Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, OCt. 1990, p. 3375-3379 Vol. 58, No. 100019-9567/90/103375-05$02.00/0Copyright C 1990, American Society for Microbiology
Vibrio cholerae HlyA Hemolysin Is Processed by ProteolysisROBERT H. HALL'* AND BOHUMIL S. DRASAR2
Center for Vaccine Development, Division of Geographic Medicine, Departments of Medicine andBiological Chemistry, University of Maryland School of Medicine, 10 South Pine St.,
Baltimore, Maryland 21201,1 and Department of Clinical Sciences,London School of Hygiene and Tropical Medicine,
London WCIE 7HT, England2
Received 21 May 1990/Accepted 22 July 1990
The leukocidal activity of the Vibrio cholerae hemolysin (HlyA) was utilized to detect, enrich, and clonehybridoma ceUs expressing neutralizing monoclonal antibody in a new survivor selection protocol. A bank of550 hybridoma clones was obtained from a mouse immunized with hemolysin by using standard techniques.The hybridoma bank was treated with a dose of HlyA hemolysin lethal to nonimmune clones. Five survivinghybridoma clones (Xl through X5) which possessed anti-HlyA activity were obtained. Western immunoblotanalysis of V. cholerae culture supernatants with monoclonal antibody from clone Xl identified proteins withMrs of 83,200, 71,600, and 60,300. Amino-terminal sequence analysis of the 71,600 Mr and 60,300-Mr formsshowed homology with the published predicted sequence of ElyA. Our data indicate that proteolytic cleavageoccurs between residues 120 and 121 (Glu-Leu) of the 83,200-Mr form, producing the 71,600-Mr form with theterminus NH2-L-L-F-T-P-F-D-Q-A-E-E-. Cleavage between residues 150 and 151 (Gly-Phe) releases the60,300-Mr form with the terminus NH2-F-A-S-P-A-P-A-N-S-E-. Calculations based on the DNA sequence andthe N termini indicated that the actual molecular masses of the 83,200-, 71,600-, and 60,300-M, forms were,respectively, 79.4 kilodaltons (kDa), 68.6 kDa, and 65.3 kDa. Survivor selection and amino-terminalmicrosequencing offer powerful tools for the analysis of leukotoxic agents.
Pathogenic strains of the bacterial genus Vibrio causegastrointestinal disorders with various degrees of severity,depending on the array of virulence determinants which theyexpress. The life-threatening diarrheal purges of choleragravis are associated with Vibrio cholerae strains expressingthe 01 antigen, the ToxR global regulation element, choleratoxin, and toxin-coregulated pili (8). Many Vibrio strainswhich cannot express cholera toxin have been isolated frompatients suffering from diarrhea, and in these cases thesecretogenic factors remain unidentified (16).One putative virulence factor is an extracellular cytolytic
protein known as the El Tor hemolysin. Although first notedfor its lytic effect on erythrocytes (hemolysis), cytolyticactivity has also been demonstrated on Vero and othermammalian cells in culture (9). Efforts to characterize the ElTor hemolysin have resulted in apparently contradictorybiochemical data among analyses of purified hemolysin andcloned hemolytic determinants (4, 6, 9, 13, 14, 21). Forexample, a gene, hlyA, implicated as the El Tor hemolysinstructural gene, has been mapped, and a translated productof hlyA has been identified as a protein with an Mr of 80,000to 84,000 (4, 5, 13). These data contrast with analyses ofpurified hemolysin, which migrates as a single subunit pro-tein species with an Mr of -60,000 (18, 20).The relationship between the El Tor hemolysin and hlyA is
of considerable significance to the study of vibrio pathogen-esis and vaccine development. Several candidate vaccinestrains have been derived by generating deletion or insertionmutations in this locus, so it is clearly important that theprotein encoded by hlyA be identified positively.We investigated several well-characterized strains of V.
cholerae by using monoclonal antibody (MAb) and amino-terminus microsequencing to reconcile these apparent con-
* Corresponding author.
tradictions. We report a model of posttranslational modifi-cation in the maturation of hemolysin.
