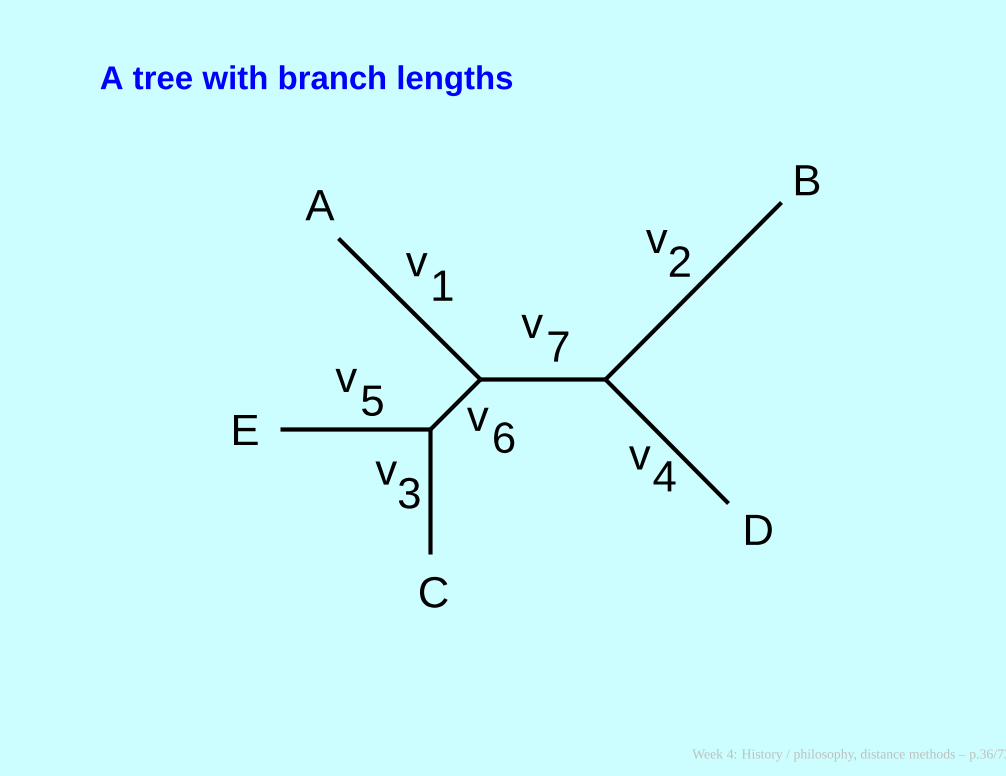
A tree with branch lengths
A B
CD
E
v1v2
v3v4
v5 v6
v7
Week 4: History / philosophy, distance methods – p.36/73

The math of least squares trees
Q =n∑
i=1
∑
j6=i
wij(Dij − dij)2
dij =∑
k
xij,kvk
Q =n∑
i=1
∑
j6=i
wij
(Dij −
∑
k
xij,kvk
)2
.
dQ
dvk
= −2
n∑
i=1
∑
j6=i
wij xij,k
(
Dij −∑
k
xij,kvk
)
= 0.
Week 4: History / philosophy, distance methods – p.37/73

Solving for least squares branch lengths
DAB + DAC + DAD + DAE = 4v1 + v2 + v3 + v4 + v5 + 2v6 + 2v7
DAB + DBC + DBD + DBE = v1 + 4v2 + v3 + v4 + v5 + 2v6 + 3v7
DAC + DBC + DCD + DCE = v1 + v2 + 4v3 + v4 + v5 + 3v6 + 2v7
DAD + DBD + DCD + DDE = v1 + v2 + v3 + 5v4 + v5 + 2v6 + 3v7
DAE + DBE + DCE + DDE = v1 + v2 + v3 + v4 + 4v5 + 3v6 + 2v7
DAC + DAE + DBC
+DBE + DCD + DDE = 2v1 + 2v2 + 3v3 + 2v4 + v5 + 6v6 + 4v7
DAB + DAD + DBC
+DBE + DCD + DDE = 2v1 + 3v2 + 3v4 + 2v5 + 4v6 + 6v7
Week 4: History / philosophy, distance methods – p.38/73

A vector of all distances, stacked
d =
DAB
DAC
DAD
DAE
DBC
DBD
DBE
DCD
DCE
DDE
Week 4: History / philosophy, distance methods – p.39/73

The design matrix and least squares equations
X =
1 1 0 0 0 0 11 0 1 0 0 1 01 0 0 1 0 0 11 0 0 0 1 1 00 1 1 0 0 1 10 1 0 1 0 0 00 1 0 0 1 1 10 0 1 1 0 1 10 0 1 0 1 0 00 0 0 1 1 1 1
XTD =(XTX
)v.
So solution of equations is
v = (XTX)−1 XTD
Week 4: History / philosophy, distance methods – p.40/73

A diagonal matrix of weights
W =
wAB 0 0 0 0 0 0 0 0 00 wAC 0 0 0 0 0 0 0 00 0 wAD 0 0 0 0 0 0 00 0 0 wAE 0 0 0 0 0 00 0 0 0 wBC 0 0 0 0 00 0 0 0 0 wBD 0 0 0 00 0 0 0 0 0 wBE 0 0 00 0 0 0 0 0 0 wCD 0 00 0 0 0 0 0 0 0 wCE 00 0 0 0 0 0 0 0 0 wDE
,
Week 4: History / philosophy, distance methods – p.41/73

Weighted least squares equations
XTWD =
(X
TWX)v,
v =
(X
TWX)−1
XTWD.
Week 4: History / philosophy, distance methods – p.42/73

