Embed Size (px)
Citation preview

WATER AND LIGHT USE IN MAIZE-MUCUNA INTERCROPPING SYSTEM 1*MWK Mburu, 2JG Mureithi and 1CKK Gachene
1Faculty of Agriculture, University of Nairobi, P .O. Box 29053-000625 Kangemi Kenya; 2Kenya Agricultural Research Institute, P. O. Box 57811 Nairobi Kenya
* Corresponding author: Email: [email protected] ABSTRACT Mucuna (Mucuna pruriens L), a high yielding and N fixing legume is adapted to a range of agro-ecological zones, has the potential to improve maize yield in low input smallholder production systems. Its Integration requires management strategies that optimise complementary resource use in the system. Mucuna planting density and the time of under-sowing in maize were assessed as potential strategies in field experiments conducted between November 2001 and November 2003 at the Kabete Field Station University of Nairobi farm. Maize was inter-cropped with mucuna at two planting densities (44,000 plants ha-1 and 88,000 plants ha-1) and three planting dates (0, 2 and 4 weeks after planting maize). The experimental design was randomised complete block design replicated three times. Maize yield was not affected by inter-planting with mucuna. Mucuna biomass in the intercrop was 25% of that of the sole crop. Delayed planting reduced mucuna biomass significantly but planting density did not have a significant effect. Maize had a comparative advantage over mucuna in light and water capture because it grew faster and taller and had a much larger root system than mucuna and therefore intercropping only reduced mucuna biomass but not maize yield. Planting mucuna and maize at the same time and at high planting density was the best option in the intercrop system when soil water was not limiting. Sole cropped mucuna produced the highest amount of biomass, indicating that maize-mucuna rotation would supply substantial N for a subsequent maize crop. Keywords: Maize, mucuna, planting density, time, yield INTRODUCTION Declining soil fertility due to nutrient mining through crop harvest and severe soil loss with little or no replenishment is a major constraint in smallholder farming in Kenya (Smaling et al., 1997). Smallholder farmers are resource poor and the high cost of chemical fertilizers restricts their use (Bohlool et al., 1992). Opportunities for maintaining and replenishing soil fertility through traditional cropping systems such as fallowing are no longer feasible because of high population pressure on land. The traditional cereal-legume production systems normally result in nutrient mining from the farms (Smaling and Braun, 1996). Integration of high biomass yielding legume cover crops (LCC) has the potential to improve soil nutrient status in the low input maize production systems by reducing soil erosion (Gachene and Haru, 1997), increasing soil organic matter through LCC incorporation (Barthes et al., 2004) and replenishment of soil nitrogen through biologically fixed nitrogen (Sanginga et al.,

1996). Besides, LCC reduce competition for nutrients from weeds by suppressing their growth and population (Vissoh et al., 1998). Integration of LCC into maize-based production system may be through relay planting, inter-planting or rotation systems. Any of these systems would be profitable if the benefits out weigh the costs e.g. if the crop yields in a season following LCC production in rotation system would be equal or greater than the total yield in the entire period the farm is out of production (Wortmann et al., 1994). Yield increases in maize-LCC systems have been documented (Gachene et al., 2002; Mureithi et al., 2005; Eilittä et al., 2004) but farmers may be reluctant to leave the land under an LCC for a season in high land pressure areas (Vissoh et al., 1998). A relay maize-LCC production system would be attractive to farmers in areas where the land is under continuous use for household food supply. For effective relay maize-LCC production system, competition for resources (water, light and nutrients) should be minimal in order to produce high legume biomass without substantially reducing maize yield. The challenge of including LCC in the system is to maximize production of LCC biomass to meet nutrient requirements for a subsequent maize crop without reducing the maize yield in the same season the LCC is grown. Maximal production on a unit area basis in the maize-LCC inter-crop system may be achieved through complementary use of resources and is indicated by yield advantage (Willey, 1979). There are two forms of complementarity, temporal and spatial complementarity. Temporal complementarity may occur when the growth patterns of component crops differ in time such that major demands on resources occur at different times. Growing crops with different growth rates and habits may result in complementary rather than competitive use of growth resources (Ofori and Stern, 1987). Temporal resource use may be achieved by plant phenotype selection (e.g. inter-cropping maize with an initially slow growing LCC) and choice of time of planting. Staggering the relative planting time of the crops in an intercrop would account for temporal differences in resource use by the crops. Light is instantaneously available and instantaneously intercepted for photosynthesis and is probably the most important factor when better temporal use of resources is achieved (Willey, 1979). Spatial complementarity in light use may occur where a tall C4 crop is combined with short C3 because the latter may have higher light use efficiency at low intensity than the former and where the under storey components are adapted to qualitative changes in light (Willey, 1979). Differences in canopy architecture i.e., fairly open tall (e.g. maize) and a denser shorter (legume) canopy may maximise the use of the solar radiation in the system. For example planting cowpea at the same time as maize depressed maize grain yield but effect was reversed when cowpea was planted four weeks after maize (Mureithi et al., 1996), possibly due improved light use. Spatial complementary resource use involves use of resources from different niches or through differences in resource use efficiency. For example, different rooting patterns and canopy architecture of the components would result in the plants exploiting different horizons of soil (for water and nutrients) and light, respectively. Spatial complementarity in light use may occur where a tall C4 crop is combined with short C3 because the former has higher light use efficiency in full light. However, the