MATERIALS AND METHODSBacterial strains and growth conditions. The V. cholerae
strains used in this study and their biotypes, serotypes,sources, and cytolytic activities are listed in Table 1. Bacte-ria were grown as described previously (6, 18) in nutrientbroth, on nutrient agar, or on nutrient agar containingwashed sheep erythrocytes (1%, vollvol).
Assays for lytic activity. The assays for cytolytic andneutralization activity were as described elsewhere (3, 6, 9).Essentially, serial twofold dilutions were made in 96-wellmicrotiter plates, and 100-pJ volumes were added to con-fluent monolayers of Vero cells. One Vero lytic unit wasdefined as the minimum amount required to lyse 100% of theVero monolayer after 16 h of incubation at 37°C. A rapidassay using radial diffusion in blood agar provided accurateestimates of hemolytic activity. In this assay, blood agarplates were poured containing 1% (wt/vol) agarose, 1%(vol/vol) sheep erythrocytes, phosphate-buffered saline, and0.05% (wtlvol) NaN3. Wells of 4-mm diameter were cut inthe blood agar, and 20-pul samples of known Vero lyticactivity were added. After 5 h of incubation at 37°C, thediameters of the zones of clearing around the wells weremeasured and plotted as a calibration curve. Samples ofunknown activity were measured by radial diffusion, and theVero lytic activity was determined from the calibrationcurve ( 6, 18).
Purification of hemolysin. Crude preparations of hemolysinwere obtained from V. cholerae 8731 by filter sterilizingculture supernatants with a 0.22-pum-pore-size asbestos filter(no longer commercially available; may be substituted with alow-protein-binding filter). Activity was followed by usingthe rapid radial diffusion hemolysis assay (6, 18). Hemolysinwas purified by sequential steps of ammonium sulfate pre-
3375

3376 HALL AND DRASAR
TABLE 1. Strains of V. cholerae possessing the hemolyticphenotype used in this study
Reactivity of proteinStrain Source Serogroup with M/'
60,300 71,600 83,200
8731 Environment 01 + + +1196 Environment 01 + - -1074 Environment 01 + + +N16961 Clinical 01 +E7946 Clinical 01 + +S7 Clinical Non-O1 +2076 Clinical Non-O1 + - +N86 Clinical 01 + + +017 Clinical 01 + + +
a Reactivity with antihemolysin MAb.
cipitation (80% saturation), isoelectric focusing, ion ex-change, and molecular exclusion chromatography, as de-scribed by Sutton et al. (18). When samples were analyzedby sodium dodecyl sulfate-polyacrylamide gel electrophore-sis (SDS-PAGE) and staining with Coomassie blue (7), purehemolysin migrated as a single band with an Mr of 60,300(Fig. 1), in close agreement with previous reports (6, 18, 20).
Preparation of hybridoma cell fusions. BALB/c mice weresubcutaneously inoculated with three 50-,ug doses of purifiedhemolysin in Freund incomplete adjuvant at 14-day intervalsand boosted with an intravenous injection of 50 ,ug ofhemolysin 3 days before fusion. Published methods (2) wereused to fuse splenocytes to Sp2/0 parental myeloma cells inthe presence of polyethylene glycol. After the fusion step,cells were suspended in culture medium supplemented withhypoxanthine, aminopterine, and thymidine to select forhybridoma fusions. Portions of the cells were placed in 1096-well microtiter plates, incubated, and fed as previouslydescribed (2). When confluent, the bank of hybridomas was
copied by subculture and the original was stored frozen at-700C.Survivor selection of immune hybridoma clones. The sen-
sitivity of nonimmune hybridoma cells was determined by
a b c d e f g h i j k
483 2- 471 6
_4603
FIG. 1. Coomassie blue-stained SDS-PAGE (lanes a and b) (10%total acrylamide) of the culture supernatant of V. cholerae 8731before (lane a) and after (lane b) purification of hemolysin. Purifiedhemolysin (Mr of 60,300) was used to immunize BALB/c mice forproduction of hybridomas by survivor selection. Western blot assaywith MAb (lanes c through k) was conducted against strains 8731 (c),1196 (d), 1074 (e), N16961 (f), E7946 (g), S7 (h), 2076 (i), N86 (j), and017 (k). Mr values of immunoreactive proteins were calculatedrelative to P-galactosidase (116.3 kDa), phosphorylase b (92.5 kDa),bovine serum albumin (66.2 kDa), and ovalbumin (45 kDa) (Sigma).
