Embed Size (px)
Citation preview

INTRODUCTION
Vertebrate intermediate filament proteins (IFPs) all share asimilar secondary and tertiary structure and all form ultra-structurally similar filaments (see Aebi et al., 1988; Steinertand Roop, 1988; Parry and Steinert, 1992; van de Klundert etal., 1993). Three distinct structural domains have been recog-nized: an N-terminal ‘head’, a central
α-helical ‘rod’ and a C-terminal ‘tail’. Previous studies have shown that the head androd domains are important in intermediate filament (IF)assembly (see van de Klundert et al., 1993, for review). Inter-actions between rod domains drive the formation of α-helicalcoiled-coil dimers. Dimers associate in an anti-parallel andstaggered manner to form tetramers (Geisler et al., 1985, 1992;Potschka et al., 1990; Steinert, 1991; Steinert et al., 1993).Tetramers, in turn, associate into higher order oligomers toform an IF (Ip et al., 1985; Aebi et al., 1988; Steinert, 1991).Isolated head domains of desmin have been shown to formdimers in solution (Saeed and Ip, 1989). The head domain of
vimentin binds with high affinity to vimentin’s rod domain(Traub et al., 1992) and a conserved nonapeptide motif in thehead has been implicated in the formation of higher orderstructures (Herrmann et al., 1992; Hofmann and Herrmann,1992).
In contrast, the role of the tail domains of IFPs are much lessclear. Even the location of the tail domain with respect to theIF axis is uncertain. Axial mass measurements of IFs have beeninterpreted to indicate that the head and tail domains arelocated at the periphery of the IF (Steven et al., 1983a,b; Engelet al., 1986). In neurofilaments the distal portion of the taildomain clearly projects centrifugally from the filament axis(Hisanaga and Hirokawa, 1989). On the other hand Birken-berger and Ip (1990) reported that an antisera directed againstthe C-terminal end of desmin’s tail bound to desmin tetramersbut not to desmin filaments, arguing that at least a portion ofthe tail domain is hidden in the assembled filament.
There have been conflicting reports on the effects ofremoving or mutating the tail domain on the ability of IFPs to
1609
The tail domain of the intermediate filament (IF) proteinvimentin is unnecessary for IF assembly in vitro
. To studythe role of vimentin’s tail in vivo, we constructed a plasmidthat directs the synthesis of a ‘myc-tagged’ version of theXenopus vimentin-1 tail domain in bacteria. This polypep-tide, mycVimTail, was purified to near homogeneity andinjected into cultured Xenopus A6 cells. In these cells thetail polypeptide co-localized with actin even in the presenceof cytochalasin. Two myc-tagged control polypeptidesargue for the specificity of this interaction. First, a similarlymyc-tagged lamin tail domain localizes to the nucleus, indi-cating that the presence of the myc tag did not itself conferthe ability to co-localize with actin (Hennekes and Nigg(1994) J. Cell Sci. 107, 1019-1029). Second, a myc-taggedpolypeptide with a molecular mass and net charge at phys-iological pH (i.e. −4) similar to that of the mycVimTailpolypeptide, failed to show any tendency to associate withactin-containing structures, indicating that the interactionbetween mycVimTail and actin-containing structures wasnot due to a simple ionic association. Franke (1987; Cell
Biol. Int. Rep. 11, 831) noted a similarity in the primarysequence between the tail of the type I keratin DG81A andvimentin. To test whether the DG81A tail interacted withactin-containing structures, we constructed and purifiedmyc-tagged DG81A tail polypeptides. Unexpectedly, thesekeratin tail polypeptides were largely insoluble under phys-iological conditions and formed aggregates at the site ofinjection. While this insolubility made it difficult todetermine if they associated with actin-containing struc-tures, it does provide direct evidence that the tails ofvimentin and DG81A differ dramatically in their physicalproperties. Our data suggest that vimentin’s tail domainhas a highly extended structure, binds to actin-containingstructures and may mediate the interaction betweenvimentin filaments and microfilaments involved in thecontrol of vimentin filament organization (Hollenbeck et al.(1989) J. Cell Sci. 92, 621; Tint et al. (1991) J. Cell Sci. 98,375).
Key words: vimentin, actin, tail domain, keratin
SUMMARY
Vimentin’s tail interacts with actin-containing structures in vivo
Robert B. Cary1,*, Michael W. Klymkowsky1,*,†, Robert M. Evans2, Alberto Domingo1,‡, Joseph A. Dent1,§ andLeilah E. Backhus1
1Molecular, Cellular and Developmental Biology, University of Colorado, Boulder, Boulder, Colorado 80309-0347, USA2Department of Pathology, University of Colorado Health Sciences Center, Denver, Colorado 80262-0216, USA
*These two authors contributed equally to this paper†Author for correspondence‡Present address: Department of Biochemistry, School of Medicine, Autonomous University of Madrid, Madrid, Spain§Present address: Department of Biochemistry, University of Texas Southwestern Medical Center, Dallas, Texas 75235-9038, USA
Journal of Cell Science 107, 1609-1622 (1994)Printed in Great Britain © The Company of Biologists Limited 1994

1610
assemble into IFs. Quinlan et al. (1989) found that deletion ofthe tail domain inhibited the ability of the IFP glial fibrillaryacidic protein (GFAP) to form filaments in vitro
. Similarly,Birkenberger and Ip (1990) found that a polypeptide derivedfrom the tail domain of desmin, as well as an antibody directedagainst this region, were able to perturb desmin filamentformation in vitro. Based on the effects of anti-tail antibodieson vimentin filament organization and in vitro binding studiesKoulkis et al. (1991) suggested that the tail domain of vimentinbound to a region at the end of the rod domain and that thisinteraction acted to inhibit the lateral association of vimentinfilaments. It has also been proposed that interactions betweenthe nuclear lamins and the tail domains of type III IFPs areinvolved in the nucleation of IF assembly (Georgatos andBlobel, 1987; Georgatos et al., 1987; Djabali et al., 1991; Papa-marcaki et al., 1991).
There have also been reports in which removal of the taildomain of vimentin (Shoeman et al., 1990; Eckelt et al., 1992;McCormick et al., 1993), desmin (Kaufmann et al., 1985), andkeratins (Wilson et al., 1992) were found to have little if anynoticeable effect on the in vitro formation of IFs. McCormicket al. (1993) found that mutations that eliminate the in vitrointeraction between the tail and rod domains of vimentin haveno apparent effect on the formation or aggregation of vimentinfilaments (see below). Tail-deleted IFPs readily integrate intopre-existing IF systems (see Eckelt et al., 1992 and McCormicket al., 1993, and references therein).
In cells with no pre-existing IF system the effects ofmutations in the tail domain of IFPs are generally much moredramatic. Deletions in the region of the desmin tail (Raats etal., 1991) lead to failure to form filaments when synthesized inhuman MCF-7 cells. Lu and Lane (1990) found that tail-lesspairs of keratins formed aberrant filament systems in mouseNIH-3T3 cells. Eckelt et al. (1992) found that the removal ofthe tail domain of Xenopus vimentin lead to aberrant filamentnetwork formation in bovine BMGE+H cells (see alsoHerrmann et al., 1993). On the other hand, Bader et al. (1991)reported that tail-less keratins formed superficially normallooking filament networks in mouse 3T3-L1 cells. In a partic-ularly striking set of observations McCormick et al. (1993)found that a human vimentin protein with its tail removed stillformed extensive filament networks in human MCF-7 cells,although some abnormalities in the organization of the networkwere observed.
As this short review indicates, the exact role of the taildomain in IF structure and organization is still unclear. Thefailure of tail-less IFPs to form extended IF networks in manycells has been interpreted to mean that ‘the establishment ofextensive IF arrays in a specific cell type could well involvethe coordinate interaction of the tail portions with one orseveral unknown cellular factor(s)’ (Eckelt et al., 1992). Whatthese cellular factors might be remains unclear. To approachthis question we have examined the behavior of the tail domainas an isolated entity. We constructed plasmids that drive thesynthesis of N-terminally epitope-tagged forms of the taildomains of Xenopus vimentin-1 and the Xenopus type Iepidermal keratin DG81A. The purified tagged tail polypep-tides were then studied both in vitro and in vivo. In vivo itappears that the tail domains of vimentin interact with actin-containing structures. The keratin tail, in contrast, behavesquite differently within the cell. These results point to previ-
ously unrecognized differences between the tail regions ofdifferent IFP types and suggests that the tail domain ofvimentin may well mediate interactions between vimentinfilaments and microfilaments.
MATERIALS AND METHODS
Construction of myc-tagged vimentin and keratin taildomain plasmidsThe tail domain of the Xenopus laevis vimentin-1 (Dent et al., 1992)and the X. laevis type I epidermal keratin DG81A (Jonas et al., 1985)(supplied by Tom Sargent, NIH) were amplified by PCR (Fig. 1). Inthe case of the vimentin domain, the upstream oligonucleotideincluded an NdeI site, the sequence encoding the myc-tagging epitope(MEQKLISEEDLN), an EcoRI site, and a sequence complementaryto the antisense strand of the 5′ end of the tail region. The downstreamoligonucleotide overlapped the 3′ end of the tail domain and containsan XbaI site. The amplification reaction was carried out as follows:94°C for 3 minutes, 5 cycles of 94°C for 2 minutes, 55°C for 2minutes, 72°C for 2 minutes followed by two cycles of 94°C for 2minutes, 72°C for 2 minutes and then 72°C for 8 minutes. The productwas restricted with NdeI and XbaI and subcloned into the pSP6.stopplasmid (Dent et al., 1992) to generate pSP6.myc-tagged vimentin tail(pSP6.mycVimTail). This plasmid was then restricted with NdeI andBamHI and the released fragment was subcloned in the pET3aplasmid (Studier et al., 1990) to create pET.mycVimTail. The originalpET3a plasmid was a gift of W. Studier (Brookhaven National Lab-oratory).
pET3a contains a single EcoRI and a single XbaI site; these siteswere removed sequentially by cutting the plasmid with the appropri-ate enzyme, filling the ends with Klenow and re-ligating to formpET*. To create a plasmid encoding the DG81A tail domain wesubcloned the vimentin-1.stop coding region from pSP6.vimentin-1.stop (Dent et al., 1992) using the NdeI and BamHI sites into pET*to form pET*vimentin-1.stop. This plasmid was then restricted withNdeI and XbaI to remove the vimentin-1 sequence and the mycVim-Tail DNA was subcloned in to form pET*mycVimTail. The taildomain of the type I keratin DG81A was amplified using oligonu-cleotide primers similar to those used for vimentin (Fig. 1) followingthe regime 95°C for 3 minutes, 5 cycles of 95°C for 2 minutes, 37°Cfor 2 minutes, 72°C for 2 minutes followed by two cycles of 95°C for2 minutes, 72°C for 2 minutes and then 72°C for 8 minutes. Theamplified DG81A DNA was restricted with EcoRI and XbaI andsubcloned into the EcoRI/XbaI-digested pET*mycVimTail plasmid togenerate pET*mycKerTail (Fig. 1B). We also subcloned the originalamplified mycVimTail DNA into the pSP6.myc plasmid (Dent et al.,1992) using the NdeI and XbaI sites to create pSP6.mycVimTailmyc.Amplified DG81A DNA was restricted with EcoRI and XbaI andsubcloned into pSP6.mycVimTailmyc to produce pSP6.mycKer-Tailmyc. The mycKerTailmyc insert was released from this plasmidby restriction with NdeI and BamHI and subcloned into pET to createpET.mycKerTailmyc. The pET plasmids were initially transformedinto Escherichia coli JM103 and plasmid DNA was sequenced usingthe T7 promoter primer and the dideoxynucleotide method (Chen andSeeburg, 1985) (Fig. 1C). The DNA was then transformed intoBL21DE3pLysS cells (Studier et al., 1990) for polypeptideexpression.
