Embed Size (px)
Citation preview

The Beagle, Records of the Northern Territory Museum of Arts and Sciences, 19929(1):35-48
WARKAIANIA, ANEW MEIOLANIID TURTLE FROM THE TERTIARYRIVERSLEIGH DEPOSITS OF QUEENSLAND, AUSTRALIA.
EUGENE s. GAFFNEYI, MICHAEL ARCHER2 AND ARTHUR WHlTE2
IDepartment of Vertebrate Paleontology, American Museum of Natural History,Central Park West at 79th St, New York, NY, USA 10024-5192.
2University of New South Wales,P.O. Box 1, Kensington, NSW 2033, Australia.
ABSTRACT
Warkalania carinaminor sp. nov., is based on posterior skull elements from the (?)Oligocene -early Miocene Carl Creek Limestone, Pancake Site of Riversleigh Station,northwest Queensland. Warkalania is a meiolaniid cryptodire because it has thesynapomorphies of the squamosal produced into posteriorly and posterolaterallydirected processes and an extensive squamosal-quadratojugal contact beneath thecavum tympani. Within meiolaniids, Warkalania is unique in having the squamosalprocesses formed into low, horizontal ridges, small and roughly similar to each otherin size, rather than having one or more of these scale areas extending prominently awayfrom the skull, as in all other meiolaniids.
KEYWORDS: Warkalania carinaminor gen et sp. nov., new genus, new species,Meiolaniidae, horned turtle, Oligocene, Miocene, Carl Creek Limestone, Queensland.
The reader should refer to Figures 5- 7 andGaffney (1983) for scale terminology ofmeiolaniid skulls. Descriptions of the skull ofMeiolania platyceps Owen can be found inGaffney (1983). Institutional prefixes to cata-logue numbers are as follows: AMF, AustralianMuseum, Sydney; BMNH, British Museum ofNatural History, London; QMF, QueenslandMuseum, Brisbane.
INTRODUCnON
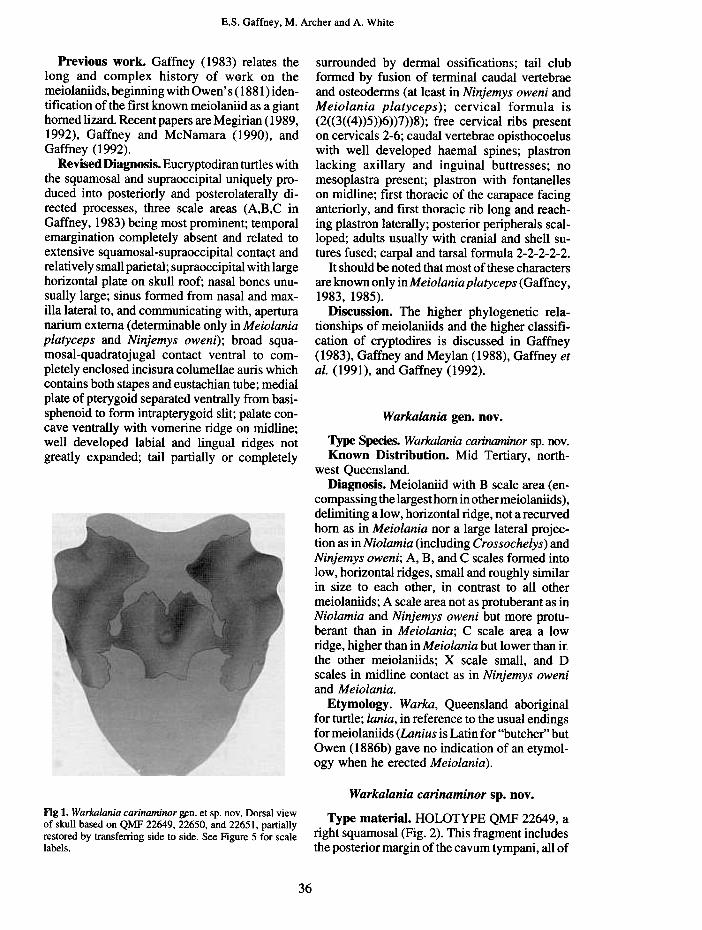
Turtles and other vertebrates were discoveredin the Carl Creek Limestone at Riversleigh Sta-tion in northwestern Queensland by Tedford in1968. Further work by teams led by M. Archer,S. Hand, and H. Godthelp resulted in the discov-ery of nearly 100 additional sites spanning intime the Oligocene to Pleistocene, and the recog-nition of a relatively rich turtle fauna (Gaffney etal. 1989). A review of the Riversleigh localitiesand an introduction to the growing literature onRiversleigh and its fossils can be found in Archeret al. ( 1989). The first turtles described fromRiversleigh Tertiary sites were three genera ofchelids based on fragmentary material (Gaffneyet al. 1989). Meiolaniids in the RiversleighTertiary were first recognized by A. Ritchie(pers. comm. ) on the basis of tail ring fragments(see below) in 1987. One of us(ESG)found moremeiolaniid fragments in Riversleigh collectionsin 1989 but it was not until Neville Pledge'sdiscovery of a partial skull, that diagnosablemeiolaniid material was recovered fromRiversleigh Tertiary deposits. This material rep-resents anew genus of meiolaniid (Fig. 1) and isdescribed here.
SYSTEMA TICS
Order TestudinesMegaorder Cryptodira
Capaxorder SelmacryptodiraHyperorder Daiocryptodira
Parvorder EucryptodiraSuborder Meiolanoidea
Family Meiolaniidae
Type Genus. Meiolania Owen, 1886b.Known Distribution. Pre-Oligocene ("Cre-
taceous or Eocene", Simpson 1938) of Argen-tina, Miocene to Pleistocene of Australia,Pleistocene (or younger) of Lord Howe Island,Walpole Island, and New Caledonia.
35
E.S. Gaffney, M. Archer and A. White
Previous work. Gaffney (1983) relates thelong and complex history of work on themeiolaniids, beginning with Owen's ( 1881) iden-tification of the first known meiolaniid as a gianthomed lizard. Recent papers are Megirian ( 1989,1992), Gaffney and McNamara (1990), andGaffney (1992).