titrating a hemolysin sample of known cytolytic activityagainst confluent monolayers of Vero and nonimmune hy-bridoma cells in parallel, as described above. After overnightincubation, treated cells were fixed and stained with Giemsa.Vero and hybridoma cells showed equal sensitivity to hemo-lysin (data not shown). Two copies of the hybridoma bankwere grown to confluence and treated with either purehemolysin or a concentrated crude culture supernatant of V.cholerae 8731 diluted to an activity of 100 Vero lytic units in100 [lI of hybridoma culture medium. Surviving clones wereidentified visually by their capacity to acidify the culturemedium after overnight incubation, indicated by the phenolred pH dye turning yellow. Survivors were fed and cloned bysurvivor selection as described above. When confluent (ifterabout 2 weeks), the clones were retreated with 100 Vero lyticunits of pure or crude hemolysin. Surviving clones wereexpanded, and culture supernatants (50-pdl samples) wereassayed for the ability to neutralize 100 Vero lytic units inthe Vero tissue culture neutralization assay, as previouslydescribed (3). Another sample of hybridoma supernatant (10Pdl) was incubated with cytotoxin (500 Vero lytic units in 10,ul of hybridoma culture medium) and assayed by radialdiffusion to confirm neutralization.Sample preparation for Western immunoblotting and ami-
no-terminal sequencing. Culture supernatants were obtainedfrom several strains of V. cholerae by centrifugation (10,000x g for 15 min). Trichloroacetic acid was added (to a finalconcentration of 10% [wt/vol]), and supernatants were incu-bated on ice for 30 min. Acid-precipitated proteins werecollected by centrifugation, washed with ethanol-ether (50:50, vol/vol), boiled in sample buffer (7), and stored at -20°C.The subsequent steps for protein staining or Western blot-ting (by using gels of the Laemmli formulation [7]) oramino-terminal sequencing (by using gels of the Jovin for-mulation [15]) were all conducted on samples prepared inthis way.
Gel electrophoresis and Western blotting. SDS-PAGE (10%total acrylamide) was conducted by using standard equip-ment and techniques (7). Western blot analysis with hybrid-oma supernatants (diluted 1/10) and alkaline phosphatase-conjugated goat anti-mouse immunoglobulin G (SigmaChemical Co., St. Louis, Mo.) was performed as previouslydescribed (1, 10, 19) but with the following modifications.Proteins were electroblotted onto nitrocellulose filters(0.2-,um pore size; Sartorius, Hayward, Calif.), and filterswere incubated in a blocking solution, defined as 0.05%(vol/vol) polyethylene sorbitan monolaurate (Tween 20) (Sig-ma), phosphate-buffered saline (Oxoid, Basingstoke, En-gland) (Dulbecco A), and 0.05% (wt/vol) NaN3, to preventnonspecific adsorption of MAb or conjugate antibody.
Amino-terminal sequence analysis. Amino-terminal micro-sequence analysis from SDS-PAGE was conducted by usingthe method and reagents described by Moos et al. (15), withthe following exceptions. Stock acrylamide solutions were
prepared immediately prior to use, TEMED (N,N,N',N',-tetramethylethylenediamine) was kept for no longer thanthree weeks, and the steps of staining and destaining poly-vinylidene difluoride membranes were accomplished in two5-min periods. These modifications minimized the exposureof protein to acrylic acid, to free radicals, and to reactiveorganics.
RESULTS AND DISCUSSION
Neutralizing MAb was prepared to the pure 60,300-Mrhemolysin form to permit the identification of precursor
INFECT. IMMUN.
