Embed Size (px)
Citation preview

a
Annals of Biomedical Engineering,Vol. 31, pp. 548–563, 2003 0090-6964/2003/31~5!/548/16/$20.00Printed in the USA. All rights reserved. Copyright © 2003 Biomedical Engineering Society
Voronoi Polyhedra Analysis of Optimized Arterial Tree Models
RUDOLF KARCH,1 FRIEDERIKE NEUMANN,1 MARTIN NEUMANN,2 PAUL SZAWLOWSKI,1
and WOLFGANG SCHREINER1
1Department of Medical Computer Sciences, University of Vienna, Spitalgasse 23, A-1090 Wien, Austria and2Institute ofExperimental Physics, Section for Computational Physics, University of Vienna, Strudlhofgasse 4, A-1090 Wien, Austri
(Received 7 June 2001; accepted 4 February 2003)
-g-
on-
tesdel.
ybu-gndfxi-lingreatedrve
so-ivew-y.
-ter
i-licastrouales-
mi-
aloftoe-in
ar-tiess,the
ndantof
lud-
althetial
ralf
cter-oia
nof
s aned
ointne
ndna,23,
Abstract—Topological and metric properties of Voronoi polyhedra~VP! generated by the distal end points of terminal sements in arterial tree models grown by the method of cstrained constructive optimization~CCO! are analyzed with theaim to characterize the spatial distribution of their supply sirelative to randomly distributed points as a reference moThe distributions of the numberNf of Voronoi cell faces, cellvolume V, surface areaS, areaA of individual cell faces, andasphericity parametera of the CCO models are all significantldifferent from the ones of random points, whereas the distritions of V, S, and a are also significantly different amonCCO models optimized for minimum intravascular volume aminimum segment length (p,0.0001). The distributions oNf , V, andS of the CCO models are reasonably well appromated by two-parameter gamma distributions. We study scaof intravascular blood volume and arterial cross-sectional awith the volume of supplied tissue, the latter being represenby the VP of the respective terminal segments. We obsescaling exponents from 1.2060.007 to 1.0860.005 for intra-vascular blood volume and 0.7760.01 for arterial cross-sectional area. Setting terminal flows proportional to the asciated VP volumes during tree construction yields a relatdispersion of terminal flows of 37% and a coefficient of skeness of 1.12. ©2003 Biomedical Engineering Societ@DOI: 10.1114/1.1566444#
Keywords—Voronoi cell, Gamma distribution, Lognormal distribution, Scaling, Blood flow, Relative dispersion, Compusimulation.
INTRODUCTION
Arterial trees serving an organ fulfill the basic physological task to deliver blood to and remove metaboend products from the tissue they supply. During the ldecades arterial trees have been the subject of numemodeling studies, ranging from self-similar fractapproaches15,56 and anatomically derived branching tubmodels7,28,55 to combined geometric and functional vacular growth models.18,32,47 Still another approach istaken by the method of constrained constructive opti
Address correspondence to Rudolf Karch, University of VienDepartment of Medical Computer Sciences, SpitalgasseA-1090 Wien, Austria. Electronic mail: [email protected]
548
s
zation ~CCO!.48,49 Based on the hypothesis of optimdesign,45 CCO allows to generate arterial tree modelssome 104 vessel segments from a feeding artery downsmall arteries without directly using anatomical data. Dspite the numerous simplifying assumptions inherentthis model, CCO trees exhibit a realistic visual appeance and quite satisfactorily reproduce various properof real arterial trees, such as branching angle statistic50
diameter ratios of parent and daughter segments, andvolume of the large arteries.26
Concerning the relationship between structure afunction of arterial trees serving an organ, one importcharacteristic of a tree model is the spatial distributionits terminal blood supply points.32 The characterizationof spatial structures is a challenging issue,16 and variousapproaches have been suggested in the literature, incing nearest-neighbor distance methods,44 second-orderanalysis ~pair correlation function!,52 and fractaldescriptions.29 In this paper, we consider the terminlocations of CCO trees as a point pattern and applymethod of Voronoi analysis to characterize its spadistribution.
The Voronoi polyhedron associated with a cent~generating! point P is defined as the convex region ospace closer toP than to any other pointP8 of a givenset of points. The Voronoi tessellation of a set of distinpoints leads to a partition of space into convex nonovlapping polyhedra that completely fill the space. Vorontessellations originally emerged in the context ofnumber-theoretical problem12,58 and have since then beesuccessfully applied as a model in such diverse fieldsscience as physics~liquid structure,37,43 crystal growth!,35
biology ~cellular patterns!,20 and medicine ~capillarydomains!.21 In particular, Kurz and Sandau32 have pro-posed the assessment of the end-point distribution aimportant characteristic of vascular trees and usVoronoi tessellations to describe the apparent end-pdistribution of arteries in the chorioallantoic membraof chicken eggs.
Here, we are interested in various topological ametric properties of the Voronoi polyhedra~VP! associ-

ofby
e-a-eIndesP.to
inalForataoodmet aetes
iono-
nstheults
ale-ctlO
gle
ootentricof
ti-di-od-fail-thethe
eg-ow-wth
intsdeleof
in-hy-sti-
et-
-
td sona-
te
rmi-he
549Voronoi Analysis of Arterial Tree Models
ated with the distal end points of terminal segmentsCCO trees. Since Voronoi tessellations generatedpurely randomly distributed points in two- and thredimensional space~so-called Poisson–Voronoi tesselltions! have been extensively studied in thliterature,19,30,31,35we use this model as a reference.particular, we address the question how different moof tree optimization affect the shape of the resulting VAs an application, the Voronoi tessellations are usedassign the region of supplied tissue to each termsegment in a quite natural and unambiguous way.comparison of model predictions with experimental dwe consider scaling relations between intravascular blvolume and arterial cross-sectional area with the voluof supplied tissue. In a further application, we presenmodification of the algorithm of CCO, namely to sterminal flows proportional to the associated VP volumduring tree generation.
This paper is organized as follows: In the next sectwe briefly summarize the method of CCO and the algrithm used for the construction of Voronoi tessellatioand describe the quantities calculated from the VP. Infollowing sections we present and discuss the resobtained and summarize the conclusions drawn.
METHODS
The Algorithm of Tree Generation
CCO trees, originally introduced in a two-dimensionframework,48,49 constitute some kind of synthesis btween rule-based fractal approaches and models direderived from the anatomy of real arterial trees: CCemploys the principle of optimal design,45 which haslong been hypothesized in the literature to hold for sinbifurcations of arterial trees.36,62 In an ‘‘ab initio’’ likefashion, CCO trees are constructed from a single rsegment by subsequently adding new terminal segmfrom randomly chosen positions within the geometmodel of the perfusion area, until a given numberterminal segmentsNterm is reached~Fig. 1!. At each stepof growth, the geometry of the new bifurcation is opmized under prescribed physiological boundary contions. This procedure allows to generate computer mels of arterial trees with an ‘‘arbitrary’’ number osegments, limited only by the computer resources avable. One of the main advantages of CCO trees isfact that at each step of construction we have at handcomplete geometric data set of the tree, including sment locations and diameters. It should be noted, hever, that no attempt is made to model the actual groprocess of real arterial trees.22
For ease of reference, we summarize the main poof the method of CCO:25,49 Arterial trees are representeas binary branching trees, vessel segments are modby rigid cylindrical tubes, and the precise geometry
y
s
d
bifurcations is not considered. Blood is assumed ancompressible, homogeneous Newtonian fluid and thedrodynamic resistance of individual segments is emated by means of Poiseuille’s law.13 Optimization ofsingle bifurcations is performed under the following sof boundary conditions: At each bifurcation, the diameters of the parent segment (d0) and the doughter segments (d1 ,d2) obey a power law of the form
d0g5d1
g1d2g ~1!
with a constant exponentg.0. At a given total perfusionflow Qperf, individual terminal flows Qterm,i with(Qterm,i5Qperf, and at a given diameter of the roosegment, all remaining segment diameters are scaleas to fulfill Eq. ~1! at each bifurcation and to maintaithe prescribed flows. The position of each new bifurction is calculated so as to minimize the quantity
T5(i
l idil , ~2!
where l i is the length of segmenti and l is a freeparameter:l52 minimizes total intravascular volume,24
whereasl50 minimizes total segment length.As of particular interest for the present work, we no
that the coordinates of new terminal positionsxt aredrawn from a random number sequence with a unifodistribution. In particular,xt is accepted as a new termnal site only if it is not too close to the segments of t
FIGURE 1. Schematic representation of the growth of a CCOtree. A new terminal positon x t is selected at random withina circular perfusion area and connected to an existing treesegment, thus forming a new terminal segment and a newbifurcation. In the next step, the geometry of the new bifur-cation is optimized with respect to a given target function T,Eq. „2…, and prescribed boundary conditions for pressuresand flows.

