Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 76, No. 4, pp. 1633-1637, April 1979Biochemistry
Avian acute leukemia viruses MC29 and MH2 share specific RNAsequences: Evidence for a second class of transforming genes
(gel electrophoresis of RNA/RNA-cDNA hybridization/fingerprinting of oligonucleotides/group-specific and specificsequences of defective viral RNA)
PETER H. DUESBERG* AND PETER K. VOGTt*Department of Molecular Biology, University of California, Berkeley, California 94720; and tDepartment of Microbiology, University of Southern California,Los Angeles, California 90007
Communicated by Heinz Fraenkel-Conrat, January 11, 1979
ABSTRACT The genome of the defective avian tumor virusMH2 was identified as a RNA of 5.7 kilobases by its presencein different MH2-helper virus complexes and its absence frompure helper virus, by its unique fingerprint pattern of RNaseTi-resistant (Ti) oligonucleotides that differed from those of twohelper virus RNAs, and by its structural analogy to the RNA ofMC29, another avian acute leukemia virus. Two sets of se-quences were distinguished in MM2 RNA: 66% hybridized withDNA complementary to helper-independent avian tumor vi-ruses, termed group-specific, and 34% were specific. The per-centage of specific sequences is considered a minimal estimatebecause the MH2 RNA used was about 30% contaminated byhelper virus RNA. No sequences related to the transforming srcgene of avian sarcoma viruses were found in MH. MH2 sharedthree large Ti oligonucleotides with MC29, two of which couldalso be isolated from a RNase A- and Ti-resistant hybrid formedbetween MH2 RNA and MC29 specific cDNA. These oligonu-cleotides belong to a group of six that define the specific seg-ment of MC29 RNA described previously. The group-specificsequences of MH2 and MC29 RNA shared only the two smallestout of about 20 T1 oligonucleotides associated with MH2 RNA.It is concluded that the specific sequences of MH2 and MC29are related, and it is proposed that they are necessary for, oridentical with, the onc genes of these viruses. These sequenceswould define a related class of transforming genes in aviantumor viruses that differs from the src genes of avian sarcomaviruses.
MC29 and MH2 are defective avian RNA tumor viruses thatcause acute leukemias and carcinomas in the animal and canalso transform fibroblasts in tissue culture (1-6). The trans-forming or onc (7) genes of these two viruses have not yet beendefined biochemically or genetically.
However, the genome of MC29, which is 5.7 kilobases (kb),was shown to contain a specific sequence of about 1.5 kb thatis unrelated to the replicative genes of avian tumor viruses andis also unrelated to the conserved src genes, which are the oncgenes of avian sarcoma viruses (8, 9). This sequence maps be-tween 2.5 and 4 kb from the 3' end of MC29 RNA and codestogether with other sequences of viral RNA for a MC29-spe-cific, nonstructural protein of 120,000 daltons (9) that is alsoexpressed in transformed cells (4). Therefore, it was suggestedthat this sequence is a candidate for the onc gene of MC29(9).A definitive identification of the onc genes of MC29 and
MH2 is complicated by the defectiveness of these viruses in allthree replicative genes of avian tumor viruses (3-5, 10, 11). Itis impractical to detect onc deletion mutants, a capability thatproved essential for the definition of the src gene of Rous sar-coma virus (RSV) (12-16), because onc deletions of MH2 orMC29 would lack detectable biological markers of any viralgene. A recombination analysis, which proved useful in defining
the src gene of RSV (17, 18), is also complicated by the func-tional defectiveness of MC29 and MH2 in all three replicativegenes. Recombinants would lack readily detectable markers,and specific sequences such as those found in MC29 (9) wouldnot be expected to recombine with other avian tumor viruseslacking them (19).
Because the oncogenic spectra of MC29 and MH2 are closelyrelated (1-6, 10, 11), it was reasoned that nucleotide sequencescoding for their related oncogenic functions might also besimilar. Further, it was hypothesized that such sequences couldbe identified, if other viral RNA sequences not directly involvedin oncogenicity were sufficiently different in both viruses.Dissimilarity in viral sequences not directly involved in onco-genic function may be expected, because MH2 was isolated inEngland in 1927 (2) and MC29 in Bulgaria in 1965 (1). It is thepurpose of this investigation to determine if MH2, like MC29and avian sarcoma viruses, contains specific sequences, definedas being unrelated to replicative genes, and if they are relatedto the specific sequences of MC29. This would be a first step inthe genetic definition of the onc genes of this class of viruses.Preliminary results of this work have been published (20-23).