A statistical justification for least squares
SSQ =n∑
i=1
∑
j6=i
(Dij − E (Dij))2
Var (Dij).
This least squares method
... is what we would get by standard statistical least squaresapproaches if the distances were normally distributed,independently varying, and had expectation and variance as shown
... but they actually aren’t independent in almost all cases (such asmolecular sequences), but ...
... it can be shown that the estimate of the tree will be a consistentestimate in the case of non-independence, just not as efficient
Week 4: History / philosophy, distance methods – p.43/73

The Jukes-Cantor model
A G
C T
u/3
u/3
u/3u/3 u/3
u/3
Week 4: History / philosophy, distance methods – p.44/73

The Jukes-Cantor modelProbability of no event:
e−43ut
Probability of some event:
1 − e−43ut
Probability of changing to C given start at A, have rate u, time t:
Prob (C|A, u, t) =1
4
(1 − e−
43ut)
fraction of sites different:
DS =3
4
(1 − e−
43ut)
.
distance as function of fraction of sites different
D = ut = −3
4ln
(1 −
4
3DS
)
Week 4: History / philosophy, distance methods – p.45/73

Branch length versus fraction of sites different
0
1
0
0.75
0.49
0.7945
per
site
diffe
renc
es
branch length
Week 4: History / philosophy, distance methods – p.46/73

If you don’t correct for multiple changes
A
B
C
0.155 0.155
0.0206
A
B
C
0.20 0.20
0.00
Week 4: History / philosophy, distance methods – p.47/73

The Minimum Evolution method
Kidd and Sgaramella-Zonta (1971) and (independently) Rzhetsky and Nei(1992ff.) came up with this method:
Search through tree space as usual
For each tree estimate branch lenghts by least squares, not allowingnegative branch lengths
Then actually evaluate the tree not by the sum of squares, but by thetotal sum of branch lengths.
This does fairly well, in spite of the mixture of two optimization criteria.
Note that it is not directly related to parsimony, in spite of its name.
Week 4: History / philosophy, distance methods – p.48/73

