Embed Size (px)
Citation preview

advantage. Any increase in benefitrecovery (via kin selection) could movethe cost/benefit ratio across the tippingpoint.
Kin selection is very likely to bea strong force helping maintain genesencoding secreted factor even in theabsence of gene transfer. Bacteriaare not randomly mixed; E. coli’sneighbors in the intestinal lumen willbe brethren produced from common(great)n-grandparents. Because celldivision places daughter cells adjacentto one another, ‘public goods’ willbe shared most frequently betweenmembers of a clone. The mechanisticquestion posed by Nogueira et al.’s [5]model is whether gene transfercan convert neighboring cells into‘relatives’ at a rate sufficient tominimize their depletion of the publicgoods. It is not clear that individual-level selection is not sufficient toexplain the data. First, we expectthe genes under strong selection tobe retained most readily; these wouldfail to show a history of loss andreacquisition. Genes under weakselection would be those detectedby Nogueira et al. [5], and these maybe biased towards those encodedsecreted proteins since their cost islarge relative to their diluted benefits.Alternatively, genes encoding externaland secreted proteins are often subjectto frequency-dependent and/ordiversifying selection, both of whichmay lead to gene loss and reacquisition[9]. It is very difficult to determine if thebenefits recovered by individual cellsfail to exceed the costs, thereforerequiring kin selection to make upthe difference. There is an associationbetween genes encoding secretedproteins and integrases and they are
more often found in recombination‘hotspots’. This would be expectedif this were a mechanism for genereacquisition, following from theirfirst observation. Yet it does notaddress the question of relativecost and benefit.
How is cooperation enforced? If kinselection were to provide sufficientbenefit to maintain cooperation, thenenforcement is not necessary. YetNogueira et al. [5] offer an additionroute, whereby addiction modulesweed out cheaters. These geneticelements are easily acquired butdifficult to lose, as their deletion fromthe genome can be lethal (hence‘addiction’ of the cell to theirpresence) [10]. A common example isbacterial restriction/modificationgenes: acquisition of the genes is easy,but sudden loss can be lethal asthe longer-lived restriction enzymebegins to degrade genomic DNA thatis left unprotected by its cognatemodification enzyme. The authorsfind an association between genesencoding secreted proteins and thosebelonging to addiction modules.Therefore, sudden loss of genesencoding social traits could alsoremove the addiction module, resultingin cell death; thus, this associationenforces cooperation by preventingrecidivism (Figure 1). The associationbetween social genes and addictionmodules may simply reflect the transitof both elements on extrachromosomalelements; it remains to be seen ifa social gene’s proximity to anaddiction module has a meaningfulimpact on its frequency in thepopulation.
As a whole, Nogueira et al. [5] do notclearly demonstrate that horizontal
gene transfer drives cooperationamong bacteria. But that is alsoa very tall order. What they have doneis provide an intriguing analysisthat forces microbiologists bothto examine forces leading to genepersistence in populations andto reconsider gene function solelyin the context of individual cells.
References1. McFall-Nagai, M. (2008). Host-microbe
symbiosis: the squid-Vibrio association — anaturally occurring, experimental model ofanimal/bacterial partnerships. Adv. Exp. Med.Biol. 635, 102–112.
2. Velicer, G.J., and Vos, M. (2009). Sociobiologyof the myxobacteria. Annu. Rev. Microbiol. 63,599–623.
3. Branda, S.S., Gonzalez-Pastor, J.E., Ben-Yehuda, S., Losick, R., and Kolter, R. (2001).Fruiting body formation by Bacillus subtilis.Proc. Natl. Acad. Sci. USA 98, 11621–11626.
4. Federla, M.J. (2009). Autoinducer-2-basedchemical communication in bacteria:complexities of interspecies signaling. Contrib.Microbiol. 16, 18–32.
5. Nogueira, T., Rankin, D.J., Touchon, M.,Taddei, F., Brown, S.P., and Rocha, E.P.C.(2009). Horizontal gene transfer of thesecretome drives the evolution of bacterialcooperation and virulence.Curr. Biol. 19, 1683–1691.