METhODS AND MATERIALSMH2 pseudotypes formed with Rous-associated virus 7 ofsubgroup C (RAV-7) or with MH2-associated viruses ofsubgroups A and C (MH2AV-A and C) were propagated inchicken or quail fibroblasts or in chicken macrophages. Culturesof yolk sac macrophages were prepared as described (24). Al-ternatively, macrophage cultures were obtained as rapidlygrowing nonadhering or readily detachable cells that appearedin secondary chicken embryo cell cultures 3-5 days after in-fection with MH2. Macrophages were grown in medium 199supplemented with 10% tryptose phosphate broth, 8% calfserum, and 2% chicken serum.
Preparation of [3H]- or [14C]uridine-labeled virus and ex-traction of viral RNA followed published procedures (8, 25).32P-Labeled virus was prepared as follows: 5-10 X 107 mac-rophages were suspended in 45 ml of phosphate-free medium199 supplemented with phosphate-free calf (3%) and chicken(1%) sera and 150 mCi (1 Ci = 3.7 X 10W becquerels) of32PO43- and incubated in three Corning tissue culture flasks(150 cm2) at 380C. Maximal uptake of 32P by cells was about50% and was reached between 2 and 4 hr, compared to 80-90%in RSV-infected fibroblast cultures (unpublished). After 8-hrincubation, cells were centrifuged and resuspended in completemedium, which was changed at 12-hr intervals. Media collectedduring 2 days were combined for virus purification, which was
The publication costs of this article were defrayed in part by pagecharge payment. This article must therefore be hereby marked "ad-vertisement" in accordance with 18 U. S. C. §1734 solely to indicatethis fact.
1633
Abbreviations: kb, kilobase(s); RSV, Rous sarcoma virus; RAV, Rous-associated virus; RPV, ringneck pheasant helper virus; PR-B, PragueRSV of subgroup B; td, transformation-defective.

1634 Biochemistry: Duesberg and Vogt
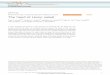
as described (8, 16, 25), except that virus was concentrated bycentrifugation for 2 hr at 19,000 rpm and 20'C in a Beckman19 rotor rather than by ammonium sulfate precipitation. ViralRNA was isolated and sedimented as described (8). Approxi-mately 10 X 106 cpm of 50-70S viral RNA was obtained, cor-responding to 60,000 cpm per mCi of 32PO43- [compared toabout 1 X 106 cpm per mCi of 32P obtained in RSV RNA fromvirus propagated in fibroblasts (16)]. About 0.9 X 106 cpm ofelectrophoretically pure 28S RNA was obtained from 10 X 106cpm of 50-70S MH2(RAV-7) RNA (Fig. 1A and ref. 8).
RESULTSThe Genome of MH2 Is a 28S RNA. The RNA of MH2
pseudotypes was isolated from purified virus as a 50-70S RNAcomplex (8, 9). After heat dissociation of the RNA complex, twoRNA monomer species were resolved by electrophoresis inpolyacrylamide gels (Fig. 1A). One of these coelectrophoresedwith 34S RNA of ringneck pheasant helper virus (RPV) ofMC29, the other with 28S MC29 RNA (8). Analogy with theRNAs of MC29 and RPV suggests that the 34S RNA was RAV-7helper virus RNA and the 28S RNA was the genome of MH2.The following evidence supported this hypothesis: the 28S RNAspecies of MH2(RAV-7) or MH2(MH2AV) was not found in theRNAs of pure RAV-7 or MH2AV (not shown). The fingerprintpatterns of T1 oligonucleotides of 28S RNA species ofMH2(RAV-7) and MH2(MH2AV) were closely related to eachother (Fig. 1; refs. 22 and 23), but different from those of RAV-7and MH2AV RNAs (Fig. 1). The electrophoretic mobility andthe fingerprint patterns of Ti oligonucleotides of the 34S RNAspecies of MH2(RAV-7) were the same as those of authenticRAV-7 RNA. We conclude that the 28S RNA species ofMH2(RAV-7) is the genome of MH2 and that the 34S RNA isthe genome of RAV-7. On the basis of their electrophoreticmobilities relative to the known MC29(RPV) RNA standards,
the 34S RAV-7RNA was estimated to measure 8.5 kb and the28S MH2 RNA to measure 5.7 kb (8, 9). More detailed sizemeasurements of these RNAs have confirmed these estimates(unpublished).The ratio of 28S MH2 to 34S helper virus (RAV-7 or
MH2AV) RNA varied with virus from different cultures ofinfected cells. Virus from about 30 different transformedchicken or quail fibroblast cultures examined contained 34S to28S RNA at molar ratios of >5:1, while this ratio was between0.5:1 and 2:1 in virus from about 10 cultures of chicken mac-rophages. Therefore, virus produced by macrophages was usedfor biochemical analyses of 28S RNA, although cell viabilityand virus production were poor, particularly in the phos-phate-free cell culture medium used to radiolabel viral RNA.These difficulties have limited our analyses of MH2 RNA.
