Embed Size (px)
Citation preview

~ E R S P E C T I V E S
Qua l i t a t i ve and quantitative control of tissue-specific gene expression is determined by 'epigenetic' modifi- cations of the DNA. Such modifications might operate at the level of the DNA bases (e.g. in mammals by methylation of cytosine), DNA configuration and higher order structure of chromosomes, timing of DNA repli- cation in the cell cycle and nuclear organization. Activation of transcription is itself a form of modifi- cation, in that it will alter the conformation of the DNA segment concerned and affect its subsequent modifi- cation (e.g. by methylation). In addition, the products of transcribed genes will exert effects, in tram, on the function of other genes whose regulatory circuits or functions are connected in some way.
In differentiated somatic tissues, epigenetic modifi- cations must be faithfully inherited to ensure stability of function. Indeed, it has been shown that methyl- ation of DNA is a stable, heritable modification propagated throughout DNA replication and cell divi- sion1, z. However, development and cell differentiation are characterized by changing patterns of epigenetic information. In the case of methylation modification, the changes may be finely tuned (such as those correlated with the onset of specific gene transcription in differentiating cells3), or they may be observed as global and more generalized (as in early develop- ment4,5).
The variation in methylation that occurs in developing systems must be time and tissue specific but the details are not yet clear. Recently, the picture appears to have become even more complex. Differences in methylation are observed as a con- sequence of inheritance via the male or female gamete (imprinting 6) or as a consequence of different parental mouse strains 7. Here, I review different examples of variability in methylation and present an hypothesis of
Variation in epigenetic inheritance MARILYN MONK
Changing patterns of ONa metbylation may malerlle dtfferential gene expression in development Additional sources of variation in allelic metbylation may be introduced by parental differences as well as by gamete of ~gs=
allele-to-allele communication as a basis for a wide range of phenomena such as allelic exclusion, random and nonrandom X-chromosome inactivation, imprint- ing, strain-dependent methylation mosaicism of trans- genes, nonrandom gene expression and develop- mental abnormalities in interspecific crosses, and tumour susceptibility. The 'cross-talk' hypothesis suggests that methylation differences may occur as a consequence of differences perceived by a mechanism that allows communication between homologous regions of chromosomes. This process would serve to compare each chromosome with its homologue, to detect differences or irregularities between them, and choose whether to respond to these differences by modifying one or both homologous regions.
Variation in DNA methylation Tissue mosaicism in methylaton has been known
for some time. To account for partial methylation pat- terns, it was assumed that a tissue DNA represented a number of different cell types. Variability in methyl- ation is also known to occur in cultured cells, where methylation appears to undergo random drifts. Others have reported a loss of methylation in aging cells9 and
/ ¢ ,..,,,,
Transgene
Transgene
Transgene
Transgene
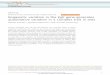
germ-line modified (leading to methylation in soma of progeny)
germ-line modified (leading to undermethylation in soma of progeny) 1 methylated in soma
E] undermethylated in soma
FIGE] hnprinting - gamete-dependent modification of transgene.
©1990 Elsevier Scie- 'ce Publ ishers Ltd (UK) 01(x8 - 9479/90/$02.00
"FIG APRIL 1990 MOL. 6 NO. 4

'~ERSPECTIVES
F;GB Strain-dependent modi- fication of transgene. In this example, when the transgenic male is mated to the female strain A, the transgene is undermethylated in the progeny (D). When he is mated to female strain B, the transgene shows intermediate, or partial, methylation in the progeny (rl).
Poren~
strain
Progeny
d 9 d
in tumours lo. Losses in methylation in cultured cells may remove the differences in methylation of CpG sequences on the active and inactive X chromosomes, normally observed in fresh tissues II. It is not known whether such random changes in methylation occur in vivo, in somatic or germ cells of the individual. If so, they could be associated with gain or loss of func- tion and this change would be heritable (a heritable abnormality in gene expression that is not the result of a change in DNA base sequence has been termed an 'epimutation' by Holliday12).
Reproducible differences in methylation of particu- lar gene sequences have recently been observed in fresh tissues. This variability can be of several types: variation between alleles on different chromosome#3a4, variation between cells within an individual (methyl- ation mosaicism) 7, variation between individuals de- pending on genetic background 15, and variation be- tween one generation and the next, that is, through the germ line ~6.
