Embed Size (px)
Citation preview

1270 NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999
ARTICLES
Cytotoxic T-lymphocyte (CTL) responses arise early after humanimmunodeficiency virus (HIV) or simian immunodeficiencyvirus (SIV) infection and are important in controlling viral repli-cation throughout the course of infection1–8. Despite vigorousCTL responses, most infected individuals eventually developAIDS. One hypothesis to explain the inability of the CTL re-sponse to contain HIV proposes that selective pressure by CTLsultimately results in the emergence of viral ‘escape’ variants. Insupport of this viral ‘escape’ hypothesis, genetic changes havebeen found in the virus populations in infected individuals thatreduce or eliminate CTL recognition in vitro9–18. However, the im-portance of CTL selection in the course of virus evolution in in-fected individuals remains controversial19. A chief criticism ofthe viral ‘escape’ hypothesis is the absence of data showing sta-tistically significant positive Darwinian selection within CTLepitopes in infected hosts19,20.
To explore the relationship between virus-specific CTL responsesand the evolution of SIV in vivo, we did a longitudinal study in fiveSIV-infected offspring from a family of major histocompatibilitycomplex (MHC)-defined rhesus macaques. Three of these animalsmade strong CTL responses to five different epitopes of the SIV Envand Nef proteins, thereby enabling us to follow the evolution oftheir virus populations in response to CTL selection. We also tested
the ability of CTL epitope variants to bind to their restricting MHCclass I molecules and to sensitize targets for CTL recognition.
Emergence of SIV Env and Nef CTL epitope variantsWe infected five members of a family of MHC-defined rhesusmacaques that were identical at their MHC class II DRB loci (Fig.
Virus-specific cytotoxic T-lymphocyte responses selectfor amino-acid variation in simian immunodeficiency
virus Env and Nef
DAVID T. EVANS1, DAVID H. O’CONNOR1, PEICHENG JING1, JOHN L. DZURIS2, JOHN SIDNEY2,JACK DA SILVA3, TODD M. ALLEN1, HELEN HORTON1, JOHN E. VENHAM1,
RICHARD A. RUDERSDORF4, THORSTEN VOGEL1, C. DAVID PAUZA1,5, RONALD E. BONTROP6, ROBERT DEMARS4, ALESSANDRO SETTE2, AUSTIN L. HUGHES3 & DAVID I. WATKINS1,5
1Wisconsin Regional Primate Research Center, University of Wisconsin, 1220 Capitol Court, Madison, Wisconsin 53715, USA
2Epimmune, 5820 Nancy Ridge Drive, San Diego, California 92121, USA3Institute of Molecular Evolutionary Genetics, The Pennsylvania State University,
University Park, Pennsylvania 16802, USA4Laboratory of Genetics, Genetics Building, University of Wisconsin, Madison, Wisconsin 53715, USA
5Department of Pathology and Laboratory Medicine, University of Wisconsin, Madison, Wisconsin 53715, USA6Biomedical Primate Research Centre-TNO, P. O. Box 5815, 2280 HV Rijswijk, The NetherlandsD.T.E. present address: New England Regional Primate Research Center, Harvard Medical School,
One Pine Hill Drive, Southborough, Massachusetts 01772, USAJ.D. present address: Department of Biology, East Carolina University, Greenville, North Carolina 27858, USA
D.T.E. and D.H.O. contributed equally to this study.Correspondence should be addressed to D.I.W.; email: [email protected]
Cytotoxic T-lymphocyte (CTL) responses to human immunodeficiency virus arise early after infec-tion, but ultimately fail to prevent progression to AIDS. Human immunodeficiency virus mayevade the CTL response by accumulating amino-acid replacements within CTL epitopes. We stud-ied 10 CTL epitopes during the course of simian immunodeficiency virus disease progression inthree related macaques. All 10 of these CTL epitopes accumulated amino-acid replacements andshowed evidence of positive selection by the time the macaques died. Many of the amino-acid re-placements in these epitopes reduced or eliminated major histocompatibility complex class Ibinding and/or CTL recognition. These findings strongly support the CTL ‘escape’ hypothesis.
Fig. 1 Pedigree of an MHC-defined family of rhesus macaques. Withinsymbols, MHC class I haplotypes for each macaque. Below, MHC class Imolecules that bind peptides from each macaque (in parentheses, haplo-type encoding these molecules). The MHC class II DRB molecules in all ofthe offspring of this family were identical.
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999 1271
ARTICLES
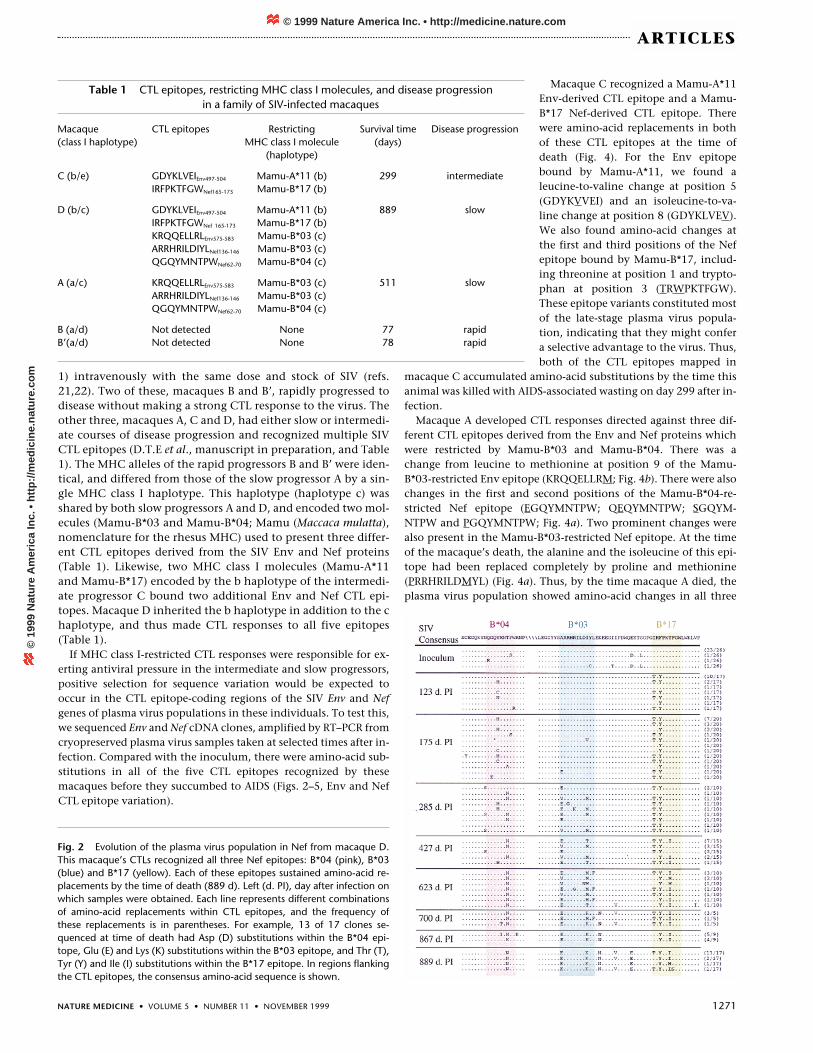
1) intravenously with the same dose and stock of SIV (refs.21,22). Two of these, macaques B and B’, rapidly progressed todisease without making a strong CTL response to the virus. Theother three, macaques A, C and D, had either slow or intermedi-ate courses of disease progression and recognized multiple SIVCTL epitopes (D.T.E et al., manuscript in preparation, and Table1). The MHC alleles of the rapid progressors B and B’ were iden-tical, and differed from those of the slow progressor A by a sin-gle MHC class I haplotype. This haplotype (haplotype c) wasshared by both slow progressors A and D, and encoded two mol-ecules (Mamu-B*03 and Mamu-B*04; Mamu (Maccaca mulatta),nomenclature for the rhesus MHC) used to present three differ-ent CTL epitopes derived from the SIV Env and Nef proteins(Table 1). Likewise, two MHC class I molecules (Mamu-A*11and Mamu-B*17) encoded by the b haplotype of the intermedi-ate progressor C bound two additional Env and Nef CTL epi-topes. Macaque D inherited the b haplotype in addition to the chaplotype, and thus made CTL responses to all five epitopes(Table 1).
