Embed Size (px)
Citation preview

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Wading Bird Foraging Habitat Selection in the FloridaEvergladesAuthor(s): Rachael L. Pierce and Dale E. GawlikSource: Waterbirds, 33(4):494-503. 2010.Published By: The Waterbird SocietyDOI: http://dx.doi.org/10.1675/063.033.0408URL: http://www.bioone.org/doi/full/10.1675/063.033.0408
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainableonline platform for over 170 journals and books published by nonprofit societies,associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should bedirected to the individual publisher as copyright holder.

494
Wading Bird Foraging Habitat Selection in the Florida Everglades
RACHAEL L. PIERCE1,* AND DALE E. GAWLIK2
1South Florida Water Management District, Restoration Sciences Department, 3301 Gun Club Road,West Palm Beach, Fl, 33406, USA
2Environmental Sciences Program, Florida Atlantic University, 777 Glades Road, Boca Raton, FL, 33431-0991, USA
*Corresponding author; E-mail: [email protected]
Abstract.—To determine how habitat structural complexity, which affects prey vulnerability, influences foraginghabitat selection by wading birds, a habitat use versus availability study was conducted throughout the Florida Ever-glades in 2005 and 2006. Also, an experiment was conducted where structural complexity was manipulated and itseffect on wading bird foraging efficiency quantified. Among-year differences in habitat selection were found, whichcorresponded to disparate hydrological conditions. In 2005, a poor hydrological year in terms of the seasonal re-cession, wading birds chose foraging sites that had less emergent vegetation, a thicker flocculent layer and higherprey density relative to random sites. In 2006, an optimal hydrological year, wading bird foraging locations weresimilar to random sites in all aspects. Submerged vegetation did not affect wading bird site selection in either year.The study indicated that hydrological conditions that affect prey density were more important to wading bird for-aging success than fine scale variation in habitat characteristics. However, in years of poor hydrology factors thataffect prey vulnerability may become increasingly important because the penalty for choosing low quality foraginghabitat is greater than in years of more optimal conditions. Elucidating habitat characteristics which create highquality foraging sites will be beneficial in planning wetland restoration projects and gauging future restorationprogress. Received 2 October 2009, accepted 18 May 2010.
Key words.—discriminant function, Everglades, foraging, habitat selection, prey availability, prey density, preyvulnerability, structural complexity, wading birds.
Waterbirds 33(4): 494-503, 2010
Numerous studies have emphasized theimportance of prey availability to the forag-ing success of wading birds and other avianpredators. Early studies highlighted the cor-relation between prey density and the func-tional and numerical response of the preda-tor (Holling 1959; Kushlan 1976a; Draulans1987; Ogden et al. 1976) but more recentwork has shown that predators and prey den-sities do not always coincide (Arengo andBaldassarre 1999; Gawlik 2002; Bennetts etal. 2006; Gwiazda and Amirowicz 2006). Onereason for the lack of correlation is thatpredators are responding to the availabilityof prey, which reflects vulnerability to cap-ture as well as prey density (Gawlik 2002). Ifprey occur in high densities, but are invul-nerable to capture, predators will respond asif prey density is low. Thus, the ability to cor-rectly predict the response of a predator tofood abundance depends on understandingfactors affecting prey vulnerability.
In the Florida Everglades, prey vulnera-bility varies seasonally in response to rainfallpatterns. During the dry season, receding
water across the landscape concentratesaquatic prey in topographic depressions.Under these conditions, prey become highlyvulnerable to capture and these high qualitypatches are strongly utilized by avian preda-tors such as wading birds (Kahl 1964; Kush-lan 1976a; Gonzalez 1997). In the pre-drain-age Everglades, this seasonal dry-down is be-lieved to have occurred predictably in timeand space. The current managed Evergladesis not only smaller in size (reduced foragingarea), but also exhibits less predictability indrydown patterns due to water managementoperations that support regional flood con-trol and water supply requirements. The un-predictable manner in which prey now be-comes available throughout the landscapemay have contributed to as much as a 78%decline of some species of Everglades wad-ing birds (Crozier and Gawlik 2003a).
Although wading birds in the Evergladesforage and nest sympatrically, individual spe-cies respond differently to changes in preyavailability (Frederick and Collopy 1989;Crozier and Gawlik 2003a). Gawlik (2002)

WADING BIRD FORAGING HABITAT SELECTION 495
showed experimentally that some species ofwading birds, termed searchers, most nota-bly Wood Stork (Mycteria americana), WhiteIbis (Eudocimus albus) and Snowy Egret(Egretta thula), are influenced more stronglyby changes in prey availability than otherspecies. These species select the highestquality patches (i.e. high prey density andhigh prey vulnerability) throughout thelandscape and abandon patches quickly asprey availability declines (Charnov 1976;Gawlik 2002). Therefore, changes in land-scape scale habitat characteristics that affectprey availability (e.g. water depth, topogra-phy, and vegetation) should be especially im-portant to these species, and it is these spe-cies that have shown the greatest declines inbreeding populations (Crozier and Gawlik2003a).
Water depth has been repeatedly demon-strated to be an important determinant ofhabitat quality (Kahl 1964; Kushlan 1976a;Powell 1987; Bancroft et al. 2002) but littledata exists describing the effects of otherhabitat characteristics, most notably, vegeta-tive structure. In aquatic ecosystems smalleraquatic prey avoid predation by larger pisciv-orous fish by seeking refuge in structurallycomplex habitats such as emergent or sub-merged macrophytes (Crowder and Cooper1982; McIvor and Odum 1988; Jordan et al.1996; Snickars et al. 2004). However, little isknown about how habitat characteristics likestructural complexity affect prey vulnerabili-ty in aquatic systems where predators in-clude avian piscivores. Factors contributingto increased prey vulnerability may be onereason wading birds forage disproportion-ately within more open habitat (Surdick1998; Bancroft et al. 2002; Stolen 2006).Studies have attempted to describe how hab-itat variables such as vegetation density orvegetation type influence the foraging deci-sions of wading birds (Hoffman et al. 1994;Smith et al. 1995; Bancroft et al. 2002); how-ever, they have done so at a coarse spatialscale with broad environmental categories.While these studies have been integral to un-derstanding the preferred habitat of wadingbirds in the Everglades, they do not offer anexplanation of the mechanisms underlying
foraging habitat selection decisions made bybirds occurring at finer spatial scales.
