Embed Size (px)
Citation preview

Vasoactive Intestinal Polypeptidein Sacral Primary Sensory Pathways
in the Cat
CHARLES W. MORGAN,1* PETER T. OHARA,2 AND DAVID E. SCOTT1
1Department of Pathology and Anatomy, Eastern Virginia Medical School,Norfolk, Virginia 23501
2Department of Anatomy, W.M. Keck Foundation Center for Integrative Neuroscience,University of California, San Francisco, California 94143-0452
ABSTRACTUnmyelinated sensory axons in the sacral spinal cord may play a role in bladder reflexes
under certain pathological conditions. Previous data suggested vasoactive intestinal polypep-tide (VIP) might be contained exclusively in sensory C-fibers, some of which innervate thebladder. This study was undertaken to describe the morphology of these VIP fibers in thesacral cord of the cat. VIP immunoreactivity was confined to unmyelinated axons observed atseveral levels of the sensory pathway including the dorsal root ganglia, dorsal roots,Lissauer’s tract, and the lateral collateral pathway. A combination of light and electronmicroscopic observations showed VIP-immunoreactive fibers with labeled varicosities andsynaptic terminals in laminae I, IIo, V, VII, and X. VIP-immunolabeled varicosities had amean diameter of 1.6 µm (range 5 0.11–7.4 µm, S.D. 5 1.01, n 5 311) with a small percentage(8%) being relatively large (3–7.4 µm). VIP varicosities contained a mixture of small clearvesicles (CLV) and large dense core vesicles (LDV). Although most varicosities contained amoderate number of LDVs (14.86 LDVs/µm2), some varicosities contained a large number ofLDVs, whereas others contained very few. Varicosities that possessed synaptic specializationswere classed as terminals and were divided into three morphological classes. Two of theseresembled Gray’s Type I terminal, whereas a third was similar to the Gray’s Type II terminal.There was no consistent relationship between vesicle content of the terminal and the type ofsynaptic contact it possessed. This study shows that in the sacral spinal cord of the cat, VIPterminals originate only from C-fibers, terminate primarily in laminae I and V, and exhibit avariety of morphologies consistent with heterogeneous origins and functions of the lowerurinary tract. J. Comp. Neurol. 407:381–394, 1999. r 1999 Wiley-Liss, Inc.
Indexing terms: C-fibers; VIP; bladder; visceral sensory; parasympathetic; pelvic nerve
The sacral spinal cord receives a major input of visceralsensory axons from the pelvic organs (Morgan et al., 1981).In the cat, these primary afferents, carried by the pelvicnerve, innervate the bladder, colon, and sex organs andterminate in the same segments that contain the sacralparasympathetic nucleus. These visceral sensory axonscontribute both to the reflexes and to perception of thepelvic organs. An interesting feature in this system is thelateral collateral pathway (LCP), a prominent distributionof axon collaterals originating from Lissauer’s tract andcoursing through the lateral extent of lamina I into laterallamina V. The LCP is composed of unmyelinated C-fibersand thinly myelinated A-d fibers, derived in large partfrom visceral and somatic primary sensory axons in thepelvic and pudendal nerves (Morgan et al., 1981; Thor etal., 1989). Both of these classes of sensory axon have been
shown to play key roles in the pelvic reflexes (de Groat andKrier, 1978; de Groat et al., 1982; de Groat, 1990). Al-though a variety of neurotransmitters are probably in-volved in this system, vasoactive intestinal polypeptide(VIP) has been proposed to act as a neurotransmitter inbladder afferents and has a unique distribution in thesacral spinal cord consistent with this role (de Groat et al.,1986).
Grant sponsor: NIDDK; Grant number: R01 DK49480.*Correspondence to: Dr. Charles W. Morgan, Department of Pathology
and Anatomy, Eastern Virginia Medical School, 700 Olney Rd., Norfolk, VA23501. E-mail: [email protected]
Received 14 September 1998; Revised 31 December 1998; Accepted 12January 1999
THE JOURNAL OF COMPARATIVE NEUROLOGY 407:381–394 (1999)
r 1999 WILEY-LISS, INC.

During normal conditions, the sensory limb of thebladder reflex is served by A-d axons (de Groat et al., 1981,1982) but in pathologic conditions, such as spinal cordinjury or painful irritation of the bladder, C-fibers areinvolved (de Groat et al., 1981, 1993). To analyze thebladder reflexes in normal and pathologic conditions it isessential to have an accurate description of the sensorypathways involved in the bladder reflex. C-fibers areparticularly difficult to trace because of their small diam-eters, and previous studies on C-fibers in the spinal cordhave led to conflicting conclusions regarding the lamina inwhich these fibers terminate (Gobel and Binck, 1977;LaMotte, 1977; Gobel and Falls, 1979; Gobel and Hock-field, 1979; Ralston and Ralston, 1979; Beal and Becknell,1981).
The present study addresses several issues with respectto sacral spinal cord afferents. First, as there is evidencethat VIP might be confined to unmyelinated axons (Hondaet al., 1983; Morgan and Ohara, 1984), we wished to usethe immunocytochemical demonstration of VIP to deter-mine in which laminae these C-fibers terminate. Secondly,we describe the distribution and ultrastructural appear-ance of VIP-containing terminals in the sacral spinal cord.This information is needed to fully understand the role ofvisceral afferents in the bladder reflex and to provide thebasis for further analysis of the role of VIP in the bladderreflexes in the normal and pathologic condition.
MATERIALS AND METHODS
Four male cats, weighing 2–5 kg were anesthetized withsodium pentobarbital (30 mg/kg, i.p.). The chest wasopened and the cat was perfused intracardially withphosphate-buffered saline (PBS), followed by a first fixa-tive of 4% paraformaldehyde and 0.1% glutaraldehyde inphosphate buffer, pH 7.4, and then a second fixative of 4%paraformaldehyde containing sodium-m-periodate (0.55g/l) and lysine (3.42 g/l). All procedures were carried outaccording to the standards established by the NIH Guidefor Care and Use of Laboratory Animals (National Insti-tutes of Health Publications No. 80–23, revised 1978) andthe protocols were approved by the UCSF Committee forAnimal Research. All efforts were made to minimizeanimal suffering and to reduce the number of animalsused.
Following a laminectomy, the S2 dorsal root was identi-fied by its location in the sacral foramen, marked with aligature, and the spinal cord was removed. The cord wascut into segments identified by using the most caudalrootlet of the marked S2 dorsal root to define the caudalborder of the S2 segment. The identified segments werepostfixed for three hours in the second fixative and storedover night, 12–16 hours in 0.1 M PBS at 4°C.
The spinal segments were cut at 30- and 50-µm sectionswith a Vibratome and collected in Tris-buffered saline(TBS) pH 7.3. The collected sections were washed in asolution of TBS, 0.02% saponin and 1% normal sheepserum, rinsed in TBS, and incubated in VIP antiserum(Immuno Nuclear, 1:1,000) at 4°C for 12–24 hours. Controlsections were processed similarly except the VIP antise-rum was preadsorbed with VIP at concentrations of 10, 50,and 100 µg/µl. Other control sections were not exposed tothe VIP antiserum or were exposed to leucine enkephalin(LENK) antiserum. All tissue was then washed in TBS andtransferred to a sheep anti-rabbit IgG, (1:20) for 30
minutes. Following thorough washing in TBS, the sectionswere incubated in rabbit peroxidase-antiperoxidase com-plex (1:100) for 30 minutes. The sections were then rinsedand reacted in 0.01% 3,38diaminobenzidine tetrahydrochlo-ride (DAB) and 0.02% hydrogen peroxide for 20 minutes.After rinsing the sections were fixed in 1% osmium tetrox-ide containing 1.5% potassium ferricyanide, stained with1% uranyl acetate, dehydrated, and infiltrated with Epon.The sections were then embedded in Epon, sandwichedbetween two plastic slides, and polymerized for 72 hours at70°C.
The hardened epon sections were cut into 4-µm serialsections and mounted onto Epon blocks that could then beexamined with the light microscope. Regions of interestwith the heaviest label were trimmed from the 4-µmsections and thin-sectioned on an ultramicrotome. Thethin sections were mounted on 1 3 2 mm slot grids, stainedwith Reynold’s lead citrate and examined with the electronmicroscope (EM).