The UPGMA algorithm
i
j
kDik
Djk
D
(ij)
(ij),k
The UPGMA algorithm
= 1
2. Pick the values of i and j thatare for the smallest of the D ij
3. Make a new group (ij)
+ wj
D(ij),k =
wi + wj
row and column for (ij).
5. Its weight is w
6. The distance between (ij) and another (say k)
7. Delete i and j from the table, put in a
8. If there is only one group left, stop. Otherwisego to step 2.
is computed asi + wj jkw
1. Assign each species weight w i
ik
ij / 2
i
D D
4. Assign time depth D to the connection between i and j.
Week 4: History / philosophy, distance methods – p.49/73
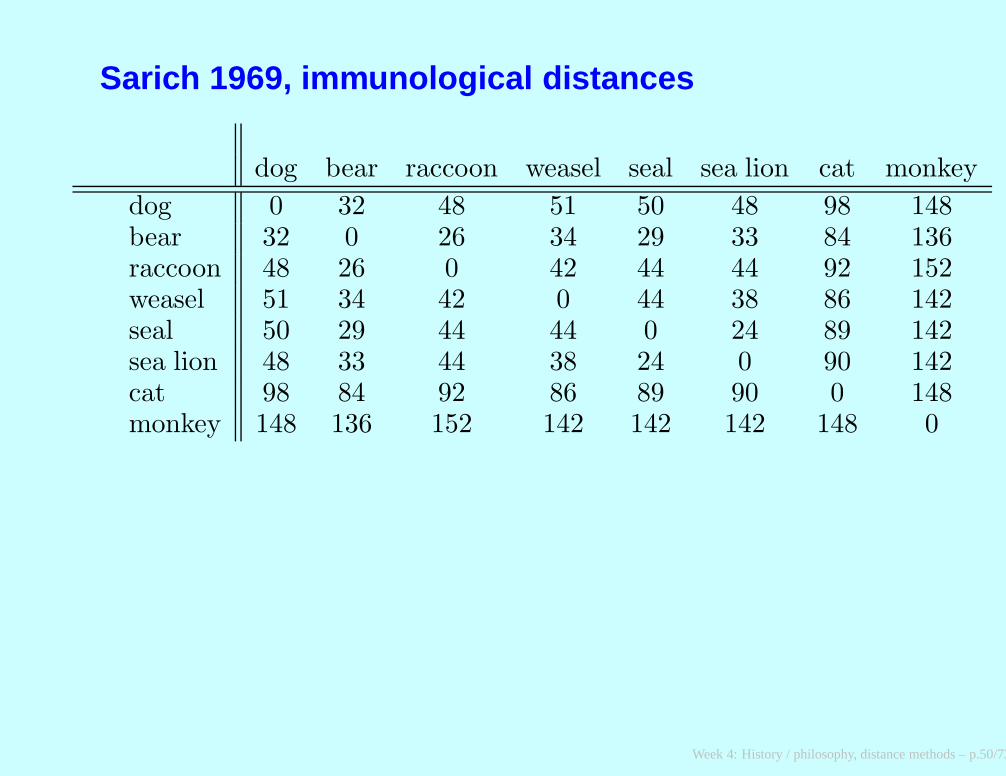
Sarich 1969, immunological distances
dog bear raccoon weasel seal sea lion cat monkey
dog 0 32 48 51 50 48 98 148bear 32 0 26 34 29 33 84 136raccoon 48 26 0 42 44 44 92 152weasel 51 34 42 0 44 38 86 142seal 50 29 44 44 0 24 89 142sea lion 48 33 44 38 24 0 90 142cat 98 84 92 86 89 90 0 148monkey 148 136 152 142 142 142 148 0
Week 4: History / philosophy, distance methods – p.50/73
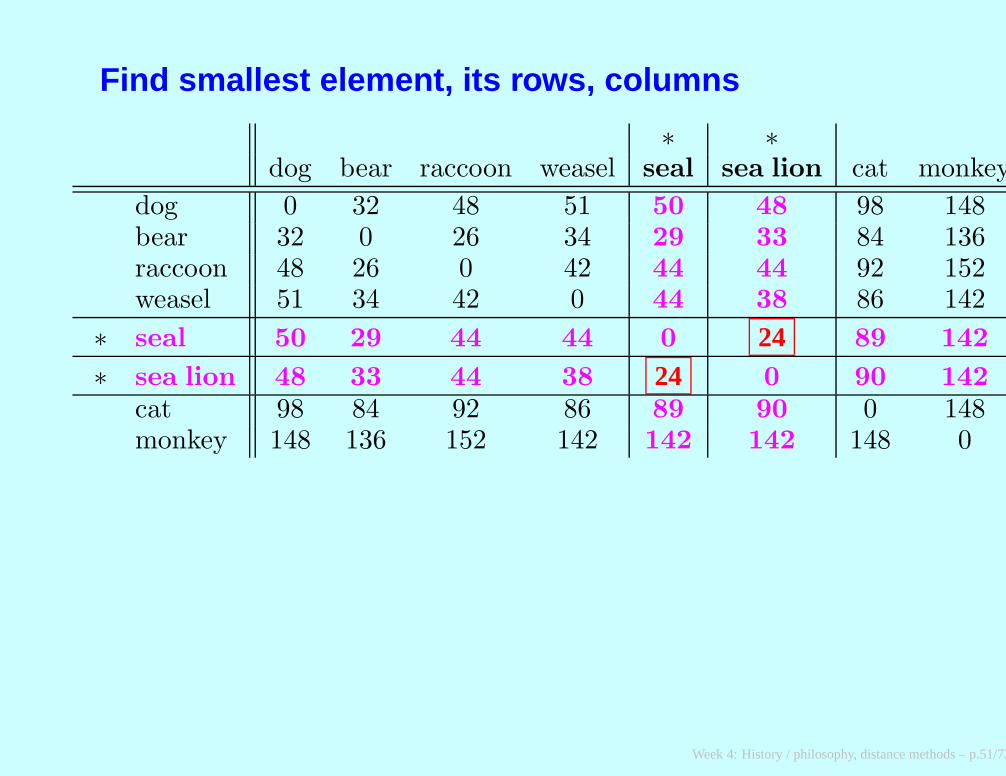
Find smallest element, its rows, columns
∗ ∗dog bear raccoon weasel seal sea lion cat monkey
dog 0 32 48 51 50 48 98 148bear 32 0 26 34 29 33 84 136raccoon 48 26 0 42 44 44 92 152weasel 51 34 42 0 44 38 86 142
∗ seal 50 29 44 44 0 24 89 142
∗ sea lion 48 33 44 38 24 0 90 142
cat 98 84 92 86 89 90 0 148monkey 148 136 152 142 142 142 148 0
Week 4: History / philosophy, distance methods – p.51/73
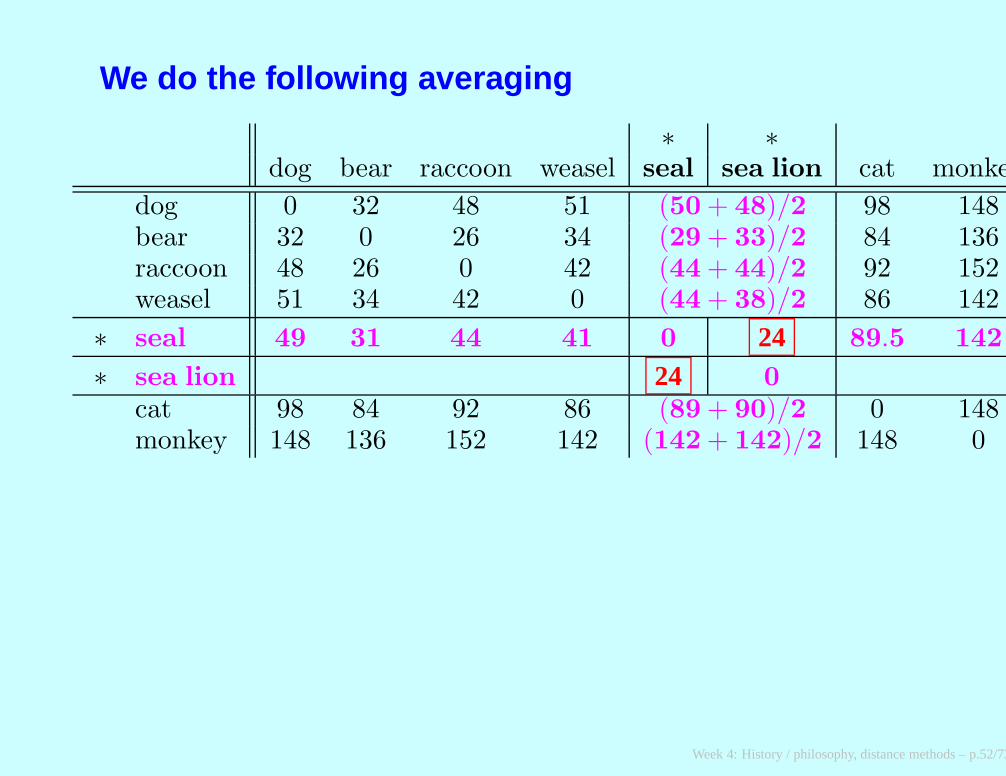
We do the following averaging
∗ ∗dog bear raccoon weasel seal sea lion cat monkey
dog 0 32 48 51 (50 + 48)/2 98 148bear 32 0 26 34 (29 + 33)/2 84 136raccoon 48 26 0 42 (44 + 44)/2 92 152weasel 51 34 42 0 (44 + 38)/2 86 142
∗ seal 49 31 44 41 0 24 89.5 142
∗ sea lion 24 0
cat 98 84 92 86 (89 + 90)/2 0 148monkey 148 136 152 142 (142 + 142)/2 148 0
Week 4: History / philosophy, distance methods – p.52/73

