Embed Size (px)
Citation preview

Variation among Desulfovibrio Species in Electron Transfer SystemsUsed for Syntrophic Growth
Birte Meyer,a Jennifer Kuehl,b Adam M. Deutschbauer,b Morgan N. Price,b Adam P. Arkin,b David A. Stahla
Department of Civil and Environmental Engineering, University of Washington, Seattle, Washington, USAa; Physical Biosciences Division, Lawrence Berkeley NationalLaboratory, Berkeley, California, USAb
Mineralization of organic matter in anoxic environments relies on the cooperative activities of hydrogen producers and consum-ers linked by interspecies electron transfer in syntrophic consortia that may include sulfate-reducing species (e.g., Desulfovi-brio). Physiological differences and various gene repertoires implicated in syntrophic metabolism among Desulfovibrio speciessuggest considerable variation in the biochemical basis of syntrophy. In this study, comparative transcriptional and mutantanalyses of Desulfovibrio alaskensis strain G20 and Desulfovibrio vulgaris strain Hildenborough growing syntrophically withMethanococcus maripaludis on lactate were used to develop new and revised models for their alternative electron transfer andenergy conservation systems. Lactate oxidation by strain G20 generates a reduced thiol-disulfide redox pair(s) and ferredoxinthat are energetically coupled to H�/CO2 reduction by periplasmic formate dehydrogenase and hydrogenase via a flavin-basedreverse electron bifurcation process (electron confurcation) and a menaquinone (MQ) redox loop-mediated reverse electronflow involving the membrane-bound Qmo and Qrc complexes. In contrast, strain Hildenborough uses a larger number of cyto-plasmic and periplasmic proteins linked in three intertwining pathways to couple H� reduction to lactate oxidation. The fastergrowth of strain G20 in coculture is associated with a kinetic advantage conferred by the Qmo-MQ-Qrc loop as an electron trans-fer system that permits higher lactate oxidation rates under elevated hydrogen levels (thereby enhancing methanogenic growth)and use of formate as the main electron-exchange mediator (>70% electron flux), as opposed to the primarily hydrogen-basedexchange by strain Hildenborough. This study further demonstrates the absence of a conserved gene core in Desulfovibrio thatwould determine the ability for a syntrophic lifestyle.
In anoxic environments depleted in inorganic electron acceptors(e.g., freshwater and marine sediments, flooded soils, landfills,
and sewage digesters), the complete mineralization of complexorganic matter to CO2 and methane relies on the cooperative ac-tivities of phylogenetically and metabolically distinct microbialgroups assembled in syntrophic consortia. In these assemblages,sulfate-reducing bacteria (SRB) function as secondary fermentersobligately linked via interspecies electron transfer to the metabolicactivity of methanogenic archaea since the oxidation of commonsubstrates (organic acids and alcohols) yields sufficient energyonly for cell maintenance and growth when the methanogensmaintain low concentrations of the products of SRB metabolism(acetate, hydrogen, and formate) (1–3). As a result, the metabo-lism of one community member is directly influenced by and de-pendent upon the activity of the other. Hydrogen and formate areconsidered the primary shuttle compounds for interspecies elec-tron transfer (1–3). Additionally, single reports suggest the in-volvement of cysteine or exogenous carriers such as humic sub-stances (2) as well as the existence of a direct electron transfer viaelectrically conductive pili called nanowires (4). Exclusive use ofhydrogen as an electron carrier has been proven for several syn-trophs, while a simultaneous formate/H2 electron shuttle was pro-posed to be operative in propionate- and butyrate-degrading co-cultures on the basis of early physiological experiments (2, 5–9)and later confirmed by biochemical and genetic studies (10, 11).Indeed, the midpoint redox potentials (E°=) of the redox couplesH2-H� and formate-CO2 are similar (�414 mV and �432 mV,respectively) (12), but hydrogen and formate have different chem-ical and physical properties. As metabolite exchange occurs viadiffusion, the intermicrobial distances have a significant influenceon the efficiency of interspecies transfer (especially for H2) and,
therefore, on biodegradation and specific growth rates (2, 7, 13,14). To ensure close proximity, cell aggregates of high complexityare often formed (e.g., sludge granules in anaerobic reactors), withflagellar proteins having a putative role in triggering the initialsyntrophic interactions (15). Nevertheless, syntrophic communi-ties exist under conditions close to thermodynamic equilibrium,and the need for reverse electron transport as a distinctive featureof syntrophic metabolism implies that biochemical mechanismsmust have been evolved to share the minimum amount of energythat is available from the conversion of the substrate (1–3, 5, 6).
SRB that are capable of switching between the sulfidogenic andthe syntrophic lifestyle (e.g., Desulfovibrio and Syntrophobacterspecies) are closely related to both sulfate reducers and obligatesyntrophs (e.g., Pelotomaculum schinkii) within the phylogeneticlineages of the Deltaproteobacteria and Firmicutes (1, 16). So far,most biochemical and genetic studies have focused on the inves-tigation of the electron transfer and energy conservation mecha-nisms in propionate and butyrate oxidizers, e.g., Syntrophobacterand Syntrophomonas (3, 5, 6, 10, 11), while there is only limitedunderstanding of the syntrophic metabolism of other representa-tive bacterial species, e.g., Desulfovibrio (17–19).
Received 5 October 2012 Accepted 4 December 2012
Published ahead of print 21 December 2012
Address correspondence to David A. Stahl, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.01959-12.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JB.01959-12
990 jb.asm.org Journal of Bacteriology p. 990–1004 March 2013 Volume 195 Number 5
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

A recent comparative analysis of Desulfovibrio vulgaris strainHildenborough grown on lactate under respiratory conditions(sulfate-limited monoculture) and syntrophic conditions (cocul-ture with Methanococcus maripaludis strain S2) concluded thatstrain Hildenborough uses largely independent energy generationpathways for syntrophic growth and sulfate respiration (17). No-tably, deletion of genes that demonstrated higher transcriptionwhen growing syntrophically (including a putative cytoplasmichydrogenase [Coo] and a membrane-bound electron-transferringcomplex [Hmc]) had little impact on the capacity for respiratorygrowth but severely impaired the capacity for syntrophy. Addi-tional physiological data and metabolic flux balance modeling in-dicated that hydrogen is the sole carrier for interspecies electrontransfer in this coculture (17, 20). However, in preliminary batchcoculture experiments, we clearly showed that different Desulfo-vibrio species vary significantly in their growth characteristics(e.g., growth rate, biomass yield, and population dynamics)when syntrophically coupled to the same hydrogenotrophicmethanogen (B. Meyer, unpublished data). Thus, physiologicaldifferences, such as the ability to ferment lactate at elevated hydro-gen concentrations, and different repertoires of genes implicatedin syntrophy (hydrogenases, formate dehydrogenases, and elec-tron transfer proteins/complexes [21, 22]) suggest considerablevariation in the biochemical basis of syntrophy.
To investigate the metabolic and genetic basis for the observedphenotypic differences in syntrophic growth of Desulfovibrio spe-cies in greater detail, we grew Desulfovibrio alaskensis strain G20 inchemostats under two culture conditions at identical dilution rate:coculture with M. maripaludis and sulfate-limited monoculture.Comparative whole-genome transcriptional analyses, comple-mented by mutant growth experiments and physiological data,established distinctive differences between the electron transfersystems and energy conservation mechanisms sustaining syn-trophic growth of Desulfovibrio species and demonstrated the useof formate by D. alaskensis strain G20 as the primary mediator ofelectron exchange, in contrast to the hydrogen exchange-basedsystem of D. vulgaris strain Hildenborough. The results of thisstudy further reveal the absence of a core of conserved genes inDesulfovibrio species that would both define and determine theability for a syntrophic lifestyle in SRB.
MATERIALS AND METHODSMicroorganisms. The D. alaskensis G20 and M. maripaludis S2 wild-typestrains used for establishing the initial cocultures were obtained from theATCC. A total of 23 D. alaskensis strain G20 mutant strains were used inphenotypic growth comparisons (Table 1; for a detailed list, see Table S1in the supplemental material).
Mono- and coculture medium composition for continuous andbatch cultures. The basal salt solution of the media was described in aprevious paper (17) and contained 30 mM sodium DL-lactate or sodiumpyruvate, resazurin as a redox indicator, and 10 mM Na2SO4 (chemostatmonoculture medium) or 30 mM Na2SO4 (batch monoculture medium).The media were prepared in bioreactor glass vessels (2 liters), mediumreservoir bottles (10 liters), and Widdel flasks (1 and 2 liters) anoxicallyunder an N2-CO2 (80:20, vol/vol) gas atmosphere as described by Widdeland Bak (16). The media were buffered using 1.1 mM KH2PO4 and 30mM NaHCO3. Nonchelated trace element mixture 1 and selenite-tung-state solution, as well as vitamin mixture, riboflavin, vitamin B1 and B12
solutions, and 1 M Na2S solution (16), were added aseptically in 1-mlaliquots per liter of medium. All stock solutions were prepared underanoxic conditions as according to Widdel and Bak (16). For batch culturecultivation, the media were dispensed anoxically in sterile N2-CO2 (80:20,
vol/vol)-pregassed 250-ml serum bottles or 25 ml Balch tubes in 130- and13-ml aliquots, respectively. After preparation, the bioreactor and me-dium reservoir bottles were connected to an N2-CO2 (90:10, vol/vol) gasmixture tank.
Chemostat operation conditions. A 0.5-ml inoculum from a glycerolstock of a previously grown coculture (10th transfer generation) or mon-oculture was used to inoculate 130 ml of medium into 250-ml serumbottles. Cultures were incubated at 37°C in a shaking incubator (300 rpm)until reaching an optical density at 600 nm (OD600) of 0.3 (� 0.02), and a100-ml inoculum was then added to a 2.45-liter bioreactor vessel (Fair-MenTec, Göttingen, Germany; Electrolab, Tewkesbury, United King-dom) containing 2 liters of culture medium. Three biological replicates ofcoculture and sulfate-limited D. alaskensis strain G20 monoculture weregrown in chemostats. Following inoculation, every chemostat was initiallyrun in batch mode at 37°C with stirring at 250 rpm. The headspace of thebioreactor was flushed through a sterile 0.2-�m-pore-size syringe filterwith N2-CO2 (90:10, vol/vol) at a constant rate of 1 ml/min using a massand volumetric precision gas flow controller (MC-20 series; Alicat Scien-tific, Tucson, AZ). The pH was maintained at 7.0 � 0.1 by using bicar-bonate buffer and small additions of 0.1 M NaOH or HCl, as needed.Headspace levels of the gases N2, O2, CH4, H2, and CO were continuouslymeasured by using an SRI 8610SC gas chromatograph with thermal con-ductivity and reducing gas detectors (carrier gas, 4.7 laser-grade helium at20 ml/min). Organic acid (lactate, acetate, and formate) concentrationswere determined using a Dionex DX500 ionic chromatograph modelLC20-1 (analytical column, IonPac AS11-HC; eluent, 0.75 to 40 mMNaOH; conductivity detector, CD20), whereas ethanol and glycerol weremeasured enzymatically as described in reference 20. The chemostatmode was initiated after the OD600 reached 0.29 � 0.01 and maintained ata dilution rate of 0.047 h�1. Coculture samples were taken regularly fordirect cell counting (Petroff-Hauser counting chamber) and protein mea-surement (determined with the Coomassie Plus assay; Pierce, Rockford,IL). Cells were harvested after three retention periods with an OD600 vari-ance of less than 10% by using an ice-chilled sterile stainless steel tubeconnected to the chemostat medium exhaust line that allowed a cell sus-pension transfer with minimal O2 contact into pregassed, ice-chilled Fal-con tubes (50 ml). After centrifugation for 15 min at 3,220 � g and 4°C,the supernatant was carefully poured off and the cell pellets were imme-diately frozen at �80°C.
For the qualitative and quantitative investigation of the interspecieselectron mediators in coculture, the methanogen was specifically inhib-ited by manually adding 5 mM bromoethanosulfonate (BES) to the che-mostat and medium reservoir bottle after three volume turnovers at con-stant OD600 values. The optical density, organic acid, and gas levels weremonitored over short time intervals for 24 h following BES addition, asdescribed above. On the basis of the data for the first 3 h after BES addi-tion, the electron-flow ratio through each electron carrier during syn-trophic lactate conversion was estimated according to the following equa-tion (simplified): percent electron flow through each electron carrier(HCOO�/H2/CO) from lactate oxidation � [�nelectron of electron carrierformed/(nelectron of lactate inflow � nelectron of lactate outflow ��nelectron of methane formed)] � 100, where nelectron is the number ofelectrons in mole and �nelectron is the difference of electrons in mole. Onlyelectrons available from conversion of lactate to acetate (four electronsper lactate molecule) were taken into consideration. The parameters ofthe chemostat runs were 30 mM lactate in media, 0.047 h�1 dilution rate,and 1 ml/min gas flow rate.
