Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Dec. 1979, p. 741-7480022-538X/79/12-0741/08$02.00/0
Vol. 32, No. 3
Herpes Simplex Virus Glycoproteins: Participation ofIndividual Herpes Simplex Virus Type 1 Glycoprotein
Antigens in Immunocytolysis and Their Correlation withPreviously Identified Glycopolypeptides
B. NORRILD,' t * S. L. SHORE,2 AND A. J. NAHMIAS'Department of Pediatrics, Emory University School ofMedicine, Atlanta, Georgia 30303,1 and Center for
Disease Control, Atlanta, Georgia 303332
Received for publication 20 March 1979
Tissue culture cells infected with herpes simplex type 1 virus express virus-specified glycoprotein antigens on the plasma membrane. Three of these havebeen previously identified and have been designated as Ag-11, Ag-8, and Ag-6. Inthe present study, immunoglobulins to each of the antigens were shown to becapable of mediating immunocytolysis in the presence of either complement(antibody-dependent complement-mediated cytotoxicity) or peripheral bloodmononuclear cells (antibody-dependent cell-mediated cytotoxicity [ADCC]). Twoherpes simplex virus type 1 strains, VR-3 and F, reacted similarly in the ADCCtest in the presence of immunoglobulins to Ag-li, Ag-8, and Ag-6 in both infectedChang liver cells and HEp-2 cells. Anti-Ag-6, however, produced a lower ADCCreaction in HEp-2 cells than in Chang liver cells, suggesting differences in the Ag-6 surface expression in, or release from, these cells. Chang liver and HEp-2 cellsinfected with the MP mutant strain of herpes simplex virus type 1 showedreduced ADCC in the presence of anti-Ag-il and anti-Ag-8, but no reactivity atall with anti-Ag-6. Crossed immunoelectrophoretic analysis showed that MP-infected cell extracts contain Ag-11 and Ag-8, but lack Ag-6. Polypeptide analysisofherpes simplex virus type 1 strains F, VR-3, andMP showed that Ag-li consistsof the glycoproteins gA and gB, that Ag-8 consists of gD, and that Ag-6 consistsof gC. In conclusion, the present study demonstrates that either one of theglycoproteins (gC, gD, and a mixture of gA and gB) can function as a target forimmunocytolysis and that the antibody preparation to gC (Ag-6) does not cross-react with any of the other glycoproteins.
Herpes simplex virus type 1 (HSV-1) inducesthe incorporation of viral proteins into the mem-branes of infected cells (6, 21). Insertion of theviral proteins in the plasma membrane impartsa new antigenic specificity to the infected cells,as demonstrated by various immunological as-says. These have included lysis of cells withantibody and complement (antibody-dependentcomplement-mediated cytotoxicity [AbC]) andlysis by antibody in the presence of peripheralblood leukocytes (K-lymphocytes, polymorpho-nuclear leukocytes, or monocytes), a processtermed antibody-dependent cell-mediated cyto-toxicity (ADCC) (8, 14, 15, 19). With the aid ofthe crossed immunoelectrophoretic technique,we have demonstrated previously that HSV-1-infected cells contain three major membrane-bound glycosylated antigens, designated as Ag-11, Ag-8, and Ag-6 (10, 24). When injected into
t Present address: Institute of Medical Microbiology, Uni-versity of Copenhagen, DK-2100 Copenhagen 0, Denmark.
rabbits, each of these antigens produced anti-body which reacted only with the respectiveantigen when tested by the crossed immunoelec-trophoretic technique, employing an intermedi-ate gel (1, 25).
In the present paper we report that: (i) eachof the glycoprotein antigens Ag-il, Ag-8, andAg-6 reacts as a target for imnmunocytolysiswhen rabbit sera reacting to each of the threeantigens are employed in either the comple-ment-dependent or the cell-dependent cytolyticassay; (ii) Ag-11, Ag-8, and Ag-6 consist of theglycopolypeptides gA plus gB, gD, and gC, re-spectively; (iii) the antibody preparation to Ag-6 (gC) only reacts to gC, as demonstrated by thelack of reactivity to cells infected with HSV-1strain MP which lacks gC.
MATERIALS AND METHODSVirus strains and cels. HSV-1 strains F and 17
(2, 22) were propagated in HEp-2 cells; HSV-1 strain
741
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

742 NORRILD, SHORE, AND NAHMIAS
VR-3 was propagated in Chang liver (CL) cells inEagle minimum essential medium (MEM) with 2%fetal calf serum (FCS) (maintenance medium). Themutant HSV-1 (MP) previously shown to lack onemajor glycopolypeptide, gC (5, 20), and used primarilyto demonstrate the specificity of the reagents, was alsopropagated in HEp-2 cells.