DNA- and RNA-expression plasmidsSince it is unclear whether the purified mycVimTail polypeptide is ina native configuration when injected into cells, we also constructedDNA- and RNA-expression vectors. pET.mycVimTail was digestedwith NdeI/BamHI to release the mycVimTail sequence, which wasthen subcloned into the pSK.tag (Dent et al., 1992) to formpSK.mycVimTail.stop. The mycVimTail.stop insert was liberated
R. B. Cary and others

1611Vimentin’s tail
with SalI/BamHI and subcloned into pCskAct.vimentin-1.myc,replacing the vimentin-1.myc sequence. This plasmid was generatedfrom a pCskAct.LacZ plasmid supplied to us by Richard Harland (UCBerkeley). pCskAct contains the X. borealis cytoskeletal actinpromoter and the SV40 polyadenylation signal (Cross et al., 1988;Smith and Harland, 1991); the pCskAct promoter is very efficient atdriving the synthesis of exogenous proteins in both Xenopus embryos(Vize et al., 1991) and Xenopus A6 cells (Cary and Klymkowsky,submitted). Supercoiled pCskAct.mycVimTail DNA was injected intothe nuclei of A6 cells at a concentration of ~1 mg/ml.
Next, we isolated the NdeI/BamHI mycVimTail fragment frompCskAct.mycVimTail and subcloned it into pT7.vimentin-1.myc,replacing the vimentin-1.myc sequence. The pT7 plasmid is derivedfrom the pSP64T plasmid (supplied by Doug Melton, Harvard Uni-versity). To construct pT7, the SP6 promoter was removed frompSP64T by digestion with NheI and HindIII and the plasmid wasisolated from an agarose gel using glass beads. Two oligonucleotideswere annealed to form the T7 RNA polymerase promoter insert.
5′ GGGGCTAGCTAATACGACTCACTATAGGGAGAAGCT-TGGG 3′
3′ CCCCGATCGATTATGCAGAGTGATATCCCTCTTCGAA-CCC 5′
where the double underlined sequence is the TATA box. NheI (dottedunderline) and HindIII (solid underline) sequences are also marked.After annealing, the oligos were digested with NheI and HindIIIovernight at 20°C and isolated using a Bio101 Mermaid® kit. The
digested oligo and plasmid were then ligated and confirmed by restric-tion digestion and by the fact that the resultant plasmid producesappropriate sized RNA when incubated with T7 RNA polymerase.The pT7.mycVimTail plasmid was confirmed by restriction digestionand by using a coupled T7 transcription/translation system (Promega).In this system, the pT7.mycVimTail drove the synthesis of a polypep-tide that co-migrated with purified mycVimTail polypeptide on SDS-PAGE (data not shown). Capped RNA was made from BamHI lin-earized pT7.mycVimTail plasmid using a T7 ‘Message Machine®’ kit(Ambion). Purified RNA was resuspended in RNase-free water andinjected into cells at a concentration of 1.3 mg/ml.
Myc-tagged control polypeptidesTo test whether the binding of mycVimTail to actin-containing struc-tures was due to the presence of the myc-epitope, we obtained aplasmid encoding an N-terminally myc-tagged lamin tail domainunder control of the cytomegalovirus promoter from H. Hennekes andEric Nigg (Swiss Institute for Experimental Cancer Research,Lausanne, Switzerland) (Hennekes and Nigg, 1994). Qiagen purifiedplasmid DNA was resuspended in distilled water and injected into thenuclei of cultured A6 cells (see below). We also constructed a myc-tagged form of the influenza hemagglutinin-derived sequenceYPYDVPDY. The original plasmid, GEPTI, was constructed by B.Futcher (Cold Spring Harbor Laboratory, NY) (see Roof et al., 1992).It was given to us by Eric Weiss and Mark Winey (UC Boulder). InGEPTI the hemagglutinin-derived sequence encoding YPYDVPDYis repeated three times. The GEPTI plasmid contains a EcoRI site that
aug¨--------tct aca atg agc ctg a------------cag cac cac gat gac ttt gag tgagtc gtg gtg cta ctg aaa ctc
5-oligo
3' oligoaga tct cccoriginal sequence
S T M S L --- Q H H D D F E stop
M E Q K L I S E E D L N S T M S L --- Q H H D D F E S R stopfinal sequence
ctg aat tct aca atg agc ctg a
cccat atg gag caa aag ctt att agc gag gaa gat
Nde IEco RI
Xba I
aug--------acc aca gtg gca------------------gtt tca tca aga gtc gag taaacc aca gtg gca
5-oligocc ccg aat tct
M E Q K L I S E E D L N T T V A --- G S S R V E S R stopfinal sequence
original sequenceT T V A --- V S S R V E stop
ca agt agt tct cag ctc
aga tct cct agg tcg
Xba I
3' oligo
Eco RI
Bam HI
mycVIMTAIL MEQKLISEEDLN---------------STMSLRETNLDSHPAETHSKRTVLIKTVETRDGQVVNESSQHHDDFE.SR.stop |||||||||||| :: | |: :| | | | | || |::mycKERTAIL MEQKLISEEDLN------------TTVANTSSVESKTESSSTSTTRTRMVKTIVEEVVDGKVGSSRVE.SR.stopmycKERTAILmyc MEQKLISEEDLN------------TTVANTSSVESKTESSSTSTTRTRMVKTIVEEVVDGKVGSSRVE.SRDST.MEQKLISEEDLN.stop
Vimentin-1
Keratin DG81A
Bam HI
codingsequence
stop
pETmyc-stop
S R stop
Tag
Bam HI
T7 promoterpETmyc-myc
Tag
Nde I Eco RI Xba I
SRDST
TagT7 promoter
Fig. 1. Schematic for construction ofpET.mycVimTail and pET.mycKerTail.Plasmids containing cDNA encoding X.laevis vimentin-1 or DG81A proteinswere amplified using theoligonucleotides shown in a. Theamplified vimentin-1 tail DNA wasdigested with NdeI and XbaI andsubcloned into pET8.vimentin-1.stop(b), replacing the original full lengthvimentin-1 coding sequence. Thisformed the pET.mycVimTail plasmid.Amplified DG81A tail DNA wasdigested with EcoRI and XbaI andsubcloned into either pET.mycVimTailor pET.mycVimTailmyc (b) replacingthe VimTail sequence. This producedthe pET.mycKerTail andpET.mycKerTailmyc plasmids. Theresulting pET plasmids were sequenced.The sequence of the mycVimTail,mycKerTail and mycKerTailmycpolypeptides are shown in c. Homologybetween mycVimTail and mycKerTailpolypeptides is noted using a ‘|’ todenote amino acid identity and a ‘:’ todenote a conservative amino aciddifference.
a
b
c
.................

1612
was removed by cutting with EcoRI, filling in with Klenow, and reli-gating. The resulting EcoRI-minus plasmid was cut with SacI, blunt-ended with mung bean nuclease and ligated to EcoRI linkers(Promega cat. no. 901028). The EcoRI/XbaI fragment from thisplasmid was then ligated into pET*mycVimTail, replacing theVimTail sequence to form pET*myc3xHa. This plasmid wassequenced using the T7 promoter of the pET plasmid. The sequenceof the encoded polypeptide was MEQKLISEEDLNSGHRGGGRI-FYPYDVPDYAGYPYDVPDYAGSYPYDVPDYPAQCGRSR.stop,where the underlined region is the myc-tag sequence and the double-underlined region the repeated hemagglutinin-derived sequence.pET*myc3xHa was then transformed in BL21DE3pLysS cells forsynthesis of the myc3xHa polypeptide (see below).
Purification of myc-tagged polypeptidesFor purification of mycVimTail, mycKerTail, mycKerTailmyc andmyc3xHa polypeptides, 1 litre cultures of bacteria were grown in LBbroth to an A600 of 0.6-1 and then induced with 0.4 mM isopropyl-β-D-thiogalactopyranoside for 2 hours. The polypeptides were followedthrough the purification by a dot-blot assay using the anti-mycantibody 9E10 (Evan et al., 1985) (see below). The tail polypeptidesdo not bind efficiently to nitrocellulose paper but can be fixed usingglutaraldehyde. Samples (2 µl) were applied to nitrocellulose paperand dried; the polypeptide was then fixed using 0.5% glutaraldehydein phosphate-buffered saline (PBS) for 10 minutes. The paper wasthen blocked for 10 minutes in 2% low fat dried milk (LFDM) in Tris-buffered saline (TBS) and incubated in 10 µg/ml 9E10 antibody for30 minutes, washed, and incubated in 1:1000 diluted horseradish per-oxidase-conjugated anti-mouse immunoglobulin antibody (Bio-Rad)for 30 minutes. Bound antibody was visualized using the diaminoben-zidine reaction.
Bacteria were recovered by centrifugation (5,000 g, 15 minutes);the cell pellet was resuspended in 100 mM Tris-HCl, 2 mM EDTA,pH 8.0, collected by centrifugation and frozen at −70°C. Afterthawing, the pellet was resuspended in 20 ml 7 M guanidine HCl in50 mM Tris-HCl, 1 mM EDTA, pH 8.0, and frozen at −70°C. Afterthawing, the solution was diluted with 80 ml of 50 mM Tris-HCl, 1mM EDTA, pH 8.0, and centrifuged at 5,000 g for 30 minutes. Inthe case of mycVimTail, mycKerTail and mycKerTailmyc polypep-tides the supernatant was recovered, 200 ml of 95% ethanol wasadded, and the solution was placed at −70°C for 2 hours or longer.After thawing, the solution was centrifuged at 5,000 g for 30minutes; the pellet was solubilized in 10 mM Tris-HCl, 0.2 mMEDTA, pH 8.8, and then centrifuged at 15,000 rpm for 30 minutesin a Sorvall SS-34 rotor. The supernatant was passed through a 0.2µm syringe filter and then applied to a HQ-5 MonoQ column(Pharmacia) at a rate of 1 ml/minute. The column was eluted witha 30 ml 0 to 130 mM NaCl gradient in 15 mM Tris-HCl, pH 8.8.To recover the purified polypeptides, fractions were combined, pre-cipitated with 2 volumes of 100% ethanol and resuspended in thebuffer of choice.