aof
re-p-ndthe
nalpuof
-m
of
se
ted
isdi-l oa
on-
theo-e-t
ed
sxi-theuau-er-ce
ly
nts,
er-
n-n
nd--
P.
eoi
the
hethe-
into
550 KARCH et al.
tree grown thus far. This ‘‘repulsion criterion’’ acts likestep potential of infinite height in the neighborhoodeach segment: Terminal positionsxt within the potentialwell are rejected, all others remain unaffected. As asult, the terminal positions of CCO trees no longer reresent a sample from a uniform distribution in space, ait is one goal of the present paper to characterizedifferences.
The Construction of Voronoi Polyhedra
The efficient construction of the Voronoi tessellatioof a set of points, in particular in three-dimensionspace, is a nontrivial task and much effort has beeninto the development of such algorithms in the fieldcomputational geometry.3,39,40 In this paper, we haveused the Qhull software package,4 which implements theQuickhull algorithm40 to construct the Voronoi tessellation of a given set of input points. The Qhull prograutilizes the following relation between convex hulls~i.e.,the smallest convex set that contains a given setpoints! and Voronoi tessellations:5,9 To obtain for a givenset of points inR2 the Delaunay triangulation~i.e., thestraight-line dual of the Voronoi tessellation of thepoints!,40 the points are lifted to a paraboloid inR3 andthe convex hull of these transformed points is compuin R3; the set of edges of the lower convex hull~i.e., thatpart of the hull which is visible from thexy plane!3 isthe Delaunay triangulation of the original points. Thalgorithm can immediately be generalized to highermensions. Since the Delaunay tessellation is the duathe Voronoi tessellation, the problem of constructingm-dimensional Voronoi diagram is thus reduced to a cvex hull problem in (m11) dimensions.
The data structure produced by Qhull includesvertex coordinates of the Voronoi polygon-edges in twdimensional ~2D! space and polyhedra-faces in thredimensional~3D! space for all pairs of adjacent inpusites; from this information we readily obtain the desirtopological~e.g., number of faces! and metric~e.g., vol-ume, surface area! properties of the VP. Since Qhull usefloating point arithmetic, the results are only appromate. As a necessary condition for the consistency oftessellations, the sum of the VP volumes must be eqto the total volume of the system. We therefore calclated the respective sum and compared it with the pfusion volume of the tree. We always found a differennot larger than approximately 1027%.
Calculated Quantities
The topology of a Voronoi polyhedron is completecharacterized by the number of verticesNv , the numberof edgesNe , and the number of facesNf . These threeparameters are related by two topological constrainamely the classical Euler formula40
t
f
l
Nv2Ne1Nf52 ~3!
and
3Nv52Ne . ~4!
The last equation follows, if we assume that the genating points are in general position,39 so that every vertexshares exactly three faces~and thus edges! and everyedge connects exactly two vertices.23,46 ~This assumptionis reasonable for CCO trees, since it is extremely ulikely for random systems that four or more points lie othe same plane and five or more points lie on the bouary of a sphere.! Hence, only one of the previous parameters is independent. Here, we have usedNf as the in-dependent quantity to describe the topology of the VFor the remaining two parameters we then haveNv52Nf24 andNe53Nf26.
In addition to the topological quantities, we havevaluated the following metric quantities of the Voronpolyhedra: The areaAk of the individual faces
Ak51
2 (j 52
n21v
~k)
u~r j(k)2r1
(k)!3~r j 11(k) 2r1
(k)!u, ~5!
wherenv(k) is the number of vertices of thekth face and
r j(k) is the position vector of thej th vertex of this face;
the total surface areaS:
S5 (k51
Nf
Ak , ~6!
and the total volumeV:
V51
3 (k51
Nf
hkAk , ~7!
wherehk denotes the normal distance of the center ofVoronoi polyhedron from itskth face. By definition of aVoronoi tessellation, each face of a cell is part of tperpendicular bisector of the line segment betweencell center and one of itsNf nearest neighbors. Therefore, thekth face exactly divides the distancer k betweenthe center and the corresponding nearest neighbortwo equal parts, and we havehk5r k /2.
Following Ruoccoet al.,46 we have also calculatedthe asphericity parametera:
a5S3
36pV2 , ~8!