Camera lucida drawings and Polaroid pictures weremade with the light microscope from the embedded sec-tions and used to track the areas from which the electronmicrographs were taken.
Large dense vesicles (LDVs) in labeled varicosities werecounted manually from enlarged micrographs by using athumb counter and marking counted vesicles with a felt tippen. Vesicle size was determined by projecting the EMnegative of labeled varicosities onto a sheet of paper anddrawing around the perimeter of the vesicles. These datawere scanned into a computer and average diameters weredetermined using Mocha software (Jandel Scientific, CorteMadera, CA). Both LDVs and clear round vesicles (CLVs)were measured. Varicosity size was determined by scan-ning their images into Mocha and measuring their averagediameters.
RESULTS
Light microscopic (LM) examination:the distribution of VIP
VIP immunoreactivity (VIP-ir) was present in the dorsalroot ganglia, dorsal roots, and spinal cord segments S1–S3.Label was highly concentrated in S1–S3 in all four cats butwas found only in trace amounts in the adjacent lumbarand coccygeal segments. As illustrated in Figure 1, VIP-irwas observed in Lissauer’s tract, throughout lateral laminaI and in lamina V, extending at times beyond the centralcanal through lamina X and the dorsal commissure to theopposite side of the cord. Infrequently, label was also seenin medial lamina I, outer lamina II, and lamina VII. Thedistribution through lateral lamina I, the location of theLCP, occurred at regular intervals in the longitudinalplane consistent with previous observations of visceralafferents (Morgan et al., 1981), VIP (Kawatani et al.,1983b; Honda et al., 1983; Basbaum and Glazer, 1983),and other peptide neurotransmitter candidates (Kawataniet al., 1985; de Groat et al., 1986; Vizzard et al., 1994). TheVIP- immunoreactive collaterals from Lissauer’s tractformed bundles of axons that traveled in a rostral andventral direction from lamina I into lamina V. The LCP inFigure 1 illustrates two partial bundles of labeled fibersand an intervening space (c), free of labeled axons. Theupper part originated from Lissauer’s tract in this sectionwhereas the lower bundle originated from Lissauer’s tractabout 150 µm caudal to this section.
382 C.M. MORGAN ET AL.

The 50 µm Epon-embedded sections were cut seriallyinto 4-µm sections and mounted onto glass slides foridentification of VIP-ir-rich locations. The density of labelseen in 50-µm sections (Fig. 1) was noticeably reducedwhen sections were cut 4-µm-thick (Fig. 2). The 4-µmsections were remounted onto Epon blocks and cut into
ultrathin sections for EM examination. Drawings andPolaroid photomicrographs were made to keep track of thelocations of the subsequent electron micrographs. Samplesof VIP labeling were taken from specific regions of Lissau-er’s tract and the LCP as demarcated in Figure 1, but noevidence of regional variations was observed.
Attached to some of the 50-µm Vibratome sections ofsacral cord were dorsal rootlets between 1 and 5 mm long.VIP-ir was easily identified in these rootlets adjacent tothe spinal cord in all tissue where they were present.Figure 2 illustrates in transverse section a rootlet prior toits merger with Lissauer’s tract. In this and other 4-µmsections viewed at the LM level, VIP-immunoreactiveelements were identified as dark, punctate structures inthe clear regions surrounding the large, myelinated axonsand were not localized to any particular side. No VIP labelwas found within the myelinated axons. In Lissauer’stract, directly adjacent to the rootlet, many small, myelin-ated axons can be seen. Surrounding these axons are theimmunoreactive VIP puncta and lightly stained areas thatcontain unmyelinated axons. VIP-immunoreactive profilescan also be seen in lamina I.
Electron microscopic examination:VIP-immunoreactive structures
Dorsal root ganglia. Evidence of VIP-ir in the dorsalroot ganglia was considerably less than in any other tissueexamined. The basis for this is unknown but is consistentwith other studies in which colchicine pretreatment wasnecessary to demonstrate the peptide content of dorsalroot ganglion neurons (Kawatani et al., 1982; de Groat etal., 1986). The only evidence of label was sparse andconsisted of immunoreactive vesicles found in unmyelin-ated afferents (Fig. 3A). The labeled axons were notconcentrated in any particular area of the roots and werefound in Remack bundles combined with other unmyelin-ated axons. VIP label was not observed in dorsal rootganglion somata or in myelinated axons.
Dorsal roots. The VIP-immunoreactive axons in thedorsal rootlets were usually bundled with small numbersof other, unlabeled unmyelinated axons as was also seen inthe dorsal root ganglia (Fig. 3A, B). As the attachedrootlets merged with the spinal cord, they continuedanother 1–2 mm to join Lissauer’s tract, thus enabling usto trace VIP- immunoreactive axons for up to 6 mm as
Figure 1
Fig. 1. Fifty-µm section of the upper quadrant of S2 spinal cord,showing the distribution of vasoactive intestinal polypeptide (VIP)immunolabel in Lissauer’s tract (LT), lateral lamina I (b,c), junction oflaminae I and V (d), lateral lamina V (e), and medial lamina V(f). Label in this region outlines the lateral collateral pathway (LCP).Lesser amounts of label were seen in medial lamina I (a), laterallamina VII (h), and bilaterally around the central canal (g). The opentriangles denote the borders of Lissauer’s tract. Closed trianglesindicate VIP reaction product in varicostities throughout Lissauer’stract and lamina I. Scale bar 5 200 µm.
Fig. 2. Semithin 4-µm Epon-embedded section of a large, rounded,dorsal rootlet on the dorsolateral surface of the spinal cord (S2).(Lateral is to the left, dorsal is up.) Vasoactive intestinal polypeptide(VIP) reaction product can be seen as dark, punctate structures(arrowheads) within the unstained areas surrounding the largermyelinated axons within the rootlet. The smallest of the myelinatedaxons and the unmyelinated axons in the unstained areas will joinLissauer’s tract (LT) located directly beneath, over the next 1–2 mmrostrally. Punctate VIP-immunolabeled structures (arrowheads) canalso be seen in LT, and lamina I (I). The borders between LT, lamina I, andlamina II (II) are indicated by the large black dots. Scale bar 5 25 µm.Figure 2
VIP IN UNMYELINATED AXONS 383
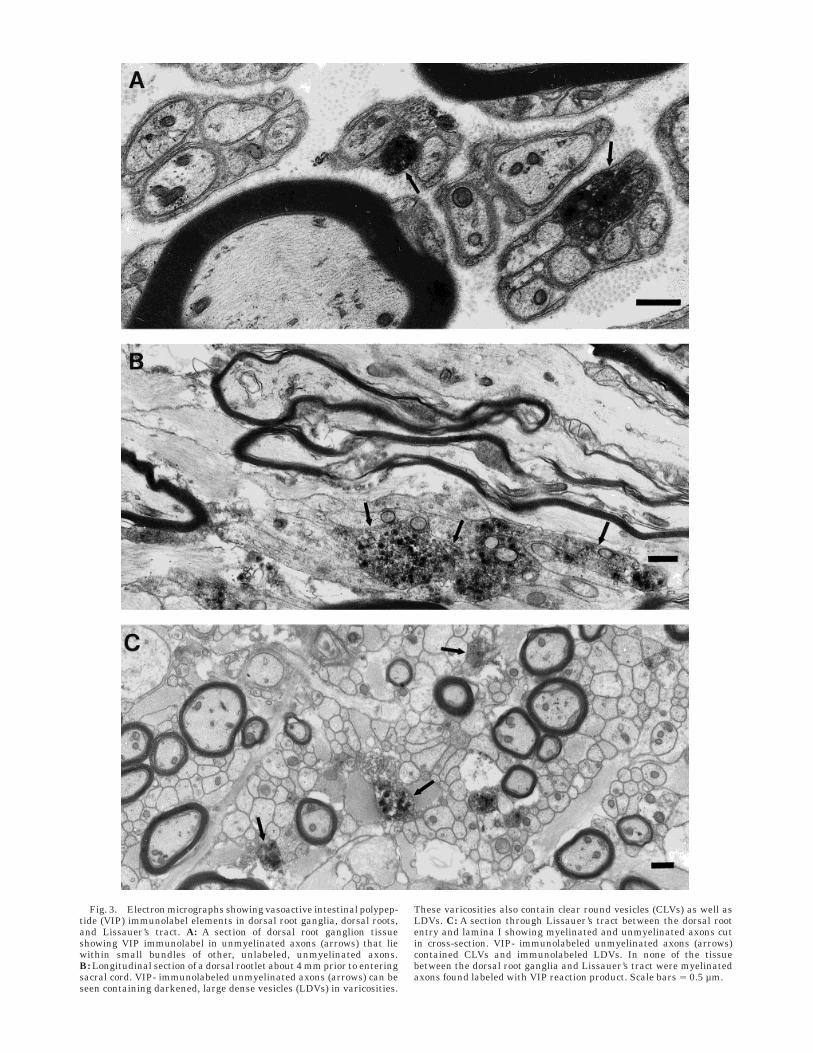
Fig. 3. Electron micrographs showing vasoactive intestinal polypep-tide (VIP) immunolabel elements in dorsal root ganglia, dorsal roots,and Lissauer’s tract. A: A section of dorsal root ganglion tissueshowing VIP immunolabel in unmyelinated axons (arrows) that liewithin small bundles of other, unlabeled, unmyelinated axons.B: Longitudinal section of a dorsal rootlet about 4 mm prior to enteringsacral cord. VIP- immunolabeled unmyelinated axons (arrows) can beseen containing darkened, large dense vesicles (LDVs) in varicosities.