In the case of myc3xHa, following the dilution and centrifugationof the guanidine fraction 400 ml of 95% ethanol was added to thesupernatant and the solution was placed at −70°C overnight; insolublematerial was collected by centrifugation and discarded. A second 400ml of 95% ethanol was then added to the supernatant, which was againincubated overnight at −70°C. The insoluble material was thencollected, dried down, resuspended in 0.1% aqueous trifluoroaceticacid (TFA) and separated on a 25 cm Vydac 214-TP546 C4 reversed-phase HPLC column. The column was run at room temperature at aflow rate of 1.5 ml/minute. The sample was applied to the column andrun in 0.1% aqueous TFA for 2 minutes. The column was eluted witha linear gradient of 0 to 80% acetonitrile, 0.1% TFA for 8 minutes,30 to 40% acetonitrile, 0.1% TFA for 10 minutes, and then 40 to 55%acetonitrile, 0.1% TFA for 7 minutes. Fractions were collected, driedunder vacuum, and resuspended in distilled water. Samples wereassayed for anti-myc reactivity by dot-blot as described above. The
major 9E10-reactive fraction was lyophilized and resuspended indistilled water for injection.
The purity of the MonoQ-purified mycVimTail polypeptide wasassayed by reverse-phase high pressure liquid chromatography(HPLC) on a 25 cm Vydac 214-TP546 C4 column. The column wasrun at room temperature with a flow rate of 1.5 ml/minute and sampleswere applied to the column and run in 0.1% aqueous TFA for 2minutes. Retained material was eluted with a linear gradient of 0 to40% acetonitrile, 0.1% TFA for 25 minutes, then 40 to 80% acetoni-trile, 0.1% TFA for 5 minutes. The purified mycVimTail was cleavedwith 10 mg/ml cyanogen bromide in 70% formic acid for 4 hours. Thesample was dried under vacuum and again analyzed by HPLC; underthese conditions it produced the two expected cleavage products (datanot shown). The purity of the mycVimTail polypeptide was alsoassayed using polyacrylamide gel electrophoresis (Laemmli, 1980).Because of the presence of only a single aromatic residue, the myc-tagged vimentin and keratin tail polypeptides have little molarabsorbency at 280 nm. Protein concentration was estimated using aCoomassie Brilliant Blue binding assay using 2 mg/ml bovine serumalbumin as a standard (Pierce).
Size-exclusion chromatography and cross-linking studiesThe apparent size of the mycVimTail polypeptide was analyzed bysize-exclusion chromatography on Sephacryl S-200 (1.5 cm × 20 cmcolumn). The column was run in TBS with increasing concentrationsof urea or 0.5% SDS. Blue dextran (Mr 2×106), bovine serumalbumin (Mr 67,000), lyzozyme (Mr 14,000) and Phenol Red (Mr 354)were used as size markers. For cross-linking studies, MonoQ-purifiedmycVimTail polypeptide fractions were precipitated with ethanoland resuspended in 50 mM sodium phosphate, 10 mM β-mercap-toethanol buffer (Saeed and Ip, 1989); 50 µl of a 0.1 to 1 mg/mlsolution of purified polypeptide was incubated at room temperature(21-23°C) for 30 minutes in the presence of 0.1% glutaraldehyde(EM grade) in the presence or absence of 0.5% SDS. Alternatively,purified polypeptide was incubated at room temperature for 60minutes in 1 mg/ml dimethylsuberimidate alone, or in the presenceof 2 M urea or 0.5% SDS. The cross-linking reactions were stoppedwith SDS sample buffer and the samples were electrophoresed on12.5 or 15% gels and the gel was stained with Coomassie BrilliantBlue.
Intracellular injection of purified protein and DNAXenopus A6 cells (obtained from American Type Culture Collection)were cultured on coverslips in 85% Leibovitz L-15 media supple-mented with 10% fetal calf serum and 50 µg/ml gentamycin. Purifiedpolypeptides were resuspended in distilled water (final concentration1 to 3 mg/ml) and injected into cells using a manually-controlledpressure injection system (Klymkowsky, 1981). The same apparatuswas used for the intranuclear injection of pCskAct.mycVimTail DNAor cytoplasmic injection of mycVimTail RNA. In some experiments,injected cells were treated with cytochalasin D (2.5 µg/ml final con-centration) or with nocodozole (5 µg/ml final concentration). At timesranging from 5 to 120 minutes after injection, the cells were washedonce with PBS, fixed with 3.7% formaldehyde for 10 minutes at roomtemperature and then extracted with 100% methanol for 1 minute. Insome cases, the cells were extracted with PHEM-0.15% Triton X-100buffer (Schliwa and van Blerkom, 1981) or MES buffer (Algrain etal., 1993) for 15-60 seconds prior to formaldehyde fixation. Fixedcells were washed in TBS and incubated with the anti-myc antibody9E10, followed by fluorescein-conjugated anti-mouse immunoglobu-lin antibody (antiMIg-Fl). For double labeling, cells were stained with9E10, fluorescein-conjugated anti-mouse γ-chain specific antibody(antiMIgγ-Fl), and a mouse monoclonal anti-actin antibody(IgM)(Amersham - cat no. N.350) and rhodamine-conjugated anti-mouse µ chain-specific antibody (antiMIgµ-Rd). 9E10 is obtainablefrom the American Type Culture Collection. Cells were examined andphotographed using a Zeiss IM35 microscope.
R. B. Cary and others
1613Vimentin’s tail
RESULTS
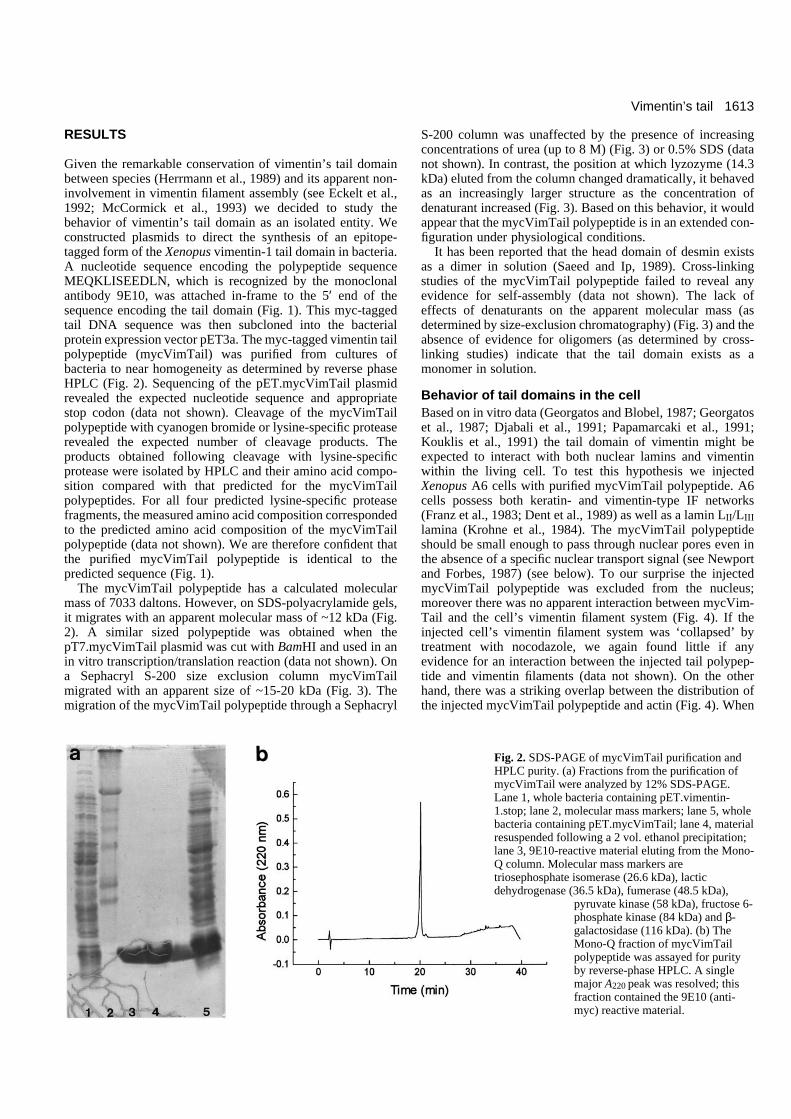
Given the remarkable conservation of vimentin’s tail domainbetween species (Herrmann et al., 1989) and its apparent non-involvement in vimentin filament assembly (see Eckelt et al.,1992; McCormick et al., 1993) we decided to study thebehavior of vimentin’s tail domain as an isolated entity. Weconstructed plasmids to direct the synthesis of an epitope-tagged form of the Xenopus vimentin-1 tail domain in bacteria.A nucleotide sequence encoding the polypeptide sequenceMEQKLISEEDLN, which is recognized by the monoclonalantibody 9E10, was attached in-frame to the 5′ end of thesequence encoding the tail domain (Fig. 1). This myc-taggedtail DNA sequence was then subcloned into the bacterialprotein expression vector pET3a. The myc-tagged vimentin tailpolypeptide (mycVimTail) was purified from cultures ofbacteria to near homogeneity as determined by reverse phaseHPLC (Fig. 2). Sequencing of the pET.mycVimTail plasmidrevealed the expected nucleotide sequence and appropriatestop codon (data not shown). Cleavage of the mycVimTailpolypeptide with cyanogen bromide or lysine-specific proteaserevealed the expected number of cleavage products. Theproducts obtained following cleavage with lysine-specificprotease were isolated by HPLC and their amino acid compo-sition compared with that predicted for the mycVimTailpolypeptides. For all four predicted lysine-specific proteasefragments, the measured amino acid composition correspondedto the predicted amino acid composition of the mycVimTailpolypeptide (data not shown). We are therefore confident thatthe purified mycVimTail polypeptide is identical to thepredicted sequence (Fig. 1).