In this study, our objectives were to iden-tify fine scale habitat features related tostructural complexity that are important towading bird foraging and to test how specifichabitat features affect prey vulnerability. Totest for the importance of foraging habitatattributes to wading birds we conducted anobservational study (hereafter habitat selec-tion study) in which we compared habitatcharacteristics at random sites throughoutthe Florida Everglades to wading bird forag-ing sites. We predicted that if structural com-plexity decreased prey vulnerability to avianpredation, birds would chose to forage inmore open habitat. We also conducted anexperiment (hereafter prey vulnerability ex-periment) in which we manipulated waterdepth and structural complexity in three en-closures in a shallow marsh to quantify theireffects on wading bird foraging success. Wepredicted that structural complexity woulddecrease prey vulnerability to avian preda-tion; therefore, wading bird foraging successwould be greater in enclosures with moreopen habitat.
METHODS
Study Area
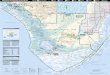
The habitat selection study was conducted through-out the freshwater marshes of the Everglades duringJanuary - May of 2005 and 2006. The Water Conserva-tion Areas (WCA-1, WCA-2A and 2B, WCA-3A and 3B),Everglades National Park (ENP), and Big Cypress Na-tional Park (BCNP) were all included in this study(Fig.1). Sampling occurred predominantly in sloughand wet prairie habitats. Slough communities were typi-cally dominated by White Water Lily (Nymphaea odorata)and were common in the WCAs, whereas wet prairiehabitats were dominated by Spikerushes (Eleocaris spp.)and Maidencane (Panicum spp.) and were typically en-countered in ENP, BCNP, and the western portion ofWCA-3A.
The prey vulnerability experiment was performedconcurrent to the habitat selection study during the dryseason (January-February) of 2006. This study was con-ducted within the Loxahatchee Impoundment Land-scape Assessment (LILA) facility at the Arthur R.Marshall Loxahatchee National Wildlife Refuge in PalmBeach County, Florida. LILA is situated directly adja-cent to WCA-1 and is an experimental facility consistingof four replicate 7-ha impoundments (hereafter macro-cosms) with topographic features that mimic key physi-cal attributes of the Everglades. A re-circulating watersystem allowed the water levels in each macrocosm to be

496 WATERBIRDS
manipulated. The advantage of conducting this study inLILA rather than in the Everglades was the control overwater depth and the large number of free-ranging wad-ing birds at the site.
Experimental Design
Habitat Selection Study. A multi stage sampling designwas implemented in this study consisting of four stratalevels; landscape units (LSU), primary sampling units(PSU), sampling sites, and 1-m2 sub-samples. The land-scape units and primary sampling units remained fixedthroughout the duration of the project and were sam-pled repeatedly throughout the two years. The sam-pling sites and the sub-samples were randomly selectedeach year. The landscape units were pre-defined andwere delineated based primarily upon hydroperiod (thenumber of days an area is inundated with water) andvegetation type.
Within each LSU, five PSUs 500 m
× 500 m wererandomly selected. Two random points within eachPSU were located by helicopter and the closest suitablehabitat to those points was chosen as the sampling lo-cations. In this study, we were not interested in com-paring used sites with random sites that were obviouslyunsuitable wading bird foraging habitat (based on ear-lier studies; Kushlan 1976a; Hoffman et al. 1994; Smithet al. 1995; Bancroft et al. 2002). Therefore, we definedsuitable habitat, a priori, as areas of sparse to moderatevegetation that had a third of the surface area coveredwith water and a water depth of
≤20 cm. Thus, not allLSUs were sampled each year because annual differ-ences in rainfall did not always produce suitable habi-tat.
The physical features of each random site werecharacterized by measuring a suite of habitat character-istics in 0.5 m2 quadrants every 5 m along a transect pass-ing through the site. Transects extended in eachbearing direction (E and W) for a maximum transectlength of 100 m. Habitat characteristics measured in-cluded: the density of emergent and submerged vegeta-tion, the height of the emergent plant above the surfaceof the water, the thickness of the flocculent layer (aloose aggregation of unconsolidated organic matter;hereafter floc), and the percent of floating periphytonmat. Vegetation density was characterized using thepoint-quarter method (Cottam and Curtis 1956), wherethe ‘nearest-neighbor’ distance is inversely related tovegetation density (i.e. large nearest neighbor distancesrepresent more open habitat). In addition to measuringfine scale habitat characteristics, the mean density offish and invertebrates at each site was estimated using a1-m2 throw trap; a mesh box open at the top and bottomwhich encloses a known volume of water when it isthrown. This sampling method has been shown to pro-vide statistically precise density information and hasproven to be the best method for sampling small fish inshallow marsh habitats (for review see Kushlan 1981b;Chick et al. 1992).
We evaluated wading bird foraging sites by sam-pling at locations with large aggregations of foragingbirds. Flocks of mixed species wading birds (>30birds) were located by helicopter within the samelandscape unit in which random sampling was beingconducted. Across all LSUs, the distance between for-aging and random sites averaged a minimum of 2.7km and a maximum of 9.8 km (range 213 m to 25.6km). Random and foraging sites were sampled within
the same timeframe (±1 wk) to ensure that hydrologyand vegetation structure was similar at both sites inorder to make relevant comparisons. We sampled thearea in which the foraging aggregation was mostheavily concentrated. Habitat characteristics weremeasured in the same manner described for randomsites.