radiation use efficiency of the shorter C3 crop is higher when shaded than when grown alone (Squire, 1990). Mucuna (Mucuna pruriens) is an annual legume cover crop (LCC) adapted to a range of agro-ecological zones in Kenya (Mureithi et al., 2003). It produces high biomass, effectively controls weeds and increases maize yields after incorporation (Amede and Kirkby, 2004; Mureithi et al., 2005) and hence has potential as an LCC in a maize-based cropping system. An understanding of the resource use patterns of the components and the trade-offs thereof in terms of the biomass production in the inter-crop system is vital for designing of a successful system. The objective of the study was to determine the effect of planting density and time of inter-planting mucuna with maize on growth, yield, light and water use of maize-mucuna intercrop. METHODOLOGY Experimental site The study was carried out at Kabete Field Station farm, University of Nairobi (1o15’S, 36o44’E; altitude 1942 m). The site has bimodal rainfall of about 1000 mm p.a. with wet seasons occurring between March to June and October to December. The average annual maximum and minimum temperature is 23.9oC and 13.0oC, respectively. The soil is deep, well-drained humic nitisol slightly acidic (soil pH in water 6.4), total soil N was 0.24% and P was 29 ppm (Mehlich) (Nkonge, 2005). The experiment was conducted between November 2001 and November 2003 for three seasons; November 12, 2001 to March 26, 2002 (season 1), April 26, to September 18, 2002 (season 2) and June 17, to November 30, 2003 (season 3). Crop management Maize (H511) spaced at 75 cm x 30 cm, 1 plant/hill (44,000 plants/ha) was inter-cropped with mucuna at two mucuna planting densities and three planting dates. The planting densities were single legume row between maize (D1 - 75 cm x 30 cm; 1 plant/hill) and two rows between maize (D2 – 37.5 cm x 30 cm; 1 plant/hill) while planting dates were 0, 2 and 4 weeks after planting maize; T1, T2 and T3, respectively. Mucuna population was 44,000 plants ha-1 in D1 and 88,000 plants ha-1 in D2. The planting density and planting time were factorially combined and laid out as randomised complete block design, replicated three times. The treatment combinations are summarized in Table 1. Phosphorus was applied (as triple super-phosphate; 46% P2O5) at 20 kg P ha-1 to both mucuna and maize but no N was added. The plot size was 8 m x 5 m. Biomass measurements Mucuna and maize biomass was determined at the end of each growing season from three rows. Maize grain and total dry matter was determined. The mucuna and stover biomass was removed from the plots at the end of each season. Root biomass and root length density were determined from soil cores collected from selected treatments (Table 1) using a root auger (Rowell, 1994). Maize and mucuna roots in the intercrop were separated based on colour in season 2 and 3. Soil water and light measurements

Aluminium neutron access tubes (40 mm internal diameter) for soil moisture determination were installed to 1.3 m depth; one tube/plot in the sole crop and 2 tubes in the intercrop plot. Soil water content was measured (16 counts/s) from 30 cm to 120 cm at a two-week interval using a neutron probe (Dicot, Abingdon, UK). The counts were converted to volumetric water content using calibration equations determined during the experimental period (Nkonge, 2005). Canopy photosynthetically active radiation (PAR) interception was measured using a sunfleck ceptometer (SF 80 Decagon, Pulman, Washington) between 11.30 am and 1.30 pm local time. At least five PAR measurements were taken per plot above and below the maize and mucuna canopies. The analysis of variance was conducted by season using Genstat Release 6.1 (Genstat Committee, 2002) and means separated by the least significant difference (p<0.05) where treatment effects were significant. Table 1: Treatment combinations in the experimental period
--------------------------------------- Seasons ------------------------------------- Cropping System November 2001 to
March 2002 (Season 1) April to September 2002 (Season 2)
June to November 2003 (Season 3)
Sole Sole maize Sole maize* Sole maize* Sole mucuna D1T1* Sole mucuna D1T1* Sole mucuna D1T1 Sole mucuna D2T1* Sole mucuna D2T1* Sole mucuna D2T1* Sole mucuna D1T2 Sole mucuna D2T2 Sole mucuna D1T3 Sole mucuna D2T3* Maize-Mucuna Intercrop D1T1* D1T1* D1T1 D1T2 D1T2* D1T2 D1T3 D1T3* D1T3 D2T1* D2T1* D2T1* D2T2 D2T2* D2T2 D2T3 D2T3* D2T3* *Treatments where root biomass and root length density were determined T1, T2 and T3 refer to mucuna planted 0, 2 and 4 weeks after planting maize respectively. D1 and D2 refer to mucuna population 44,000 plants ha-1 and 88,000 plants ha-1 respectively RESULTS Rainfall The total rainfall in season 1 (Nov 2001 to March 2002), season 2 (April to September 2002) and three (June to November 2003) was 296 mm, 285 and 263 mm, respectively. In season 3, supplemental irrigation water was applied (60 mm) between July-August 2003 to facilitate establishment. Total rainfall received two weeks prior to planting was 115 mm, 150 mm and 55 mm in season 1, 2 and 3, (S1, S2, S3)
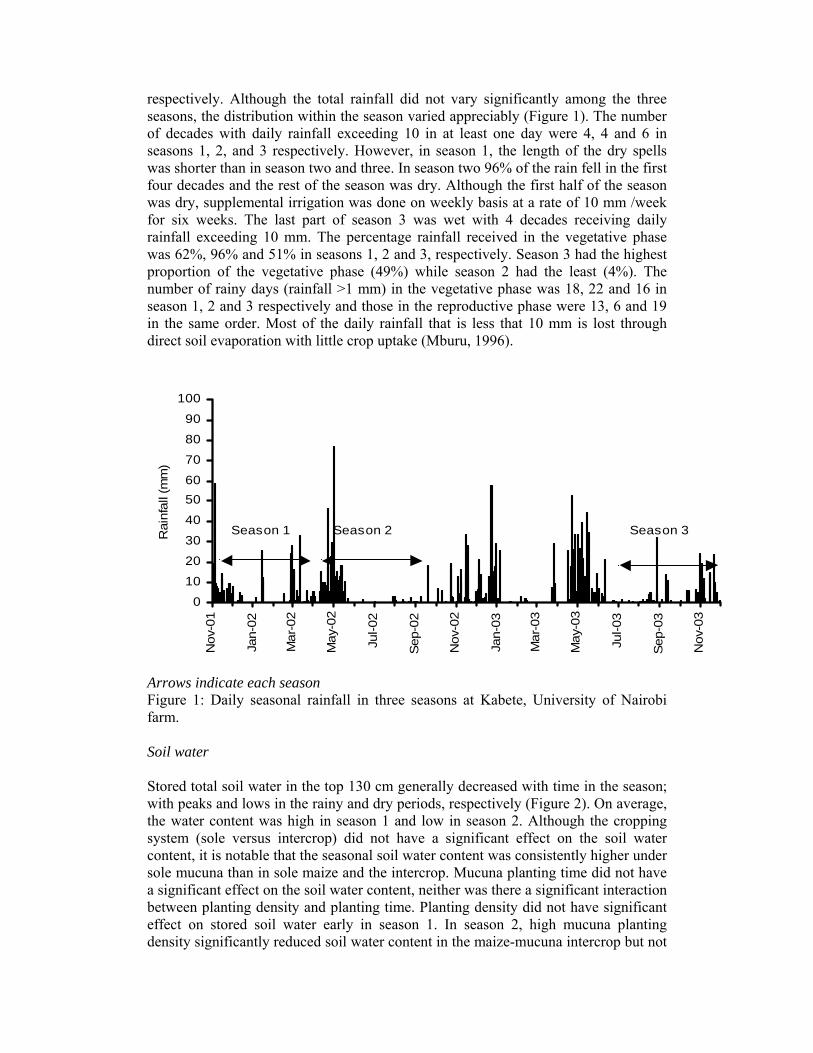
respectively. Although the total rainfall did not vary significantly among the three seasons, the distribution within the season varied appreciably (Figure 1). The number of decades with daily rainfall exceeding 10 in at least one day were 4, 4 and 6 in seasons 1, 2, and 3 respectively. However, in season 1, the length of the dry spells was shorter than in season two and three. In season two 96% of the rain fell in the first four decades and the rest of the season was dry. Although the first half of the season was dry, supplemental irrigation was done on weekly basis at a rate of 10 mm /week for six weeks. The last part of season 3 was wet with 4 decades receiving daily rainfall exceeding 10 mm. The percentage rainfall received in the vegetative phase was 62%, 96% and 51% in seasons 1, 2 and 3, respectively. Season 3 had the highest proportion of the vegetative phase (49%) while season 2 had the least (4%). The number of rainy days (rainfall >1 mm) in the vegetative phase was 18, 22 and 16 in season 1, 2 and 3 respectively and those in the reproductive phase were 13, 6 and 19 in the same order. Most of the daily rainfall that is less that 10 mm is lost through direct soil evaporation with little crop uptake (Mburu, 1996).
0
10
20
30
40
5060
70
80
90
100
Nov
-01
Jan-
02
Mar
-02
May
-02
Jul-0
2
Sep
-02
Nov
-02
Jan-
03
Mar
-03
May
-03
Jul-0
3
Sep
-03
Nov
-03
Rai
nfal
l (m
m)
Season 1 Season 2 Season 3
Arrows indicate each season Figure 1: Daily seasonal rainfall in three seasons at Kabete, University of Nairobi farm. Soil water Stored total soil water in the top 130 cm generally decreased with time in the season; with peaks and lows in the rainy and dry periods, respectively (Figure 2). On average, the water content was high in season 1 and low in season 2. Although the cropping system (sole versus intercrop) did not have a significant effect on the soil water content, it is notable that the seasonal soil water content was consistently higher under sole mucuna than in sole maize and the intercrop. Mucuna planting time did not have a significant effect on the soil water content, neither was there a significant interaction between planting density and planting time. Planting density did not have significant effect on stored soil water early in season 1. In season 2, high mucuna planting density significantly reduced soil water content in the maize-mucuna intercrop but not

