Embed Size (px)
Citation preview

PertanikaJ. Trap. Agric. Sci. 21(2): 113 -122 (1998) ISSN: 1511-3701© Universiti Puu"a Malaysia Press
Variability in Eggplant (Solanum melongena L.) and Its Nearest Wild Speciesas Revealed by Polyacrylamide Gel Electrophoresis of Seed Protein
SAYED M. ZAI HASAN and MIMI LINDA ISAIFaculty of Science and Professional ArtUniversiti Putra Malaysia Terengganu
21030 Mengabang Telipot, Kuala Terengganu, Malaysia
IFaculty of Agriculture,Universiti Putra Malaysia,
43400 UPM Serdang, Selangor, Malaysia
Keywords: Solanum melongena L., SDS-PAGE, seed protein, variability
ABSTRAK
Satu kajian telah dilakukan untuk menentukan dan membezakan genotip-genotip terong (Solanum melongenaL.) dan spesis-spesis liar terdekatnya dengan elektroforesis gel poliakrilamid natrium dodesilsulfat (SDS-PAGE)keatas protin biji boleh larut. Lima puloh empat aksesi kumpulan-kumpulan tanaman dan spesis liar terongtelah digunakan untuk analisis. Hasil dari corak penjaluran elektroforetik menunjukan bahawa terdapat satuvariasi yang besar diantara dan didalam kumpulan-kumpulan terong dari segi bilangan, saiz, kedudukan,keamatan warna dan hadir atau tak hadir jalur-jalur protin didalam profil, yang mana boleh digunakan untukpencaman dan pencirian kultivar-kultivar terong. Kultivar terong jenis buah panjang telah menunjukkanbanyak jalur-jalur khusus-sepunya yang memisahkanya dari kumpulan-kumpulan lain. Manakala, satukevariabelan besar profil protin biji terdapat didalam kultivar-kultivar jenis buah bulat dan primitif dankumpulan-kumpulan rumpai dan liar terong, termasuk yang paling liar dari S. incanum, yang menunjukanbahawa kumpulan-kumpulan ini mengandungi pelbagai genotip yang berbeza.
ABSTRACT
A study was undertaken to determine and distinguish the genotypes of eggplant (Solanum melongena L.) andits nearest wild species by sodium dodecylsulphate polyacrylamide gel electrophoresis (SDS-PAGE) of soluble seedproteins. Fiftyfour accessions of cultivated and wild groups of eggplant were subjected to the analysis. The resultsof electrophoretic banding patterns show that there was a great deal of variation within and between groups ofeggplant in terms of numbers, sizes, positions, staining intensities and presence or absence of protein bands inthe profile, which can be used for characterization and identification of eggplant cultivars. Elongated fruiteggplant cultivars exhibited nine common group-specific bands which distinguish them from the other groups.Meanwhile, a great variability in seed protein profiles was detected in the round fruit type and primitive cultivars,weedy and wild groups ofeggplant including the truly wild S. incanum, which indicated that these groups consistof diverse genotypes.
INTRODUCTION
Solanum melongena L. (eggplant) has a wide rangeof morphological diversity ranging from wildand weedy to semi- or fully-cultivated forms.Many characters are overlapping from one groupto another group, which makes it difficult todistinguish between and within groups (Lesterand Hasan 1991; Karihaloo and Rai 1995).
Horticulturally, it is important to be able torecognize and distinguish the different genotypes
• and, in particular, different cultivars of this speciesbecause the cultivars often differ in their fruitquality and other important agronomic characterswhich are obscure and difficult to detect.
Morphological and physiological charactershave traditionally been used for the identification

VARIABILITY I EGGPlANT (SOLANUM MELONGENA L.)
of eggplant cultivars (Brezhnev 1958). Many ofthese characters, particularly leaf and stempigmentation and fruit size, shape and colour,which are often important in the definition ofan eggplant cultivar, can be discerned only inadult plants. Thus, it is necessary to grow outplants, which is very laborious and timeconsuming. Furthermore, the phenotypiccharacters are strongly influenced by climaticfactors and soil conditions, and their use maylead to unreliable or erroneous determinations.
These problems can be avoided by usingseed storage protein electrophoresis becauseprotein banding patterns produced through thistechnique are generally unaffected by externalfactors and the genotype of each cultivar can berapidly determined. The seed storage proteinsare also known to be highly polymorphic withrespect to their size and charge in almost allspecies investigated (Cooke 1984). Hence, theyprovide sufficient variability in protein compositionto detect differences between genotypes.Furthermore, the seed protein profiles are knownfor their stability, uniformity and additive nature(Smith 1976). For these reasons, SDS-PAGE ofseed proteins is used as a method for cultivaridentification in many cultivated species (Sathaiahand Reddy 1985; Gupta and Robbelen 1986;Gardiner and Forde 1988; Huaman andStegemann 1989; Rao et al1990; Stegemann et. al1992; Yupsanis et. al 1992; Wang et. al 1994).However, this method has not been widely usedfor the identification of cultivars and varieties ofmany vegetables, particularly the eggplant.