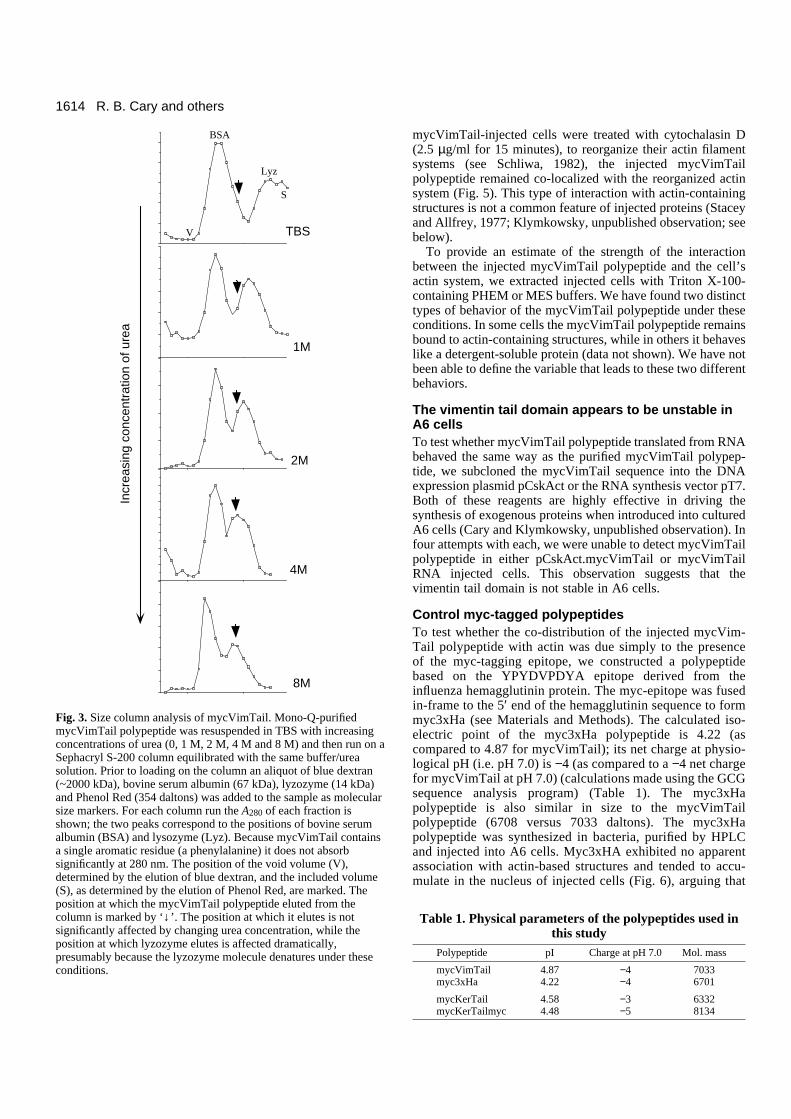
The mycVimTail polypeptide has a calculated molecularmass of 7033 daltons. However, on SDS-polyacrylamide gels,it migrates with an apparent molecular mass of ~12 kDa (Fig.2). A similar sized polypeptide was obtained when thepT7.mycVimTail plasmid was cut with BamHI and used in anin vitro transcription/translation reaction (data not shown). Ona Sephacryl S-200 size exclusion column mycVimTailmigrated with an apparent size of ~15-20 kDa (Fig. 3). Themigration of the mycVimTail polypeptide through a Sephacryl
S-200 column was unaffected by the presence of increasingconcentrations of urea (up to 8 M) (Fig. 3) or 0.5% SDS (datanot shown). In contrast, the position at which lyzozyme (14.3kDa) eluted from the column changed dramatically, it behavedas an increasingly larger structure as the concentration ofdenaturant increased (Fig. 3). Based on this behavior, it wouldappear that the mycVimTail polypeptide is in an extended con-figuration under physiological conditions.
It has been reported that the head domain of desmin existsas a dimer in solution (Saeed and Ip, 1989). Cross-linkingstudies of the mycVimTail polypeptide failed to reveal anyevidence for self-assembly (data not shown). The lack ofeffects of denaturants on the apparent molecular mass (asdetermined by size-exclusion chromatography) (Fig. 3) and theabsence of evidence for oligomers (as determined by cross-linking studies) indicate that the tail domain exists as amonomer in solution.
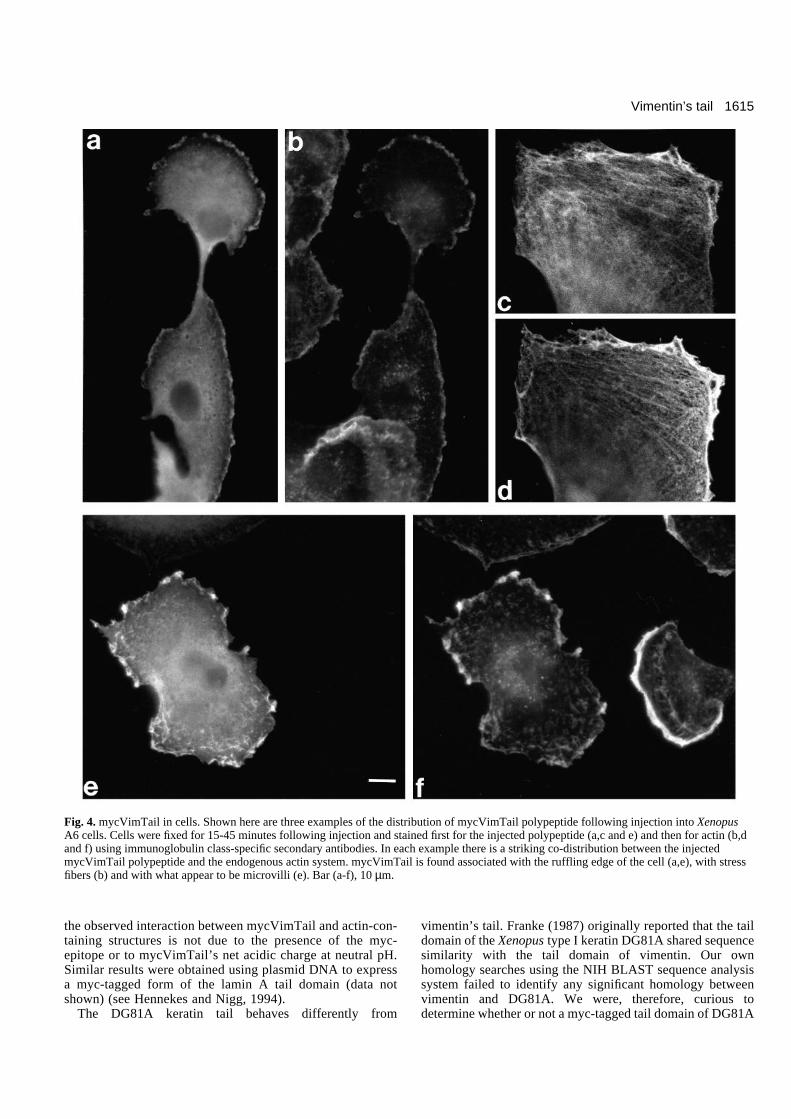
Behavior of tail domains in the cellBased on in vitro data (Georgatos and Blobel, 1987; Georgatoset al., 1987; Djabali et al., 1991; Papamarcaki et al., 1991;Kouklis et al., 1991) the tail domain of vimentin might beexpected to interact with both nuclear lamins and vimentinwithin the living cell. To test this hypothesis we injectedXenopus A6 cells with purified mycVimTail polypeptide. A6cells possess both keratin- and vimentin-type IF networks(Franz et al., 1983; Dent et al., 1989) as well as a lamin LII/LIIIlamina (Krohne et al., 1984). The mycVimTail polypeptideshould be small enough to pass through nuclear pores even inthe absence of a specific nuclear transport signal (see Newportand Forbes, 1987) (see below). To our surprise the injectedmycVimTail polypeptide was excluded from the nucleus;moreover there was no apparent interaction between mycVim-Tail and the cell’s vimentin filament system (Fig. 4). If theinjected cell’s vimentin filament system was ‘collapsed’ bytreatment with nocodazole, we again found little if anyevidence for an interaction between the injected tail polypep-tide and vimentin filaments (data not shown). On the otherhand, there was a striking overlap between the distribution ofthe injected mycVimTail polypeptide and actin (Fig. 4). When
Fig. 2. SDS-PAGE of mycVimTail purification andHPLC purity. (a) Fractions from the purification ofmycVimTail were analyzed by 12% SDS-PAGE.Lane 1, whole bacteria containing pET.vimentin-1.stop; lane 2, molecular mass markers; lane 5, wholebacteria containing pET.mycVimTail; lane 4, materialresuspended following a 2 vol. ethanol precipitation;lane 3, 9E10-reactive material eluting from the Mono-Q column. Molecular mass markers aretriosephosphate isomerase (26.6 kDa), lacticdehydrogenase (36.5 kDa), fumerase (48.5 kDa),
pyruvate kinase (58 kDa), fructose 6-phosphate kinase (84 kDa) and β-galactosidase (116 kDa). (b) TheMono-Q fraction of mycVimTailpolypeptide was assayed for purityby reverse-phase HPLC. A singlemajor A220 peak was resolved; thisfraction contained the 9E10 (anti-myc) reactive material.
1614
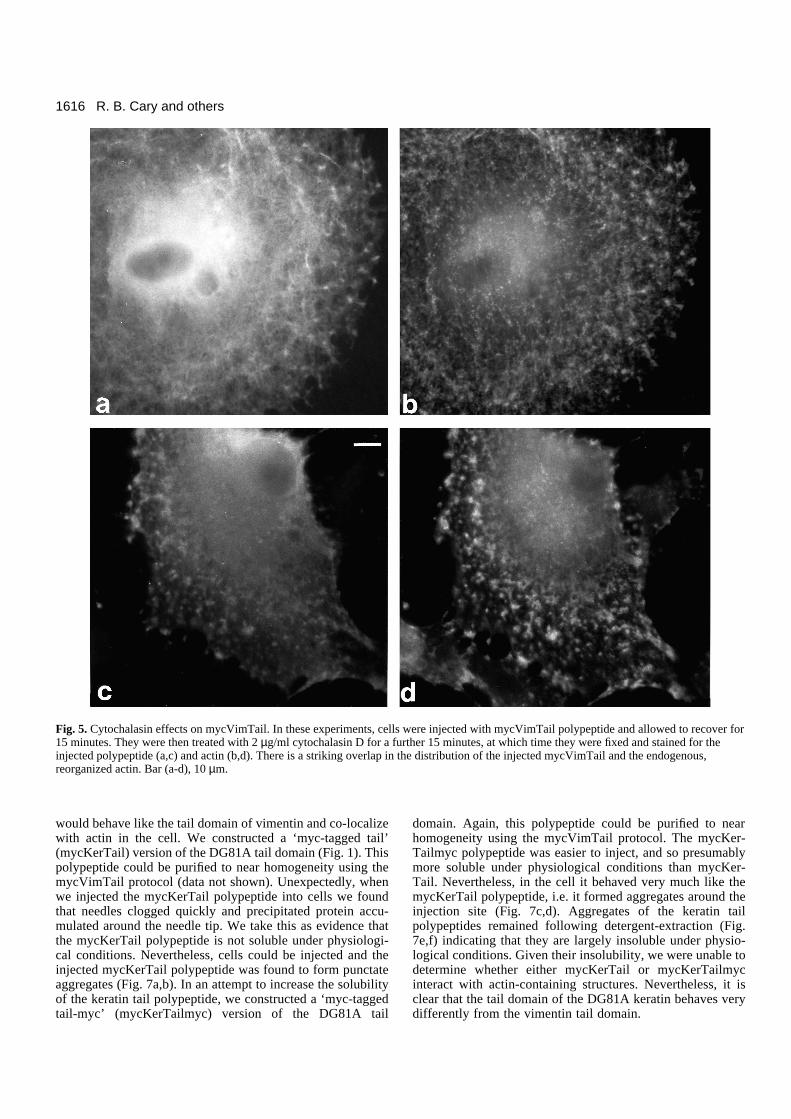
mycVimTail-injected cells were treated with cytochalasin D(2.5 µg/ml for 15 minutes), to reorganize their actin filamentsystems (see Schliwa, 1982), the injected mycVimTailpolypeptide remained co-localized with the reorganized actinsystem (Fig. 5). This type of interaction with actin-containingstructures is not a common feature of injected proteins (Staceyand Allfrey, 1977; Klymkowsky, unpublished observation; seebelow).