Allele-specific variation in methylation One reason why variation in methylation inheri-
tance has come to the fore is the discovery of differen- tial methylation of transgenes inherited either from the father or from the motherlT-20. This was possible to observe because of the hemizygosity of the transgene allele.
:~.t zro-nd the same time, Chandler et al. 14 report- ed allele-specific metifflztion of an endogenous gene, c-Ha-ras-1, in human fibrGbiast cell lines (the differences were not seen in fresh tissues, which were highly methylated). Each copy of the gene in a diploid cell could be looked at because the cell was heterozygous for linked arrays of tandem repeat sequences which served as markers. This meant that the HpalI fragments from each allele could be identi- fied on the basis of size on the gels. In a further study, Silva and White~3 identified heritable, allele- specific methylation patterns in blood cell DNA for each of ten human loci, distributed over six chromo- somes. Again, each allele at each locus was identified by a variable tandem repeat sequence. The methyl- ation pattern (whether partially or fully methylated, or unmethylated) for all alleles tested was found to be heritable in family studies through three gener- ations. The allele-specific methylation patterns were
reproduced after transmission through the germ line (male or female) and showed no differential parental imprinting. However, in sperm at least (oocytes could not be studied in this way), all alleles showed the same invariant methylation pattern (usually fully methylated, in a few cases unmethylated, never par- tially methylated). This me.~ns that information governing the later allele-specific methylation pat- terns in blood DNA was carried through the germ cells by some means other than methylation itself, either as some other modification or as sequence variation due to variable repeat number. As well as the heritable differences in methylation between 'homologous' alleles within an heterozygous in- dividual, three of the ten alleles showed different methylation patterns among different individuals.
Thus, we can assume that homologous alleles within the genome may commonly be differently methylated and that such differences may be heritable. This raises the question of whether endogenous gene expression may vary according to the specific methylation pattern of a particular allele in heritable fashion. Do individu- als carry genes silenced by epimutation and do they transmit such epimutations through the germ linei2? Could the 'unmasking' of a previously silent epi- mutation lead to the sudden unexpected appearance of a new phenotype?
Whatever the case, it is clear that the partial methylation patterns generally seen within DNA ex- tracted from a tissue could arise from allele variability. In the case of a hemizygous locus, allele variability will give rise to cellular mosaicism.
Genetic background effects and cell-specific methylation mosaicism
Hemizygous transgene loci in mice present the opportunity to investigate parent-specific methylation patterns (in~tpdnting), genetic background effects on transgene methylation and cellular mosaicism in methylation. (The hypothesis outlined below suggests that these seemingly different phenomena may all arise from a common mechanism.) Six out of 21 trans- genes studied have shown imprinting, with the trans- gene inherited from the egg being more methyl- ated !7-z0. A simplified view of the transmission of methylation imprinting via the male or the female gamete is shown in Fig. 1. In several cases, the
Tm APRIL 1990 VOL. 6 NO. 4
1 1~1

~IBERSPECTIVES
difference in methylation, dependent on the parent of origin of the transgene, is not seen in the extra-embry- onic lineages of the mouse conceptus, suggesting that the methylation imprint may be established after these lineagt:a have been set aside.
Superimposed on the differential methylation seen for imprinted transgenes (dependent on whether the transgene was inherited via the egg or the sperm) is a further trans effect on the degree of methylation (increase or decrease) dependent on the mouse strain of the other parenfl 5. An illustration of strain- dependent variation in methylation is shown in Fig. 2. The interstrain effect on transgene methylation has been observed for a number of different transgenes and a number of different strains 7,i5,16. It may occur also for transgenes that do not show parental origin (imprinting) effects. The interstrain differences in methylation must be established after fertilization, perhaps at the time of de novo methylation. The strain- dependent trans effect, observed initially with the quail troponin I transgene, operates from a single locus - a 1:1 segregation of transgene methylation phenotypes is produced by F l female animals 15.
The basis of the intermediate methylation patterns seen as partial HpaII sensitivity of a transgene-associated CpG site is the subject of a recent paper by McGowan et al.Z. In this paper, the authors report .hat a lacZ-containing transgene, transmitted by the male, may show hypomethylation, hypermethylation or partial methylation in tissue DNA of the offspring, depending on the strain of the female used in the cross. Sit filar observations have been reported by Surani et al. 16.
