Embed Size (px)
Citation preview

Vesicle Transport: The Role of Actin Filamentsand Myosin MotorsANA S. DEPINA and GEORGE M. LANGFORD*Department of Biological Sciences, Dartmouth College, Hanover, New Hampshire 03755-3576
KEY WORDS actin filaments; myosin; vesicle transport; organelle/vesicle movement
ABSTRACT The transport of vesicles and the retention of organelles at specific locations arefundamental processes in cells. Actin filaments and myosin motors have been shown to be requiredfor both of these tasks. Most of the organelles in cells associate with actin filaments and some of themyosin motors required for movement on actin filaments have been identified. Myosin V has beenshown to transport endoplasmic reticulum (ER) vesicles in neurons, pigment granules in melano-cytes, and the vacuole in yeast. Myosin I has been shown to be involved in the transport ofGolgi-derived vesicles in epithelial cells. Myosin VI has been shown to be associated with Golgi-derivedvesicles, and cytoplasmic vesicles in living Drosophila embryos. Myosin II may be a vesicle motor butits role in vesicle transport has not been resolved. Secretory vesicles, endosomes and mitochondriaappear to be transported on actin filaments but the myosin motors on these organelles have not beenidentified. Mitochondria in yeast may be transported by the dynamic assembly of an actin ‘‘tail.’’ Themodel that has unified all of these findings is the concept that long-range movement of vesiclesoccurs on microtubules and short-range movement on actin filaments. The details of how themicrotubule-dependent and the actin-dependent motors are coordinated are important questions inthe field. There is now strong evidence that two molecular motors, kinesin and myosin V, interactwith each other and perhaps function as a complex on vesicles. An understanding of theinterrelationship of microtubules and actin filaments and the motors that move cargo on them willultimately establish how vesicles and organelles are transported to their specific locations in cells.Microsc. Res. Tech. 47:93–106, 1999. r 1999 Wiley-Liss, Inc.
INTRODUCTIONOne of the principal functions of molecular motors is
to transport vesicles and to retain organelles at specificlocations in the cell. It is now well established thatvesicle budding and fusion represent the principalmechanisms of membrane traffic in cells. These pro-cesses occur constitutively and, most often, the donorand acceptor organelles are located in different regionsof the cell. The transport of vesicles and organelles onactin filaments by myosin motors and especially thetransfer of vesicles from microtubules to actin filamentswill be reviewed in this article. The specific role of theactin cytoskeleton in the transport of smooth endoplas-mic reticulum (ER), Golgi-stacks, endosomes, lyso-somes, melanosomes, and mitochondria will be ad-dressed.
The orientation of the plus and minus ends of micro-tubules and actin filaments establishes the direction ofvesicle movement. In most cells microtubules are nucle-ated at the centrosome, with their minus ends towardthe center of the cell and plus ends at the cell periphery.Therefore, vesicles with plus-end directed motors willmove outward on microtubules and those with minus-end directed motors inward. The nucleation and attach-ment of microtubules at the centrosome ensures thatthe polarity of microtubules is preserved. There arespecialized cells, however, in which microtubules be-come detached from the centrosome. Noncentrosomalmicrotubules have been observed in a variety of mam-malian cells grown in culture (Vorobjev et al., 1997), in
polarized epithelial cells (Mogensen et al., 1997), and inaxons and dendrites of neurons (Bray and Bunge, 1981;Chalfie and Thomson, 1979). In dendrites, the polarityof microtubules is mixed with about 50% of plus-endsoriented toward the cell body (Baas et al., 1988). Actinfilaments, on the other hand, are nucleated at multiplesites in the cell, and are not associated with a nucle-ation center. The best-characterized site of nucleationfor actin filaments is the plasma membrane (Shariffand Luna, 1990; Svitkina et al., 1986; Wang, 1985). Anetwork of cortical actin filaments, some of whichremain membrane-attached, assembles just below theplasma membrane. Actin filaments that nucleate at themembrane attach by their plus or barbed ends (Tilneyet al., 1981), and therefore vesicles carried on theseactin filaments by plus-end-directed myosin motorsmove toward the membrane. Actin filaments that formthe cortical network are most often of mixed polarity(Lewis and Bridgman, 1992), but may be of uniformpolarity (Begg et al., 1978; Isenberg and Small, 1978;Small and Celis, 1978). Actin filaments in stress fibersmay support vesicle transport but they function primar-ily as contraction fibers. The polymerization (and bun-dling) of actin filaments in stress fibers, lamellipodia, andfilopodia is controlled by members of the Rho-GTPase
*Correspondence to: George M. Langford, Department of Biological Sciences,Dartmouth College, 6044 Gilman Laboratory, Hanover, NH 03755-3576.E-mail: [email protected].
Received 20 March 1999; accepted in revised form 19 June 1999.
MICROSCOPY RESEARCH AND TECHNIQUE 47:93–106 (1999)
r 1999 WILEY-LISS, INC.

family of proteins (Hall, 1998; He et al., 1998; Ma-chesky and Hall, 1997; Symons et al., 1996).
In addition to the cortex, actin filaments are foundwithin the cytoplasm in association with specific organ-elles and often in association with microtubules. Themode of nucleation of actin filaments in these regions ofthe cell is not well understood. Evidence to suggest thatsome organelles have actin filament nucleating-activityis beginning to accumulate (Valderrama et al., 1998;Boldogh et al., 1999). The involvement of actin fila-ments and myosin motors in the transport of each of theintracellular organelles will be reviewed below.
ACTIN FILAMENTS ARE REQUIREDFOR TRANSPORT OF ER
The role of actin filaments in the transport of ER hasbeen firmly established by a variety of studies in bothplant and animal cells. For example, light-inducedreorganization of ER in plant cells has been shown todepend on the presence of actin filaments. In quiescentmesophyll cells in the leaves of the aquatic plantVallisneria, ER is located along the wall facing the topof the leaf (the periclinal wall) but upon illumination,ER moves down along the side walls (anticlinal wall)(Liebe and Menzel, 1995). ER transport in these cellswas inhibited when treated with cytochalasin, indicat-ing that actin filaments were required for ER transport.
Studies of ER transport in photoreceptor cells ofinsect eyes provide similar evidence for actin-depen-dent ER reorganization. In dark-adapted locust photo-receptor cells, ER is located at the periphery of the cellnear the base of the photoreceptive microvilli, but inlight-adapted cells, ER moves toward the interior of thecell (Sturmer et al., 1995). The submicrovillar region ofthe cell contains actin filaments but no microtubulesand treatment with cytochalasin blocked light-inducedtransport of ER. Actin filaments, therefore, appeared tobe required for transport and for retention of ER in thesubmicrovillar region. By electron microscopy, the sub-microvilli actin filaments were shown to be extensionsof the actin cables in the microvilli, and thereforeoriented with their plus-end at the microvillar tips(Arikawa et al., 1990; Baumann, 1992; Hafner et al.,1991). Plus-end-directed myosin motors would facili-tate the transport of ER during dark adaptation, buttransport toward the cell body during light adaptationwould require that some of the actin filaments have anorientation of opposite polarity since minus-end di-rected myosin motors have not been found.
Studies have shown that light does not stimulaterelocation of ER in the photoreceptor cells of honeybeeeyes; however, actin filaments are required for theretention of ER in the submicrovillar region (Baumannand Lautenschlager, 1994). Treatment of these cellswith cytochalasin caused disorganization of ER locatedat the base of the microvilli, an actin-rich regionwithout microtubules. Therefore, retention of ER in thesubmicrovillar region rather than its transport towardthe cell interior required the presence of actin fila-ments. The mechanism by which ER attaches to actinfilaments in these cells has not been established, butspectrin, an actin cross-linking protein, is thought to beinvolved (Baumann, 1998). Cytochalasin treatment ofthese cells caused the actin filaments to depolymerize,leaving spectrin on the ER membranes. One interpreta-
tion of these data is that actin filaments are linked toER by spectrin, thereby retaining ER in the submicrovil-lar region. An equally likely explanation is that linkageof ER to actin filaments is via a myosin motor. In thelatter case, the role of spectrin may be to facilitate theattachment of molecular motors to the vesicle surface.For example, the dynein/dynactin complex, a microtu-bule-motor, is predicted to interact with spectrin on themembrane (Holleran et al., 1996). Additional evidencefor spectrin’s role in motor attachment comes fromstudies of myosin V. This actin-based motor has aputative membrane-docking motif called DIL (for di-lute) (Ponting, 1995) that is homologous to a motif inhuman AF-6 (Prasad et al., 1993) and Drosophila canoe(Miyamoto et al., 1995). The membrane protein towhich AF-6 binds has been shown to be a member of theprotein 4.1 family of cytoskeleton-associated proteinsand the complex is part of a signal transduction path-way (Ponting and Phillips, 1995; Kennedy, 1995; Marfa-tia et al., 1995). This family of proteins include protein4.1, talin, ezrin, merlin, and one unconventional myo-sin, myosin VIIA (Chen et al., 1996; Weil et al., 1996). Amember of the protein 4.1 family may be the membraneprotein to which the DIL motif of myosin V binds,although this region has been shown to bind to kinesin(Huang et al., 1999). Protein 4.1 also functions to linkthe spectrin cytoskeleton to membranes (Becker et al.,1990; Cohen and Langley, 1984). Therefore, spectrinmay be a principal component in the attachment ofmotors to vesicles through its interaction with themotor-membrane protein complex and the vesicle mem-brane cytoskeleton. (Although there is no direct evi-dence demonstrating myosin V-protein 4.1 association,binding of myosin II to protein 4.1 has been shown[Pasternack and Racusen, 1989].)