in sole mucuna. The largest variation in soil water content among treatments corresponded to the periods with low rainfall. Volumetric water content in the profile varied between 0.25 and 0.45 cm3 cm-3 in the three seasons (Figure 3), which concurred with previous finding where the available soil water was 0.25 to 0.4 of the volumetric water content (Mburu, 1996). Late season soil water content was highest in season three and lowest in season 2. Soil volumetric water content changes at different measurement times and depths indicated that planting density and time did not have consistent effects on profile water content and neither was there significant interaction between the two. Soil water content was lower in the intercrop than in the sole crops in three seasons at lower depths than 70 cm but the difference was only significant in season 1 at 110 cm and 130 cm (Figure 3a). Planting time significantly influenced profile soil water content in the intercrop at 90-130 cm late in the season. Soil water content was significantly lower in early-planted mucuna (T1) than in late-planted mucuna (T3) in season 3 and the former took up more water at 90 cm (Figure 3c). Soil water content under sole maize was lowest at about 50 cm depth but was higher deeper in the profile. Soil water content was slightly higher in the sole mucuna treatment (D2) in season 1. Although this difference was not significant, it may be indicative of microclimate change by mucuna canopy. This may have been as a result of accumulated litter and live biomass insulating the soil surface thus reducing direct evaporation from the soil. This effect was not apparent in season 2, possibly because mucuna canopy growth was restricted by water stress. The pooled seasonal evapotranspiration was 259 mm and 221 mm in season 1 and 2 respectively.

a: Daily rainfall
0
20
40
60
80
100
1-Nov
-01
22-N
ov-01
13-D
ec-01
3-Ja
n-0224
-Jan-0
214
-Feb
-02
7-Mar
-02
28-M
ar-0
218
-Apr
-02
9-May
-0230
-May
-02
20-Ju
n-02
11-Ju
l-02
1-Aug
-02
22-A
ug-02
12-S
ep-02
Date
Rai
nfal
l (m
m)
Season 1Season 2
b: Change in soil water content in season 1 and 2 in the top 130 cm
340
360
380
400
420
440
460
1-Nov
-01
22-N
ov-01
13-D
ec-01
3-Jan
-02
24-Ja
n-02
14-F
eb-02
7-Mar-
02
28-M
ar-02
18-A
pr-02
9-May
-02
30-M
ay-02
20-Ju
n-02
11-Ju
l-02
1-Aug
-02
22-A
ug-02
12-S
ep-02
Date
Sto
red
soil
wat
er (m
m)
Sole MaizeSole MucunaIntercrop S1Intercrop D1 S2intercrop D2 S2
Season 1 Season 2
Figure 2: Seasonal change in (a) daily rainfall and (b) stored soil water top 130 cm in season 1 (Nov 2001-April 2002) and season 2 (May to October 2002). S1 = season 1, S2 = Season 2, D1 = 4.4 plants m-2, D2 = 8.8 plants m-2.