In addition, 28S MH2 RNA prepared electrophoretically (Fig1A) was always contaminated to various degrees with degradedhelper virus RNA of the same electrophoretic mobility. Thepurity of MH2 RNA was estimated to be 60-70% for a typicalpreparation of 28S RNA (Fig 1A). This estimate was obtainedby drawing a hypothetical base line of degraded 34S RNAacross the peak of 28S RNA (see broken line Fig. 1A). The areaabove this line was assumed to represent the fraction of pure28S MH2 RNA in the peak. It was assumed that both 34S and28S RNAs have the same distribution of intact and degradedspecies.
Sequence Relationship between MH2 and MC29 andOther Avian Tumor Viruses Measured by Hybridization. Totest if 28S MH2 RNA contained specific sequences, defined asbeing unrelated to helper-independent avian tumor virus RNAs,we have hybridized MH2 RNA with DNAs complementary(cDNA) to Prague RSV of subgroup B (PR-B), which containsthe src gene of RSV (16, 17), to a transformation-defective (td)src deletion of PR-B, to a combination of PR-B and td PR-B,
10 w IV11 A'--
q. 191JO95) a0l040
- ,;I 1r3o
2#3 4
eI
4
0l0 20 30 40Distance Moved (mm) Electrophoresis
FIG. 1. Identification of the RNA species of MH2(RAV-7) (A) and fingerprint patterns of the RNase Tl-resistant oligonucleotides of 34SRAV-7 RNA (B) and 28S MH2 RNA ofan MH2(RAV-7) pseudotype (C), ofMH2RNA isolated from a RNase-resistantMH2 RNA-MC29 cDNAhybrid (D), and ofMH2AV RNA (E). (A) The 50-70S RNA complex ofMH2(RAV-7) propagated in chicken macrophages was mixed with 50-70SRNA of MC29(RPV) propagated in a quail cell line (4) to provide 34S RPV RNA and 28S MC29 RNA standards (8). After heat dissociation,the RNA monomer species were subjected to electrophoresis in 2% polyacrylamide gel. Preparation of viral RNA is described in Methods andMaterials. Electrophoresis followed published procedures (8, 12, 17). The broken line is a hypothetical base line of degraded 34S RNA underthe peak of 28S RNA (see text). (B) The 34S RAV-7 [32PIRNA component ofMH2(RAV-7) was eluted from a polyacrylamide gel like that shownin A and after exhaustive digestion with RNase T1 was subjected to two-dimensional fingerprint analysis and autoradiographed (8,9, 16). Numbersrefer to large T1 oligonucleotides whose RNase A-resistant fragments were determined (Table 3). (C) Fingerprint analysis ofthe T1 oligonucleotidesof the 28S MH2 [32PJRNA component prepared electrophoretically as shown in A. The RNase-A resistant fragments of large T1 oligonucleotidesare reported in Table 2. Oligonucleotides, for which homologous counterparts were found in RAV-7 RNA (Table 2) are parenthesized. The numberof G-terminating constituents of multiple oligonucleotide spots is indicated alphabetically. (D) The T1 oligonucleotides of a RNase A and T1-resistant hybrid formed between 288 MH2 [32P]RNA and MC29-specific cDNA. MC29-specific cDNA was made by hybridizing MC29(RPV)cDNA to an excess ofRPV and PR-B (RSV) RNA. MC29-specific cDNA was then hybridized to 28S MH2 [32PJRNA [devoid of poly(A)-taggedRNA fragments that were used for oligonucleotide mapping, see text]. Hybridization was in 50% formamide and 0.5M NaCl as detailed (8,9,22). After digestion with RNases A and T1 in 0.3 M NaCl, the hybrid was purified by P100 Bio-Gel (BioRad) chromatography and phenol ex-traction; subsequently it was heat dissociated and the RNA was digested with RNase Ti (9, 22, 30). The two large T1 oligonucleotides 4a and9 were identified by their chromatographic locations and their RNase-A-resistant fragments (Table 2). (E) The 60-70S [32P]RNA ofMH2AV-Aand C propagated in chicken embryo fibroblasts was fingerprinted as for A.