To provide an estimate of the strength of the interactionbetween the injected mycVimTail polypeptide and the cell’sactin system, we extracted injected cells with Triton X-100-containing PHEM or MES buffers. We have found two distincttypes of behavior of the mycVimTail polypeptide under theseconditions. In some cells the mycVimTail polypeptide remainsbound to actin-containing structures, while in others it behaveslike a detergent-soluble protein (data not shown). We have notbeen able to define the variable that leads to these two differentbehaviors.
The vimentin tail domain appears to be unstable inA6 cellsTo test whether mycVimTail polypeptide translated from RNAbehaved the same way as the purified mycVimTail polypep-tide, we subcloned the mycVimTail sequence into the DNAexpression plasmid pCskAct or the RNA synthesis vector pT7.Both of these reagents are highly effective in driving thesynthesis of exogenous proteins when introduced into culturedA6 cells (Cary and Klymkowsky, unpublished observation). Infour attempts with each, we were unable to detect mycVimTailpolypeptide in either pCskAct.mycVimTail or mycVimTailRNA injected cells. This observation suggests that thevimentin tail domain is not stable in A6 cells.
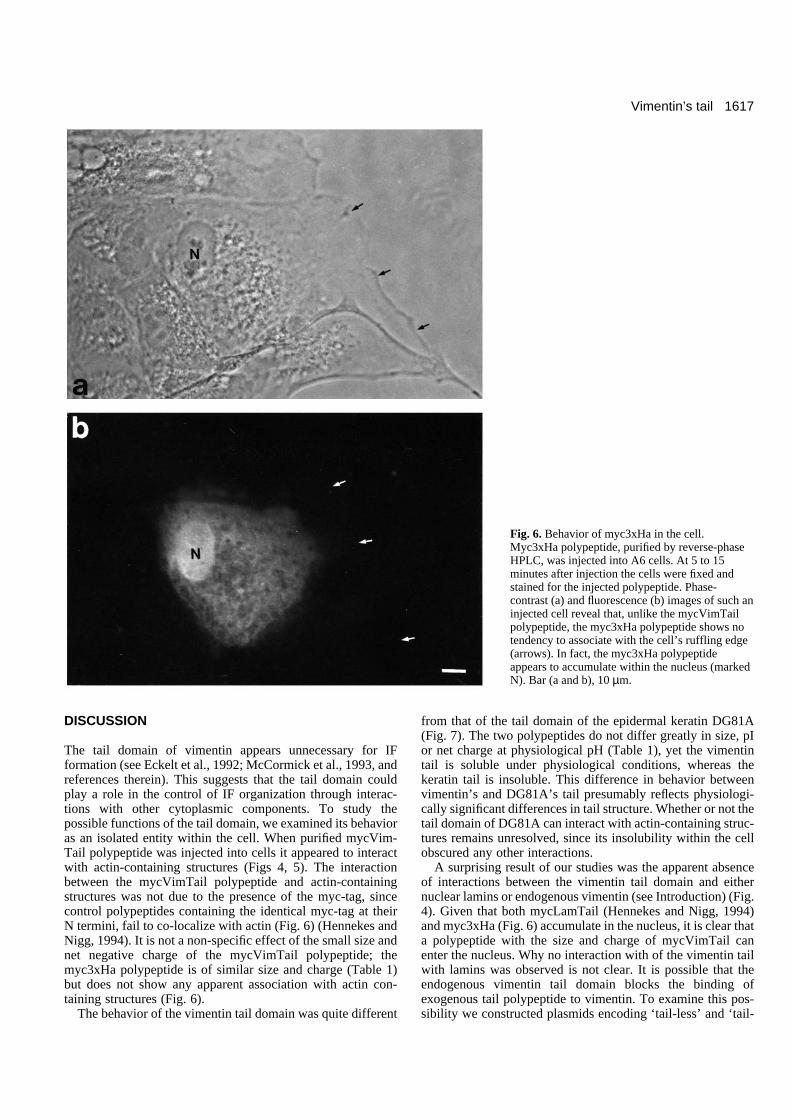
Control myc-tagged polypeptidesTo test whether the co-distribution of the injected mycVim-Tail polypeptide with actin was due simply to the presenceof the myc-tagging epitope, we constructed a polypeptidebased on the YPYDVPDYA epitope derived from theinfluenza hemagglutinin protein. The myc-epitope was fusedin-frame to the 5′ end of the hemagglutinin sequence to formmyc3xHa (see Materials and Methods). The calculated iso-electric point of the myc3xHa polypeptide is 4.22 (ascompared to 4.87 for mycVimTail); its net charge at physio-logical pH (i.e. pH 7.0) is −4 (as compared to a −4 net chargefor mycVimTail at pH 7.0) (calculations made using the GCGsequence analysis program) (Table 1). The myc3xHapolypeptide is also similar in size to the mycVimTailpolypeptide (6708 versus 7033 daltons). The myc3xHapolypeptide was synthesized in bacteria, purified by HPLCand injected into A6 cells. Myc3xHA exhibited no apparentassociation with actin-based structures and tended to accu-mulate in the nucleus of injected cells (Fig. 6), arguing that
R. B. Cary and others
BSA
Lyz
V
S
Incr
easi
ng c
once
ntra
tion
of u
rea
8M
4M
2M
TBS
1M
Fig. 3. Size column analysis of mycVimTail. Mono-Q-purifiedmycVimTail polypeptide was resuspended in TBS with increasingconcentrations of urea (0, 1 M, 2 M, 4 M and 8 M) and then run on aSephacryl S-200 column equilibrated with the same buffer/ureasolution. Prior to loading on the column an aliquot of blue dextran(~2000 kDa), bovine serum albumin (67 kDa), lyzozyme (14 kDa)and Phenol Red (354 daltons) was added to the sample as molecularsize markers. For each column run the A280 of each fraction isshown; the two peaks correspond to the positions of bovine serumalbumin (BSA) and lysozyme (Lyz). Because mycVimTail containsa single aromatic residue (a phenylalanine) it does not absorbsignificantly at 280 nm. The position of the void volume (V),determined by the elution of blue dextran, and the included volume(S), as determined by the elution of Phenol Red, are marked. Theposition at which the mycVimTail polypeptide eluted from thecolumn is marked by ‘↓’. The position at which it elutes is notsignificantly affected by changing urea concentration, while theposition at which lyzozyme elutes is affected dramatically,presumably because the lyzozyme molecule denatures under theseconditions.
Table 1. Physical parameters of the polypeptides used inthis study
Polypeptide pI Charge at pH 7.0 Mol. mass
mycVimTail 4.87 −4 7033myc3xHa 4.22 −4 6701
mycKerTail 4.58 −3 6332mycKerTailmyc 4.48 −5 8134
1615Vimentin’s tail
the observed interaction between mycVimTail and actin-con-taining structures is not due to the presence of the myc-epitope or to mycVimTail’s net acidic charge at neutral pH.Similar results were obtained using plasmid DNA to expressa myc-tagged form of the lamin A tail domain (data notshown) (see Hennekes and Nigg, 1994).
The DG81A keratin tail behaves differently from
vimentin’s tail. Franke (1987) originally reported that the taildomain of the Xenopus type I keratin DG81A shared sequencesimilarity with the tail domain of vimentin. Our ownhomology searches using the NIH BLAST sequence analysissystem failed to identify any significant homology betweenvimentin and DG81A. We were, therefore, curious todetermine whether or not a myc-tagged tail domain of DG81A
Fig. 4. mycVimTail in cells. Shown here are three examples of the distribution of mycVimTail polypeptide following injection into XenopusA6 cells. Cells were fixed for 15-45 minutes following injection and stained first for the injected polypeptide (a,c and e) and then for actin (b,dand f) using immunoglobulin class-specific secondary antibodies. In each example there is a striking co-distribution between the injectedmycVimTail polypeptide and the endogenous actin system. mycVimTail is found associated with the ruffling edge of the cell (a,e), with stressfibers (b) and with what appear to be microvilli (e). Bar (a-f), 10 µm.