a. Season 1 - 95 DAP
0
20
40
60
80
100
120
140
0.2 0.25 0.3 0.35 0.4 0.45 0.5
Volumetric water content (cm3 cm-3)
Dep
th (c
m)
Sole MaizeSole mucunaIntercrop
b. Season 2 - 144 DAP
0
20
40
60
80
100
120
140
0.2 0.25 0.3 0.35 0.4 0.45 0.5
Volumetric water content (cm3 cm-3)
Dep
th (c
m)
Sole MaizeSole mucunaIntercrop
c. Season 3 - 114 DAP
0
20
40
60
80
100
120
140
0.2 0.25 0.3 0.35 0.4 0.45 0.5
Volumetric water content (cm3 cm-3)
Dep
th (c
m)
Sole MaizePooled IntercropT1T3
Figure 3: Soil water content in intercropped and sole cropped maize and mucuna during the dry spells in season 1, 2 and 3 at 95, 144 and 114 days after planting (DAP) respectively. Standard error bars shown. Root biomass and root length density

Root biomass decreased with increasing soil depth (Table 2). Intercropping reduced maize and mucuna root biomass by approximately 50% in the entire soil profile (Table 2). However, maize had the highest root biomass both in the sole and intercrop. The total root mass of the intercrop components was comparable to that of sole maize. In the intercrop, maize root biomass comprised 70% of the total intercrop root mass in the entire profile. Table 2: Cropping systems effects on maize and mucuna root mass distribution in soil profile in season 3.
---------------------------- Root biomass (mg) ------------------------- Depth (cm) Sole maize Intercropped maize Sole mucuna Intercropped
mucuna 0-20 253.0 152.0 175.0 65.0 20-40 97.0 70.0 70.0 28.3 40-60 56.7 35.0 31.7 15.0 60-90 40.0 11.7 11.7 10.0 Total root length density (RLD) in the soil profile ranged between 130 and 738 m m-3 in the three seasons in sole maize and between 117 to 377 m m-3 in sole mucuna; it was highest in season 1 and lowest in season 2 for both mucuna and maize (Figure 4). There was no significant effect of planting density on mucuna RLD in the sole crop in season 1. Maize and mucuna roots in the intercrop system were not separated is season 1. Sole mucuna had significantly lower RLD than sole maize and the intercrop while intercrop RLD was comparable to sole maize RLD (Figure 4a). There was no significant difference in RLD in all the treatments in season 2 (Figure 4b). There was no significant effect of planting time on the mucuna RLD in both the sole and intercrop systems. Intercropping reduced total RLD of mucuna and maize by 47% and 64%, respectively in season three (Figure 4c). There were no significant differences between root length density in the intercropped maize and sole cropped mucuna in the soil profile in season 3.

Figure 4: Maize (Mz) and Mucuna (Muc) root length density at Kabete, Kenya. Standard error bars indicated. Note scales in season 1 and season 2 & 3 are different. Canopy photosynthetically active radiation (PAR) interception
a: Season 1
0
20
40
60
80
100
0 50 100 150 200 250 300 350 400 450 500
Root length density (m/m3)
Dep
th (c
m)
Mz+Muc intercropSole MucSole Mz
b: Season 2
0
20
40
60
80
100
0 50 100 150 200
Root length density (m/m3)
Dep
th (c
m)
MaizeMucuna
c: Season 3
0
20
40
60
80
100
0 50 100 150 200
Root length density (m/m3)
Dep
th (c
m)
Sole MucIntercrop MucSole MzIntercrop Mz

Canopy photosynthetically active radiation (PAR) interception increased as the season progressed (Figure 5). However, interception was not affected by planting or density nor was there any interaction between the two in all the seasons. Seasonal differences in the amount of PAR intercepted were evident late in the season; the first and third seasons were comparable and second season had the least (Figure 5a). The cropping system significantly influenced PAR interception throughout the season. The maize-mucuna intercrop intercepted more light than either sole maize or mucuna early in the season but sole mucuna intercepted more light late in the season (Figure 5b). Intercropping did not influence the amount of PAR intercepted by maize, which varied between 54% and 60% of the incident PAR in all the seasons. Mucuna under-sown in maize intercepted less than 40% of the incident PAR. The planting time effect was only significant late in the season; delayed mucuna planting reduced PAR interception (Figure 5c). Mucuna planted same time as maize (T1) intercepted the most light while that planted two or four weeks (T2 and T3 respectively) intercepted similar amounts. Partitioning of the late season (134 DAP) total PAR interception between the intercrop components in season 3 showed that the sole mucuna, sole and intercropped maize and mucuna intercepted 94%, 65% and 80%, respectively (Figure 6). Maize and mucuna in the intercrop intercepted 54% and 26%, respectively of the total incident PAR. The amount of light transmitted through the maize over-storey was reasonably high (46% of the total) but the size of the understorey mucuna canopy limited interception. Light supply per se did not limit mucuna growth, possibly the canopy size and the efficiency of utilization of the light for growth.
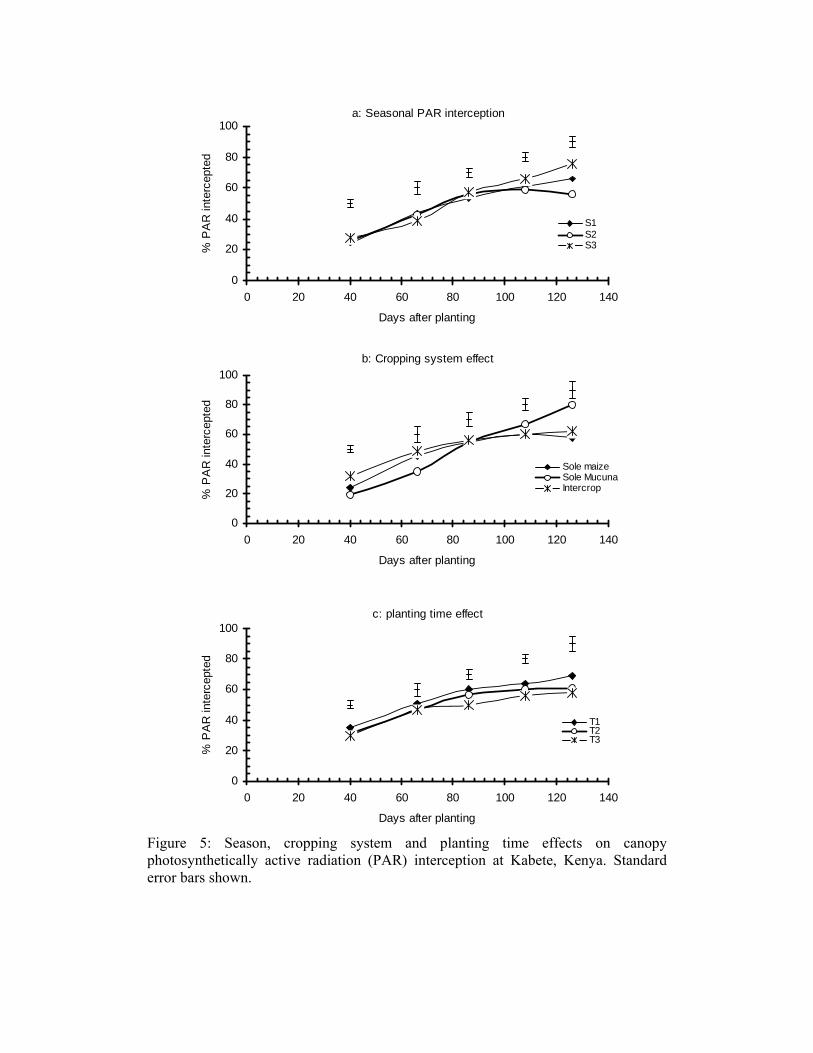
Figure 5: Season, cropping system and planting time effects on canopy photosynthetically active radiation (PAR) interception at Kabete, Kenya. Standard error bars shown.
a: Seasonal PAR interception
0
20
40
60
80
100
0 20 40 60 80 100 120 140
Days after planting
% P
AR
inte
rcep
ted
S1S2S3
b: Cropping system effect
0
20
40
60
80
100
0 20 40 60 80 100 120 140
Days after planting
% P
AR
inte
rcep
ted
Sole maizeSole MucunaIntercrop
c: planting time effect
0
20
40
60
80
100
0 20 40 60 80 100 120 140
Days after planting
% P
AR
inte
rcep
ted
T1T2T3