I0x
I
0
a:
aE
00
4
3%2of
Proc. Natl. Acad. Sci. USA 76 (1979)

Proc. Natl. Acad. Sci. USA 76 (1979) 1635
and to MH2AV. Maximal levels of hybridizations betweenhomologous RNA and cDNAs were about 90% under OutrO-frditions (Table 1). About 60%, normalized 66%, of 28S RNA washybridized by each of these cDNAs at the plateau level ofmaximal hybridization reached by varying cDNA-to-RNAratios (Table 1). As with MC29 RNA, these sequences weretermed group-specific (9). Hence about 34% of MH2 RNAappeared to be MH2-specific. We conclude MH2 RNA is or-ganized like MC29 RNA, of which 60% is group-specific and40% specific (8, 9, 22). The slightly lower percentage of specificsequences in MH2 RNA compared to MC29 RNA is thoughtto reflect a higher degree of contamination by degraded helperviral RNA, which would increase the percentage of group-specific sequences. Further, the result that approximately thesame percentage of 28S MH2 RNA was hybridized by PR-BcDNA, td PR-B cDNA, and a combination of the two cDNAs(Table 1) indicates the absence of sarc gene-related sequencesin 28S MH2 RNA. However, it may be argued that, due tounequal representation of sequences in cDNA (26), the PR-BcDNA used here did not contain sufficient DNA complemen-tary to the sarc gene to detect src sequences in MH2 RNA. It mayalso be argued that contamination of 28S MH2 RNA by de-graded RAV-7 RNA would render our test too insensitive todetect src sequences. It is shown in Table 1 that the PR-B cDNAused detected the expected amount of src-specific sequencesof PR-B RNA-i.e., 12% that were not hybridized by td PR-BcDNA (15). Because the src gene measures about 1.5 kb (15-18),it would, if present, represent 26% of the 28S MH2 RNA, which
Table 1. Hybridizationa of viral RNAs with viral cDNAsb
% RNA hybridized at variouscDNA/RNA ratios, mean 1: SD
RNA cDNA 5:1 20:1 50:1 100:1
28S MH2C PR-B 60.5 ± 1 62tdPR-B 58I5 59.5PR-B +
tdPr-Bd 62 + 3MC29(RPV) 61.5 + 5 64 4- 1MC29(RPV) + 671 3 71 6PR-Bed
28S MH2f MH2AV-A 58:I5 60i 2and C
28S MC299 MC29(RPV) 65 6 84 + 4 88 + 1 93i 7
MH2AV-A MH2AV-A 75 + 4 96and Ch and C
PR-Be PR-B 93 + 0 93 + 1td PR-B 81 81
a Each reaction mixture contained about 1 ng of [32P]RNA (2000cpm/ng) and 5-100 ng of [3H]cDNA (50 cpm/ng). Hybridizationswere in 4-5,ul of70% (wt/vol) deionized formamide/0.3M NaCl/0.03M Na citrate/15mM Na phosphate (pH 7.0)/0.05% sodium dodecylsulfate at 400C for 12 hr. Percentage nuclease resistance is expressedas the radioactivity recovered in aliquots digested with nucleaserelative to that found in undigested aliquots. Each value is the meanof two or three experiments using, in some cases, independentpreparations of RNA and cDNA. Digestion was with RNases A (5Ag/ml), T1 (10 units/ml), and T2 (10 units/ml) for 30 min at 40°Cin 0.3 M NaCl/0.03 M Na citrate (pH 7.0). The background of nu-clease resistance of an aliquot heated at 1000C in 0.01 M Na+ was<0.5%.
b cDNA was prepared as described (8, 9).c Prepared from MH2(RAV-7) propagated in macrophages as in Fig.1A.
d Each cDNA was present at the DNA-to-RNA ratio indicated.e Shown electrophoretically to be free of td PR-B RNA (16, 25).f Prepared from MH2(MH2AV-A and C) as for c.gAs in ref. 8.h Sixty-70S RNA from MH2AV-A and C was used.
was estimated at 5.7 kb. Therefore, our assay would have de-tectedire -in MH2 RNA even if the RNA were 50% contami-nated by helper RAV-7 RNA.
Further experiments designed to detect a relationship be-tween the specific sequences of MH2 and MC29 indicated that64%, normalized 70%, of MH2 RNA was hybridized byMC29(RPV) cDNA and 71%, normalized 78%, by a combina-tion of MC29(RPV) and PR-B cDNAs (Table 1). One inter-pretation of this experiment is that Pr-B cDNA hybridizes withgroup-specific sequences of MH2 RNA not represented inMC29(RPV) cDNA and that MC29-specific cDNA hybridizeswith related MH2-specific sequences and consequently raisesthe level of maximal hybridization to 78%. An alternative in-terpretation of the 71% hybridization of MH2 RNA describedabove is that the group-specific sequences of MH2 and MC29are more closely related to each other than to any of the otherviruses tested and that no hybridization of specific sequenceswas observed.