Whole-genome transcriptional analysis. Microarray gene expressionmeasurements were performed on three biological replicates of cocultureand monoculture chemostats (the effect of time for harvesting on tran-scription results was examined for two reactor studies by two cell collec-tions separated by 15 min at the end of the chemostat run). Total RNA wasextracted from frozen cell paste (stored at �80°C) using an RNeasy kitaccording to the manufacturer’s protocol (the lysozyme cell wall digestionstep was omitted). Total RNA concentration and quality were determined
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 991
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
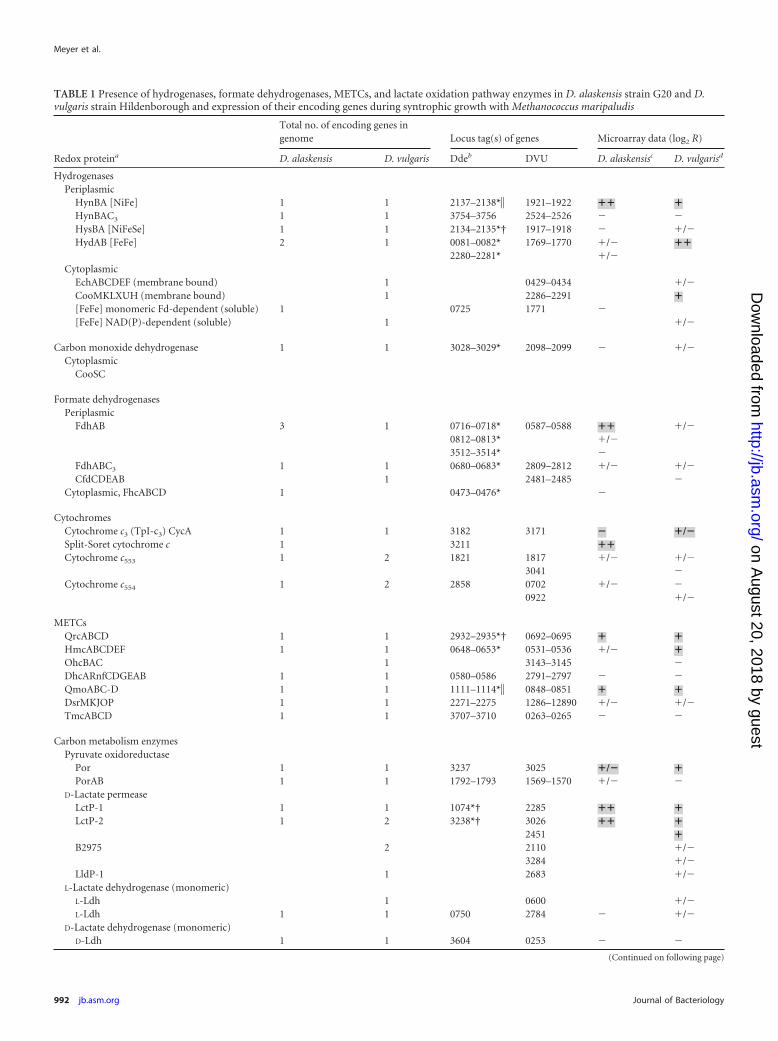
TABLE 1 Presence of hydrogenases, formate dehydrogenases, METCs, and lactate oxidation pathway enzymes in D. alaskensis strain G20 and D.vulgaris strain Hildenborough and expression of their encoding genes during syntrophic growth with Methanococcus maripaludis
Redox proteina
Total no. of encoding genes ingenome Locus tag(s) of genes Microarray data (log2 R)
D. alaskensis D. vulgaris Ddeb DVU D. alaskensisc D. vulgarisd
HydrogenasesPeriplasmic
HynBA [NiFe] 1 1 2137–2138*� 1921–1922 �� �HynBAC3 1 1 3754–3756 2524–2526 � �HysBA [NiFeSe] 1 1 2134–2135*† 1917–1918 � �/�HydAB [FeFe] 2 1 0081–0082* 1769–1770 �/� ��
2280–2281* �/�Cytoplasmic
EchABCDEF (membrane bound) 1 0429–0434 �/�CooMKLXUH (membrane bound) 1 2286–2291 �[FeFe] monomeric Fd-dependent (soluble) 1 0725 1771 �[FeFe] NAD(P)-dependent (soluble) 1 �/�
Carbon monoxide dehydrogenase 1 1 3028–3029* 2098–2099 � �/�Cytoplasmic
CooSC
Formate dehydrogenasesPeriplasmic
FdhAB 3 1 0716–0718* 0587–0588 �� �/�0812–0813* �/�3512–3514* �
FdhABC3 1 1 0680–0683* 2809–2812 �/� �/�CfdCDEAB 1 2481–2485 �
Cytoplasmic, FhcABCD 1 0473–0476* �
CytochromesCytochrome c3 (TpI-c3) CycA 1 1 3182 3171 � �/�Split-Soret cytochrome c 1 3211 ��Cytochrome c553 1 2 1821 1817 �/� �/�
3041 �Cytochrome c554 1 2 2858 0702 �/� �
0922 �/�
METCsQrcABCD 1 1 2932–2935*† 0692–0695 � �HmcABCDEF 1 1 0648–0653* 0531–0536 �/� �OhcBAC 1 3143–3145 �DhcARnfCDGEAB 1 1 0580–0586 2791–2797 � �QmoABC-D 1 1 1111–1114*� 0848–0851 � �DsrMKJOP 1 1 2271–2275 1286–12890 �/� �/�TmcABCD 1 1 3707–3710 0263–0265 � �
Carbon metabolism enzymesPyruvate oxidoreductase
Por 1 1 3237 3025 �/� �PorAB 1 1 1792–1793 1569–1570 �/� �
D-Lactate permeaseLctP-1 1 1 1074*† 2285 �� �LctP-2 1 2 3238*† 3026 �� �
2451 �B2975 2 2110 �/�
3284 �/�LldP-1 1 2683 �/�
L-Lactate dehydrogenase (monomeric)L-Ldh 1 0600 �/�L-Ldh 1 1 0750 2784 � �/�
D-Lactate dehydrogenase (monomeric)D-Ldh 1 1 3604 0253 � �
(Continued on following page)
Meyer et al.
992 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
using an RNA nanochip run on a Bioanalyzer (Agilent, Santa Clara, CA).First-strand cDNA (labeled with Alexa 555) was generated using aSuperScript indirect cDNA labeling system (Life Technologies, Carlsbad, CA)with random hexamers and hybridized to a custom Nimblegen microarrayslide containing four independent microarrays (�1 �g per array) com-prised of 72,000 probes each (60-mers). Most genes were represented bynine unique oligonucleotide probes, with each probe arrayed in duplicate.Following overnight incubation in a hybridization station, the array slideswere washed and scanned using a GenPix Pro 4200A scanner followingthe Nimblegen standard protocol (minimum of three photomultipliertube [PMT] gains). The scanned array images were analyzed for probeintensities using the Nimblegen Scan (version 2.4) program with the localalignment turned off and a border of �1. Feature files containing theprobe intensities and probe identifiers were exported for further analysis.PMT settings were used, with the values for �1% of probes being abovehalf of the maximum value. Each scan was normalized by forcing thedistribution of values to conform to the distribution of log2 intensities ofmonoculture control chemostats (average of two monoculture chemo-stats). After excluding probes that were in the lowest 2% of the control,initial log2 levels and log2 ratios (log2 R) for each probe were computed byaveraging the replicates. Since the separate harvests for each of the twococulture chemostats had no influence on the transcription results, wetreated these samples as replicates and averaged the five coculture assaysand the three monoculture assays. To remove intensity-dependent effects,we used local regression (loess in R) of each probe’s log2 ratio versus log2
level and subtracted out these estimated effects. The OpWise program(24) was used to assess whether genes had statistically significant changesin expression.
Transposon mutant construction. The complete description of a D.alaskensis strain G20 transposon mutant collection is in preparation andwill be described elsewhere (A. M. Deutschbauer et al., unpublished data).Briefly, a Tn5 delivery vector was engineered to contain 1 of 4,280 DNAbar codes (TagModule) (25). D. alaskensis strain G20 was mutagenizedwith DNA-bar-coded transposons via conjugation with Escherichia coli
(strain WM3064) on lactate-sulfate agar plates (LS4; conjugations wereincubated overnight at 30°C under anoxic conditions). Matings were re-covered in liquid LS4 medium for 4 to 6 h, and transposon mutants wereselected by plating on LS4 agar plates with 400 �g/ml G418. For each of�15,000 mutant strains, we identified the insertion location of the trans-poson using a two-step arbitrary PCR protocol as previously described(25). The correct insertion of the transposon was further confirmed byPCR amplification with a transposon and genome-specific primer pair foreach mutant strain used in this study.
Phenotypic growth analyses. Dilution series of M. maripaludis and D.alaskensis strain G20 wild-type and mutant strains were prepared fromglycerol stocks in 25-ml Balch tubes containing 13 ml monoculture me-dium (the methanogen was grown in medium containing 5 mM acetatewith an H2-CO2 [80:20, vol/vol] headspace at an overpressure of 30 lb/in2). Cocultures were established by mixing 1 ml of the highest dilution ofexponentially grown cultures of M. maripaludis with the D. alaskensisstrain G20 wild type or a mutant strain in Balch tubes containing 13 mlcoculture medium. Initial batch mono- and cocultures were transferred atleast two times (1%, vol/vol) before inoculation of triplicate tubes formonitoring growth (OD600). All cultures were incubated at 37°C withhorizontal shaking at 300 rpm.
Microarray data accession number. The microarray data are availableunder GEO reference no. GSE42571.
RESULTSGrowth characteristics of D. alaskensis strain G20 in coculturewith M. maripaludis. Continuous cultures of D. alaskensis strainG20 on 30 mM lactate were grown in sulfate-limited monocultureand coculture with M. maripaludis under identical conditions at aconstant dilution rate of 0.047 h�1 (maximum growth rate ofcoculture, 0.051 h�1). During chemostat operation, the D.alaskensis/M. maripaludis coculture cell ratio stabilized at �1.4after a short period of slight fluctuation (Fig. 1). The lactate level
TABLE 1 (Continued)
Redox proteina
Total no. of encoding genes ingenome Locus tag(s) of genes Microarray data (log2 R)
D. alaskensis D. vulgaris Ddeb DVU D. alaskensisc D. vulgarisd
GlcD 1 1 0182 0390 � �/�D-LdhA 1 1 1681 1412 �/� �
Lactate dehydrogenase (oligomeric withHdrD-like subunits)
LldEFG 1 1 1842–1844 1781–1783 �/� �/�LdhA � Ldh-HdrD-1 (LdhB-1) 1 1 3239–3240*† 3027–3028 �� �/�Ldh3 1 1 1085�1087 0826–0827 � �/�
Phosphotransacetylase Pta 3241 3029 �� �Acetate kinase AckA 1 1 3242 3030 � �/�Pyruvate formate lyase Pfl-I 1 1 1273 2824 �/� �Hdr-like proteins
HdrABC-FloxABCD 2 1 1207–1213 2398 � �3523–3530 2405 �
Ldh-HdrD-2 (LdhB-2) 1 1 3245*† 3033 �� �/�FAD-HdrD 1 1 0312 3071 �/�Mop-HdrD 1 1 3540 1558 � �
a Gene annotation according to http//www.microbesonline.org (23), Pereira et al. (22), and Keller and Wall (83).b D. alaskensis strain G20 transposon insertion mutants of respective genes investigated in comparative growth experiments are marked with asterisks (mutants withinhibited/impaired growth in coculture on lactate are highlighted as follows: �, inhibited growth relative to reference strains; †, no growth observed).c Log2 R of D. alaskensis strain G20 grown in coculture with M. maripaludis (30 mM lactate) relative to D. alaskensis strain G20 grown in monoculture (30 mM lactate, 10 mMsulfate) at 0.047 h�1 dilution rate. Log2 R values for enzymes/protein complexes are summarized with the following symbols: ��, log2 R 1; �, log2 R 0.5; �/�, log2 R 0; �,log2 R 0. Redox proteins putatively involved in the syntrophic electron transfer system are highlighted with bold symbols and grey shading. See Tables S2 and S3 in thesupplemental material for detailed lists of log2 R values.d Log2 R of D. vulgaris strain Hildenborough grown in coculture with M. maripaludis (30 mM lactate) relative to D. vulgaris strain Hildenborough grown in monoculture (30 mMlactate, 10 mM sulfate) at 0.039 h�1 dilution rate (data collected from Walker et al. [17]). The definitions of the symbols are as described in footnote c.