Figure 6: Canopy photosynthetically active radiation interception in sole and intercropped maize and mucuna. Standard error bars shown. Maize total dry matter (TDM), grain yield and harvest index There was significant variation in total dry matter, grain and harvest index among the three seasons (Table 3) reflecting variation in rainfall among the three seasons. Mucuna planting density did not have a significant effect on total dry matter, maize grain yield and harvest index in the three seasons. Intercropping significantly reduced maize yield in season 1 but not in the other two seasons. The maize yield was 3989 kg ha-1 and 2901 kg ha-1 in the sole and intercrop respectively. Yield was highest in the maize under-sown with mucuna at 4 weeks after maize (2798 kg ha-1) compared with 1947 kg ha-1, 1682 kg ha-1, 2798 at 0 and 2 weeks after maize respectively. The harvest index in season 2 was significantly higher in maize planted at the same time with mucuna and that sown in 2 and 4 weeks latter; the latter were comparable. Total dry matter was significantly lower in season 2 compared to the other two seasons, which were comparable. Grain yield variation was significant in the three seasons. Yield was highest in season 1 and lowest in season 2 (Table 3). The maize harvest index (HI) varied significantly among the three seasons; it was highest in season 1 and lower in season 2 and 3 but comparable between the latter. Table 3: Effect of under sowing mucuna in maize on maize total dry matter, grain yield, harvest index and relative yield total in three seasons
Total dry matter
Grain Yield
Harvest index
Relative yield total Season
-------- (kg ha-1) -------
0
10
20
30
40
50
60
70
80
90
100
0 20 40 60 80 100 120 140
Time (Days after planting)
% P
AR
inte
rcep
ted
Intercrop totalSole MaizeIntercrop mucunaSole mucuna

Season 1 (Nov. 2001 – March 2002) 11,069 3,056 0.36 1.01 Season 2 (April - Sept. 2002) 2,510 487 0.18 0.86 Season 3 (June – Nov. 2003) 12,283 2,232 0.19 1.01 F(p) <0.001 <0.001 <0.001 Ns LSD 1,782 396 0.049 0.17 CV (%) 33 32 32 27
Mucuna under-sowing time did not affect grain yield in season 1 and 2 but in season 3, it was significantly higher where mucuna was sown four weeks after maize. Harvest index was comparable at the three planting times is seasons 1 and 3 but it was higher at in maize sown at the same time with mucuna. Table 4: Maize grain yield, total dry matter (TDM) and harvest index response to mucuna under sowing time (weeks after planting maize). Planting time ---------- Season 1 -------- -------- Season 2 ------ ------- Season 3 -------
Grain Yield
TDM
HI Grain yield
TDM
HI Grain yield
TDM
HI (Weeks after maize)
----- (kg/ha) ---- ----- (kg/ha) ------ ----- (kg/ha) ------
T1 (0 WAM) 2937 9427 0.33 539 2214 0.24 1947 11087 0.21
T2 (2 WAM) 2601 10370 0.33 301 2270 0.12 1682 11616 0.17
T3 (4 WAM) 3164 13487 0.41 549 2865 0.18 2798 14327 0.20
F(p) Ns Ns Ns Ns Ns 0.022 0.031 Ns Ns
LSD 591 3726 0.099 327 723 0.081 806 3389 0.087CV% 16 26 22 34 23 35 28 21 35
WAM - weeks after maize; Ns – not significant Mucuna biomass Mucuna biomass varied among the seasons and the mean biomass was 2020, 656 and 2012 kg ha-1 in the first, second and third seasons, respectively. Biomass was highest in season 1 and lowest in season 2 both in sole and intercropped mucuna (Table 5). Intercropping significantly and drastically depressed mucuna biomass yields compared to sole mucuna yields in the three seasons (Table 5). The reduction was by 69%, 46% and 74 % in seasons 1, 2 and 3, respectively. Averaged over the seasons, higher planting density increased biomass significantly; 575 kg ha-1 and 1096 kg ha-1
in 44,000 plants ha-1 and 88,000 plants ha-1, respectively. There was no interaction between planting density and time of planting in the three seasons. Planting density did not have a significant effect in the sole planted mucuna in the three seasons but biomass was significantly higher in treatments with high planting density in season 1 and 3 in the intercrop (Table 6). High planting density was only beneficial in the intercrop when rainfall did not limit growth. Mucuna may have branched profusely in the sole crop resulting in comparable biomass accumulation in the two planting