These varicosities also contain clear round vesicles (CLVs) as well asLDVs. C: A section through Lissauer’s tract between the dorsal rootentry and lamina I showing myelinated and unmyelinated axons cutin cross-section. VIP- immunolabeled unmyelinated axons (arrows)contained CLVs and immunolabeled LDVs. In none of the tissuebetween the dorsal root ganglia and Lissauer’s tract were myelinatedaxons found labeled with VIP reaction product. Scale bars 5 0.5 µm.

dorsal root axons. In this region, labeled axons werenumerous and were distributed among the other unmyelin-ated axons where they were easy to find. Figure 3Billustrates unmyelinated axons containing VIP- immunore-active vesicles and forming varicosities adjacent to unla-beled myelinated axons.
Lissauer’s tract. Within Lissauer’s tract, darkenedvesicles indicating VIP-ir were observed in unmyelinatedaxons (Fig. 3C) and in occasional varicosities and termi-nals (Fig. 4A). In S2, VIP label was distributed evenlythroughout the tract without predominance medially orlaterally. Although VIP-ir was observed in S1–S3, it wasmost prevalent in S2 and most of the EM data werecollected in S2.
Some of the unmyelinated VIP-immunoreactive axonswere larger in diameter than many of the small, myelin-ated axons. A survey of the myelinated axons in Lissauer’stract near upper lateral lamina I (Fig.3B) revealed diam-eters ranging from 0.2 to 1.7 µm (mean 5 0.7, S.D. 5 0.32,n 5 90). VIP-labeled varicosities and unmyelinated axondiameters ranged from 0.21 to 0.96 µm (mean 5 0.49,S.D. 5 0.2 , n 5 15). Unlabeled unmyelinated axondiameters, which ranged from 0.2 to 1.1 µm (mean 5 0.3,S.D. 5 0.16, n 5 704), also overlapped those of themyelinated axons in this region.
Gray matter: varicosities and vesicles. VIP-immu-noreactive varicosities were observed throughout the dis-tribution of the LCP from Lissauer’s tract through laterallamina I to medial lamina V. Over 500 varicosities, includ-ing some followed serially, were examined in spinal tissuefrom four cats. Of these varicosities, 290 were used in thecounts and measurements reported below. The numberswe have reported do not reflect random sampling, as it wasour intent to locate and examine synaptic specializations.The specific regions of the LCP from which the varicositieswere taken were recorded to determine if there weresignificant differences between regions, but no such differ-ences were apparent.
VIP-immunoreactive varicosities were seen as dark,punctate areas which were easily visible at the LM and atlow magnification EM levels (Figs. 2, 3C, 4C). The labeledvaricosities were small, with a mean of 1.6 µm in diameter(range 5 0.11–7.4 µm, S.D. 5 1.01, n 5 311), though asmall percentage, 8%, were relatively large (3– 7.4 µm).Immunolabeled varicosities in the gray matter (Figs. 4–6)were small compared to those in the dorsal roots (Fig. 3B).Several varicosities were followed over 10–20 serial sec-tions in order to determine the degree of consistency oftheir composition and also to find synaptic specializations.From analysis of these serial sections it was concludedthat individual sections are fairly representative of theentire varicosity.
Tightly packed, evenly distributed vesicles were theprimary inclusions in the labeled varicosities (Figs. 4–6).At higher magnifications it was seen that the varicositiescontained darkened LDVs and a dark, flocculent materialcovering the membranes of all inclusions, clear vesiclesCLVs, mitochondria, and the internal membrane of thevaricosity itself (Figs. 5, 6). In a few cases, the axonsleading to the darkened varicosities appeared to be unla-beled (Fig. 3B, C). In the unlabeled part of the axons, therewere only CLVs, whereas the labeled part of the axon andthe varicosities contained both LDVs and CLVs.
The LDVs ranged in size from 64.7 to 139.29 nm(mean 5 95.71, S.D. 5 16.03, n 5 142) and were round
with a darkened core. The CLVs were mostly round,though some were irregular and ovoid with diametersranging from 23.16 to 65.1 nm (mean 5 44.27, S.D. 5 6.87,n 5 219). These measurements must be considered to bereasonable approximations as the vesicle membranes wereoften covered with flocculent reaction product that im-paired the precision of these measurements.
Immunolabeled varicosities containing LDVs and CLVswere seen at all levels of the sensory distribution (Figs. 3,4–6) and presented a variety of morphologies. Althoughthe appearance of individual varicosities were quite simi-lar when followed over multiple serial sections, the appear-ance of different varicosities could be quite variable,suggesting a heterogeneous population. The principle vari-ability was related to the ratio of CLVs to LDVs within avaricosity and to the density of packing of these vesicles(for example, Figs. 5E, 6A). The majority of varicositiescontained both CLVs and LDVs but the percentage of eachclass of vesicle in each varicosity varied such that somevaricosities contained largely CLVs (Fig. 5A, E), whereasothers contained largely LDVs (Fig. 6A, B). These lattertypes of varicosity seemed to represent the extreme ends ofa distribution which in the middle included more of amixture of the two vesicle types (Figs. 4A, 5C, F). Inaddition to the darkly stained LDVs, some of these varicosi-ties also displayed a dark background (Fig. 6A, B, F).
In order to establish more quantitative and objectivecriteria to evaluate these different immunoreactive varicosi-ties, the LDVs in each were counted and the area of thevaricosity in which they were contained was determined.From these data it was established that density of LDVsvaried from 0 to 64 LDV/µm2 for 290 varicosities. Afrequency histogram of these data (Fig. 7) shows a skeweddistribution with a mean density of 14.86 LDV/µm2 (n 5290; S.D. 10.04) and with the preponderance (93%) of theprofiles having less than 30 LDV/µm2. The varicositiesshown in Figures 4A and 5F have an LDV density (13.16and 15.23) close to the mean.
Normally, CLVs were not counted because their smallsize and overlap within a section made this impracticaland potentially inaccurate. However, CLVs appeared tohave an inverse relationship with the LDVs, i.e., a lowLDV number indicates large numbers of CLVs. In anumber of labeled varicosities where the vesicles wereclear, both types were counted to test this idea. Forexample, in Figure 6E all vesicles were counted and CLVstotaled 324 (104.18/µm2) compared to 15 LDVs (4.8/µm2).It was found that the varicosities that appear to containmostly CLVs (Figs. 4B; 5A, B, E; 6C, E ) have an LDV countless than 9 LDV/µm2. Varicosities with less than 9 LDV/µm2 make up 31% of the entire population in the currentstudy and insinuate a functionally different type of termi-nal than those with a higher density of LDVs. Figure 6Aillustrates a varicosity filled with LDVs with an LDVdensity of 49.19/µm2 and with very few CLVs visible exceptfor those near the synapse.