Infection of cells. (i) For ADCC and AbC tests,HEp-2 or CL cells were infected with HSV-1 as follows.Monolayers in 25-cm2 plastic flasks were infected at amultiplicity of infection of 1 to 2 PFU ofVR-3 per cell,5 to 10 PFU of F per cell, and 20 to 30 PFU ofMP percell. After 1 h of absorption at 37°C, the monolayerwas washed twice in phosphate-buffered saline, andreplenished with 10 ml of maintenance medium. (ii)For antigen production and polypeptide analysis, 150-cm2 monolayers of rabbit kidney (RK), CL, or HEp-2cells were exposed to either VR-3, F, MP, or 17 at 10to 20 PFU/cell. After 1 h of adsorption, the monolayercultures were overlaid with 50 ml of maintenancemedium.Labeling of cells with radioisotopes. The main-
tenance medium was replaced at 9 h postinfection bymedium containing 1/10 the normal amount ofleucine,isoleucine, and valine, but supplemented with 5 ,uCi ofeach of the amino acids [U-'4C]leucine, [U-'4C]isoleu-cine, and [U- 4C]valine per ml of maintenance medium(300 mCi/mmol for each amino acid; New EnglandNuclear Corp., Boston, Mass.). The cells were proc-essed at 10 h postinfection. For labeling with glucosa-mine, the maintenance medium was replaced at 9 hpostinfection with medium containing 1/10 the normalconcentration of glucose, but supplemented with 1 uCiof D-[1-'4C]glucosamine per ml of medium (45 to 55mCi/mmol; New England Nuclear Corp.). The cellswere processed at 20 h postinfection.
Preparation of viral protein and polypeptides.(i) Triton X-100 solubilization. For Triton X-100solubilization, HSV-1-infected cells were scraped offthe flasks, washed once in Hanks balanced salt solu-tion, and suspended in 4 volumes of 5% Triton X-100in 0.020 M glycine-0.0076 M Tris buffer, pH 8.6. Thesuspension was disrupted by sonication and centri-fuged as described elsewhere (10).
(ii) SDS-mercaptoethanol solubilization. Forsodium dodecyl sulfate (SDS)-mercaptoethanol solu-bilization, monolayers of infected cells were washedonce in Hanks balanced salt solution and disrupted in2% (wt/vol) SDS-5% (vol/vol) 2-mercaptoethanol, asreported by Morse et al. (9).
Antibodies. Polyspecific antibodies were producedby inoculation of rabbits with HSV-1 (F)-infected rab-bit cornea cells disrupted in distilled water (25). Anti-bodies to Ag-li, Ag-8, and Ag-6 were obtained afterimmunization of rabbits with individual immunopre-cipitates, obtained from lysates of HSV-1 (VR-3)-in-fected RK cells, as described elsewhere (25). Theglobulin fractions of the sera were obtained by am-monium sulfate precipitation, followed by serial di-alysis against water, 0.05 M acetate buffer (pH 5.0),and phosphate-buffered salt solution, pH 7.2 (3). Allpurified globulin fractions were absorbed with unin-fected HEp-2 or CL cells as follows. Globulin aliquots(1 ml each) were absorbed three times for 1 h at 37°Cwith 5 x 107 cells. All serum globulins were heat
J. VIROL.
inactivated at 56°C for 30 min before use in thecytotoxicity assays.Crossed immunoelectrophoresis. Crossed im-
munoelectrophoresis, including the intermediate gelprocedure, has been detailed elsewhere (10, 25).Briefly, 30 A1 of Triton X-100-solubilized antigens fromHSV-1 (F)- or HSV-1 (MP)-infected cells were elec-trophoresed in a 1% (wt/vol) agarose gel (Indubiose,Vitry-sur-Seine, France) in 0.18 M Tris-0.06 M barbi-tal buffer, pH 8.6. First-dimension electrophoresis wasperformed at 5 V/cm for 1.5 h. For second-dimensionelectrophoresis, the agarose gel contained 15 tll ofpolyspecific rabbit immunoglobulin per cm2 (30 to 40mg of protein per ml). The intermediate gel containedeither preimmune rabbit globulin or monospecific im-munoglobulin at a concentration of 10 Pd/cm2. Thesecond electrophoresis was performed at 2 V/cm for18 h. According to the experiment, individual immu-noprecipitates were either (i) examined only afterstaining, (ii) cut out for antibody preparation, or (iii)extracted and solubilized for polypeptide analysis (11).SDS-polyacrylamide gel electrophoresis. HSV-
1-infected cell extracts and solubilized immunoprecip-itates were treated with 2% (wt/vol) SDS-5% (vol/vol) 2-mercaptoethanol and boiled for 1 min. Electro-phoresis was performed in a 9.25% (wt/vol) acrylamideseparation gel cross-linked with 0.24% (wt/vol) N,N-diallyltartardiamide (Bio-Rad Laboratories, Rich-mond, Calif.). The stacking gel was 3.0% (wt/vol)acrylamide-0.08% (wt/vol). N,N-diallyltartardiamide.Buffer conditions were as specified previously (9).Autoradiographs of the dried gels were obtained onCronex X-ray film (E. I. du Pont de Nemours & Co.,Inc., Wilmington, Del.).AbC. HSV (VR-3)-infected CL cells were sus-