densities. High planting density was only beneficial in the intercrop, where branching was possibly more restricted. Table 5: Mucuna biomass (kg ha-1) in sole and intercrop system
Mucuna biomass (kg ha-1) Season Sole crop Intercrop 1. Nov 2001 – Apr 2002 4072 1278 2. May – Sept 2002 998 542 3. June – Nov 2003 3193 830
F test 0.001 0.001 LSD season 1274 333 CV% 37 57
Table 6: Effect of mucuna planting density on mucuna biomass
Plant density Mucuna biomass (kg ha-1) System (Plants/ha) Season 1 Season 2 Season 3 Sole Crop 4072 998 3193 Intercrop 44,000 677 395 656 88,000 1614 690 1004 F test 0.004 0.089 0.058 LSD (0.05%) 528 350 364 CV% 41 61 42 Delayed planting mucuna in maize drastically reduced mucuna biomass, resulting in a strong linear negative correlation between time of planting and mucuna biomass production in the three seasons (Figure 7). Averaged over the three seasons, biomass at 0, 2 and 4 weeks after planting maize in the intercrop was 1192, 815 and 499 kg ha-
1, respectively. The planting time in sole mucuna was only varied in season 3 and the variation was insignificant among the planting dates. In season 1 and 3, time of planting accounted for 98% of the variation in biomass compared to 72% in the drier season 2 (Figure 7). There was no clear relationship between total seasonal rainfall and maize grain and mucuna biomass yield but there was a linear relationship between yield and total rainfall in the vegetative phase (Figure 8).

Figure 7: Effect of time of under sowing mucuna in maize on mucuna biomass. Standard error bars shown.
y = -246x + 1638R2 = 0.9897
y = -115.5x + 764.33R2 = 0.7224
y = -143.25x + 1116.8R2 = 0.9829
0
400
800
1200
1600
2000
0 1 2 3 4 5Weeks after planting maize
Bio
mas
s (k
g ha
-1)
Season 1Season 2Season 3

Figure 8: Maize and mucuna yield response to total rainfall in the maize reproductive phase (tussling to physiological maturity). Data pooled within three seasons. Trend lines in (a) represent the mean values for sole and intercropped maize biomass and grain yield. Both maize and mucuna yield increased with increasing late season rainfall. Intercropping did not influence maize yield response to late season rainfall (3 months after sowing). Approximately 97% and 69% of the maize biomass and grain,
a: Maize biomass and grain
y = 63.219x + 1427.9R2 = 0.9658
y = 14.415x + 545.88R2 = 0.6914
0
2000
4000
6000
8000
10000
12000
14000
0 50 100 150 200
Rainfall (mm)
Yie
ld (k
g/ha
)
Sole Mz biomass
Intercrop Mz biomass
Grain
Maize biomass
Grain
b: Mucuna biomass
y = 2.6836x + 601.28R2 = 0.3878
y = 17.376x + 1116.6R2 = 0.6814
0
1000
2000
3000
4000
5000
0 50 100 150 200
Rainfall (mm)
Bio
mas
s (k
g/ha
)
Sole Muc Biomass
Intercrop Muc biomass

respectively, was accounted for by variation in rainfall (Figure 8). Late season rainfall accounted for 68% and 39% of the variation in sole and intercropped mucuna biomass, respectively. The mucuna rainfall use efficiency was approximately 6 times higher in the sole crop compared to the intercrop, indicating other factors than rainfall reduced biomass production in the intercrop. Maize biomass rainfall use efficiency was 3.6 times higher than sole mucuna rainfall use efficiency. However, the maize biomass rainfall use efficiency was 4.4 times higher than grain use efficiency. The maize-mucuna intercrop biomass relative yield total (RYT) was 1.01, 0.86 and 1.01 in season 1, 2 and 3 respectively. Planting density and time did not have a significant effect on the RYT. DISCUSSION Change in soil profile water content was a function of inputs (rainfall and irrigation) and evapotranspiration. In cases where the daily total rainfall is less than the daily pan evaporation, then the rainfall is lost through direct soil evaporation, hence not effective (available) for crop uptake. The mean daily evaporation in Kabete is 5 mm (Anonymous, 1984). The number of rainy days in season 1, 2, 3 was 31, 28 and 35, respectively but rainfall exceeded 5 mm in 15, 14 and 10 days respectively. This indicates that effective rainfall was available in only a few days in the growing season. Seasonal variation of maize yield and mucuna biomass was largely attributable to differences in late season rainfall amount and distribution (i.e. in the maize reproductive phase) (Figure 8). On average approximately 70% of the rain fell early in the season, thus facilitating reasonable vegetative growth corroborated by PAR interception at 100 days after planting in the three seasons (Figure 5). Plant biomass is a function of radiation intercepted and the efficiency of its conversion into dry matter. The amount of radiation intercepted is a function of canopy size and architecture while the efficiency is influenced by factors like crop photosynthetic pathway and plant nutrition, especially nitrogen (Squire, 1990). Both canopy expansive growth and photosynthesis are reduced by water stress but the former is more sensitive than the latter (Hsiao, 1984). Maize yield is a function of accumulated biomass and partitioning into the grain and neither of these was significantly affected by under-sowing maize with mucuna. Drought reduces maize yield (NeSmith and Ritchie 1992; Rosen and Scott, 1992) through reduction in leaf area index, leaf photosynthetic rate and impaired assimilate translocation into kernels (Schussler and Westgate, 1995) resulting in low harvest index. In maize the critical period is the reproductive phase, which includes the periods immediately before and after anthesis/silking (Calvino et al., 2003). Water stress delays silk emergence relative to pollen shed thereby reducing grain set (Otegui and Slafer, 2004) due to increased ovary abortion (Carcova et al., 2000). Although total maize biomass was comparable in seasons 1 and 3, the harvest index in season 3 was approximately 53% compared to season 1 (Table 3). This indicates that assimilate partitioning played a major role in limiting maize yield. Although 49% of the seasonal rain (158 mm) fell during the reproductive phase in season 3 compared to 38% (12 mm) in season 1, its use for dry matter production in both maize and mucuna