Revised Diagnosis. Eucryptodiran turtles withthe squamosal and supraoccipital uniquely pro-duced into posteriorly and posterolaterally di-rected processes, three scale areas (A,B,C inGaffney, 1983) being most prominent; temporalemargination completely absent and related toextensive squamosal-supraoccipital contact andrelatively small parietal; supraoccipital with largehorizontal plate on skull roof; nasal bones unu-sually large; sinus formed from nasal and max-illa lateral to, and communicating with, aperturanarium extema (determinable only in Meiolaniaplatyceps and Ninjemys oweni); broad squa-mosal-quadratojugal contact ventral to com-pletely enclosed incisura columellae auris whichcontains both stapes and eustachian tube; medialplate of pterygoid separated ventrally from basi-sphenoid to form intrapterygoid slit; palate con-cave ventrally with vomerine ridge on midline;well developed Iabial and Iingual ridges notgreatly expanded; tail partially or completely
surrounded by dennal ossifications; tail clubfonned by fusion of tenninal caudal vertebraeand osteodenns (at least in Ninjemys oweni andMeiolania platyceps); cervical formula is(2«3«4»5»6»7»8); free cervical ribs presenton cervicals 2-6; caudal vertebrae opisthocoeluswith well developed haemal spines; plastronlacking axillary and inguinal buttresses; nomesoplastra present; plastron with fontanelleson midline; first thoracic of the carapace facinganteriorly, and first thoracic rib long and reach-ing plastron laterally; posterior peripherals scal-loped; adults usually with cranial and shell su-tures fused; carpal and tarsal fonnula 2-2-2-2-2.
It should be noted that most of these charactersare known only in M eiolania platyceps ( Gaffney ,1983, 1985).
Discussion. The higher phylogenetic rela-tionships of meiolaniids and the higher classifi-cation of cryptodires is discussed in Gaffney(1983), Gaffney and Mey1an (1988), Gaffney etal. (1991), and Gaffney (1992).
Warkalania gen. nov.
Type Speci~ Warkalania carinaminor sp. nov.Known Distribution. Mid Tertiary , north-
west Queensland.Diagnosis. Meiolaniid with B scale area ( en-
compassing the largest horn in other meiolaniids ),delimiting a low, horizontal ridge, not a recurvedhorn as in Meiolania nor a large lateral projec-tion as in Niolamia (including Crossochelys) andNinjemys oweni; A, B, and C scales formed intolow, horizontal ridges, small and roughly similarin size to each other, in contrast to all othermeiolaniids; A scale area not as protuberant as inNiolamia and Ninjemys oweni but more protu-berant than in Meiolania; C scale area a lowridge, higher than in Meiolania but lower than ir.the other meiolaniids; X scale small, and Dscales in midline contact as in Ninjemys oweniand Meiolania.
Etymology. Warka, Queensland aboriginalfor turtle; lania, in reference to the usual endingsfor meiolaniids (Lanius is Latin for "butcher" butOwen (1886b) gave no indication of an etymol-ogy when he erected Meiolania).
Warkalania carinaminor sp. nov.
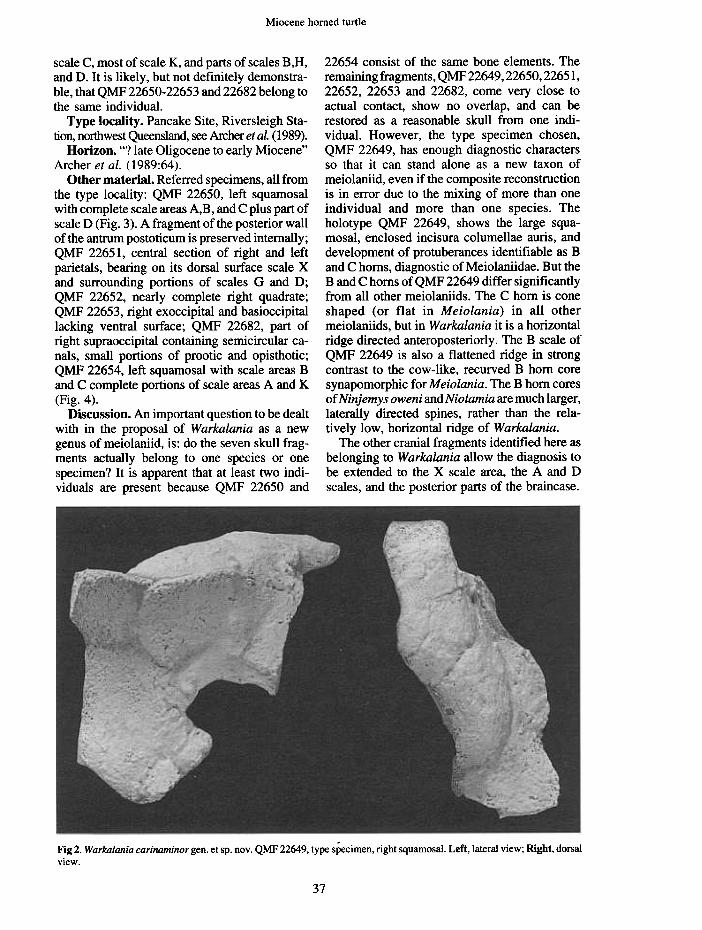
Type material. HOLOTYPE QMF 22649, aright squamosal (Fig. 2). This fragment includesthe posterior margin of the cavum tympani, all of
Fig I. Warkalania carinaminor ~n. et sp. nov. Dorsal viewof skull based on QMF 22649, 22650, and 22651, partiallyrestored by transferring side to side. See Figure 5 for scalelabels.
36
Miocene homed turtle
22654 consist of the same bone elements. Theremaining fragments, QMF22649, 22650,22651,22652, 22653 and 22682, come very close toactual contact, show no overlap, and can berestored as a reasonable skull from one indi-vidual. However, the type specimen chosen,QMF 22649, has enough diagnostic charactersso that it can stand alone as a new taxon ofmeiolaniid, even if the composite reconstructionis in error due to the mixing of more than oneindividual and more than one species. Theholotype QMF 22649, shows the large squa-mosal, enclosed incisura columellae auris, anddevelopment of protuberances identifiable as Band C horns, diagnostic of Meiolaniidae. But theBand C horns of QMF 22649 differ significantlyfrom all other meiolaniids. The C horn is coneshaped (or flat in Meiolania} in all othermeiolaniids, but in Warkalania it is a horizontalridge directed anteroposteriorly. The B scale ofQMF 22649 is also a flattened ridge in strongcontrast to the cow-like, recurved B horn coresynapomorphic for Meiolania. The B horn coresofNinjemys oweni and Niolamia are much larger,laterally directed spines, rather than the rela-tively low, horizontal ridge of Warkalania.