1616
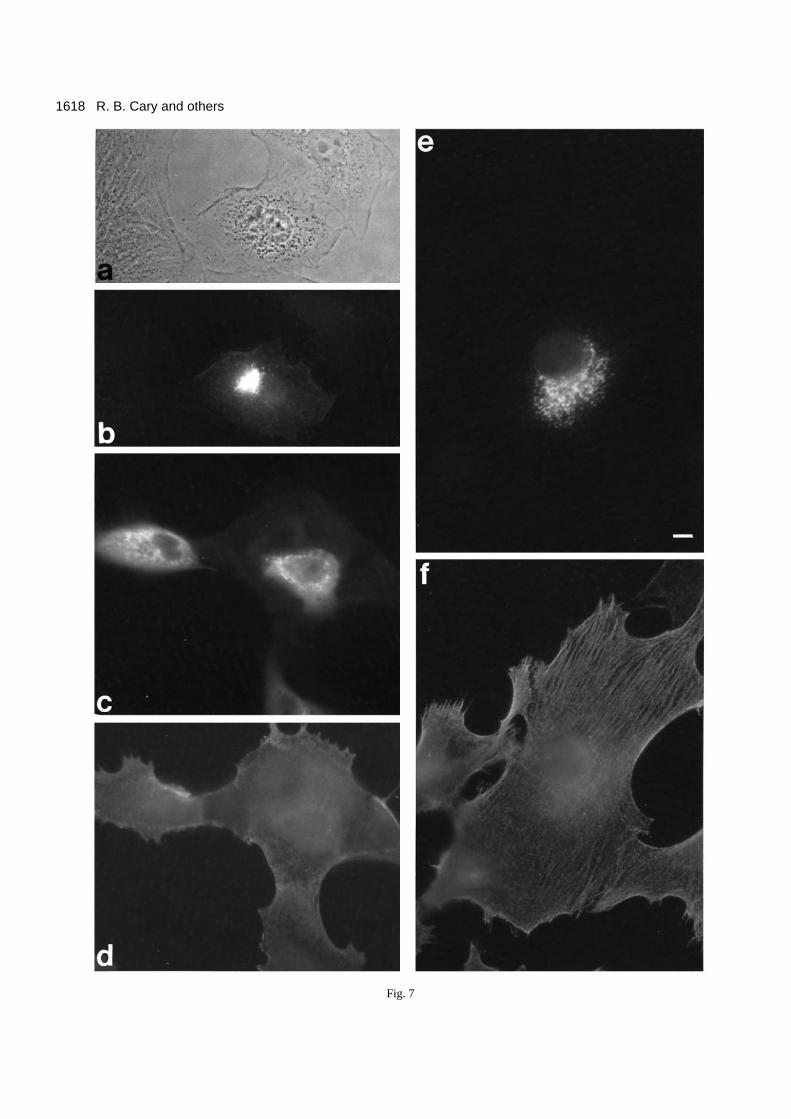
would behave like the tail domain of vimentin and co-localizewith actin in the cell. We constructed a ‘myc-tagged tail’(mycKerTail) version of the DG81A tail domain (Fig. 1). Thispolypeptide could be purified to near homogeneity using themycVimTail protocol (data not shown). Unexpectedly, whenwe injected the mycKerTail polypeptide into cells we foundthat needles clogged quickly and precipitated protein accu-mulated around the needle tip. We take this as evidence thatthe mycKerTail polypeptide is not soluble under physiologi-cal conditions. Nevertheless, cells could be injected and theinjected mycKerTail polypeptide was found to form punctateaggregates (Fig. 7a,b). In an attempt to increase the solubilityof the keratin tail polypeptide, we constructed a ‘myc-taggedtail-myc’ (mycKerTailmyc) version of the DG81A tail
domain. Again, this polypeptide could be purified to nearhomogeneity using the mycVimTail protocol. The mycKer-Tailmyc polypeptide was easier to inject, and so presumablymore soluble under physiological conditions than mycKer-Tail. Nevertheless, in the cell it behaved very much like themycKerTail polypeptide, i.e. it formed aggregates around theinjection site (Fig. 7c,d). Aggregates of the keratin tailpolypeptides remained following detergent-extraction (Fig.7e,f) indicating that they are largely insoluble under physio-logical conditions. Given their insolubility, we were unable todetermine whether either mycKerTail or mycKerTailmycinteract with actin-containing structures. Nevertheless, it isclear that the tail domain of the DG81A keratin behaves verydifferently from the vimentin tail domain.
R. B. Cary and others
Fig. 5. Cytochalasin effects on mycVimTail. In these experiments, cells were injected with mycVimTail polypeptide and allowed to recover for15 minutes. They were then treated with 2 µg/ml cytochalasin D for a further 15 minutes, at which time they were fixed and stained for theinjected polypeptide (a,c) and actin (b,d). There is a striking overlap in the distribution of the injected mycVimTail and the endogenous,reorganized actin. Bar (a-d), 10 µm.
1617Vimentin’s tail
DISCUSSION
The tail domain of vimentin appears unnecessary for IFformation (see Eckelt et al., 1992; McCormick et al., 1993, andreferences therein). This suggests that the tail domain couldplay a role in the control of IF organization through interac-tions with other cytoplasmic components. To study thepossible functions of the tail domain, we examined its behavioras an isolated entity within the cell. When purified mycVim-Tail polypeptide was injected into cells it appeared to interactwith actin-containing structures (Figs 4, 5). The interactionbetween the mycVimTail polypeptide and actin-containingstructures was not due to the presence of the myc-tag, sincecontrol polypeptides containing the identical myc-tag at theirN termini, fail to co-localize with actin (Fig. 6) (Hennekes andNigg, 1994). It is not a non-specific effect of the small size andnet negative charge of the mycVimTail polypeptide; themyc3xHa polypeptide is of similar size and charge (Table 1)but does not show any apparent association with actin con-taining structures (Fig. 6).
The behavior of the vimentin tail domain was quite different
from that of the tail domain of the epidermal keratin DG81A(Fig. 7). The two polypeptides do not differ greatly in size, pIor net charge at physiological pH (Table 1), yet the vimentintail is soluble under physiological conditions, whereas thekeratin tail is insoluble. This difference in behavior betweenvimentin’s and DG81A’s tail presumably reflects physiologi-cally significant differences in tail structure. Whether or not thetail domain of DG81A can interact with actin-containing struc-tures remains unresolved, since its insolubility within the cellobscured any other interactions.
A surprising result of our studies was the apparent absenceof interactions between the vimentin tail domain and eithernuclear lamins or endogenous vimentin (see Introduction) (Fig.4). Given that both mycLamTail (Hennekes and Nigg, 1994)and myc3xHa (Fig. 6) accumulate in the nucleus, it is clear thata polypeptide with the size and charge of mycVimTail canenter the nucleus. Why no interaction with of the vimentin tailwith lamins was observed is not clear. It is possible that theendogenous vimentin tail domain blocks the binding ofexogenous tail polypeptide to vimentin. To examine this pos-sibility we constructed plasmids encoding ‘tail-less’ and ‘tail-
Fig. 6. Behavior of myc3xHa in the cell.Myc3xHa polypeptide, purified by reverse-phaseHPLC, was injected into A6 cells. At 5 to 15minutes after injection the cells were fixed andstained for the injected polypeptide. Phase-contrast (a) and fluorescence (b) images of such aninjected cell reveal that, unlike the mycVimTailpolypeptide, the myc3xHa polypeptide shows notendency to associate with the cell’s ruffling edge(arrows). In fact, the myc3xHa polypeptideappears to accumulate within the nucleus (markedN). Bar (a and b), 10 µm.
1618 R. B. Cary and others
Fig. 7

1619Vimentin’s tail
truncated’ vimentins. In both IF-minus human SW13 clone 2cells (Sarria et al., 1990) and in vimentin-minus human MCF-7 cells (supplied by E. Fuchs, University of Chicago) these tail-deleted vimentins failed to form extended filament networks(Cary and Klymkowsky, unpublished observation) (see Eckeltet al., 1992). It therefore remains possible that the vimentin taildomain does bind to vimentin’s rod domain as suggested byKouklis et al. (1991). On the other hand, the observation thatthis interaction can be disrupted by mutation without apparenteffect on vimentin filament formation or behavior (McCormicket al., 1993) makes the significance of this interaction unclear.
The role of the tail in vimentin filament organizationIn most vimentin-containing cells, vimentin filaments form anextended network that stretches from the vicinity of the nucleusto the cellular periphery (see Goldman et al., 1986). Thisextended vimentin filament network appears to be establishedprimarily through interactions with microtubules (Geiger andSinger, 1980; Ball and Singer, 1981; Gyoeva and Gelfand,1991). Depolymerization of the microtubule system leads tothe collapse of the vimentin network into a dense coil near thecell center (Goldman and Knipe, 1972; Croop and Holtzer,1975; Blose and Chako, 1976; Franke et al., 1978; Hynes andDestree, 1978; Wehland and Willingham, 1983; Blose et al.,1984).
This centripetal collapse of the vimentin filament networkappears to be mediated by the interaction of IFs with themicrofilament system(s) of the cell (Hollenbeck et al., 1989;Tint et al., 1991). Direct interactions between vimentinfilaments and microfilaments/microfilament-associated struc-tures have been reported (Knapp et al., 1983; Magneat andBurridge, 1985; Langley and Cohen, 1987; Granger andLazarides, 1982; Green and Goldman, 1986; Green et al.,1986; Bershadsky et al., 1987). Disruption of normal micro-filament organization also leads to the reorganization of IFs(Croop and Holtzer, 1975).
These observations indicate that IFs interact with microfila-ments; why then are IFs and microfilaments not co-localizedin the cell? The answer would appear to involve the relativestrengths of the interactions between IFs, microtubules andmicrofilaments. The IF-microtubule interaction appears to bedominant, as demonstrated by the co-alignment of the IF andmicrotubule systems in many cell types. An IF-microfilamentinteraction is generally apparent only upon the disruption ofthe microtubule system. In the absence of microtubules,vimentin filaments undergo a microfilament-dependent cen-tripetal collapse (see Hollenbeck et al., 1989; Tint et al., 1991).In addition, the connected nature of the IF itself imposes con-straints on the distribution of IFs within the cell. The isolatedtail domain of vimentin can not form IFs and does not appearto interact with microtubules, so its distribution in the cellreflects the distribution of its highest affinity partner, i.e. themicrofilament system.
Physical properties of the vimentin tail domainOn size-exclusion columns the mycVimTail polypeptidebehaves as a much larger structure than its molecular masswould predict (Fig. 3). Since its behavior is not changed bydenaturants (Fig. 3), it would appear that the mycVimTailpolypeptide has a highly extended configuration in solution.Whether it is also highly extended when it is part of the intactvimentin polypeptide remains to be determined. Solid stateNMR measurements (Mack et al., 1988) suggest that the tailand head domains of keratins are not structured, which wouldbe consistent with an extended configuration. STEM measure-ments of vimentin, vimentin/desmin and keratin filaments(Steven et al., 1983a,b; Engel et al., 1986) suggest that the truediameter of an IF is ~16 nm of which ~8-10 nm is accountedfor by the central portion, which is composed primarily of rod
Fig. 7. Injection of mycKerTail and mycKerTailmyc into cells.Based on the appearance of precipitated material around the injectionneedle tip (not shown), it appears that the mycKerTail polypeptidewas insoluble under physiological conditions. Nevertheless, it waspossible to inject the purified mycKerTail polypeptide into A6 cellsand examine its behavior. After fixation, the cells were stained with9E10 and anti-MIgFl to visualize the injected polypeptide. Theinjected mycKerTail appeared to be concentrated at the injection site(a, phase image; b, fluorescein optics). The addition of a secondmyc-region at the C-terminal end of the polypeptide made thepolypeptide mycKerTailmyc easier to inject into the cell. InjectedmycKerTailmyc was found in punctate aggregate scattered throughout the injected cell (c); double staining with anti-actin antibody (d)showed little overlap between actin and the injected polypeptide.When cells injected with either mycKerTail (not shown) ormycKerTailmyc (e) were extracted with PHEM/T buffer prior tofixation and staining, the injected polypeptide (e) was found to beinsoluble and not apparently associated with the cell’s actin system(f). Bar (a-f), 10 µm.
Fig. 8. A cartoon of the IF↔microfilamentinteraction: based on a recent model for IF structure(Steinert et al., 1993) we have constructed a cartoonof a vimentin filament showing the positions of thetail domains. We assume, for the purposes of thismodel, that both tails project from a dimer in thesame direction and both point outward, i.e. away fromthe filament axis. Interactions between tails andmicrofilaments will, therefore, occur in pairs. It isquite possible that the vimentin tail domain interactsnot with actin, but with actin-associated proteins.

1620
domains. This would leave 6 to 8 nm of the diameter to beoccupied by head and tail domains.