pended with 0.25% trypsin-0.05% EDTA, washed threetimes in MEM containing 2% heat-inactivated FCS (I-FCS), and labeled in suspension at 18 h postinfectionwith 50 to 100 ytCi of 51Cr (sodium chromate, 200 to500 Ci/g of chromium; New England Nuclear Corp.)for 1 h. After labeling, the cells were washed througha cushion of 4 ml of I-FCS and resuspended in MEMwith 10% I-FCS. The AbC assay was performed in atotal volume of 0.3 ml. One-tenth of the target cells(7 x 103) was incubated with 0.1 ml ofvarious antibodydilutions and 0.1 ml of undiluted guinea pig comple-ment. The complement source was a pool of freshguinea pig sera. After incubation for 2 h at 37°C, 0.7ml of MEM with 10% I-FCS was added. The 51Crrelease was measured in triplicate samples in a gammacounter, as previously described (19).ADCC. The ADCC test, detailed elsewhere (8, 14,
19), was done as follows. CL or HEp-2 cells infectedwith VR-3, F, or MP for 18 h were trypsinized andlabeled in suspension with 50 to 100,uCi of 51Cr for 1h. The target cells were washed as described for theAbC assay and resuspended in MEM with 10% I-FCSto a concentration of 5 x 104 cells per ml. The mono-nuclear effector cells were prepared from the periph-eral blood of healthy HSV antibody-positive or -neg-ative human donors by the Ficoll-Hypaque densitycentrifugation technique (4). The cells were washedthree times in Hanks balanced salt solution and resus-pended in MEM with 10% I-FCS at a concentration of1.5 x 106 cells per ml. The assay was performed in a
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

HSV GLYCOPROTEINS 743
1.0-ml volume with 0.4 ml of target cells, 0.4 ml ofeffector cells, and 0.2 ml of rabbit antibodies dilutedas specified in the figure legends; the effector/targetcell ratio was 30:1. After 6 h of incubation at 37°C, thespecific 51Cr release was measured in triplicate sam-ples, as previously described (19).
Calculations. For AbC, percent specific 5"Cr re-lease was calculated as the percent release in thepresence of rabbit immunoglobulin and complementminus the percent release in the presence of preim-munoglobulin and complement. For ADCC, percentspecific 51Cr release was calculated as the percentrelease in the presence of immunoglobulin and mono-nuclear effector cells minus the percent release in thepresence of pre-immunoglobulin and mononuclear ef-fector cells. The release from target cells alone intothe medium was between 9 and 11% for the 2-h incu-bation time used in the AbC measurements and be-tween 7 and 14% for the 6-h incubation period used inthe ADCC assay. In the latter assay, the release ob-tained after the addition of effector cells alone neverexceeded 21%. The maximal cell-associated isotoperelease was 85%. With either the AbC or the ADCCassay, antibody titers were defined as the reciprocal ofthe dilution yielding half of the maximal specific re-lease (23).
RESULTSSurface expression of individual viral
glycoprotein antigens as measured by im-munocytolysis. Previous experiments witheither AbC or ADCC assays utilized polyspecificantibodies to HSV-1 obtained by immunizationof rabbits (14) or from human convalescent se-rum (19). The recent availability of antibodiesto three HSV glycoprotein antigens (Ag-11, Ag-8, and Ag-6) has enabled us to measure thecytotoxicity due to the reaction of each of theseantigens with the corresponding antibody (26,27).
(i) Reactivity of individual antigens inthe AbC reaction. HSV-1 (VR-3)-infected CLcells were labeled with 5"Cr at 18 h postinfection.The 51Cr-labeled target cells were reacted withpolyspecific immunoglobulin, as well as withanti-Ag-11, Ag-8, and Ag-6 rabbit immunoglob-ulins diluted from 1:30 to 1:1,000. All of theimmunoglobulin preparations initiated immu-nocytolysis, giving a maximal percent specificrelease ranging from 30 to 65% (Fig. 1). Thisfinding indicates that all three antigens are ex-posed at the surface of the cells and are able tobind antibody and complement. The comple-ment was used in a final concentration of 1:3, ashigher dilutions were found to give a lower spe-cific 5'Cr release (data not shown).
(ii) Reactivity of individual antigens inthe ADCC reaction. Previous studies with hu-man sera have suggested that the ADCC assayis more sensitive than the AbC method for thedetection of small amounts of antibodies (19,23). To confirm participation of all three anti-gens in the ADCC reaction and to compare thedifference in sensitivity between the two meth-ods, we diluted the various immunoglobulinsfrom 1:1,000 to 1:300,000 and reacted them withtarget HSV-1 (VR-3)-infected CL cells labeledwith 51Cr at 18 h postinfection (Fig. 2). Theresults show that all of the anti-HSV sera testedin these assays mediated cytolysis and that theADCC assay was more than 100-fold more sen-sitive in detecting HSV antibodies than was theAbC method (Table 1). Therefore, in all subse-quent experiments, only the ADCC reaction wasused.