he-
e o
s oinan-
Theideeseerie00l-een
seisis
he
ofin
.,
thels.
ruclatetalwetheus
ichure
cov-the
at
551Voronoi Analysis of Arterial Tree Models
which measures the deviation of the shape of a polydron from that of a perfect sphere. By definition,aequals 1 for a sphere, and larger values ofa indicate amore aspheric polyhedron, e.g.,a51.91 for a cube.
RESULTS
Generation of Model Trees
We have generated ten model trees for each valuthe optimization parameterl50 and l52. The modelrealizations were obtained by means of different seedthe random number sequences for tossing new termlocations; all remaining model parameters were left uchanged for the set of ten trees within each series.trees were generated in a cubic slab of 4.642 cm slength. Simulation parameters are listed in Table 1. Thparameters were chosen so as to model the large artof the coronary arterial tree supplying approximately 1g of myocardial tissue.26 Figure 2 shows a specific reaization for each of the generated models. As can be strees optimized for minimum segment length (l50) ex-hibit a strongly meandering structure. Although the cal50 is not exceedingly realistic, we have included thmodel in order to study if and how such a structurereflected in the topological and metric properties of tcorresponding VP.
As a reference model for the spatial arrangementterminal end-points in CCO trees, we have generatedthe same geometry~cubic slab! ten samples of purelyrandom points drawn from a uniform distribution, i.erealizations of a so-called Poisson point process.39 ThenumberN of random points was chosen the same asnumberNterm of terminals in the respective tree mode
Construction of Voronoi Polyhedra
In the present paper we are concerned with tree sttures grown in a bounded region of space. To calcuthe Voronoi tessellations of such regions with the disend points of terminal segments as generator pointsmirror these points along the boundaries and performVoronoi decomposition on the extended point set thobtained. Finally, we keep only those polyhedra whare associated with the original point set. This proced
TABLE 1. Simulation parameters.
Parameter Meaning Value Reference
pperf Perfusion pressure 100 mm Hg 10Qperf Perfusion flow 500 ml/min 11h Viscosity of blood 3.6 cp 34rroot Radius of the root segment 2 mm 27g Bifurcation exponent 2.55 2Nterm Number of terminals 6000Vperf Volume of perfusion area 100 cm3
f
fl
s
,
-
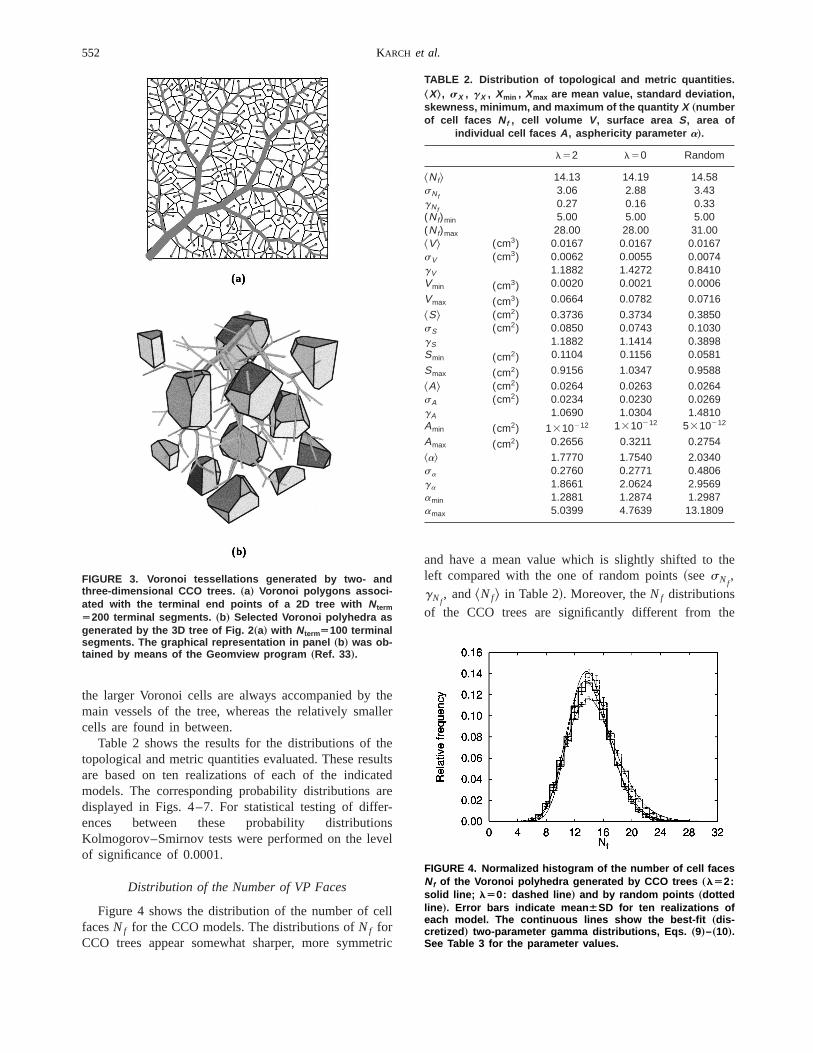
guarantees that the region boundaries are exactly reered as parts of the calculated VP. Figure 3 illustratesresulting tessellations for a 2D@panel~a!# and 3D@panel~b!# CCO tree withNterm5200 andNterm5100 terminalsegments, respectively. In Fig. 3~a! we immediatly rec-ognize a distinctive feature of CCO trees, namely th
FIGURE 2. Visual representation of CCO trees optimized forminimum intravascular volume „a… and minimum total seg-ment length „b…. Simulation parameters are given in Table 1.Visualization was performed by representing the vessel seg-ments as the isosurface of a pseudopotential assigned to thewhole tree „see Neumann et al. , Ref. 38….
theller
heultstedrer-nsvel
ell
etri
he
e
552 KARCH et al.
the larger Voronoi cells are always accompanied bymain vessels of the tree, whereas the relatively smacells are found in between.
Table 2 shows the results for the distributions of ttopological and metric quantities evaluated. These resare based on ten realizations of each of the indicamodels. The corresponding probability distributions adisplayed in Figs. 4–7. For statistical testing of diffeences between these probability distributioKolmogorov–Smirnov tests were performed on the leof significance of 0.0001.
Distribution of the Number of VP Faces
Figure 4 shows the distribution of the number of cfacesNf for the CCO models. The distributions ofNf forCCO trees appear somewhat sharper, more symm
FIGURE 3. Voronoi tessellations generated by two- andthree-dimensional CCO trees. „a… Voronoi polygons associ-ated with the terminal end points of a 2D tree with NtermÄ200 terminal segments. „b… Selected Voronoi polyhedra asgenerated by the 3D tree of Fig. 2 „a… with NtermÄ100 terminalsegments. The graphical representation in panel „b… was ob-tained by means of the Geomview program „Ref. 33….
c
and have a mean value which is slightly shifted to tleft compared with the one of random points~seesNf
,gNf
, and^Nf& in Table 2!. Moreover, theNf distributionsof the CCO trees are significantly different from th
TABLE 2. Distribution of topological and metric quantities.ŠX‹, sX , gX , Xmin , Xmax are mean value, standard deviation,skewness, minimum, and maximum of the quantity X „numberof cell faces Nf , cell volume V, surface area S, area of
individual cell faces A , asphericity parameter a….
l52 l50 Random
^Nf& 14.13 14.19 14.58sNf
3.06 2.88 3.43gNf
0.27 0.16 0.33(Nf)min 5.00 5.00 5.00(Nf)max 28.00 28.00 31.00^V& (cm3) 0.0167 0.0167 0.0167sV (cm3) 0.0062 0.0055 0.0074gV 1.1882 1.4272 0.8410Vmin (cm3) 0.0020 0.0021 0.0006
Vmax (cm3) 0.0664 0.0782 0.0716
^S& (cm2) 0.3736 0.3734 0.3850sS (cm2) 0.0850 0.0743 0.1030gS 1.1882 1.1414 0.3898Smin (cm2) 0.1104 0.1156 0.0581
Smax (cm2) 0.9156 1.0347 0.9588
^A& (cm2) 0.0264 0.0263 0.0264sA (cm2) 0.0234 0.0230 0.0269gA 1.0690 1.0304 1.4810Amin (cm2) 1310212 1310212 5310212
Amax (cm2) 0.2656 0.3211 0.2754
^a& 1.7770 1.7540 2.0340sa 0.2760 0.2771 0.4806ga 1.8661 2.0624 2.9569amin 1.2881 1.2874 1.2987amax 5.0399 4.7639 13.1809
FIGURE 4. Normalized histogram of the number of cell facesNf of the Voronoi polyhedra generated by CCO trees „lÄ2:solid line; lÄ0: dashed line … and by random points „dottedline …. Error bars indicate mean ÁSD for ten realizations ofeach model. The continuous lines show the best-fit „dis-cretized … two-parameter gamma distributions, Eqs. „9…–„10….See Table 3 for the parameter values.

t,ce
ter
D
the
he
maesnd-
s aot
553Voronoi Analysis of Arterial Tree Models
random points~two-sample Kolmogorov–Smirnov tesp,0.0001), whereas there is no significant differenbetween the tree models withl52 and l50 at the0.0001 level of statistical significance.
Kumar et al.30 suggested a discretized two-paramegamma distribution
f ~Nf !5ENf21/2
Nf11/2
p~x!dx ~9!
to approximate the distribution of cell faces in a 3Poisson–Voronoi tessellation, where
p~x!5xa21
baG~a!expS 2
x
bD , x.0, ~10!
FIGURE 5. „a… Probability density distribution „meanÁSD… ofthe normalized cell volume V*ÄVÕŠV‹ of the Voronoi polyhe-dra generated by CCO trees „lÄ2: solid circles; lÄ0: opencircles … and by random points „open squares …. The continu-ous lines show the corresponding best-fit two-parametergamma distributions „lÄ2: solid line; lÄ0: dashed line;random points: dotted line …, Eq. „10…. See Table 3 for theparameter values. „b… Distribution of normalized cell volumesV* of CCO trees relative to the result p „Vrnd* … for randompoints.
is the probability density function of the~continuous!two-parameter gamma distribution.G(a) denotes thegamma function and the mean and variance ofdistribution in Eq. ~10! are given by ab and ab2,respectively.
We have fitted the above distribution, Eq.~9!, to ourdata, both for random points and for CCO models. Tresults for the best-fit parametersa and b together withthe mean and standard deviation of the fitted gamdistribution are listed in Table 3. Comparing the valufor the mean and standard deviation with the correspoing results of the original data~Table 2!, we concludethat a two-parameter gamma distribution representgood approximation to the distribution of cell faces, n
FIGURE 6. Probability density distribution „meanÁSD… of thenormalized total surface area S*ÄSÕŠS‹ of the Voronoi poly-hedra generated by CCO trees „lÄ2: solid circles; lÄ0:open circles … and by random points „open squares …. Thecontinuous lines show the corresponding best-fit two-parameter gamma distributions „lÄ2: solid line; lÄ0:dashed line; random points: dotted line …, Eq. „10….
FIGURE 7. Probability density distribution „meanÁSD… of theasphericity parameter a, Eq. „8…. Solid circles: CCO treeswith lÄ2. Open circles: CCO trees with lÄ0. Open squares:Poisson point process.