Based on a length of ~0.34 nm/amino acid residue wecalculate the maximum extended length of the vimentin taildomain is ~15 nm. Given STEM mass measurements of intactIFs (see above), it would seem unlikely that the tail is fullyextended; it may well fold back upon itself at least once,perhaps twice. This could place the highly conserved end ofthe tail domain in close proximity to the filament axis and mayexplain why the extreme C-terminal region of the tail domainof desmin is inaccessible to antibody when assembled intofilaments (Birkenberger and Ip, 1990). It could also provide astructural explanation for why mutations in the tail domain inthis highly conserved region could effect filament structure(McCormick et al., 1993). In fact, Franke’s (1987) originalrecognition of conservation in the C-termini of vimentin,desmin, GFAP, and keratin K8 tail domains noted that thissequence motif resembled a ‘loop’ similar to that seen in Ca2+-binding proteins (see also Branden and Tooze, 1991). Such aloop could be involved in the folding of the tail domain (Fig.8). A folded tail would still project ~5-7 nm from the centralaxis of the filament and would be well positioned to mediateinteractions with microfilaments, either directly or throughother actin-binding proteins.
While the tail domain is associated with actin-containingstructures in formaldehyde-fixed cells, in detergent-extractedcells we often found that it behaved as a detergent-solubleprotein. This would imply that the interaction between the taildomain and microfilaments is of relatively low affinity. Thisdoes not mean that within the context of the cell it is not sig-nificant. Based on structural studies, it appears that there are16 IFP subunit dimers per cross-section in an IF (Aebi et al.,1988; Steinert et al., 1993). If the two tails of a vimentindimer both project outward from the filament axis they couldco-operate with one another and with tails elsewhere in thefilament to form more stable interactions with microfila-ments. A number of such low affinity interactions couldmediate the vimentin filament ↔ microfilament interaction(Fig. 8).
We thank Michael Yarus for the use of his FPLC machine; CraigTuerk for sharing his early results on the behavior of the in vitro syn-thesized mycVimTail polypeptide; Tom Sargent for the DG81AcDNA; Doug Melton for the pSP64T plasmid; Richard Harland forpCskAct.LacZ; and Danielle Przychodzin for technical assistance inthe early stages of the project. Sequence analysis was performed atthe NCBI using the BLAST network service; we thank Robin Gutellfor introducing us to BLAST and Mark Winey for carrying out theGCG analysis of tail polypeptides. This work was supported by agrant from the National Science Foundation (DCB91-05523) withadditional support from the Colorado Chapter of the American HeartAssociation and the Council for Research and Creative Work of theUniversity of Colorado, Boulder. A half-year sabbatical granted toM.W.K. by the University of Colorado made this work possible.R.B.C. was supported in part by a training grant from the NIH. R.M.E.was supported by NIH grant HL51850; A.D. was supported by a fel-lowship from the Spanish Ministry of Science; J.A.D. was supportedin part by an NSF pre-doctoral fellowship.
REFERENCES
Aebi, U., Haner, M., Troncoso, J., Eichner, R. and Engel, A. (1988).
Unifying principles in intermediate filament (IF) structure and assembly.Protoplasma 145, 73-81.
Algrain, M., Turunen, O., Vaheri, A., Louvard, D. and Arpin, M. (1993).Ezrin contains cytoskeleton and membrane binding domains accounting forits proposed role as a membrane-cytoskeletal linker. J. Cell Biol. 120, 129-139.
Bader, B. L., Magin, T. M., Freudenmann, M., Stumpp, S. and Franke, W.W. (1991). Intermediate filaments formed de novo from tail-less cytokeratinsin the cytoplasm and in the nucleus. J. Cell Biol. 115, 1293-1307.
Ball, E. H. and Singer, S. J. (1981). Association of microtubules andintermediate filaments in normal fibroblasts and its disruption upontransformation by a temperature-sensitive mutant of Rous sarcoma virus.Proc. Nat. Acad. Sci. USA 78, 6986-6990.
Bershadsky, A. D., Tint, I. S. and Svitkina, T. M. (1987). Association ofintermediate filaments with vinculin-containing adhesion plaques offibroblasts. Cell Motil. Cytoskel. 8, 274-283.
Birkenberger, L. and Ip, W. (1990). Properties of the desmin tail domain:studies using synthetic peptides and antipeptide antibodies. J. Cell Biol. 111,2063-2075.
Blose, S. H. and Chako, S. (1976). Rings of intermediate (100Å) filamentbundles in the perinuclear region of vascular endothelial cells. Theirmobilization by colcemid and mitosis. J. Cell Biol. 70, 459-466.
Blose, S. H., Meltze, D. I. and Feramisco, J. R. (1984). 10-nm filaments areinduced to collapse in living cells microinjected with monoclonal andpolyclonal antibodies against tubulin. J. Cell Biol. 98, 847-859.
Branden, C. and Tooze, J. (1991). Introduction to Protein Structure. NewYork, Garland Press.
Chen, E. Y. and Seeburg, P. H. (1985). Supercoil sequencing: a fast andsimple method for sequencing plasmid DNA. DNA 4, 165-170.
Croop, J. and Holtzer, H. (1975). Response of myogenic and fibrogenic cellsto cytochalasin and colcemid. J. Cell Biol. 65, 271-285.
Cross, G. S., Wilson, C., Erba, H. P. and Woodland, H. R. (1988).Cytoskeletal actin gene families of Xenopus borealis and Xenopus laevis. J.Mol. Evol. 27, 17-28.
Dent, J. A., Polson, A. G. and Klymkowsky, M. W. (1989). A whole-mountimmunocytochemical analysis of the expression of the intermediate filamentprotein vimentin in Xenopus. Development 105, 61-74.
Dent, J. A., Cary, R. B., Bachant, J. B., Domingo, A. and Klymkowsky, M.W. (1992). Host cell factors controlling vimentin organization in theXenopus oocyte. J. Cell Biol. 119, 855-866.
Djabali, K., Portier, M. M., Gros, F., Blobel, G. and Georgatos, S. D.(1991). Network antibodies identify nuclear lamin B as a physiologicalattachment site for peripherin intermediate filaments. Cell 64, 109-121.
Eckelt, A., Herrmann, H. and Franke, W. W. (1992). Assembly of a tail-lessmutant of the intermediate filament protein, vimentin, in vitro and in vivo.Eur. J. Cell Biol. 58, 319-330.
Engel, A., Eichner, R. and Aebi, U. (1986). Polymorphism of reconstitutedhuman epidermal keratin filament: determination of their mass-per-lengthand width by scanning transmission electron microscopy (STEM). J.Ultrastruct. Res. 90, 323-335.
Evan, G. I., Lewis, G. K., Ramsay, G. and Bishop, J. M. (1985). Isolation ofmonoclonal antibodies specific for human c-myc proto-oncogene product.Mol. Cell. Biol. 5, 3610-3616.
Franke, W. W., Schmid, E., Osborn, M. and Weber, K. (1978). Differentintermediate-sized filaments distinguished by immunofluorescencemicroscopy. Proc. Nat. Acad. Sci. USA 75, 5034-5038.
Franke, W. W. (1987). Homology of a conserved sequence in the tail domainof intermediate filament proteins with the loop region of calcium bindingproteins. Cell Biol. Int. Rep. 11, 831.
Franz, J. K., Gall, L., Williams, M. A., Picheral, B. and Franke, W. W.(1983). Intermediate-size filaments in a germ cell: expression of cytokeratinsin oocytes and eggs of the frog Xenopus. Proc. Nat. Acad. Sci. USA 80, 6254-6258.
Geiger, B. and Singer, S. J. (1980). Association of microtubules andintermediate filaments in chicken gizzard cells as detected by doubleimmunofluorescence. Proc. Nat. Acad. Sci. USA 77, 4769-4773.
Geisler, N., Kaufmann, E. and Weber, K. (1985). Antiparallel orientation ofthe double-stranded coiled-coils in the tetrameric protofilament unit ofintermediate filaments. J. Mol. Biol. 182, 173-177.
Geisler, N., Schunemann, J. and Weber, K. (1992). Chemical cross-linkingindicates a staggered and antiparallel protofilament of desmin intermediatefilaments and characterizes one higher-level complex betweenprotofilaments. Eur. J. Biochem. 206, 841-852.
Georgatos, S. D. and Blobel, G. (1987). Two distinct attachment sites for
R. B. Cary and others

1621Vimentin’s tail
vimentin along the plasma membrane and the nuclear envelope in avianerythrocytes: A basis for vectorial assembly of intermediate filaments. J. CellBiol. 105, 105-115.
Georgatos, S. D., Weber, K., Geisler, N. and Blobel, G. (1987). Binding oftwo desmin derivatives to the plasma membrane and the nuclear envelope ofavian erythrocytes: evidence for a conserved site-specificity in intermediatefilament-membrane interactions. Proc. Nat. Acad. Sci. USA 84, 6780-6784.
Goldman, R. D. and Knipe, D. M. (1972). Functions of cytoplasmic fibers innonmuscle cell motility. Cold Spring Harb. Symp. Quant. Biol. 37, 523-534.
Goldman, R. D., Goldman, A. E., Green, K. J., Jones, J. C. R., Jones, S. M.and Yang, H. (1986). Intermediate filament networks: organization andpossible functions of a diverse group of cytoskeletal elements. J. Cell Sci.Suppl. 5, 69-97.
Granger, B. L. and Lazarides, E. (1982). Structural associations of syneminand vimentin filaments in avian erythrocytes revealed by immunoelectronmicroscopy. Cell 30, 263-275.
Green, K. J. and Goldman, R. D. (1986). Evidence for an interaction betweenthe cell surface and intermediate filaments in cultured fibroblasts. Cell Motil.Cytoskel. 6, 389-405.
Green, K. J., Talian, J. C. and Goldman, R. D. (1986). Relationship betweenintermediate filaments and microfilaments in cultured fibroblasts: Evidencefor common foci during cell spreading. Cell Motil. Cytoskel. 6, 406-418.
Gyoeva, F. K. and Gelfand, V. I. (1991). Coalignment of vimentinintermediate filaments with microtubules depends on kinesin. Nature 353,445-448.
Hennekes, H. and Nigg, E. A. (1994). The role of isoprenylation in membraneattachment of nuclear lamins: a single point mutation prevents proteolyticcleavage of the lamin A precursor and confers membrane binding properties.J. Cell Sci. 107, 1019-1029.
Herrmann, H., Fouquet, B. and Franke, W. W. (1989). Expression ofintermediate filament proteins during development of Xenopus laevis. I.cDNA clones encoding different forms of vimentin. Development 105, 279-298.
Herrmann, H., Hofmann, I. and Franke, W. W. (1992). Identification of anonapeptide motif in the vimentin head domain involved in intermediatefilament assembly. J. Mol. Biol. 223, 637-650.