VIBRIO CHOLERAE HlyA CYTOLYSIN 3377
A
B
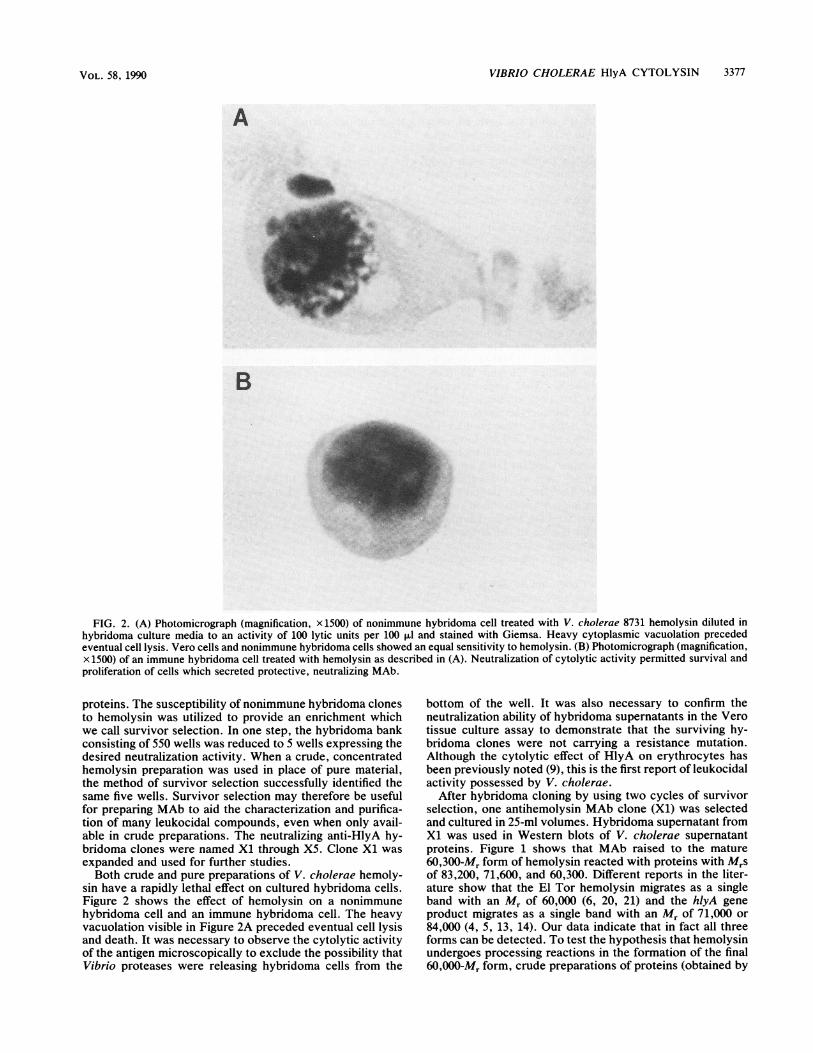
FIG. 2. (A) Photomicrograph (magnification, x 1500) of nonimmune hybridoma cell treated with V. cholerae 8731 hemolysin diluted inhybridoma culture media to an activity of 100 lytic units per 100 p1l and stained with Giemsa. Heavy cytoplasmic vacuolation precededeventual cell lysis. Vero cells and nonimmune hybridoma cells showed an equal sensitivity to hemolysin. (B) Photomicrograph (magnification,x 1500) of an immune hybridoma cell treated with hemolysin as described in (A). Neutralization of cytolytic activity permitted survival andproliferation of cells which secreted protective, neutralizing MAb.