Prey Vulnerability Experiment. To determine the effectof structural complexity on prey vulnerability, we con-structed three 5m
× 5m enclosures, 50 m apart fromone another in one of the LILA macrocosms. Each en-closure was constructed with polyethylene mesh whichwas attached to a frame and buried beneath the sedi-ment so that each enclosure was self-contained. Blad-derwort (Utricularia sp.), one of the most commonsubmerged plants in the Everglades, was added to eachenclosure at a stocking rate of 0 L/m2, 2 L/m2, or 5 L/m2. Treatments were randomly assigned to the enclo-sures and remained the same throughout the durationof the experiment. Each enclosure was stocked with1,500 fish (60 fish/m2), a moderate density in the natu-ral Everglades system during the dry season. We usedEastern Mosquitofish (Gambusia holbrooki), one of themost abundant fish species in the Everglades (Trexleret al. 2002) and one that is important in the diet ofmany wading birds (Jenni 1969; Smith 1997). Only fish
≥2 cm were used because this is the size typically pre-ferred by wading birds (Niethammer and Kaiser 1983;Kent 1986; Kushlan and Bildstein 1992). Existing resi-dent fish, as well as any vegetation, were removed be-fore stocking.
The effects of structural complexity on prey vul-nerability can be mediated by water depth. As waterdepth decreases during the dry season, submergedvegetation collapses, creating a mat of vegetation.Dense vegetation can confound visual perceptionand reduce the foraging efficiency of some predators(Jordan et al. 1996; Priyadarshana et al. 2001). Con-versely, structural complexity at low water depths mayincrease prey vulnerability to predation by inhibitingprey movement. This relationship between structuralcomplexity and water depth has the potential to ben-efit the predator or the prey and could affect howwading birds select a foraging patch. Consequently,this experiment was conducted at three different wa-ter depths; 10 cm, 16 cm, and 22 cm. The 10-cmdepth treatment was chosen as the minimum be-cause, in a pilot study, shallower water depths in-duced fish mortality due to high water temperatureand low dissolved oxygen (R. P., pers. obs.). The 22-cm depth treatment was chosen as the maximum be-cause we did not want to eliminate smaller heronsfrom the study due to treatments exceeding theirmaximum foraging depth capacity. Further, a depththreshold exists beyond which deep water providesfish refuge from avian predation (Gawlik 2002).
To attract wading birds to the experimental site,we used 1-2 white wading bird decoys per enclosure(Crozier and Gawlik 2003b). Birds foraging withinthe enclosures were observed using a 60x spottingscope from the nearest levee of the macrocosm andthe time interval (sec) between captures was mea-sured and averaged for each foraging bout. The ob-servation session for each animal continued for aslong as that individual remained in the area unlessanother bird stopped to forage (Altman 1973). Then,observations lasted for a minimum of 15 min at whichtime the other individual was observed. The number

WADING BIRD FORAGING HABITAT SELECTION 497
of birds observed on any day ranged between one andfive, for a total of 20 birds observed throughout theduration of the experiment. Observations were con-sidered independent because there was never an in-stance when there was more than one bird foragingwithin the same enclosure at the same time and therewas never an instance where more than two birdswere foraging at the site at the same time. We ob-served foraging birds for three consecutive days ateach of the three water depth treatment intervals fora total of nine experimental observation days.
Statistical Analysis
Habitat Selection Study. Discriminant function analysis(DFA; PROC DISCRIM; SAS Institute 2003) was used todetermine which habitat characteristics were importantto foraging site selection by wading birds. The analysiswas conducted on the 2005 data only since small samplesizes in 2006 precluded the use of this procedure. Allhabitat variables were assessed for equal variance (Lev-ene’s homogeneity of variance test) and univariate nor-mality (Shapiro-Wilks test statistic). Homogeneity andnormality were not violated and the original variableswere used in all analyses. Multicollinearity problemswere detected using the Pearson correlation procedure.The F-values of intercorrelated variables were com-pared and the variable with the larger value remained inthe dataset.
To establish the optimal combination of discrimi-nating variables to be used in the DFA we utilized astepwise-selection procedure (PROC STEP; SAS Insti-tute 2003). All groups met the minimum sample sizerequirements (N
≥ 3P, where P is the number of dis-criminating variables; Williams and Titus 1988) andthe normality of the canonical scores was assessed.Validation of our results was determined using thejackknife resampling procedure (PROC DISCRIM;SAS Institute 2003). Since a certain percentage ofsamples in any dataset can be expected to be correctlyclassified based upon chance alone, we applied Co-hen’s kappa statistic which provided a chance-cor-rected classification measure and supplied a morerealistic measure of discrimination (Cohen 1960; Ti-tus et al. 1984).
In addition to the DFA, we also present means and95% CI for all measured variables to identify interannu-al differences in habitat characteristics and wading birdresponses during both years. Means represent a singlepooled estimate for all LSUs combined. To calculate themeans and variance, we used PROC SURVEYMEANS(SAS Institute 2003) because this procedure takes intoaccount that the sites were clustered in space due to thestratified sampling design. PROC SURVEYMEANS usesthe Taylor expansion method to estimate error, whichhas the benefit of being able to handle unbalanced da-ta, unlike the traditional methods of variance estima-tion.
Prey Vulnerability Experiment. To determine the effectof structural complexity and water depth on wadingbird foraging efficiency we used a two-way factorialANOVA (PROC GLM; SAS Institute 2003). The re-sponse variable was the mean time interval between cap-tures and the fixed effects were water depth, vegetationdensity, and the interaction between water depth andvegetation. Capture intervals were square root trans-formed to meet the assumption of normality. We as-sumed that the data obtained between days was
independent because wading birds have been shown toreassess foraging conditions on a daily basis (Gawlik2002; Master et al. 2005).