MYOSIN V HAS BEEN SHOWN TOTRANSPORT ER VESICLES IN NERVE CELLS
Direct evidence for the role of actin filaments in thetransport of ER comes from studies of axonal transportin nerve cells. Langford and colleagues used the giantaxon of the squid to show that ER vesicles are trans-ported on both microtubules and actin filaments invitro (Kuznetsov et al., 1994) and that myosin V is theactin-dependent motor (Tabb et al., 1998). In thesestudies, axoplasm of the giant axon was extruded into abuffer that promoted the assembly of actin filamentsfrom subunits that diffused out of the axoplasm. Byvideo microscopy, vesicles were observed to move onmicrotubules and then on the newly assembled actinfilaments adsorbed to the glass coverslip. This in vitromotility assay permitted the first direct observation ofvesicle transport on individual actin filaments (Kuz-netsov et al., 1992). By electron microscopy, vesiclesthat moved on actin filaments were found to consist ofan interconnected network of vesicles tethered to eachother by thin membrane tubules (Tabb et al., 1998; Fig.1). An antibody to an ER-resident protein, proteindisulfide isomerase (PDI), was used to identify thesevesicles as ER and immunogold electron microscopywas used to show that myosin V and kinesin motorswere present on ER vesicles (Tabb et al., 1998; Fig. 2).The involvement of myosin V in the transport of vesicleswas established by using myosin V-specific antibodies thatinhibited movement. Two antibodies were used to in-
94 A.S. DEPINA AND G.M. LANGFORD

Fig. 1. ER vesicles in the squid giant axon. Electron micrographs ofnegatively stained ER vesicles from squid axoplasm at high magnifica-tion (A–C) and low magnification (D–E). The vesicles were isolated byincubating extruded axoplasm on ice for 2 hours in excess buffer toremove soluble proteins, actin filaments, microtubules, and diffusibleorganelles (Morris and Lasek, 1984). The resulting axoplasmic rem-nant of vesicles and neurofilaments (NF) was applied to Formvar/
carbon-coated copper grids and negatively stained using 1% aqueousuranyl acetate. The ER, identified by an antibody to the ER-residentprotein PDI, consisted of vesicles (V) ranging between 100–500 nm indiameter connected by tubular membranes (T) 20–25 nm in diameter.Vesicles were usually present at the ends of the tubular elements. Thetubular membrane branched to form Y-shaped intersections (Y), andcoiled into spirals (asterisks in D). Bars 5 100 nm.
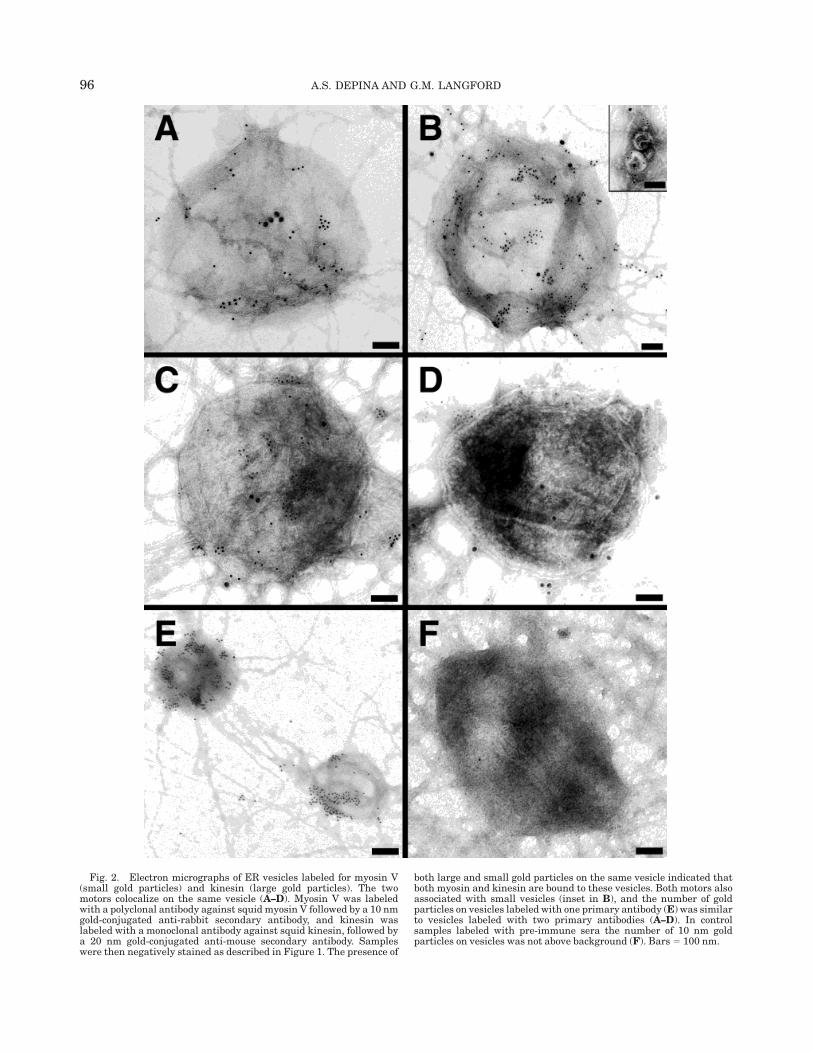
Fig. 2. Electron micrographs of ER vesicles labeled for myosin V(small gold particles) and kinesin (large gold particles). The twomotors colocalize on the same vesicle (A–D). Myosin V was labeledwith a polyclonal antibody against squid myosin V followed by a 10 nmgold-conjugated anti-rabbit secondary antibody, and kinesin waslabeled with a monoclonal antibody against squid kinesin, followed bya 20 nm gold-conjugated anti-mouse secondary antibody. Sampleswere then negatively stained as described in Figure 1. The presence of
both large and small gold particles on the same vesicle indicated thatboth myosin and kinesin are bound to these vesicles. Both motors alsoassociated with small vesicles (inset in B), and the number of goldparticles on vesicles labeled with one primary antibody (E) was similarto vesicles labeled with two primary antibodies (A–D). In controlsamples labeled with pre-immune sera the number of 10 nm goldparticles on vesicles was not above background (F). Bars 5 100 nm.
96 A.S. DEPINA AND G.M. LANGFORD

hibit movement: one directed against the head domainof myosin V and the other against the tail domain. Bothmyosin V specific antibodies inhibited movement ofvesicles on actin filaments (Tabb et al., 1998). Theantibody-inhibition experiments provided the first di-rect evidence for the involvement of myosin V in thetransport of vesicles on actin filaments.
The association of myosin V with synaptic vesicles inneurons also supports its role in vesicle transport(Evans et al., 1997; Prekeris and Terrian, 1997). In ratcerebrocortical synaptosomes, myosin V was found tocopurify specifically with synaptic vesicles, and coimmu-noprecipitate with synaptobrevin II and synaptophy-sin, two different synaptic vesicle proteins (Prekerisand Terrian, 1997). Treatment with BDM, an agentthat blocks myosin activity, inhibited exocytosis ofglutamate-containing synaptic vesicles in the presynap-tic terminal, suggesting that myosin ATPase activitywas required for synaptic vesicle exocytosis. Althoughdirect observation of the movement of synaptic vesicleson actin filaments was not reported, this was the firstdemonstration of myosin V binding to synaptic vesicles.
The in vitro observations of ER transport on actinfilaments by myosin V in the squid giant axon aresupported by studies of ER localization in the neuronsof mice and rats that are functional nulls for myosin Va(dilute). Dilute rats and mice appear normal at birthexcept for a light (dilute) coat color, but neurologicaldefects appear soon thereafter (Searle, 1952) (see be-low). By day 12, mutant animals develop symptoms ofataxia that become more severe by 3 weeks, and theyexhibit convulsive movement or opisthotonus that leadto death. When the Purkinje cells in the cerebellum ofthese animals were examined by electron microscopy,small tubular profiles of ER, normally found in den-dritic spines, were missing (Dekker-Ohno et al., 1996;Takagishi et al., 1996). ER was observed in the den-dritic trunks and tapered branches of ataxic animals,but not in the spines. The dendritic spines have beenshown to contain actin filaments but not microtubules.Therefore, the absence of ER in the dendritic spinessuggested that movement into these specialized regionsof the neuron required the activity of myosin V, aconclusion that supports the observations that ER istransported on actin filaments by myosin V. Movementof ER into the trunks of dendrites can be explained bythe presence of microtubule-motors on ER vesicles, asdemonstrated in the squid axon (Tabb et al., 1998, Fig.2). The lack of myosin V activity would account for theabsence of ER in the spines of dendrites. The absence ofER, the principal calcium store in spines, would impaircalcium signaling in the postsynaptic cell, the conse-quence of which would be the neurological defectscharacteristic of dilute mutants. A mutation in myosinV producing similar neurological defects known as Gris-celli disease has been identified in humans, althoughthis disease has other complications, including immuno-deficiency and myelodysplastic syndrome (Pastural etal., 1997).