proteins. The susceptibility of nonimmune hybridoma clonesto hemolysin was utilized to provide an enrichment whichwe call survivor selection. In one step, the hybridoma bankconsisting of 550 wells was reduced to 5 wells expressing thedesired neutralization activity. When a crude, concentratedhemolysin preparation was used in place of pure material,the method of survivor selection successfully identified thesame five wells. Survivor selection may therefore be usefulfor preparing MAb to aid the characterization and purifica-tion of many leukocidal compounds, even when only avail-able in crude preparations. The neutralizing anti-HlyA hy-bridoma clones were named Xl through X5. Clone Xl wasexpanded and used for further studies.Both crude and pure preparations of V. cholerae hemoly-
sin have a rapidly lethal effect on cultured hybridoma cells.Figure 2 shows the effect of hemolysin on a nonimmunehybridoma cell and an immune hybridoma cell. The heavyvacuolation visible in Figure 2A preceded eventual cell lysisand death. It was necessary to observe the cytolytic activityof the antigen microscopically to exclude the possibility thatVibrio proteases were releasing hybridoma cells from the
bottom of the well. It was also necessary to confirm theneutralization ability of hybridoma supernatants in the Verotissue culture assay to demonstrate that the surviving hy-bridoma clones were not carrying a resistance mutation.Although the cytolytic effect of HlyA on erythrocytes hasbeen previously noted (9), this is the first report of leukocidalactivity possessed by V. cholerae.
After hybridoma cloning by using two cycles of survivorselection, one antihemolysin MAb clone (Xl) was selectedand cultured in 25-ml volumes. Hybridoma supernatant fromXl was used in Western blots of V. cholerae supernatantproteins. Figure 1 shows that MAb raised to the mature60,300-Mr form of hemolysin reacted with proteins with Mrsof 83,200, 71,600, and 60,300. Different reports in the liter-ature show that the El Tor hemolysin migrates as a singleband with an Mr of 60,000 (6, 20, 21) and the hlyA geneproduct migrates as a single band with an Mr of 71,000 or84,000 (4, 5, 13, 14). Our data indicate that in fact all threeforms can be detected. To test the hypothesis that hemolysinundergoes processing reactions in the formation of the final60,000-Mr form, crude preparations of proteins (obtained by
VOL. 58, 1990

3378 HALL AND DRASAR
trichloroacetic acid precipitation of culture supernatants)were fractionated by SDS-PAGE, using the Jovin formula-tion described by Moos et al. (15). The amino-terminalsequence of bands identified by survivor-selected antihemol-ysin X1 MAb was determined by transblotting from JovinSDS gels onto polyvinylidene difluoride membrane supportsand sequencing excised bands shown in Western blots toreact with antihemolysin X1 MAb. Amino-terminal microse-quence analysis of the 71,600-Mr form revealed the sequenceNH2-Leu-Leu-Phe-Thr-Pro-Phe-Asp-Gln-Ala-Glu-Glu-.Amino-terminal microsequence analysis of the 60,300-Mrform yielded the sequence NH2-Phe-Ala-Ser-Pro-Ala-Pro-Ala-Asn-Ser-Glu-.The two decapeptide sequences which we identified from
immunoreactive proteins corresponded exactly with pre-dicted amino acid sequences internal to the hlyA gene fromV. cholerae, as reported by Rader and Murphy (17) andYamamoto et al. (21). Our data suggest the following modelof posttranslational proteolytic processing. The 83,200-MrhlyA gene product is processed to the 71,600-M, form byproteolytic cleavage between residues 120 and 121 (Glu-Leu). The 71,600-Mr form or the 83,200-Mr form is cleavedbetween residues 150 and 151 (Gly-Phe) to release the finalform with an Mr of 60,300. Localizing the amino terminiwithin the predicted sequence of the hlyA gene product (17)shows that the 60,300-, 71,600-, and 83,200-Mr forms areencoded by the same gene. Predicting the actual molecularmass from the N termini (this paper) and the DNA sequence(17) suggests that the 83,200-Mr form is 79.4 kilodaltons(kDa), the 71,600-Mr form is 68.6 kDa, and the 60,300-Mrform is 65.3 kDa. The discrepancy between the Mr valuesdetermined by SDS-PAGE and the kDa values determinedby summation of the molecular masses of the amino acidsfrom the predicted primary structure could be due to unde-tected carboxy-terminus processing or anomalous migrationin the gel systems utilized.