Because none of the viruses (including MH2AV; see Fig. 1,Table 1) used here to prepare cDNAs are known to be congenicpredecessors of MH2, specific RNA sequences defined by theirfailure to hybridize with cDNAs of helper-independent virusesmay include two different classes of sequences: one, repre-senting the sum of minor strain-specific elements, scatteredbetween largely group-specific sequences; another may rep-resent a contiguous MH2-specific sequence totally unrelatedto group-specific elements. Hybridization cannot by itselfdistinguish between these alternatives.MH2 Shares Specific Oligonucleotides with MC29 That
Are Not Related to Replicative Genes of Avian Tumor Vi-ruses. To determine directly if the specific sequences of MH2and MC29 are related and to estimate the extent of their rela-tionship, we have investigated if the two viral RNAs share largeTi oligonucleotides. Shared Tl-oligonucleotides provide sen-sitive signals of sequence identity among related RNAs, becausea single base change may eliminate a common or generate aspecific oligonucleotide without detectably affecting theirhomology measured by hydridization. In addition, the com-plexity of large oligonucleotides renders fortuitous coincidencesimprobable.
About 20 large oligonucleotides were associated with 28SMH2 RNA and about 20 also with 34S RAV-7 (Fig. 1, Tables2 and 3). Of the 20 oligonucleotides associated with 28S MH2RNA, 10 were shared with RAV-7 RNA (parenthesized in Fig.1C and Table 2). Five of these 10 were also shared with ho-mologs of MH2AV RNA, including the 5' terminal cap IV oh-gonucleotide (19) and two to three of these with PR-B RNA(Tables 2, 3, and 4). After oligonucleotides shared with helpervirus RNA had been subtracted, at least 11 of the remainingoligonucleotides associated with 28S MH2 RNA, including twomultiple oligonucleotide spots, were specific for 28S MH2 RNAand were therefore termed MH2 oligonucleotides. None ofthese was identical to one of the conserved src gene oligonu-cleotides of RSV (16-18), confirming the notion that MH2 lackssequences related to the src gene.The oligonucleotides associated with 28S MH2 RNA were
not present at equimolar ratios (Fig. IC). This may be explainedif one considers that they represent three nonequimolar poolsof RNA sequences deriving from: 28S MH2 RNA, contami-nating helper virus RNA, and sequences shared by MH2 andhelper virus RNA. Further work using MH2 RNA of higherpurity or from a different pseudotype will be necessary to de-termine which of the oligonucleotides shared with RAV-7 areintegral parts of 28S MH2 RNA and which reflect contami-nating RAV-7 RNA.
Four of the 11 MH2-oligonucleotides had homologouscounterparts [defined as chromatographically and composi-
Biochemistry: Duesberg and Vogt

1636 Biochemistry: Duesberg and Vogt
Table 2. T1 oligonucleotides of 28S MH2 RNAa and homologsin other viruses
28S RNase A digestion MC- RAV- MH2AV-A PR-MH2b products 29c 7c and Cc B
1 U, 2 C, G,5AU,AAU,AAAC
(2) 9U,7C,2AC,2AU, 1 4AG, AAC, AAU
3 4U,5C,G,3AU,2 AAU, AAAU
4ad 2U,3C,G, 12 AC, AU, 2 AAC
4bd U,6C,G,2AC,2AU,AAC, AAAC
(5) 6U,9C,2AC,AU,AAC, 2 2 4AAG
6ae 3U,8C,G,AC,AU,AAC, A5N
(6b)e 6 U, 8 C, G, AC, AU, 3 3 6AAAC
(7) 5U,5C,G,2AC,3AU 4 6(8) 4 U, 5 C, AC, AAU, 5
AAAC, AAAG9 3 U, 10 C, AC, AAG 7b10 2 U, 3 C, AC, AG, AAU, 8bf
AAAC11a,b,c 10 U, 15 C, G, 3 AC, 9? 8?
5 AU, AG, AAG,AAAU, AAAG, AnN
12 2U,4C,2ACAU,AG13 U, 3 C, 2 AC, AAG,
AAAU(14) 4 C, 2 AC, AU, A4G 1515a,b 2 U, 4 C, G, 2 AC, AU, 11? 14?