RESULTS
Habitat Selection Study
In 2005, we conducted a total of 95transects at 69 random sites and 27 foragingsites (Fig. 1) distributed throughout twelveand seven landscape units, respectively. In2006, we characterized sites along 72transects at 59 random sites and 13 foragingsites (Fig. 1) in seven and six landscape units,respectively.
The discriminant function analysis re-vealed clear differences in habitat character-istics between used and random sites in 2005(Fig. 2). The canonical discriminant func-tion explained 41% of the variation in thedata and the overall correct classification
Figure 1. Spatial extent over which sampling occurredduring the dry seasons (Jan-May) of 2005 and 2006 inthe Florida Everglades. Sampling sites north of Tami-ami Trail are part of the Water Conservation Areas(WCA) and south of Tamiami Trail are part of Ever-glades National Park (ENP) or Big Cypress NationalPark (BCNP). Sampling sites include both random andforaging locations.

498 WATERBIRDS
rate was 81%, which was 37% better thanchance alone. Three of the four discriminat-ing variables that entered the model wereidentified as being the most important togroup separation; these were the thicknessof the flocculent layer, emergent stem densi-ty, and prey density (Table 1). Foraging siteshad thicker floc layers, less emergent vegeta-tion, and higher prey densities than did ran-dom sites.
Interannual Differences
A comparison of means (± 95% CI) indi-cated that in 2005, wading birds selected for-
aging sites with a thicker floc layer than atrandom sites [x– = 2.2 cm ± 0.8 (random)and 5.0 cm ± 1.6 (forage)]. In 2006, overlap-ping confidence intervals indicated thatthere was no significant difference in flocthickness between foraging and randomsites [x– = 0.5 cm ± 1.2 (random) and 1.7 cm± 2.1 (forage)], but it tended to be greater atforaging sites, as it was in 2005 (Table 2).
Birds selected foraging sites with signifi-cantly less dense emergent vegetation thanrandom sites [x– = 9.1 cm ± 0.9 (random)and 13.6 cm ± 2.5 (forage); larger values rep-resent more open habitat] in 2005; in 2006,the density of emergent vegetation was simi-lar between random and foraging sites [x– =13.0 cm ± 1.1 (random) and 13.6 cm ± 3.7(forage)]. In both years birds selected forag-ing sites with identical vegetation densities(Table 2).
In 2005, birds selected foraging sites witha higher mean prey density than at randomsites, but the high variability resulted in over-lapping 95% confidence intervals [x– = 81prey/m2 ± 27(random) and 184 prey/m2 ±196 (forage)]. In 2006, birds selected siteswith a prey density similar to random sites[x– = 142 prey/m2 ± 70.8 (random) and 126prey/m2 ± 70.5 (forage)]. However, prey atrandom sites was considerably higher in2006 than in 2005 (Table 2).
Similar to the DFA, a comparison of themean density of submerged vegetation be-tween random and foraging sites indicatedthat there was no major difference in 2005 or2006 [2005: [x– = 24.1 cm ± 13.5 (random)and 20.4 cm ± 8.6 (forage); 2006: 11.9 cm ±4.3 (random) and 8.8 cm ± 4.7 (forage); larg-
Figure 2. Distribution of discriminant function canoni-cal scores for random and foraging sites in the dry sea-son of 2005 in the Florida Everglades. Larger canonicalscores are associated with habitat characteristics thathad larger measurements. Floc thickness and emergentvegetation are listed (in order of importance) becausethey contributed the most to group separation (loadings≥0.57).
Table 1. Habitat variables for wading bird foraging sites and random sites throughout the Florida Everglades withthe corresponding correlation coefficients for the canonical discriminant function. Bold values represent signifi-cant differences between the two groups.
Habitat Variables Forage Sites (N = 26)a Random Sites (N = 67) DFA correlation coefficientb
Floc Thickness (cm) 5.0 ± 1.6c 2.2 ± 0.8 0.91Emergent Vegetation (cm)d 13.6 ± 2.5 9.1 ± 0.9 0.57Prey Density (prey/m2) 183.7 ± 196 80.6 ± 27 0.44Floating Periphyton (%) 4.8 ± 3.9 1.8 ± 1.3 0.34
aSample size.bCoefficients < 0.44 were deemed unimportant.cMean ± 95% Confidence Interval.dHigher values for emergent vegetation = more open habitat.

WADING BIRD FORAGING HABITAT SELECTION 499
er values represent more open habitat].However, the nonsignificant tendency wassimilar in both years; the trend being thatbirds chose foraging sites that had more sub-merged vegetation.
Prey Vulnerability Study
Over the observation period of nine days,the Snowy Egret (N = 20) was the only speciesto consistently forage within the experimen-tal enclosures so our analysis was restricted tothis species. We were unable to detect a differ-ence in Snowy Egret capture intervals amongany of the submerged vegetation treatments(F2, 30 = 2.27, P = >0.05) or the water depthtreatments (F2, 30 = 1.41, P > 0.05; Fig. 3).
DISCUSSION
Field observations demonstrated that, inaddition to coarse scale habitat selectiondocumented by other studies (Hoffman et al.1994; Smith et al. 1995; Bancroft et al. 2002),wading birds also select habitat characteris-tics at fine spatial scales. The discriminantfunction analysis indicated that wading birdsselected foraging sites with a thicker floc lay-er and less emergent vegetation than thatgenerally available in the landscape. Despitesupport for fine scale habitat selection, thecanonical scores also revealed that there wassimilarity between random and foragingsites. This similarity exists because of howavailable habitat was defined in this study.We chose sites based on prior knowledge ofthe habitat in which wading birds typicallyforage (Hoffman et al. 1994; Smith et al.1995; Bancroft et al. 2002). The predefinedcriteria precluded sites that were unsuitableforaging habitat so that within each LSU allsampled locations were similar to each otherin regard to hydrology and vegetative struc-ture. Therefore, the differences that weredetected among these similar sites are morelikely to be biologically meaningful insteadof an artifact of a random sampling design.