If MHC class I-restricted CTL responses were responsible for ex-erting antiviral pressure in the intermediate and slow progressors,positive selection for sequence variation would be expected tooccur in the CTL epitope-coding regions of the SIV Env and Nefgenes of plasma virus populations in these individuals. To test this,we sequenced Env and Nef cDNA clones, amplified by RT–PCR fromcryopreserved plasma virus samples taken at selected times after in-fection. Compared with the inoculum, there were amino-acid sub-stitutions in all of the five CTL epitopes recognized by thesemacaques before they succumbed to AIDS (Figs. 2–5, Env and NefCTL epitope variation).
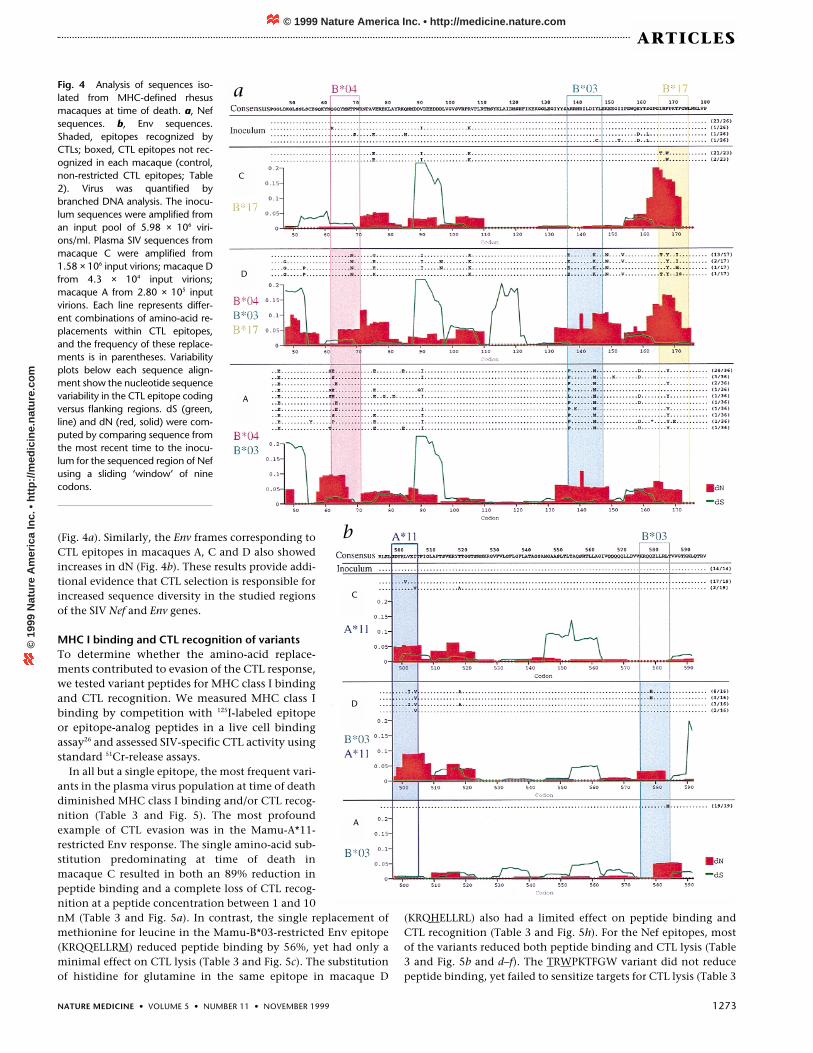
Macaque C recognized a Mamu-A*11Env-derived CTL epitope and a Mamu-B*17 Nef-derived CTL epitope. Therewere amino-acid replacements in bothof these CTL epitopes at the time ofdeath (Fig. 4). For the Env epitopebound by Mamu-A*11, we found aleucine-to-valine change at position 5(GDYKVVEI) and an isoleucine-to-va-line change at position 8 (GDYKLVEV).We also found amino-acid changes atthe first and third positions of the Nefepitope bound by Mamu-B*17, includ-ing threonine at position 1 and trypto-phan at position 3 (TRWPKTFGW).These epitope variants constituted mostof the late-stage plasma virus popula-tion, indicating that they might confera selective advantage to the virus. Thus,both of the CTL epitopes mapped in
macaque C accumulated amino-acid substitutions by the time thisanimal was killed with AIDS-associated wasting on day 299 after in-fection.