(iii) Possible influence ofhost cells on thesurface expression of an individual anti-
80
170
1:10 1:30 1:100 1:300 1:1000Ab. dilution
FIG. 1. Titration of the immunoglobulin preparations in the AbC assay with HSV-I (VR-3)-infected CLcells. Symbols: x, polyspecific anti-HSV-I;@*, anti-Ag-li;E, anti-Ag-8; A, anti-Ag-6. Arrows representantibody titers of the various immunoglobulins.
VOL. 32, 1979
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

744 NORRILD, SHORE, AND NAHMIAS
30U2!4 30._
220
1:1000 1:3000 1:10.000 1:30.000 1:100.000 1:300.000Ab. dilution
FIG. 2. Titration of the immunoglobulin preparations in the ADCC assay with HSV-1 (VR-3)-infected CLcells. Symbols: x, polyspecific anti-HSV-I; 0, Anti-Ag-11; , anti-Ag-8; A, anti-Ag-6. Arrows representantibody titers of the various immunoglobulins.
TABLE 1. Comparison of antibody titers in the AbCand ADCC assays
Antibody titer Ratio ofImmunoglob- ADCC/AbCulin AbC ADCC titers
Anti-HSV-1 240 47,000 195Anti-Ag-1l 160 27,000 168Anti-Ag-8 440 270,000 613Anti-Ag-6 700 110,000 157
gen. Experiments were done to ascertainwhether the host cell might influence surfaceviral antigen expression. This was relevant asmost of our previous studies with immunocyto-lytic assays were performed in CL cells infectedwith HSV-1 (VR-3) (8, 14, 19), whereas most ofthe information regarding HSV-1 polypeptidesand antigens has been obtained in HEp-2 cellsinfected with other strains of HSV-1 (5, 11). Theimmunocytolysis obtained with HEp-2 cells in-fected with HSV-1 (VR-3) or (F) was comparedwith that measured with CL cells infected withthe same strains. Table 2 gives the maximumpercent specific release for each of the immu-noglobulins. In both types of cells infected witheither HSV-1 (VR-3) or (F), similar percent spe-cific releases were obtained in the presence ofthe polyspecific globulin and the anti-Ag-li andanti-Ag-8 immunoglobulins. In contrast, lowerspecific 51Cr releases were obtained with anti-Ag-6 globulin when this was employed in thetest with HEp-2 target cells, but not when em-ployed with CL cells, infected with either strainF or VR-3.Antigenic analysis of cells infected with
TABLE 2. Comparison of different host cells andHSV-1 strains in ADCC mediated by polyspecific
and monospecific immunoglobulinsSpecific release (%)
HSV-1 (VR- HSV-1 (F)-Immunoglobulin 3)-infected infected cels
cells
HEp-2 CL HEp-2 CL
Anti-HSV-1 32.0 27.7 29.3 36.7Anti-Ag-li 27.5 24.5 26.5 34.7Anti-Ag-8 32.2 25.8 30.5 34.8Anti-Ag-6 13.4 23.7 12.0 27.8
HSV-1 (MP). Inasmuch as a mutant strain ofHSV-1, (MP), fails to accumulate glycoproteingC (20), we investigated antigen reactivity ofcells infected with this strain in the ADCC testand by crossed immunoelectrophoresis. TheADCC test was done with both HEp-2 and CLinfected cells. The results (Table 3) show thatanti-Ag-6 failed to mediate the ADCC reactionwith either type of infected cells. It is of interestthat polyspecific anti-Ag-11 and anti-Ag-8 im-munoglobulins also produced a lower specific5"Cr release in MP-infected cells than in F- orVR-3-infected cells.The lack of reactivity of the anti-Ag-6 immu-
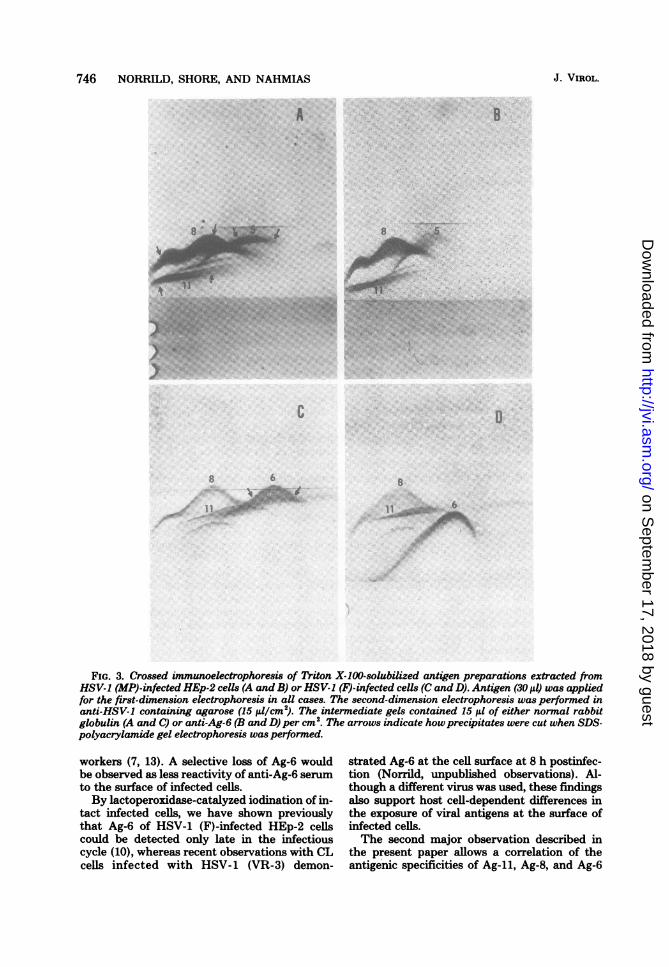
noglobulin with MP-infected target cellsprompted us to study HSV-1 (MP)-infected cellsby the crossed immunoelectrophoresis tech-nique to determine whether all three antigens(Ag-li, Ag-8, and Ag-6) are expressed. Analysisof Triton X-100-solubilized antigens by thistechnique showed that Ag-11 and Ag-8 were