6. Hamilton, W.D. (1964). The genetical evolutionof social behaviour. I. J. Theor. Biol. 7, 1–16.
7. Hamilton, W.D. (1964). The genetical evolutionof social behaviour. II. J. Theor. Biol. 7, 17–52.
8. MacLean, R.C. (2008). The tragedy of thecommons in microbial populations: insightsfrom theoretical, comparative and experimentalstudies. Heredity 100, 471–477.
9. Lipsitch, M., and O’Hagan, J.J. (2007). Patternsof antigenic diversity and the mechanisms thatmaintain them. J. R. Soc. Interface 4, 787–802.
10. Engelberg-Kulka, H., and Glaser, G. (1999).Addiction modules and programmed cell deathand antideath in bacterial cultures. Annu. Rev.Microbiol. 53, 43–70.
Department of Biological Sciences,University of Pittsburgh, Pittsburgh,PA 15260, USA.E-mail: [email protected]
DOI: 10.1016/j.cub.2009.09.045
DispatchR945
Visual Maps: To Merge or Not ToMerge
A recent study of a child born with one cerebral hemisphere has revealed anextreme developmental reorganization of visual cortex. Self-organizing visualmaps demonstrate a surprisingly flexible restructuring in response to corticalloss.
Alyssa A. Brewer
Primate visual cortex consists ofmultiple maps of visual space
which follow the organization of theretina — hence these maps aretermed retinotopic [1]. Under normalconditions, these retinotopic maps
are stable throughout life; however,there are several examples ofhow these retinotopic corticalmaps can be altered by geneticmutations and/or early visualexperience [2–4]. A recent study byMuckli et al. [5] provides a strikingillustration of the flexibility of thedevelopment of these retinotopicmaps in human visual cortex. Intheir case study, a 10 year old girlpresents with a congenital lossof her right cerebral hemispherebut with a visual field of view that

Current Biology Vol 19 No 20R946
unexpectedly spans both hemifieldsof visual space. Such a drasticchange in early in utero corticaldevelopment has rarely beeninvestigated.
To understand why the preservationof her bilateral vision is so surprising,one must examine the typical patternof spatial organization in visualcortex. Normally, the retinotopicmaps of primate visual cortex spanthe occipital lobes of both corticalhemispheres (Figure 1A) [6]. Theinformation from the left hemifieldof visual space is represented in theprimary visual cortex (V1) of theright occipital lobe, and theinformation from the right hemifieldis represented in V1 of the leftoccipital lobe. A coherentrepresentation of visual space isvery carefully conserved in each V1,with the retinotopic map smoothlyrepresenting visual space from theupper to the lower vertical meridian
of each hemifield. At therepresentations of the dorsal andventral vertical meridia in V1, thespatial representations reverse backtowards the horizontal meridianto form the dorsal and ventralquarterfield representations thatcomprise area V2 (Figure 1B).This repetition of distinct maps ofvisual space is repeated throughoutlarge regions of visual cortex.Within a single visual field map,each position in visual space is onlyrepresented once, and each mapis assumed to perform a set ofunique computations across visualspace [7].
In accordance with this division ofvisual image processing into distincthemispheres, image informationfrom each hemifield normally reachesvisual cortex along very separatepathways [8]. An image falling on theretinae is separated into nasal andtemporal components in each eye.
Consider in Figure 2A how theimage information from just the lefthemifield is transmitted from theretinae to visual cortex. The part ofthe image within the left hemifieldfalls on the nasal retina in the lefteye and the temporal retina in theright eye. The information fromthese two sections of retinaerepresenting the left hemifield istransmitted through the optic nerveto the optic chiasm. To keep thecorrect hemifield informationtogether for collective corticalprocessing, the axons fromthe nasal retina of the left eye crossover to the right optic tract at theoptic chiasm, while the axons fromthe temporal retina of the righteye remain on the right side.Together, these sets of fiberscarrying left hemifield imageinformation then project first tothe right lateral geniculate nucleus(LGN) of the thalamus and thenthrough the optic radiations toV1 at the occipital pole of theright cortical hemisphere.Retinotopic organization ispreserved along these pathwaysand then integrated into specificmaps within visual cortex underthe guidance of molecular cuesand patterns of coordinatedactivity [9].