At the EM level, individual VIP-immunoreactive axonscould not be traced very far before they were lost. It wasnoted that successive varicosities clearly belonging to thesame axon, as well as those possibly belonging to that axonby following the same trajectory, had a similar appearancein terms of their vesicle content. At the same time indifferent axons with parallel trajectories, varicosities witha different vesicle content were observed. These observa-tions imply that single VIP axons may have similar,
VIP IN UNMYELINATED AXONS 385

Fig. 4. A: A vasoactive intestinal polypeptide (VIP)-immunola-beled terminal surrounding a dendritic spine in Lissauer’s tract (LT).Label is indicated by the darkened large dense vesicles (LDVs;13.16/µm2). The synaptic specialization (arrowhead) is tangentiallycut in this section. This terminal is surrounded by neural structuresincluding vesicle containing profiles (VCPs, marked by asterisks) withno intervening glial elements for nearly all of its outer perimeter.B: An unmyelinated axon in the lateral collateral pathway (LCP) iscontinuous with a labeled, darkened varicosity packed with clear,round vesicles (CLVs) and LDVs (18.59/µm2). C: A VIP- immunola-beled terminal (18.55 LDVs per µm2) in the LCP, lateral lamina I. The
synaptic specialization (white arrow) is a type I or Is. The postsynapticstructure contains a variety of clear vesicles and could be either anaxon or a dendrite. The arrowheads mark unmyelinated axons thattravel in parallel bundles and which are characteristic of the LCP inboth lamina I and V. Note in both B and C that at the upper arrows theprevaricose region of the axon and the CLVs it contains are notdarkened by VIP reaction product, whereas at the lower arrows a darklabel begins and continues into the varicosity. Also note that in eachexample there are no LDVs in the unlabeled part of the axon. Scalebars 5 0.5 µm.
386 C.M. MORGAN ET AL.

Fig. 5. A: A type I terminal with asymmetric specialization, thathas a well-developed subsynaptic web (arrow), and contains mostlyclear round vesicles (CLVs; 2.44 large dense vesicles [LDVs]/µm2). B: Atype I terminal with an asymmetric specialization, presynaptic grid(arrow), subsynaptic web, and Taxi bodies. This terminal containsmostly CLVs (8.53 LDVs/µm2). C: A type I terminal with an asymmet-ric specialization, a loose knit subsynaptic web (arrow), and a largenumber of LDVs (18.67/µm2). D: A type Is terminal with presynaptic
and postsynaptic thickenings that are of equal size, giving it asymmetric appearance. This terminal contains 7.92 LDVs/µm2. E: Atype II symmetric terminal contacting a dendritic spine. This is anexample of a terminal with few LDVs (0.94 LDVs/µm2). F: A type Iterminal contacting a dendritic spine and containing close to theaverage number of LVDs, (15.23/µm2). This terminal lies close to anumber of other vesicle containing profiles (VCPs). Scale bars 50.25 µm.

Figure 6
388 C.M. MORGAN ET AL.

multiple varicosities; whereas in the same region, otherVIP axons may have varicosities with different characteris-tics.
Synaptic specializations and other contacts. Fourdifferent specialized contacts were seen between VIP-immunoreactive profiles and other structures (Table 1). Ofthese four types of contact, three were considered to besynaptic specializations. A total of 290 immunolabeledvaricosities were surveyed (Fig. 8). Of these, 113 (39%) had
synaptic contacts and were then classed as synaptic termi-nals. The synaptic specializations in the current studyhave been categorized as type I, type Is, and type IIbecause of their similarity to Gray’s type I and II (Gray andGuillery, 1966). The type I contact (Fig. 5A–C) had apostsynaptic density that was thicker than the presynap-tic. This type of specialization often had a subsynaptic web(Fig. 5A, C), density, or Taxi bodies (Fig. 5B). In some casesa presynaptic density or grid composed of a series ofregularly spaced triangles was present (Fig. 5B). The typeI contact had a widened synaptic cleft, was often curved,and had round CLVs adjacent to the presynaptic mem-brane. Forty-seven synaptic specializations were type I.
The type Is (Fig. 5D) contact was a variation of the type Ispecialization, identical in most attributes except that itdisplayed a symmetrical appearance. This was the resultof the presynaptic thickening being equal to, or largerthan, the postsynaptic thickening. Thirty type Is synapticspecializations were observed in this study.
The third type of synaptic specialization, type II, alsohad a symmetric appearance (Figs. 5E, 6A, C). The postsyn-aptic density was only slightly thicker (no more than twotimes) than the postsynaptic membrane and appeared as aslight thickening of the postsynaptic membrane. Typically,there were no associated subsynaptic specializations. Therewas a slightly widened synaptic cleft with parallel mem-branes, usually straight, with CLVs clustered on thepresynaptic side. Unlike the conventional Gray’s type IIsynapse which is normally associated with flattenedvesicles, in the present case only round CLVs were seen inthe VIP-immunolabeled type II synapses. It should benoted that all the synaptic vesicles within the labeledvaricosities appeared to be round, even in the type IIterminals with symmetric synaptic contacts. Flattenedvesicles were present in adjacent unlabeled profiles, whichsuggests that the absence of flattened vesicles in thelabeled terminals was not an artifact of fixation or theembedding process. In adjacent serial sections, flattenedand round vesicles were also seen (Fig. 6D) in unlabeled
Fig. 6. A: A type II terminal with a symmetric specialization, andsubsynaptic cisterns (arrow). Note that although the majority of thevesicles are large dense vesicles (LDVs; 49.19/µm2), it is clear roundvesicles (CLVs) that are found near the presynaptic thickening. B: Twovaricosities containing mostly LDVs (19.41/µm2). A dendritic spine(arrow) has a few CLVs surrounding it. C: A type II terminal (arrow)contacting a dendrite. This contact has a thin postsynaptic thickeningand the terminal contains mostly CLVs (2.26 LDVs/µm2). D: Anunlabeled type II terminal (arrow) from a section serial to those shownabove but processed with leucine enkephalin (LENK) antisera as acontrol. This terminal contains flattened vesicles, whereas roundvesicles are present in an adjacent vesicle containing profiles (VCP;asterisk). This indicates that the lack of flattened vesicles at type IIendings in the VIP-immunoreactive terminals is not an artifact offixation or immunohistochemistry. E: Puncta adherentia (arrows)between a VIP-immunoreactive varicosity (4.82 LDVs/µm2) and twounlabeled VCPs. F: A punctum adherens (arrow) between a type Iterminal (7.09 LDVs/µm2) and a VCP. The labeled terminal exhibits adarkened background and is in synaptic contact with a dendritic spine(asterisk). Scale bars 5 0.25 µm.
Fig. 7. Histogram illustrating the numbers of large dense vesicles(LDVs) counted in each varicosity relative to the size of the varicosity.There were 290 varicosities measured and the average ratio ofLDVs/µm2 was 14.86.
Fig. 8. Histogram showing the numbers of large dense vesicles(LDVs) counted in each of the three terminal types from 103 termi-nals. Terminal types were based on the morphology of the synapticspecializations. The majority of terminals were of the type 1 and type1s variety.
TABLE 1. Types of Neural Contacts Made by 99 Vasoactive IntestinalPolypeptide (VIP)-Containing Terminals
SpecializationDendrites
(%)
Dendritic andsomatic
spines (%)
Vesicle-containingprofiles (%)
Somata(%)
Type I 81 16 3 0Type Is 81 14 0 5Type II 65 30 0 5Puncta Adherentia 18 0 67 11Indeterminate 22 67 0 11
VIP IN UNMYELINATED AXONS 389

profiles processed using the same immunocytochemicalprotocol as used for the VIP tissue but substituting LENKantisera. Thus, it also seems unlikely that the lack offlattened vesicles in the VIP-labeled profiles was a result ofthe immunocytochemical procedures. We conclude that thevesicles in VIP terminals are all round (or spherical) andthe classical relationship between round vesicles and Graytype I contacts and flat vesicles and Gray type II contactsdoes not hold true for this tissue.
As not all synaptic contacts could be unambiguouslyidentified and placed in a particular class, serial sectionanalysis was used to reduce the possibility of misclassifica-tion. Some type I specializations followed in serial sectionwere found in a few sections to resemble type II specializa-tions. Therefore, the type II specializations were followedserially whenever possible to ensure they were not type I.At other times, interpretation of terminal type was madedifficult by the plane of cut or because the dark reactionproduct obscured the terminal. In those cases where therewas obviously a terminal but its type was not clear, thespecialization was categorized as ‘‘indeterminate’’ (Fig. 4C).There were nine indeterminate specializations in thecurrent sample.