The other cranial fragments identified here asbelonging to Warkalania allow the diagnosis tobe extended to the X scale area, the A and Dscales, and the posterior parts of the braincase.
scale c, most of scale K, and parts of scales B,H,and D. It is likely, but not definitely demonstra-ble, that QMF 22650-22653 and 22682 belong tothe sanle individual.
Type locality. Pancake Site, Riversleigh sta-tion, northwest Queensland, see Archer et aL ( 1989).
Horizon. .'? late Oligocene to early Miocene"Archer et at. (1989:64).
Other material. Referred specimens, all fromthe type locality: QMF 22650, left sqUanlosalwith complete scale areas A,B, and C plus part ofscale D (Fig. 3). A fragment of the posterior wallof the antrum postoticum is preserved internally;QMF 22651, central section of right and leftparietals, bearing on its dorsal surface scale Xand surrounding portions of scales G and D;QMF 22652, nearly complete right quadrate;QMF 22653, right exoccipital and basioccipitallacking ventral surface; QMF 22682, part ofright supraoccipital containing semicircular ca-nals, small portions of prootic and opisthotic;QMF 22654, left sqUanlosal with scale areas Band C complete portions of scale areas A and K(Fig. 4).
Discussion. An important question to be dealtwith in the proposal of Warkalania as anewgenus of meiolaniid, is: do the seven skull frag-ments actually belong to one species or onespecimen? It is apparent that at least two indi-viduals are present because QMF 22650 and
Fig 2. Warkalania carinaminor gen. et sp. nov. QMF 22649, type specimen, right squamosal. Left, lateral view; Right, dorsal
view.
37

E.S. Gaffney, M. Archer and A. White
The areas preserved in Warkalania only in-clude the posterior parts of the skull. The overallproportions and size of the Warkalania speci-mens are roughly the same as Meiolania platyceps(Figs 5,6). In most Meiolania platyceps skullsthe scute areas are delimited by raised ridges,while in Ninjemys oweni and Niolamia the scales
Diagnosis. Same as for genus.Description. Because there are no bone su-
tures preserved in the Warkalania material, thedescription uses the scute areas as homologousentities that are hypothesized as suitable forcomparisons. The scute terms and the order isfrom Gaffney (1983).
Fig 3. Warkalania carinaminor gen. et sp. nov. QMF 22650, left squamosal. Left, lateral view; Right. dorsal view.
Fig. 4. Warkalania carinaminor gen. et sp. nov. QMF 22654, left squamosal. Left, lateral view; Right, dorsal view.
38

Miocene horned turtle
are bordered by shallow grooves. Warkalaniahas grooves rather than ridges and thus differsfrom Meiolania platyceps, and resemblesNinjemys oweni and Niolamia in this feature.
The midline area of the skull in Warkalania ispreserved in QMF 22651, a fragment that con-sists of scale X and surrounding portions ofscales G and D. It probably represents the ante-rior part of both parietals and the posterior partof both frontals. On its dorsal surface there is aclear, oblong scale area in the midline, scale X,with a small, cone-shaped projection just left ofthe midline. Although not as well defined as inmost Meiolania platyceps specimens, scale X inWarkalania is similar in size and shape to theLord Howe species. The small cone is alsopresent in most Meiolania, but smaller. Thedorsal surface is somewhat rugose in QMF 22651,and there are vague grooves that could be thesulci separating scales G and D from each other.However, if they are present, they are poorlydifferentiated from other surface irregularities.
In Ninjemys oweni, the X scale area is not wellpreserved, but it is smaller than in Niolamia andmay be much as in Meiolania platyceps. The Dscales of Warkalania and Meiolania platycepsmeet in the midline. In Niolamia they are sepa-rated by a large X scale. In Ninjemys oweni theX scale margins are not clear but the X scale areais definitely not as large as in Niolamia, and theD scales probably do meet in the midline inNinjemys oweni.
The ventral surface of QMF 22651 preservesthe dorsalmost portion of the cavum cranii. Mostof this region is formed by the parietal but a smallpart of the anterior extension of the supraoccipitalis preserved on the midline. The area of thecavum cranii preserved includes the space usu-ally filled in life by the cartilaginous remnant ofthe synotic tectum (Gaffney 1979). This area ispreserved in a number of specimens of M eiolaniaplatyceps and in Ninjemys oweni but not inNiolamia. Although this region is similar in allturtles, there are differences between Ninjemysoweni and Meiolania platyceps. Unfortunately,not enough is preserved in QMF 22651 to deter-mine to which one it is most similar. The proc-essus inferior parietalis is broken on both sides,but anterolaterally on the outside of each proces-sus, is a concavity also seen in Meiolaniapla'tyceps but not in Ninjemys oweni. This con-cavity is apparently involved in the attachmentarea of the M. adductor mandibulaepseudotemporalis. The broken edge of the cristasupraoccipitalis is preserved in QMF 22651. To
the extent that it is preserved, it agrees withMeiolania platyceps.
The fragment of cranial roof, QMF 22651,taken alone, could not be distinguished fromMeiolania platyceps, except in the absence ofraised scale edges. But even this feature varies inMeiolania platyceps; AMF 57984 also lacksraised scale edges.
Scute D is a large, paired scale covering muchof the posterior portion of the skull, and consistsmostly of parietal and squamosal. Parts of ScuteD are preserved in QMF 22649,22650, 22651and 22654. Even when the fragments are com-bined, the D scale area is not completely pre-served in Warkalania. The D scale area is bestpreserved on the left posterolateral portion ofQMF 22651 and the left anteromedial portion ofQMF 22650. The two areas do not have a goodcontact but, when restored using QMF 22652 and22653 (braincase elements) for control of theskull width, only a small amount of bone appearsto be missing between the two cranial roofsections. Posteriorly and laterally, the D scalelies against the A, B, and C scales, as in othermeiolaniids. An anterior sulcus with scale Hisnot discernible in QMF 22649, which has enoughbone preserved to expect the presence of thesulcus. It is likely, however, that poor preserva-tion is the cause of the absence of the sulcus,rather than the actual absence of this sulcus. Theposteromedial limits of the D scale are notpreserved on any of the fragments. The region ofthe D scale is a distinctive convexity in bothNinjemys oweni andNiolamia. Warkalania, how-ever, agrees with Meiolania in having the D scalearea relatively low.