e-larO
elle-onith
ees
andt, so
isu-
d inon–
u-
-fted
theOini-f
eststhee
the
. Intreeol-ar-
for(ar
-nheveerist-esa
fer-re-
tal-
r
m
ecee
of
554 KARCH et al.
only for random points, but also for CCO trees~see alsoFig. 4!. Moreover, the moments of theNf distribution~Table 2! reflect the differences in the spatial arrangment of the corresponding generating points, in particubetween random points and terminal locations in CCtrees.
Distribution of Cell Volumes
Figure 5~a! shows the distributions of the reduced cvolumes,V* 5V/^V&, for the CCO models studied. Thcell volume distributionsp(V* ) of the tree models appear markedly different from the respective distributiof the Poisson–Voronoi tessellation, in particular wrespect to their spread around the mean~seesV in Table2!: tree models are more sharply peaked around^V* &51, and this behavior is even more pronounced for trwith l50 than for threes withl52. ~Note that for themean value we haveV* &51 for each model. This isbecause we use a simulation box of fixed geometrywe keep the number of the generating points constanthat the mean cell volume always takes the value^V&5Vbox/N, and therefore^V* &51, independent of thespecific model.! The p(V* ) distributions are significantlydifferent between CCO models withl52 andl50 andbetween CCO models and random points~Kolmogorov–Smirnov test,p,0.0001).
The cell volume distribution of the random pointswell approximated by a two-parameter gamma distribtion, Eq. ~10!.30 The best-fit parameter values fora andb are listed in Table 3. These values are consistent anreasonable agreement with previous studies for PoissVoronoi tessellations.30 Deviations ina andb are due toboundary effects. Fitting two-parameter gamma distribtions to thep(V* ) values of the tree models~see Table
TABLE 3. Parameters of best-fit two-parameter gammadistributions. Entries for each variable „number of faces Nf ,normalized volume V*ÄVÕŠV‹ and surface area S*ÄSÕŠS‹…
denote the estimated parameter values for a and b „includingthe 68.3% confidence interval … as well as the mean ab andstandard deviation Aab 2 of the respective best-fit gamma
distribution, see Eq. „10….
l52 l50 Random
Nf 21.4360.51 24.8960.82 17.4660.350.6660.02 0.5760.02 0.8460.0214.17 14.24 14.713.06 2.85 3.52
V* 8.448960.2998 10.824660.3618 5.106260.04850.116960.0043 0.091660.0031 0.196060.00180.9876 0.9914 1.00070.3398 0.3013 0.4429
S* 21.874360.8567 30.081961.0022 13.932160.27370.045460.0018 0.032960.0011 0.074160.00140.9927 0.9907 1.03190.2123 0.1806 0.2765
3 for the estimated parameters! also yields a good approximation for these data, in particular along the leand the right tails; small differences can be observonly near the peaks of the distributions, see Fig. 5~a!.
To further evaluate the features ofp(V* ) for CCOtrees, we have plotted the differences betweenp(V* ) ofthe tree models andp(Vrnd* ) of the correspondingPoisson–Voronoi tessellation, see Fig. 5~b!. We observepronounced peaks in these distributions, illustratingdifferent environment of the generating points in CCtrees and in a purely random arrangement: The first mmum nearV50.5 V& illustrates a reduced number osmaller cells present in tree models. This result suggsome kind of repulsive effect of the tree structure onspatial distribution of terminal end points. Likewise, thsecond minimum nearV51.5 V& shows a reduction inthe number of larger cells in CCO trees, whereasnumber of cells near the mean cell volumeV5^V& isincreased at the expense of smaller and larger cellssummary, these results indicate a tendency of themodels to more homogeneously distribute supply vumes than could be achieved by a purely randomrangement of terminal locations. Moreover, Fig. 5~b!shows that the above effects are more pronouncedCCO trees optimized for minimum segment lengthl50) than for trees optimized for minimum intravasculvolume (l52).
It is interesting to note that Montoroet al.17 observeda similar behavior of the Voronoi cell volume distribution for certain physical conditions in their simulatiostudy on ionic association in electrolyte solutions. Tauthors attributed their VP distributions to the repulsiaction of Coulomb forces in the system. This furthsupports our notion that the repulsive effect of an exing tree on the location of new terminal sites producdifferences in their spatial distribution relative topurely random arrangement of points, and these difences are directly reflected in the properties of thespective Voronoi polyhedra.
Distribution of the Total Surface Area of the VP
Figure 6 shows the distribution of the normalized tosurface area,S* 5S/^S&, of the Voronoi cells. Each distribution p(S* ) is fairly symmetric around its meanvalue. Moreover,p(S* ) appears distinctly narrower fothe tree models than for random points~see also thevalues ofsS in Table 2!. This narrowing effect inp(S* )is more pronounced for trees optimized for minimulength (l50) than for trees optimized for minimumvolume (l52). In comparison to random points, thVoronoi cells of the tree models having a total surfaarea near the mean value^S& are favored at the expensof cells with relatively smaller and largerS. This behav-ior of p(S* ) is consistent with the above observations

w-i-
v–
rethes.
eaarsns
m-
er,
oisectdis-
ar-ery
dels
e
by
ingof
d
of
t in aied.
t
etheirs.
555Voronoi Analysis of Arterial Tree Models
p(V* ) concerning a certain balancing effect of the groing tree structure on the spatial distribution of its termnal supply points. Based on a two-sample KolmogoroSmirnov test, the differences between thep(S* )distributions are statistically significant (p,0.0001),both between CCO models withl52 and l50 andbetween CCO models and random points.
Again, a two-parameter gamma distribution, Eq.~10!,represents a good fit to the reference distributionp(S* )of the corresponding Poisson–Voronoi tessellation.30 Fit-ting a gamma distribution to thep(S* ) data of the CCOtrees, also yields a fairly good approximation. Yet, theare noticeable differences between the fitted andoriginal distributions, in particular near their peak valueThe best-fit parametersa and b are given in Table 3.
Distribution of the Asphericity Parameter
The parameter of asphericitya, Eq. ~8!, combinesvolume and surface area into a single descriptor to msure how different the shape of a polyhedron appefrom that of a sphere. Figure 7 shows the distributiop(a) for the models studied: Markedly different fromthe Poisson point process, which exhibits a long asymetric tail and has a mean value neara'2, the distri-butions of the CCO models are considerably narrowmore symmetric, and their mean value ata'1.7 liesclose to that of a sphere (a51). The VP of CCO treesare thus more regular than the ones generated by a Pson point process, again confirming the balancing effalready mentioned for VP volume and surface areatributions. In contrast to that, the differences in thep(a)distributions between the CCO models are only mginal: Both mean values and standard deviations are vclose, only the tree forl50 is slightly more symmetric.However, these differences inp(a) are statistically sig-nificant, as are the differences between the CCO moand the random points~Kolmogorov–Smirnov test,p,0.0001).
DISCUSSION
Distribution of Cell Volumes
The volume of a Voronoi cell is determined by thareasAk of its faces, by the distancesr k of its centerfrom the corrresponding nearest neighbor points, andthe numberNf of its faces @cf. Eq. ~7!#. Hence, theproperties ofAk , r k , and Nf in CCO models and in aPoisson point process provide further insight concernthe observed differences in the respective distributionscell volumes.
Figure 8~a! shows the PDFsdk(r ) of the kth nearestneighbor distances fork51,2,3 for the CCO models an
-
-
for a Poisson point process with the same densitypointsr5N/Vbox. For the latter,dk(r ) can be written inclosed form~see also Ref. 52!:
dkP~r !5r4pr 2
~rbr !k21
~k21!!exp~2rbr !, ~11!
wherebr5(4p/3)r 3. Figure 8~a! illustrates that the firstthree nearest neighbor distances are always abundanPoisson point process relative to the CCO trees studRecalling that the mean number of cell facesNf ~i.e., thenumber of nearest neighbors for each generating poin! isabout 15, with maximum values ranging up to 31~seeTable 2!, it is by far not sufficient to consider only thfirst three nearest neighbor distances to characterizecontribution to the observed cell volume distribution
FIGURE 8. „a… Probability density distribution „meanÁSD… ofthe k th nearest neighbor distances for kÄ1 „left curves …, kÄ2 „middle curves …, and kÄ3 „right curves … for ten realiza-tions of CCO trees with lÄ2 „solid circles with solid lines …
and lÄ0 „open circles with dashed lines …. The dotted linesshow the theoretical curves for a Poisson point process withthe same density r, see Eq. „11…. „b… Radial pair distributionfunctions g „r … for the CCO models „symbols as earlier … andfor the corresponding Poisson point process with g „r …Æ1.