Why do sites in the trar~sgene show partial methyl- ation in the progeny of certain interstrain crosses? In these progeny, the same site in the transgene may be methylated or unmethylated. Since this variability is observed in mid-gestation fetuses, it is unlikely to be due to random drift, as seen in cell culture. One possi- bility is that the partial methylation patterns, seen for the quail troponin I and the lacZ-containing trans- genes in the progeny of the different parent inbred strains, are due to variable methylation among the multiple copies of the transgene inserted in tandem. To approach this question, McGowan et al. 7 correlated methylation of the lacZ-containing transgene with an in situ histological examination of the number of lacZ- expressing cells in the neural tube in 12 day old fetuses. Individuals with an intermediate (or partial) methyl- ation of the transgene in tissue DNA showed a mosaic expression phenotype in situ; that is, a variable number of expressing cells (cellular mosaicism). Furthermore, the proportion of expressing cells was inversely correlated with the degree of methylation of the transgene. The authors did not see identical inter- mediate levels of expression in all the cells, as might be expected if the partial methylation patterns were due to variable methylation of the different copies in the tandem array of transgenes within the cells. McGowan et al. 7 conclude that a cell-to-cell variation in methylation and expression (mosaicism) exists between apparently identical cells in the developing fetus (this conclusion would not necessarily be correct if only one copy of the tandem repeats was capable of
The cross.talk hypothesis A unifying hypothesis that incorporates the diverse
phenomena so far reviewed is depicted in F!g. 3. It is suggested that homologous regions of the genome communicate and compare the epigenetic modifi- cations that recall their past history. The 'cross-talk' enables homologues to compare their degree of relatedness (e.g. species or strain) or similarity (e.g. gamete of origin). Irregularities observed between homologous alleles may result in 'repair' (e.g. mitotic recombination, nondisjunction, conversion) or differential modification of one or the other allele, and, possibly, differential expression of :he regions con- cerned. Genes involved in the action taken would qualify as 'modifier genes'. The outcome may be absolute (preferential maternal or paternal allele expression) or it may result in cellular mosaicism, with different proportions of cells expressing either allele.
The 'cross-talk' hypothesis implies that regions of each pair of homologous chromosomes are in communication. Random X-inactivation in females would be a special case where an irregularity would be 'set up' in the course of development, perhaps by an event that occurs first on one or the other X chro- mosome, and then is rapidly followed by the silencing of or, e of die two chromosome#l.aZ. (A similar mechanism may operate in allelic exclusion in antibody-producing cells of the immune system.) The strain-dependent variation in methvlation and expression described here shows many similarities to
I
expressing lacZ). The mosaicism results from the parental strain difference and hence is genetically controlled.
It has not been established whether all or most of the copies of lacZ are methylated in some cells and undermethylated in others. However, a cell-to-cell difference in expression may not require such an- abso lu ted i f f e rence in methylation; one could imagine that methylation of a DNA segment may be continuously variable, whereas expression of the transgene array may be an on/off step function with a switch probability affected by the overall degree of methylation. This would mean that methylation dif- ferences could be very common (though difficult to observe for endogenous homologous genes which cannot be looked at separately) whereas the effects of these differences on gene expression would be rare.
What could be the basis of cellular variation in methylation? The authors suggest that, early in devel- opment, an event occurs such that differences in methylation occur at the transgene locus in different cells. The event would be mediated by 'modifier genes' wl;ich il: ~,ome way interpret the genetic back- ground environment of the locus and modify it accord- ingly. The methylation mosaicism, once established, would be stable and give rise to similar mosaic pro- portions in all tissues of that individual. (An alternative to this 'one event' hypothesis is that an allele's history and current situation may reside in some modification other than methylation and be subject to continuous assessment.)
TIG APRIL 1990 VOL. 6 NO. 4

[ ~ E R S P E C T I V E S
X-chromosome inactivation. Ran- dom X-chromosome inactivation in somatic tissues results in cellular X- inactivation mosaicism with differ- ent proportions of cells with one or the other X chromosome active in different individualsZ3. As for trans- genes, there are also strain effects on the randomness of X inacti- vation (the Xce effect24), and the two X chromosomes show allele- specific variation in methylation (reviewed in Ref. 25). In the extra- embryonic tissues of rodents, the outcome of the cross-talk between the two X chromosomes is deter- mined by the gamete of origin. However, unlike transgenes, X- chromosome inactivation spreads to inactivate the whole X chromo- some. Very little can be said about a possible mechanism of cross-talk between chromosomes and the action exerted in trans. Indeed, little is known of the communi- cation between the two X chromo- somes despite many years of in- vestigation.