Clustering seal and sea lion
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
12 12
Week 4: History / philosophy, distance methods – p.53/73
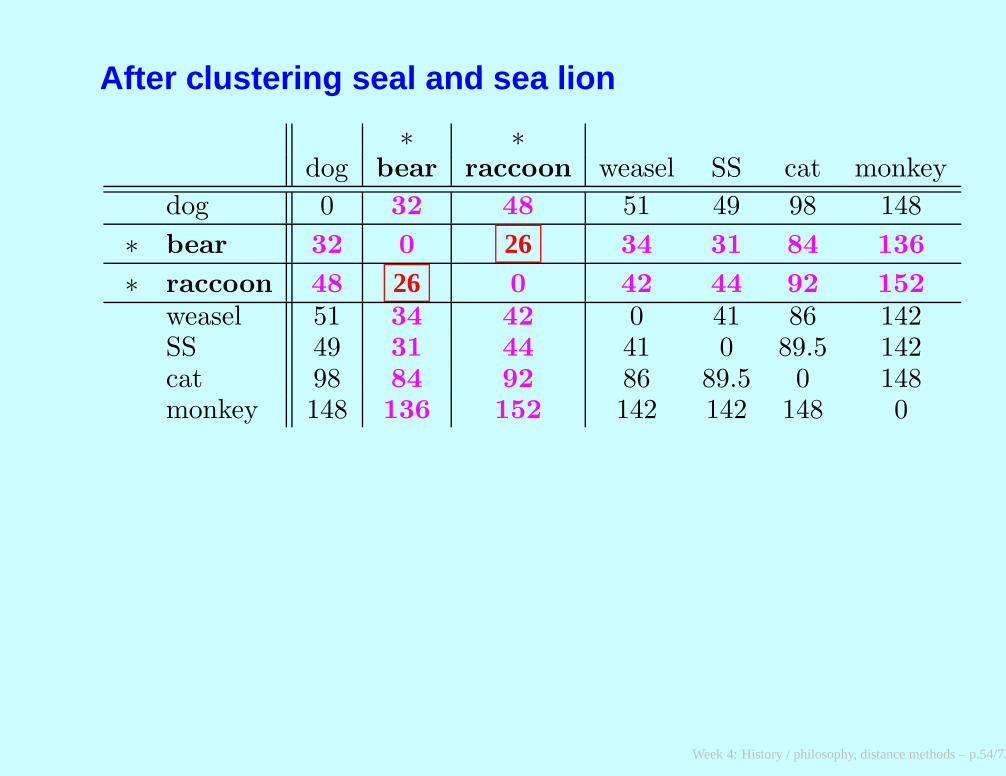
After clustering seal and sea lion
∗ ∗dog bear raccoon weasel SS cat monkey
dog 0 32 48 51 49 98 148
∗ bear 32 0 26 34 31 84 136
∗ raccoon 48 26 0 42 44 92 152
weasel 51 34 42 0 41 86 142SS 49 31 44 41 0 89.5 142cat 98 84 92 86 89.5 0 148monkey 148 136 152 142 142 148 0
Week 4: History / philosophy, distance methods – p.54/73

Clustering bear and racoon
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
13 13 12 12
Week 4: History / philosophy, distance methods – p.55/73
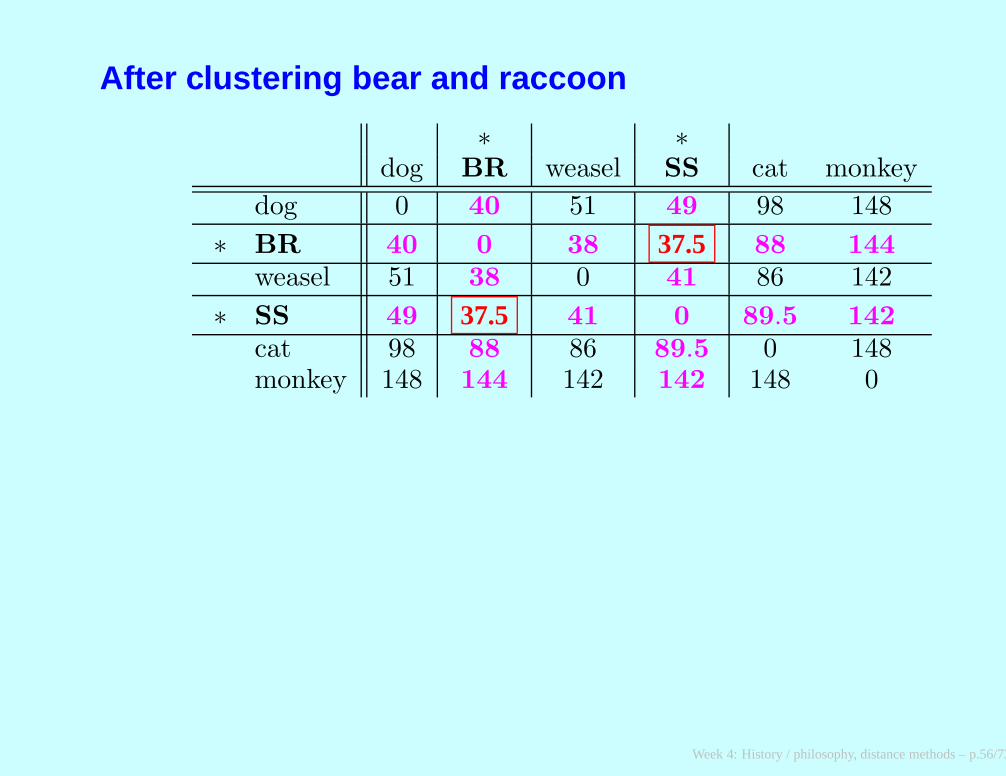
After clustering bear and raccoon
∗ ∗dog BR weasel SS cat monkey
dog 0 40 51 49 98 148
∗ BR 40 0 38 37.5 88 144
weasel 51 38 0 41 86 142
∗ SS 49 37.5 41 0 89.5 142
cat 98 88 86 89.5 0 148monkey 148 144 142 142 148 0
Week 4: History / philosophy, distance methods – p.56/73