It is conceivable that the 71,600- and 84,000-Mr proteinspreviously reported from maxicell experiments (4, 5, 13)have not been purified from V. cholerae because of rapidproteolytic cleavage during purification. The fully mature60,300-Mr form may not have been detected during geneticstudies possibly because the cleavage reaction requiresspecific protease activity absent in the Escherichia colistrains used for cloning experiments. It is notable nonethe-less that hemolytic activity has been used by several inves-tigators to screen gene banks of V. cholerae DNA in E. colias a means of identifying and cloning hlyA, demonstratingthat hlyA in an E. coli background supports detectablehemolytic activity. The method of survivor selection pro-vides a means to detect immunologically cross-reactiveprecursors, even if such precursors do not possess leuko-cidal activity.
Correlating the amino-terminal sequences of the immuno-reactive protein bands with the nucleotide sequence demon-strates for the first time that the El Tor hemolysin is, in fact,encoded by hlyA. Several V. cholerae strains expressedhemolysin precursors which were detected by Western blots(Table 1), although in some cases with slightly different Mrvalues (Fig. 1). The three bands described above weredetected in strains N86, 017, 1074, and 8731 (Fig. 1). Strain017 was the strain from which the gene hlyA was originallycloned and characterized by minicell analysis (13). StrainsN86 and 8731 were, respectively, the sources of the 60-kDahemolysin reported by Yamamoto et al. (20) and Hall et al.(6; this paper). We presume that the observed differences inthe banding patterns between strains are a consequence of
differences in the sequence of the structural gene or differ-ences in processing. The published sequences of hlyA fromthree different strains show some heterogeneity, which mayaccount for the differences in the Mr reported in Fig. 1.
V. cholerae 01 strains can express several proteolyticenzymes which might catalyze the cleavages of HlyA (22). Itshould be interesting to investigate the regulation, expres-sion, and specificity of proteases involved in processingcholera toxin and HlyA.The role of the HlyA El Tor hemolysin in the biology of V.
cholerae remains unclear. Expression of hlyA does notcorrelate with disease symptoms; there are both virulentHlyA- strains (e.g., classical biotype strains) and avirulentHlyA+ strains (e.g., 1074 and 1196 in Fig. 1) (11, 12). It hasbeen noted previously that in most Vibrio infections anerythrolytic activity is pathogenically irrelevant (9). Theactivity of the El Tor hemolysin on lymphocyte-derived cellsmay be of greater interest in vivo. We are currently investi-gating the capacity of bacterial cytotoxins in general tointerfere with the protein expression, proliferation, andcommunication of lymphocytes.
ACKNOWLEDGMENTS
We thank I. Wise, J. H. Collins, and J. Theibert for their excellenttechnical assistance.
This work was financed by the Wellcome Trust and the MedicalBiotechnology Center of the Maryland Biotechnology Institute.
LITERATURE CITED1. Blake, M. S., K. H. Johnston, G. J. Russell-Jones, and E. C.
Gotschlich. 1984. A rapid and sensitive method for detection ofalkaline phosphatase-conjugated anti-antibody on westernblots. Anal. Biochem. 136:175-179.
2. Fazekas de St. Groth, S., and D. Scheidegger. 1980. Productionof monoclonal antibodies: strategy and tactics. J. Immunol.Methods 35:1-21.
3. Giugliano, L. G., G. F. Mann, and B. S. Drasar. 1982. Responseof mammalian cell lines to the toxins of Escherichia coli. J.Med. Microbiol. 15:531-539.
4. Goldberg, S. L., and J. R. Murphy. 1984. Molecular cloning ofthe hemolysin determinant from Vibrio cholerae El Tor. J.Bacteriol. 160:239-244.
5. Goldberg, S. L., and J. R. Murphy. 1985. Cloning and charac-terization of the hemolysin determinants from Vibrio choleraeRV79(Hly+), RV79(Hly-), and S69B. J. Bacteriol. 162:35-41.
6. Hall, R. H., A. C. Hall, J. C. Ellory, C. W. Sutton, and D. A.Broadbent. 1985. The molecular mass of Vibrio cholerae hae-molysin measured by radiation inactivation. Biochem. Soc.Trans. 13:1251-1252.
7. Hames, B. D. 1981. An introduction to polyacrylamide gelelectrophoresis, p. 1-91. In B. D. Hames and D. Rickwood(ed.), Gel electrophoresis of proteins. IRL Press, Oxford.