Homologous alleles on the autosomes may differ in a number of ways: by retaining a 'memory' of their gamete of origin or even more distant ancestors, or through parental strain differences (perhaps with different chromatin organi- zation in the affected region) and local differences that cause mod- ification to nearby sequences (position-effect variegation; e.g. rearrangements, transgene insertions or variable numbers of tandem repeats). As well as the modifying effects on DNA sequences, alleles will differ in DNA base sequence itself (apart from inbred individuals). Sequence variation is present in interstrain and random crosses and even more so in interspecific crosses, where phenomena reminiscent of imprinting occu# 6, that is, preferen- tial parental allele expression and nonreciprocal developmental abnor- malities and lethality. The mule and the hinny clearly demonstrate the differential roles of paternal and maternal genomes in the interspecific cross between horse and donkey. The greater the genetic distance in interspecific crosses, the greater the effects27. 28.
cross-talk hypothesis Q) Modification/activation oF both alleles
b ) Random modification/activation of" either allele
¢ ) Preferential modification/activation of one allele
d) Preferential modification/activolion of one allele via
I (i ~ "modifier"gene
2.~ ~~'"! other alleles?
FIG[]
The cross-talk hypothesis. The figure shows various modes of activation of homologous alleles (-~-) via their controlling sites (-o-) on a pair of chromosomes. An event occurs in development that normally modifies/activates homologous alleles on both chromosomes (a). This event may occur in such a way that one homologous allele is activated before the other (b, c, d). The difference in timing could arise because the first event is slow or inefficient (b) e.g. de nova methylation2], 22. Alternatively, an irregularity or difference between the loci (c, d) could be the basis of the difference in timing. Modification of the second allele may occur ff prior activity of the first allele results in suppression of the second allele, directly (b, c), or if activation of the first allele leads to suppression of the second allele indirectly via a 'modifier' gene product (d). The first modification event may be transcriptional activation itself; the second modification event could be a consequence of the continuance of the developmental programme.
Germ-line variation in methylation Can the epigenetic modifications that generate the
differences in methylation in development be transmit- ted through the germ line? Clearly, the answer is yes. In the classical imprinting situation, a transgene may
switch from one methylation state to the other in the progeny by transmission through the male or female germ line. Expression of a transgene may also be changed; the c-myc transgene is only expressed after transmission through the male 20.
Irrespective of the methylation status determined in the offspring, endogenous alleles or imprin:ed transgenes tend to show an invarianr methylation pattern in sperm. ,'l,us, the epigenetic modification that determines methylation patterns later in develop-
Tm APRU. 1990 VOL. 6 NO. 4
117

~'~ERSPECTIVES
ment (perhaps at the time of de novo methylation), is carried in sorae form other than methylation itself. The memory which recalls the history of a DNA sequence may shift from one form of modification to another -'6.
The gamete-transmitted memory that determines methylation patterns in development - whether parent specific (imprinted), allele specific, or recipient strain specific - is generally reproducible, but there are exceptions. In one case, the imprint was not reversible following transmission through the female germ line, after which the transgene became irreversibly methyl- ated and silenced TM. A change in epigenetic inheritance had occurred (a germ-line epimutation). Another note- worthy case of germ-line variation in inheritance of methylation has been reported recently. Surani et al) 6 have shown increased or decreased methylation associ- ated with low and high lacZ expression transmitted through the germ line during repeated backcrosses to one or other inbred line. But now these changes are associated with a change in the methylation status of sperm. These results show that progressive changes in methylation and expression of genes can occur from generation to generation.
Conclusions Methylation variation in somatic tissue may be very
common, though perhaps only rarely affecting gene expression. Many methylation differences may be inconsequential with respect to gene expression; others might occur in mortal somatic tissues that will be eliminated from the body. Other changes, albeit less common, may be critical and block development, or occur in stem cell populations or in the germ line. These will have important consequences. The exper- imental systems discussed, as well as our existing knowledge of X-chromosome inactivation, show that m~,chanism~ exist that make such important changes possible for endogenous genes.