The three scale areas, A, B, and C, are the mostuseful skull regions within meiolaniids for taxo-nomic comparisons. Fortunately, nearly all ofthese scale areas are preserved in Warkalania.Scute Cis a paired scale area on the posterola-teral part of the skull, and is formed mostly by thesquamosal bone. In QMF 22649, 22650, and22654 are preserved significant parts of the Cscale area. In these specimens the C scale formsa laterally projecting ridge that is roughly hori-zontal but with a slight anterodorsal trend. Thisridge projects laterally to a lesser extent thaneither of the other scale areas, A and B. In QMF22649 and 22650, the C scale has a more acuteedge than in QMF 22654 but the entire scale areaprojects laterally to a greater extent in QMF22654. In Meiolania platyceps, Ninjemys oweni,and Niolamia the C scale is more rounded andcone-shaped than in Warkalania. Meiolania
39
E.S. Gaffney. M. Archer and A. White
platyceps varies from nearly flat in AMF 43183to a well developed cone in AMF 57984, but it isnever a horizontal ridge as in Warkalania.
The B scale area is preserved completely inQMF 22650 and QMF 22654, and in QMF 22649the anterior half is preserved. The B scale islarger and projects more than the A scale, and itis flattened in the same plane as the A scale. InQMF 22654 the B scale is thicker and has ablunter lateral edge than in QMF 22649 and22650. The most striking differences in skullmorphology of meiolaniids can be found in theshapes of scale B. InNinjemys oweni and Niolamiathe B scale areas are very large, laterally project-ing processes that extend laterally beyond therest of the skull. In Meiolania platyceps the Bscale also forms a projection but in this case it isthe cow-like horn core distinctive of this genus.Warkalania, with its relatively small B scalearea, forms a projection that is much less exten-sive than in Ninjemys oweni or Niolamia. The Bhorn core of some individuals of Meiolania
platyceps (e.g. AMF 43183, and AMF 18368)approaches the size of the B scale in Warkalania,but in none of the Meiolania platyceps speci-mens is the B scale as small as in Warkalania.Meiolania platyceps also has a B horn core thatis conical, usually recurved, and not flattened inany plane. In both Niolamia and Ninjemys owenithe B horn is flattened in the horizontal plane asin Warkalania but not to the degree seen inWarkalania.
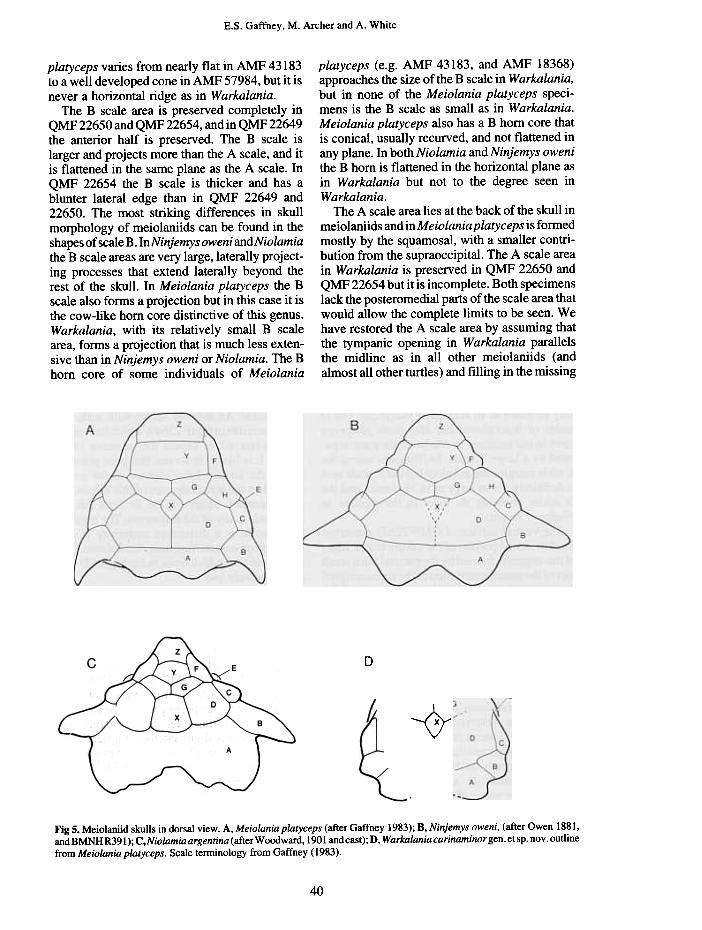
The A scale area lies at the back of the skull inmeiolaniids and in Meiolania platyceps is formedmostly by the squamosal, with a smaller contri-bution from the supraoccipital. The A scale areain Warkalania is preserved in QMF 22650 andQMF 22654 but it is incomplete. Both specimenslack the posteromedial parts of the scale area thatwould allow the complete limits to be seen. Wehave restored the A scale area by assuming thatthe tympanic opening in Warkalania parallelsthe midline as in all other meiolaniids (andalmost all other turtles) and filling in the missing
D
~
Fig 5. Meiolaniid skulls in dorsal view. A, Meiolaniaplatyceps (after Gaffney 1983); B, Ninjemys oweni, (after Owen 1881,and BMNHR391); C,Niolamia argentina (after Woodward, 1901 and cast); D, Warkalania carinaminorgen. etsp. nov. outline
from Meiolania platyceps. Scale ternlinology from Gaffney (1983).