ACTIN FILAMENTS ARE REQUIRED TOMAINTAIN THE ORGANIZATION ANDLOCATION OF THE GOLGI COMPLEX
In addition to ER localization and transport, actinfilaments are involved in the organization of the Golgi
stacks. Satiat-Jeunemaitre et al. (1996) and Boevink etal. (1998) studied the organization of the Golgi stacks inepidermal cells of plant leaves. When treated withcytochalasin, the Golgi elements that were dispersed inthe cell collapsed or aggregated. These studies showedthat actin filaments were required for Golgi localizationand movement. Similarly, in photoreceptor cells of thehoneybee eyes, Golgi stacks aligned with actin fila-ments in the submicrovillar region of the cell wheremicrotubules were absent (Baumann, 1998). In thecellular slime mold Dictyostelium discoideum, treat-ment with agents that disrupted cortical actin fila-ments led to the fragmentation of the Golgi and itsdispersal throughout the cytoplasm (Weiner et al.,1993). Comitin, a 24-kDa actin binding protein associ-ated with the Golgi stacks, was thought to connect actinfilaments to the Golgi complex, thereby forming astabilizing scaffold that maintained the organization ofthe Golgi stacks. In normal rat kidney (NRK) cellstreated with cytochalasin, the dispersed perinuclearGolgi complexes collapsed into clusters that were re-stricted to the centrosome region (Valderrama et al.,1998). The Golgi clusters induced by disruption of actinfilaments required an intact microtubule network, i.e.,if nocodazole was used to depolymerize microtubulesthe clusters became fragmented and dispersed through-out the cytoplasm. These observations suggest thatactin filaments are involved in maintaining the shapeand subcellular localization of the Golgi stacks, al-though more direct evidence is needed.
MYOSINS I AND VI HAVE BEEN IMPLICATEDIN THE TRANSPORT OF GOLGI-DERIVEDVESICLES TO THE PLASMA MEMBRANE
Both the localization of Golgi stacks and the trans-port of vesicles derived from the trans-Golgi networkappear to require actin filaments and myosin motors.Using myosin-specific antibodies, Fath et al. (1994)showed that Golgi stacks and Golgi-derived vesiclesisolated from chicken intestinal crypt cells possessedmyosin I. They proposed that Golgi-derived vesicleswere delivered to the submicrovillar region of epithelialcells on microtubules. Within the submicrovillar regionof the cell, however, vesicles were transported on actinfilaments by myosin I. In a similar study, Montes deOca et al. (1997) showed that Golgi-derived vesiclesisolated from MDCK cells possessed myosin I. Actinfilaments co-sedimented with the isolated vesicles invitro in the absence of ATP. They, too, proposed thatmyosin I transported Golgi-derived vesicles to and fromthe apical membrane in these cells.
In addition to myosin I, myosin VI has been found tobe associated with the Golgi complex (Buss et al., 1998).It was identified at the leading ruffling edge, suggestingthat it may be involved in vesicle transport from theGolgi to the leading edge. In both NRK and A431 cells(human carcinoma cell line) myosin VI localized to theleading edge, the juxtanuclear area, and throughoutthe cytoplasm. Myosin VI in the juxtanuclear regioncolocalized with Golgi markers for the cis-, medial-, andtrans-compartments, confirming that the staining inthis region was due to the Golgi stacks. In A431 cells,EGF stimulation caused ruffling and recruitment ofmyosin VI to the ruffles. This recruitment was accompa-nied by an increase in the phosphorylation of myosin VI
97ACTIN FILAMENTS AND MYOSIN MOTORS

head domain. Upon phosphorylation, myosin VI at-tached to actin filaments in the presence of ATP asefficiently as it did in the absence of ATP. In anotherstudy, myosin VI was shown to transport cytoplasmicvesicles in living Drosophila embryos (Mermall et al.,1994). These studies suggested that myosin VI is likelyto be involved in vesicle transport from the Golgi stacksto the leading edge and its ATPase activity may beregulated by phosphorylation.
Isolated Golgi elements from liver cells, however,have been reported to contain very little myosin VI(Hasson and Mooseker, 1994), while Golgi isolated fromkidney cells contained a significant amount of myosinVI (Buss et al., 1998). The fact that comitin (Wiener etal., 1993), spectrin (Beck et al., 1994; Fath et al., 1997),and the unconventional motors myosin I and VI wereshown to be associated with the Golgi (Fath andBurgess, 1993; Fath et al., 1994; Buss et al., 1998),suggest that actin-dependent processes are involved inthe secretory pathway at the Golgi complex.
MYOSIN II AND THE TRANSPORT OF GOLGISTACKS AND GOLGI-DERIVED VESICLES
Myosin II has been implicated as a motor for vesicletransport but the issue remains unsettled. Severalstudies from cultured mammalian cells suggested thatmyosin II is involved in the localization and transport ofGolgi stacks and Golgi-derived vesicles (Stow et al.,1998). Recent studies have shown that a monoclonalantibody (AD7) that recognizes nonmuscle myosin II-precipitated Golgi-derived vesicles in extracts of MDCKcells (Musch et al., 1997). Studies from two differentlaboratories designed to clarify the role of myosin II inthe formation of Golgi-derived vesicles and transport ofvesicles from the trans-Golgi network (TGN) to theplasma membrane produced contradictory results. Onestudy concluded that myosin II is required for vesicleformation (Musch et al., 1997), while the other found noevidence for a role of myosin II in transport from theTGN to the plasma membrane or its presence inTGN-derived vesicles (Ikonen et al., 1996). A morerecent study has shown that the AD7 antibody used inthese studies was not specific for myosin but alsocross-reacted with coatamer proteins (Simon et al.,1998), calling into question the role of myosin in Golgivesicle budding and transport (Stow and Heimann,1998). The specific role of myosin II in vesicle transportremains an unanswered question but its associationwith Golgi membranes has been established (Ikonen etal., 1997).
Recent studies of DePina and Langford have shownthat vesicles in extracts of clam oocytes (Spisula solidis-sima) are transported on actin filaments and myosin II
is present on these vesicles (DePina and Langford,manuscript in preparation). These vesicles moved onactin filaments at speeds that were about twice as fastas the speed of vesicle transport on actin filaments inthe giant axon of the squid (Fig. 3). This vesicle-
Fig. 3. Vesicle transport on an actin filament in an extract of clamoocytes. A: A series of images showing the movement of a vesicle alongan actin filament (invisible) adsorbed to the coverslip surface (AVEC-DIC microscopy). The direction of movement is toward the right(intervals, 0.1 seconds). Extracts were prepared as described byWalker et al. (1996), clarified by a high speed spin, and activated bychanging the pH from 6.8 to 7.2 and incubating at 18°C for 45 minutes.Samples were then stained with rhodamine-phalloidin and observedby AVEC-DIC and conventional fluorescence microscopy. B: Distancevs. time plot of the vesicle moving on an actin filament shows that thevesicle travels 2.2 µm in 2 seconds. C: The instantaneous velocity ofthe vesicle reached a maximum of 2.9 µm/s. Bar 5 2.5 µm.
98 A.S. DEPINA AND G.M. LANGFORD

associated myosin was purified and identified as myo-sin II based on the microsequence data of 11 tryptic-peptides. The peptide sequence data permitted thepositive identification of the motor protein as an iso-form of nonmuscle myosin II.
Additional evidence that myosin II is the vesiclemotor in these extracts was obtained by inhibitingvesicle transport with ML-7, a selective inhibitor ofmyosin light chain kinase (MLCK) (Ruchhoeft andHarris, 1997). Vesicle transport on actin filaments wasinitiated in extracts of clam oocytes by shifting the pHfrom 6.8 to 7.2 and incubating at 18°C. High levels ofmotile activity, 65 6 14 vesicles/field/m (n 5 4), wereobserved in the control samples but in samples treatedwith ML-7, motility on actin filaments was inhibited. At10 µM ML-7 the motile activity decreased by 31% whileat 50–150 µM ML-7, motile activity decreased by85–95%. Motility was completely inhibited at 200 µM.ML-7 did not affect the formation of actin networksduring the observation period (60 minutes) as revealedby immunofluorescence microscopy (Fig. 4). Immunob-lots of the vesicle fractions showed that myosin IIremained associated with the vesicles in the presence ofML-7. These data showed that the inhibition of MLCKby ML-7 blocked vesicle movement presumably byinhibiting the activity of myosin II. Therefore thesedata support the conclusion that myosin II functions asa vesicle motor in extracts of clam oocytes. The identityof these vesicles has not been determined but the role ofmyosin II as a vesicle motor is supported by thesestudies.
ACTIN FILAMENTS AND MYOSINARE REQUIRED FOR EXOCYTOSIS
IN SECRETORY CELLSActin filaments and myosin motors have been shown
to be involved in the release of exocytic vesicles insecretory cells. The initial evidence for the involvementof actin filaments in secretion came from genetic stud-ies in yeast where mutations in the actin gene resultedin the accumulation of secretory vesicles in the mothercell (Novick and Botstein, 1985). Evidence for myosinmotors came from studies of yeast mutants lackingMyo2p, a class V myosin. In these mutant cells, vesiclesaccumulated in the cytoplasm and the cells failed tobud (Johnston et al., 1991; Govindan et al., 1995),suggesting a role for myosin V in vesicle transport.Myosin motors have been shown to be required for therecruitment of vesicles to exocytic sites in early seaurchin embryonic cells (Bi et al., 1997).