Clustering bear-raccoon with seal-sealion
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
5.75 6.75
13 13 12 12
Week 4: History / philosophy, distance methods – p.57/73

After clustering those two clusters
∗ ∗dog BRSS weasel cat monkey
dog 0 44.5 51 98 148
∗ BRSS 44.5 0 39.5 88.75 143
∗ weasel 51 39.5 0 86 142
cat 98 88.75 86 0 148monkey 148 143 142 148 0
Week 4: History / philosophy, distance methods – p.58/73
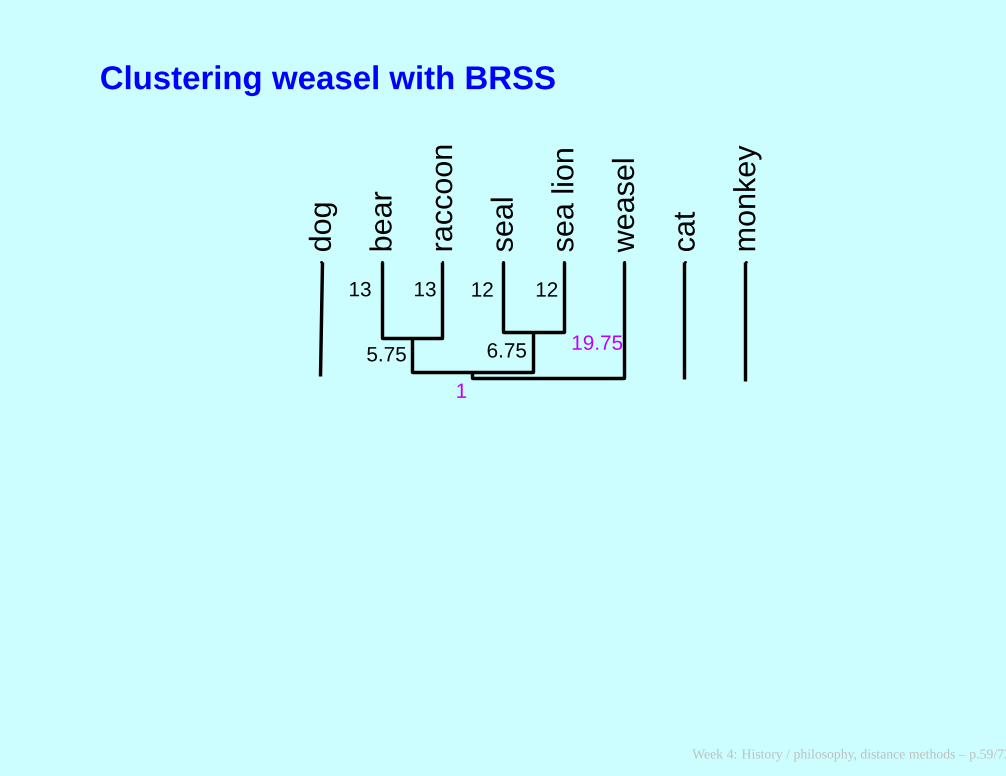
Clustering weasel with BRSS
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
5.75 6.75 19.75
13 13 12 12
1
Week 4: History / philosophy, distance methods – p.59/73

After adding weasel to that cluster
∗ ∗dog BRSSW cat monkey
∗ dog 0 45.8 98 148
∗ BRSSW 45.8 0 88.2 142.8cat 98 88.2 0 148monkey 148 142.8 148 0
Week 4: History / philosophy, distance methods – p.60/73
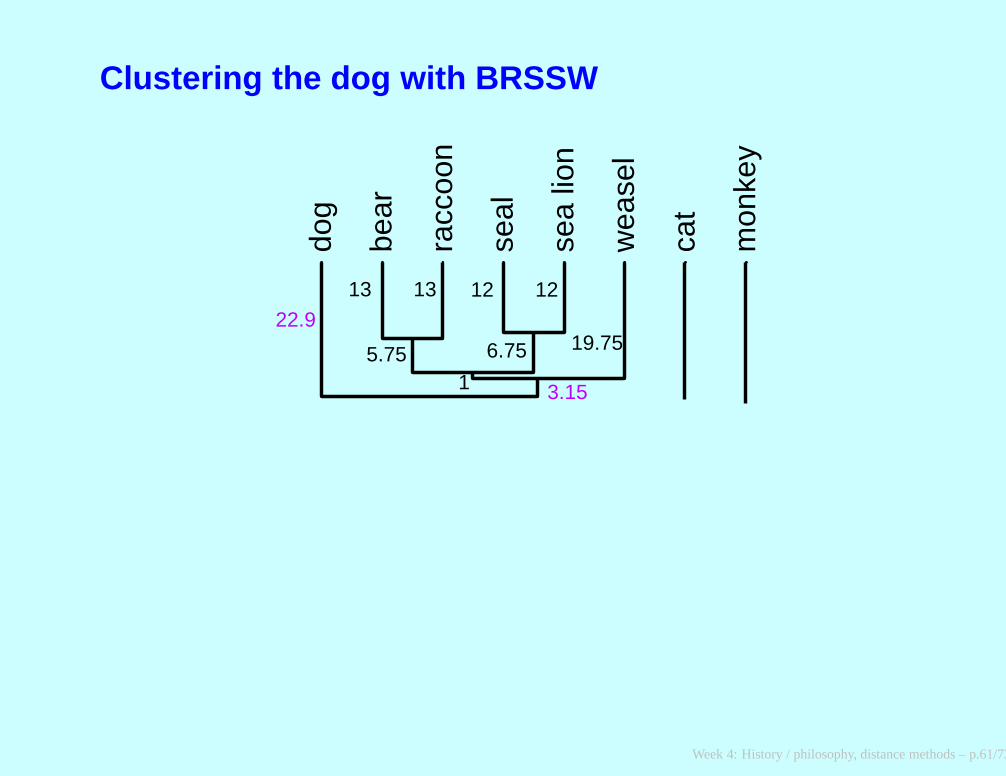
Clustering the dog with BRSSW
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
5.75 6.75 19.75
3.15
22.9
1
13 13 12 12
Week 4: History / philosophy, distance methods – p.61/73