40
Miocene homed turtle
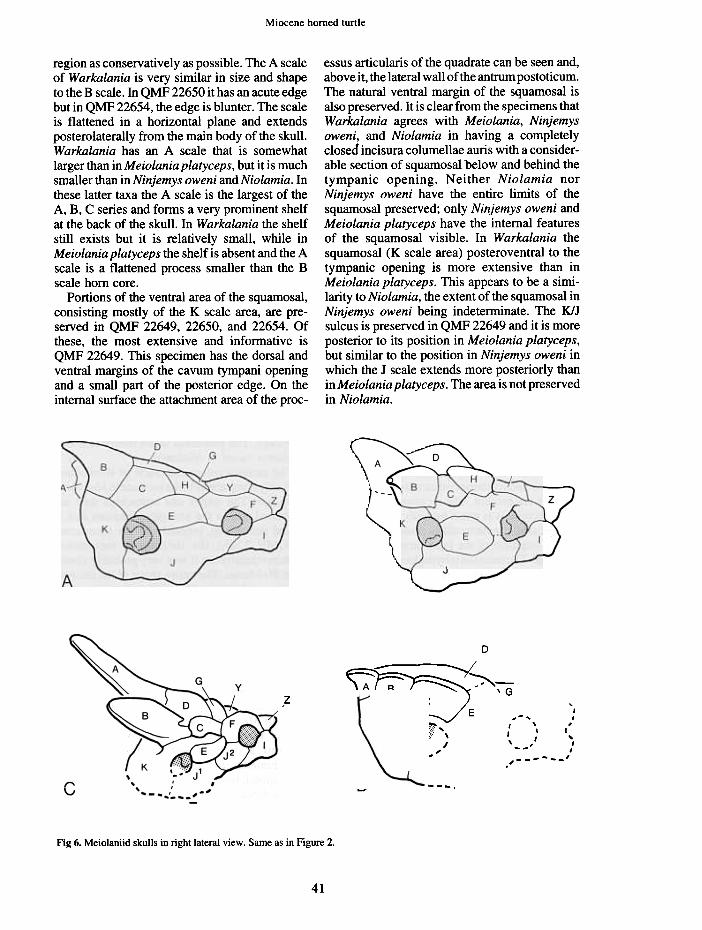
essus articularis of the quadrate can be seen and,above it, the lateral wall of the antrum postoticum.The natural ventral margin of the squamosal isalso preserved. It is clear from the specimens thatWarkalania agrees with Meiolania, Ninjemysoweni, and Niolamia in having a completelyclosed incisura columellae auris with a consider-able section of squamosal below and behind thetympanic opening. Neither Niolamia norNinjemys oweni have the entire limits of thesquamosal preserved; only Ninjemys oweni andMeiolania platyceps have the internal featuresof the squamosal visible. In Warkalania thesquamosal (K scale area) posteroventral to thetympanic opening is more extensive than inMeiolania platyceps. This appears to be a simi-larity to Niolamia, the extent of the squamosal inNinjemys oweni being indeterminate. The K/Jsulcus is preserved in QMF 22649 and it is moreposterior to its position in Meiolania platyceps,but similar to the position in Ninjemys oweni inwhich the J scale extends more posteriorly thanin Meiolania platyceps .The area is not preservedin Niolamia.
region as conservatively as possible. The A scaleof Warkalania is very similar in size and shapeto the B scale. In QMF 22650 it has an acute edgebut in QMF 22654, the edge is blunter. The scaleis flattened in a horizontal plane and extendsposterolaterally from the main body of the skull.Warkalania has an A scale that is somewhatlarger than in Meiolania platyceps, but it is muchsmaller than in Ninjemys oweni and Niolamia. Inthese latter taxa the A scale is the largest of theA, B, C series and forms a very prominent shelfat the back of the skull. In Warkalania the shelfstill exists but it is relatively small, while inMeiolania platyceps the shelf is absent and the Ascale is a flattened process smaller than the Bscale horn core.
Portions of the ventral area of the squamosal,consisting mostly of the K scale area, are pre-served in QMF 22649, 22650, and 22654. Ofthese, the most extensive and informative isQMF 22649. This specimen has the dorsal andventral margins of the cavum tympani openingand a small part of the posterior edge. On theinternal surface the attachment area of the proc-
D yA
z
B
D
~ ~~A- c 'G
E,-..
\I
z ,I,
K I' ,, ' I, I'
-~, II
."- ,
D ---,
Fig 6. Meiolaniid skulls in right lateral view. Same as in Figure 2.
41

E.S. Gaffney, M. Archer and A. White
The three braincase fragments, QMF 22652,22653, and 22682, all belong to the right side andprobably are from the same individual. All threeof these pieces very nearly come into contactwhen restored on the basis of Meiolania platycepsand are separated by relatively narrow areas ofbone. Nonetheless, they do not have any con-tacts.
QMF 22682 is the part of the rightsupraoccipital that contains the semicircularcanals, plus some small portions of the prooticand opisthotic. The fragment consists of the areaof bone just anterior to the foramen magnum (themargin of that structure is not preserved) andposterior to the foramen nervi trigemini. Theposterior margin of the foramen nervi trigeminiis preserved. On the lateral surface of this pieceof supraoccipital is the shallow groove thatwould lead to the foramen stapedio-temporale.Everything described so far is quite similar toMeiolania platyceps and does not vary a greatdeal from other generalized cryptodires.
The internal portion of QMF 22682 containsthe dorsal impression of the canalis semicircularishorizontalis (see Gaffney 1979: Figs 49,52, forterminology of the cavum labyrinthicum in tur-tles and Gaffney 1983: Figs 49-51, for the innerear in Meiolania platyceps) with the ventralregion missing. The recessus labyrinthicusprooticus and its connected canalis semicircularisanterior are present. The recessus labyrinthicussupraoccipitalis is intact but only the dorsal partof the canalis semicircularis posterior is presentwith a fragment of the bony strap defining thisstructure. The recessus labyrinthicus opisthoticusis missing also. The cavum labyrinthicum aspreserved in Warkalania agrees with Meiolaniaplatyceps except that the canals are smaller thanin AMF 57984, presumably a function of thesmaller skull in Warkalania.
QMF 22682 has two grooves preserved in theregion of the dorsal edge of the hiatus acusticus.This opening is usually nearly closed in M eiolaniaplatyceps and the edges are broken in QMF22682. The grooves do not appear to be part ofthe fossa acustico-facialis but rather may beinterpreted as the foramen aquaducti vestibuliand one of the foramen nervi acustici. Bothgrooves lead directly from the cavum cranii intothe cavum labyrinthicum and their contents can-not be definitely determined. QMF 22652 is aright qlfadrate separated from the more medialpart of the otic chamber at roughly the positionof the quadrate-opisthotic/pterygoid suture, ex-
42

Miocene horned turtle
Etymology. Carina, ridge, and minor, small,in reference to this species having scale areasA,B, and C as small ridges, a condition uniqueamong meiolaniids.
Relationships of Warkalania. Warkalania isclearly a meiolaniid. It possesses two of the threesynapomorphies proposed for the family byGaffney ( 1983) and Gaffney and Meylan ( 1988):1. Squamosal and supraoccipital produced into
large posteriorly and posterolaterally directedprocesses that extend clear of skull.