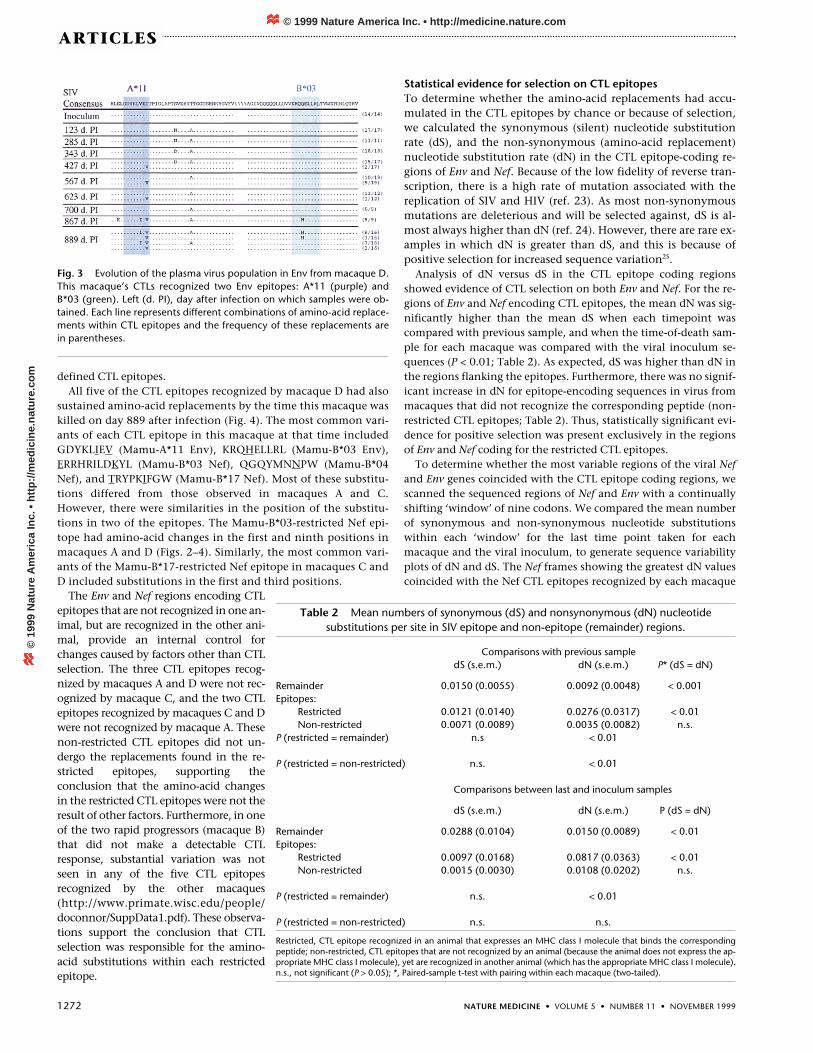
Macaque A developed CTL responses directed against three dif-ferent CTL epitopes derived from the Env and Nef proteins whichwere restricted by Mamu-B*03 and Mamu-B*04. There was achange from leucine to methionine at position 9 of the Mamu-B*03-restricted Env epitope (KRQQELLRM; Fig. 4b). There were alsochanges in the first and second positions of the Mamu-B*04-re-stricted Nef epitope (EGQYMNTPW; QEQYMNTPW; SGQYM-NTPW and PGQYMNTPW; Fig. 4a). Two prominent changes werealso present in the Mamu-B*03-restricted Nef epitope. At the timeof the macaque’s death, the alanine and the isoleucine of this epi-tope had been replaced completely by proline and methionine(PRRHRILDMYL) (Fig. 4a). Thus, by the time macaque A died, theplasma virus population showed amino-acid changes in all three
Fig. 2 Evolution of the plasma virus population in Nef from macaque D.This macaque’s CTLs recognized all three Nef epitopes: B*04 (pink), B*03(blue) and B*17 (yellow). Each of these epitopes sustained amino-acid re-placements by the time of death (889 d). Left (d. PI), day after infection onwhich samples were obtained. Each line represents different combinationsof amino-acid replacements within CTL epitopes, and the frequency ofthese replacements is in parentheses. For example, 13 of 17 clones se-quenced at time of death had Asp (D) substitutions within the B*04 epi-tope, Glu (E) and Lys (K) substitutions within the B*03 epitope, and Thr (T),Tyr (Y) and Ile (I) substitutions within the B*17 epitope. In regions flankingthe CTL epitopes, the consensus amino-acid sequence is shown.
Table 1 CTL epitopes, restricting MHC class I molecules, and disease progression in a family of SIV-infected macaques
Macaque CTL epitopes Restricting Survival time Disease progression(class I haplotype) MHC class I molecule (days)
(haplotype)
C (b/e) GDYKLVEIEnv497-504 Mamu-A*11 (b) 299 intermediateIRFPKTFGWNef165-173 Mamu-B*17 (b)
D (b/c) GDYKLVEIEnv497-504 Mamu-A*11 (b) 889 slowIRFPKTFGWNef 165-173 Mamu-B*17 (b)KRQQELLRLEnv575-583 Mamu-B*03 (c)ARRHRILDIYLNef136-146 Mamu-B*03 (c)QGQYMNTPWNef62-70 Mamu-B*04 (c)
A (a/c) KRQQELLRLEnv575-583 Mamu-B*03 (c) 511 slowARRHRILDIYLNef136-146 Mamu-B*03 (c)QGQYMNTPWNef62-70 Mamu-B*04 (c)
B (a/d) Not detected None 77 rapidB’(a/d) Not detected None 78 rapid
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
1272 NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999
ARTICLES
defined CTL epitopes.All five of the CTL epitopes recognized by macaque D had also
sustained amino-acid replacements by the time this macaque waskilled on day 889 after infection (Fig. 4). The most common vari-ants of each CTL epitope in this macaque at that time includedGDYKLIEV (Mamu-A*11 Env), KRQHELLRL (Mamu-B*03 Env),ERRHRILDKYL (Mamu-B*03 Nef), QGQYMNNPW (Mamu-B*04Nef), and TRYPKIFGW (Mamu-B*17 Nef). Most of these substitu-tions differed from those observed in macaques A and C.However, there were similarities in the position of the substitu-tions in two of the epitopes. The Mamu-B*03-restricted Nef epi-tope had amino-acid changes in the first and ninth positions inmacaques A and D (Figs. 2–4). Similarly, the most common vari-ants of the Mamu-B*17-restricted Nef epitope in macaques C andD included substitutions in the first and third positions.
The Env and Nef regions encoding CTLepitopes that are not recognized in one an-imal, but are recognized in the other ani-mal, provide an internal control forchanges caused by factors other than CTLselection. The three CTL epitopes recog-nized by macaques A and D were not rec-ognized by macaque C, and the two CTLepitopes recognized by macaques C and Dwere not recognized by macaque A. Thesenon-restricted CTL epitopes did not un-dergo the replacements found in the re-stricted epitopes, supporting theconclusion that the amino-acid changesin the restricted CTL epitopes were not theresult of other factors. Furthermore, in oneof the two rapid progressors (macaque B)that did not make a detectable CTLresponse, substantial variation was notseen in any of the five CTL epitopesrecognized by the other macaques(http://www.primate.wisc.edu/people/doconnor/SuppData1.pdf). These observa-tions support the conclusion that CTLselection was responsible for the amino-acid substitutions within each restrictedepitope.