ighhe
of
sity
teeairns
of
of-
totheewyra-ntst oas
COy isst
beur-
the
fo
556 KARCH et al.
Since it is very cumbersome to analyze all nearest nebor distances in the way given earlier, we apply twell-known concept of the~radial! pair distribution func-tion g(r ).1 This function measures the average portionpairs of points having a distancer from each other rela-tive to a purely random arrangement at the same denr. For spatially homogeneous and isotropic systemsg(r )can—without loss of information—be written as
g~r !51
r
1
4pr 2
1
N (i
(j Þ i
d~r 2r i j !, ~12!
where r i j denotes the separation between pointi andpoint j and d(r 2r i j ) is the Dirac delta function. Notethat in Eq.~12! any features ofg(r ) arising from spatialinhomogeneities or anisotropies are averaged out.g(r ) isrelated to the distributionsdk(r ) of thekth nearest neigh-bor distances by
g~r !51
r
1
4pr 2 (k
dk~r !. ~13!
By definition, and by means of Eqs.~11! and ~13!, wehave g(r )[1 for a Poisson point process. To evaluag(r ) for the CCO models, we follow the usual practicand construct a histogram of the respective pseparations.1 The results, averaged over ten realizatioof each CCO model, are shown in Fig. 8~b!. We firstnotice a distinctive maximum in the relative frequencypair separations aroundr 50.23 cm. This is a broadpeak, indicating the contribution of many ordersneighbor separations, see Eq.~13!. The greater abundance of such pair separations in CCO trees relativerandom points is caused by the larger segments oftrees: since the interior of segments is excluded for nterminal positions, large~i.e., thick! segments necessarilgive rise to a relatively higher frequency of pair sepations near or beyond the diameter of these segmeThis behavior is best recognized from the arrangemenlarge tree segments and neighboring Voronoi cellsshown in the 2D example of Fig. 3~a!.
The low values ofg(r ) for small r illustrate a pro-nounced deficiency of these pair separations in Ctrees relative to a Poisson point process. This decamainly due to the distribution of the first three neareneighbor distances, see Fig. 8~a!, and originates from therepulsion criterion, which suppresses small distancestween new terminal locations and existing segments ding tree generation.
The second important parameter characterizingdistribution of cell volumes is the areaA of individualVP faces. Figure 9 shows the distribution ofA groupedby selected classes of nearest neighbor separations
-
.f
-
r
FIGURE 9. Probability density distribution „meanÁSD… of thearea A of individual polyhedra faces grouped by selectedclasses k of ascending nearest neighbor separations forwhich the respective faces form the perpendicular bisectors.Solid circles with solid lines: CCO trees with lÄ2. Opencircles with dashed lines: CCO trees with lÄ0. Opensquares with dotted lines: Poisson point process.

ec-n-
rest
inrs,intsntlyeig-
ssoisnecy.-v
of
tri-
v-
r
for
of
ed
m
t
inothtedo-nO
of
e-
cectsIn
we
d-
ofblenalthe
nceersentth.
557Voronoi Analysis of Arterial Tree Models
which the respective faces form the perpendicular bistors: For smallA the Poisson point process always cotributes with a relatively larger number of faces toV,whereas for midrangedA the situation is reversed. FolargeA, only the face areas pertaining to the first nearneighbors are noticeably abundant for random pointscomparison with CCO trees; for higher-order neighbothe differences between CCO trees and random poare almost negligible. These properties are consistereflected in the overall distribution of individual facareasA, irrespective of the nearest neighbor index: Fure 10 shows the results for the distribution ofA*5A/^A&. The smallest face areas and—though lepronounced—also the largest areas are favored in a Pson point process, whereas in CCO trees face areasthe mean value appear with a relatively higher frequenThis difference inp(A* ) between CCO trees and random points is confirmed by a Kolmogorov–Smirnotest (p,0.0001), whereas CCO models withl52 andl50 are not discriminated at the 0.0001 levelsignificance.
As to the third parameter which determines the disbution of V, namely the number of facesNf per poly-hedron, we first note thatV and Nf are positively corre-lated: The correlation coefficient betweenNf and V isrNf ,V50.74 for CCO models withl52, rNf ,V50.70 forCCO models withl50, and rNf ,V50.65 for randompoints. Figure 4 illustrates that the frequency of VP haing a large number of facesNf.16 ~and therefore, alarge cell volume! is significantly higher for randompoints than for CCO trees. On the contrary, VP withNf
near the meanNf& are found more frequently in CCOtrees. For smallNf , all models behave fairly similaconcerning the frequency ofNf .
In summary, the abundance of small cell volumes
FIGURE 10. Probability density distribution „meanÁSD… ofthe normalized area A *ÄA ÕŠA ‹ of individual polyhedrafaces. Solid circles with solid lines: CCO trees with lÄ2.Open circles with dashed lines: CCO trees with lÄ0. Opensquares with dotted lines: Poisson point process.
-ar
random points is due to a relatively higher frequencysmall pair distances@g(r ), see Fig. 8~b!# as well as ofsmall individual facet areasA ~Fig. 10! in these models.For intermediate cell volumes, the situation is reverswith an additional effect ofNf in favor of CCO trees.The higher frequency of larger cell volumes for randopoints is mainly due toA ~Fig. 10! and to Nf ~Fig. 4!.From g(r ) and the distributions ofA grouped by nearesneighbors ~Fig. 9! it is evident, that only low-orderneighbors significantly contribute to the differencesthe cell volume distributions. These observations—bthe distribution of nearest neighbor distances as reflecin g(r ) as well as the one of individual face areas tgether with Nf—consistently explain the differences ithe distributions of Voronoi cell volumes between CCtrees and random points as displayed in Fig. 5.
Dependence of Cell Volume Statistics on the NumberEnd Points
For ideal ~infinite! Poisson–Voronoi tessellations, thdistribution p(V* ) of reduced cell volumes is independent of the point densityr due to the invariance of thegenerating Poisson point process under scaling.19 How-ever, Voronoi tessellations in a finite region of spabecome more and more dominated by boundary effeas the densityr of the generating points decreases.particular, for CCO trees it isa priori unclear, how cellvolume statistics vary as a function ofr5Nterm/Vbox andif there exists a ‘‘limiting’’ distribution of cell volumesfor large Nterm. Figure 11~a! shows thep(V* ) distribu-tions for CCO models withl52 and with differentnumbers of end points ranging fromNterm5600 toNterm532,000. There is a modest effect ofNterm on therespectivep(V* ) distributions, in particular forNterm
,6000. The relative dispersions RDV5sV /^V& tend todecrease asNterm increases with values between RDV
50.42 for Nterm5600 to RDV50.36 for Nterm532,000.This dependence onNterm is mainly due to boundaryeffects: excluding boundary cells from the analysis,obtain p(V* ) distributions as illustrated in Fig. 11~b!.Now the variability between the distributions is consierably reduced and we find values for RDV in the rangeRDV50.3460.01. In summary, the chosen valueNterm56000 for the model trees proves a reasonacompromise between the requirements for computatioresources during tree generation and the validity ofreported moments ofp(V* ) in the limit of largeNterm.These results suggest a certain kind of scaling invariaor self-similarity of CCO trees in the sense that evfiner tree structures reproduce properties already preat coarser levels of resolution in earlier stages of grow

ieda
inal
m-e
dio--d-
r adta
n-tionthea-of
uethe
b-
t,
l-therspo-is
a
talhe
558 KARCH et al.
Comparison with Experimental Data
Scaling of Blood Volume with the Volume of SupplTissue.Scaling relations in biology have long beensubject of discussion since their advent in the semwork of D’Arcy Thompson.54 Only recently, Westet al.60
have presented a general model for the origin of alloetric scaling laws in biology, i.e., relationships of thform
Y5Y0Xs, ~14!
where Y and X are some biological observables~e.g.,metabolic rate and mass of an organism!, Y0 is a nor-
FIGURE 11. The effect of the number Nterm of end points onthe cell volume distribution. „a… Probability density distribu-tion of the normalized cell volume V*ÄVÕŠV‹ of the Voronoipolyhedra generated by CCO tree models „lÄ2… for variousvalues of Nterm „triangles: NtermÄ600, squares: NtermÄ2000,solid circles: NtermÄ6000, open circles: NtermÄ18,000, dia-monds: NtermÄ32,000…. The symbols indicate mean valuesfor 10 „NtermÄ600, 2000, 6000… and 2 „NtermÄ18,000, 32,000…model realizations, respectively. The dotted lines „solid linefor NtermÄ6000… show the corresponding best-fit two-parameter gamma distributions, Eq. „10…. „b… Same as „a…,boundary cells excluded from the analysis.
malization constant, ands the scaling exponent. Baseon a few basic principles concerning the design of blogical transport networks~such as a hierarchical branching structure, invariant terminal units, and minimizeenergy dissipation!, this model predicts many scaling relations of mammalian circulatory systems, in particulalinear relationship~i.e., s51) between blood volume antotal body volume in agreement with experimental dareported by Prothero.41
To evaluate this relation for the CCO models we cosider the intravascular volume of subtrees as a funcof the tissue volume supplied by each subtree. SinceVoronoi tessellation induced by the tree’s terminal loctions provides a convenient and unambiguous wayassigning to each terminal location a region of tisssupplied with blood, we consider—for each subtree—total volume of the Voronoi polyhedraVVP correspond-ing to the terminal supply points of the respective sutree. The results for the more realistic models withl52 are shown in Fig. 12 in a double-logarithmic plowhere the maximum value ofVVP represents the wholeperfusion volume. Single terminal segments~i.e., degen-erated subtrees! and corresponding single polyhedra voumes have been excluded from the analysis. Forremaining data points, which still range over four ordeof magnitude, a best-fit regression yields a scaling exnent of s51.2060.007. The obvious departure of thresult from the value ofs51 predicted by Westet al. isnot too surprising, given the fact that we have usedconstant bifurcation exponentg52.55 throughout thetree and an optimization target which minimizes tointravascular volume. Both settings are different from t
FIGURE 12. Scaling of blood volume Vb with the volume VVPof supplied tissue for ten realizations of CCO trees with lÄ2 in a double-logarithmic representation. y axis: total intra-vascular volume of subtrees, excluding single terminal seg-ments. x axis: total volume of Voronoi polyhedra generatedby corresponding distal end points of terminal segments.The solid line is the best-fit regression to the data points.