So how does the child in the casestudy from Muckli et al. [5] havevision that spans both hemifields,when she is missing her entire rightvisual cortex? Muckli et al. [5]suggest that all of the axons of theintact left eye in their case studyproject to the left hemisphere,with the few axons of theunderdeveloped right eye also allcrossing at the chiasm to the lefthemisphere (Figure 2B). Therepresentations of visual space inthis subject show a massiverestructuring of the normalretinotopic maps to represent bothhemifields in a single corticalhemisphere. Alternative patternsof map organization forthe coherent merging of therepresentations of the left and righthemifields into a single hemisphereinclude: directly overlappingcontralateral and ipsilateral mapsand/or the insertion of additional,discrete ipsilateral spatialrepresentations [10]. In the first case,the same region of cortex wouldrepresent both hemifields of space
3°
B
A
V1
V2vV3v
V2d
FuG
ColS
CalS
POS
20°
FuGColS
CalS
POS CC
STS
CS
IPS
V1
V2vV3v
V2d
FuG
ColS
CalS
POS
FuGColS
CalS
POS CC
STS
CS
IPS
Current Biology
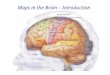
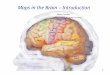
Figure 1. Visual field maps in human cortex.
(A) The span of cortical responses in posterior visual cortex to an expanding ring stimulus isshown across the left hemisphere of a single example subject. The lateral view is shown on theleft, medial on the right. Eccentricity measurements were made using a 3� radius stimulus. Thecolor overlay depicts the region of visual space (see inset) that most effectively stimulates thatregion of cortex. (B) A view of the posterior occipital lobe of a single subject’s left hemisphereillustrates the position of the first three posterior visual field maps: V1, V2, V3. The color over-lay indicates the visual field angle (left) or eccentricity (right) that produces the most powerfulresponse at each cortical location (see the colored legends on the right). For clarity,only responses within these visual field maps are colored, and only locations with a powerfulresponse are shown (statistical coherence >0.20). CalS, calcarine sulcus; POS, parieto-occipital sulcus; ColS, collateral sulcus; FuG, fusiform gyrus; IPS, intraparietal sulcus; STS,superior temporal sulcus; CS, central sulcus; CC, corpus callosum.

DispatchR947
smoothly across the corticalrepresentation, much like theinterspersed input to oculardominance columns. In the second,distinct islands of ipsilateralmaps would be present within thedistorted representation of thecontralateral hemifield. FunctionalMRI (fMRI) measurements by Muckliet al. [5] suggest that both occur intheir case study across areas V1, V2and V3.
What might be the driving forcesbehind this unusual integration ofboth hemifields of visual space intoone hemisphere? The axons ofthe optic nerve originating fromthe retinal ganglion cells usuallyfind their way through this mazeof pathways by following precisemolecular guidance cues.These guidance cues directspecific axons to one hemisphereor the other, depending on theparticular receptor expressed byeach retinal cell interacting withreceptor cues at the optic chiasm(for example, Eph-B1, Ephrin-B2 [11]).The visual pathways are further
refined by synchronizedneuronal activity, both spontaneouswaves and experience-drivenresponses [9].
The disruption of normal guidancecues has been studied in severalgenetic models that demonstratethe possibility of the integrationof contra- and ipsilateral visualfields into a single hemisphere(for example, albino mutationswith complete chiasmatic crossing[3]). One outstanding question hasbeen the importance of potentialcontralateral and ipsilateralguidance cues in the properdevelopment of visual pathways. Inthe case study by Muckli et al. [5],the normal guidance cues fromone entire hemisphere are lost. Theredirection of the left nasal axonsto the ipsilateral left hemispheredemonstrates that there are norepressor cues from the ipsilateralhemisphere preventing thisrestructuring of the visualpathways. With no changes inmolecular expression thought toexist in the intact hemisphere in
this subject, changes in thepatterns of input activity are thelikely organizing forces for theflexible restructuring of theinternal representations ofvisual space in visual field mapslike V1.