AG, AAC, A4N(17) 2U,4C,G,2AC,AU 11(19) 10 U, 7 C, G, AU 8(20) 6U,2C,G,3AU,AAU 7269 5 C, 2 AC, AG 26(Cap) m7GpppGmC, 2U, C, cap IV cap IV cap IV
AU, G (Class IV)a MH2 RNA of 28S was eluted from a polyacrylamide gel as in Fig.1A and fingerprinted (8). Oligonucleotides were prepared fromfingerprints shown in Fig. 1 and their RNase A-resistant fragmentswere determined as described (16, 17). Some, but not all, oligonu-cleotide numbers are the same as those used previously for morepreliminary analyses ofMH2 RNA (20-23). Because of the scarcityofMH2 [32P]RNA, some numbers are semiquantitative, combiningscintillation counting of RNase A-resistant fragments and reflectingvisual estimates from autoradiograms.
b Numbers as in Fig. 1C. Oligonucleotides with homologous coun-terparts in RAV-7 or MH2AV-A and C helper virus RNAs are par-enthesized (see text).
c Number denotations of homologous oligonucleotides found in 28SMC29 RNA (8,9), RAV-7 RNA (Fig. 1 and Table 3), and MH2AV-Aand C RNA (Fig. 1 and Table 4).
d The composition of unique T1 oligonucleotides in 4a,b was deter-mined by subtracting 4a, identified as in Fig. 1D, from 4a,b.
e The composition of unique T1 oligonucleotides of 6a,b was deter-mined from a fingerprint (not shown) in which 6a and 6b were sep-arated.
f No. 10 ofMH2 and 8b ofMC29 are closely related but may differ inU or C (see text).
g No. 26 has recently been identified as an MC29 oligonucleotide thatmaps at the 5' border of the MC29-specific map segment (unpub-lished) and its composition has been revised from that describedin ref. 8.
tionally indistinguishable but possibly differing in sequence(27)] in 28S MC29 RNA (Table 2). The three larger ones (nos.4a, 9, and 10) belong to a group of six MC29 oligonucleotides
Table 3. T1 oligoriucleotides of RAV-7 RNAa and PR-Bhomologs
RAV-7b RNase A digestion products PR-Bc
1 7U,4C,2AC,2AU,AG,AAC,AAU2 4U,7C,2AC,AU,AAC,AAG 43 6U,8C,G,AC,AU,AAAC 64 5U,5C,G,2AC,3AU5 4U,5C,AC,AAU,AAAC,AnN6 6U,6C,3AC,AU,AAG,AAAN7 6U,2C,G,3AU,AAU8 10 U, 7 C, G, AU9 3U,G,AC,2AU10 4U,C,AC,AU,AnG11 2U,4C,G,2AC,AU12 2U,4C,G,2AAU,AAAN13 2U,4C,G,2AC,AU,AAC 1414a,b 3 U, 5 C, G, 3 AC, AU, AG, AAC, AAAN15 4C,2AC, AU, AnG16 U,2C,2AC,AAG17 2C,AU,AG,AnN18 4U,2C,G,AU,AAU19 2 U, C, G, 3AU,AAU20 2U,4C,G,AC,2AU,AACCap T1 Class IV
a As in Table 2, but sufficient [32P]RNA was used for a quantitationof each RNase A-resistant fragment.
b Numbers refer to oligonucleotide spots shown in Fig. 1B.c Oligonucleotides of the same composition were found in PR-B (16).
that define the specific segment of MC29 RNA (9). The fourthand least complex one (no. 26) maps near the 5' border of thespecific segment of MC29 RNA (unpublished) and is group-specific, because it is also found in RPV RNA (i.e., no. 23, ref.8). In addition, the two RNAs shared their class IV cap oligo-nucleotides and possibly constituents of the multiple MH2oligonucleotide spots lla,b,c and 15a,b (Table 2).To measure more directly homology between the specific
sequences of MH2 and MC29, the oligonucleotides of MH2[32P]RNA isolated from a hybrid formed with MC29-specificcDNA (9) were analyzed (28). The MC29-specific DNA used
Table 4. T1 oligonucleotides of MH2AV-A and C RNAa andPR-B homologs
MH2AV-A and Cb RNase A digestion products PR-BC
1 7U,10C,G,3AC,AAC2 5 U, 10 C, 2 AC, AU, AAC, AAG 43 6U,8C,G,AC,AU,AAAC 64 8U,6C,2AC,2AU,AG,AAC,AAU5 3U,6C,G,2AC,AU6 6U,6C,G,2AC,4AU7a,b 4U,3C,2AC,2AU,AG,AAU,
AAAN, A4N8 6U,6C,AAG,A5N 89 6U,9C,G,AAC10 8U,3C,4AU,AG,AAU11 6U,2C,G,4AU,AAC12 U,2C,G,AC,2AAAN13 4U,3C,G,2AU,AAAN14 5 U, 4 C, G, AC, AAAC, AAAN15 4U,4C,AC,AAAC,A4G16 2U,5C,G,2AC,AU,AAC 14Cap T1 Class IV
a As in Table 3.b Numbers refer to oligonucleotide spots shown in Fig. 1E.c Oligonucleotides of the same composition were found in PR-B (16).