Statistical evidence for selection on CTL epitopesTo determine whether the amino-acid replacements had accu-mulated in the CTL epitopes by chance or because of selection,we calculated the synonymous (silent) nucleotide substitutionrate (dS), and the non-synonymous (amino-acid replacement)nucleotide substitution rate (dN) in the CTL epitope-coding re-gions of Env and Nef. Because of the low fidelity of reverse tran-scription, there is a high rate of mutation associated with thereplication of SIV and HIV (ref. 23). As most non-synonymousmutations are deleterious and will be selected against, dS is al-most always higher than dN (ref. 24). However, there are rare ex-amples in which dN is greater than dS, and this is because ofpositive selection for increased sequence variation25.
Analysis of dN versus dS in the CTL epitope coding regionsshowed evidence of CTL selection on both Env and Nef. For the re-gions of Env and Nef encoding CTL epitopes, the mean dN was sig-nificantly higher than the mean dS when each timepoint wascompared with previous sample, and when the time-of-death sam-ple for each macaque was compared with the viral inoculum se-quences (P < 0.01; Table 2). As expected, dS was higher than dN inthe regions flanking the epitopes. Furthermore, there was no signif-icant increase in dN for epitope-encoding sequences in virus frommacaques that did not recognize the corresponding peptide (non-restricted CTL epitopes; Table 2). Thus, statistically significant evi-dence for positive selection was present exclusively in the regionsof Env and Nef coding for the restricted CTL epitopes.
To determine whether the most variable regions of the viral Nefand Env genes coincided with the CTL epitope coding regions, wescanned the sequenced regions of Nef and Env with a continuallyshifting ‘window’ of nine codons. We compared the mean numberof synonymous and non-synonymous nucleotide substitutionswithin each ‘window’ for the last time point taken for eachmacaque and the viral inoculum, to generate sequence variabilityplots of dN and dS. The Nef frames showing the greatest dN valuescoincided with the Nef CTL epitopes recognized by each macaque
Table 2 Mean numbers of synonymous (dS) and nonsynonymous (dN) nucleotidesubstitutions per site in SIV epitope and non-epitope (remainder) regions.
Comparisons with previous sampledS (s.e.m.) dN (s.e.m.) P* (dS = dN)
Remainder 0.0150 (0.0055) 0.0092 (0.0048) < 0.001Epitopes:
Restricted 0.0121 (0.0140) 0.0276 (0.0317) < 0.01Non-restricted 0.0071 (0.0089) 0.0035 (0.0082) n.s.
P (restricted = remainder) n.s < 0.01
P (restricted = non-restricted) n.s. < 0.01
Comparisons between last and inoculum samples
dS (s.e.m.) dN (s.e.m.) P (dS = dN)
Remainder 0.0288 (0.0104) 0.0150 (0.0089) < 0.01Epitopes:
Restricted 0.0097 (0.0168) 0.0817 (0.0363) < 0.01Non-restricted 0.0015 (0.0030) 0.0108 (0.0202) n.s.
P (restricted = remainder) n.s. < 0.01
P (restricted = non-restricted) n.s. n.s.
Restricted, CTL epitope recognized in an animal that expresses an MHC class I molecule that binds the correspondingpeptide; non-restricted, CTL epitopes that are not recognized by an animal (because the animal does not express the ap-propriate MHC class I molecule), yet are recognized in another animal (which has the appropriate MHC class I molecule).n.s., not significant (P > 0.05); *, Paired-sample t-test with pairing within each macaque (two-tailed).
Fig. 3 Evolution of the plasma virus population in Env from macaque D.This macaque’s CTLs recognized two Env epitopes: A*11 (purple) andB*03 (green). Left (d. PI), day after infection on which samples were ob-tained. Each line represents different combinations of amino-acid replace-ments within CTL epitopes and the frequency of these replacements arein parentheses.
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999 1273
ARTICLES
(Fig. 4a). Similarly, the Env frames corresponding toCTL epitopes in macaques A, C and D also showedincreases in dN (Fig. 4b). These results provide addi-tional evidence that CTL selection is responsible forincreased sequence diversity in the studied regionsof the SIV Nef and Env genes.
MHC I binding and CTL recognition of variantsTo determine whether the amino-acid replace-ments contributed to evasion of the CTL response,we tested variant peptides for MHC class I bindingand CTL recognition. We measured MHC class Ibinding by competition with 125I-labeled epitopeor epitope-analog peptides in a live cell bindingassay26 and assessed SIV-specific CTL activity usingstandard 51Cr-release assays.
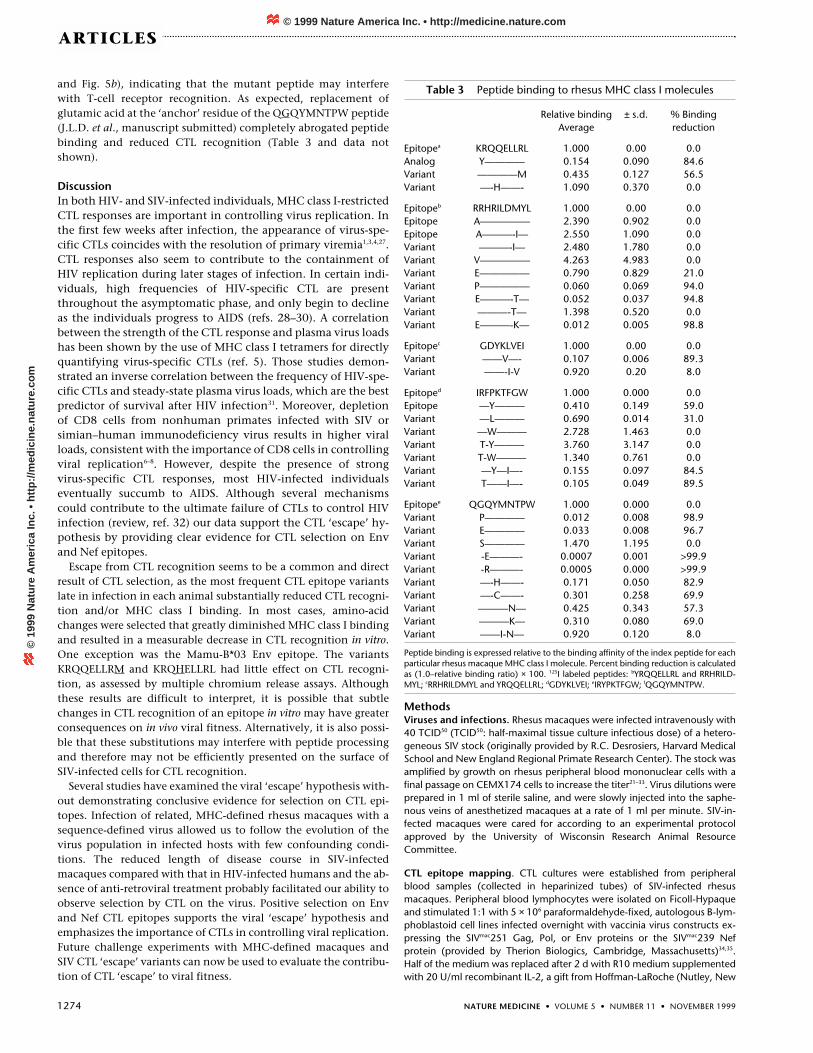
In all but a single epitope, the most frequent vari-ants in the plasma virus population at time of deathdiminished MHC class I binding and/or CTL recog-nition (Table 3 and Fig. 5). The most profoundexample of CTL evasion was in the Mamu-A*11-restricted Env response. The single amino-acid sub-stitution predominating at time of death inmacaque C resulted in both an 89% reduction inpeptide binding and a complete loss of CTL recog-nition at a peptide concentration between 1 and 10nM (Table 3 and Fig. 5a). In contrast, the single replacement ofmethionine for leucine in the Mamu-B*03-restricted Env epitope(KRQQELLRM) reduced peptide binding by 56%, yet had only aminimal effect on CTL lysis (Table 3 and Fig. 5c). The substitutionof histidine for glutamine in the same epitope in macaque D
(KRQHELLRL) also had a limited effect on peptide binding andCTL recognition (Table 3 and Fig. 5h). For the Nef epitopes, mostof the variants reduced both peptide binding and CTL lysis (Table3 and Fig. 5b and d–f). The TRWPKTFGW variant did not reducepeptide binding, yet failed to sensitize targets for CTL lysis (Table 3