J. VIROL.
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

VOL. 32, 1979
identified by electrophoresis through interme-diate gels containing antibodies to either Ag-11or Ag-8 (data not shown). On the other hand,Ag-6 could not be demonstrated, since the anti-Ag-6 in the intermediate gel did not shift any ofthe precipitates to a lower position (Fig. 3A andB). It should be noted that an antigen that wedesignate as Ag-5 is present and that its electro-phoretic mobility is sinilar to that of Ag-6 inHSV-1 (F)-infected cells (Fig. 3C and D). Ag-5was slightly glycosylated by D-[1-14C]glucosa-mine under labeling conditions used in the pres-ent study (data not shown).Correlation between viral antigens and
glycopolypeptides of HSV-1 (MP)-infectedcells. The polypeptides present in the individualTriton X-100-solubilized antigens of HSV-1(MP)-infected HEp-2 cells were identified witha 4C-amino acid-labeled antigen preparation.Individual immunoprecipitates were cut fromthe agarose gel after crossed immunoelectropho-resis, as shown with arrows in Fig. 3A, solubilizedin SDS and 2-mercaptoethanoL and analyzed onSDS-polyacrylamide gels (Fig. 4). The Ag-liprecipitate contained three polypeptides (Fig. 4,slot d), two of which correspond to the glyco-polypeptides gA and gB (Fig. 4, slots b and c)found by other workers (5, 20). The Ag-8 precip-itate showed one broad band corresponding tothe glycopolypeptide gD2 and a minor band cor-responding to gD, (Fig. 4, slots b, c, and e). Onlyone band, corresponding to the nonglycosylatedICP-5, was demonstrated in Ag-5 (Fig. 4, slots aand f). As a control for Ag-6, an Ag-6 immuno-precipitate was cut from an HSV-1 (F) referenceantigen preparation as shown in Fig. 4, slot g.Only one band corresponding to the glycopoly-peptide gC could be detected. HSV-1 strains 17and F have been used as reference strains inter-changeably, as no difference with respect toeither virus-specified polypeptides or antigenshas been demonstrated (9; Norrild, unpublisheddata). The reference bands for D-[14C]glucosa-mine-labeled HSV-1 (MP) polypeptides areshown in Fig. 4, slot c, which demonstrates theabsence of the glycopolypeptide gC.