may have been greatly limited by a smaller root system (Figure 5). It is probable a large proportion of the rain may have been lost through evaporation rather than transpiration; evaporative losses of up to 80% of the seasonal rainfall have been documented at Kabete (Mburu, 1996). Intercropping maize and mucuna reduced mucuna biomass drastically possibly through reduced light supply to the under storey mucuna (Figure 6), which may have reduced the rate of photosynthesis (Egli and Yu, 1991), assimilate allocated to the root system (Passioura, 1977) and the amount of nitrogen fixed biologically. Planting density may not have increased mucuna biomass possibly because of its prolific growth. However, the growth occurred when water supply was adequate. A two-week and four-week delay in planting reduced mucuna biomass significantly probably due to strong shading effect by the maize canopy. The decline was higher in drier season than in the wetter season. Root length density plays an important role in water and nutrient uptake (Smucker and Aiken, 1992). Plant ability to transport soil water to the canopy depends on the root size and its distribution in the soil profile relative to the soil water. Root expansion is closely related to leaf development (Squire, 1990). Plants with a large root length density explore a larger soil volume. The maize roots were finer than those of mucuna, an attribute that may have given the former a competitive advantage for water and nutrients (George et al., 1996). It is notable that both RLD and PAR interception (an indicator of leaf area index) were lowest in the driest season and highest in the wettest season, indication of water supply effect on the growth of both. It is probable that although maize RLD was higher than that of mucuna, the latter may be few but efficient in water transport because of a bigger diameter. Maize is a C4 plant that has higher growth rate (16 g m-2 day –1) (Nkonge 2005) than mucuna; 3.4 g m-2 day –1 (calculated from Eilittä et al., 2004) consequently maize canopy established faster and highly shaded mucuna. Maize, which is taller, intercepted more radiation (over 50%) than intercropped mucuna (less than 40%) and hence accumulated more biomass. Mucuna planting density did not have a significant effect on canopy radiation interception possibly due to the ability of mucuna to branch prolifically and the spreading habit could have increased PAR interception at the low planting density. Mucuna planting time effect was more pronounced in season 1 than the other seasons. In the second season canopy expansion, possibly both emergence and expansion may have been restricted by water stress (Mburu et al., 1999). The amount of mucuna biomass produced under the intercrop, when sown at the same time with maize was only about 25% compared to sole crop system (Table 5), which was comparable to 40-70% reduction reported by Gachene and Wortmann (2004). Assuming a tissue nitrogen concentration of 3% in the mucuna (Mureithi and Gitahi, 2004), the mucuna biomass intercrop (approximately 1000 kg ha-1) would contribute approximately 30 kg N ha-1 for the subsequent crop. The N amount may not be sufficient to supply the amount of nitrogen required by the maize crop but may require supplementation with inorganic fertilizer. Although the N effect from intercropped mucuna may not translate into improved grain maize yield (Maobe, 2008), it may improve the soil organic matter and the N supply may help ensure sustainable productivity over several seasons in the future. Relay cropping (delayed planting) would reduce mucuna biomass production to very low levels (less than 1 t

ha-1); which would lead to significantly low increase in maize yield in a subsequent maize crop. Late planted mucuna is likely to produce substantial biomass for incorporation in the subsequent season if there is adequate water supply after maize is harvested. Intercropping would be most profitable in the wet seasons and when the maize is sown at the same time with mucuna. Maize-mucuna rotation system where the two crops are sown as sole crops in different seasons would yield the highest biomass and would contribute significant amount of nitrogen (approximately 80 kg N/ha, assuming an average biomass yield of 2750 kg ha-1 at 3% N concentration). This would be expected to increase maize yield significantly. A rotation would have the added benefit of breaking pest build up and minimizing tillage effects of soil. However, the opportunity cost of foregoing an alternative food crop should be considered. CONCLUSIONS Maize had a comparative advantage over mucuna in light and water capture because it grew faster and taller and had a much larger root system than mucuna. Intercropping only reduced mucuna biomass but not maize biomass in the three seasons; the relative yield ratio was 1 in two seasons out of three. Although the amount of mucuna biomass produced in the intercrop was approximately 25% of that in the sole crop, incorporation of that biomass is likely to improve N supply for maize. Long term application of mucuna biomass plus maize stover would be expected to improve the soil organic matter status in the long run, thus sustain maize production. Planting mucuna at high planting density and at the same time as maize was the best option in the intercrop system at Kabete. Sole cropped mucuna produced the highest amount of biomass, indicating that maize-mucuna rotation would supply large enough amounts of biomass for a subsequent maize crop. REFERENCES Amede, T., and R. Kirkby. 2004. Guidelines for integration of legumes into farming
systems of East African Highlands. In: A. Bationo (Ed). Managing nutrient cycles to sustain soil fertility in Sub-Saharan African Highlands. Academy of Science Publishers. Nairobi.
Anonymous, 1984. Climatological statistics of Kenya. Kenya Meteorological Department. Nairobi.
Barthės, B., A. Azontonde., E. Blanchart., C. Girardin., C. Villenave., S. Lesaint., R. Oliver and C. Feller. 2004. Effect of a legume cover crop (Mucuna pruriens var utilis) on soil carbon in an Utisol under maize cultivation in southern Benin. Soil use and management 20:231-239.
Bohlool, B.B., L.K. Ladha., D.P. Garrity, and T. George. 1992. Biological nitrogen fixation for sustainable agriculture: A perspective. Kluwer Academic Publishers, London.
Calvino P.A., F.H. Andrade and V.O. Sadras. 2003. Maize yields as affected by water availability, soil depth and crop management. Agronomy Journal 95:275-281.
Carcova J., M. Uribelarrea., L. Borras., M.E. Otegui and M.E. Westgate. 2000. Crop physiology and metabolism. Synchronous pollination within and between ears improves kernel set in maize. Crop Science 40:1056-1061.