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 993
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
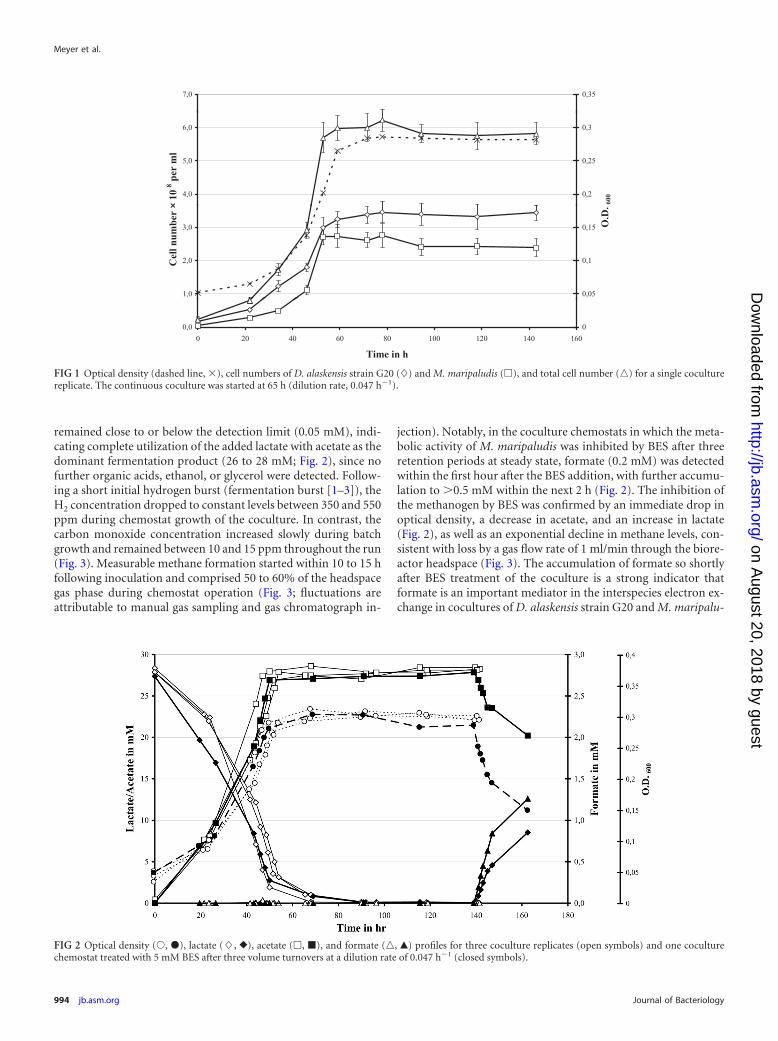
remained close to or below the detection limit (0.05 mM), indi-cating complete utilization of the added lactate with acetate as thedominant fermentation product (26 to 28 mM; Fig. 2), since nofurther organic acids, ethanol, or glycerol were detected. Follow-ing a short initial hydrogen burst (fermentation burst [1–3]), theH2 concentration dropped to constant levels between 350 and 550ppm during chemostat growth of the coculture. In contrast, thecarbon monoxide concentration increased slowly during batchgrowth and remained between 10 and 15 ppm throughout the run(Fig. 3). Measurable methane formation started within 10 to 15 hfollowing inoculation and comprised 50 to 60% of the headspacegas phase during chemostat operation (Fig. 3; fluctuations areattributable to manual gas sampling and gas chromatograph in-
jection). Notably, in the coculture chemostats in which the meta-bolic activity of M. maripaludis was inhibited by BES after threeretention periods at steady state, formate (0.2 mM) was detectedwithin the first hour after the BES addition, with further accumu-lation to 0.5 mM within the next 2 h (Fig. 2). The inhibition ofthe methanogen by BES was confirmed by an immediate drop inoptical density, a decrease in acetate, and an increase in lactate(Fig. 2), as well as an exponential decline in methane levels, con-sistent with loss by a gas flow rate of 1 ml/min through the biore-actor headspace (Fig. 3). The accumulation of formate so shortlyafter BES treatment of the coculture is a strong indicator thatformate is an important mediator in the interspecies electron ex-change in cocultures of D. alaskensis strain G20 and M. maripalu-
7,0 0,35
4,0
5,0
6,0
× 10
8 per
ml
0,2
0,25
0,3
600
1,0
2,0
3,0
Cel
l num
ber
×
0,05
0,1
0,15 O.D
.
0,00 20 40 60 80 100 120 140 160
Time in h
0
FIG 1 Optical density (dashed line, �), cell numbers of D. alaskensis strain G20 (�) and M. maripaludis (�), and total cell number (o) for a single coculturereplicate. The continuous coculture was started at 65 h (dilution rate, 0.047 h�1).
FIG 2 Optical density (Œ, �), lactate (�, }), acetate (�, �), and formate (o, Œ) profiles for three coculture replicates (open symbols) and one coculturechemostat treated with 5 mM BES after three volume turnovers at a dilution rate of 0.047 h�1 (closed symbols).
Meyer et al.
994 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
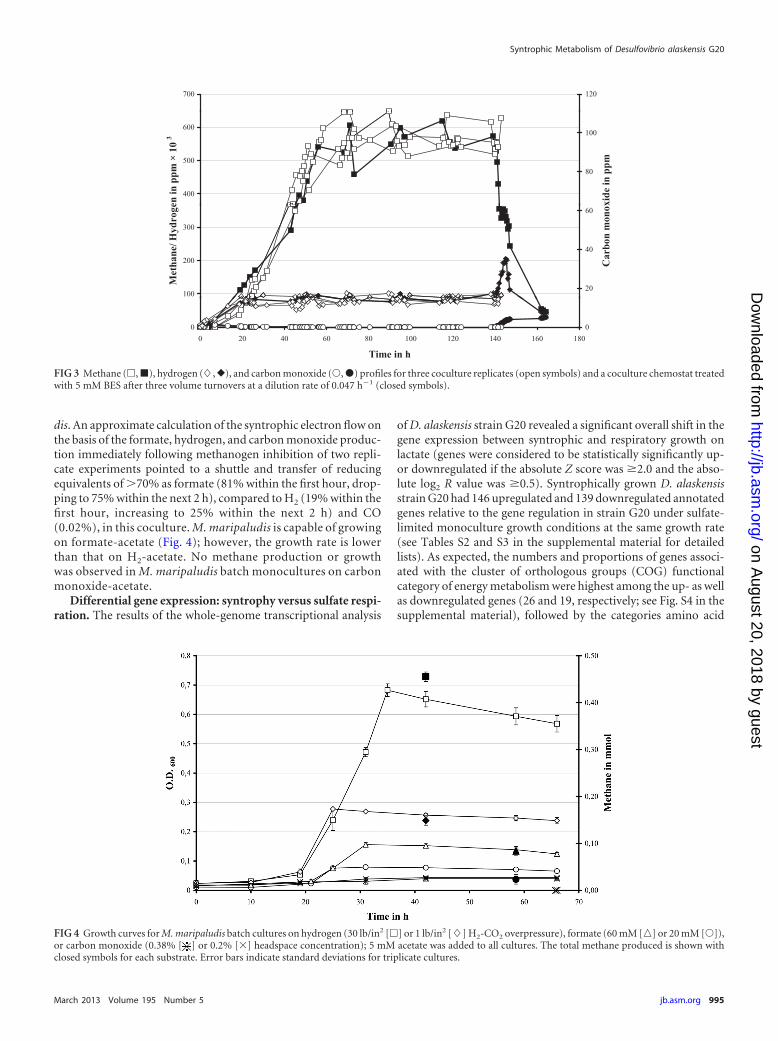
dis. An approximate calculation of the syntrophic electron flow onthe basis of the formate, hydrogen, and carbon monoxide produc-tion immediately following methanogen inhibition of two repli-cate experiments pointed to a shuttle and transfer of reducingequivalents of 70% as formate (81% within the first hour, drop-ping to 75% within the next 2 h), compared to H2 (19% within thefirst hour, increasing to 25% within the next 2 h) and CO(0.02%), in this coculture. M. maripaludis is capable of growingon formate-acetate (Fig. 4); however, the growth rate is lowerthan that on H2-acetate. No methane production or growthwas observed in M. maripaludis batch monocultures on carbonmonoxide-acetate.
Differential gene expression: syntrophy versus sulfate respi-ration. The results of the whole-genome transcriptional analysis
of D. alaskensis strain G20 revealed a significant overall shift in thegene expression between syntrophic and respiratory growth onlactate (genes were considered to be statistically significantly up-or downregulated if the absolute Z score was �2.0 and the abso-lute log2 R value was �0.5). Syntrophically grown D. alaskensisstrain G20 had 146 upregulated and 139 downregulated annotatedgenes relative to the gene regulation in strain G20 under sulfate-limited monoculture growth conditions at the same growth rate(see Tables S2 and S3 in the supplemental material for detailedlists). As expected, the numbers and proportions of genes associ-ated with the cluster of orthologous groups (COG) functionalcategory of energy metabolism were highest among the up- as wellas downregulated genes (26 and 19, respectively; see Fig. S4 in thesupplemental material), followed by the categories amino acid
700 120
400
500
600
e n in
ppm
× 1
0 3
80
100
x ide
in p
pm
100
200
300
Met
hane
/ Hyd
roge
20
40
60
Car
bon
mon
ox
00 20 40 60 80 100 120 140 160 180
Time in h
0
FIG 3 Methane (�, �), hydrogen (�, }), and carbon monoxide (Œ, �) profiles for three coculture replicates (open symbols) and a coculture chemostat treatedwith 5 mM BES after three volume turnovers at a dilution rate of 0.047 h�1 (closed symbols).
FIG 4 Growth curves for M. maripaludis batch cultures on hydrogen (30 lb/in2 [�] or 1 lb/in2 [�] H2-CO2 overpressure), formate (60 mM [o] or 20 mM [Œ]),or carbon monoxide (0.38% [ ] or 0.2% [�] headspace concentration); 5 mM acetate was added to all cultures. The total methane produced is shown withclosed symbols for each substrate. Error bars indicate standard deviations for triplicate cultures.
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 995
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
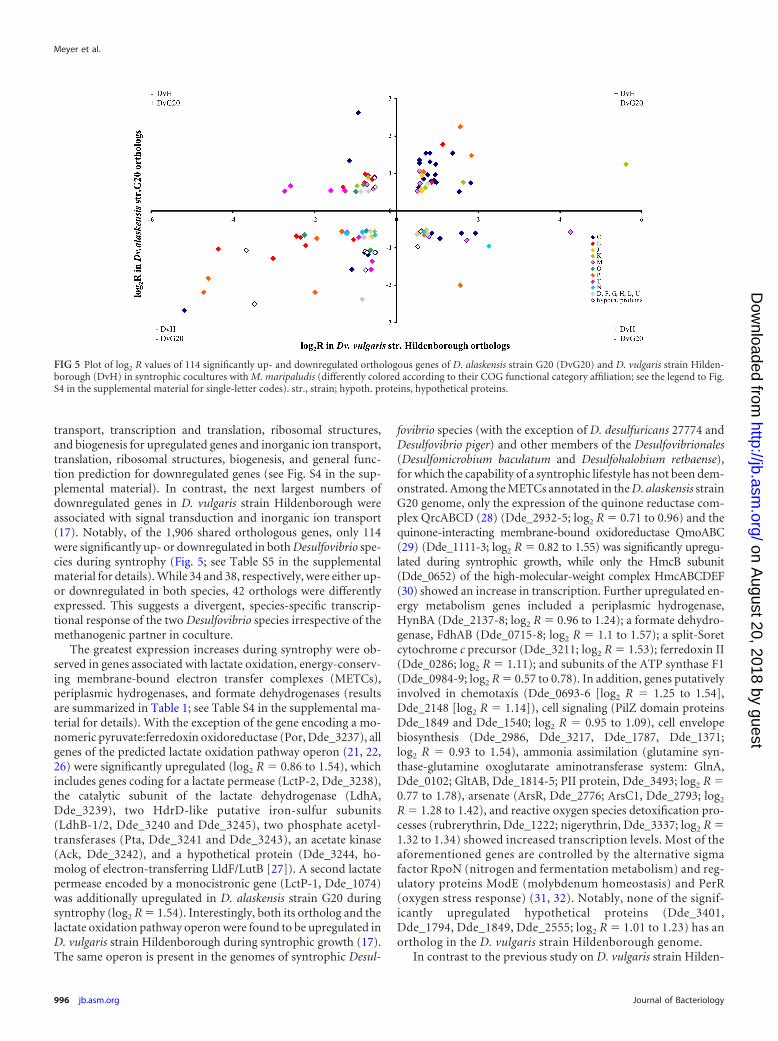
transport, transcription and translation, ribosomal structures,and biogenesis for upregulated genes and inorganic ion transport,translation, ribosomal structures, biogenesis, and general func-tion prediction for downregulated genes (see Fig. S4 in the sup-plemental material). In contrast, the next largest numbers ofdownregulated genes in D. vulgaris strain Hildenborough wereassociated with signal transduction and inorganic ion transport(17). Notably, of the 1,906 shared orthologous genes, only 114were significantly up- or downregulated in both Desulfovibrio spe-cies during syntrophy (Fig. 5; see Table S5 in the supplementalmaterial for details). While 34 and 38, respectively, were either up-or downregulated in both species, 42 orthologs were differentlyexpressed. This suggests a divergent, species-specific transcrip-tional response of the two Desulfovibrio species irrespective of themethanogenic partner in coculture.