Additional evidence for the involvement of actinfilaments in secretory cells came from studies of insulinsecretion in pancreatic acinar cells where partial break-down of cortical actin increased secretion while com-plete disassembly inhibited secretion (Muallem et al.,1995). These authors proposed that disassembly ofactin was the final trigger for exocytosis in permeabi-lized pancreatic acinar cell, since introduction of lowconcentrations of actin monomer-binding proteins in-duced amylase release. Valentijn et al. (1998) per-formed similar studies and their results indicated thatthe subapical actin filaments of pancreatic acinar cells
Fig. 4. Actin filaments assemble spontaneously in extracts of clam oocytes. Extracts were prepared asdescribed in Figure 3. Actin filaments were visualized by fluorescence microscopy after staining withrhodamine-phalloidin (0.4 µM). Each fine line represents a single actin filament. Treatment of extractswith ML-7 did not affect actin filament assembly. Bar 5 10 µm.
99ACTIN FILAMENTS AND MYOSIN MOTORS

regulated the balance between exocytosis and compen-satory membrane retrieval.
ACTIN FILAMENTS ASSOCIATE WITHENDOSOMES AND LYSOSOMES
The mechanism by which endosomal vesicles aredelivered to lysosomes remains to be established, butthere is some evidence that both the actin and microtu-bule cytoskeletons are involved. It has been shown thatmicrotubules are involved in early to late stage endo-cytic transport (Aniento et al., 1993; Gruenberg et al.,1989), and that microtubule motors regulate the posi-tion and movement of endosomes (Aniento et al., 1993;Bomsel et al., 1990; Heuser, 1989; Matteoni and Kreis,1987; Hollenbeck and Swanson, 1990; Parton et al.,1991). However, myosin motors have also been reportedto be associated with endocytic organelles (Baines et al.,1992; Yonemura and Pollard, 1992; D’Andrea et al.,1994), and appear to be required for vacuole inheri-tance in yeast (Hill et al., 1996). Therefore, not onlymicrotubules, but also actin filaments are thought tomediate endosomal vesicle transport.
The association of actin and myosin motors withendosomes and lysosomes and their role in endocytosiswere investigated recently in cultured cells and inisolated rat liver fractions. In Caco-2 cells, a humancolon adenocarcinoma cell line, cytochalasin inhibitedapical endocytosis of ricin or folate in a concentration-dependent manner (Jackman et al., 1994). Binding ofricin to the cell surface and basolateral endocytosis wasnot affected. Cytochalasin treatment did not alter brushborder morphology nor disrupt microvillar actin fila-ments, but completely depolymerized terminal webactin filaments. The disruption of actin filaments in theterminal web at the base of microvilli by cytochalasinwas thought to be responsible for the inhibition ofendocytosis. In human carcinoma cells HEp-2 (vanDeurs et al., 1995), endocytosis was monitored byincubating cells with cationized gold particles for vari-ous time points and processing for EM. Ultrathincryosections were probed secondarily with immunogold-labeled lysosome-specific markers using smaller goldparticles. The colocalization of the two gold probes wasdetermined. In the presence of nocodazole, colocaliza-tion was not affected, but cytochalasin decreased colocal-ization, suggesting that actin filaments were involvedin the delivery of endosomes to lysosomes. Cytochalasinalso slowed the fusion of mature endosomes and lyso-somes. In addition, treatment with cytochalasin inhib-ited short excursions of endosomes at the periphery ofthe cell. Nocodazole treatment had the opposite effect,inhibiting the long excursions and causing perinuclearaggregation of endosomes. These data suggested thatboth actin filaments and microtubules were necessaryto maintain the normal distribution and movement ofendocytic compartments in HEp-2 cells.
Studies in BWTG3 cells derived from a mouse hepa-toma cell line (Durrbach et al., 1996a) showed thatendocytosed fluorescently tagged transferrin and a-2-macroglobulin localized to one pole of the nucleus.Treatment with either colchicine or cytochalasin priorto endocytosis prevented the perinuclear localizationand caused dispersal of the fluorescently labeled com-partments throughout the cytoplasm. Treatment withcolchicine after endocytosis and localization to the
juxtanuclear position caused dispersal, but treatmentwith cytochalasin at this time point did not lead todispersal from the perinuclear region. In untreatedcells, the endosomes and some recycling endosomeslocalized to the juxtanuclear region (Salzman andMaxfield, 1989; Hopkins et al., 1994), whereas lyso-somes dispersed throughout the cytoplasm. Therefore,these studies with colchicine and cytochalasin sug-gested that microtubules mediated transport of ligandsamong endosomal compartments, and actin filamentsmediated transport between endosomes and lysosomes.Treatment with cytochalasin delayed transferrin up-take, suggesting that actin filaments were also involvedin the uptake of ligands.
In rat embryonic fibroblasts (Nakagawa and Miya-moto, 1998), actin filaments specifically localized tointact sorting endosomes. Immunofluorescence studiesusing an actin antibody revealed punctate structuresthroughout the cytoplasm of these cells. Antibodiesagainst a specific sorting endosome protein colocalizedwith the punctate structures, suggesting that actinlocalized to sorting endosomes. Furthermore, stainingwith antitransferrin receptor antibody, a marker for thesorting and recycling endosomes, colocalized with actinon vesicular structures at the cell periphery. The actinantibody did not stain endosomal vesicles in the peri-nuclear region where recycling endosomes were pres-ent, confirming that actin localized to sorting endo-somes. Tropomyosin, an actin-binding protein, colocalizedwith actin filaments on sorting endosomes; therefore,actin filaments and tropomyosin were components ofthe membrane cytoskeleton of sorting endosomes. Thefact that sorting endosomes are located beneath the cellmembrane (Gruenberg et al., 1989) in regions rich inactin filaments with few microtubules (Heuser andKirschner, 1980; Rinnerthaler et al., 1991) supports theidea that sorting endosomes utilize actin filaments forlocalization and transport.
Pol et al. (1997) identified actin in association withendosomes by isolating three highly purified rat liverendosomal fractions and determining the amount ofactin in each fraction. The MVB fraction (late endo-somes or multivesicular bodies) contained the lowestamount of actin. The CURL fraction (early endosomesor compartment of uncoupling receptors and ligands)contained 15% of the actin, while the RCC fraction(receptor-recycling compartment) contained 84% of theactin. These results provided additional evidence thatrat liver endosomes associated with actin. RhoD GT-Pase has been shown to regulate early endosome dynam-ics and actin filament rearrangements (Murphy et al.,1996).
MYOSIN I AND ENDOSOMESThe expression of chicken brush border myosin I
(BBMI) in a mouse hepatoma cell line (BWTG3), re-vealed a role for myosin I in the transport of endosomes.The most significant results were obtained when trun-cated myosin I constructs were expressed. For example,the expression of cDNA encoding a truncated myosin-Ithat lacked the ATP binding site or the entire motordomain revealed colocalization of myosin I with endo-cytic compartments (Durrbach et al., 1996b). Expres-sion of the full-length cDNA revealed codistribution ofmyosin I with cortical actin, a distribution similar to
100 A.S. DEPINA AND G.M. LANGFORD

that in fibroblasts (Wagner et al., 1992; Conrad et al.,1993), and Cos or 3T3 cells (Collins and Matsudaira,1995; Footer and Brestcher, 1994). These results demon-strated that when the amino-terminal sequence contain-ing the ATP binding site was present, myosin-I colocal-ized with actin filaments, whereas when the actinbinding region was absent the tail domain colocalizedwith endocytic vesicles. Furthermore, expression oftruncated myosin I disrupted the morphology and positionof the juxtanuclear endocytic compartments, resultingin fragmented structures scattered throughout thecytoplasm, a dominant negative effect (Durrbach et al.,1996b). Also, expression of BBMI lacking the ATPbinding domain decreased the rate of a-2-macroglobu-lin degradation, whereas expression of myosin I lackingthe entire motor domain increased its rate of degrada-tion. Presumably, myosin I lacking the ATP binding sitewould still be able to link endocytic compartments toactin filaments, but would not be able to move themalong actin in an ATP-dependent manner, whereasmyosin I lacking the entire motor domain would not beable to link endocytic compartments to actin filaments.These findings suggest that an unidentified myosin Ibinds the juxtanuclear endosomes to actin and regu-lates delivery of ligands to degradative compartments.The fact that the antibodies against chicken brushborder myosin I recognized three polypeptides inBWTG3 cell extracts indicated that other myosins Iwere present in these cells.
A more recent study provides biochemical and immu-noelectron microscopic data that myosin Ia is themyosin I associated with endosomes and lysosomes inmammalian cells (Raposo et al., 1999). The authors alsoshow that the delivery of fluid phase markers fromendosomes to lysosomes is impaired in cells producingtruncated myosin I, suggesting that myosin I mightmediate vesicle transport between endosomes and lyso-somes.
Additional evidence for the participation of myosin Iin endocytosis was obtained from studies of mutantDictyostelium. Five myosin I heavy chain genes havebeen cloned and sequenced in Dictyostelium, myoA,myoB, myoC, myoD, and myoE. MyoB, myoC, andmyoD contain all three tail homology regions identifiedpreviously in Acanthamoeba myosins I and are referredto as ‘‘classic’’ myosins I. Analysis of myoB, myoC, andmyoD single mutants and myoB/myoC/myoD triplemutants reveal that all three isoforms contributed toendocytosis, and that their contributions were synergis-tic (Jung et al., 1996). Triple mutants exhibited approxi-mately 60% inhibition in fluid phase pinocytosis rate,while the sum of the inhibition by the three singlemutants was about 25%. It has also been reported thatDictyostelium myosin I double-mutants (myoA/B andmyoB/C) exhibited a decreased rate of endocytosis andabnormal intracellular vesicle profiles (Novak et al.,1995). MyoB, myoB/C, myoA, and myoA/B mutantswere found to oversecrete the lysosomal enzymes a-man-nosidase and acid phosphatase, but transport, localiza-tion, and processing of a-mannosidase were unaffected.These results suggested that myosin I isoforms controlendocytosis at the actin-rich cortical region rather thanintracellular movement of endocytic vesicles (Temes-vari et al., 1996).