The response of birds choosing foragingsites that had thicker floc layers is counterin-tuitive and the opposite of what was expect-ed (e.g. that increased floc thickness wouldreduce foraging site quality by decreasingprey vulnerability). Prey species appear touse the floc layer to reduce their visibilityand decrease the likelihood of capture (R.P., pers. obs.). One explanation for the con-
Table 2: Means (± 95% CI) for measured habitat characteristics at random and foraging sites throughout the Ever-glades in 2005 and 2006.
Habitat Variables
2005 Sites 2006 Sites
Random(N = 69)
Forage(N = 27)
Random(N = 59)
Forage(N = 13)
Emergent Vegetation (cm) 9.1 ± 0.9 13.6 ± 2.5 13.0 ± 1.1 13.6 ± 3.7Submerged Vegetation (cm) 24.1 ± 13.5 20.4 ± 8.6 11.9 ± 4.3 8.8 ± 4.7Floc Thickness (cm) 2.2 ± 0.8 5.0 ± 1.6 0.5 ± 1.2 1.7 ± 2.1Emergent Height (cm) 45.4 ± 2.3 40.8 ± 5.5 36.4 ± 2.7 30.7 ± 5.8Floating Periphyton (%) 1.8 ± 0.6 4.8 ± 1.9 6.0 ± 2.0 8.3 ± 4.9Prey Density (prey/m2) 80.6 ± 27 183.7 ± 196 142 ± 70.8 126 ± 70.5
Figure 3. Mean time interval (s) between captures madeby Snowy Egrets (n = 20) foraging within three vegeta-tion treatments at three different water depths. Blackbars within each box plot denote the median; boxes en-compass 50% of the data while whiskers (SE) encom-pass 90% of the data; open circles represent the 5th and95th percentile outliers. The missing box plot at the 16-cm water depth indicates that no birds foraged withinthe open vegetation treatment.

500 WATERBIRDS
tradictory response by birds is that the thick-ness of the floc layer is indicative of other fac-tors that increase the quality of a foragingsite. Flocculent material is rich in organicmatter and has been shown to be an impor-tant source of nutrients in an oligotrophicsystem (Wood 2005) and nutrient rich areasin the Everglades have higher aquatic animaldensities (Turner et al. 1999; McCormick etal. 2004; Rehage and Trexler 2006). Further,parts of the system having the thickest floclayers are often open slough areas that areassociated with intermediate to long hydro-period marshes (Leonard et al. 2006). In-creased hydroperiod has been linked withincreased fish densities and increased fishbiomass (Loftus and Eklund 1994; Trexler etal. 2002; Chick et al. 2004). Many studieshave documented that birds use visual cueswithin the landscape to reduce search timewhen choosing foraging locations (Kushlan1976b; Custer and Osborn 1978; Gawlik andCrozier 2007; Master et al. 2005). Thus, thethickness of the floc layer could act as a visu-al cue indicating favorable foraging habitat.Another explanation could be that sites thathad the thickest flocculent layers were foundin parts of the system that had increased mi-crotopographic variation (e.g. ridge andslough habitat, peat pop-ups, or solutionholes). These areas often become denselyconcentrated with prey as the landscapedries. In this case, wading birds may not beresponding to floc thickness, but instead tothe increased microtopography, which was aparameter not considered in this study.
In both study years, wading birds selectedforaging sites with less emergent vegetationthan random sites and with similar densitiesof emergent vegetation despite the fact thatthis habitat characteristic varied greatly inthe landscape. These results are consistentwith other habitat selection studies that havesuggested that emergent vegetation is an im-portant variable for foraging habitat selec-tion at coarse (Hoffman et al. 1994; Smith etal. 1995; Bancroft et al. 2002) as well as finespatial scales (Surdick 1998). Wading birdsmay be selecting areas of sparse emergentvegetation to reduce the risk of predation(Metcalfe 1984; Cresswell 1994). However,
healthy adult wading birds are rarely preyedupon and the explanation for this lack of pre-dation, to date, has never been tested (Kush-lan 1981a). It is more likely that wading birdsavoid areas of dense emergent vegetation be-cause they inhibit foraging success by reduc-ing the vulnerability of prey. Studies of wad-ing birds foraging in rice fields have indicat-ed that birds expend more energy per unittime in this habitat type and also have de-creased capture rates as a result of the densenature of these plants (Campos and Lukuona2001; Richardson et al. 2001). Batzer andShurtleff (1999) found that a pond beingmanaged for feeding Wood Storks was un-derutilized by this species because stockedprey sought refuge in nearby macrophytebeds and were not vulnerable to capture.
Conversely, some species of waterbirdshave been found to forage at the interface ofopen water and vegetation (Safran 2000;Bennetts et al. 2006; Stolen et al. 2007) and topreferentially select low stature wet-prairiehabitats despite the existence of more openareas (Smith et al. 1995). If wading birds areresponding to availability, they must assessprey density and prey vulnerability simulta-neously. Therefore, if birds choose to foragein or near highly vegetated areas, prey densi-ty must be high enough to offset reducedprey vulnerability.
Unlike the response to emergent vegeta-tion, birds seemed to be inhibited little bysubmerged vegetation. This variable did notenter the discriminant model as being a sig-nificant contributor to group separation norwas there any difference in either 2005 or2006 in the mean density of submerged vege-tation between random sites and sites selectedby birds. Further, this result was also support-ed experimentally; we were unable to detectan effect of submerged vegetation on captureintervals of foraging Snowy Egrets. This sug-gests that prey vulnerability was not sufficient-ly decreased due to increased submergedstructural complexity to affect foraging effi-ciency. These results were consistent with oth-er studies that have shown that wading birdforaging success is not negatively affected bysubmerged vegetation (see Safran 2000; Ad-ams and Frederick 2008; Lantz 2008).