The greatest expression increases during syntrophy were ob-served in genes associated with lactate oxidation, energy-conserv-ing membrane-bound electron transfer complexes (METCs),periplasmic hydrogenases, and formate dehydrogenases (resultsare summarized in Table 1; see Table S4 in the supplemental ma-terial for details). With the exception of the gene encoding a mo-nomeric pyruvate:ferredoxin oxidoreductase (Por, Dde_3237), allgenes of the predicted lactate oxidation pathway operon (21, 22,26) were significantly upregulated (log2 R � 0.86 to 1.54), whichincludes genes coding for a lactate permease (LctP-2, Dde_3238),the catalytic subunit of the lactate dehydrogenase (LdhA,Dde_3239), two HdrD-like putative iron-sulfur subunits(LdhB-1/2, Dde_3240 and Dde_3245), two phosphate acetyl-transferases (Pta, Dde_3241 and Dde_3243), an acetate kinase(Ack, Dde_3242), and a hypothetical protein (Dde_3244, ho-molog of electron-transferring LldF/LutB [27]). A second lactatepermease encoded by a monocistronic gene (LctP-1, Dde_1074)was additionally upregulated in D. alaskensis strain G20 duringsyntrophy (log2 R � 1.54). Interestingly, both its ortholog and thelactate oxidation pathway operon were found to be upregulated inD. vulgaris strain Hildenborough during syntrophic growth (17).The same operon is present in the genomes of syntrophic Desul-
fovibrio species (with the exception of D. desulfuricans 27774 andDesulfovibrio piger) and other members of the Desulfovibrionales(Desulfomicrobium baculatum and Desulfohalobium retbaense),for which the capability of a syntrophic lifestyle has not been dem-onstrated. Among the METCs annotated in the D. alaskensis strainG20 genome, only the expression of the quinone reductase com-plex QrcABCD (28) (Dde_2932-5; log2 R � 0.71 to 0.96) and thequinone-interacting membrane-bound oxidoreductase QmoABC(29) (Dde_1111-3; log2 R � 0.82 to 1.55) was significantly upregu-lated during syntrophic growth, while only the HmcB subunit(Dde_0652) of the high-molecular-weight complex HmcABCDEF(30) showed an increase in transcription. Further upregulated en-ergy metabolism genes included a periplasmic hydrogenase,HynBA (Dde_2137-8; log2 R � 0.96 to 1.24); a formate dehydro-genase, FdhAB (Dde_0715-8; log2 R � 1.1 to 1.57); a split-Soretcytochrome c precursor (Dde_3211; log2 R � 1.53); ferredoxin II(Dde_0286; log2 R � 1.11); and subunits of the ATP synthase F1(Dde_0984-9; log2 R � 0.57 to 0.78). In addition, genes putativelyinvolved in chemotaxis (Dde_0693-6 [log2 R � 1.25 to 1.54],Dde_2148 [log2 R � 1.14]), cell signaling (PilZ domain proteinsDde_1849 and Dde_1540; log2 R � 0.95 to 1.09), cell envelopebiosynthesis (Dde_2986, Dde_3217, Dde_1787, Dde_1371;log2 R � 0.93 to 1.54), ammonia assimilation (glutamine syn-thase-glutamine oxoglutarate aminotransferase system: GlnA,Dde_0102; GltAB, Dde_1814-5; PII protein, Dde_3493; log2 R �0.77 to 1.78), arsenate (ArsR, Dde_2776; ArsC1, Dde_2793; log2
R � 1.28 to 1.42), and reactive oxygen species detoxification pro-cesses (rubrerythrin, Dde_1222; nigerythrin, Dde_3337; log2 R �1.32 to 1.34) showed increased transcription levels. Most of theaforementioned genes are controlled by the alternative sigmafactor RpoN (nitrogen and fermentation metabolism) and reg-ulatory proteins ModE (molybdenum homeostasis) and PerR(oxygen stress response) (31, 32). Notably, none of the signif-icantly upregulated hypothetical proteins (Dde_3401,Dde_1794, Dde_1849, Dde_2555; log2 R � 1.01 to 1.23) has anortholog in the D. vulgaris strain Hildenborough genome.
In contrast to the previous study on D. vulgaris strain Hilden-
FIG 5 Plot of log2 R values of 114 significantly up- and downregulated orthologous genes of D. alaskensis strain G20 (DvG20) and D. vulgaris strain Hilden-borough (DvH) in syntrophic cocultures with M. maripaludis (differently colored according to their COG functional category affiliation; see the legend to Fig.S4 in the supplemental material for single-letter codes). str., strain; hypoth. proteins, hypothetical proteins.
Meyer et al.
996 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

borough, all key genes for sulfate respiration (sulfate permeases,ATP sulfurylase, dissimilatory adenosine 5=-phosphosulfate[APS] and sulfite reductase, and pyrophosphatase) are downregu-lated in D. alaskensis strain G20 under syntrophic conditions.Other significantly downregulated genes included the iron-re-pressed flavodoxin (fld, Dde_3667; log2 R � �2.67), ferrous ironuptake transporters (feoAB, Dde_2669-73; log2 R � �1.81 to�2.19), the Tmc complex (tmcABCD, Dde_3707-10; log2 R ��0.91 to �1.19), periplasmic [NiFeSe] hydrogenase (hysAB,Dde_2134-5; log2 R � �1.08 to �1.25), chaperones (mopAB,Dde_2473-4; dnaK, Dde_1023; copZ, Dde_1312; log2 R � �0.54to �2.13), three operons encoding ABC-type transporters (twoTonB/TolQR-associated biopolymer transport systems,Dde_0447-57 and Dde_2152-57; log2 R � �0.72 to �1.47),Zn2�-dependent peptidase (pqqL, Dde_0444-46; log2 R � �1.06to �1.47), and a cluster of genes coding for putative dinitrogenaseiron-molybdenum cofactor biosynthesis proteins (Dde_3197 toDde_3202; log2 R � �2.42 to �2.67). In addition, an operon oftwo-component system response regulators (Dde_3711 toDde_3715; log2 R � �1.12 to �1.6) and several monocistronicgenes encoding hypothetical proteins with D. vulgaris strainHildenborough orthologs (Dde_2020, Dde_2315, Dde_1145,Dde_2212, Dde_1143, Dde_0283) are negatively expressed duringsyntrophy relative to respiratory growth (log2 R � �0.93 to�2.00). Notably, most of these genes are part of the Rex (energymetabolism) or Fur (iron homeostasis) regulon (31, 33, 34).
Comparative phenotypic growth experiments of mutantstrains. On the basis of the previous results, phenotypic growthanalyses of 23 D. alaskensis strain G20 transposon mutants withinsertions in genes coding for subunits of formate dehydroge-nases, hydrogenases, METCs, lactate metabolism enzymes, andtwo hypothetical proteins as additional reference strains forgrowth (Dde_1072 and Dde_2939) were carried out in mono- andcoculture on lactate or pyruvate to evaluate the function of therespective genes in syntrophy (Fig. 6). Surprisingly, all mutants,including both aforementioned reference strains, showed an ex-tended lag phase in lactate-based coculture growth relative to thewild type, whereas no differences were observed when grown incoculture on pyruvate or in monoculture on both substrates. Thephenotypic growth behavior of all mutants in coculture on lactateis therefore discussed with reference to the mutant strainDde_1072 (Fig. 6).
None of the mutants of formate dehydrogenases were inhib-ited in syntrophic growth on lactate or pyruvate. Among the mu-tants of periplasmic hydrogenases, the HysA mutant (Dde_2135)was impaired in coculture growth on lactate, although the hysABgenes showed significant repression under steady-state conditionsin the chemostat (negative absolute log2 values; data not shown).Furthermore, the HynA mutant (Dde_2138) showed an extendedlag phase and reduced growth rate during syntrophy but reached amaximum cell density comparable to that of the reference strain.None of the investigated formate dehydrogenase and hydrogenasemutants demonstrated growth inhibition during sulfate respira-tion on lactate or pyruvate. Among the METC subunit mutants,the QrcD mutant (Dde_2935) showed no growth in coculture onlactate, while mutants for QmoD (Dde_1114) and HmcB(Dde_0652) were inhibited with lower growth rates and maxi-mum cell densities (74 and 66%, respectively) relative to those forthe reference strain. The QmoD mutant demonstrated inhibitedrespiratory growth on both substrates, consistent with a putative
function within the QmoABC complex (35) (L. R. Krumholz etal., submitted for publication), whereas the QrcD mutant wasimpaired in monoculture only on pyruvate (27% of the wild-typeyield). The mutants of both lactate permeases (LctP-1 [Dde_1074]and LctP-2 [Dde_3238]) and the HdrD-like putative beta sub-units of the lactate dehydrogenase (LdhB-1 [Dde_3240] andLdhB-2 [Dde_3245]) showed no growth in coculture on lactate,confirming their essential role during syntrophy. However, asdemonstrated by slightly inhibited (LdhB-1/2) and impaired(LctP-2) growth or reduced maximum cell density (LctP-1, with acell density 20% of that of the wild type) on lactate in monocul-ture, they are also involved in the lactate uptake/oxidation pro-cesses during sulfate respiration. Notably, the mutant for the lac-tate oxidation operon integral permease LctP-2 also showedinhibited growth in mono- and coculture on pyruvate.
DISCUSSION
On the basis of the whole-genome transcription and mutantgrowth experiment data, a conceptual model for the D. alaskensisstrain G20 electron transfer system has been developed for syn-trophic growth with M. maripaludis on lactate (Fig. 7A). Lactate isoxidized by a cytoplasmic lactate dehydrogenase and pyruvate:ferredoxin oxidoreductase to acetyl coenzyme A (acetyl-CoA),generating a reduced thiol-disulfide redox pair and reducedferredoxin. The membrane-bound Qmo complex couples theoxidation of both reduced electron carriers in a flavin-basedreverse electron bifurcation process (electron confurcation) tothe reduction of periplasmic cytochromes, formate dehydroge-nases, and hydrogenases via menaquinone (MQ) and mem-brane-bound Qrc complex (reverse electron flow), yieldingboth formate and H2 as final products for the exchange with themethanogen.
Metabolic model for syntrophic growth of D. alaskensisstrain G20. Lactate is transported in symport with one proton intothe cell via two putative lactate permeases, Lct-P1 and Lct-P2(sharing only 26% sequence identity). Remarkably, the concertedfunction of both permeases is required for syntrophy, since inser-tional inactivation of either gene resulted in loss of the ability forsyntrophic growth of D. alaskensis strain G20 on lactate (Fig. 7).Since all so far characterized lactate permeases have been demon-strated to transport both isomeric forms (e.g., LldP of E. coli) (36,37), this phenotype is not attributable to each permease beingdedicated to either L- or D-lactate uptake. The poor growth of theLctP-2 mutant on pyruvate in both mono- and coculture is sug-gestive of a functioning as a low-affinity lactate permease with anunusual double function as a pyruvate permease, while LctP-1might be a high-affinity lactate permease expressed under lowlactate concentrations. However, both lactate transporters mightbe required to drive the flux of lactate needed to support syn-trophic growth. Alternatively, LctP-2 could be a misannotatedacetate permease, since the sequence identity to the known lactatepermeases is low (30%) and homologs to acetate transporters ofbacteria (e.g., ActP in E. coli [38, 39]) are missing in the D. alasken-sis strain G20 genome.