DISCUSSIONViral antigens expressed on the surface of
HSV-1-infected cells have been shown to initiatedifferent immunocytolytic reactions in the pres-ence of human, as well as polyspecific rabbit,antibodies to HSV-1 (15, 19). The present studyhas extended these observations by analyzingthe ability of individual glycoprotein antigens(Ag-l, Ag-8, and Ag-6) to initiate immunocy-tolysis. Two different immunocytolytic mecha-nisms were used: AbC and ADCC. The antibody
HSV GLYCOPROTEINS 745
TABLE 3. ADCC mediated bypolyspecific andmonospecific immunoglobulins, using different host
cells infected with the HSV-1 (MP) strainSpecific release (%)
ImmunoglobulinHEp-2 cells CL cells
Anti-HSV-1 14.6 7.7Anti-Ag-ll 13.9 3.5Anti-Ag-8 19.4 7.1Anti-Ag-6 -2.8 -2.6
directed to the Ag-il, Ag-8, and Ag-6 prepara-tions mediated immunocytolysis in both assays,thus demonstrating the surface expression of thethree antigens late in the infectious cycle andthe ability of each of the viral antigens to partic-ipate independently as a target for the cytolyticreactions (Fig. 1 and 2). Titration of the immu-noglobulins by both methods showed that thesensitivity of the ADCC test for the detection ofantibodies was more than 100-fold that of theAbC reaction (Table 1), leading us to use thismethod for later studies. The interaction be-tween the membrane antigens and the antibod-ies to Ag-il, Ag-8, and Ag-6 is in agreement withprevious findings demonstrating the neutralizingcapacity of these antibodies (27) and also con-firms earlier findings demonstrating the pres-ence of identical viral antigenic determinants atthe surface of the plasma membrane of HSV-1-infected cells and at the surface of the viralenvelope (15, 16). The ability of individual viralglycoprotein antigens to participate in the AbCandADCC cytolytic reactions was demonstratedwith two wild-type HSV-1 strains, VR-3 and F.No difference was noted with the two types ofHSV-1 when the same host cell line was used,but the expressions of Ag-il, Ag-8, and Ag-6 atthe surface of infected cells appeared to be hostcell dependent when tested in the ADCC assay(Table 2). Thus, the specific release obtainedwith HSV-1-infected HEp-2 or CL cells wassimiar for the polyspecific, anti-Ag-11 and anti-Ag-8 antibodies. The reactivity of anti-Ag-6 an-tibody was lower in HEp-2 cells than in CL cellsinfected with HSV-1 (VR-3) or (F). The reasonfor this difference is unknown at present. Threepossibilities should be considered. First, theexpression of Ag-6 at the surface of infected cellsvaries depending on the host cell used. Second,the plasma membrane of tissue culture cellsvaries in intrinsic properties, as recently re-ported by Schlehofer et al., who observed differ-ent permeabilities ofHSV-1-infected cells to 50Cr(18). Third, the possibility also exists that theOOviral antigen Ag-6 is preferentially shed fromthe surface of HEp-2 cells into the tissue culturemedium, as previously suggested by several
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
746 NORRILD, SHORE, AND NAHMIAS
A
):
c
6.\...iFIG. 3. Crossed immunoelectrophoresis of Triton X-100-solubilized antigen preparations extracted from
HSV-1 (MP)-infected HEp-2 cells (A and B) or HSV-1 (F)-infected cells (C and D). Antigen (30 Id) was appliedfor the first-dimension electrophoresis in all cases. The second-dimension electrophoresis was performed inanti-HSV-I containing agarose (15 ,ul/cm2). The intermediate gels contained 15 il of either normal rabbitglobulin (A and C) or anti-Ag-6 (B and D) per cm2. The arrows indicate howprecipitates were cut when SDS-polyacrylamide gel electrophoresis was performed.
workers (7, 13). A selective loss of Ag-6 wouldbe observed as less reactivity of anti-Ag-6 serumto the surface of infected cells.By lactoperoxidase-catalyzed iodination of in-
tact infected cells, we have shown previouslythat Ag-6 of HSV-1 (F)-infected HEp-2 cells
could be detected only late in the infectiouscycle (10), whereas recent observations with CLcells infected with HSV-1 (VR-3) demon-
strated Ag-6 at the cell surface at 8 h postinfec-tion (Norrild, unpublished observations). Al-though a different virus was used, these findingsalso support host cell-dependent differences inthe exposure of viral antigens at the surface ofinfected cells.
The second major observation described inthe present paper allows a correlation of theantigenic specificities of Ag-11, Ag-8, and Ag-6
B
J. VIROL.
ADI
I I G:!:... AM116
--s..
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

HSV GLYCOPROTEINS 747
ICP a b c d-. % "R
5
7
8
11
20
25
29
4UM
__
-mma
43 400*
44 q0m
FIG. 4. SDS-polyacrylamideIa 9.25% (wt/vol) separation g0.24% (wt/vol) N,N-diallyltartHSV-1 (MP) "4C-amino acid latin disruption buffer; (slot b) BIglucosamine labeled and soluJbuffer; (slot c) HSV-1 (MP) D -[lbeled and solubilized in disruptand f) Ag-li, Ag-8, and Ag-5 Xbeled HSV-1 (MP) solubilizedtates; (slot g) Ag-6 of '4C-aminc(F) solubilized from immunopre