hisortul-
-
nd,
d-
theode
heon-ndye oalver,of
ricryO
heycec-exttricrea
notre-
aveloiedord
ml
al:e
ed
e-
re-
Inels
oly-re-
eri-ri-
o-ta
mthe
are
ameurell
rees
uceod
559Voronoi Analysis of Arterial Tree Models
assumptions and results of the model cited earlier: Tmodel minimizes the energy dissipation of the transpnetwork and predicts—for the more realistic case of psatile flow—a bifurcation exponentg which varies fromg52 for larger arteries tog53 for smaller arteries. Wehave also evaluated the scaling exponents for the caseg53. ~Minimizing total intravascular volume is equivalent to energy minimization only forg53.)61 Note thatincreasingg in Eq. ~1! slows down the rate at whichsegmental volumes drop along bifurcation orders ahence, has the effect of decreasings. Accordingly, weobtain s51.1660.004, a value closer to the reportevalue of s51. This result further improves if we consider only larger subtrees: ForVVP.0.1 cm3 the value ofthe scaling exponent decreases tos51.0860.005, sug-gesting that CCO trees quite satisfactorily reproducepredicted and experimentally observed scaling of blovolume with the volume of tissue supplied, provided wdo not take too close a look at all the details of tsmallest subunits of the model. This observation is csistent with recent results reported by Wang aBassingthwaighte,59 who showed that capillary supplregions, i.e., regions based on the functional measurcapillary diffusion, cannot—apart from very specicases—be approximated by Voronoi polygons; moreoappropriate modeling of transport and consumptionoxygen in ~cardiac! tissue demands a realistic geometmodel for the three-dimensional nontree-like capillanetwork.8 On the other hand, terminal segments of CCtrees do not represent real terminal arterioles, nor do tmodel capillaries, and the subdivision of tissue spainto VP induced by the terminals is not based on funtional considerations, but should in the present contbe understood as an attempt—on purely geomegrounds—to assign to the model’s terminal segmentssonable areas of blood supply.
Finally, we note that the volume of the vessels canbe considered part of the terminal supply regions repsented by the associated VP. Therefore, we hestimated—by a straight-forward Monte Carsimulation—the portions of polyhedra volumes occupby tree segments and corrected these volumes accingly. For the models withl52 and Nterm56000 theaverage intravascular volume is approximately 1.5~i.e., only 1.5% of the total perfusion volume!. Conse-quently, the effects of tree volume are only marginNeither the distribution of Voronoi cell volumes nor thscaling of blood volume with~now corrected! supplyvolume are noticeably different from the uncorrectcase.
Scaling of Arterial Cross-Sectional Area with the Volumof Supplied Tissue.Seileret al.51 have reported a power
f
-
-
law relationship between luminal cross-sectional areaaof human coronary artery segments and perfusedgional myocardial mass
a}VVPb , ~15!
with exponentsb in the range between 0.62 and 0.82.good agreement with these results we find for the modwith l52 a scaling exponent ofb50.7760.01 ~Fig.13!, where terminal segments and associated single phedra volumes have again been excluded from thegression analysis.
Voronoi Cell Area Distribution of Arterial End Points inthe Human Retina.Two-dimensional CCO models arconvenient for a comparison of Voronoi cell area distbutions with physical data. Here, we attempt a compason with the major arteries of the approximately twdimensional layer of the human retina. From dapublished by Zamiret al.,63 we digitized just visible ar-terial end points within a square region of about 5 msidelength centered around the macular area, usingoriginal fundus photograph~their Fig. 1! and a map ofmajor retinal vessels~their Fig. 2!. The resulting set ofapparent end points and its Voronoi tessellationshown in Fig. 14~a!. In order to provide comparablemodel data, we have pruned the model trees to the snumber of terminal segments as points digitized. Fig14~b! shows the distributions of normalized Voronoi ceareas of the experimental data and pruned 2D CCO twith original Nterm53000 and l52. In both cases,boundary cells were excluded from the analysis to rededge effects. The model distribution appears in go
FIGURE 13. Scaling of segmental cross-sectional area a „ter-minals excluded … with the volume VVP of tissue supplied bythe respective subtrees for ten realizations of CCO modelswith lÄ2. The solid line represents the least-squares fit of apower-law relation, Eq. „15…, to the data points.

vertsin
entof
nointsliedllusthe
thewsaf-
tersd
ted
an-anenal
tow-rm-
g-
-yxi-
-
into-emsrder
560 KARCH et al.
qualitative agreement with the measured data. Howedue to the difficulties in identifying segmental end poinin the retinal fundus photograph, the results shownFig. 14 currently do not allow a quantitative assessmof model predictions regarding the spatial distributionend points in the retinal arterial system.
A Modified Algorithm for Tree Construction
In the previous section we have used the Voropolyhedra associated with individual terminal segmeas a geometric model for the regions of tissue suppby these segments. The reported scaling properties itrate that this model is a reasonable approach withinframework of CCO trees.
These observations suggest a modification ofpresent method of CCO, namely to set terminal floproportional to their associated Voronoi cell volumes
FIGURE 14. „a… Voronoi tessellation of apparent arterial endpoints digitized from data of the human retina published byZamir et al. „Ref. 63…. „b… Probability density distribution„meanÁSD… of normalized Voronoi cell area A *ÄA ÕŠA ‹ gen-erated by ten realizations of pruned 2D CCO trees „solidcircles … and by the points of panel „a… „open circles …. Thecontinuous line shows the best-fit two-parameter gammadistribution for the model trees.
,
-
ter each step of growth and to rescale segment diameof the whole tree accordingly to meet the modifieboundary conditions. This approach is further supporby a recent paper of Qian and Bassingthwaighte,42 whoshowed that asymmetric branching tree models with rdom flow variations at each bifurcation give rise toasymptotic lognormal flow distribution, in favorablagreement with experimental measurements of regioblood flow distributions in the heart and lung.~Note thatwe have used two-parameter gamma distributionscharacterize Voronoi cell volumes in CCO trees; hoever, Vaz and Fortes57 have shown that two-parametegamma and lognormal distributions with suitable paraeters are very similar.!
Figure 15 shows the distributions of normalized semental flows,q* 5q/^q&, for CCO trees (l52) grownby the modified algorithm and with other model parameters left unchanged~Table 1!. The data are grouped bStrahler orders53 of the respective segments and appromated by best-fit lognormal distributions
p~x!51
A2px logsg
expF2~ logx2 log^x&0.5!
2
2 log2 sgG , x.0,
~16!
where^x&0.5 is the median andsg is the geometric standard deviation.@Qian and Bassingthwaighte42 consideredflow distributions as a function of generation numbersstrictly dichotomous binary trees; for the asymmetricpology of CCO trees the Strahler ordering scheme semore appropriate: terminal segments are assigned o
FIGURE 15. Probability density distribution „meanÁSD… ofnormalized segmental flows q *Äq ÕŠq ‹ for three realizationsof CCO tree models „lÄ2… grown by the modified algorithm.Data are grouped by Strahler orders i S of the respectivesegments. The continuous lines show best-fit lognormal dis-tributions, Eq. „16…. i SÄ1 „terminals …: solid circles and solidline. i SÄ2 „preterminals …: open circles and dashed line. i SÄ3: diamonds and dashed-dotted line. i SÄ4: triangles anddotted line.