Herrmann, H., Eckelt, A., Brettel, M., Grund, C. and Franke, W. W.(1993). Temperature-sensitive intermediate filament assembly: Alternativestructure of Xenopus laevis vimentin in vitro and in vivo. J. Mol. Biol. 234,99-113.
Hisanaga, S. and Hirokawa, N. (1989). The effects of dephosphorylation onthe structure of the projections of neurofilament. J. Neurosci. 9, 959-966.
Hofmann, I. and Herrmann, H. (1992). Interference in vimentin assembly invitro by synthetic peptides derived from the vimentin head domain. J. CellSci. 101, 687-700.
Hollenbeck, P. J., Bershadsky, A. D., Pletjushkina, O. Y., Tint, I. S. andVasiliev, J. M. (1989). Intermediate filament collapse is an ATP-dependentand actin-dependent process. J. Cell Sci. 92, 621-631.
Hynes, R. O. and Destree, A. T. (1978). 10nm filaments in normal andtransformed cells. Cell 13, 151-163.
Ip, W., Hartzer, M. K., Pang, S. and Robson, R. M. (1985). Assembly ofvimentin in vitro and its implications concerning the structure of intermediatefilaments. J. Mol. Biol. 183, 365-375.
Jonas, E., Sargent, T. D. and Dawid, I. B. (1985). Epidermal keratin geneexpressed in embryos of Xenopus laevis. Proc. Nat. Acad. Sci. USA 82,5413-5417.
Kaufmann, E., Weber, K. and Geisler, N. (1985). Intermediate filamentforming ability of desmin derivatives lacking either the amino-terminal 67 orthe carboxyl-terminal 27 residues. J. Mol. Biol. 185, 733-742.
Klymkowsky, M. W. (1981). Intermediate filaments in 3T3 cells collapse afterintracellular injection of a monoclonal anti-intermediate filament antibody.Nature 291, 249-251.
Knapp, L. W., O’Guin, W. M. and Sawyer, R. H. (1983). Drug-inducedalterations of cytokeratin organization in cultured epithelial cells. Science219, 501-503.
Kouklis, P. D., Papamarcaki, T., Merdes, A. and Georgatos, S. D. (1991). Apotential role for the COOH-terminal domain in the lateral packing of type IIIintermediate filaments. J. Cell Biol. 114, 773-786.
Krohne, G., Debus, E., Osborn, M., Weber, K. and Franke, W. W. (1984).A monoclonal antibody against nuclear lamina proteins reveals cell-typespecificity in Xenopus laevis. Exp. Cell Res. 150, 47-59.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227, 680-683.
Langley, R. C. and Cohen, C. M. (1987). Cell type-specific association
between two types of spectrin and two types of intermediate filaments. CellMotil. Cytoskel. 8, 165-173.
Lu, X. and Lane, E. (1990). Retrovirus-mediated transgenic keratin expressionin cultured fibroblasts: specific domain functions in keratin stabilization andfilament formation. Cell 62, 681-696.
Mack, J. W., Torchia, D. A. and Steinert, P. M. (1988). Solid-state NMRstudies of the dynamics and structure of mouse keratin intermediatefilaments. Biochemistry 27, 5418-5426.
Magneat, P. H. and Burridge, K. (1985). Immunoprecipitation of non-erythrocyte spectrin within live cells following microinjection of specificantibodies: relation to cytoskeletal structures. J. Cell Biol. 98, 1363-1377.
McCormick, M. B., Kouklis, P., Syder, A. and Fuchs, E. (1993). The roles ofthe rod end and the tail in vimentin IF assembly and IF network formation. J.Cell Biol. 122, 395-407.
Newport, J. W. and Forbes, D. J. (1987). The nucleus: structure, function, anddynamics. Annu. Rev. Biochem. 56, 535-565.
Papamarcaki, T., Kouklis, P. D., Kreis, T. E. and Georgatos, S. D. (1991).The ‘lamin B-fold’. Anti-idiotypic antibodies reveal a structuralcomplementarity between nuclear lamin B and cytoplasmic intermediatefilament epitopes. J. Biol. Chem. 266, 21247-21251.
Parry, D. A. and Steinert, P. M. (1992). Intermediate filament structure. Curr.Opin. Cell Biol. 4, 94-98.
Potschka, M., Nave, R., Weber, K. and Geisler, N. (1990). The two coiledcoils in the isolated rod domain of the intermediate filament protein desminare staggered. A hydrodynamic analysis of tetramers and dimers. Eur. J.Biochem. 190, 503-508.
Quinlan, R. A., Moir, R. D. and Stewart, M. (1989). Expression inEscherichia coli of fragments of glial fibrillary acidic protein:characterization, assembly properties and paracrystal formation. J. Cell Sci.93, 71-83.
Raats, J. M., Henderik, J. B., Verdijk, M., van, Oort, F. L. G., Gerards, W.L., Ramaekers, F. C. and Bloemendal, H. (1991). Assembly of carboxy-terminally deleted desmin in vimentin-free cells. Eur. J. Cell Biol. 56, 84-103.
Roof, D. M., Meluh, P. B. and Rose, M. D. (1992). Kinesin-related proteinrequired for assembly of the mitotic spindle. J. Cell Biol. 118, 95-108.
Sarria, A. J., Nordeen, S. K. and Evans, R. M. (1990). Regulated expressionof vimentin cDNA in cells in the presence and absence of a preexistingvimentin filament network. J. Cell Biol. 111, 553-565.
Saeed, T. and Ip, W. (1989). Assembly properties of two CNBr fragments ofavian desmin that correspond to the headpiece domain and helix 1B.Biochem. Biophys. Res. Commun. 165, 1059-1066.
Schliwa, M. and van Blerkom, J. (1981). Structural interaction of cytoskeletalcomponents. J. Cell Biol. 90, 222-235.
Schliwa, M. (1982). Action of cytochalasin D on cytoskeletal networks. J. CellBiol. 92, 79-91.
Shoeman, R., Mothes, E., Kesselmeier, C. and Traub, P. (1990).Intermediate filament assembly and stability in vitro: effect and implicationsof the removal of head and tail domains of vimentin by humanimmunodeficiency virus type I protease. Cell Biol. Int. Rep. 14, 583-594.
Smith, W. C. and Harland, R. M. (1991). Injected Xwnt-8 RNA acts early inXenopus embryos to promote formation of a vegetal dorsalizing center. Cell67, 753-765.
Stacey, D. W. and Allfrey, V. G. (1977). Evidence for the autophagy ofmicroinjected proteins in HeLa cells. J. Cell Biol. 75, 807-817.
Steinert, P. M. and Roop, D. R. (1988). Molecular and cellular biology ofintermediate filaments. Annu. Rev. Cell Biol. 57, 593-625.
Steinert, P. M. (1991). Organization of coiled-coil molecules in native mousekeratin 1/keratin 10 intermediate filaments: evidence for alternating rows ofantiparallel in-register and antiparallel staggered molecules. J. Struct. Biol.107, 157-174.
Steinert, P. M., Marekov, L. N., Fraser, R. D. and Parry, D. A. (1993).Keratin intermediate filament structure. Crosslinking studies yieldquantitative information on molecular dimensions and mechanism ofassembly. J. Mol. Biol. 230, 436-452.
Steven, A. C., Hainfeld, J. F., Trus, B. L., Wall, J. S. and Steinert, P. M.(1983a). The distribution of mass in heteropolymer intermediate filamentsassembled in vitro. Stem analysis of vimentin/desmin and bovine epidermalkeratin. J. Biol. Chem. 258, 8323-8329.
Steven, A. C., Hainfeld, J. F., Trus, B. L., Wall, J. S. and Steinert, P. M.(1983b). Epidermal keratin filaments assembled in vitro have masses-per-unit-length that scale according to average subunit mass: structural basis forhomologous packing of subunits in intermediate filaments. J. Cell Biol. 97,1939-1944.

1622
Studier, F. W., Rosenberg, A. H., Dunn, J. J. and Dubendorff, J. W. (1990).Use of T7 RNA polymerase to direct the expression of cloned genes. Meth.Enzymol. 185, 60-89.
Tint, I. S., Hollenbeck, P. J., Verkhovsky, A. B., Surgucheva I. G., andBershadsky, A. D. (1991). Evidence that intermediate filamentreorganization is induced by ATP-dependent contraction of the actomyosincortex in permeabilized fibroblasts. J. Cell Sci. 98, 375-384.
Traub, P., Scherbarth, A., Wiegers, W. and Shoeman, R. L. (1992). Saltstable interaction of the amino-terminal head region of vimentin with the α-helical rod domain of cytoplasmic intermediate filament proteins and itsrelevance to protofilament structure and filament formation and stability. J.Cell Sci. 101, 363-381.
van de Klundert, F. A. J. M., Raats, J. M. H. and Bloemendal, H. (1993).Intermediate filaments: regulation of gene expression and assembly. Eur. J.Biochem. 214, 351-366.
Vize, P. D., Melton, D. A., Hemmati-Brivanlou, A. and Harland, R. M.(1991). Assays for gene function in developing Xenopus embryos. Xenopuslaevis: Practical Uses in Cell and Molecular Biology. Academic Press. 367-387.
Wehland, J. and Willingham, M. C. (1983). A rat monoclonal antibodyreacting specifically with the tyrosylated form of α-tubulin. II. Effects on cellmovement, organization of microtubules, and intermediate filaments, andarrangement of Golgi elements. J. Cell Biol. 97, 1476-1490.
Wilson, A. K., Coulombe, P. A. and Fuchs, E. (1992). The roles of K5 andK14 head, tail, and R/KLLEGE domains in keratin filament assembly invitro. J. Cell Biol. 119, 401-414.
(Received 27 December 1993 - Accepted 15 February 1994)
R. B. Cary and others






![CYTOSKELETON NEWS - fnkprddata.blob.core.windows.net · Dynamic remodeling of the actin cytoskeleton [i.e., rapid cycling between filamentous actin (F-actin) and monomer actin (G-actin)]](https://img.pdfslide.us/doc/110x75/609edd2b88630103265d18ee/cytoskeleton-news-dynamic-remodeling-of-the-actin-cytoskeleton-ie-rapid-cycling.jpg)









![Heavy and light roles: myosin in the morphogenesis of the ... · myosin so that it interacts with actin [17]. At the N-ter-minus, each MHC folds onto itself forming a globular head](https://img.pdfslide.us/doc/110x75/609836677fc95b2dbd12fb1d/heavy-and-light-roles-myosin-in-the-morphogenesis-of-the-myosin-so-that-it.jpg)

![Review Actin-targeting natural products: structures ... · actin-binding proteins actively break or ‘sever’ actin filaments [e.g. actin-depolymerizing factor (ADF) and cofilin]](https://img.pdfslide.us/doc/110x75/5f0f85bd7e708231d44494d0/review-actin-targeting-natural-products-structures-actin-binding-proteins-actively.jpg)