to the glycoproteins described by other workers(17, 20). In addition, a new antigenic determi-nant not previously described was identified anddesignated as Ag-5. The polypeptides of antigensAg-il, Ag-8, and Ag-5 were identified from "4C-amino acid-labeled extracts of HSV-1 (MP)-in-fected cells. As Ag-5 in crossed immunoelectro-phoresis showed an electrophoretic mobilitysimilar to that of Ag-6 from extracts of HSV-1(F)-infected cells (Fig. 3A and C), the polypep-tides of Ag-6 were included as a reference (Fig.4, slot g). The polypeptide analysis of Ag-ilidentified three polypeptides (Fig. 4, slot d), ofwhich the two with the highest molecular weight
B corresponded to glycoproteins gA and gB ofOk HSV-1 demonstrated by other workers (Fig. 4,
slots b, c, and d). This antigen has previouslybeen demonstrated to be in common with HSV-1, HSV-2, B-virus, and bovine herpes mammil-litis virus. Although the Ag-il immunocomplexconsists of different polypeptides when isolatedfrom extracts of cells infected with these differ-ent viruses, one glycoprotein was in commonwith the Ag-11 of the four viruses, thus indicat-ing that the major antigenic determinants wereprobably carried on this glycoprotein (12). Thepolypeptides of Ag-8 of HSV-1 (MP) showedseveral bands. A minor band corresponds to gDi,and the broad band observed above gD, corre-sponds to gD2 of Spear (20) (Fig. 4, slots c ande). Inasmuch as the different bands of 14C-aminoacid-labeled Ag-8 all correspond to various formsof gD, it might be concluded that the antigenicdeterminant sites are specified within gD. Thepolypeptide analysis of Ag-5 of HSV-1 (MP)demonstrated a significant difference from thatof Ag-6 of HSV-1 (F) (Fig. 4, slots f and g). Thus,Ag-5 predominantly consists of ICP-5, whichis identified as the major capsid polypeptide(5). The observation that Ag-5 is labeled by[I4C]glucosamine under certain conditions ismost likely because glucosamine labeling wasdone in medium with 1/10 of the normal con-centration of glucose. Reduced glucose allowslabeling of non-glycosylated proteins, and a 1/10concentration of glucose was near the criticalvalue where additional proteins showed label
gel electrophoresis in (Spear, personal communication). As the pres-,el cross-linked with ent study also identified the major polypeptideardiamide. (Slot a) of Ag-6 as gC (Fig. 4, slots b and g), the corre-beled and solubilized lation of Ag-il, Ag-8, and Ag-6 with the majorFSV-1 (17) D [1 _c]- glycoproteins gA, gB, gC, and gD can be sum-bilized in disruption marized as follows: Ag-il consists of gA and gB,-_ 14C]glucosamine la- Ag-8 consists of gD, and Ag-6 consists of gC.~ion buffer; (slots d, e,Df 14C-amino acid-la- Regarding the specificity of the antibody prep-from immunoprecipi- arations employed in the present study, we can) acid-labeled HSV-1 conclude that the Ag-6 (gO) antibody reacts onlycipitates. to gC (26). This is based on the observation that
VOL. 32, 1979
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

748 NORRILD, SHORE, AND NAHMIAS
Ag-6 antibody did not react to HSV-1 (MP)-infected cells when employed in the ADCC re-action. As the test is very sensitive to the con-centration of antibody, as discussed above, mi-nor amounts of antibody directed to other viralantigenic specificities would be detected. Theanti-Ag-li and anti-Ag-8 antibodies cannot betested for mono-specificity at present inasmuchas no mutants which lack either gA plus gB orgD are available.
In conclusion, the present paper demonstratesthat gA, gB, gC, and gD carry different antigenicspecificities and that either one of them canparticipate as a target for immunocytolytic re-actions in the presence of either complement ormononuclear leukocytes from human peripheralblood.
ACKNOWLEDGMENTSWe thank Theresa Cromeans and Phyllis Wood for excel-
lent technical assistance.This work was performed while B.N. was an International
Fogarty Fellow (grant Fo5TW2501) and was supported in partby a grant from the National Foundation-March of Dimes.
LITERATURE CITED1. Axelsen, N. H. 1973. Intermediate gel in crossed and
fused rocket immunoelectrophoresis. Scand. J. Immu-nol. 2(Suppl. 1):71-77.
2. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968.Characterization of herpes simplex virus strains differ-ing in their effects on social behavior of infected cells.J. Gen. Virol. 2:357-364.
3. Harboe, N., and A. Ingild. 1973. Immunization, isolationof immunoglobulins, estimation of antibody titers.Scand. J. Immunol. 2(Suppl. 1):161-164.
4. Harris, R., and E. 0. Ukaejiofo. 1969. Rapid prepara-tion of lymphocytes for tissue typing. Lancet ii:327.
5. Heine, J. W., R. W. Honess, E. Cassai, and B. RoIz-man. 1974. Proteins specified by herpes simplex virus.Xfl. The virion polypeptides of type 1 strains. J. Virol.14:640-651.
6. Heine, J. W., P. G. Spear, and B. Roizman. 1972.Proteins specified by herpes simplex virus. VI. Viralproteins in the plasma membrane. J. Virol. 9:431439.
7. Kaplan, A. S., J. S. Erickson, and T. Ben-Porat. 1975.Synthesis of proteins in cells infected with herpesvirus.X. Proteins excreted by cells infected with herpes sim-plex virus, types 1 and 2. Virology 64:132-143.
8. Kohl, S., S. E. Starr, J. M. Oleske, S. L Shore, R. B.Ashman, and A. J. Nahmias. 1977. Human mono-cyte-macrophage-mediated antibody-dependent cyto-toxicity to herpes simplex virus-infected cells. J. Im-munol. 118:729-735.