Fig. 4 Analysis of sequences iso-lated from MHC-defined rhesusmacaques at time of death. a, Nefsequences. b, Env sequences.Shaded, epitopes recognized byCTLs; boxed, CTL epitopes not rec-ognized in each macaque (control,non-restricted CTL epitopes; Table2). Virus was quantified bybranched DNA analysis. The inocu-lum sequences were amplified froman input pool of 5.98 × 106 viri-ons/ml. Plasma SIV sequences frommacaque C were amplified from1.58 × 106 input virions; macaque Dfrom 4.3 × 104 input virions;macaque A from 2.80 × 105 inputvirions. Each line represents differ-ent combinations of amino-acid re-placements within CTL epitopes,and the frequency of these replace-ments is in parentheses. Variabilityplots below each sequence align-ment show the nucleotide sequencevariability in the CTL epitope codingversus flanking regions. dS (green,line) and dN (red, solid) were com-puted by comparing sequence fromthe most recent time to the inocu-lum for the sequenced region of Nefusing a sliding ‘window’ of ninecodons.
a
b
C
C
D
D
A
A
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
1274 NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999
ARTICLES
and Fig. 5b), indicating that the mutant peptide may interferewith T-cell receptor recognition. As expected, replacement ofglutamic acid at the ‘anchor’ residue of the QGQYMNTPW peptide(J.L.D. et al., manuscript submitted) completely abrogated peptidebinding and reduced CTL recognition (Table 3 and data notshown).
DiscussionIn both HIV- and SIV-infected individuals, MHC class I-restrictedCTL responses are important in controlling virus replication. Inthe first few weeks after infection, the appearance of virus-spe-cific CTLs coincides with the resolution of primary viremia1,3,4,27.CTL responses also seem to contribute to the containment ofHIV replication during later stages of infection. In certain indi-viduals, high frequencies of HIV-specific CTL are presentthroughout the asymptomatic phase, and only begin to declineas the individuals progress to AIDS (refs. 28–30). A correlationbetween the strength of the CTL response and plasma virus loadshas been shown by the use of MHC class I tetramers for directlyquantifying virus-specific CTLs (ref. 5). Those studies demon-strated an inverse correlation between the frequency of HIV-spe-cific CTLs and steady-state plasma virus loads, which are the bestpredictor of survival after HIV infection31. Moreover, depletionof CD8 cells from nonhuman primates infected with SIV orsimian–human immunodeficiency virus results in higher viralloads, consistent with the importance of CD8 cells in controllingviral replication6–8. However, despite the presence of strongvirus-specific CTL responses, most HIV-infected individualseventually succumb to AIDS. Although several mechanismscould contribute to the ultimate failure of CTLs to control HIVinfection (review, ref. 32) our data support the CTL ‘escape’ hy-pothesis by providing clear evidence for CTL selection on Envand Nef epitopes.
Escape from CTL recognition seems to be a common and directresult of CTL selection, as the most frequent CTL epitope variantslate in infection in each animal substantially reduced CTL recogni-tion and/or MHC class I binding. In most cases, amino-acidchanges were selected that greatly diminished MHC class I bindingand resulted in a measurable decrease in CTL recognition in vitro.One exception was the Mamu-B*03 Env epitope. The variantsKRQQELLRM and KRQHELLRL had little effect on CTL recogni-tion, as assessed by multiple chromium release assays. Althoughthese results are difficult to interpret, it is possible that subtlechanges in CTL recognition of an epitope in vitro may have greaterconsequences on in vivo viral fitness. Alternatively, it is also possi-ble that these substitutions may interfere with peptide processingand therefore may not be efficiently presented on the surface ofSIV-infected cells for CTL recognition.
Several studies have examined the viral ‘escape’ hypothesis with-out demonstrating conclusive evidence for selection on CTL epi-topes. Infection of related, MHC-defined rhesus macaques with asequence-defined virus allowed us to follow the evolution of thevirus population in infected hosts with few confounding condi-tions. The reduced length of disease course in SIV-infectedmacaques compared with that in HIV-infected humans and the ab-sence of anti-retroviral treatment probably facilitated our ability toobserve selection by CTL on the virus. Positive selection on Envand Nef CTL epitopes supports the viral ‘escape’ hypothesis andemphasizes the importance of CTLs in controlling viral replication.Future challenge experiments with MHC-defined macaques andSIV CTL ‘escape’ variants can now be used to evaluate the contribu-tion of CTL ‘escape’ to viral fitness.
MethodsViruses and infections. Rhesus macaques were infected intravenously with40 TCID50 (TCID50: half-maximal tissue culture infectious dose) of a hetero-geneous SIV stock (originally provided by R.C. Desrosiers, Harvard MedicalSchool and New England Regional Primate Research Center). The stock wasamplified by growth on rhesus peripheral blood mononuclear cells with afinal passage on CEMX174 cells to increase the titer21–33. Virus dilutions wereprepared in 1 ml of sterile saline, and were slowly injected into the saphe-nous veins of anesthetized macaques at a rate of 1 ml per minute. SIV-in-fected macaques were cared for according to an experimental protocolapproved by the University of Wisconsin Research Animal ResourceCommittee.