To date, seed protein SDS-PAGE of S.melongena had been conducted as part of ataxonomic study of the genus Solanum (Edmondsand Glidewell 1977; Pearce and Lester 1979) butthere was no serious attempt to determinedifferences between cultivars of eggplant on thebasis of protein profiles. The objectives of thisstudy were to determine seed protein bandingpatterns for 54 accessions of eggplants and toascertain the level of protein variation presentin the species as well as to determine thepossibility of using SDS-PAGE of seed proteinsfor cultivar identification and characterizationin eggplants.
MATERIAL AND METHODS
Seed Materials
The seeds of 54 accessions of eggplant complexrepresenting a range of geographical origins
and morphological varieties were used (Table1). They were grouped as i) most advancedcultivars with elongated large fruit (fruit lengththree times width), ii) advanc~d cultivars withround fruit (fruit either round, obovate or pearshape) with diameter greater than 5 cm, iii)primitive cultivars with small fruit (less than 5cm diameter), iv) prickly plant of weedy (S.cumingii Dun.) or wild (S. insanum L.) groupsand v) the truly wild relative of S. incanum L.group (Lester and Hasan 1990).
Preparation of Seed Flour
Dry seeds (0.5 g) were dehulled using a microFeinmuhle-Culatti sample mill, followed byseparation through a Hearson's blower to obtainclean seeds. The seeds were ground in a microdisembrator for 10 - 15 min to produce very fine
. seed flour, which was then defatted in fivechanges of diethyl ether and allowed to dry onfilter paper.
Protein Extraction
Soluble proteins were extracted by soaking 0.05g of defatted protein flour in 0.5 ml H
20 in a
centrifuge vial. The suspension was centrifugedand 100 III of protein solution (supernatant) wasincubated with 100 III Tris-HCl buffer (pH =6.8) consisting of 20% glycerol and 5 mMmercaptoethanol. 0.1 ml of bromophenol blue(WIV) was added into the samples as a trackingdye in order to mark the moving protein front.
Gel Preparation
The crude protein was fractionated by themethod of SDs-discontinuous polyacrylamide slabgel electrophoresis (SDS-PAGE) in a verticalformat in a Studier-type slab gel apparatus. Theseparations were performed on 2.5% stackinggel and 12.5% separation gel consisting of 10%sodium dodecyl sulphate (SDS) and Tris-HCl atpH of 6.8 and 8.8 respectively. Fifteen wells weremade in the stacking gel by inserting a combinto the gel before polymerization. Ammoniumpersulphate (10%) was used to initiate thepolymerization and N ,N,N' , tetramethylethylenediamine (TEMED) was used as a catalyst(Hames and Rickwood 1986).