9. Morse, L. S., L Pereira, B. Roizman, and P. A. Schaf-fer. 1978. Anatomy of herpes simplex virus (HSV)DNA. X. Mapping of viral genes by analysis of polypep-tides and functions specified by HSV-1 x HSV-2 recom-binants. J. Virol. 26:389-410.
10. Norrild, B., 0. J. Bjerrum, H. Ludwig, and B. F.Vestergaard. 1978. Analysis of herpes simplex virustype 1 antigens exposed on the surface of infected tissueculture cells. Virology 87:307-316.
11. Norrild, B., 0. J. Bjerrum, and B. F. Vestergaard.1977. Polypeptide analysis of individual immunoprecip-itates from crossed immunoelectrophoresis. Anal. Bio-
J. VIROL.
chem. 81:432-441.12. Norrild, B., IL Ludwig, and R. Rott. 1978. Identification
of a common antigen of herpes simplex virus, bovineherpes mammillitis virus, and B virus. J. Virol. 26:712-717.
13. Norrild, B., and B. F. Vestergaard. 1979. Immunoelec-trophoretic identification and purification of herpessimplex virus antigens released from infected cells intissue culture. Intervirology 11:104-110.
14. Oleske, J. M., R. B. Ashman, S. Kohl, S. L Shore, S.E. Starr, P. Wood, and A. J. Nahmias. 1977. Humanpolymorp: .iuclear leucocytes as mediators of anti-body-dependent cellular cytotoxicity to herpes simplexvirus-infected cells. Clin. Exp. Immunol. 27:446-453.
15. Roane, P. R., Jr., and B. Roizman. 1964. Studies of thedeterminant antigens of viable cells. II. Demonstrationof altered antigenic reactivity of HEp-2 cells infectedwith herpes simplex virus. Virology 22:1-8.
16. Roizman, B., and P. G. Spear. 1971. Herpesvirus anti-gens on cell membranes detected by centrifugation ofmembrane-antibody complexes. Science 171:298-300.
17. Ruyechan, W. T., L. S. Morse, D. M. Knipe, and B.Roizman. 1979. Molecular genetics of herpes simplexvirus. II. Mapping of the major viral glycoproteins andof the genetic loci specifying the social behavior ofinfected cells. J. Virol. 29:677-697.
18. Schlehofer, J. R., K. 0. Habermehl, W. Diefenthal,and H. Hampl. 1979. Reduction of 51Cr-permeabilityof tissue culture cells by infection with herpes simplextype 1. Intervirology 11:158-166.
19. Shore, S. L., C. M. Black, F. M. Melewicz, P. A. Wood,and A. J. Nahmias. 1976. Antibody-dependent cell-mediated cytotoxicity to target cells infected with type1 and type 2 herpes simplex virus. J. Immunol. 116:194-201.
20. Spear, P. G. 1976. Membrane proteins specified by herpessimplex virues I. Identification of four glycoproteinprecursors and their products in type 1-infected cells. J.Virol. 17:991-1008.
21. Spear, P. G., J. M. Keller, and B. Roizman. 1970.Proteins specified by herpes simplex virus. II. Viralglycoproteins associated with cellular membranes. J.Virol. 5:123-131.
22. Subak-Sharpe, J. H., J. M. Brown, D. A. Ritchie, M.C. Thmbury, J. C. M. Macnab, H. S. Marsden, andJ. H. Hay. 1974. Genetic and biochemical studies withherpesvirus. Cold Spring Harbor Symp. Quant. Biol.39:717-730.
23. Subramanian, T., and W. E. Rawls. 1977. Comparisonofantibody-dependent cellular cytotoxicity and comple-ment-dependent antibody lysis of herpes simplex virus-infected cells as methods of detecting antiviral antibod-ies in human sera. J. Clin. Microbiol. 5:551-558.
24. Vestergaard, B. F. 1973. Crossed immunoelectrophoreticcharacterization of herpesvirus hominis type 1 and 2antigens. Acta Pathol. Microbiol. Scand. Sect. B 81:808-810.
25. Vestergaard, B. F., 0. J. Bjerrum, B. Norrild, and P.C. Grauballe. 1977. Crossed immunoelectrophoreticstudies of the solubility and immunogenicity of herpessimplex virus antigens. J. Virol. 24:82-90.
26. Vestergaard, B. F., and B. NonRild. 1978. Crossed im-munoelectrophoresis of a herpes simplex virus type 1-specific antigen: immunological and biochemical char-acterization. J. Infect. Dis. 138:639-643.
27. Vestergaard, B. F., and B. Norrild. 1979. Crossed im-munoelectrophoretic analysis and viral neutralizing ac-tivity of five monospecific antisera against five differentherpes simplex virus glycoproteins. Boston Symposium1977. IARC (Int. Agency Res. Cancer) Sci. Publ. 24:225-234.
on Septem
ber 17, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from





