CTL epitope mapping. CTL cultures were established from peripheralblood samples (collected in heparinized tubes) of SIV-infected rhesusmacaques. Peripheral blood lymphocytes were isolated on Ficoll-Hypaqueand stimulated 1:1 with 5 × 106 paraformaldehyde-fixed, autologous B-lym-phoblastoid cell lines infected overnight with vaccinia virus constructs ex-pressing the SIVmac251 Gag, Pol, or Env proteins or the SIVmac239 Nefprotein (provided by Therion Biologics, Cambridge, Massachusetts)34,35.Half of the medium was replaced after 2 d with R10 medium supplementedwith 20 U/ml recombinant IL-2, a gift from Hoffman-LaRoche (Nutley, New
Table 3 Peptide binding to rhesus MHC class I molecules
Relative binding ± s.d. % BindingAverage reduction
Epitopea KRQQELLRL 1.000 0.00 0.0Analog Y———— 0.154 0.090 84.6Variant ————M 0.435 0.127 56.5Variant —-H——- 1.090 0.370 0.0
Epitopeb RRHRILDMYL 1.000 0.00 0.0Epitope A————— 2.390 0.902 0.0Epitope A———-I— 2.550 1.090 0.0Variant ———-I— 2.480 1.780 0.0Variant V————— 4.263 4.983 0.0Variant E————— 0.790 0.829 21.0Variant P————— 0.060 0.069 94.0Variant E———-T— 0.052 0.037 94.8Variant ———-T— 1.398 0.520 0.0Variant E———-K— 0.012 0.005 98.8
Epitopec GDYKLVEI 1.000 0.00 0.0Variant ——V—- 0.107 0.006 89.3Variant ——-I-V 0.920 0.20 8.0
Epitoped IRFPKTFGW 1.000 0.000 0.0Epitope —Y——— 0.410 0.149 59.0Variant —L——— 0.690 0.014 31.0Variant —W——— 2.728 1.463 0.0Variant T-Y——— 3.760 3.147 0.0Variant T-W——— 1.340 0.761 0.0Variant —Y—I—- 0.155 0.097 84.5Variant T——I—- 0.105 0.049 89.5
Epitopee QGQYMNTPW 1.000 0.000 0.0Variant P———— 0.012 0.008 98.9Variant E———— 0.033 0.008 96.7Variant S———— 1.470 1.195 0.0Variant -E———- 0.0007 0.001 >99.9Variant -R———- 0.0005 0.000 >99.9Variant —-H——- 0.171 0.050 82.9Variant —-C——- 0.301 0.258 69.9Variant ———N— 0.425 0.343 57.3Variant ———K— 0.310 0.080 69.0Variant ——I-N— 0.920 0.120 8.0
Peptide binding is expressed relative to the binding affinity of the index peptide for eachparticular rhesus macaque MHC class I molecule. Percent binding reduction is calculatedas (1.0–relative binding ratio) × 100. 125I labeled peptides: bYRQQELLRL and RRHRILD-MYL; cRRHRILDMYL and YRQQELLRL; dGDYKLVEI; eIRYPKTFGW; fQGQYMNTPW.
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999 1275
ARTICLES
Jersey). On day 7, viable cells were isolated on Ficoll-Hypaque and againstimulated 1:1 with paraformaldehyde-fixed, vaccinia-infected B-lym-phoblastoid cell lines. CTLs were then expanded in the presence of recom-binant IL-2, and were tested for CTL activity after a total of 13 d in culture.
CTL epitopes of the SIV Env and Nef proteins were mapped using sets ofpeptides synthesized according to the predicted amino-acid sequences ofthese proteins for SIVmac251. Autologous B-lymphoblastoid cell lines werefirst pulsed with ‘pools’ of overlapping peptides 20 amino acids in length,and used as CTL targets in 51Cr-release assays. In subsequent CTL assays,individual peptides 20 amino acids in length from positive pools weretested for CTL recognition. These were then followed by testing overlap-ping peptides 9 amino acids in length within each positive peptide 20amino acids in length. Fine mapping was completed by ‘titrating’ addi-tional peptides with single amino-acid additions or subtractions at the N-and C-termini (peptides 8–10 amino acids in length) to precisely definethe ends of each CTL epitope. SIV-specific CTL activity was assessed usinga standard 51Cr-release assay36.
Plasma virus sequencing. SIV was isolated from 1- to 2-ml frozen plasmasamples collected in tubes containing EDTA. After being thawed, each sam-ple was centrifuged at 10,000g for 10 min to remove any contaminatingcellular debris. The virus was then pelleted in an ultracentrifuge at 35,000rpm for 33 min at 4 °C, using a Beckman SW60 rotor. Viral RNA was ex-tracted from the pellets using 0.5 ml RNA STAT-60 (Tel-Test) according tothe manufacturer’s instructions. After being precipitated overnight at –20°C with an equal volume of isopropanol and 20 µg glycogen, the viral RNAwas collected by centrifugation at 12,500g, washed with 75% ethanol andresuspended in 20 µl RNAse-free water.
Total SIV cDNA was synthesized from 2–3 µl viral RNA in a 20-µl reversetranscription reaction using Superscript II according to the manufacturer’sinstructions (Life Technologies). Fragments approximately 700 bp in lengthof the viral Env and Nef genes were amplified in a 100-µl PCR reaction bythe addition of 80 µl PCR reaction buffer containing 1 mM MgCl2, 2.5 UTaq polymerase (Perkin-Elmer, Norwalk, Connecticut), and 25 pmolprimers (Env-specific: ENVF793, 5’–GCCTCCAAGAGAGGGAGA–3’ and ENVR8637, 5’–AAGGTCAAACCAATTGCC–3’; or Nef-specific:NEFF9047, 5’–CTACCTACAATATGGGTG–3’ and NEFR9762, 5’–GCTTC-CAACTCTTCTGGG–3’). After an initial denaturation step at 94 °C for 1 min,reactions consisted of 40 cycles of denaturation (30 s at 94 °C), annealing(30 s at 53 °C) and extension (45 s at 72 °C), followed by a final extensionstep at 72 °C for 10 min.
To preserve more material for future analysis, viral RNA from macaque Dwas isolated using a QIAgen Viral RNA kit (QIAgen, Valencia, California) ac-cording to the manufacturer’s instructions. Four 140-µl reactions werecombined onto a single purification column to increase the viral RNA yield.
Viral RNA (2 µl) was then used in the SuperScript One-Step RT–PCR System(Life Technologies) with the Env and Nef primers described above. ThecDNA was obtained in a 30-minute step at 50 °C and was amplified with a2-minute 94 °C denaturation step followed by 35 cycles of denaturation (15s at 94 °C), annealing (30 s at 53 °C), and extension (90 s at 72 °C). The PCRwas completed with a final extension at 72 °C for 10 min.