Proc. Nati. Acad. Sci. USA 76 (1979)

Proc. Natl. Acad. Sci. USA 76 (1979) 1637
was prepared by hybridization of MC29(RPV) cDNA to a largeexcess of RPV and PR-B RNA (9). Thus, group-specific se-quences, including small strain-specific elements of group-specific sequences, of MC29 cDNA were heteroduplexed withunlabeled RPV and PR-B RNAs and could not hybridize withMH2 [32P]RNA (9, 22, 28). The MH2 RNA-MC29 specificcDNA hybrid was treated with RNases A and Ti in order todigest unhybridized and partially hybridized RNA sequencesat U, C, and G residues, then purified by gel chromatographyand phenol extraction (9, 28). After heat dissociation of thehybrid, the RNA was digested with RNase Ti and the digestwas subjected to fingerprint analysis. Two large MH2 oligo-nucleotides (nos. 4a and 9) and one less complex (no. 26) MH2oligonucleotide were recovered and identified by their chro-matographic location (Fig. 1D) and by their RNase A-resistantfragments (Table 2). We deduce that MH2 oligonucleotides 4aand 9, have identical counterparts in the specific segment ofMC29 (nos. 1 and 7b). MH2 oligonucleotide no. 10 was not re-covered from the hybrid, although it has a homolog in MC29RNA (Table 2). Hence it either differs slightly from its MC29counterpart (no. 8b) in a U or C residue or it has an identicalcounterpart that may have been lost from the hybrid becausemismatched neighboring sequences were digested by RNase(28). It is concluded that the specific sequences of MC29 andMH2 share three identical or closely related large Ti oligonu-cleotides and that their group-specific sequences share only thetwo smnallest out of 20 T1 oligonucleotides associated with MH2RNA (no. 26 and cap IV). The multiple oligonucleotide spotsof MH2 (i.e., ila,b,c and 15a,b) may contain further specificor group-specific oligonucleotides.
DISCUSSIONOur data indicate that MH2, like MC29, has a 28S RNA ge-nome; 66% of it is sequence-related to the RNA of helper-in-dependent members of the avian tumor virus group and termedgroup-specific, and 34% is MH2-specific. About 78% of MH2RNA, 12% more than its group-specific portion, was hybridizedby a combination of MC29 cDNA and cDNAs of otherhelper-independent avian tumor viruses, suggesting that thespecific sequences of MH2 and MC29 may be related. This wasdirectly proven, because MH2 shared with MC29 three (twoidentical and one identical or closely related) of six largeoligonucleotides that define the specific segment of MC29RNA. Further work using RNA cDNA hybridization betweenthe specific sequences of MH2 and MC29 would be necessaryto quantitate their relationship. Preliminary data indicate thatthe four MH2 oligonucleotides shared with MC29 map between1.5 and 5.7 kb from the 3' end of the RNA, as do their coun-terparts in MC29 (9).
Because the large MH2 oligonucleotides shared with MC29have not been found in any other avian tumor virus studied(Tables 2, 3, and 4; refs. 16-19) and are not related by hybrid-ization to replicative genes of helper-independent avian tumorviruses, it would appear that these oligonucleotides identify aspecific class of closely related sequences shared by MH2 andMC29. Moreover, because the group-specific sequences ofMH2and MC29 are different by the same criteria, we propose thatthe nucleotide sequences shared by the two viruses are necessaryfor their related oncogenic functions and define a class of oncgenes that differs from the src genes of avian sarcoma viruses(12-18). This suggestion does not rule out that group-specificsequences are also involved in transformation. It is consistentwith our previous proposal that the specific sequences of MC29code together with some group-specific sequences for a non-structural protein of 120,000 daltons that is also found in
transformed cells (4) and may function as a transforming pro-tein (9). An analogous protein has also been found in cellstransformed by MH2 (5). According to our present data, theseproteins ought to be related in their specific sequences.Note Added in Proof. A third avian acute leukemia virus, CMII, alsoshares specific sequences with MC29 and MH2 (K. Bister, H. C.LUliger, and P. H. Duesberg, unpublished data).