After adding dog to it
∗ ∗DBRWSS cat monkey
∗ DBRWSS 0 89.833 143.66
∗ cat 89.833 0 148
monkey 143.66 148 0
Week 4: History / philosophy, distance methods – p.62/73
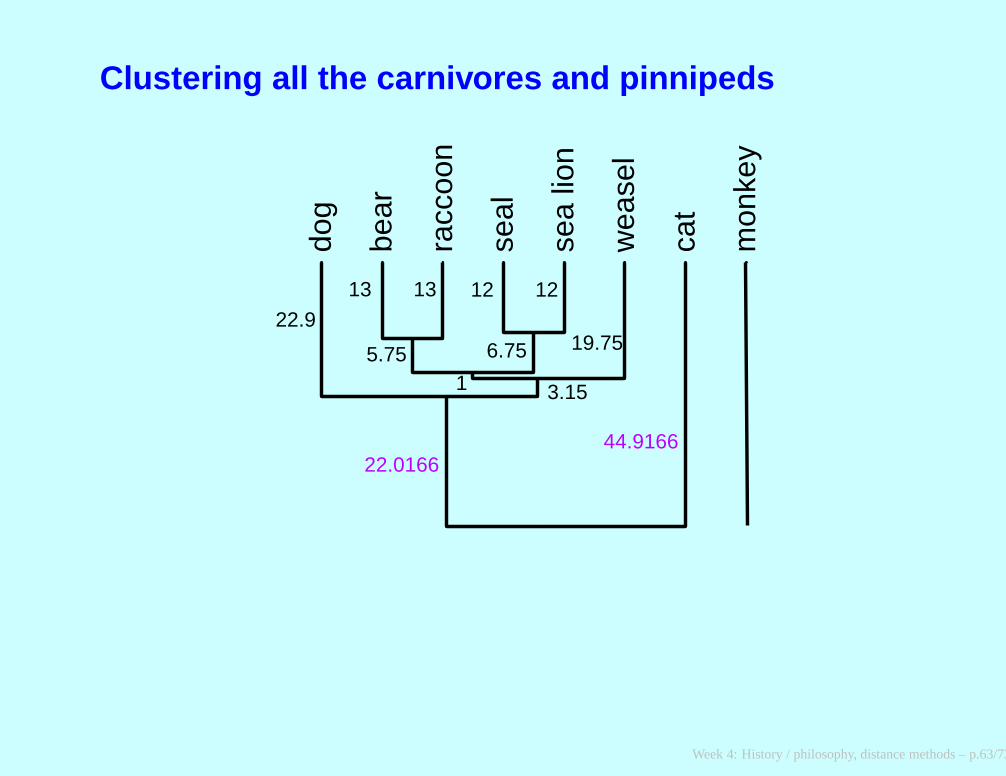
Clustering all the carnivores and pinnipeds
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
5.75 6.75 19.75
3.15
22.9
22.016644.9166
1
13 13 12 12
Week 4: History / philosophy, distance methods – p.63/73

Finally, just monkey remaining
DBRWSSC monkey
DBRWSSC 0 144.2857monkey 144.2857 0
Week 4: History / philosophy, distance methods – p.64/73