Of the seven candidate gene loci encoding mono- or oligo-meric lactate dehydrogenases in D. alaskensis strain G20, only oneis significantly upregulated during syntrophy (Dde_3239-40 andDde_3245). This cytoplasmic lactate dehydrogenase, LdhAB, isnot homologous to the previously described monomeric, qui-none-interacting lactate oxidases of E. coli (27, 40) but instead
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 997
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
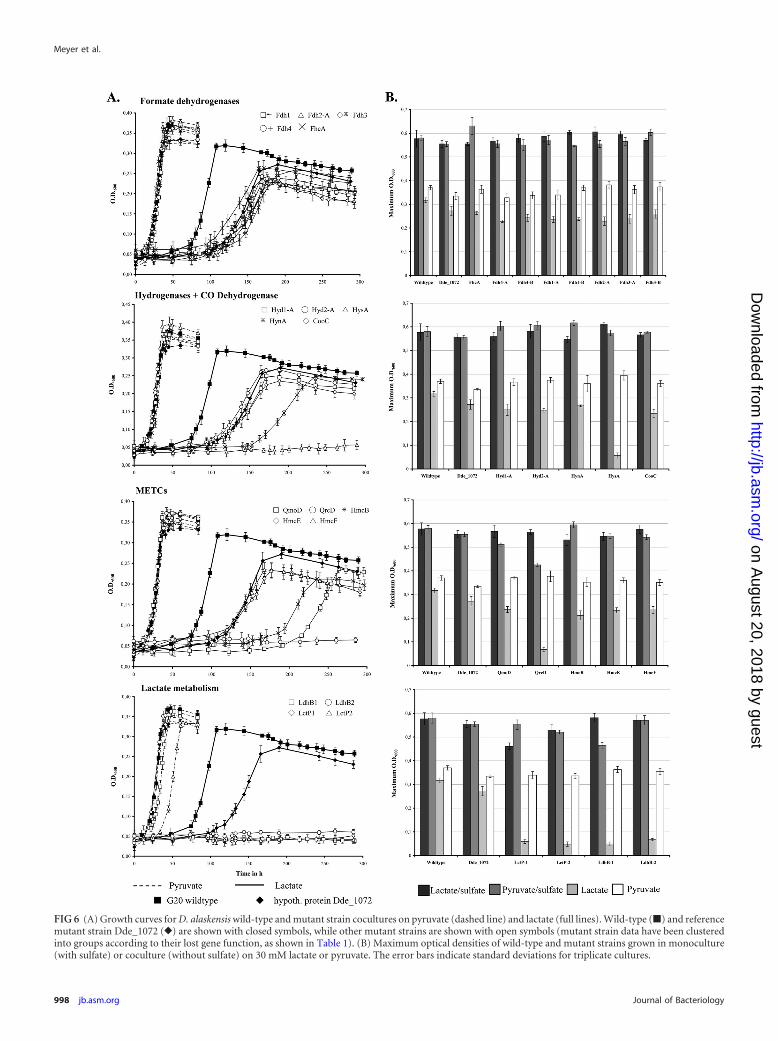
FIG 6 (A) Growth curves for D. alaskensis wild-type and mutant strain cocultures on pyruvate (dashed line) and lactate (full lines). Wild-type (�) and referencemutant strain Dde_1072 (}) are shown with closed symbols, while other mutant strains are shown with open symbols (mutant strain data have been clusteredinto groups according to their lost gene function, as shown in Table 1). (B) Maximum optical densities of wild-type and mutant strains grown in monoculture(with sulfate) or coculture (without sulfate) on 30 mM lactate or pyruvate. The error bars indicate standard deviations for triplicate cultures.
Meyer et al.
998 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
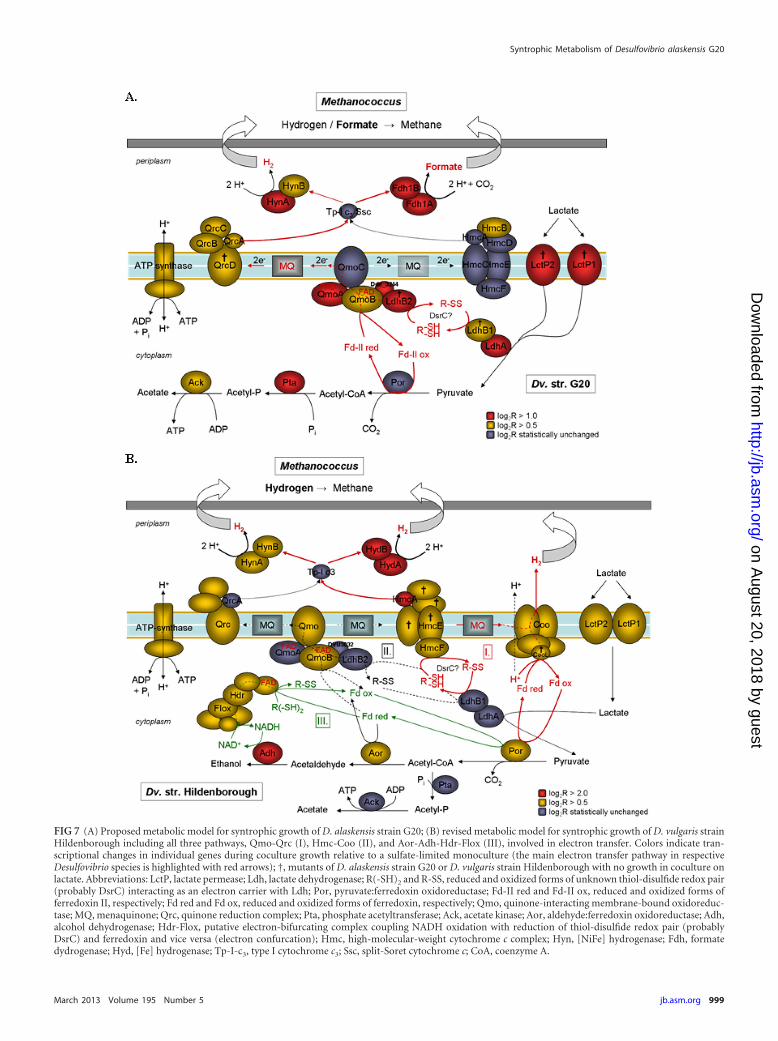
FIG 7 (A) Proposed metabolic model for syntrophic growth of D. alaskensis strain G20; (B) revised metabolic model for syntrophic growth of D. vulgaris strainHildenborough including all three pathways, Qmo-Qrc (I), Hmc-Coo (II), and Aor-Adh-Hdr-Flox (III), involved in electron transfer. Colors indicate tran-scriptional changes in individual genes during coculture growth relative to a sulfate-limited monoculture (the main electron transfer pathway in respectiveDesulfovibrio species is highlighted with red arrows); †, mutants of D. alaskensis strain G20 or D. vulgaris strain Hildenborough with no growth in coculture onlactate. Abbreviations: LctP, lactate permease; Ldh, lactate dehydrogenase; R(-SH)2 and R-SS, reduced and oxidized forms of unknown thiol-disulfide redox pair(probably DsrC) interacting as an electron carrier with Ldh; Por, pyruvate:ferredoxin oxidoreductase; Fd-II red and Fd-II ox, reduced and oxidized forms offerredoxin II, respectively; Fd red and Fd ox, reduced and oxidized forms of ferredoxin, respectively; Qmo, quinone-interacting membrane-bound oxidoreduc-tase; MQ, menaquinone; Qrc, quinone reduction complex; Pta, phosphate acetyltransferase; Ack, acetate kinase; Aor, aldehyde:ferredoxin oxidoreductase; Adh,alcohol dehydrogenase; Hdr-Flox, putative electron-bifurcating complex coupling NADH oxidation with reduction of thiol-disulfide redox pair (probablyDsrC) and ferredoxin and vice versa (electron confurcation); Hmc, high-molecular-weight cytochrome c complex; Hyn, [NiFe] hydrogenase; Fdh, formatedydrogenase; Hyd, [Fe] hydrogenase; Tp-I-c3, type I cytochrome c3; Ssc, split-Soret cytochrome c; CoA, coenzyme A.
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 999
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

shows sequence similarity to the oligomeric lactate dehydrogenaseLldEFG/LutABC (27, 41, 42). During syntrophic growth, LdhABoxidizes lactate to pyruvate and shuttles the electrons to the Qmocomplex. As both iron-sulfur subunits LdhB-1 and LdhB-2 arerelated to the catalytically active subunits (HdrD/HdrB) of theheterodisulfide reductases of methanogens (43, 44) and each con-tains two CCG domains (catalytic sites), the electron carrier be-tween Ldh and Qmo is most likely a low-molecular-weight thiol-disulfide redox pair or a redox-active cysteine residue containingprotein like DsrC (45, 46). Both proteins could operate as a thiol-disulfide-dependent shuttle system that is essential for linking thecytoplasmic lactate oxidation (LdhB-1) to the membrane-boundMQ-reducing complex Qmo (via LdhB-2 and Dde_3244) duringsyntrophy, which is consistent with the impaired growth of bothLdhB mutants in coculture. Interestingly, the activities of mostcharacterized lactate dehydrogenases from Desulfovibrio/Desulfo-microbium species were demonstrated to be tightly associated withthe membrane (despite the absence of a membrane-integral sub-unit) and to require reducing conditions (thiol compoundsneeded for assays), while the in vivo electron acceptor remainedunknown (47–51). The immediate product of lactate oxidation,pyruvate, is oxidized by a pyruvate:ferredoxin oxidoreductase(ortholog of homodimeric Por of D. africanus [52–54]),thereby generating reduced ferredoxin II (upregulatedDde_0286) and acetyl-CoA. The latter compound is furtherconverted to acetate via acetyl phosphate mediated by phos-photransacetylase and acetate kinase (upregulated genes of thelactate oxidation pathway operon), which sustains energy gener-ation by substrate-level phosphorylation. Since arsenate has beendemonstrated to uncouple the oxidation of lactate from ATP syn-thesis by the affecting Pta activity (50), the upregulation of itsdetoxification system (arsC1, arsRBCC [55]) ensures a rapid re-sponse to a potential arsenate influx to maintain the main energyconservation process during syntrophic growth.
The major energetical difficulty in the lactate utilization undersyntrophic conditions is that the oxidation of lactate to pyruvate(E°=, �190 mV) with protons or CO2 as electron acceptors (E°=,�414 mV and �432 mV, respectively) is highly endergonic evenat low H2 levels (E°=, �270 mV at H2 partial pressure of 1 Pa).Therefore, oxidation of lactate requires energy input by reverseelectron transport (12, 50) which must be derived from the pyru-vate oxidation to acetyl-CoA and CO2 and reduced ferredoxin(E°=, �498 mV), with the latter having a sufficiently low redoxpotential to drive H2/formate generation (Fdox/Fdred redox coupleE°=, �420 mV) (12). The additional energetic costs are reflected inthe lower biomass yield and growth rate of the syntrophic cocul-ture on lactate compared to pyruvate (Fig. 6). Interestingly, D.alaskensis strain G20 does not possess cytoplasmic membrane-bound hydrogenases like the Coo complex, which is essential forsyntrophic growth in D. vulgaris strain Hildenborough (17). Itsalternative enzymes, a soluble putative [FeFe] hydrogenase(Dde_0725) and putative formate:H2 lyase (Dde_0473-76), aredownregulated during syntrophy. Instead, the experimental dataof this study implicate two METCs to be functionally important inD. alaskensis strain G20 during syntrophy on lactate, the Qmo andQrc complexes (28, 29). The Qmo complex consists of three sub-units, QmoABC, that are essential for the sulfate reduction path-way in SRB by channeling electrons from the MQ pool to thecytoplasmic APS reductase (AprBA) (29, 35, 56). A fourth protein,QmoD (Dde_1114), has recently been found to be QmoABC as-
sociated in two-dimensional protein gels of lactate-sulfate-grownDesulfovibrio species (57) (Krumholz et al., submitted), indicatinga possible function as a structural scaffold for the other subunits.The membrane-bound QmoC (HdrE-HdrC fusion protein) linksthe electron flow between the MQ pool and the cytoplasmic sub-units of the complex, QmoA and QmoB, which are both iron-sulfur flavoproteins homologous to HdrA, the flavin-containingsubunit of the soluble heterodisulfide reductase (29, 44, 56). Inmethanogens, the HdrA is involved in a flavin-based electron bi-furcation process that couples the exergonic reduction of a het-erodisulfide with H2/formate to the endergonic reduction of ferre-doxin in an Mvh hydrogenase/formate dehydrogenase-HdrABCcomplex (44, 58–60). Interestingly, QmoB has another domainhomologous to the Mvh hydrogenase subunit MvhD which is re-sponsible for electron transfer to HdrABC (29, 44, 56). In sulfaterespiration, QmoABC proteins were recently suggested to interactwith AprBA, the MQ pool, and a third physiological partner of lowredox potential to couple the cytoplasmic APS reduction tochemiosmotic energy conservation by a reverse electron bifurca-tion mechanism (56). As an explanation for its essential functionin syntrophic growth on lactate, we hypothesize that the oxidationof the thiol-disulfide redox pair (serving as a soluble electron car-rier between LdhAB-1 and the Qmo complex) is mediated by theHdrD-like LdhB-2 (possibly via Dde_3244), which transfers theelectrons to QmoB. The FAD group of the QmoB subunit operatesas a coupling site that confurcates electrons from the low-poten-tial electron donor ferredoxin II and the high-potential thiol-di-sulfide pair to drive the reduction of menaquinol via a reducedQmoC (redox potentials of hemes b are �75 and �20 mV for D.desulfuricans ATCC 27774 [29]). We propose that an efficientlinkage of the periplasmic proton/CO2 reduction to the cytoplas-mic thiol-disulfide redox pair oxidation (via the MQ pool, Qrccomplex, and type I cytochrome c3 [TpIc3]) at elevated H2 levels(460 ppm measured, on average, at a high coculture growth rate)is possible only via the QmoB-mediated electron confurcationmechanism. Alternatively, the ferredoxin oxidation could be me-diated by QmoA, since AprB interacts with the QmoA via a ferre-doxin-like domain; however, the proposed specific docking site(61) is missing in the ferredoxin II sequence.