2. Broad squamosal-quadratojugal contact ven-tral to quadrate.The third character, the intrapterygoid slit, is
indeterminate in Warkalania.Relationships within the Meiolaniidae have
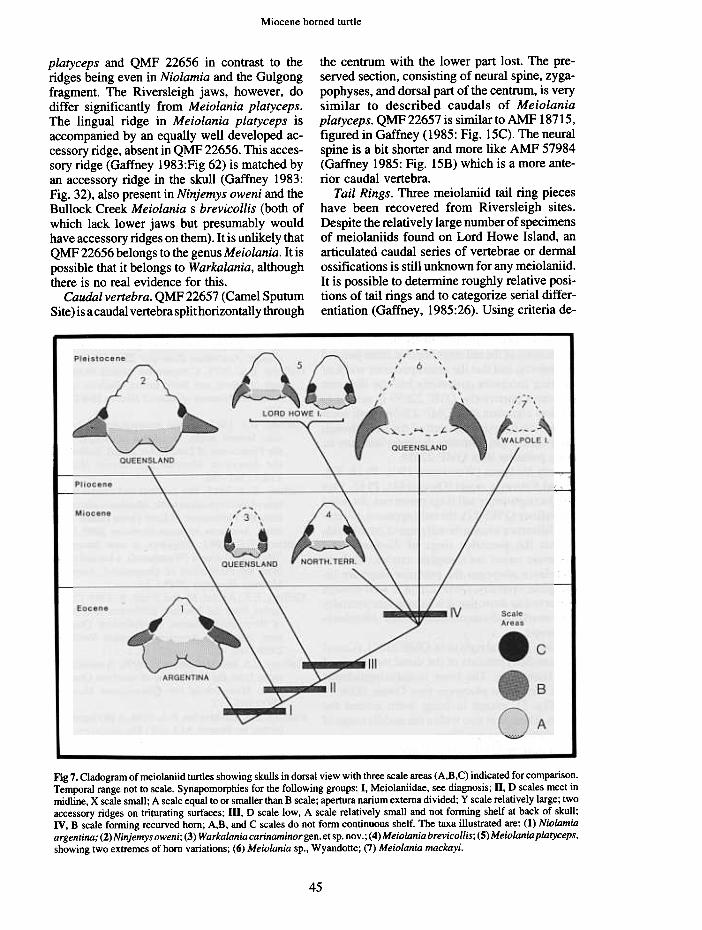
only been briefly referred to by Gaffney andMeylan (1988:181 and Figs 5,8). They groupedNinjemys oweni (now named Ninjemys owni,Gaffney 1992) and Meiolania platyceps as thesister group to Niolamia on the basis of A scutearea relatively small in comparison to the A scutearea in Niolamia. However, the squamosal-supraoccipital scale characters cannot be readilypolarized by reference to a cryptodiran outgroupbecause the very presence of these elements is ameiolaniid synapomorphy unique to this group.The entire polarizing of the scales in meiolaniidsrests on the argument that Niolamia is the sistergroup of all other meiolaniids. This argument isbased on a character that can be polarized byoutgroup comparison, the structure of theintrapterygoid slit. Gaffney (1983:431-435, Fig.60) argued that "Crossochelys" (Simpson 1938,congeneric with Niolamia) has an intrapterygoidslit that is primitive with respect to theintrapterygoid slit of Meiolania platyceps. Thisis only one character, but, at present, it is the onlycharacter with any degree of complexity that canbe used to resolve the polarity problem within themeiolaniids. It is then possible to hypothesizeNiolamia as the sister taxon to other meiolaniidsand to use this hypothesis to resolve the varioussquamosal scale characters. In addition to theinherent weaknesses of this character, it shouldbe noted that the intrapterygoid slit character isdeterminable only in Niolamia ( ="Crossochelys")and Meiolania platyceps, the region is absent inNinjemys oweni, Warkalania, and other pre-sumed species of Meiolania. Nonetheless, itseems reasonable to adopt the idea of Niolamiaas the sister taxon to all other meiolaniids (Fig.7) as a working hypothesis and accept the limi-tations.
ally to a greater degree in Meiolania platycepsthan in Warkalania. Unfortunately;- this area isnot well enough preserved in other meiolaniidtaxa for rigorous comparisons.
QMF 22653 consists of most of the basioccipi-tal plus the right exoccipital. As in all the otherWarkalania fragments, sutures are not distin-guishable. The condylus basioccipitalis ofWarkalania is similar in shape and proportions toMeiolania platyceps, the neck is relatively shortand stout in both taxa, in contrast to most otherturtles. The articular surface of the "ondylus isslightly concave in Meiolania platyceps andslightly convex in Warkalania. The degree towhich the condylus occipitalis is made up ofbasioccipital versus exoccipitals is not deterrni-nable in Warkalania. The ventral surface of thebasioccipital is broken off.
The foramen magnum of Warkalania slopesanterodorsally as in Meiolania platyceps, a rela-tively unusual chelonian condition but indeter-minate in the other meiolaniids. There are twopairs of foramina nervi hypoglossi, one pairpenetrating each exoccipital. In some Meiolaniaplatyceps a very small, third foramen nervihypoglossi is present (Gaffney 1983) but mostspecimens have two prominent foramina on eachside as in Warkalania. The area of the exoccipitalbetween the foramen magnum and the medialedge of the foramen jugulare posterius containsthe two hypoglossal foramina, and in Warkalaniathis forms a relatively flat surface roughly paral-lel to a transverse plane. Both posterior foraminanervi hypoglossi open at an acute angle to thissurface. In Meiolania platyceps this area be-tween foramen magnum and foramen jugulareposterius is more curved, so that the more poste-rior of the foramina nervi hypoglossi opens atnearly right angles to the bone surface. In theabsence of the rest of the cavum acustico-jugulareand fenestra postotica in Warkalania, and theabsence of comparable regions in the othermeiolaniids, the significance of this differencecannot as yet be determined.
In QMF 22653 the anterior and posterioropenings of the foramina nervi hypoglossi can beseen on the right side, and on the left side someof the internal parts of the canals for these nervescan be seen in the broken surface. The anteriorand lateral margins of QMF 22653 are mostlybroken edges but the posterior margin of theforamen jugulare anterius and the posterior wallof the recessus scalae tympani (fide Gaffney,1979) are identifiable.