The UPGMA tree
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat
mon
key
5.75 6.75 19.75
3.15
22.9
22.016644.9166
27.22619 72.1428
1
13 13 12 12
Week 4: History / philosophy, distance methods – p.65/73
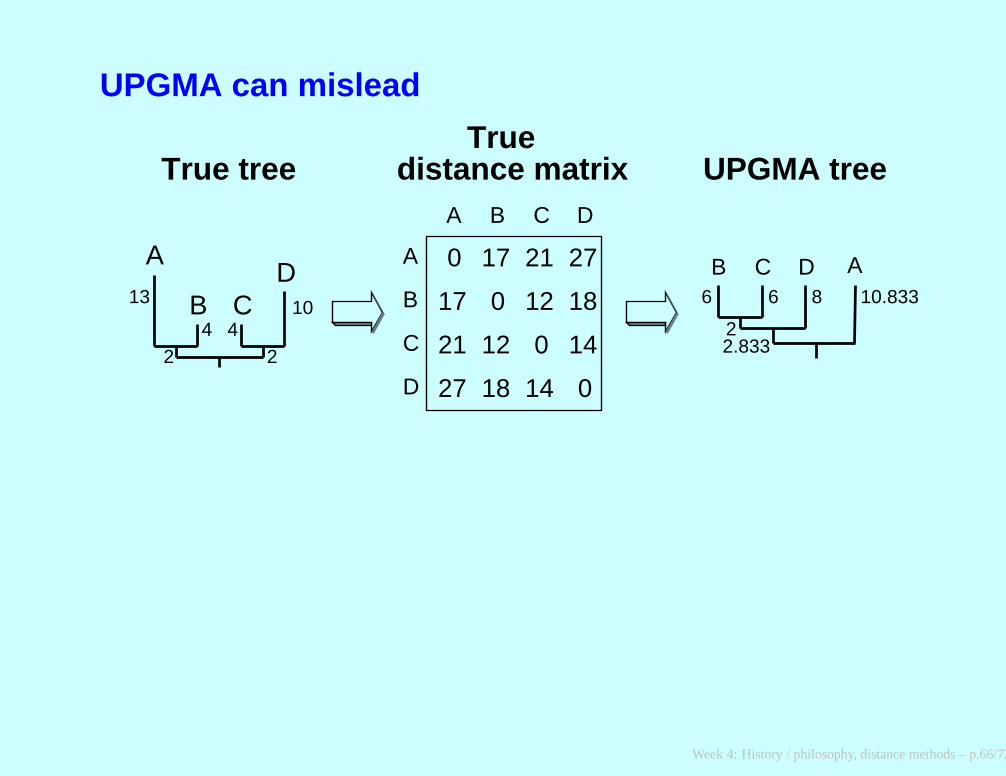
UPGMA can mislead
A
B CD
13
4 410
2 2
A
B
C
D
A B C D
0 17 21 27
17
21
27
0 12 18
12
18
0 14
14 0
B C D A
True tree UPGMA tree
6 6 8 10.833
2.8332
Truedistance matrix
Week 4: History / philosophy, distance methods – p.66/73

Neighbor-Joining
For each tip, compute ui =∑n
j:j6=i Dij/(n − 2). Note that thedenominator is (deliberately) not the number of items summed.
Choose the i and j for which Dij − ui − uj is smallest.
Join items i and j. Compute the branch length from i to the newnode (vi) and from j to the new node (vj) as
vi = 12Dij +
12(ui − uj)
vj = 12Dij +
12(uj − ui)
(continued on next slide)
Week 4: History / philosophy, distance methods – p.67/73

continued ...Compute the distance between the new node (ij) and each of theremaining tips as
D(ij),k = (Dik + Djk − Dij)/
2
Delete tips i and j from the tables and replace them by the newnode, (ij) , which is now treated as a tip.
If more than two nodes remain, go back to step 1. Otherwise,connect the two remaining nodes (say, ℓ and m ) by a branch oflength Dℓm.
Week 4: History / philosophy, distance methods – p.68/73
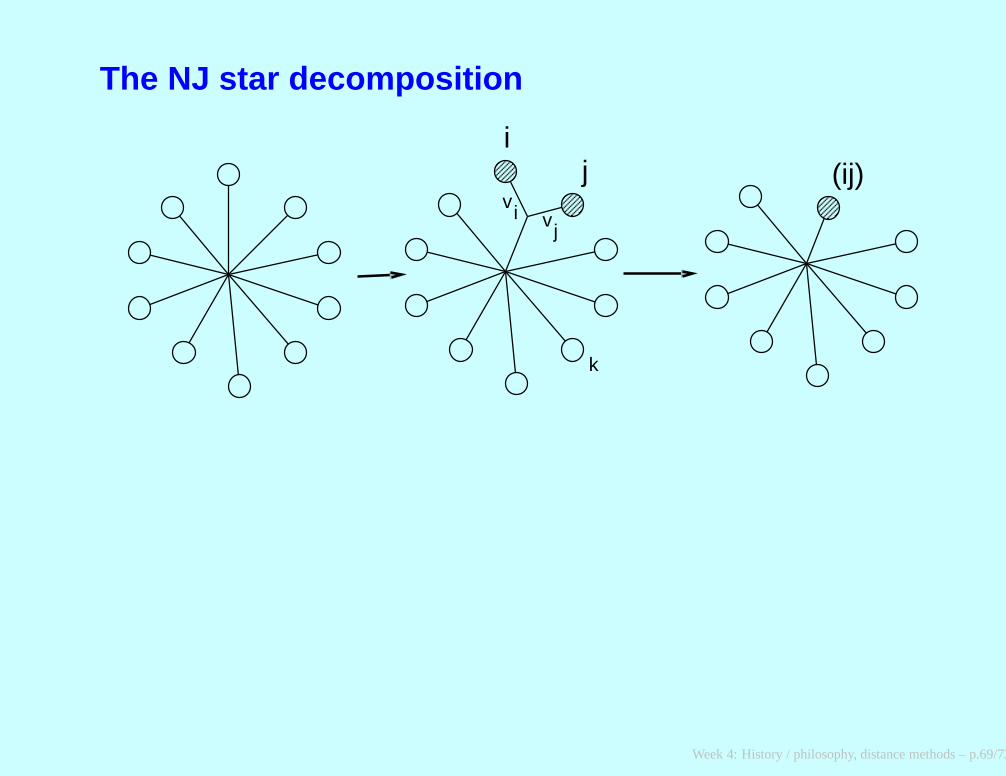
The NJ star decomposition
ij (ij)
vi vj
k
Week 4: History / philosophy, distance methods – p.69/73