WADING BIRD FORAGING HABITAT SELECTION 501
The lack of a response to submerged veg-etation in our studies may be the result ofseveral factors. Submerged vegetation maynot provide enough structural complexity toinhibit foraging success. Gawlik (2002) andMaster et al. (2005) demonstrated that wad-ing birds respond strongly to habitat with ac-tive and accessible prey. Submerged vegeta-tion likely does not prevent the birds fromrecognizing disturbance of the water’s sur-face caused by surface breathing fish or anescape response by fish to an approachingwading bird. Coupled with the fact that preyare easily flushed from cover by the typicalegret behavior of foot-stirring, this could al-low wading birds to be effective foragers inareas of dense submerged vegetation. Papa-kostas et al. (2005) found that Squacco Her-ons (Ardeola ralloides) had higher successrates foraging in open water with submergedvegetation relative to other microhabitattypes, suggesting that submerged vegetationmay be beneficial to foraging wading birds.In addition, birds are probably highly visibleto their prey in open water habitats. Sub-merged macrophyte beds may act as a bufferagainst moving water caused by herons’ wad-ing which may allow birds to approach closerto prey without being noticed (Matsunaga2000).
The comparison of means between thetwo study years indicated apparent differenc-es in interannual habitat selection. In 2005,wading birds appeared to be selecting forag-ing sites that were different from what wasavailable in the landscape; in 2006, birds se-lected sites that were more similar to thelandscape. One explanation for this was thedisparate hydrological conditions betweenthe two years. The 2005 dry season experi-enced several rainfall events which resultedin the reversal of the seasonal water reces-sion. In 2006, ideal hydrological conditionsexisted throughout the dry season. In yearswith ideal hydrology, high densities of preythroughout the landscape are probablymore important to wading bird success thanfine scale variations in habitat characteris-tics. However, in years with poor hydrologi-cal conditions the penalty for choosing a lowquality foraging site may be much greater
than in years with more optimal conditions.Under these conditions, habitat structurewould become an increasingly importantcomponent of habitat quality.
Species that reside in highly dynamic,patchy environments must rely heavily onsocial and environmental cues to locatehigh quality foraging habitat (Weins 1976;Green and Leberg 2005; Gawlik and Cro-zier 2007). This study identified specificfine scale habitat variables that are impor-tant to wading bird foraging site selectionand also identified landscape characteris-tics which may act as visual cues guidingbirds to higher quality foraging sites. Al-though strong habitat selection was appar-ent in this study, it is still unclear how thesehabitat variables may influence foragingsuccess. Habitat preference is thought to beadaptive and affect fitness (Pulliam andDanielson 1991; Martin 1998). Therefore, itis likely that birds benefit from choosingforaging locations having specific habitatcharacteristics even though these benefitshave not yet been quantified. The advan-tage of foraging at sites with specific habitatcharacteristics may not be realized until for-aging conditions are sub-optimal, such asthey are during times of high water condi-tions, extreme drought, or in areas with lowprey density.
Wading birds are considered to be an in-dicator of ecosystem health. Thus, many res-toration plans, including the Comprehen-sive Everglades Restoration Plan (CERP),use healthy wading bird populations as a tar-get for restoration success. However, beforewading birds can be reliably used as a perfor-mance measure it is necessary to have a clearunderstanding of the linkage between wad-ing birds and the ecosystem processes thatare being restored (Gawlik 2006). Knowl-edge of the type of habitat characteristicsthat create high quality wading bird foragingsites in the Florida Everglades is an impor-tant step in beginning to elucidate this link-age. Incorporating this type of informationinto continually evolving conceptual modelsin an adaptive management framework willassist managers in gauging future restorationprogress.

502 WATERBIRDS
ACKNOWLEDGMENTS
The South Florida Water Management District pro-vided funding for this project. We are grateful to thestaff of the Arthur R. Marshall Loxahatchee NationalWildlife Refuge and the Florida Fish and Wildlife Con-servation Commission Non-native Fish Research Centerfor cooperation and logistical support. We thank all whoassisted in the field: B. Botson, P. Heideman, H. Her-ring, M. Johnston, S. Lantz, J. Nagy and K. Simpson; wefurther thank B. Botson for data management and assis-tance with statistical analysis. G. Herring, H. K. Herringand two anonymous reviewers commented on themanuscript.
LITERATURE CITED
Adams, E. M. and P. C. Frederick. 2008. Effects of meth-ylmercury and spatial complexity on foraging behav-ior and foraging efficiency in juvenile White Ibises(Eudocimus albus). Environmental Toxicology andChemistry 27: 1708-1712.
Altman, J. 1973. Observational study of behavior: sam-pling methods. Behaviour 49: 226-267.
Arengo, F. and G. A. Baldassarre. 1999. Resource vari-ability and conservation of American Flamingos incoastal wetlands of Yucatan, Mexico. Journal of Wild-life Management 63: 1201-1212.
Bancroft, G. T., D. E. Gawlik and K. Rutchey. 2002. Dis-tribution of wading birds relative to vegetation andwater depths in the northern Everglades of Florida,USA. Waterbirds 25: 265-277.
Batzer, D. P. and A. S. Shurtleff. 1999. Managing fishand invertebrate resources in a Wood Stork feedingpond. Journal of Freshwater Ecology 14: 159-165.
Bennetts, R. E., P. C. Darby and L. B. Karunaratne. 2006.Foraging patch selection by Snail Kites in responseto vegetation structure and prey abundance andavailability. Waterbirds 29: 88-94.