Qrc is present in all SRB that possess periplasmic hydrogenasesand formate dehydrogenases lacking a membrane subunit for qui-none interaction (22). Therefore, Qrc was suggested to link theperiplasmic enzymes to the quinone pool by acting as a TpIc3:menaquinone oxidoreductase during respiratory growth on H2/formate (22, 28, 62). Recently, its essential function in syntrophywas proposed on the basis of growth defects of qrcB and cycAtransposon mutants in coculture (19). The higher redox potentialof the MQ relative to the H�/H2 and CO2/formate redox pairsrequires a second energy input to efficiently drive the electron flowfrom the MQ pool (�80 mV [12, 29]) to the periplasmic hydro-genases and formate dehydrogenases via the membrane-boundQrc complex and TpIc3 (heme c group redox potentials in D.vulgaris strain Hildenborough, �80 to �110 mV and �325to �170 mV, respectively [28]). This energy investment is mostlikely derived from proton intrusion using reverse electron trans-port mediated by a Qmo- and Qrc-linked MQ redox loop. As theexact location for proton uptake of the MQ-interacting QrcD sub-unit is still unresolved (sequence-based prediction models posi-tion the quinone-binding site at the periplasmic site of the mem-brane [28, 29]), a reverse electron transport from Qmo to Qrc via
Meyer et al.
1000 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

MQ during syntrophy would result in the intrusion of either twoprotons (two proton motive force-consuming half-redox loops;H�/e� � �2 [63]) or one proton per transferred electron (elec-troneutral quinol oxidation by Qrc; H�/e� � �1). Irrespective ofthis, the inward movement of protons by reverse MQ cyclingwould need to be compensated for by proton extrusion mediatedby the membrane-bound ATPase using the energy generated inthe substrate-level phosphorylation step.
During syntrophic growth on lactate at a constant high growthrate, the [NiFe] hydrogenase HynBA seems to be the main enzymeresponsible for periplasmic H2 production in D. alaskensis strainG20 (confirmed by absolute log2 data; not shown). The rela-tively minor growth defect conferred by the hyn mutants incoculture may reflect compensation by the other functionallysimilar periplasmic hydrogenases. Indeed, D. alaskensis strainG20 has four periplasmic hydrogenases (Hys, Hyn, and two Hydhydrogenases) that could potentially be involved in H2 generationunder syntrophic conditions. The impaired growth of the hys mu-tants might indicate the importance of the [NiFeSe] hydrogenasefor lactate fermentation during the early stages of coculturegrowth with M. maripaludis, while the [NiFe] hydrogenase ismore important under steady-state conditions in the chemostat.The hydrogenases possess unique kinetic properties as a result ofdissimilar metal compositions in their active sites and are differ-ently regulated in D. vulgaris strain Hildenborough during sulfaterespiration as a function of medium composition and growth con-ditions (64–66). Both, hyn and hys were suggested to functionbidirectionally (67); however, the recently discovered supercom-plex between the [NiFe] hydrogenase, TpIc3, and Qrc (68) mightensure an optimized electron transfer that is necessary for syn-trophic H2 production under constantly elevated H2 levels. Theupregulation of the split-Soret cytochrome c (Ssc) gene duringsyntrophy points to a possible involvement in an alternative path-way for electron transport, in addition to that served by TpIc3.Reduction of the Ssc by periplasmic dehydrogenases has indeedbeen demonstrated (69), but previous studies support a role onlyin nitrate respiration (70).
The periplasmic formate dehydrogenases of Desulfovibrio dif-fer in their kinetic properties as a result of various subunit andcofactor compositions (71, 72). If tungsten is available, D. alasken-sis strain G20 uses mainly the W-formate dehydrogenase (Fdh-1)during respiratory growth on formate but downregulates thegenes for the Mo-/W-incorporating enzyme (Fdh-3, Dde_3512-14) (71). The increased transcription also implicates Fdh-1 as amajor source of formate during syntrophy, but the loss of one Fdhcan apparently be compensated for by the isoenzymes (possibly bysubunit substitution), as shown in the mutant growth studies.Notably, at least 70% of electron flux is directed from the reducedQrc complex/TpIc3 (or Ssc) to the periplasmic Fdh during syn-trophic growth. A cytoplasmic production of formate in syn-trophic D. alaskensis strain G20 is unlikely because of the lowtranscription levels of the pyruvate:formate and formate:H2 lyases(fhc mutants have no growth defect in coculture), absence of aformate transporter, and slow excretion rates of the protonatedform by a simple diffusion process that would be inconsistent withthe measured growth rate of the coculture.
Revised metabolic model for syntrophic growth of D. vul-garis strain Hildenborough. Compared to D. alaskensis strainG20, the electron transfer system of D. vulgaris strain Hildenbor-ough is more complex, involving a larger number of cyto- and
periplasmic proteins that appear to be linked in three intertwiningpathways to energetically couple reduction of protons to lactateoxidation: pathway I, Qmo-Qrc; pathway II, Hmc-Coo; and path-way III, Aor-Adh-Hdr-Flox (Fig. 7B). Besides the Qmo-Qrc loop,the thiol-disulfide redox pair derived from the LdhAB-mediatedlactate oxidation is mainly reoxidized by the HmcF subunit(HdrD homolog) of the membrane-bound Hmc complex andchanneled via the MQ pool (putative energy input by reverse re-dox loop) to the membrane-bound Coo hydrogenase that usesreduced ferredoxin for cytoplasmic H2 production linked to pro-ton/sodium translocation (12, 22). The endergonic formation ofH2 (via TpIc3 and the periplasmic hydrogenases Hyd and Hyn)with electrons from MQ might be directly energetically coupled tothe exergonic proton reduction from the reduced ferredoxin inthe Coo complex or indirectly coupled through proton intrusion.Alternatively, a periplasmic proton reduction could beachieved by an MQ-based reverse redox loop involving HmcEFas menaquinone reductase and HmcABCD as menaquinol oxi-dase. HmcE is homologous to the cytochrome b integral mem-brane subunit HdrE (43, 44), while HmcC belongs to a family ofproteins that interact with the quinone pool without the involve-ment of heme b, as for QrcD (28). Indeed, previous studies havedemonstrated the Hmc complex to be involved in bidirectionalelectron transport between the cytoplasm and periplasm (73, 74).On the basis of the mutant growth results of Walker et al. (17), theHmc-Coo loop represents the main route for syntrophic electrontransfer in D. vulgaris strain Hildenborough. At high growth rate,a transient disposal of excess reducing equivalents by reduction ofacetyl-CoA to ethanol occurs (via upregulated Aor and Adh), withthe NADH being generated by the cytoplasmic HdrABC-Flox-ABCD complex via electron bifurcation (22). The function of theAor-Adh-Hdr-Flox-mediated pathway might be to maintain suf-ficiently low intracellular reducing equivalents/H2 levels for theoperating cytoplasmic Coo hydrogenase.
Effect of alternative electron transfer systems and interspe-cies electron-exchange mediators on growth characteristics ofsyntrophic Desulfovibrio species. According to the metabolicmodels presented here, D. alaskensis strain G20 relies on energyconservation via substrate-level phosphorylation for syntrophicgrowth. In D. vulgaris strain Hildenborough, however, the H2-forming cytoplasmic Coo hydrogenase could contribute to protonmotive force by its cytoplasmic H2 production (and putative H�/Na� translocation); thus, it might be more energy efficient andflexible in adapting to various conditions under syntrophy. Thenevertheless faster coculture growth rate of strain G20 comparedto that of strain Hildenborough might be based on two factors: (i)a kinetic advantage conferred by the Qmo-MQ-Qrc loop as anelectron transfer system that permits higher lactate oxidation ratesunder elevated H2 levels (thereby enhancing the growth of M.maripaludis) and (ii) use of formate as the main electron-ex-change mediator (70% electron flux) by involving periplasmicformate dehydrogenases (as opposed to the exclusively H2-basedexchange by strain Hildenborough). Diffusion kinetics based onFick’s law showed that interspecies formate transfer could sustaina 100-fold higher conversion rate than interspecies hydrogentransfer (7, 8). Indeed, several studies have indicated the impor-tance of formate as an electron shuttle in propionate- and bu-tyrate-oxidizing syntrophs (3, 5–11). However, to our knowledge,this is the first description of its involvement in lactate oxidationof syntrophic Desulfovibrio species.
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 1001
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

Divergent transcriptional profiles of syntrophic Desulfovi-brio species. The comparative transcriptional analysis of thisstudy showed highly divergent gene expression profiles in bothDesulfovibrio species, which might indicate the absence of a con-served pool of genes that sustain syntrophy (Fig. 5; see Table S5 inthe supplemental material). The observed common up- or down-regulation of certain orthologs appears to be related to environ-mental conditions not directly related to the syntrophic lifestyle.For example, the downregulation of Fur regulon-associated genes,e.g., iron-repressed flavodoxin, ferrous iron, and molybdenumtransporters (FeoAB, ModAB), and TonB-associated ABC-typebiopolymer transport systems (COGs C, P, and E) is a direct con-sequence of the lack of sulfide production by syntrophically grownDesulfovibrio, which results in a greater availability of reducedmetals like Fe2� and Mo2�. The reduced metals work as corepres-sors of the Fur protein (31, 33). Most of the commonly upregu-lated genes are part of the RpoN regulon and code for lactateoxidation-associated energy metabolism (COG C), general tran-scription and translation factors (tuf, nusG, and infA; COGs K andJ), ammonia assimilation (glnN and gluA; COG E), envelope bio-synthesis (kdsA, gmD, and lpxL; COG M), and FeS cluster biosyn-thesis (SufA/IscA-like proteins, rhodanese-related sulfur trans-ferase, and DoxD-like membrane protein [75–77]). Strikingly,there were no significantly upregulated orthologs coding for pro-teins of unknown function. The overall physiological switch fromrespiratory to fermentative metabolism in both investigated spe-cies is most likely coordinated by the alternative sigma factorRpoN in combination with RexA (repressor of the sulfate reduc-tion pathway [34]). Indeed, recent studies have shown that RpoNfunctions in Deltaproteobacteria as a global regulator controlling avariety of cellular processes, including flagellum biosynthesis andmotility/biofilm formation (32, 78). While both syntrophic Desul-fovibrio species differed in expression of the individual flagellarsystem components, the regulatory module FliW-CsrA (posttran-scriptional control of flagellin synthesis [79]) was upregulated inboth species. This indicates that bacterial flagellum-mediatedcell-cell communication might be a relatively common mech-anism to initiate and synchronize syntrophic interactions innature (14, 15).