MYOSIN V, MELANOSOMES, AND LYSOSOMESTransport of pigment granules in melanocytes has
been shown to involve myosin V. In addition to theneurological defects described above, dilute mice ex-hibit pigmentation defects resulting in a ‘‘diluted’’ orlight coat color (Searle, 1952). Melanocytes, the den-dritic cells near the base of the hair bulb within theepidermis (reviewed by Boissy, 1988; Hearing and King,1993; Jimbow et al., 1993) are responsible for deliveringmelanosomes via their dendritic processes to keratino-cytes. Melanocytes in dilute animals fail to deliverpigment granules to the hair shaft. Dilute melanocyteshave been shown to have normal dendritic morphology,melanosome biogenesis, and melanin content, but themelanosome distribution is altered (Provance et al.,1996; Wu et al., 1997; Wei et al., 1997). Melanosomesare distributed throughout the cell body and dendritesin wild-type cells, but is concentrated in the cell bodyand excluded from dendrites in mutant melanocytes.
Myosin V is associated with melanosomes in normalmouse melanocyte cell lines at the light and electronmicroscope level, and is enriched in partially purifiedmelanosome fractions (Wu et al., 1997; Evans et al.,1997; Nascimento et al., 1997). Myosin V colocalizedwith actin filaments in dendrites and dendritic tips,regions where melanosomes accumulated. Wu et al.(1998) demonstrated that melanosomes movement wasmyosin-V dependent, and that the role of myosin V wasprimarily to capture melanosome in the actin-richperiphery. The capture of melanosomes at the actin richperiphery occurred in the presence of bidirectionalmovement of melanosomes on microtubules, and servedto drive the peripheral accumulation of melanosomes.The evidence from these studies favor a mechanismwhereby melanosomes are transported from the centerof the cell by microtubule motors and are then distrib-uted within dendritic extensions by myosin V.
Transport of melanosomes on actin filaments in fishand Xenopus melanophores after treatment with cyclicAMP has been shown (Rodionov et al., 1998; Rogers andGelfand, 1998). In these studies, myosin V was shownto be associated with isolated melanosomes and theproper distribution of melanosomes was shown to re-quire the functional coordination of transport on actinfilaments and microtubules.
In yeast, a class V myosin (Myo2p) has been impli-cated in vacuole (lysosome) inheritance (Hill et al.,1996). Actin cables and Myo2p were found to colocalizewith vacuole membranes, and a temperature-sensitivemutation in the actin-binding domain of Myo2p blockedvacuole inheritance. The defect was not due to adisruption of the actin cytoskeleton, but several actinmutations in regions of actin that were important formyosin binding impaired vacuole inheritance. There-fore, in addition to melanosomes and ER, myosin V ishypothesized to act as a molecular motor for vacuoletransport on actin filaments during bud formation inyeast.
MITOCHONDRIA TRANSPORTON ACTIN FILAMENTS
The transport of mitochondria on actin filamentsremains controversial (Yaffe, 1999); however, there isabundant evidence that mitochondria co-align with
101ACTIN FILAMENTS AND MYOSIN MOTORS

actin filaments. For example, in the photoreceptor cellsof the locust eye mitochondria have been shown to alignwith actin filaments (Sturmer et al., 1995). In thesecells, mitochondria moved and aggregated at the baseof the photoreceptive microvilli when illuminated. Treat-ment with cytochalasin caused displacement of mito-chondria and blocked the light-induced transport. Mor-ris and Hollenbeck (1995) used chick sympatheticneurons to study the transport of mitochondria alongactin filaments in axons. They found that mitochondriacontinued to move in the presence of drugs that depoly-merized microtubules. The microtubule-independentmovement was cytochalasin sensitive. The actin-dependent movement showed net retrograde displace-ment and occurred at a slower speed. These studiesprovided strong evidence for the role of actin filamentsin the movement of mitochondria. Although the myosinmotor has not been identified in these cells, evidencefrom studies of mitochondria transport in Drosophilanurse cells suggest that myosin VI may be involved(Bohrmann, 1997).
The role of actin filaments in the transport of mito-chondria in yeast has been studied in wild-type and inmutant cells (reviewed by Simon and Pon, 1996).Drubin et al. (1993) showed that mitochondria colocal-ize with actin cables in yeast and the alignment patternwas defective in cells carrying a mutant actin gene.Mutations in the actin gene that had the most severeeffect on mitochondria organization were located in ornear the myosin binding site on the actin monomer. Ponand colleagues (Lazzarino et al., 1994) isolated yeastmitochondria and studied actin-binding activity in vitro.They showed that mitochondria bound to actin fila-ments in an ATP-dependent manner and co-sedimentedwith actin filaments in the absence of ATP. They alsoshowed that isolated mitochondria immobilized on glasscoverslips supported unidirectional gliding of actinfilaments (Simon et al., 1995). The in vitro motilityassay demonstrated that actin-dependent motor activ-ity is associated with mitochondria. Surprisingly, mito-chondria transport was not affected in cells bearingmutations in either of the genes for myosins I, II, and V.More recently, actin polymerization similar to that usedby the bacterial pathogen Listeria monocytogenes wasreported to be required for mitochondrial inheritance inyeast (Boldogh et al., 1999). Therefore, the movement ofmitochondria in yeast may be driven by the assembly ofactin filaments mediated by the Arp2/3 complex ratherthan a myosin motor.
Several mitochondrial outer membrane proteins havebeen identified as essential for mitochondrial transportfrom the mother cell to the daughter bud (Berger et al.,1997). Cells bearing mutations in genes identified asMDM10, MMM1, and MDM12 have giant, round mito-chondria that do not move into the daughter buds(Berger et al., 1997; Sogo and Yaffe, 1994; Burgess et al.,1994; Boldogh et al., 1998). The products of these genes,Mdm10p, Mmm1p, and Mdm12p are thought to func-tion as attachment sites for mitochondria to the actincytoskeleton (Boldogh et al., 1999).
CONCLUSIONSThere is abundant evidence that organelle localiza-
tion and vesicle transport in cells require both microtu-
bules and actin filaments. The evidence is also clearthat microtubules function as tracks for long-distancemovement while actin filaments function as tracks forthe local movement and positioning of organelles/vesicles (Langford and Molyneaux, 1998). The specificroles of these two filament systems in the localizationand transport of each organelle represent one of themajor areas of research. The other major area ofresearch is the identification of the specific motors thatgenerate movement of vesicles and organelles. Theidentification of 15 classes of myosin motors (Mermallet al., 1998) require the identification of those membersthat function as vesicle motors. It is clear that many ofthese myosins are vesicle motors and they may haveoverlapping roles (Table 1). The motors for which thereis compelling evidence for vesicle transport are myosinI and myosin V. These motors have been purified,biochemically characterized, and shown to bind and totransport vesicles. The evidence is strong that myosin Itransports a subset of Golgi-derived vesicles to theapical cell surface of epithelial cells (Fig. 5B). Myosin Vtransports melanosomes into the dendritic tips of mela-nocytes and transports ER into the dendritic spines ofneuronal cells (Fig. 5A,C). Myosin V has been shown tocopurify with synaptic vesicles, and is involved invacuole inheritance in yeast. Myosin II is anotherimportant potential vesicle motor that may transportGolgi-derived vesicles. The motors required for move-ment of secretory vesicles to the docking site on theplasma membrane and the compensatory recycling ofvesicles are unknown. Other unconventional myosins,including myosin IXb, have been demonstrated to ex-hibit mechanochemical activity in vitro and are candi-dates for vesicle transport (Post et al., 1998).
In addition to the role of actin filaments as tracks formyosin motors, they serve as scaffolds for organellesand organelle localization. The linkage of actin fila-ments to organelles is poorly understood, but severalorganelle-associated actin-binding proteins have beenidentified. The site of nucleation of actin filaments atorganelles and within microtubule domains has yet tobe established. These are all exciting areas of currentand future research.
TABLE 1. Association of myosin with organelles
Organelle Myosin Reference
ER V Dekker-Ohno et al., 1996; Takagishiet al., 1996; Tabb et al., 1998
Golgi I, II, VI Fath and Burgess, 1993; Fath et al.,1994; 1997; Montes de Oca et al.,1997; Ikonen et al., 1997; Muschet al., 1997; Buss et al., 1998
Golgi-derived &Secretory vesicles
V Johnson, 1991; Espreafico et al.,1992; Govindan et al., 1995;Prekeris and Terrian, 1997
Endosome I Novak et al., 1995; Durrbach et al.,1996b; Geli and Riezman, 1996;Goodson et al., 1996; Jung et al.,1996; Raposo et al., 1999
Lysosome V Hill et al. 1996; Raposo et al., 1999Melanosome V Provance et al., 1996; Nascimento
et al., 1997; Wei et al., 1997; Wuet al., 1997; Rogers and Gelfand,1998
Mitochondria VI Bohrmann, 1997
102 A.S. DEPINA AND G.M. LANGFORD

REFERENCESAniento F, Emans N, Griffiths G, Gruenberg J. 1993. Cytoplasmic
dynein-dependent vesicular transport from early to late endosomes.J Cell Biol 123:1373–1387.