Campos, F. and J. M. Lekuona. 2001. Are rice fields asuitable foraging habitat for Purple Herons duringthe breeding season? Waterbirds 24: 450-452.
Charnov, E. L. 1976. Optimal foraging, the marginal valuetheorem. Theoretical Population Biology 9: 129-136.
Chick, J. H., F. Jordan, J. P. Smith and C. C. McIvor.1992. A comparison of four enclosure traps andmethods used to sample fishes in aquatic macro-phytes. Journal of Freshwater Ecology 7: 353-361.
Chick, J. H., C. R. Ruetz III and J. C. Trexler. 2004. Spa-tial scale and abundance patterns of large fish com-munities in freshwater marshes of the FloridaEverglades. Wetlands 24: 652-664.
Cohen, J. 1960. A coefficient of agreement for nominalscales. Educational and Psychological Measurement20: 37-46.
Cottam, G. and J. T. Curtis. 1956. The use of distancemeasure in phytosociological sampling. Ecology 37:45-460.
Cresswell, W. 1994. Age-dependent choice of Redshank(Tringa totanus) feeding location: profitability orrisk? Journal of Animal Ecology 63: 589-600.
Crowder, L. B. and W. E. Cooper. 1982. Habitat structur-al complexity and the interaction between Bluegillsand their prey. Ecology 63: 1802-1813.
Crozier, G. E. and D. E. Gawlik. 2003a. Wading bird nest-ing effort as an index to wetland ecosystem integrity.Waterbirds 26: 303-324.
Crozier, G. E. and D. E. Gawlik. 2003b. The use of de-coys as a research tool for attracting wading birds.Journal of Field Ornithology 74: 53-58.
Custer, T. W. and R. G. Osborn. 1978. Feeding habitatuse by colonially-breeding herons, egrets, and ibisesin North Carolina. Auk 95: 733-743.
Draulans, D. 1987. The effect of prey density on forag-ing behaviour and success of adult and first-yearGrey Herons (Ardea cinerea). Journal of Animal Ecol-ogy 56: 479-493.
Frederick, P. C. and M. W. Collopy. 1989. Nesting suc-cess of five Ciconiiform species in relation to waterconditions in the Florida Everglades. Auk 106: 625-634.
Gawlik, D. E. 2002. The effects of prey availability on thenumerical response of wading birds. EcologicalMonographs 72: 329-346.
Gawlik, D. E. 2006. The role of wildlife science in wet-land ecosystem restoration: lessons from the Ever-glades. Ecological Engineering 26: 70-83.
Gawlik, D. E. and G. E. Crozier. 2007. A test of cues af-fecting habitat selection by wading birds. Auk 124:1075-1082.
Gonzalez, J. A. 1997. Seasonal variation in the foragingecology of the Wood Stork in the southern Ilanos ofVenezuela. Condor 99: 671-680.
Green, M. C. and P. L. Leberg. 2005. Flock formationand the role of plumage colouration in Ardeidae.Canadian Journal of Zoology 83: 683-693.
Gwiazda, R. and A. Amirowicz. 2006. Selective foragingof Grey Heron (Ardea cinerea) in relation to densityand composition of the littoral fish community in asubmontane dam reservoir. Waterbirds 29: 226-232.
Hoffman, W., G. T. Bancroft and T. J. Sawicki. 1994. For-aging habitat of wading birds in the water conserva-tion areas of the Everglades. Pages 585-614 inEverglades: the Ecosystem and its Restoration (S. M.Davis and J. C. Ogden, Eds.). St. Lucie Press, DelrayBeach, Florida.
Holling, C. S. 1959. Some characteristics of simple typesof predation and parasitism. Canadian Entomolo-gist 91: 385-398.
Jenni, D. A. 1969. A study of the ecology of four speciesof herons during the breeding season at Lake AliceAlachua County, Florida. Ecological Monographs39: 245-270.
Jordan, F., C. J. DeLeon and A. C. McCreary. 1996. Pre-dation, habitat complexity, and distribution of thecrayfish Procambarus alleni within a wetland habitatmosaic. Wetlands 16: 452-457.
Kahl, M. P., Jr. 1964. Food ecology of the Wood Stork(Mycteria americana) in Florida. Ecological Mono-graphs 34: 97-117.
Kent, D. M. 1986. Behavior, habitat use, and food ofthree egrets in a marine habitat. Colonial Water-birds 9: 25-30.
Kushlan, J. A. 1976a. Wading bird predation in a season-ally fluctuating pond. Auk 93: 464-476.
Kushlan, J. A. 1976b. Feeding behavior of North Ameri-can herons. Auk 93: 86-94.
Kushlan, J. A. 1981a. Resource use strategies of wadingbirds. Wilson Bulletin 93: 145-163.
Kushlan, J. A. 1981b. Sampling characteristics of enclo-sure fish traps. Transactions of the American Fisher-ies Society 110: 557-562.
Kushlan, J. A. and K. L. Bildstein. 1992. White Ibis. InThe Birds of North America, No. 9 (A. Poole, P.Stettenheim and F. Gill, Eds.). Philadelphia: The

WADING BIRD FORAGING HABITAT SELECTION 503
Academy of Natural Sciences; Washington, D.C.:The American Ornithologists’ Union.
Lantz, S. 2008. The effects of water depth and vegeta-tion on wading bird foraging habitat selection andforaging success in the Everglades. UnpublishedM.Sc.Thesis, Florida Atlantic University, Boca Raton,Florida.
Leonard, L., A. Croft, D. Childers, S. Mitchell-Bruker,H. Solo-Gabriele and M. Ross. 2006. Characteristicsof surface-water flows in the ridge and slough land-scape of Everglades National Park: implications forparticulate transport. Hydrobiologia 569: 5-22.