We thank Pamela Mellon for providing MC29-specific cDNA andfor critical discussions, L. Evans and K. Bister for review of the man-uscript, and Lorrine Chao for assistance with these experiments. Wealso gratefully acknowledge J. M. Bishop and D. Sheiness for com-municating to us their independent experiments which indicated thatspecific sequences of MC29 and MH2 are related. This work wassupported by National Institutes of Health Grants CA 11426 and CA19725.
1. Ivanov, X., Mladenov, Z., Nedyalhov, S. & Bozhkov, S. (1965)Dokl. BuWg. Akad. Nauk. 18,593-595.
2. Begg, A. M. (1927) Lancet i, 912-915.3. Ishizaki, R., Langlois, A. J., Chabot, J. & Beard, J. W. (1971) J.
Virol. 8, 821-827.4. Bister, K., Hayman, M. & Vogt, P. K. (1977) Virology 82,
431-448.5. Hu, S. S. F., Moscovici, C. & Vogt, P. K. (1978) Virology 89,
162-178.6. Moscovici, C., Gazzolo, L. & Moscovici, M. G. (1978) Micro-
blologica 1, 1-13.7. Baltimore, D. (1975) Cold Spring Harbor Symp. Quant. Biol.
39, 1187-1200.8. Duesberg, P. H., Bister, K. & Vogt, P. K. (1977) Proc. Nat!. Acad.
Sci. USA 74,4320-4324.9. Mellon, P., Pawson, A., Bister, K., Martin, G. S. & Duesberg, P.
H. (1978) Proc. Natl. Acad. Sci. USA 75,5874-5878.10. Bister, K. & Vogt, P. K. (1978) Virology 88,213-221.11. Hu, S. S. F. & Vogt, P. K. (1979) Virology, in press.12. Duesberg, P. H. & Vogt, P. K. (1970) Proc. Natl. Acad. Sci. USA
67, 1675-1680.13. Vogt, P. K. (1971) Virology 46,939-946.14. Martin, G. S. & Duesberg, P. H. (1972) Virology 47,494-497.15. Lai, M. M. C., Duesberg, P. H., Horst, J. & Vogt, P. K. (1973) Proc.
Natl. Acad. Sci. USA 70,2266-2270.16. Wang, L. H., Duesberg, P. H., Beemon, K. & Vogt, P. K. (1975)
J. Virol. 16, 1051-1070.17. Wang, L. H., Duesberg, P. H., Mellon, P. & Vogt, P. K. (1976)
Proc. Natl. Acad. Sci. USA 73,1073-1077.18. Joho, R. H., Billeter, M. A. & Weissmann, C. (1975) Proc. Nat!.
Acad. Sci. USA 72,4772-4776.19. Wang, L. H., Duesberg, P. H., Robins, T., Yokota, H. & Vogt, P.
K. (1977) Virology 82,472-492.20. Duesberg, P. H., Vogt, P. K., Bister, K., Troxler, D. & Scolnick,
E. M. (1978) in Avian RNA Tumor Viruses, eds. Barlati, S. & deGiuli-Morghen, C. (Piccin, Padua, Italy), pp. 95-111.
21. Duesberg, P. H., Vogt, P. K., Bister, K., Troxler, D. & Scolnick,E. M. (1979) in Oncogenic Viruses and Host Cell Genes, OjiInternational Seminar on Friend Virus and Friend Cells, ed.Ikawa, Y. (Academic, New York), in press.
22. Duesberg, P. H., Mellon, P., Pawson, A., Martin, G. S., Bister, K.& Vogt, P. K. (1978) in 4th ICN-UCLA Symposium on AnimalViruses, eds. Stevens, J., Todaro, G. & Fox, C. F. (Academic, NewYork), pp. 245-266.
23. Duesberg, P. H., Mellon, P., Pawson, A., Bister, K. & Vogt, P. K.(1979) in Modem Trends in Human Leukemia, Mls. Neth, R.& Mannweiler, K. (Springer, New York), Vol. 3, in press.
24. Moscovici, C., Gazzolo, L. & Moscovici, M. G. (1975) Virology68, 173-181.
25. Duesberg, P. H. & Vogt, P. K. (1973) Virology 54,207-219.26. Duesberg, P. H. & Canaani, E. (1970) Virology 42,783-788.27. Barrell, B. G. (1971) in Procedures in Nucleic Acid Research, eds.
Cantoni, S. L. & Davies, D. R. (Harper & Row, New York), Vol.2, pp. 751-795.
28. Coffin, J. M., Champion, M. & Chabot, F. (1978) J. Virol. 28,972-991.
Biochemistry: Duesberg and Vogt