43

E.S. Gaffney, M. Archer and A. White
at least two meiolaniid taxa makes a less likelyalternative possible, namely that this is a B plusC scale area with the gutter between the tworepresented by the broken area. However, on thedorsal surface there is no sign of a trough ordepression as in Warkalania, and, most impor-tantly, on the internal surface there is one con-tinuous sheet of finished bone with no indica-tions of a depression for each scale area as inWarkalania. QMF 22655 does differ fromMeiolania platyceps in having more acute poste-rior edges, which are rounded in Meiolaniaplatyceps. The A horn of the Bullock CreekMeiolania s brevicollis (Megirian 1989, 1992) islower than in either Meiolania platyceps or QMF22655 and does not have acute edges either. Atpresent, however, the best interpretation of thisspecimen is that of Meiolania in the strict sense,species indet
Lower jaws. A lower jaw of a meiolaniid fromRiversleigh is QMF 22656 (Camel Sputum Site).The specimen consists of most of the fuseddentaries, the right side being more brokenposteriorly. The jaws are very deep, much deeperthan any chelid, but relatively narrow. In addi-tion to Meiolania platyceps (Gaffney, 1983),partial lower jaws are known for Niolamia (Wood-ward 1901) and for an unnamed meiolaniidknown from very fragmentary material fromGulgong, NSW (Mining Museum of Sydney13898). A11 the jaws are relatively deep, havefused mandibular symphyses, and parallel labialand lingual ridges separated by a trough. InNiolamia and the Gulgong fragment, the ridgesare low and the trough shallow (probably exag-gerated by breakage) and there is no symphysealor median cusp. In Meiolania platyceps andQMF 22656, the ridges are sharp and well de-fined and a median cusp is present (Gaffney1983:449). The lingual (inner) ridge is distinctlyhigher than the labial ( outer) ridge in M eiolania
The genus Meiolania can be restricted to justthose taxa with cylindrical, recurved, cow-like Bhorns, following Gaffney (1983) and Megirian(1989,1992). The species oweninarnedbyWood-ward (1901) and placed in Meiolania by himshould be removed to its own genus, Ninjemys ,characterized by small, cone shaped C scales,laterally projecting B horns, and A scales inter-mediate in size between Niolamia and Meiolania(see also Table 1). Given these taxa and Niolamiabeing a senior synonym of Crossochelys, thereare three generic level taxa of meiolaniids tocompare with Warkalania (Table 1).
The small size of the B scales is unique toWarkalania but the small size of the A scalesmay be interpreted as a synapomorphy in com-mon with Meiolania and not found in Ninjemysoweni and Niolamia. The absence of a laterallyprojecting B scale, found in Ninjemys oweni andNiolamia, and presumably a primitive meiolaniidfeature, could also be argued as a synapomorphyuniting Meiolania and Warkalania. However,the B scales of Meiolania and Warkalania arevery different from each other. A better characteris the low D scale of Warkalania and Meiolania,produced into a raised tubercle in Ninjemysoweni and Niolamia. This analysis of the fewcharacters available at present is reflected in thecladograrn in Figure 7.
Other meiolaniid specimens fromRiversleigh sites. Skull fragment. QMF 22655appears to be the left A scale area of a speciesvery similar to Meiolania platyceps. The speci-men is unfortunately very limited in useful fea-tures, and a break in the middle of the scale areamakes other interpretations possible. As pre-served, the fragment is a triangular piece ofbonewith the apex-divided by breakage. It looks as ifthe broken area was originally filled with boneforming a single, large projection. However, thepresence in the Miocene of northern Australia of
Table I. Comparison of the cranial morphology meiolaniid genera.
Warkalania Meiolania Ninjemysoweni
Niolamia
probablyintermediate
high
yes
yes
no
yes
yes
yes
yes
yes
no
low
yes
no
yes
no
yes
no
indet
no
yes
high
yes
yes
no
yes
yes
yes
no
yes
no
low
no
yes (when present)
no
no
no
no
yes
.D scales meet in midline " ! , i
X scale large
D scale
A, B, and C scales fonning a continuous shelf
C scale cone-shaped
C scale a horizontal ridge
B scale projecting laterally
B scale flattened
A scale very large and fonning shelf at back of skull
y scale relatively large
44
Miocene homed turtle
the centrum with the lower part lost. The pre-served section, consisting of neural spine, zyga-pophyses, and dorsal part of the centrum, is verysimilar to described caudals of Meiolaniaplatyceps. QMF22657 is similartoAMF 18715,figured in Gaffney (1985: Fig. 15C). The neuralspine is a bit shorter and more like AMF 57984(Gaffney 1985: Fig. 15B) which is a more ante-rior caudal vertebra.
Tail Rings. Three meiolaniid tail ring pieceshave been recovered from Riversleigh sites.Despite the relatively large number of specimensof meiolaniids found on Lord Howe Island, anarticulated caudal series of vertebrae or dermalossifications is still unknown for any meiolaniid.It is possible to determine roughly relative posi-tions of tail rings and to categorize serial differ-entiation (Gaffney, 1985:26). Using criteria de-
platyceps and QMF 22656 in contrast to theridges being even in Niolamia and the Gulgongfragment. The Riversleigh jaws, however, dodiffer significantly from Meiolania platyceps.The lingual ridge in Meiolania platyceps isaccompanied by an equally well developed ac-cessory ridge, absent in QMF 22656. This acces-sory ridge (Gaffney 1983:Fig 62) is matched byan accessory ridge in the skull (Gaffney 1983:Fig. 32), also present in Ninjemys oweni and theBullock Creek Meiolania s brevicollis (both ofwhich lack lower jaws but presumably wouldhave accessory ridges on them). It is unlikely thatQMF 22656 belongs to the genus Meiolania. It ispossible that it belongs to Warkalania, althoughthere is no real evidence for this.
Caudal vertebra. QMF 22657 (Camel SputumSite) is a caudal vertebra split horizontally through
~,
Fig 7. Cladogram of meiolaniid turtles showing skulls in dorsal view with three scale areas (A,B,C) indicated for comparison.Temporal range not to scale. Synapomorphies for the following groups: I, Meiolaniidae, see diagnosis; II, D scales meet inmidline, X scale small; A scale equal to or smaller than B scale; apertura narium externa divided; y scale relatively large; twoaccessory ridges on triturating surfaces; m, D scale low, A scale relatively small and not forming shelf at back of skull;IV, B scale forming recurved horn; A,B, and C scales do not form continuous shelf. The taxa illustrated are: (I) Niolamiaargentina; (2)Ninjemys oweni; (3) Warkalania carinaminorgen. et sp. nov .; (4) Meiolania brevicollis; (5) Meshowing two extremes of horn variations; (6) Meiolania sp., Wyandotte; (7) Meiolania mackayi.