Most differently expressed orthologs are associated with cellenvelope biogenesis, cyto- or periplasmic transport, intracellularsignal transduction, and regulatory processes, which furtherstresses the species specificity of syntrophic interactions amongDesulfovibrio species (differently regulated genes include, e.g.,lipid A core ligase, six undescribed membrane proteins, threeABC-type transporters for phosphate and amino acids, and sixuncharacterized response regulators; COGs M, P, E, K, and T).Notably, none of these regulatory proteins have an assigned func-tion, despite recent progress (78), but their CheY-like receiverdomains are indicative for a function in alternative chemosensorypathways (e.g., exopolysaccharide production) or energy taxis(80, 81) (e.g., Dde_3246 in the lactate oxidation pathway operon).As shown in E. coli (82), the interacting regulators UvrY(Dde_0977) and CsrA (Dde_3150) could be involved in a quorumsensing-dependent regulation of motility and biofilm formationof D. alaskensis strain G20. The ability to navigate to positionswhere metabolic rates are optimized and to control the transitionbetween the motile and biofilm lifestyles is especially importantunder conditions close to the thermodynamic equilibrium.
In summary, this study demonstrated the involvement of al-
ternative electron transfer systems and energy conservation mech-anisms in syntrophic Desulfovibrio species, the importance of for-mate as an interspecies electron-exchange mediator in syntrophicD. alaskensis strain G20 relative to the primarily H2-based systemin D. vulgaris strain Hildenborough, and the species specificity ofsyntrophic Desulfovibrio transcriptional profiles. These differ-ences may reflect the alternative adaptive strategies used by Desul-fovibrio species in environments that fluctuate in electron accep-tor availability. Despite the absence of a single strategy forsyntrophy, which would be reflected by a conserved core of syn-trophy-associated genes, the results of this study confirm the func-tional importance of thiol-disulfide redox pairs and electron bi-furcation/confurcation processes as common mechanisms forlinking exergonic and endergonic reaction in sulfate reducers.
ACKNOWLEDGMENT
This work, conducted by ENIGMA—Ecosystems and Networks Inte-grated with Genes and Molecular Assemblies (http://enigma.lbl.gov), aScientific Focus Area program at Lawrence Berkeley National Laboratory,was supported by the Office of Science, Office of Biological and Environ-mental Research, of the U.S. Department of Energy under contract no.DE-AC02-05CH11231.
REFERENCES1. McInerney MJ, Struchtemeyer CG, Sieber J, Mouttaki H, Stams AJM,
Schink B, Rohlin L, Gunsalus RP. 2008. Physiology, ecology, phylogeny,and genomics of microorganisms capable of syntrophic metabolism. AnnN. Y. Acad. Sci. 1125:58 –72.
2. Schink B, Stams AJM. 2006. Syntrophism among prokaryotes, p 309 –335. In Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E(ed), The prokaryotes, 2nd ed, vol 2. Springer-Verlag, Berlin, Germany.
3. Stams AJM, Plugge CM. 2009. Electron transfer in syntrophic commu-nities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 7:568 –577.
4. Gorby YA, Yanina S, McLean JS, Rosso KM, Moyles D, Dohnalkova A,Beveridge TJ, Chang IS, Kim BH, Kim KS, Culley DE, Reed SB, RomineMF, Saffarini DA, Hill EA, Shi L, Elias DA, Kennedy DW, Pinchuk G,Watanabe K, Ishii S, Logan B, Nealson KH, Fredrickson JK. 2006.Electrically conductive bacterial nanowires produced by Shewanella one-idensis strain MR-1 and other microorganisms. Proc. Natl. Acad. Sci.U. S. A. 103:11358 –11363.
5. McInerney MJ, Sieber JR, Gunsalus RP. 2009. Syntrophy in anaerobicglobal carbon cycles. Curr. Opin. Biotechnol. 20:623– 632.
6. Müller N, Worm P, Schink B, Stams AJM, Plugge CM. 2010. Syntrophicbutyrate and propionate oxidation processes: from genomes to reactionmechanisms. Environ. Microbiol. Rep. 2:489 – 499.
7. Boone DR, Johnson Liu Y. 1989. Diffusion of the interspecies electroncarriers H2 and formate in methanogenic ecosystems and its implicationsin the measurement of Km for H2 or formate uptake. Appl. Environ. Mi-crobiol. 55:1735–1741.
8. Thiele JH, Zeikus JG. 1988. Control of interspecies electron flow duringanaerobic digestion: significance of formate transfer versus hydrogentransfer during syntrophic methanogenesis in flocs. Appl. Environ. Mi-crobiol. 54:20 –29.
9. Stams AJM, Dong X. 1995. Role of formate and hydrogen in the degra-dation of propionate and butyrate by defined suspended cocultures ofacetogenic and methanogenic bacteria. Antonie Van Leeuwenhoek 68:281–284.
10. Worm P, Stams AJM, Cheng X, Plugge CM. 2011. Growth- and sub-strate-dependent transcription of formate dehydrogenase and hydroge-nase coding genes in Syntrophobacter fumaroxidans and Methanospirillumhungatei. Microbiology 157:280 –289.
11. de Bok FAM, Luijten MLGC, Stams AJM. 2002. Biochemical evidencefor formate transfer in syntrophic propionate-oxidizing cocultures of Syn-trophobacter fumaroxidans and Methanospirillum hungatei. Appl. Environ.Microbiol. 68:4247– 4252.
12. Thauer RK, Stackebrandt E, Hamilton WA. 2007. Energy metabolismand phylogenetic diversity of sulphate-reducing bacteria, p 1–37. InBarton LL, Hamilton WA (ed), Sulphate-reducing bacteria: environ-
Meyer et al.
1002 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

mental and engineered systems, 1st ed. Cambridge University Press,New York, NY.
13. Ishii S, Kosaka T, Hotta Y, Watanabe K. 2006. Simulating the contri-bution of coaggregation to interspecies hydrogen fluxes in syntrophicmethanogenic consortia. Appl. Environ. Microbiol. 72:5093–5096.
14. Ishii S, Kosaka T, Hori K, Hotta Y, Watanabe K. 2005. Coaggregationfacilitates interspecies hydrogen transfer between Pelotomaculum thermo-propionicum and Methanothermobacter thermautotrophicus. Appl. Envi-ron. Microbiol. 71:7838 –7845.
15. Shimoyama T, Kato S, Ishii S, Watanabe K. 2009. Flagellum mediatessymbiosis. Science 323:1574.
16. Widdel F, Bak F. 1992. Gram-negative mesophilic sulfate-reducing bac-teria, p 3352–3378. In Balows A, Trüper HG, Dworkin M, Harder W,Schleifer K.-H (ed), The prokaryotes, 2nd ed, vol 4. Springer-Verlag, NewYork, NY.
17. Walker CB, He Z, Yang ZK, Ringbauer JA, He Q, Zhou J, Voordouw G,Wall JD, Arkin AP, Hazen TC, Stolyar S, Stahl DA. 2009. The electrontransfer system of syntrophically grown Desulfovibrio vulgaris. J. Bacteriol.191:5793–5801.
18. Plugge CM, Scholten JCM, Culley DE, Nie L, Brockman FJ, Zhang W.2010. Global transcriptomics analysis of the Desulfovibrio vulgaris changefrom syntrophic growth with Methanosarcina barkeri to sulfidogenic me-tabolism. Microbiology 156:2746 –2756.
19. Li X, McInerney MJ, Stahl DA, Krumholz LR. 2011. Metabolism of H2
by Desulfovibrio alaskensis G20 during syntrophic growth on lactate. Mi-crobiology 157:2912–2921.
20. Stolyar S, van Dien S, Hillesland KL, Pinel N, Lie TJ, Leigh JA, StahlDA. 2007. Metabolic modeling of a mutualistic microbial community.Mol. Syst. Biol. 3:92. doi:10.1038/msb4100131.
21. Pereira IAC, Haveman SA, Voordouw G. 2007. Biochemical, genetic andgenomic characterization of anaerobic electron transport pathways in sul-phate-reducing delta-proteobacteria, p 215–240. In Barton LL, Hamilton,WAA (ed), Sulphate-reducing bacteria: environmental and engineeredsystems. Cambridge University Press, Cambridge, United Kingdom.
22. Pereira IAC, Ramos AR, Grein F, Marques MC, Marques da Silva S,Venceslau SS. 2011. A comparative genomic analysis of energy metabo-lism in sulfate reducing bacteria and archaea. Frontiers Microbiol. 2:69.doi:10.3389/fmicb.2011.00069.
23. Dehal PS, Joachimiak MP, Price MN, Bates JT, Baumohl JK, Chivian D,Friedland GD, Huang KH, Keller K. 2010. MicrobesOnline: an inte-grated portal for comparative and functional genomics. Nucleic Acids Res.38:D396 –D400. doi:10.1093/nar/gkp919.
24. Price MN, Arkin AP, Alm EJ. 2006. OpWise: operons aid the identifica-tion of differentially expressed genes in bacterial microarray experiments.BMC Bioinformatics 7:19. doi:10.1186/1471-2105-7-19.
25. Oh J, Fung E, Price MN, Dehal PS, Davis RW, Giaever G, Nislow C,Arkin AP, Deutschbauer AM. 2010. A universal TagModule collectionfor parallel genetic analysis of microorganisms. Nucleic Acids Res. 38:e146. doi:10.1093/nar/gkq419.
26. Wall JD, Arkin AP, Rapp-Giles B. 2008. Genetics and genomics of sulfaterespiration in Desulfovibrio, p 1–12. In Dahl C, Friedrich, CG. (ed), Mi-crobial sulfur metabolism. Springer-Verlag, Heidelberg, Germany.
27. Pinchuk GE, Rodiono DA, Yang C, Li X, Osterman A, Dervyn E,Geydebrekht OV, Reed SB, Romine MF, Collart FR, Scott JH, Fredrick-son JK, Beliaev AS. 2009. Genomic reconstruction of Shewanella oneiden-sis MR-1 metabolism reveals a previously uncharacterized machinery forlactate utilization. Proc. Natl. Acad. Sci. U. S. A. 106:2874 –2879.
28. Venceslau SS, Lino RR, Pereira IAC. 2010. The Qrc membrane complex,related to the alternative complex III, is a menaquinone reductase in-volved in sulfate respiration. J. Biol. Chem. 285:22774 –22783.
29. Pires RH, Lourenco AI, Morais F, Teixeira M, Xavier AV, SaraivaLM, Pereira IAC. 2003. A novel membrane-bound respiratory com-plex from Desulfovibrio desulfuricans ATCC 27774. Biochim. Biophys.Acta 1605:67– 82.
30. Rossi M, Pollock WBR, Reij MW, Keon RG, Fu RD, Voordouw G. 1993.The hmc operon of Desulfovibrio vulgaris subsp. vulgaris Hildenboroughencodes a potential transmembrane redox protein complex. J. Bacteriol.175:4699 – 4711.
31. Rodionov DA, Dubchak I, Arkin A, Alm E, Gelfand MS. 2004. Recon-struction of regulatory and metabolic pathways in metal-reducing Delta-proteobacteria. Genome Biol. 5:R90. doi:10.1186/gb-2004-5-11-r90.
32. Leang C, Krushkal J, Ueki T, Puljic M, Sun J, Juarez K, Nunez C,Reguera G, DiDonato R, Postier B, Adkins RM, Lovley DR. 2009.
Genome-wide analysis of the RpoN regulon in Geobacter sulfurreducens.BMC Genomics 10:331. doi:10.1186/1471-2164-10-331.
33. Bender KS, Yen H-C, Hemme CL, Yang Z, He Z, He Q, Zhou J, HuangKH, Alm EJ, Hazen TC, Arkin AP, Wall JD. 2007. Analysis of ferricuptake regulator (Fur) mutant of Desulfovibrio vulgaris Hildenborough.Appl. Environ. Microbiol. 73:5389 –5400.
34. Ravcheev DA, Li X, Latif H, Zengler K, Leyn SA, Korostelev YD,Kazakov AE, Novichkov PS, Osterman AL, Rodionov DA. 2012. Tran-scriptional regulation of central carbon and energy metabolism in bacteriaby redox-responsive repressor Rex. J. Bacteriol. 194:1145–1157.
35. Zane GM, Yen HB, Wall JD. 2010. Effect of the deletion of qmoABC andthe promoter-distal gene encoding a hypothetical protein on sulfate re-duction in Desulfovibrio vulgaris Hildenborough. Appl. Environ. Micro-biol. 76:5500 –5509.
36. Nunez MF, Pellicer M-T, Badia J, Aguilar J, Baldoma L. 2001. The geneyghK linked to the glc operon of Escherichia coli encodes a permease forglycolate that is structurally and functionally similar to L-lactate permease.Microbiology 147(Pt 4):1069 –1077.
37. Nunez MF, Kwon O, Wilson TH, Aguilar J, Baldoma L, Lin ECC. 2002.Transport of L-lactate, D-lactate, and glycolate by the LldP and GlcA mem-brane carriers of Escherichia coli. Biochem. Biophys. Res. Commun. 290:824 – 829.