Arikawa K, Hicks JL, Williams DS. 1990. Identification of actinfilaments in the rhabdomeral microvilli of Drosophila photorecep-tors. J Cell Biol 110:1993–1998.
Baas PW, Deitch JS, Black MM, Banker GA. 1988. Polarity orientationof microtubules in hippocampal neurons: uniformity in the axon andnonuniformity in the dendrite. Proc Natl Acad Sci USA 85:8335–8339.
Baines IC, Brzeska H, Korn ED. 1992. Differential localization ofAcanthamoeba myosin I isoforms. J Cell Biol 119:1193–1203.
Baumann O. 1992. Structural interactions of actin filaments andendoplasmic reticulum in honeybee photoreceptor cells. Cell TissueRes 268:71–79.
Baumann O. 1998. The Golgi apparatus in honeybee photoreceptorcells: structural organization and spatial relationship to microtu-bules and actin filaments. Cell Tissue Res 291:351–361.
Baumann O, Lautenschlager B. 1994. The role of actin filaments in theorganization of the endoplasmic reticulum in honeybee photorecep-tor cells. Cell Tissue Res 278:419–432.
Beck KA, Buchanan JA, Malhotra V, Nelson WJ. 1994. Golgi spectrin:identification of an erythroid beta-spectrin homolog associated withthe Golgi complex. J Cell Biol 127:707–723.
Becker PS, Schwartz MA, Morrow JS, Lux SE. 1990. Radiolabel-transfer cross-linking demonstrates that protein 4.1 binds to theN-terminal region of beta spectrin and to actin in binary interac-tions. Eur J Biochem 193:827–836.
Begg DA, Rodewald R, Rebhun LI. 1978. The visualization of actinfilament polarity in thin sections. Evidence for the uniform polarityof membrane-associated filaments. J Cell Biol 79:846–852.
Berger KH, Sogo LF, Yaffe MP. 1997. Mdm12p, a component requiredfor mitochondrial inheritance that is conserved between buddingand fission yeast. J Cell Biol 136:545–553.
Bi GQ, Morris RL, Liao G, Alderton JM, Scholey JM, Steinhardt RA.1997. Kinesin- and myosin-driven steps of vesicle recruitment forCa21-regulated exocytosis. J Cell Biol 138:999–1008.
Fig. 5. Composite diagram of a cell illustrating organelle andvesicle transport on actin filaments. Microtubules are used for long-distance transport of vesicles and actin filaments are used for localtransport of vesicles. Actin filaments form the core of cellular projec-tions, such as microvilli (B), lamellipodia (D), and filopodia (E). Actinfilaments serve as tracks for transport of melanosomes into thedendritic tips of melanocytes (A) and transport of ER into the dendritic
spines of neuronal cells (C). The presence of myosin motors onGolgi-derived vesicles suggest that actin filaments also support move-ment of secretory vesicles to the apical surface of epithelial cells (B).Mitochondria, ER, and Golgi stacks align with actin filaments, andactin filaments are found in association with the Golgi apparatus andendosomal compartments.
103ACTIN FILAMENTS AND MYOSIN MOTORS

Boevink P, Oparka K, Santa Cruz S, Martin B, Betteridge A, Hawes C.1998. Stacks on tracks: the plant Golgi apparatus traffics on anactin/ER network. Plant J 15:441–447.
Bohrmann J. 1997. Drosophila unconventional myosin VI is involvedin intra- and intercellular transport during oogenesis. Cell Mol LifeSci 53:652–662.
Boissy RE. 1988. The melanocyte. Its structure, function, and subpopu-lations in skin, eyes, and hair. Dermatol Clin 6:161–173.
Boldogh I, Vojtov N, Karmon S, Pon LA. 1998. Interaction betweenmitochondria and the actin cytoskeleton in budding yeast requirestwo integral mitochondrial outer membrane proteins, Mmm1p andMdm10p. J Cell Biol 141:1371–1381.
Boldogh I, Nowakowski D, Karmon S, Vojtov N, Pon L. 1999. Actin-based mitochondrial motility in budding yeast. Biophys J 76:A1.
Bomsel M, Parton R, Kuznetsov SA, Schroer TA, Gruenberg J. 1990.Microtubule- and motor-dependent fusion in vitro between apicaland basolateral endocytic vesicles from MDCK cells. Cell 62:719–731.
Bray D, Bunge MB. 1981. Serial analysis of microtubules in culturedrat sensory axons. J Neurocytol 10:589–605.
Burgess SM, Delannoy M, Jensen RE. 1994. MMM1 encodes amitochondrial outer membrane protein essential for establishingand maintaining the structure of yeast mitochondria. J Cell Biol126:1375–1391.
Buss F, Kendrick-Jones J, Lionne C, Knight AE, Cote GP, Luzio JP.1998. The localization of myosin VI at the Golgi complex and leadingedge of fibroblasts and its phosphorylation and recruitment intomembrane ruffles of A431 cells after growth factor stimulation. JCell Biol 143:1535–1545.
Chalfie M, Thomson JN. 1979. Organization of neuronal microtubulesin the nematode Caenorhabditis elegans. J Cell Biol 82:278–289.
Chen ZY, Hasson T, Kelley PM, Schwender BJ, Schwartz MF, Ra-makrishnan M, Kimberling WJ, Mooseker MS, Corey DP. 1996.Molecular cloning and domain structure of human myosin-VIIa, thegene product defective in Usher syndrome 1B. Genomics 36:440–448.
Cohen CM, Langley RC Jr. 1984. Functional characterization ofhuman erythrocyte spectrin alpha and beta chains: association withactin and erythrocyte protein 4.1. Biochemistry 23:4488–4495.
Collins K, Matsudaira PT. 1995. Recombinant expression of the brushborder myosin I heavy chain. Cell Motil Cytoskeleton 32:151–161.
Conrad PA, Giuliano KA, Fisher G, Collins K, Matsudaira PT, TaylorDL. 1993. Relative distribution of actin, myosin I, and myosin IIduring the wound healing response of fibroblasts. J Cell Biol120:1381–1391.
D’Andrea L, Danon MA, Sgourdas GP, Bonder EM. 1994. Identifica-tion of coelomocyte unconventional myosin and its association within vivo particle/vesicle motility. J Cell Sci 107:2081–2094.
Dekker-Ohno K, Hayasaka S, Takagishi Y, Oda S, Wakasugi N,Mikoshiba K, Inouye M, Yamamura H. 1996. Endoplasmic reticu-lum is missing in dendritic spines of Purkinje cells of the ataxicmutant rat. Brain Res 714:226–230.
Drubin DG, Jones HD, Wertman KF. 1993. Actin structure andfunction: roles in mitochondrial organization and morphogenesis inbudding yeast and identification of the phalloidin-binding site. MolBiol Cell 4:1277–1294.
Durrbach A, Louvard D, Coudrier E. 1996a. Actin filaments facilitatetwo steps of endocytosis. J Cell Sci 109:457–465.
Durrbach A, Collins K, Matsudaira P, Louvard D, Coudrier E. 1996b.Brush border myosin-I truncated in the motor domain impairs thedistribution and the function of endocytic compartments in ahepatoma cell line. Proc Natl Acad Sci USA 93:7053–7058.
Espreafico EM, Cheney RE, Matteoli M, Nascimento AA, De CamilliPV, Larson RE, Mooseker MS. 1992. Primary structure and cellularlocalization of chicken brain myosin-V (p190), an unconventionalmyosin with calmodulin light chains. J Cell Biol 119:1541–1557.
Evans LL, Hammer J, Bridgman PC. 1997. Subcellular localization ofmyosin V in nerve growth cones and outgrowth from dilute-lethalneurons. J Cell Sci 110:439–449.
Fath KR, Burgess DR. 1993. Golgi-derived vesicles from developingepithelial cells bind actin filaments and possess myosin-I as acytoplasmically oriented peripheral membrane protein. J Cell Biol120:117–127.
Fath KR, Trimbur GM, Burgess DR. 1994. Molecular motors aredifferentially distributed on Golgi membranes from polarized epithe-lial cells. J Cell Biol 126:661–675.
Fath KR, Trimbur GM, Burgess DR. 1997. Molecular motors and aspectrin matrix associate with Golgi membranes in vitro. J Cell Biol139:1169–1181.
Footer M, Bretscher A. 1994. Brush border myosin-I microinjected into
cultured cells is targeted to actin-containing surface structures. JCell Sci 107:1623–1631.
Geli MI, Riezman H. 1996. Role of type I myosins in receptor-mediatedendocytosis in yeast. Science 272:533–535.
Goodson HV, Anderson BL, Warrick HM, Pon LA, Spudich JA. 1996.Synthetic lethality screen identifies a novel yeast myosin I gene(MYO5): myosin I proteins are required for polarization of the actincytoskeleton. J Cell Biol 133:1277–1291.
Govindan B, Bowser R, Novick P. 1995. The role of Myo2, a yeast classV myosin, in vesicular transport. J Cell Biol 128:1055–1068.
Gruenberg J, Griffiths G, Howell KE. 1989. Characterization of theearly endosome and putative endocytic carrier vesicles in vivo andwith an assay of vesicle fusion in vitro. J Cell Biol 108:1301–1316.
Hafner GS, Tokarski TR, Kipp J. 1991. Changes in the microvilluscytoskeleton during rhabdom formation in the retina of the crayfishProcambarus clarkii. J Neurocytol 20:585–596.