45

E.S. Gaffney. M. Archer and A. White
ACKNOWLEDGEMENTS
We would like to thank Neville Pledge of theSouth Australia Museum for collecting the typespecimen of Warkalania and bringing it to ourattention. One ofus (ESG) is very grateful to thefollowing individuals and institutions for mak-ing their specimens of meiolaniids available: R.Jones, A. Ritchie, Australian Museum, Sydney;J. Picket, Mining and Geological Museum, Syd-ney; A. Milner, C. Walker, S. Chapman, BritishMuseum (Natural History ), London; D. Megirian,Northern Territory Museum, Darwin.
REFERENCES
veloped from Meiolania platyceps, the mostanterior tail ring is QMF 22660 (Camel SputumSite), with more posterior rings being QMF22658 (Ringtail Site), and QMF 22659 (StickyBeak Site). The anterior ring, QMF 22660, hastwo projections, probably the two on the rightside, and is similar to the fragment figured byOwen (1888: PI. 36, Figs 7-9). The projections inQMF 22660 are more acute than in the figuredspecimen, but this is probably due to the Riversleighspecimen being from a more posterior position.
A more posterior tail ring is QMF 22658 (thespecimen originally identified by A. Ritchie),which is the only Riversleigh tail ring to beessentially complete. It is similar to AMF 50635,figured in Gaffney (1985: Fig. 21), but theprojections are lower in QMF 22658. Again,however, this is probably due to QMF 22658being more anterior than AMF 50635. InMeiolania platyceps, it is likely that the pairedprojections of the tail rings become more pointedposteriorly, and that the anteroposterior width ofthe ring increases posteriorly but the diameterdecreases posteriorly. QMF 22659 is similar tobut less complete than QMF 22658. Both seemto be from the posterior part of the tail althoughQMF 22659 has a smaller diameter and may bemore posterior than QMF 22658.
Both Niolamia (Woodward 1901: P118, Fig.2) and Ninjemys oweni (Owen 1881: P165, Figs1-4) have posterior tail rings preserved. As notedby Gaffney (1985:27), the only apparent system-atic difference among the tail rings of meiolaniidsis that the posterior rings of Niolamia andNinjemys oweni are complete circles, while inMeiolania platyceps the posterior rings are in-complete ventrally. QMF 22658 is well enoughpreserved to show that it is incomplete ventrallyand nearly indistinguishable from Meiolaniaplatyceps.
Tibia. A partial right tibia, QMF 22661, (CamelSputum Site) consists of the distal two thirds ofthis limb bone. The bone is indistinguishablefrom Meiolania platyceps (see Owen 1888: PI36, Fig. 2), except in being worn around theedges. The size is also within the middle range ofMeiolania platyceps tibias.
Osteoderm. QMF 22662 (Camel Sputum Site)is an asymmetrical, tear-drop shaped osteoderm,typical of meiolaniids and nearly identical toones found on Lord Howe Island belonging toMeiolania platyceps. A figure of a Meiolaniaplatyceps osteoderm that is very similar to thisRiversleigh specimen can be found in Owen(1888: PI. 36, Fig. 10).
Archer, M., Godthelp, H., Hand, S. and Megirian, D.1989. Fossil mammals of Riversleigh, north-western Queensland: preliminary overview ofbiostratigraphy, correlation and environmentalchange. Australian Zoologist 25(2):29-65.
Gaffney, E.S. 1979. Comparative cranial morphol-ogy of recent and fossil turtles. Bulletin of theAmerican Museum of Natural History 164(2):65-376.
Gaffney, E.S. 1983. Cranial morphology of the ex-tinct horned turtle, Meiolania platyceps, fromthe Pleistocene of Lord Howe Island. Bulletin ofthe American Museum of Natural History
174(4):361-480.Gaffney, E. S. 1985. The cervical and caudal verte-
brae of the cryptodiran turtle, Meiolania platyceps,from the Pleistocene of Lord Howe Island, Aus-tralia. American Museum Novitates 2805:1-29.
Gaffney, E.S. 1992. Ninjemys, a new name for"Meiolania" oweni (Woodward), a horned turtlefrom the Pleistocene of Queensland. AnericanMuseum Novitates 3049:1-10.
Gaffney, E.S., Archer, M. and White, A. 1989. Chelidturtles from the Miocene freshwater limestonesof Riversleigh Station, Northwestern Queens-land, Australia. American Museum Novitates2959: 1-10.
Gaffney, E.S. and McNamara G. 1990. A meiolaniidturtle from the Pleistocene of northern Queens-land. Memoirs of the Queensland Museum28(1): 107-113.
Gaffney, E.S. and Meylan, P .A. 1988. A phylogeny ofturtles. In: Benton, M.j. (ed.) The phylogeny andclassification of tetrapods. Pp 157-219.Clarendon Press: Oxford.
Gaffney, E.S., Meylan, P.A. and Wyss, A. 1991. Acomputer assisted analysis of the relationshipsof the higher categories of turtles. Cladistics7:313-335.
Megirian, D. 1989. A description of horned-turtleremains (Testudines:Meiolaniidae) from the mid-Miocene Camfield Beds of northern Australia.
46

Miocene homed turtle
Owen, R. 1888. On parts of the skeleton of Meiolaniaplatyceps (Owen). Philosophical Transactionsof the Royal Society, London, serial B 179:181-191.
Simpson, G. G. 1938. Crossochelys, Eocene homedturtle from Patagonia. Bulletin of the AmericanMuseum of Natural History 74:221-254.
Woodward, A. S. 190 I. On some extinct reptiles fromPatagonia, of the genera Miolania, Dinilysia,and Genyodectes. Proceedings of the ZoologicalSociety, London: 169-184.
The Beagle, Records of the Northern TerritoryMuseum of Arts and Sciences 6(1): 105-113.
Megirian, D. 1992. Meiolania brevicollis sp.nov.(Testudines: Meiolaniidae): a new homed turtlefrom the Australian Miocene. Alcheringa 3:93-106.
Owen, R. 1881. Description of some remains of thegigantic land-Iizard (Megalania prisca, Owen)from Australia. Part II. Philosophical Transac-tions of the Royal Society, London ( 1880)171:1037-1050.
Owen, R. 1886. Description of fossil remains of twospecies of Megalanian genus (Meiolania) from"Lord Howe's Island." Philosophical Transac-tions of the Royal Society, London 177:471-480. Accepted 23 April, 1992
47





