The fourth type of specialization encountered was classedas punctum adherens (Peters et al., 1976). These struc-tures (Fig. 6E, F) had symmetrical, thick membranethickenings and had a punctate nature. The lack ofvesicles adjacent to the membrane indicates they were nota docking site for vesicles and were unlikely to be involvedin synaptic transmission. Twenty-three puncta adherentiawere counted in the current experiments. Most punctaadherentia were formed between the VIP-immunoreactivevaricosities and a nonlabeled vesicle-containing profile(Table 1).
Nonsynaptic membrane associations
VIP-immunoreactive terminals and varicosities wereoften observed in close contact with dendrites, axons, andnonimmunoreactive vesicle-containing profiles (VCP) fromunidentified sources (Figs. 4; 5F; 6E, F). Estimates of theproportion of perimeters directly apposed to neural struc-tures, glia and/or to empty space were made of 268varicosities. Of the varicosities examined, 192 (73%) had75% or more of their membrane perimeters in contact withneural structures. Forty-one percent of the varicositieswere apposed to neural structures for their entire visibleperimeters. A few varicosities (8.6%) were in direct contactwith glia. For these few varicosities, glial contact rangedfrom 5% to 55% coverage.
DISCUSSION
VIP in unmyelinated sensory axons
In the sacral spinal cord of the cat, VIP is largely, andperhaps entirely, contained in unmyelinated sensory axons.Light microscopic evidence shows VIP in small to mediumdiameter dorsal root ganglion neurons (Honda et al., 1983;Kawatani et al., 1986), indicating that VIP is contained inA-d fibers and/or C-fibers. Honda et al. (1983) reportedobserving VIP in unmyelinated sensory axons in the catsacral cord, though it was not clear that they tried to ruleout the presence of VIP in myelinated axons. In ourexperiments in which tissue was examined at both the LMand EM levels, there was no evidence of VIP-ir in myelin-ated axons. In the dorsal root entry zone near Lissauer’s
tract, punctate VIP-ir was seen in areas surrounding themyelinated axons (Fig. 2), similar to the visceral sensorydistribution from the pelvic nerve in the same location(Morgan et al., 1981). We examined VIP-ir in sacral dorsalroot ganglia, dorsal roots, Lissauer’s tract, and the dorsalhorn, and label was only observed in unmyelinated axons,terminals, and varicosities. Considering these data andthe fact that VIP has never been seen in myelinated axons,it is reasonable to conclude that VIP is only present inunmyelinated axons. It is possible that some VIP-immuno-reactive fibers and varicosities could arise from sourcesintrinsic to the spinal cord, but transection of the sacraldorsal roots leads to nearly complete loss of VIP-contain-ing axons, showing there is little or no intrinsic source ofVIP with the exception of a small amount in the ependy-mal layer of the central canal (Kawatani et al., 1983b,1985; Honda et al., 1983). These observations enable VIPto be used as a marker for unmyelinated, predominantlysensory, axons in the sacral spinal cord.
There is a close correspondence between the sacraldistribution of VIP within Lissauer’s tract and LCP inlaminae I, V, and X (Basbaum and Glazer, 1983; Honda etal., 1983; Kawatani et al., 1985, 1986; LaMotte anddeLanerolle, 1986) and the sacral distribution of sensoryaxons from the bladder and urethra carried by the pelvicand pudendal nerves (Morgan et al., 1981; de Groat et al.,1986; Thor et al., 1989). The pelvic nerve consists predomi-nantly of A-d and C-fibers, many of which employ peptidessuch as VIP, substance P, cholecystokinin, and LENK asneurotransmitters (Kawatani et al., 1985, 1986). VIP-containing sensory axons, which compose a specific, unmy-elinated subpopulation of this distribution, have beentraced from the urinary bladder and colon and are thoughtto have a role in micturition and defecation (Keast and deGroat, 1992). The distribution of sensory axons from thepelvic nerve in the LCP has been shown by horseradishperoxidase tracing studies and it has long been a reason-able assumption that, in addition to axons of passage, axonterminals are also present in the LCP (Morgan et al.,1981). The present results confirm this assumption by thedemonstration of VIP-immunoreactive axons and termi-nals throughout the entire LCP.
Our finding that VIP-immunoreactive terminals occurin laminae I, V, VII, and X is at odds with the conclusions ofothers (Rethelyi, 1977; LaMotte, 1977; Ralston and Rals-ton, 1979; Beal and Bicknell, 1981) that C-fibers terminateonly in lamina II. Although not all of these other studieswere carried out in the sacral spinal cord, it was impliedthat such an organization may be a general feature of thespinal cord. The difference between our results and othersmay be a consequence of the different methodologiesemployed. The conclusions of Ralston and Ralston (1979)were based on the electron microscopic appearance ofdegenerating terminals in the dorsal horn. Such analysisis complicated by the different and overlapping rates ofdegeneration of different sizes of axon. Further, the possi-bility of secondary degeneration caused by the deafferenta-tion also has to be taken into account. The Golgi methodused by Rethelyi (1977) and Beal and Bicknell (1981) cannot distinguish between afferent axons that are unmyelin-ated and myelinated afferent axons that lose their myelinin the dorsal root entry zone. The method we employed ismore direct than the previous studies. At the LM level (Fig.1), the laminar location of VIP-ir structures can be easilyand accurately determined. These same structures viewed
390 C.M. MORGAN ET AL.

with the EM were identified as unmyelinated axons in thedorsal root ganglia, dorsal root entry zone, and Lissauer’stract and as synaptic profiles in laminae I, V, and X. Thetermination of C-fibers in these laminae does not excludethe possibility that other, larger diameter sensory fibersterminate in the same laminae. For example, other studiesshow that A-d sensory axons also terminate in these samelaminae (Light and Perl, 1979; Morgan et al., 1981, 1986;Nadelhaft et al., 1983; Thor et al., 1989).
Our conclusions regarding the termination of C-fibersapply to the sacral spinal cord of the cat but might beextended to other species at an equivalent level of thespinal cord. For example, the morphology of the sensoryinnervation of the pelvic organs in the lumbosacral spinalcord in humans, baboons, monkeys, guinea pigs, and ratsis similar in many respects to that in the cat (Anand et al.,1983; Kawatani et al., 1983a; Nadelhaft, 1983; Roppolo etal., 1983; Charnay et al., 1984; Gibson et al., 1984; deGroat et al., 1986; LaMotte and deLanerolle, 1986; Bradyet al., 1988; Chung et al., 1989). It would be reasonable toexpect that the pelvic reflexes in these species might bedependent upon similar sensory mechanisms. In addition,VIP-ir has been identified in Lissauer’s tract and the LCPin these various species. Thus the presence of A-d andC-fibers and their terminals in the same laminae as in thecat might be predicted.
On the other hand, notwithstanding some similarities,our conclusions do not apply to the entire spinal cord.Although VIP-ir has been observed at all levels of thespinal cord, it is in considerably reduced quantities com-pared to the sacral cord (Kawatani et al., 1985; LaMotteand deLanerolle, 1986). In the upper segments, smalldiameter axons, including those containing VIP, servedifferent functions from those in the sacral cord and thustheir termination in different laminae is not surprising.For example, at the cervical, thoracic, and lumbar levels inmonkeys, cats, and rats, VIP is found in laminae II and IIIas well as in laminae I, V, VII, and X (Kawatani et al.,1985; LaMotte and deLanerolle, 1986). Although most ofthis VIP is probably sensory in origin, some of it isapparently derived from a small number of propriospinalneurons (LaMotte and deLanerolle, 1986). If the VIP-containing axons at these higher levels were also unmyelin-ated, this would surely dispel the idea of exclusive laminatermination for sensory C-fibers. However, the diametersof the VIP-containing axons in the upper segments havenot yet been determined.