h.
g-ly
witha-ntre-
ernp-asorkoel,ol-the
trictheandithhe
tyofapu-
r-ts,
ith
ru-
a-O
eted
onas
celO
ol-llOs.n-odol-
adthe
ex-
withthe
thes-hed
of
w
atizeheis-r-re-iesnalas-ingofO
onsar-s.ra-in
d-iti-
of
561Voronoi Analysis of Arterial Tree Models
1; if two first-order segments meet, the parent~i.e., pre-terminal! segment is assigned order 2, and so fort#Figure 15 illustrates that the distributionp(q* ) of nor-malized terminal flows is well approximated by a lonormal distribution, and this property is approximatealso observed for higher-order segments, consistentthe results of Ref. 42. For the terminal flows we findrelative dispersion of RDq537% and a skewness coefficient of gq51.12. Both values are in good agreemewith data reported for experimental measurements ofgional blood flow in the heart and the lung.6,14 Moreover,RDq and gq are roughly independent on the numbNterm of terminal segments~see the previous discussioon Nterm dependence!, suggesting that these values reresent an invariant property of CCO tree models, at lefor the parameter space considered in the present w
We finally note that the modified algorithm has nsignificant effect on other characteristics of the mode.g., the distribution of segment diameters and VP vumes, the pressure profile, the total tree volume, anddistribution of segments over Strahler orders.
SUMMARY AND CONCLUSIONS
In this paper we have analyzed topological and meproperties of the Voronoi polyhedra generated bydistal end points of terminal segments in CCO treesby randomly distributed points as a reference model wthe aim to characterize the spatial distribution of trespective point sets.
We have evaluated the distributions of the numberNf
of Voronoi cell faces, the cell volumeV, surface areaS,the areaA of individual cell faces, and the asphericiparametera. We have characterized the distributionsthese quantities by their respective moments and byproximation with best-fit two-parameter gamma distribtions ~Tables 2 and 3!. The distributions ofNf , V, S, A,anda of the CCO models all proved significantly diffeent from the ones obtained for the random poinwhereas the distributions ofV, S, and a were also sig-nificantly different between CCO models generated wdifferent modes of optimization ~two-sampleKolmogorov–Smirnov test,p,0.0001). Two-parametegamma distributions not only characterize the distribtions of Nf , V, and S for Poisson–Voronoitessellations,30 but also provide a reasonable approximtion for the distributions of these quantities in CCmodels.
Both the distribution of Voronoi cell volumes and thdistribution ofkth nearest neighbor distances as reflecin the pair distribution functiong(r ) consistently showedthe repulsive effect of a growing CCO tree structurethe spatial arrangement of its terminal supply pointswell as the tendency of such trees to balance thevolume distribution observed for random points: CC
t.
-
l
trees increase the number of Voronoi cells having vumes near the mean value^V& at the expense of smaand large cells, i.e., the cell volume distribution for CCmodels is more homogeneous than for random point
For comparison of model predictions with experimetal data we have studied scaling of intravascular blovolume and of arterial cross-sectional area with the vume of supplied tissue, which we have modeled ongeometrical~not functional! basis by the VP generateby the model’s terminal segments. Depending onparticular value of the bifurcation exponentg and on thesize of subtrees considered, we have found scalingponents betweens51.2060.007 ands51.0860.005 forintravascular blood volume andb50.7760.01 for arte-rial cross-sectional area. These results are consistentexperimental data and model predictions reported inliterature.41,51,60
Based on these observations we have extendedCCO algorithm to set terminal flows proportional to asociated VP volumes. As a major improvement of tmodel we now obtain terminal flows well approximateby a lognormal distribution with a relative dispersionRDq537% and a skewness coefficient ofgq51.12, ingood agreement with theoretical considerations42 and ex-perimental measurements of regional blood flodistributions.6,14
The first conclusion we draw from this study is thVoronoi analysis proved a valuable tool to characterthe spatial distribution of end points governed by tpresence of a growing CCO tree structure and to dcriminate such distributions from a purely random arangement of points. From the comparison of model pdictions with experimentally observed scaling propertof intravascular blood volume and arterial cross-sectioarea with supplied tissue volume we conclude thatsigning to each terminal segment its correspondVoronoi polyhedron reasonably defines the regionstissue supplied by individual terminal segments in CCtrees. Finally, a modified algorithm of tree constructiwith terminal flows set proportional to Voronoi volumeyields a considerable improvement of the model, in pticular with respect to the distribution of terminal flowFor future developments, this new mode of tree genetion may prove useful to study flow heterogeneity withthe framework of CCO trees.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the valuable avice from the editor regarding the literature and the crcal comments provided by the anonymous reviewersthis study.