Eilittä, M., A. Etèka and R. Catsky. 2004. Integrating Mucuna in the maize-based systems of southern Benin. In: (Eds. Eilittä, M; JG Mureithi and R Derpsch.) Green manure/cover crop systems of smallholder farmers: Experiences from tropical and subtropical regions. pp 237-261. Kluwer Academic Publishers. Dordrecht.
Gachene C.K.K. and R. Haru 1997. Controlling seasonal soil loss using purple vetch (Vicia benghalensis). African Crop Science Conference Proceedings. 3:369-373.
Gachene, C.K.K., and C.S. Wortmann. 2004. Green manure/cover crop technology in eastern and central Uganda: development and dissemination. In: (Eds. Eilitta, M; J G Mureithi and R Derpsch.) Green manure/cover crop systems of smallholder farmers: Experiences from tropical and subtropical regions. pp 219-236. Kluwer Academic Publishers. Dordrecht.
Gachene, C.K.K., J.G. Mureithi., F. Anyika and M. Makau. 2002. Incorporation of green manure cover crops in maize based cropping system in semi arid and sub-humid environments of Kenya. In: Participatory Technology Development for Soil management by small Holders in Kenya. Proceedings of the 2nd Scientific Conference of the Soil Management and Legume research network Projects (June 2000, Mombasa Kenya). Published by Kenya Agricultural Research Institute. Pp 145-151.
GenStat Committee 2002. GenStat Release 6.1 Reference Manuals. Published by VSN International, Oxford, UK.
George, S.J., B.M. Kumar., P.A. Wahid and N.V. Kamalan. 1996. Root competition for phosphorous between the tree and herbaceous components of silvo-pastoral systems in Kerala, India. Plant and Soil. 179:189-196.
Maobe, S.N. 2008. Maize response to mucuna green manure. PhD thesis. University of Nairobi.
Mburu, M.W.K. 1996. The effects of irrigation, fertilizer nitrogen and planting density on bean (Phaseolus vulgaris) yield under different weather conditions. PhD thesis. University of Reading.
Mburu, M.W.K., L.P. Simmonds and C.J. Pilbeam. 1999. Bean canopy response to irrigation, nitrogen fertilizer and planting density under temperate and tropical conditions. East African Agriculture and Forestry Journal. 65:7-20.
Mureithi J.G., and Gitahi F.M., 2004. Legume Screening Database and instructional manual. Legume Research Network Project. Kenya Agricultural Research Institute, pp 23.
Mureithi, J.G., C.K.K. Gachene, and J. Ojiem. 2003. The role of green manure legumes in smallholder systems in Kenya: The Legume Research Network Project. Tropical and Subtropical Agroecosystems. 1:57-70.
Mureithi, J.G., C.K.K. Gachene and J.W. Wamuongo. 2005. Participatory evaluation of residue management effects of green manure legumes on maize yield in central Kenya highlands. Journal of Sustainable Agriculture, 25: 49-68.
Mureithi, J.G., R.S. Taylor and W. Thorpe. 1996. Effect of the application of dairy slurry and intercropping with cowpea on the performance of maize in coastal lowland Kenya. African Crop Science Journal 4:315-324
NeSmith, D.S. and J.T. Ritchie. 1992. Effects of soil water-deficits during tassel emergence on development and yield components of maize (Zea mays L.). Field Crops Research 28:251-256.
Nkonge, I.G. 2005. Light, water, and nitrogen use in low input of maize pigeon pea intercrop in sub-humid Kenya. MSc. Thesis. University of Nairobi.

Ofori, F., and Stern. W.R. 1987. Cereal-legume intercropping systems. Advances in Agronomy 41:41-90.
Otegui M.E. and G.A. Slafer. 2004. Increasing cereal yield potential by modifying developmental traits. "New directions for a diverse planet". In: Proceedings of the International Crop Science Congress, 26 Sep-1 Oct 2004, Brisbane, Australia (www.cropscience.org.au) 1-11
Passioura, J.B. 1977. Grain yield harvest index and water use of wheat. Journal of Australian. Institute of Agricultural Science. 43: 117-120
Rosen S. and L. Scott. 1992. Famine grips sub-Saharan Africa. Agricultural Outlook 191:20-24.
Rowell, D.L 1994. Soil science: Methods and applications. 350 pp. John Wiley and Sons, New York, USA.
Sanginga N., B. Ibewiro., P. Hougnandan., B. Vanlauwe., and J.A. Okogun. 1996. Evaluation of symbiotic properties and nitrogen contribution of mucuna to maize growth in the derived savannas of West Africa. Plant and Soil. 179, 119–129.
Schussler J.R. and M.E Westgate. 1991. Maize kernel set at low water potential. II. Sensitivity to reduced assimilates at pollination. Crop Science 31:1196-1203.
Schussler, J.R. and M.E. Westgate. 1995. Assimilate flux determines kernel set at low water potential in maize. Crop Science 35:1196-1203.
Smaling, E.M.A, and A.R. Braun. 1996. Soil fertility research in Sub Saharan Africa: New dimensions, new challenges. Communications in Soil Science and Plant Analysis. 27:356-386
Smaling, E.M.A., S.M. Nandwa and B.H. Janssen. 1997. Soil fertility in Africa is at stake. (In: Buresh, R.J., P.A. Sanchez, and F. Calhoun. (Eds)). Replenishing Soil Fertility in Africa. SSSA Special Publication No. 51. Soil Science Society of America, Madison, WI. pp 47-61.
Smucker, A.J.M. and R.M. Aiken. 1992. Dynamic root responses to water deficits. Soil Science. 154:281-289.
Squire G.R. 1990. The physiology of Tropical Crop Production. C.A.B. International. Wallingford, UK.
Vissoh, P., V.M. Manyong., J.R. Carsky, P. Osei-Bonsu and M. Galiba. 1998. Experiences with Mucuna in West Africa. http://network.idrc.ca/en/ev-31912-201-1-DO_TOPIC.html
Willey, R.W. Intercropping – Its importance and Research needs. Part 1. Competition and yield advantages. Field Crop Abstracts. 32:1-10.
Wilson, J.B. 1988. A review of evidence on the control of shoot:root ratio, in relation to models. Annals of Botany 61, 433–449.
Wortmann, C.S., M. Isabirye., and S. Musa. 1994. Crotalaria ochrelueca as a green manure in Uganda. African Crop Science Journal. 2:55-61.










![A Review On Maize- Legume Intercropping For … yield in okra/cowpea intercropping system [11]. Other advantages of intercropping include: insurance against crop failure thereby minimizing](https://img.pdfslide.us/doc/110x75/5ac18fda7f8b9a5a4e8d4a51/a-review-on-maize-legume-intercropping-for-yield-in-okracowpea-intercropping.jpg)


