For most of us, brought up on the secure con- cepts and 'laws' of Darwin and Mendel, a new con- ceptual framework based on allele-to-allele, cell-to- cell, and generation-to-generation variation and change in the processes responsible for specific pat- terns of gene expression is difficult to formulate in our minds. However, it is often the exception to the rule that opens the door to a new understanding. A significant question arises in connection with the pos- sibility of variation in epigenetic inheritance through the germ line (see Refs 12, 29). Could the potential for expression of the genes we pass on to our off- spring change? An opportunity exists for heritable epigenetic variation before sequestration of the germ line in development, and then in the 10--20 cell divisions that generate the female gametes, and the many more cell divisions that generate the male gametes (variation via the male might be more common). If such changes happen, are they always random? Could some variability be directed by hor- monal and environmental factors, such as individual and environmental stress? How many silent genes in the form of epimutations might we carry? The possi- bility that inheritance may be affected by the way we
live our lives individually, as well as collectively in the environment we create for ourselves, is food for thought.
Acknowledgements I thank Bruce Ponder and Azim Surani for helpful com-
ments on the manuscript and A. Lee for illustrations.
References 1 Wiglet, M., Levy, D. and Perucho, M. (1981) Cell24,
33-40 2 Stein, R. etal. (1982) Proc. NatlAcad. Sci. USA79,
61-65 3 Jost, J.P. et al. (1990) Phil. Trans. R. Soc. London Ser. B
326, 231-240 4 Monk, M., Boubelik, M. and Lehnert, S. (1987)
Development 99, 371-382 5 Monk, M. (1990) Phil. Tram. R. Soc. London Ser. B 326,
299-312 6 Surani, M.A., Reik, W. and Alien, N. (1988) Trends
Genet. 4, 59-62 7 McGowan, R., Campbell, R., Peterson, A. and Sapienza,
C. (1989) Genes Dev. 3, 1669-1676 8 Reis, R.J.S. and Goldstein, S. (1982) Proc. NatlAcad. Sci.
USA 79, 3949-3953 9 Wilson, V.L. and Jones, P.A. (1983) Science 220,
1055-1057 10 Feinberg, A.P. and Vogelstein, B. (1983) Nature301,
89-91 11 Lindsay, S. et al. (1985)Ann. Hum. Genet. 49,
115-127 12 Holliday, R. (1987) Science 238, 163-170 13 Silva, A.J. and White, R. (1988) C01154, 145-152 14 Chandler, L.A. etal. (1987) Cell50, 711-717 15 Sapienza, C., Paquette, J., Tran, T.H. and Peterson, A.
(1989) Development 107, 165-168 16 Surani, M.A. et al. (1990) Phil. Trans. R. Soc. London Ser.
B 326, 313-327 17 Reik, W. et al. (1987) Nature328, 248-251 18 Hadchouel, M. et a/. (1987) Nature 329, 454-456 19 Sapienza, C., Peterson, A., Rossant, J. and Bailing, R.
(1987) Nature 328, 251-254 20 Swain, J.L., Stewart, T.A. and Leder, P. (1987) Cell 50,
719-727 21 Riggs, A. (1975) Cytogenet. Cell Genet. 14, 9-25 22 Holliday, R. and Pugh, J.E. (1975) Science 187,
226-232 23 McMahon, A., Fosten, M. and Monk, M. (1983) I.
Embryo/. Exp. Morphol. 74, 207-2.?0 24 Cattanach, B.M. and Williams, C.E. (1972) Genet. ICes. 19,
229-240 25 Monk, M. (1986) BioEssal,s 4, 204-208 26 Monk, M. (1988) Genes Dev. 2, 921-925 27 Whitt, G.S., Philipp, D.P. and Childers, W.F. (1977)
Differentiation 9, 97-109 28 Ohno, S. (1969) in Heteroapec~'c Genome
Interact.ion ( Wistar Institute Monograph No. 9), (Defendi, V., ed.), pp. 137-150, Wistar Institute Press
29 Jablonka, E. and Lamb, M. (1989)/. Theor. Biol. 139, 69-83
M. MONK IS IN THE MRC MAMMALIAN DEVELOPMENT UNITp I [4 STEPHENSON WAY, LONDON NWI 2HE, UK. ]
TnC, At'mL 1990 VOL. 6 NO. 4
11~i