Loftus, W. F. and A. Eklund. 1994. Long-term dynamicsof an Everglades small-fish assemblage. Pages 461-484 in Everglades: the ecosystem and its restorations(S. M. Davis and J. C. Ogden, Eds.). St. Lucie Press,Delray Beach, Florida.
Martin, T. E. 1998. Are microhabitat preferences of co-existing species under selection and adaptive? Ecol-ogy 79: 656-670.
Master, T. L., J. K. Leiser, K. A. Bennett, J. K. Bretsch andH. J. Wolfe. 2005. Patch selection by Snowy Egrets.Waterbirds 28: 220-224.
Matsunaga, K. 2000. Effects of tidal cycle on the feedingactivity and behavior of Grey Herons in a tidal flat inNotsuke Bay, northern Japan. Waterbirds 23: 226-235.
McCormick, P. V., R. B. E. Shuford III and P. S. Rawlik.2004. Changes in macroinvertebrate communitystructure and function along a phosphorus gradientin the Florida Everglades. Hydrobiologia 529: 113-132.
McIvor, C. C. and W. E. Odum. 1988. Food, predationrisk, and microhabitat selection in a marsh fish as-semblage. Ecology 69: 1341-1351.
Metcalfe, N. B. 1984. The effects of habitat on the vigi-lance of shorebirds: is visibility important? AnimalBehavior 32: 981-985.
Niethammer, K. R. and M. S. Kaiser. 1983. Late summerfood habits of three heron species in northeasternLouisiana. Colonial Waterbirds 6: 148-153.
Ogden, J. C., J. A. Kushlan and J. T. Tilmant. 1976. Preyselectivity by the Wood Stork. Condor 78: 324-330.
Papakostas, G., S. Kazantzidis, V. Goutner and I. Charal-ombidou. 2005. Factors affecting the foraging be-havior of the Squacco Heron. Waterbirds 28: 28-34.
Powell, G. V. N. 1987. Habitat use by wading birds in asubtropical estuary: Implications of hydrography.Auk 104: 740-749.
Priyadarshana, T., T. Asaeda and J. Manatunge. 2001.Foraging behaviour of planktivorous fish in artificialvegetation: the effects on swimming and feeding.Hydrobiologia 442: 231-239.
Pulliam, H. R. and B. J. Danielson. 1991. Sources, sinks,and habitat selection: A landscape perspective onpopulation dynamics. American Naturalist 137(Supplement): 50-66.
Rehage, J. S. and J. C. Trexler. 2006. Assessing the net ef-fect of anthropogenic disturbance on aquatic com-munities in wetlands: Community structure relativeto distance from canals. Hydrobiologia 569: 359-373.
Richardson, A. J., I. R. Taylor and J. E. Growns. 2001.The foraging ecology of egrets in rice fields in south-ern New South Wales, Australia. Waterbirds 24: 255-264.
Safran, R. J., M. A. Colwell, C. R. Isola and O. E. Taft.2000. Foraging site selection by nonbreeding White-faced Ibis. Condor 102: 211-215.
SAS Institute. 2003. SAS/STAT user’s guide. Version9.1. SAS Institute, Cary, North Carolina, USA.
Smith, J. P., J. R. Richardson and M. W. Collopy. 1995.Foraging habitat selection among wading birds(Ciconiiformes) at Lake Okeechobee, Florida, in re-lation to hydrology and vegetative cover. Archive ofHydrobiology Special Issues in Advanced Limnology45: 247-285
Smith, J. P. 1997. Nesting season food habits of 4 speciesof herons and egrets at Lake Okeechobee, Florida.Colonial Waterbirds 20: 198-220.
Snickars, M., A. Sandström and J. Mattila. 2004. Anti-predator behaviour of 0+ year Perca fluviatilis: Effectof vegetation density and turbidity. Journal of FishBiology 65: 1604-1613.
Stolen, E. D. 2006. Habitat selection and foraging suc-cess of wading birds in impounded wetlands in Flor-ida. Unpublished Ph.D. Dissertation, University ofFlorida, Gainesville, Florida.
Stolen E. D., J. A. Collazo and H. F. Percival. 2007. Scale-dependent habitat selection of nesting Great Egretsand Snowy Egrets. Waterbirds 30: 384-393.
Surdick, J. A., Jr. 1998. Biotic and abiotic indicators offoraging site selection and foraging success of fourCiconiiform species in the freshwater Everglades ofFlorida. Unpublished M.Sc.Thesis, University ofFlorida, Gainesville, Florida.
Titus, K., J. A. Mosher and B. K. Williams. 1984. Chance-corrected classification for use in discriminant anal-ysis: ecological applications. American Midland Nat-uralist 111: 1-7.
Trexler, J. C., W. F. Loftus, F. Jordan, J. H. Chick, K. L.Kandl, T. C. McElroy and O. L. Bass, Jr. 2002. Ecolog-ical scale and its implications for freshwater fisheriesin the Florida Everglades. Pages 153-181 in The Flor-ida Everglades, Florida Bay, and Coral Reefs of theFlorida Keys: an Ecosystem Sourcebook (J. W. Porterand K. G. Porter, Eds.). CRC Press, New York, NewYork.
Turner, A. M., J. C. Trexler, C. F. Jordan, S. J. Slack, P.Geddes, J. H. Chick and W. F. Loftus. 1999. Target-ing Ecosystem Features for Conservation: StandingCrops in the Florida Everglades. Conservation Biol-ogy 13: 898-911.
Weins, J. A. 1976. Population responses to patchy envi-ronments. Annual Review of Ecology and Systemat-ics 7: 80-120.
Williams, B. K. and K. Titus. 1988. Assessment of sam-pling stability in ecological application of discrimi-nant analysis. Ecology 69: 1275-1285.
Wood, A. D. 2005. Dynamics of detrital particulate or-ganic material in the ridge and slough landscape ofthe Everglades. Unpublished M. Sc. Thesis, FloridaInternational University, Miami, Florida.