PCR products were cloned into the pCRII vector in a 5-minute ligation re-action at room temperature using the TOPO TA Cloning kit according tothe manufacturer’s instructions (Invitrogen, Carlsbad, California). The liga-tion reactions were then used to transform Escherichia coli strain TOP10(Invitrogen, Carlsbad, California), and colonies were grown overnight on LBplates containing 50 µg/ml ampicillin and 50 µg/ml X-gal. Plasmid DNAwas prepared by ‘miniprep’ from selected white colonies, and sequencedusing the Taq DyeDeoxy Terminator Cycle Sequencing kit and an ABI 373automated sequencer (both from Perkin-Elmer, Norwalk, Connecticut).Primers used for sequencing included the PCR primers ENVF7935,ENVR8637, NEFF9047 and NEFR9762, and nested primer pairs (ENVF7977,5’–CAGTCTCATAGCAAACAT–3’ and ENVR8446, 5’–CAGTAGTGTGGCA-GACTTG–3’; and NEFF9115, 5’–ACTCTTGCGGGCGCGTGG–3’ andNEFR9689, 5’–GAACCTCTCCCCAAGGGT–3’). The consensus was derivedby manually determining the predominant nucleotide at a given position inthe inoculum.
MHC class I binding assay. Peptides were obtained as lyophilized productsfrom Chiron Mimotopes (San Diego, California). Peptides were resus-pended at a concentration of either 10 mg/ml or 20 mg/ml in 10% DMSOand PBS. Peptide GDYKLVEI was used for binding to Mamu-A*11,; peptideIRYPKTFGW. for Mamu-B*17 binding; peptide QGQYMNTPW, for Mamu-B*04 binding, and peptides RRHRILDMYL; and YRQQELLRL, for binding toMamu-B*03. The live cell binding assay was done as described26,36.Macaque MHC class I-transfected 721.221 cells (1 × 106 cells/ml) werepreincubated overnight in RPMI 1640 supplemented with 15% FBS, L-glut-amine, 100 IU/ml penicillin and 100 µg/ml streptomycin at room tempera-ture. Cells were then washed twice in RPMI 1640 without supplements andresuspended at a concentration of 1.25 × 107 cells/ml in RPMI supple-mented with 3 µg/ml β2-microglobulin (Scripps Clinic and ResearchFoundation, La Jolla, California). Peptides purified with high-performanceliquid chromatography were radiolabeled with 125I according to the chlo-ramine-T method37. Cells (2 × 106 cells/well in a 96-well U-bottom mi-crotiter plate) were incubated at 20 °C for 4 h in the presence of 1 × 105
counts per minute (c.p.m.) 125I-radiolabeled peptides, titrating concentra-tions of unlabeled inhibitor peptide (10-fold dilutions, 100 µg–0.1 ng), andprotease inhibitors (final concentrations: 250 µg/ml PMSF, 1.07 mg/mlEDTA, 62.5 µg/ml pepstatin A, 60 µg/ml TLCK and 325 mg/ml phenan-throline). After this incubation, unbound peptide was removed by washing
Fig. 5 Recognition of epitope variants by CTLsfrom macaques C, A and D. Epitope variants thatreduced or eliminated CTL recognition were iden-tified in all three macaques. Amino-acid differ-ences between the epitope sequences of theinoculum and these variants are underlined.Although we tested many mutant peptides in CTLassays, only those peptides corresponding to themost frequent epitope variants identified in theplasma virus populations of each animal at time ofdeath are presented. CTL recognition of thesemutant peptides (� and �) were compared tothe mapped peptide (�) and an irrelevant controlpeptide (�) for each epitope. Autologous B-lym-phoblastoid cell line targets were pulsed with 10-fold dilutions of each peptide and were tested atan effector:target ratio of 20:1 E:T in chromiumrelease assays.
% S
pec
ific
51C
r re
leas
e
Peptide concentration [nM]
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

1276 NATURE MEDICINE • VOLUME 5 • NUMBER 11 • NOVEMBER 1999
ARTICLES
cells three times in serum-free medium, then passed through a FBS gradi-ent. Labeled peptide bound to the cells was then determined by measuringthe radioactivity of pelleted cells on a gamma scintillation counter, and theconcentration of peptide yielding 50% inhibition of the binding of the radi-olabeled probe peptide was calculated (IC50).
Statistical analysis of sequence data. Numbers of synonymous nucleotidesubstitutions per synonymous site (dS) and of nonsynonymous nucleotidesubstitutions per site (dN) were estimated by a published method24, andmean dS and dN were estimated between each sample and the previoussample and between the last sample and inoculum. These were estimatedfor separate regions of the sequenced portions of Env and Nef: epitopes re-stricted by the host; epitopes not restricted by the host; and the flanking re-gions. Mean dS and dN were computed in a sliding ‘window’ of ninecodons along the sequence for comparisons between the last sample takenand the original inoculum.
AcknowledgmentsWe thank L. Smith for help in preparing this manuscript and B. Becker for helpwith illustration. We thank J. Scheffler for initially identifying this family, and J.Mitchen, M. Dykhuizen and L. Acker for infecting the macaques, collectingblood and monitoring disease progression. We also thank J. Malter, S.Wolinsky and G. Watkins for critical review. This work was supported by grantsfrom the National Institutes of Health (AI32426, AI42641, and AI41913 toD.I.W.; AI36643 to C.D.P.; AI15486 to R.D.; GM34940 to A.L.H.; AI38081 toEpimmune; and RR00167 to the Wisconsin Regional Primate Research Center).D.I.W. is an Elizabeth Glaser Scientist.
RECEIVED 24 MAY; ACCEPTED 31 AUGUST 1999
1. Borrow, P., Lewicki, H., Hahn, B., Shaw, G. & Oldstone, M. Virus-specific CD8+ cy-totoxic T-lymphocyte activity associated with control of viremia in primary humanimmunodeficiency virus type 1 infection. J. Virol. 68, 6103–6110 (1994).
2. Koup, R. & Ho, D. Shutting down HIV. Nature 370, 416 (1994).3. Yasutomi, Y., Reimann, K., Lord, C., Miller, M. & Letvin, N. Simian immunodefi-
ciency virus-specific CD8+ lymphocyte response in acutely infected rhesus mon-keys. J. Virol. 67, 1707–1711 (1993).
4. Reimann, K. et al. Immunopathogenic events in acute infection of rhesus monkeyswith simian immunodeficiency virus of macaques. J. Virol. 68, 2362–2370 (1994).