The interplay of changes inmolecular guidance cues andself-organizing cortical activityproduce a merged corticalrepresentation of visual space inthis unique subject. Her ability tosee across both hemifieldsof visual space demonstratesthe astonishing range of flexiblerestructuring of the visualpathways possible underextreme changes of corticaldevelopment.
References1. Van Essen, D.C. (2003). Organization of visual
areas in macaque and human cerebral cortex.In The Visual Neurosciences, L.M. Chalupa andJ.S. Werner, eds. (Boston: Bradford Books),pp. 507–521.
2. Baseler, H.A., Brewer, A.A., Sharpe, L.T.,Morland, A.B., Jagle, H., and Wandell, B.A.(2002). Reorganization of human corticalmaps caused by inherited photoreceptorabnormalities. Nat. Neurosci. 5,364–370.
3. Hoffmann, M.B., Tolhurst, D.J., Moore, A.T.,and Morland, A.B. (2003). Organization of thevisual cortex in human albinism. J. Neurosci.23, 8921–8930.
4. Fine, I., Wade, A.R., Brewer, A.A., May, M.G.,Goodman, D.F., Boynton, G.M., Wandell, B.A.,and MacLeod, D.I. (2003). Long-termdeprivation affects visual perception andcortex. Nat. Neurosci. 6, 915–916.
5. Muckli, L., Naumer, M.J., and Singer, W. (2009).Bilateral visual field maps in a patient with onlyone hemisphere. Proc. Natl. Acad. Sci. USA106, 13034–13039.
6. Wandell, B.A., Dumoulin, S.O., andBrewer, A.A. (2007). Visual field maps in humancortex. Neuron 56, 366–383.
7. Wandell, B.A., Brewer, A.A., andDougherty, R.F. (2005). Visual field mapclusters in human cortex. Philos. Trans. R. Soc.Lond. B. 360, 693–707.
8. Horton, J.C., and Hoyt, W.F. (1991). Therepresentation of the visual field in humanstriate cortex. A revision of the classicHolmes map. Arch. Ophthalmol. 109,816–824.
9. Katz, L.C., and Shatz, C.J. (1996).Synaptic activity and the constructionof cortical circuits. Science 274,1133–1138.
10. Krubitzer, L. (2007). The magnificentcompromise: cortical field evolution inmammals. Neuron 56, 201–208.
11. Polleux, F., Ince-Dunn, G., and Ghosh, A.(2007). Transcriptional regulation ofvertebrate axon guidance and synapseformation. Nat. Rev. Neurosci. 8,331–340.
Department of Cognitive Sciences, 3151Social Sciences Plaza, University ofCalifornia, Irvine, Irvine, CA 92617, USA.E-mail: [email protected]
DOI: 10.1016/j.cub.2009.09.016
LGN
V1 V1
LGNLGNLGN
V1 V1
A B
LGN
V1 V1
LGN
VVVVVVVV
LGGGGGGGGGGGGGG
Current Biology
Figure 2. Visual pathways from retina to V1.
(A) The pathway of visual information from retina to primary visual cortex is shown overlaid onan axial T1 image. Note that the information from the left hemifield of visual space (green) ineach eye travels along pathways in the right hemifield, and vice versa for the right hemifield(red). ‘V1’ denotes the approximate locations of primary visual cortex in each hemisphere.Similarly, ‘LGN’ denotes the approximate locations of the lateral geniculate nucleus of the thal-amus. (B) The visual pathways of the case study from Muckli et al. [5] are estimated. Note thatall information in the left eye remains in the left hemisphere, with no crossing at the opticchiasm. Residual fibers from the subject’s underdeveloped right eye are shown as the dottedline crossing completely at the chiasm to the left hemisphere. The right cortical hemisphere isabsent in this subject.