VIP terminals and varicosities
The sources of VIP to the sacral spinal cord are the pelvicnerve, which supplies about 42% of dorsal root ganglionneurons, and the pudendal nerve carrying 10% of theseprimary afferents. About half of those VIP afferents fromthe pelvic nerve innervate the urinary bladder, the otherssupplying the colon and sex organs (Kawatani et al., 1985;de Groat, 1986; de Groat et al., 1986, 1987). The morpho-logical variability of the VIP- immunoreactive varicositiesand terminals in the present study are consistent withthese multiple sources and probably underlie a variety ofdifferent functions.
Different vesicle types. The variety of VIP-immunore-active terminals and varicosities in the sacral cord is anindication of a heterogeneous population of sensory axons.One easily recognized example of this heterogeneity is thevariation in vesicle types and quantity in the labeled
terminals. VIP-immunolabeled varicosities were character-istically densely packed with CLVs and LDVs (Figs. 4–6),consistent with earlier reports (Honda et al., 1983; Morganand Ohara, 1984). However, it is clear that a number ofvesicles varied from very few LDVs (less than 1 per µm2) toalmost all LDVs, as high as 64 LDVs per µm2 (Figs. 5–7).This difference in LDV counts is likely to have functionalsignificance, as it indicates a variation in the amount ofVIP available at each terminal. Those terminals contain-ing only a small amount of VIP (as evidenced by containingfew LDVs), are likely to have a different function thanthose terminals with a higher concentration of VIP (i.e.,those containing many LDVs). Although CLVs were notcounted, there appeared to be a reciprocal relationshipsuch that varicosities with few LDVs had many and thosewith many LDVs had very few CLVs.
Different neurotransmitters. The presence of twotypes of vesicles (LDVs and CLVs) in VIP varicositiessuggests the presence of at least two neurotransmitters,one of which we know is VIP. Fractionation and EMmorphology studies indicate VIP is contained in the LDVs,not in the CLVs (Emson et al., 1978; Pelletier et al., 1981).Consistent with this, most LDVs in this study were darklystained with reaction product, indicating the presence ofVIP. Those few LDVs that were not stained might bedepleted of VIP or the lack of staining might indicate thatother peptides are also present (Fig. 5A, B). Others haveshown additional peptides including substance P, andLENK can be colocalized with VIP in dorsal root ganglionneurons (de Groat et al., 1986). Therefore, we speculate inthe present study that some of the varicosities may containtwo excitatory neurotransmitters, VIP and substance P,whereas others may contain VIP and the inhibitory neuro-transmitter, LENK. This multiplicity of transmitters inthe same terminal increases the functional possibilities ofthese axons (de Groat et al., 1986).
Furthermore, an additional neurotransmitter, the iden-tity of which is unknown, is very likely located in the CLVs.One possible candidate is glutamate, which has beenproposed as an excitatory neurotransmitter in sensoryaxons and is contained in CLVs (Battaglia and Rustioni,1988; Westlund et al., 1989; Broman et al., 1993). Termi-nals containing mostly CLVs (Fig. 5A, 2.44 LDV/µm2)probably serve a different function than those with mostlyLDVs (Fig. 6A, 49.19 LDV/µm2). However, regardless ofthe number of LDVs within a terminal, CLVs rather thanLDVs were always the vesicles present at the presynapticspecialization. CLVs are thought to be dependent uponcalcium channels which are in high concentration at thespecializations (Thureson-Klein and Klein, 1990). Thisassociation with the synaptic specialization suggests arapid, limited effect upon the postsynaptic membrane. Onthe other hand, LDVs are less reliant upon calciumchannels and are seldom seen near the synaptic specializa-tions. The LDVs may have their effects by fusing atnonspecialized parts of the terminal membrane and releas-ing VIP (Thureson-Klein and Klein, 1990). Thus VIP, likeother neuropeptides, may act in a paracrine manner byreleasing into the extracellular space and diffusing oversome distance and time to have a slower, more lastingeffect. In the analyses of functional neural connectivity,lack of a visible synaptic specialization may not be anaccurate indicator of lack of a functional relationship.Consequently, the actions of some of the VIP terminals
VIP IN UNMYELINATED AXONS 391

may be presynaptic and may not involve a synapticspecialization. In this regard it is interesting that 192 VIPvaricosities in this study were in close apposition to neuralstructures over 75–100% of their surface, and 111 varicosi-ties were apposed to other neural elements over theirentire surface. Often the contact was with other terminalsand varicosities of unknown origin.
In earlier reports, synaptic specializations in VIP termi-nals were observed infrequently (Pelletier et al., 1981;Honda et al., 1983; Morgan and Ohara, 1984). Theseobservations contrast with the present experiments inwhich synaptic specializations were found for 39% of VIPimmunolabeled varicosities (113 synaptic specializationson 290 varicosities). Even this figure might represent anunderestimation. Thus, in the present study, varicositiesthat were followed in serial section were often found tohave one or more synaptic specializations, each of whichwas present in only a few sections. As most of the varicosi-ties were not traced in their entirety, it is likely that somespecializations were missed.
Different synaptic specializations. We separatedthe VIP-immunolabeled terminals into three classes basedon the type of synaptic specialization they each possessed.Although our classification was based on that originallyproposed by Gray (1959), they are not entirely equivalentto those described by Gray (1959). The most frequentlyobserved synaptic contact was very similar to the Graytype I contact (Fig. 5A–C), but we also distinguished asubclass we designated as type Is (Fig. 5D). These synapticspecializations had well-developed pre- and postsynapticthickenings giving them a symmetric appearance. How-ever, the width of the thickenings was far greater thanthat found in either ours or the Gray type II symmetriccontact. As these contacts resemble the type I contact morethan the type II (Figs. 5E, 6C) but are significantlydifferent than the type I contact, we have given them thedesignation Is. Most previous studies (e.g., see Peters etal., 1976 for review) equate the Gray type I contact withexcitatory synaptic actions, in part because these special-izations are usually associated with spherical synapticvesicles. If this relationship can be extrapolated to thepresent study, it would seem reasonable to assume thatboth the type I and type Is described here indicated anexcitatory contact.
The VIP type II terminal we observed differed from thetypical Gray’s type II (Gray, 1959; Peters et al., 1976)Characteristically, Gray’s type II terminals are associatedwith boutons that contain flattened synaptic vesicles. Inthe present tissue, the VIP- immunoreactive boutons withtype II specializations differed from the typical Gray’s typeII because all the vesicles contained in the terminal wereround rather than flattened (Figs. 5E, 6A, C; Gray, 1959;Peters et al., 1976). It is possible that the presence of roundvesicles in Gray type II boutons were the result of fixationartifact. This did not seem to be the case, however, as inadjacent profiles, unlabeled by VIP, flattened and roundvesicles were present (Fig. 6D), which indicates that theabsence of flattened vesicles in the VIP-labeled terminalswas not an artifact of tissue processing.
We conclude that the classical relationship betweenround vesicles and Gray type I terminals and between flatvesicles and Gray type II terminals does not hold true forthis tissue. This includes the association of the flattenedvesicles with inhibitory synaptic actions of the Gray type IIterminal (Uchizono, 1965). Nonetheless, the VIP terminals
in the sensory pathway may involve some inhibitoryactions. It is interesting that the inhibitory neurotransmit-ter LENK is colocalized in some VIP sensory neurons (deGroat et al., 1986) and that some LENK terminals bear aresemblance to the VIP type II terminal (Morgan, 1984),suggesting that these may be inhibitory.
Relationship to parasympatheticpreganglionic neurons
Different postsynaptic contacts. Most of the struc-tures contacted by VIP-ir terminals were dendrites ordendritic spines (Table 1; Figs. 4–6). These can originatefrom a large variety of neurons in this region includingparasympathetic PGNs innervating the bladder, colon,and sex organs; interneurons serving the excretory andsexual reflexes; and spinothalamic interneurons process-ing normal and painful sensations from the pelvic organsand somatic structures of the perineal region (de Groat etal., 1978, 1979, 1981; Morgan et al., 1979, 1993; Nadelhaftet al., 1980; McMahon and Morrison, 1982; Milne et al.,1981).