8. Herrington, D. A., R. H. Hall, G. Losonsky, J. J. Mekalanos,R. K. Taylor, and M. M. Levine. 1988. Toxin, toxin co-regulatedpili and the toxR regulon are essential for Vibrio choleraepathogenesis in humans. J. Exp. Med. 168:1487-1492.
9. Honda, T., and R. A. Finkelstein. 1979. Purification and charac-terization of a hemolysin produced by Vibrio cholerae biotypeEl Tor: another toxic substance produced by cholera vibrios.Infect. Immun. 26:1020-1027.
10. Leary, J. J., D. J. Brigati, and D. C. Ward. 1983. Rapid andsensitive colorimetric method for visualizing biotin-labeledDNA probes hybridized to DNA or RNA immobilized on
nitrocellulose: Bio-blots. Proc. Natl. Acad. Sci. USA 80:4045-4049.
11. Levine, M. M., R. E. Black, M. L. Clements, L. Cisneros, A.Saah, D. R. Nalin, D. M. Gill, J. P. Craig, C. R. Young, and P.Ristaino. 1982. The pathogenicity of non-enterotoxigenic Vibriocholerae serogroup 01 biotype El Tor isolated from sewagewater in Brazil. J. Infect. Dis. 145:296-299.
INFECT. IMMUN.

VIBRIO CHOLERAE HlyA CYTOLYSIN 3379
12. Levine, M. M., J. B. Kaper, D. Herrington, G. Losonsky, J. G.Morris, M. L. Clements, R. E. Black, B. Tall, and R. Hall. 1988.Volunteer studies of deletion mutants of Vibrio cholerae 01prepared by recombinant techniques. Infect. Immun. 56:161-167.
13. Manning, P. A., M. H. Brown, and M. W. Heuzenroeder. 1984.Cloning of the structural gene (hly) for the haemolysin of Vibriocholerae El Tor strain 017. Gene 31:225-231.
14. Mercurio, A., and P. A. Manning. 1985. Cellular localization andexport of the soluble haemolysin of Vibrio cholerae El Tor. Mol.Gen. Genet. 200:472-475.
15. Moos, M., N. Y. Nguyen, and T.-Y. Liu. 1988. Reproduciblehigh yield sequencing of proteins electrophoretically separatedand transferred to an inert support. J. Biol. Chem. 263:6005-6008.
16. Morris, J. G., Jr., and R. E. Black. 1985. Cholera and othervibrioses in the United States. N. Engl. J. Med. 312:343-350.
17. Rader, A. E., and J. R. Murphy. 1988. Nucleotide sequencesand comparison of the hemolysin determinants of Vibrio chol-erae El Tor RV79(Hly+) and RV79(Hly-) and classical 569B(Hly-). Infect. Immun. 56:1414-1419.
18. Sutton, C. W., R. H. Hall, and D. A. Broadbent. 1985. Purifica-tion and characterization of haemolysin from Vibrio choleraebiotype El Tor. J. Med. Microbiol. 19(Suppl.):iii.
19. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
20. Yamamoto, K., Y. Ichinose, N. Nakasone, M. Tanabe, M.Nagahama, J. Sakurai, and M. Iwanaga. 1986. Identity ofhemolysins produced by Vibrio cholerae non-O1 and V. chol-erae 01, biotype El Tor. Infect. Immun. 51:927-931.
21. Yamamoto, K., M. Iwanaga, T. Honda, T. Miwatani, H. Shina-gawa, and A. Nakata. 1988. Genetic and biochemical studies onthe hemolysin of Vibrio cholerae 01, biotype El Tor, p. 175-184. In N. Ohtomo and R. B. Sack (ed.), Advances in researchon cholera and related diarrheas, vol. 6. KTK Scientific Pub-lishers, Tokyo.
22. Young, D. B., and D. A. Broadbent. 1982. Biochemical charac-terization of extracellular proteases from Vibrio cholerae. In-fect. Immun. 37:875-883.
VOL. 58, 1990





