Hall A. 1998. Rho GTPases and the actin cytoskeleton. Science279:509–514.
Hasson T, Mooseker MS. 1994. Porcine myosin-VI: characterization ofa new mammalian unconventional myosin. J Cell Biol 127:425–440.
He H, Watanabe T, Zhan X, Huang C, Schuuring E, Fukami K,Takenawa T, Kumar CC, Simpson RJ, Maruta H. 1998. Role ofphosphatidylinositol 4,5-bisphosphate in Ras/Rac-induced disrup-tion of the cortactin-actomyosin II complex and malignant transfor-mation. Mol Cell Biol 18:3829–3837.
Hearing VJ, King RA. 1993. Determinants of skin color: melanocytesand melanization. In: Levine N, editor. Pigmentation and pigmen-tary disorders. London: CRC Press. p 4–18.
Heuser J. 1989. Changes in lysosome shape and distribution corre-lated with changes in cytoplasmic pH. J Cell Biol 108:855–864.
Heuser JE, Kirschner MW. 1980. Filament organization revealed inplatinum replicas of freeze-dried cytoskeletons. J Cell Biol 86:212–234.
Hill KL, Catlett NL, Weisman LS. 1996. Actin and myosin function indirected vacuole movement during cell division in Saccharomycescerevisiae. J Cell Biol 135:1535–1549.
Hollenbeck PJ, Swanson JA. 1990. Radial extension of macrophagetubular lysosomes supported by kinesin. Nature 346:864–866.
Holleran EA, Tokito MK, Karki S, Holzbaur EL. 1996. Centractin(ARP1) associates with spectrin revealing a potential mechanism tolink dynactin to intracellular organelles. J Cell Biol 135:1815–1829.
Hopkins CR, Gibson A, Shipman M, Strickland DK, Trowbridge IS.1994. In migrating fibroblasts, recycling receptors are concentratedin narrow tubules in the pericentriolar area, and then routed to theplasma membrane of the leading lamella. J Cell Biol 125:1265–1274.
Huang JD, Brady ST, Richards BW, Stenolen D, Resau JH, CopelandNG, Jenkins NA. 1999. Direct interaction of microtubule- andactin-based transport motors. Nature 397:267–270.
Ikonen E, Parton RG, Lafont F, Simons K. 1996. Analysis of the role ofp200-containing vesicles in post-Golgi traffic. Mol Biol Cell 7:961–974.
Ikonen E, de Almeid JB, Fath KF, Burgess DR, Ashman K, Simons K,Stow JL. 1997. Myosin II is associated with Golgi membranes:identification of p200 as nonmuscle myosin II on Golgi-derivedvesicles. J Cell Sci 110:2155–2164.
Isenberg G, Small JV. 1978. Filamentous actin, 100 A filaments andmicrotubules in neuroblastoma cells. Their distribution in relationto sites of movement and neuronal transport, Cytobiologie 16:326–344.
Jackman MR, Shurety W, Ellis JA, Luzio JP. 1994. Inhibition of apicalbut not basolateral endocytosis of ricin and folate in Caco-2 cells bycytochalasin D. J Cell Sci 107:2547–2556.
Jimbow K, Quevedo WC Jr, Fitzpatrick TB, Szabo G. 1993. Biology ofmelanocytes. In: Fitzpatrick TB et al., editors. Dermatology ingeneral medicine, 4th ed. New York: McGraw-Hill. p 261–288.
Johnston GC, Prendergast JA, Singer RA. 1991. The Saccharomycescerevisiae MYO2 gene encodes an essential myosin for vectorialtransport of vesicles. J Cell Biol 113:539–551.
Jung G, Wu X, Hammer JA III. 1996. Dictyostelium mutants lackingmultiple classic myosin I isoforms reveal combinations of sharedand distinct functions. J Cell Biol 133:305–323.
Kennedy MB. 1995. Origin of PDZ (DHR, GLGF) domains. TrendsBiochem Sci 20:350.
Kuznetsov SA, Langford G, Weiss DG. 1992. Actin-dependent organ-elle movement in squid axoplasm. Nature 356:722–725.
Kuznetsov SA, Rivera DT, Severin FF, Weiss DG, Langford GM. 1994.Movement of axoplasmic organelles on actin filaments from skeletalmuscle. Cell Motil Cytoskeleton 28:231–242.
Langford GM, Molyneaux BJ. 1998. Myosin V in the brain: mutationslead to neurological defects. Brain Res Rev 28:1–8.
104 A.S. DEPINA AND G.M. LANGFORD

Lazzarino DA, Boldogh I, Smith MG, Rosand J, Pon LA. 1994. Yeastmitochondria contain ATP-sensitive, reversible actin-binding activ-ity. Mol Biol Cell 5:807–818.
Lewis AK, Bridgman PC. 1992. Nerve growth cone lamellipodiacontain two populations of actin filaments that differ in organizationand polarity. J Cell Biol 119:1219–1243.
Liebe S, Menzel D. 1995. Actomyosin-based motility of endoplasmicreticulum and chloroplasts in Vallisneria mesophyll cells. Biol Cell85:207–222.
Machesky LM, Hall A. 1997. Role of actin polymerization and adhesionto extracellular matrix in Rac- and Rho-induced cytoskeletal reorga-nization. J Cell Biol 138:913–926.
Marfatia SM, Leu RA, Branton D, Chishti AH. 1995. Identification ofthe protein 4.1 binding interface on glycophorin C and p55, ahomologue of the Drosophila discs-large tumor suppressor protein. JBiol Chem 270:715–719.
Matteoni R, Kreis TE. 1987. Translocation and clustering of endo-somes and lysosomes depends on microtubules. J Cell Biol 105:1253–1265.
Mercer JA, Seperack PK, Strobel MC, Copeland NG, Jenkins NA.1991. Novel myosin heavy chain encoded by murine dilute coatcolour locus. Nature 349:709–713.
Mermall V, McNally JG, Miller KG. 1994. Transport of cytoplasmicparticles catalysed by an unconventional myosin in living Dro-sophila embryos. Nature 369:560–562.
Mermall V, Post PL, Mooseker MS. 1998. Unconventional myosins incell movement, membrane traffic, and signal transduction. Science279:527–533.
Miyamoto H, Nihonmatsu I, Kondo S, Ueda R, Togashi S, Hirata K,Ikegami Y, Yamamoto D. 1995. Canoe encodes a novel proteincontaining a GLGF/DHR motif and functions with Notch andscabrous in common developmental pathways in Drosophila. GenesDev 9:612–625.
Mogensen MM, Mackie JB, Doxsey SJ, Stearns T, Tucker JB. 1997.Centrosomal deployment of gamma-tubulin and pericentrin: evi-dence for a microtubule-nucleating domain and a minus-end dock-ing domain in certain mouse epithelial cells. Cell Motil Cytoskeleton36:276–290.
Montes de Oca G, Lezama RA, Mondragon R, Castillo AM, Meza I.1997. Myosin I interactions with actin filaments and trans-Golgi-derived vesicles in MDCK cell monolayers. Arch Med Res 28:321–328.
Morris RL, Hollenbeck PJ. 1995. Axonal transport of mitochondriaalong microtubules and F-actin in living vertebrate neurons. J CellBiol 131:1315–1326.
Muallem S, Kwiatkowska K, Xu X, Yin HL. 1995. Actin filamentdisassembly is a sufficient final trigger for exocytosis in nonexcitablecells. J Cell Biol 128:589–598.
Murphy C, Saffrich R, Grummt M, Gournier H, Rybin V, Rubino M,Auvinen P, Lutcke A, Parton RG, Zerial M. 1996. Endosomedynamics regulated by a Rho protein. Nature 384:427–432.
Musch A, Cohen D, Rodriguez-Boulan E. 1997. Myosin II is involved inthe production of constitutive transport vesicles from the TGN. JCell Biol 138:291–306.
Nakagawa H, Miyamoto S. 1998. Actin-filaments localize on thesorting endosomes of 3Y1 fibroblastic cells. Cell Struct Funct23:283–290.
Nascimento AA, Amaral RG, Bizario JC, Larson RE, Espreafico EM.1997. Subcellular localization of myosin-V in the B16 melanomacells, a wild-type cell line for the dilute gene. Mol Biol Cell8:1971–1988.
Novak KD, Peterson MD, Reedy MC, Titus MA. 1995. Dictyosteliummyosin I double mutants exhibit conditional defects in pinocytosis. JCell Biol 131:1205–1221.
Novick P, Botstein D. 1985. Phenotypic analysis of temperature-sensitive yeast actin mutants. Cell 40:405–416.
Parton RG, Dotti CG, Bacallao R, Kurtz I, Simons K, Prydz K. 1991.pH-induced microtubule-dependent redistribution of late endo-somes in neuronal and epithelial cells. J Cell Biol 113:261–274.
Pasternack GR, Racusen RH. 1989. Erythrocyte protein 4.1 binds andregulates myosin. Proc Natl Acad Sci USA 86:9712–9716.
Pastural E, Barrat FJ, Dufourcq-Lagelouse R, Certain S, Sanal O,Jabado N, Seger R, Griscelli C, Fischer A, de Saint Basile G. 1997.Griscelli disease maps to chromosome 15q21 and is associated withmutations in the myosin-Va gene. Nat Genet 16:289–292.
Pol A, Ortega D, Enrich C. 1997. Identification of cytoskeleton-associated proteins in isolated rat liver endosomes. Biochem J327:741–746.