Among the most interesting target neurons in thecontext of the present study are the PGNs located inlateral lamina VII of the sacral spinal cord. Electrophysi-ological and axon tracing experiments have shown that 2/3of these PGN innervate the urinary bladder (Morgan et al.,1979; Nadelhaft et al., 1980; de Groat et al., 1982). Datafrom single cell injection experiments suggest there maybe two cell types in the lateral sacral parasympatheticnucleus that innervate the bladder (Morgan et al., 1993).Each of these cell types has dendrites that extend well intothe lateral and dorsolateral funiculus where it is believedthey may be excited by axons descending from the pontinemicturition center. The normal bladder reflexes are servedby A-d sensory fibers in the LCP that do not terminatedirectly upon bladder PGN but instead contact spinobul-bar interneurons which project to the pons. Thus, thepresence of dendrites of these two bladder PGN candidatesin the LCP (in lamina V for one type and in both laminae Iand V for the other) is inconsistent with current conceptsof normal bladder reflexes. We speculate that the den-drites of these bladder PGNs in the LCP are probablycontacted by the VIP-containing C-fibers. The C-fibers inthe LCP are high threshold axons, many of which areimportant to pain perception from the bladder, but are notnormally activated by bladder distension. The inactivity ofthe C-fibers in the normal condition is most likely becausethe A-d afferents have a lower threshold and trigger thebladder reflex long before the C-fibers would be activated.However, under certain conditions, such as experimentalspinal cord transection, when A-d axons no longer producea bladder reflex, C-fiber stimulation will lead to bladdercontraction (de Groat et al., 1981). Therefore, we suggestthat C-fibers may contribute to some bladder reflexes thatemerge following spinal cord injury (Cheng et al., 1995; deGroat, 1997). These C-fibers are always present in thesacral cord but they usually remain silent and are notnormally active in the bladder reflex pathways. Conditionssuch as the painful irritation of the bladder as seen ininterstitial cystitis may also involve activation of thenormally silent C-fibers, perhaps by altering their thresh-olds so that they become abnormally responsive to disten-sion. Recent experimental and clinical application of capsa-icin or dimethyl sulfoxide, both of which act upon C-fibersand reduce bladder hyperreflexia, is consistent with this
392 C.M. MORGAN ET AL.

(Geirsson et al., 1995; Lazzeri et al., 1996; Birder et al.,1997).
CONCLUSION
This study has described the morphology of afferentaxons to the sacral spinal cord that contain VIP asdetermined by immunocytochemistry. It is clear that theVIP-immunoreactive terminals originate from C-fibersand form a heterogeneous population as determined bothby the morphology of the synaptic contact and by thevesicle content of individual terminals. This heterogeneityof terminal morphology suggests that the VIP- containingafferents might also have a variety of functions. Theunequivocal determination that VIP-immunoreactive ter-minals represent C-fiber terminations has allowed us toshow that C-fibers do terminate in laminae other thanlamina II, as had been earlier suggested. Overall, the datapresented here show the VIP innervation of the sacral cordto be more complex than previously recognized and willserve as a basis for further studies on the role of thisneurotransmitter in autonomic function.
ACKNOWLEDGMENTS
This research was supported by grants from the Na-tional Institutes of Health to Dr. Morgan including NRSAF32 NS07057 and NIDDK R01 DK49480. The authorsthank Frank Haigler and Michael E. Brown for theirexcellent technical support.
LITERATURE CITED
Anand P, Gibson SJ, McGregor GP, Blank MA, Ghatei MA, Bacarese-Hamilton AJ, Polak JM, Bloom SR. 1983. A VIP-containing systemconcentrated in the lumbosacral region of human spinal cord. Nature305:143–145.
Basbaum AI, Glazer EJ. 1983. Immunoreactive vasoactive intestinalpolypeptide is concentrated in the sacral spinal cord: a possible markerfor pelvic visceral afferent fibers. Somatosen Res 1:69–82.
Battaglia G, Rustioni A. 1988. Coexistence of glutamate and substance P indorsal root ganglion neurons of the rat and monkey. J Comp Neurol277:302–312.
Beal JA, Bicknell HR. 1981. Primary afferent distributions pattern in themarginal zone (lamina I) of adult monkey and cat lumbosacral spinalcord. J Comp Neurol 202:255–263.
Birder LA, Kanai AJ, de Groat WC. 1997. DMSO: effect on bladder afferentneurons and nitric oxide release. J Urol 158:1989–1995.
Brady A, de Groat WC, Nadelhaft I. 1988. Electron microscopic examina-tion of vasoactive intestinal polypeptide in the baboon lumbosacralspinal cord. Soc Neurosci Abstr 14:355.
Broman J, Anderson S, Ottersen OP. 1993. Enrichment of glutamate-likeimmunoreactivity in primary afferent terminals throughout the spinalcord dorsal horn. Eur J Neurosci 5:1050–1061.
Charnay Y, Chayvialle JA, Said SI, Dubois PM. 1984. Localization ofvasoactive intestinal peptide immunoreactivity in human foetus andnewborn infant spinal cord. Neuroscience 14:195–205.
Cheng CL, Ma CP, de Groat WC. 1995. Effect of capsaicin on micturitionand associated reflexes in chronic spinal rats. Brain Res 678:40–48.
Chung K, Briner RP, Carlton SM, Westlund KN. 1989. Immunohistochemi-cal localization of seven different peptides in the human spinal cord. JComp Neurol 280:158–170.
de Groat WC. 1986. Spinal cord projections and neuropeptides in visceralafferent neurons. In: Cervero F, Morrison JFB, editors. Progress inbrain research, vol 67. London: Elsevier. p 165–185.
de Groat WC. 1990. Central neural control of the lower urinary tract. CibaFoundation Symposium 151:27–56.
de Groat WC. 1997. A neurologic basis for the overactive bladder. Urology50:36–52.
de Groat WC, Booth AM, Milne RJ, Roppolo JR. 1982. Parasympatheticpreganglionic neurons in the sacral spinal cord. J Auton Nerv Syst5:23–43.
de Groat WC, Krier J. 1978. The sacral parasympathetic reflex pathwayregulating colonic motility and defaecation in the cat. J Physiol (Lond)276:481–500.
de Groat WC, Nadelhaft I, Morgan CW, Schauble T. 1978. Horseradishperoxidase tracing of visceral efferent and primary afferent pathways inthe cat’s sacral spinal cord using benzidine processing. Neurosci Lett10:103–108.
de Groat WC, Krier J, Booth AM, Morgan CW, Nadelhaft I. 1979. Neuralcontrol of the urinary bladder and large intestine. In: Brooks McC,Koizumi K, Sato A, editors. Integrative functions of the autonomicnervous system. Tokyo: Tokyo University Press. p 50–67.
de Groat WC, Nadelhaft I, Milne RJ, Booth AM, Morgan CW, Thor K. 1981.Organization of the sacral parasympathetic reflex pathways to theurinary bladder and large intestine. J Auton Nerv Syst 3:135–160.
de Groat WC, Booth AM, Milne RJ, Roppolo JR. 1982. Parasympatheticpreganglionic neurons in the sacral spinal cord. J Auton Nerv Syst5:23–43.
de Groat WC, Kawatani M, Hisamitsu T, Booth AM, Roppolo JR, Thor K,Tuttle P, Nagel J. 1986. Neural control of micturition: the role ofneuropeptides. J Auton Nerv Syst 369–387.
de Groat WC, Kawatani M, Houston M B, Rutigliano M, Erdman S. 1987.Identification of neuropeptides in afferent pathways to the pelvicviscera of the cat. In: Ciriello J, Calaresu FR, Renaud LP, Polosa C,editors. Neurology and neurobiology, vol 31. New York: Alan R. Liss. p81–90.
de Groat WC, Booth AM, Yoshimura N. 1993. Neurophysiology of micturi-tion and its modification in animal models of human disease. In: MaggiCA, editor. Nervous control of the urogenital system. Cher, Switzwer-land: Harwood Academic Publishers.
Emson PC, Fahernkrug J, Schaffalitzky de Muckadell O, Jessel TM,Iverson LL. 1978. Vasoactive intestinal polypeptide (VIP): vesicularlocalization and potassium evoked release from rat hypothalamus.Brain Res 143:174–178.
Geirsson G, Fall M, Sullivan L. 1995. Clinical and urodynamic effects ofintravesical capsaicin treatment in patients with chronic traumaticspinal detrusor hyperreflexia. J Urol 154:1825–1829.
Gibson SJ, Polak JM, Anand P, Blank MA, Morrison JFB, Bloom SR. 1984.The distribution and origin of VIP in the spinal cord of six mammalianspecies. Peptides 5:219–223.