of5.s,
of
d-d J.1–
heis,
ck-
tal
uredel
ec-s.
,lar
the
s-
nd
ofe-
on
y-or-
icra
la-
hella-
o-
.is-
in
uc-
of
. Aen-
er.
g.
ofal
te.
a.ela-
al
el-lity
.we-
i-owseo-
of
.
ster.y.
er.In:r-ta-
tialia-
etry.
als.
r-
562 KARCH et al.
REFERENCES
1Allen, M. P., and D. J. Tildesley. Computer SimulationLiquids. Oxford: Oxford University Press, 1987, pp. 54–5
2Arts, T., R. T. I. Kruger, W. VanGerven, J. A. C. Lambregtand R. S. Reneman. Propagation velocity and reflectionpressure waves in the canine coronary artery.Am. J. Physiol.237:H469–H474, 1979.
3Aurenhammer, F., and R. Klein. Voronoi diagrams. In: Hanbook of Computational Geometry, edited by J. R. Sack anUrrutia. Amsterdam: Elsevier Science B. V., 2000, pp. 20290.
4Barber, C. B., and H. Huhdanpaa. Qhull version 2.6. TGeometry Center, University of Minnesota, Minneapol1998. http://www.geom.umn.edu/locate/qhull.
5Barber, C. B., D. P. Dobkin, and H. Huhdanpaa. The quihull algorithm for convex hulls.ACM Trans. Math. Softw.22:469–483, 1996.
6Bassingthwaighte, J. B., R. B. King, and S. A. Roger. Fracnature of regional myocardial blood flow heterogeneity.Circ.Res.65:578–590, 1989.
7Beard, D. A., and J. B. Bassingthwaighte. The fractal natof myocardial blood flow emerges from a whole-organ moof arterial network.J. Vasc. Res.37:282–296, 2000.
8Beard, D. A., and J. B. Bassingthwaighte. Modeling advtion and diffusion of oxygen in complex vascular networkAnn. Biomed. Eng.29:298–310, 2001.
9Brown, D. Voronoi diagrams from convex hulls.Inf. Process.Lett. 9:223–228, 1979.
10Chilian, W. M., S. M. Layne, E. C. Klausner, C. L. Easthamand M. L. Marcus. Redistribution of coronary microvascuresistance produced by dipyridamole.Am. J. Physiol.256:H383–H390, 1989.
11Chilian, W. M. Microvascular pressures and resistances inleft ventricular subepicardium and subendocardium.Circ.Res.69:561–570, 1991.
12Dirichlet, G. L. Uber die Reduction der positiven quadratichen Formen mit drei unbestimmten ganzen Zahlen.J. ReineAngew. Math.40:209–227, 1850.
13Fung, Y. C. Biomechanics: Motion, Flow, Stress, aGrowth. New York: Springer, 1990, pp. 155–195.
14Glenny, R. W., and H. T. Robertson. Fractal propertiespulmonary blood flow: Characterization of spatial heterogneity. J. Appl. Physiol.69:532–545, 1990.
15Glenny, R. W., and H. T. Robertson. A computer simulatiof pulmonary perfusion in three dimensions.J. Appl. Physiol.79:357–369, 1995.
16Gil Montoro, J. C., and J. L. F. Abascal. The Voronoi polhedra as tools for structure determination in simple disdered systems.J. Phys. Chem.97:4211–4215, 1993.
17Gil Montoro, J. C., F. Bresme, and J. L. F. Abascal. Ionassociation in electrolyte solutions: A Voronoi polyhedanalysis.J. Chem. Phys.101:10892–10898, 1994.
18Godde, R., and H. Kurz. Structural and biophysical simution of angiogenesis and vascular remodeling.Dev. Dyn.220:387–401, 2001.
19Hinde, A. L., and R. E. Miles. Monte Carlo estimates of tdistributions of the random polygons of the Voronoi tessetion with respect to a Poisson process.J. Stat. Comput.Simul. 10:205–223, 1980.
20Honda, H. Description of cellular patterns by Dirichlet dmains: The two-dimensional case.J. Theor. Biol. 72:523–543, 1978.
21Hoofd, L., Z. Turek, K. Kubat, B. E. M. Ringnalda, and SKazda. Variability of intercapillary distance estimated on h
tological sections of rat heart.Adv. Exp. Med. Biol.191:239–247, 1985.
22Hudlicka, O., A. J. Wright, and A. M. Ziada. Angiogenesisthe heart and skeletal muscle.Can. J. Cardiol.2:120–123,1986.
23Jedlovszky, P. Voronoi polyhedra analysis of the local strture of water from ambient to supercritical conditions.J.Chem. Phys.111:5975–5985, 1999.
24Kamiya, A., and T. Togawa. Optimal branching structurethe vascular tree.Bull. Math. Biophys.34:431–438, 1972.
25Karch, R., F. Neumann, M. Neumann, and W. Schreinerthree-dimensional model for arterial tree representation, gerated by constrained constructive optimization.Comput.Biol. Med. 29:19–38, 1999.
26Karch, R., F. Neumann, M. Neumann, and W. SchreinStaged growth of optimized arterial model trees.Ann.Biomed. Eng.28:495–511, 2000.
27Kassab, G. S., C. A. Rider, N. J. Tang, and Y.-C. B. FunMorphometry of pig coronary arterial trees.Am. J. Physiol.265:H350–H365, 1993.
28Kassab, G. S., J. Berkley, and Y. C. B. Fung. Analysispig’s coronary arterial blood flow with detailed anatomicdata.Ann. Biomed. Eng.25:204–217, 1997.
29King, R. B., L. J. Weissman, and J. B. BassingthwaighFractal descriptions for spatial statistics.Ann. Biomed. Eng.18:111–121, 1980.
30Kumar, S., S. K. Kurtz, J. R. Banavar, and M. G. SharmProperties of a three-dimensional Poisson–Voronoi tesstion: A Monte Carlo study.J. Stat. Phys.67:523–551, 1992.
31Kumar, S., and S. K. Kurtz. Properties of a two-dimensionPoisson–Voronoi tesselation: A Monte-Carlo study.Mater.Charact. 31:55–68, 1993.
32Kurz, H., and K. Sandau. Modelling of blood vessel devopment. Bifurcation pattern and hemodynamics, optimaand allometry.Comments Theor. Biol.4:261–291, 1997.
33Levy, S., T. Munzer, M. Phillips, C. Fowler, N. Thurston, DKrech, S. Wisdom, D. Meyer, and T. Rowley. Geomvieversion 1.6.1. The Geometry Center, University of Minnsota, Minneapolis, 1998. http://www.geomview.org.
34Lipowsky, H. H., and B. W. Zweifach. Methods for the smultaneous measurement of pressure differentials and flin single unbranched vessels of the microcirculation for rhlogical studies.Microvasc. Res.14:345–361, 1977.
35Meijering, J. L. Interface area, edge length, and numbervertices in crystal aggregates with random nucleation.PhilipsRes. Rep.8:270–290, 1953.
36Murray, C. D. The physiological principle of minimum workI. The vascular system and the cost of blood volume.Proc.Natl. Acad. Sci. U.S.A.12:207–214, 1926.
37Neumann, M., F. J. Vesely, O. Steinhauser, and P. SchuSolvation of large dipoles. I. A molecular dynamics studMol. Phys.35:841–855, 1978.
38Neumann, F., M. Neumann, R. Karch, and W. SchreinVisualization of computer-generated arterial model trees.Simulation Modelling in Bioengineering, edited by M. Cerolaza, D. Jugo, and C. A. Brebbia. Southampton: Computional Mechanics, 1996, pp. 259–268.
39Okabe, A., B. Boots, K. Sugihara, and S. N. Chiu. SpaTessellations: Concepts and Applications of Voronoi Dgrams, Second Edition. Chichester: Wiley, 1999.
40Preparata, F. P., and M. I. Shamos. Computational GeomAn Introduction. New York: Springer, 1985.
41Prothero, J. W. Scaling of blood parameters in mammComparative Biochem. Physiol.67A:649–657, 1980.
42Qian, H., and J. B. Bassingthwaighte. A class of flow bifu

r-
s.
r-
eby
o-an
ree
of
A.es o
e-ee.
and
or-
A:
thene-
th-ed
e
s a.
ly
del
ng
al
563Voronoi Analysis of Arterial Tree Models
cation models with lognormal distribution and fractal dispesion. J. Theor. Biol.205:261–268, 2000.
43Rahman, A. Liquid structure and self-diffusion.J. Chem.Phys.45:2584–2592, 1966.
44Ripley, B. D. Test of ‘randomness’ for spatial point patternJ. R. Statist. Soc. B41:368–374, 1979.
45Rosen, R. Optimality Principles in Biology. London: Butteworth, 1967, pp. 40–60.
46Ruocco, G., M. Sampoli, and R. Vallauri. Analysis of thnetwork topology in liquid water and hydrogen sulphidecomputer simulation.J. Chem. Phys.96:6167–6176, 1992.
47Sandau, K., and H. Kurz. Modelling of vascular growth prcesses: A stochastic biophysical approach to embryonicgiogenesis.J. Microsc.175:205–213, 1994.
48Schreiner, W. Computer generation of complex arterial tmodels.J. Biomed. Eng.15:148–149, 1993.
49Schreiner, W., and P. Buxbaum. Computer-optimizationvascular trees.IEEE Trans. Biomed. Eng.40:482–491, 1993.
50Schreiner, W., M. Neumann, F. Neumann, S. M. Roedler,End, P. Buxbaum, M. R. Mu¨ller, and P. Spieckermann. Thbranching angles in computer-generated optimized modelarterial trees.J. Gen. Physiol.103:975–989, 1994.
51Seiler, C., R. L. Kirkeeide, and K. L. Gould. Basic structurfunction relations of the epicardial coronary vascular trCirculation 85:1987–2003, 1992.
52Stoyan, D., and H. Stoyan. Fractals, Random ShapesPoint Fields. Chichester: Wiley, 1994.
53Strahler, A. N. Quantitative analysis of watershed geomphology. Trans. Am. Geophys. Union38:913–920, 1957.
-
f
54Thompson, D. W. On Growth and Form. Cambridge, MCambridge University Press, 1917.
55Van Bavel, E., and J. A. E. Spaan. Branching patterns inporcine coronary arterial tree. Estimation of flow heterogeity. Circ. Res.71:1200–1212, 1992.
56Van Beek, J. H. G. M., S. A. Roger, and J. B. Bassingwaighte. Regional myocardial flow heterogeneity explainwith fractal networks.Am. J. Physiol.257:H1670–H1680,1989.
57Vaz, M. F., and M. A. Fortes. Grain size distribution: Thlognormal and the gamma distribution functions.Scr. Metall.22:35–40, 1988.
58Voronoi, G. Nouvelles applications des parametres continula theorie des formes quadratiques.J. Reine Angew. Math134:198–287, 1908.
59Wang, C. Y., and J. B. Bassingthwaighte. Capillary suppregions.Math. Biosci.173:103–114, 2001.
60West, G. B., J. H. Brown, and B. J. Enquist. A general mofor the origin of allometric scaling laws in biology.Science276:122–126, 1997.
61Woldenberg, M. J., and K. Horsfield. Relation of branchiangles to optimality for four cost principles.J. Theor. Biol.122:187–204, 1986.
62Zamir, M. Optimality principles in arterial branching.J.Theor. Biol.62:227–251, 1976.
63Zamir, M., J. A. Medeiros, and T. K. Cunningham. Arteribifurcations in the human retina.J. Gen. Physiol.74:537–548, 1979.





