The NJ tree on the Sarich data
dog
bear
racc
oon
seal
sea
lion
wea
sel
cat mon
key
19.5625
20.437547.0833
100.9166
1.56253.4375
25.25
1.75
6.87519.125
7.8125
12.35 11.65
Week 4: History / philosophy, distance methods – p.70/73
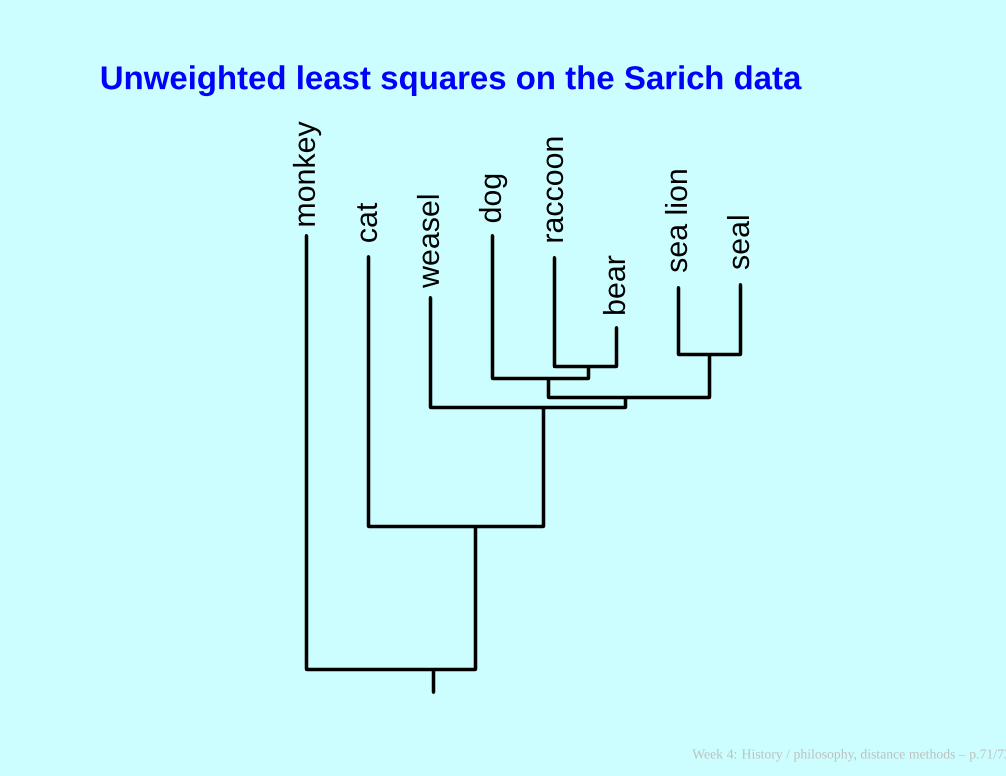
Unweighted least squares on the Sarich data
mon
key
cat
wea
sel
dog
racc
oon
bear se
a lio
n
seal
Week 4: History / philosophy, distance methods – p.71/73
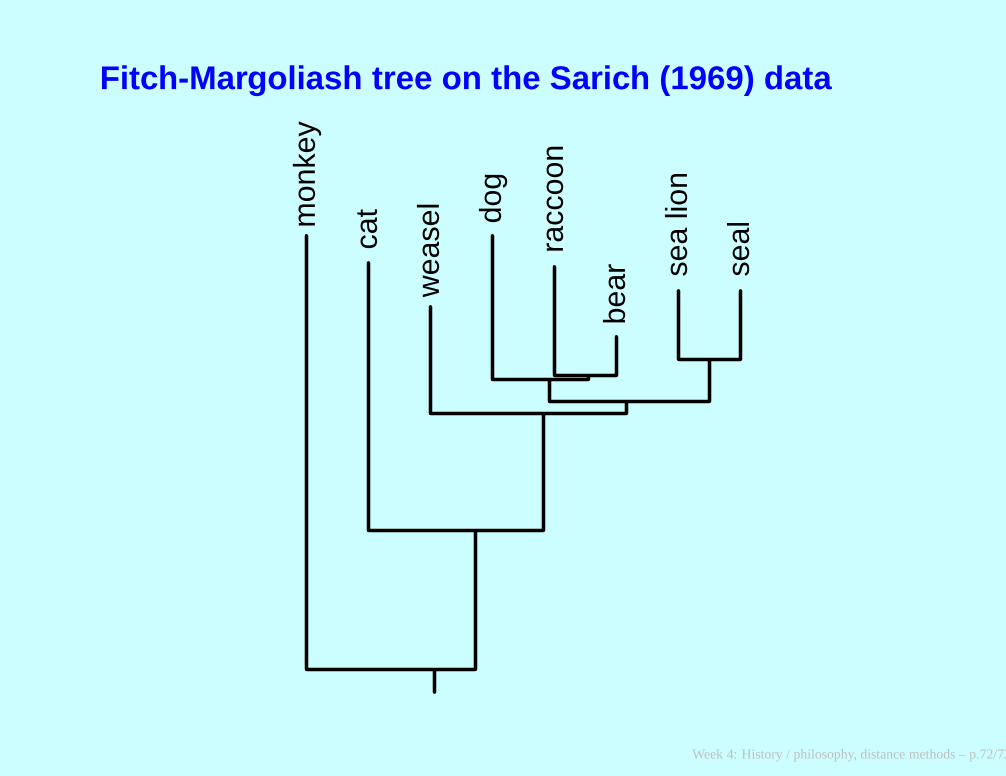
Fitch-Margoliash tree on the Sarich (1969) data
mon
key
cat
wea
sel do
g
racc
oon
bear se
a lio
n
seal
Week 4: History / philosophy, distance methods – p.72/73

Minimum evolution tree on the Sarich (1969) data
mon
key
cat
wea
sel
racc
oon
dog
bear se
a lio
n
seal
Week 4: History / philosophy, distance methods – p.73/73





