Gobel S, Binck JM. 1977. Degenerative changes in primary trigeminalaxons and in neurons in nucleus caudalis following tooth pulp extirpa-tions in the cat. Brain Res 132:347–354.
Gobel S, Falls W. 1979. Anatomical observations of horseradish peroxidase-filled terminal primary axonal arborizations in layer II of the substan-tia gelatinosa of Rolando. Brain Res 175:335–340.
Gobel S, Hockfield S. 1979. An anatomical analysis of the synaptic circuitryof layers I, II and III of trigeminal nucleus caudalis in the cat. In:Anderson DJ, Mathews B, editors. Pain in the trigeminal region.Amsterdam: Elsevier. p 203–211.
Gray EG. 1959. Axo-somatic and axo-dendritic synapses of the cerbralcortex: an electron microscope study. J Anat (Lond) 93:420–433.
Gray EG, Guillery RW. 1966. Synaptic morphology in the normal anddegenerating nervous system. Int Rev Cytol 19:111–182.
Honda CN, Rethelyi M, Petruz P. 1983. Preferential immunohistochemicallocalization of vasoactive intestinal polypeptide (VIP) in the sacralspinal cord of the cat: light and electron microscopic observations. JNeurosci 11:2183–2196.
Kawatani M, Lowe I, Morgan CW, Nadelhaft I, Erdman S, de Groat WC.1982. Vasoactive intestinal polypeptide (VIP) in primary afferentprojections to the sacral spinal cord of the cat. Soc Neurosci Abstr 8:585.
Kawatani M, Lowe I, Moosy J, Martinez J, Nadelhaft I, Eskay R, de GroatWC. 1983a. Vasoactive intestinal polypeptide (VIP) is localized to thelumbosacral segments of the human spinal cord. Soc Neurosc Abstr9:294.
Kawatani M, Lowe I, Nadelhaft I, Morgan CW, de Groat WC. 1983b.Vasoactive intestinal polypeptide in visceral afferent pathways to thesacral spinal cord of the cat. Neurosci Lett 42:311–316.
Kawatani M, Erdman S, de Groat WC. 1985. Vasoactive intestinal polypep-tide and substance P in primary afferent pathways to the sacral spinalcord of the cat. J Comp Neurol 241:327–347.
Kawatani M, Nagel J, de Groat WC. 1986. Identification of neuropeptides inpelvic and pudendal nerve afferent pathways to the sacral spinal cord ofthe cat. J Comp Neurol 249:117–132.
VIP IN UNMYELINATED AXONS 393

Keast JR, de Groat WC. 1992. Segmental distribution and peptide contentof primary afferent neurons innervating the urogenital organs andcolon of male rats. J Comp Neurol 319:615–623.
LaMotte C. 1977. Distribution of the tract of Lissauer and the dorsal rootfibers in the primate spinal cord. J Comp Neurol 172:529–562.
LaMotte CC, deLanerolle NC. 1986. VIP terminals, axons, and neurons:distribution throughout the length of monkey and cat spinal cord. JComp Neurol 249:133–145.
Lazzeri M, Beneforti P, Benaim G, Maggi CA, Lecci A, Turini D. 1996.Intravesical capsaicin for treatment of severe bladder pain: a random-ized placebo controlled study. J Urol 156:947–952.
Light AR, Perl ER. 1979. Spinal termination of functionally identifiedprimary afferent neurons with slowly conducting myelinated fibers. JComp Neurol 186:133–150.
McMahon SB, Morrison JFB. 1982. Spinal neurones with long projectionsactivated from the abdominal viscera of the cat. J Physiol (Lond)332:1–20.
Milne RJ, Foreman RD, Giesler GJ, Willis WD. 1981. Convergence ofcutaneous and pelvic visceral nocicepetive inputs onto primate spinotha-lamic neurons. Pain 11:163–183.
Morgan CW. 1984. Ultrastructure of leucine enkephalin terminals onneurons in the sacral parasympathetic nucleus of the cat. Soc NeurosciAbstr 10:590.
Morgan CW, Ohara P. 1984. Electron microscopic identification of vasoac-tive intestinal polypeptide (VIP) in visceral primary afferent axons inthe sacral spinal cord of the cat. Anat Rec 208:121A.
Morgan CW, Nadelhaft I, de Groat WC. 1979. Localization of bladderpreganglionic neurons within the sacral parasympathetic nucleus of thecat. Neurosci Lett 14:189–194.
Morgan CW, Nadelhaft I, de Groat WC. 1981. The distribution of visceralprimary afferents from the pelvic nerve to Lissauer’s tract and thespinal gray matter and its relationship to the sacral parasympatheticnucleus. J Comp Neurol 201:415–440.
Morgan CW, de Groat WC, Nadelhaft I. 1986. The spinal distribution ofsympathetic preganglionic and visceral primary afferent neurons whichsend axons into the hypogastric nerves of the cat. J Comp Neurol243:23–40.
Morgan CW, de Groat WC, Felkins LA, Zhang S-J. 1993. Intracellularinjection of neurobiotin or horseradish peroxidase reveals separate
types of preganglionic neurons in the sacral parasympathetic nucleus ofthe cat. J Comp Neurol 331:161–182.
Nadelhaft I. 1983. The distribution of vasoactive intestinal polypeptide(VIP) in the lumbosacral spinal cord of the rat. Soc Neurosci Abstr9:293.
Nadelhaft I, de Groat WC, Morgan CW. 1980. Location and morphology ofparasympathetic preganglionic neurons in the sacral spinal cord of thecat revealed by retrograde axonal transport of horseradish peroxidase.J Comp Neurol 193:265–281.
Nadelhaft I, Roppolo J, Morgan CW, de Groat WC. 1983. Parasympatheticpreganglionic neurons and visceral primary afferents in monkey sacralspinal cord revealed following application of horseradish peroxidase topelvic nerve. J Comp Neurol 216:36–52.
Pelletier G, Leclarc R, Puviani R, Polak JM. 1981. Electron immunocyto-chemistry in vasoactive intestinal peptide (VIP) in the rat brain. BrainRes 210:356–360.
Peters A, Palay S, Webster HD. 1976. The fine structure of the nervoussystem. Phildelphia: W.B. Saunders Company.
Ralston HJ, Ralston DD. 1979. The distribution of dorsal root axons inlaminae I, II and III of the macaque spinal cord: a quantitative electronmicroscopic study. J Comp Neurol 184:643–684.
Rethelyi M. 1977. Preterminal and terminal axon arborizations in thesubstantia gelatinosa of cat’s spinal cord. J Comp Neurol 172:511–528.
Roppolo JR, Nadelhaft I, de Groat WC. 1983. The preferential distributionof vasoactive intestinal polypeptide VIP) in the sacral spinal cord of therhesus monkey. Soc Neurosci Abstr 9:293.
Thor KB, Morgan CW, Nadelhaft I, Houston MB, de Groat WC. 1989.Organization of afferent and efferent pathways in the pudendal nerve ofthe female cat. J Comp Neurol 288:263–279.
Thureson-Klein AK, Klein RL. 1990. Exocytosis from neuronal largedense-cored vesicles. Internat Rev Cytol 121:67–126.
Uchizono K. 1965. Characteristics of excitatory and inhibitory synapses inthe central nervous system of the cat. Nature 207:642–643.
Vizzard MA, Erdman S, Erikson VL, Stewart RJ, Roppolo JR, de Groat WC.1994. Localization of NADPH diaphorase in the lumbosacral spinal cordand dorsal root ganglia of the cat. J Comp Neurol 339:62–75.
Westlund KN, McNeill DL, Coggeshall RE. 1989. Glutamate immunoreac-tivity in rat dorsal root axons. Neurosci Lett 96:13–17.
394 C.M. MORGAN ET AL.



![Terlipressin versus other vasoactive drugs for hepatorenal ...[Intervention Review] Terlipressin versus other vasoactive drugs for hepatorenal syndrome Mads Israelsen 1, Aleksander](https://img.pdfslide.us/doc/110x75/60a7d279a5295c2888291ae4/terlipressin-versus-other-vasoactive-drugs-for-hepatorenal-intervention-review.jpg)

