38. Gimenez R, Nunez MF, Badia J, Aguilar J, Baldoma L. 2003. The geneyjcG, cotranscribed with the gene acs, encodes an acetate permease inEscherichia coli. J. Bacteriol. 185:6448 – 6455.
39. Hosie AHF, Allaway D, Poole PS. 2002. A monocarboxylate permease ofRhizobium leguminosarum is the first member of a new subfamily of trans-porters. J. Bacteriol. 184:5436 –5448.
40. Dym O, Pratt EA, Ho C, Eisenberg D. 2000. The crystal structure ofD-lactate dehydrogenase, a peripheral membrane respiratory enzyme.Proc. Natl. Acad. Sci. U. S. A. 97:9413–9418.
41. Lord JM. 1972. Glycolate oxidoreductase in Escherichia coli. Biochim.Biophys. Acta 267:227–237.
42. Pellicer M-T, Badia J, Aguilar J, Baldoma L. 1996. glc locus of Escherichiacoli: characterization of genes encoding the subunits of glycolate oxidaseand the glc regulator protein. J. Bacteriol. 178:2051–2059.
43. Hedderich R, Hamann N, Bennati M. 2005. Heterodisulfide reductasefrom methanogenic archaea: a new catalytic role for an iron-sulfur cluster.Biol. Chem. 386:961–970.
44. Thauer RK, Kaster AK, Seedorf H, Buckel W, Hedderich R. 2008.Methanogenic archaea: ecologically relevant differences in energy conser-vation. Nat. Rev. Microbiol. 6:579 –591.
45. Mander GJ, Weiss MS, Hedderich R, Kahnt J, Ermler U, Warkentin E.2005. X-ray structure of the gamma-subunit of a dissimilatory sulfite re-ductase: fixed and flexible C-terminal arms. FEBS Lett. 579:4600 – 4604.
46. Oliveira TF, Vonrhein C, Matias PM, Venceslau SS, Pereira IAC. 2008.The crystal structure of Desulfovibrio vulgaris dissimilatory sulfite reduc-tase bound to DsrC provides novel insights into the mechanism of sulfaterespiration. J. Biol. Chem. 283:34141–34149.
47. Czechowski M, Rossmoore HW. 1990. Purification and partial charac-terization of a D(�)-lactate dehydrogenase from Desulfovibrio desulfuri-cans (ATCC 7757). J. Ind. Microbiol. 6:117–122.
48. Hansen TA. 1994. NAD(P)-independent lactate dehydrogenase from sul-fate-reducing prokaryotes. Methods Enzymol. 243:21–23.
49. Ogata M, Arihara K, Yagi T. 1981. D-Lactate dehydrogenase of Desulfo-vibrio vulgaris. J. Biochem. 89:1423–1431.
50. Pankhania IP, Spormann AM, Hamilton WA, Thauer RK. 1988. Lactateconversion to acetate, CO2 and H2 in cell suspensions of Desulfovibriovulgaris (Marburg): indications for the involvement of an energy drivenreaction. Arch. Microbiol. 150:26 –31.
51. Stams AJM, Hansen TA. 1982. Oxygen-labile L(�) lactate dehydroge-nase activity in Desulfovibrio desulfuricans. FEMS Microbiol. Lett. 13:389 –394.
52. Chabriere E, Charon M-H, Volbeda A, Pieulle L, Hatchikian EC,Fontecilla-Camps J-C. 1999. Crystal structures of the key anaerobic en-zyme pyruvate:ferredoxin oxidoreductase, free and in complex with pyru-vate. Nat. Struct. Biol. 6:182–190.
53. Pieulle L, Charon M-H, Bianco P, Bonicel J, Petillot Y, Hatchikian EC.1999. Structural and kinetic studies of the pyruvate-ferredoxin oxi-doreductase/ferredoxin complex from Desulfovibrio africanus. Eur. J.Biochem. 264:500 –508.
54. Pieulle L, Guigliarelli B, Asso M, Dole F, Bernadec A, Hatchikian EC.1995. Isolation and characterization of the pyruvate-ferredoxin oxi-
Syntrophic Metabolism of Desulfovibrio alaskensis G20
March 2013 Volume 195 Number 5 jb.asm.org 1003
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

doreductase from sulfate-reducing bacterium Desulfovibrio africanus.Biochim. Biophys. Acta 1250:49 –59.
55. Li X, Krumholz LR. 2007. Regulation of arsenate resistance in Desulfo-vibrio desulfuricans G20 by an arsRBCC operon and an arsC gene. J. Bac-teriol. 189:3705–3711.
56. Ramos AR, Keller KL, Wall JD, Pereira IAC. 2012. The membraneQmoABC complex interacts directly with the dissimilatory adenosine 5=-phosphosulfate reductase in sulfate reducing bacteria. Front. Microbiol.3:137. doi:10.3389/fmicb.2012.00137.
57. Haveman SA, Brunelle JK, Voordouw G, Voordouw JK, Heidelberg JF,Rabus R. 2003. Gene expression analysis of energy metabolism mutants ofDesulfovibrio vulgaris Hildenborough indicates an important role for al-cohol dehydrogenase. J. Bacteriol. 185:4345– 4353.
58. Nitschke W, Russell MJ. 2011. Redox bifurcations: Mechanisms andimportance to life now, and at its origin. Bioessays 34:106 –109.
59. Kaster A-K, Moll J, Parey K, Thauer RK. 2011. Coupling of ferredoxinand heterodisulfide reduction via electron bifurcation in hydro-genotrophic methanogenic archaea. Proc. Natl. Acad. Sci. U. S. A. 108:2981–2986.
60. Costa KC, Wong PM, Lie T, Dodsworth TJ, Swanson JA, Burn I,Hackett JA, Leigh JA. 2010. Protein complexing in a methanogen suggestselectron bifurcation and electron delivery from formate to heterodisulfidereductase. Proc. Natl. Acad. Sci. U. S. A. 107:11050 –11055.
61. Meyer B, Kuever J. 2008. Homology modeling of dissimilatory APSreductases (AprBA) of sulfur-oxidizing and sulfate-reducing prokaryotes.PLoS One 3:e1514. doi:10.1371/journal.pone.0001514.
62. Li X, Luo Q, Wofford NQ, Keller KL, McInerney MJ, Wall JD, Krum-holz LR. 2009. A molybdopterin oxidoreductase is involved in H2 oxida-tion in Desulfovibrio desulfuricans G20. J. Bacteriol. 191:2675–2682.
63. Simon J, van Spanning RJM, Richardson DJ. 2008. The organisation ofproton motive and non-proton motive redox loops in prokaryotic respi-ratory systems. Biochim. Biophys. Acta 1777:1480 –1490.
64. Caffrey SM, Park H-S, Voordouw JK, He Z, Zhou J, Voordouw G. 2007.Function of periplasmic hydrogenases in the sulfate-reducing bacteriumDesulfovibrio vulgaris Hildenborough. J. Bacteriol. 189:6159 – 6167.
65. Valente FMA, Almeida CC, Pacheco I, Carita J, Saraiva LM, PereiraIAC. 2006. Selenium is involved in regulation of periplasmic hydrogenasegene expression in Desulfovibrio vulgaris Hildenborough. J. Bacteriol. 188:3228 –3235.
66. Pohorelic BKJ, Voordouw JK, Lojou E, Dolla A, Harder J, Voordouw G.2002. Effects of deletion of genes encoding Fe-only hydrogenase of Desul-fovibrio vulgaris Hildenborough on hydrogen and lactate metabolism. J.Bacteriol. 184:679 – 686.
67. Valente FMA, Oliveira ASF, Gnadt N, Pacheco I, Coelho AV, XavierAV, Teixeira M, Soares CM, Pereira IAC. 2005. Hydrogenases in Desul-fovibrio vulgaris Hildenborough: structural and physiologic characterisa-tion of the membrane-bound (NiFeSe) hydrogenase. J. Biol. Inorg. Chem.10:667– 682.
68. Venceslau SS, Matos D, Pereira IAC. 2011. EPR characterization of the
new Qrc complex from sulfate reducing bacteria and its ability to form asupercomplex with hydrogenase and TpIc3. FEBS Lett. 585:2177–2181.
69. da Silva SM, Pacheco I, Pereira IAC. 2012. Electron transfer betweenperiplasmic formate dehydrogenase and cytochromes c in Desulfovibriodesulfuricans ATCC27774. J. Biol. Inorg. Chem. 17:831– 838.
70. Abreu IA, Lourenco AI, Xavier AV, LeGall J, Coelho AV, Matias PM,Pinto DM, Carrondo MA, Teixeira M, Saraiva LM. 2003. A novel ironcentre in the split-Soret cytochrome c from Desulfovibrio desulfuricansATC27774. J. Biol. Inorg. Chem. 8:360 –370.
71. Mota CS, Valette O, Gonzalez PJ, Brondino CD, Moura JJG, Moura I,Dolla A, Rivas MG. 2011. Effects of molybdate and tungstate on expres-sion levels and biochemical characteristics of formate dehydrogenasesproduced by Desulfovibrio alaskensis NCIMB 13491. J. Bacteriol. 193:2917–2923.
72. da Silva SM, Pimentel C, Valente FMA, Rodrigues-Pousada C, PereiraIAC. 2011. Tungsten and molybdenum regulation of formate dehydroge-nase expression in Desulfovibrio vulgaris Hildenborough. J. Bacteriol. 193:2909 –2916.
73. Dolla A, Pohorelic BKJ, Voordouw JK, Voordouw G. 2000. Deletion ofthe hmc operon of Desulfovibrio vulgaris subsp. vulgaris Hildenboroughhampers hydrogen metabolism and low-redox-potential niche establish-ment. Arch. Microbiol. 174:143–151.
74. Voordouw G. 2002. Carbon monoxide cycling by Desulfovibrio vulgarisHildenborough. J. Bacteriol. 184:5903–5911.
75. Müller FH, Bandeiras TM, Urich T, Teixeira M, Gomes CM, Kletzin A.2004. Coupling of the pathway of sulphur oxidation to dioxygen reduc-tion: characterization of a novel membrane-bound thiosulphate:quinoneoxidoreductase. Mol. Microbiol. 53:1147–1160.
76. Cipollone R, Ascenzi P, Visca P. 2007. Common themes and variationsin the rhodanese superfamily. IUBMB Life 59:51–59.
77. Xu XM, Moeller SG. 2011. Iron-sulfur clusters: biogenesis, molecularmechanisms, and their functional significance. Antioxid. Redox Signal.15:271–307.
78. Rajeev L, Luning EG, Dehal PS, Price MN, Arkin AP, MukhopadhyayA. 2011. Systematic mapping of two component response regulators togene targets in a model sulfate reducing bacterium. Genome Biol. 12:R99.doi:10.1186/gb-2011-12-10-r99.
79. Mukherjee S, Yakhnin H, Kysela D, Sokoloski J, Babitzke P, Kearns DB.2011. CsrA-FliW interaction governs flagellin homeostasis and a check-point on flagellar morphogenesis in Bacillus subtilis. Mol. Microbiol. 82:447– 461.
80. Alexandre G. 2010. Coupling metabolism and chemotaxis-dependentbehaviours by energy taxis receptors. Microbiology 156:2283–2293.
81. Porter SL, Wadhams GH, Armitage JP. 2011. Signal processing in com-plex chemotaxis pathways. Nat. Rev. Microbiol. 9:153–165.
82. Timmermans J, van Melderen L. 2010. Post-transcriptional global reg-ulation by CsrA in bacteria. Cell. Mol. Life Sci. 67:2897–2908.
83. Keller KL, Wall JD. 2011. Genetics and molecular biology of the electronflow for sulfate respiration in Desulfovibrio. Front. Microbiol. 2:135.
Meyer et al.
1004 jb.asm.org Journal of Bacteriology
on August 20, 2018 by guest
http://jb.asm.org/
Dow
nloaded from














![Molecular Dynamics Simulation of Bacterial [FeNi]-Hydrogenase Found in Desulfovibrio fructosovorans Solvated in a Water Sphere Introduction Hydrogenase](https://img.pdfslide.us/doc/110x75/56649f1a5503460f94c2fabc/molecular-dynamics-simulation-of-bacterial-feni-hydrogenase-found-in-desulfovibrio.jpg)





![Diverse hydrogen production and consumption pathways … · ing dissimilatory sulfate reduction (e.g., Desulfovibrio desulfuricans)[50, 51], fumarate and nitrate reduction (e.g.,](https://img.pdfslide.us/doc/110x75/5f478d2813f91a6e9825a474/diverse-hydrogen-production-and-consumption-pathways-ing-dissimilatory-sulfate-reduction.jpg)