Ponting CP. 1995. AF-6/cno: neither a kinesin nor a myosin, but a bit ofboth. Trends Biochem Sci 20:265–266.
Ponting CP, Phillips C. 1995. DHR domains in syntrophins, neuronal
NO synthases and other intracellular proteins. Trends Biochem Sci20:102–103.
Post PL, Bokoch GM, Mooseker MS. 1998. Human myosin-IXb is amechanochemically active motor and a GAP for rho. J Cell Sci111:941–950.
Prasad R, Gu Y, Alder H, Nakamura T, Canaani O, Saito H, HuebnerK, Gale RP, Nowell PC, Kuriyama K, Miyazaki Y, Croce E, CanaaniE. 1993. Cloning of the ALL-1 fusion partner, the AF-6 gene,involved in acute myeloid leukemias with the t(6;11) chromosometranslocation. Cancer Res 53:5624–5628.
Prekeris R, Terrian DM. 1997. Brain myosin V is a synaptic vesicle-associated motor protein: evidence for a Ca21-dependent interactionwith the synaptobrevin-synaptophysin complex. J Cell Biol 137:1589–1601.
Provance DW Jr, Wei M, Ipe V, Mercer JA. 1996. Cultured melanocytesfrom dilute mutant mice exhibit dendritic morphology and alteredmelanosome distribution. Proc Natl Acad Sci USA 93:14554–14558.
Raposo G, Cordonnier MN, Tenza D, Menichi B, Durrbach A, LouvardD, Coudrier E. 1999. Association of myosin I alpha with endosomesand lysosomes in mammalian cells. Mol Biol Cell 10:1477–1494.
Rinnerthaler G, Herzog M, Klappacher M, Kunka H, Small JV. 1991.Leading edge movement and ultrastructure in mouse macrophages.J Struct Biol 106:1–16.
Rodionov VI, Hope AJ, Svitkina TM, Borisy GG. 1998. Functionalcoordination of microtubule-based and actin-based motility in mela-nophores. Curr Biol 8:165–168.
Rogers SL, Gelfand VI. 1998. Myosin cooperates with microtubulemotors during organelle transport in melanophores. Curr Biol8:161–164.
Ruchhoeft ML, Harris WA. 1997. Myosin functions in Xenopus retinalganglion cell growth cone motility in vivo. J Neurobiol 32:567–578.
Salzman NH, Maxfield FR. 1989. Fusion accessibility of endocyticcompartments along the recycling and lysosomal endocytic path-ways in intact cells. J Cell Biol 109:2097–2104.
Satiat-Jeunemaitre B, Steel C, Hawes C. 1996. Golgi-membranedynamics are cytoskeleton dependent. Protoplasma 191:21–23.
Searle AG. 1952. A lethal allele of dilute in the house mouse. Heredity6:395–401.
Shariff A, Luna EJ. 1990. Dictyostelium discoideum plasma mem-branes contain an actin-nucleating activity that requires ponticulin,an integral membrane glycoprotein. J Cell Biol 110:681–692.
Simon VR, Pon LA. 1996. Actin-based organelle movement. Experien-tia 52:1117–1122.
Simon VR, Swayne TC, Pon LA. 1995. Actin-dependent mitochondrialmotility in mitotic yeast and cell-free systems: identification of amotor activity on the mitochondrial surface. J Cell Biol 130:345–354.
Simon JP, Shen TH, Ivanov IE, Gravotta D, Morimoto T, Adesnik M,Sabatini DD. 1998. Coatomer, but not P200/myosin II, is requiredfor the in vitro formation of trans-Golgi network-derived vesiclescontaining the envelope glycoprotein of vesicular stomatitis virus.Proc Natl Acad Sci USA 95:1073–1078.
Small JV, Celis JE. 1978. Filament arrangements in negativelystained cultured cells: the organization of actin. Cytobiologie 16:308–325.
Small JV, Isenberg G, Celis JE. 1978. Polarity of actin at the leadingedge of cultured cells. Nature 272:638–639.
Sogo LF, Yaffe MP. 1994. Regulation of mitochondrial morphology andinheritance by Mdm10p, a protein of the mitochondrial outermembrane. J Cell Biol 126:1361–1373.
Stow JL, Heimann K. 1998. Vesicle budding on Golgi membranes:regulation by G proteins and myosin motors. Biochim Biophys Acta1404:161–171.
Stow JL, Fath KR, Burgess DR. 1998. Budding roles for myosin II onthe Golgi. Trends Cell Biol 8:138–141.
Sturmer K, Baumann O, Walz B. 1995. Actin-dependent light-inducedtranslocation of mitochondria and ER cisternae in the photoreceptorcells of the locust Schistocerca gregaria. J Cell Sci 108:2273–2283.
Svitkina TM, Neyfakh AA Jr, Bershadsky AD. 1986. Actin cytoskeletonof spread fibroblasts appears to assemble at the cell edges. J Cell Sci82:235–248.
Symons M, Derry JM, Karlak B, Jiang S, Lemahieu V, McCormick F,Francke U, Abo A. 1996. Wiskott-Aldrich syndrome protein, a noveleffector for the GTPase CDC42Hs, is implicated in actin polymeriza-tion. Cell 84:723–734.
Tabb JS, Molyneaux BJ, Cohen DL, Kuznetsov SA, Langford GM.1998. Transport of ER vesicles on actin filaments in neurons bymyosin V. J Cell Sci 111:3221–3234.
Takagishi Y, Oda S, Hayasaka S, Dekker-Ohno K, Shikata T, InouyeM, Yamamura H. 1996. The dilute-lethal (dl) gene attacks a Ca21
105ACTIN FILAMENTS AND MYOSIN MOTORS

store in the dendritic spine of Purkinje cells in mice. Neurosci Lett215:169–172.
Temesvari LA, Bush JM, Peterson MD, Novak KD, Titus MA, Cardelli,JA. 1996. Examination of the endosomal and lysosomal pathways inDictyostelium discoideum myosin I mutants. J Cell Sci 109:663–673.
Tilney LG, Bonder EM, DeRosier DJ. 1981. Actin filaments elongatefrom their membrane-associated ends. J Cell Biol 90:485–494.
Valderrama F, Babia T, Ayala I, Kok JW, Renau-Piqueras J, Egea G.1998. Actin microfilaments are essential for the cytological position-ing and morphology of the Golgi complex. Eur J Cell Biol 76:9–17.
Valentijn KM, Gumkowski FD, Jamieson JD. 1998. The subapicalactin cytoskeleton regulates secretion and membrane retrieval inpancreatic acinar cells. J Cell Sci 112:81–96.
Van Deurs B, Holm PK, Kayser L, Sandvig K. 1995. Delivery tolysosomes in the human carcinoma cell line HEp-2 involves an actinfilament-facilitated fusion between mature endosomes and preexist-ing lysosomes. Eur J Cell Biol 66:309–323.
Vorobjev IA, Svitkina TM, Borisy GG. 1997. Cytoplasmic assembly ofmicrotubules in cultured cells. J Cell Sci 110:2635–2645.
Wagner MC, Barylko B, Albanesi JP. 1992. Tissue distribution andsubcellular localization of mammalian myosin I. J Cell Biol 119:163–170.
Walker J, Dale M, Standart N. 1996. Unmasking mRNA in clamoocytes: role of phosphorylation of a 3’ UTR masking element-binding protein at fertilization. Dev Biol 173:292–305.
Wang YL. 1985. Exchange of actin subunits at the leading edge of
living fibroblasts: possible role of treadmilling. J Cell Biol 101:597–602.
Wei Q, Wu X, Hammer JA III. 1997. The predominant defect in dilutemelanocytes is in melanosome distribution and not cell shape,supporting a role for myosin V in melanosome transport. J MuscleRes Cell Motil 18:517–527.
Weil D, Levy G, Sahly I, Levi-Acobas F, Blanchard S, El-Amraoui A,Crozet F, Philippe H, Abitbol M, Petit C. 1996. Human myosin VIIAresponsible for the Usher 1B syndrome: a predicted membrane-associated motor protein expressed in developing sensory epithelia.Proc Natl Acad Sci USA 93:3232–3237.
Weiner OH, Murphy J, Griffiths G, Schleicher M, Noegel AA. 1993.The actin-binding protein comitin (p24) is a component of the Golgiapparatus. J Cell Biol 123:23–34.
Wu X, Bowers B, Wei Q, Kocher B, Hammer JA III. 1997. Myosin Vassociates with melanosomes in mouse melanocytes: evidence thatmyosin V is an organelle motor. J Cell Sci 110:847–859.
Wu X, Bowers B, Rao K, Wei Q, Hammer JA III. 1998. Visualization ofmelanosome dynamics within wild-type and dilute melanocytessuggests a paradigm for myosin V function in vivo. J Cell Biol143:1899–1918.
Yaffe MP. 1999. The machinery of mitochondrial inheritance andbehavior. Science 283:1493–1497.
Yonemura S, Pollard TD. 1992. The localization of myosin I andmyosin II in Acanthamoeba by fluorescence microscopy. J Cell Sci102:629–642.
106 A.S. DEPINA AND G.M. LANGFORD


![The Physical Interaction of Myoblasts with the ... … · is believed to increase the interaction of FA proteins with actin filaments and lead to integrin clustering [12]. Myosin-II](https://img.pdfslide.us/doc/110x75/5f2c3d62797c78131c4e3513/the-physical-interaction-of-myoblasts-with-the-is-believed-to-increase-the.jpg)


