5. Ogg, G.S. et al. Quantitation of HIV-1-specific cytotoxic T lymphocytes andplasma load of viral RNA. Science 279, 2103–2106 (1998).
6. Schmitz, J.E. et al. Control of viremia in simian immunodeficiency virus infectionby CD8+ lymphocytes. Science 283, 857–860 (1999).
7. Jin, X. et al. Dramatic rise in plasma viremia after CD8+ T cell depletion in simianimmunodeficiency virus-infected macaques. J. Exp. Med. 189, 991–998 (1999).
8. Matano, T. et al. Administration of an anti-CD8 monoclonal antibody interfereswith the clearance of chimeric simian/human immunodeficiency virus during pri-mary infections of rhesus macaques. J. Virol. 72, 164–169 (1998).
9. Couillin, I. et al. Impaired cytotoxic T lymphocyte recognition due to genetic vari-ations in the main immunogenic region of the human immunodeficiency virus 1NEF protein. J. Exp. Med. 180, 1129–34 (1994).
10. Goulder, P.J.R. et al. Late escape from an immunodominant cytotoxic T-lympho-cyte response associated with progression to AIDS. Nature Med. 3, 212–217(1997).
11. McMichael, A.J. & Phillips, R.E. Escape of human immunodeficiency virus from im-mune control. Annu. Rev. Immunol. 15, 271–296 (1997).
12. Borrow, P. et al. Antiviral pressure exerted by HIV-1-specific cytotoxic T lympho-
cytes (CTLs) during primary infection demonstrated by rapid selection of CTL es-cape virus. Nature Med. 3, 205–211 (1997).
13. Wolinsky, S.M. et al. Adaptive evolution of human immunodeficiency virus-type 1during the natural course of infection. Science 272, 537–542 (1996).
14. Koenig, S. et al. Transfer of HIV-1-specific cytotoxic T lymphocytes to an AIDS pa-tient leads to selection for mutant HIV variants and subsequent disease progres-sion. Nat. Med. 1, 330–336 (1995).
15. Dai, L.C., West, K., Littaua, R., Takahashi, K. & Ennis, F.A. Mutation of human im-munodeficiency virus type 1 at amino acid 585 on gp41 results in loss of killing byCD8+ A24-restricted cytotoxic T lymphocytes. J. Virol. 66, 3151 (1992).
16. Haas, G. et al. Dynamics of viral variants in HIV-1 Nef and specific cytotoxic T lym-phocytes in vivo. J. Immunol. 157, 4212–4221 (1996).
17. Mortara, L. et al. Selection of virus variants and emergence of virus escape mutantsafter immunization with an epitope vaccine. J. Virol. 72, 1403–1410 (1998).
18. Price, D.A. et al. Positive selection of HIV-1 cytotoxic T lymphocyte escape variantsduring primary infection. Proc. Natl. Acad. Sci. USA 94, 1890–1895 (1997).
19. Balter, M. Modest Briton stirs up storm with views on role of CTLs. Science 280,1860–1861 (1998).
20. Brander, C. et al. Lack of strong immune selection pressure by the immunodomi-nant, HLA-A*0201-restricted cytotoxic T lymphocyte response in chronic humanimmunodeficiency virus-1 infection. J. Clin. Invest. 101, 2559–2566 (1998).
21. Dykhuizen, M. et al. Determinants of disease in the simian immunodeficiencyvirus-infected rhesus macaque: characterizing animals with low antibody re-sponses and rapid progression. J. Gen. Virol. 79, 2461–2467 (1998).
22. Pauza, C.D. et al. Pathogenesis of SIVmac251 after atraumatic inoculation of therectal mucosa in rhesus monkeys. J. Med. Primatol. 22, 154–61 (1993).
23. Coffin, J.M. HIV population dynamics in vivo: Implications for genetic variation,pathogenesis, and therapy. Science 267, 483–489 (1995).
24. Nei, M. & Gojobori, T. Simple methods for estimating the numbers of synony-mous and nonsynonymous nucleotide substitutions. Molec. Biol. Evol. 3, 418–426(1986).
25. Hughes, A.L. & Nei, M. Pattern of nucleotide substitution at major histocompati-bility complex class I loci reveals overdominant selection. Nature 335, 167–170(1988).
26. delGuercio, M.F. et al. Binding of a peptide antigen to multiple HLA alleles allowsdefinition of an A2-like supertype. J. Immunol. 154, 685–693 (1995).
27. Koup, R.A. et al. Temporal association of cellular immune responses with the initialcontrol of viremia in primary human immunodeficiency virus type 1 syndrome. J.Virol. 68, 4650–4655 (1994).
28. Walker, B.D. et al. HIV-specific cytotoxic T lymphocytes in seropositive individuals.Nature 328, 345–348 (1987).
29. Koup, R.A. et al. Limiting dilution analysis of cytotoxic T lymphocytes to humanimmunodeficiency virus gag antigens in infected persons: In vitro quantitation ofeffector cell populations with p17 and p24 specificities. J. Exp. Med. 174,1593–1600 (1991).
30. Carmichael, A., Jin, X., Sissons, P. & Borysiewicz, L. Quantitative analysis of thehuman immunodeficiency virus type 1 (HIV-1)-specific cytotoxic T lymphocyte(CTL) response at different stages of HIV-1 infection: Differential CTL responses toHIV-1 and Epstein-Barr virus in late disease. J. Exp. Med. 177, 249–256 (1993).
31. Mellors, J.W. et al. Prognosis in HIV-1 infection predicted by the quantity of virusin plasma. Science 272, 1167–1170 (1996).
32. McMichael, A. T cell responses and viral escape. Cell 93, 673–676 (1998).33. Trivedi, P. et al. Selective amplification of simian immunodeficiency virus geno-
types after intrarectal inoculation of rhesus monkeys. J. Virol. 62, 7649–7653(1994).
34. van Baalen, C.A. et al. Selective in vitro expansion of HLA class I-restricted HIV-1gag-specific CD8+ T cells: Cytotoxic T-lymphocyte epitopes and precursor fre-quencies. AIDS 7, 781–786 (1993).
35. Voss, G. et al. Human immunodeficiency virus type 1 envelope glycoprotein-spe-cific cytotoxic T lymphocytes in simian-human immunodeficiency virus-infectedrhesus monkeys. Virology 208, 770–775 (1995).
36. Allen, T.M. et al. Characterization of the peptide-binding motif of a rhesus MHCclass I molecule (Mamu-A*01) that binds an immunodominant CTL epitope fromSIV. J. Immunol. 160, 6062–6071 (1998).
37. Greenwood, F., Hunter, W. & Glover, J. The preparation of 131I-labeled humangrowth hormone of high specific radioactivity. Biochem. J. 89, 114 (1963).
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m





