Electrophoresis
The 20-lll sample solution of each accession wasloaded in the well. Sample solutions of twu
114 PERTANlKAJ. TRap. AGRIC. SCI. VOL. 21 0.2,1998
SAYED M. ZAIN HASAN AND MIMI LINDA ISA
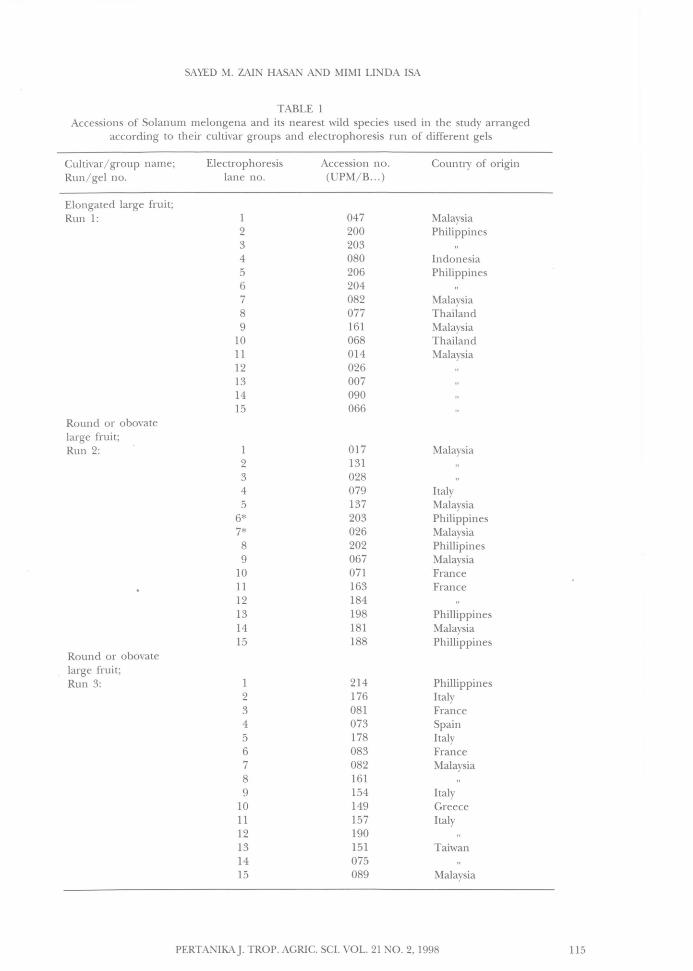
TABLE 1Accessions of Solanum melongena and its nearest wild species used in the study arranged
according to their cultivar groups and electrophoresis run of different gels
CUltivar/ group name; Electrophoresis Accession no. Country of originRun/gel no. lane no. (UPM/B... )
Elongated large fruit;Run 1: 1 047 Malaysia
2 200 Philippines3 2034 080 Indonesia5 206 Philippines6 2047 082 Malaysia8 077 Thailand9 161 Malaysia
10 068 Thailand11 014 Malaysia12 02613 00714 09015 066
Round or obovatelarge fruit;Run 2: 1 017 Malaysia
2 1313 0284 079 Italy5 137 Malaysia
6* 203 Philippines7* 026 Malaysia8 202 Phillipines9 067 Malaysia
10 071 France11 163 France12 18413 198 Phillippines14 181 Malaysia15 188 Phillippines
Round or obovatelarge frui t;Run 3: 1 214 Phillippines
2 176 Italy3 081 France4 073 Spain5 178 Italy6 083 France7 082 Malaysia8 1619 154 Italy
10 149 Greece11 157 Italy12 19013 151 Taiwan14 07515 089 Malaysia
PERT lKA]. TRap. AGRIC. SCI. VOL. 21 0.2,1998 115
VARIABILITY IN EGGPLANT (SOLANUM MELONGENA L.)
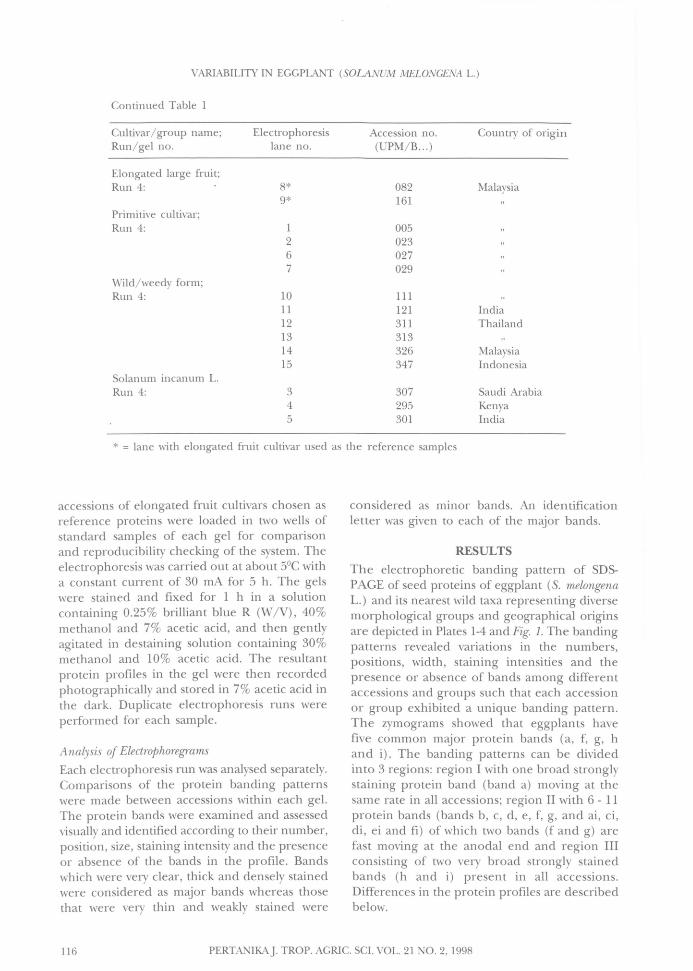
Continued Table 1
Cultivar/ group name;Run/gel no.
Elongated large fruit;Run 4:
Primitive cultivar;Run 4:
Wild/weedy form;Run 4:
Solanum incanum L.Run 4:
Electrophoresis Accession no.lane no. (UPM/B... )
8* 0829* 161
1 0052 0236 0277 029
10 11111 12112 31113 31314 32615 347
3 3074 2955 301
Country of origin
Malaysia
IndiaThailand
MalaysiaIndonesia
Saudi ArabiaKenyaIndia
* = lane with elongated fruit cultivar used as the reference samples
accessions of elongated fruit cultivars chosen asreference proteins were loaded in two wells ofstandard samples of each gel for comparisonand reproducibility checking of the system. Theelectrophoresis was carried out at about 5°C witha constant current of 30 rnA for 5 h. The gelswere stained and fixed for 1 h in a solutioncontaining 0.25% brilliant blue R (WIV), 40%methanol and 7% acetic acid, and then gentlyagitated in destaining solution containing 30%methanol and 10% acetic acid. The resultantprotein profiles in the gel were then recordedphotographically and stored in 7% acetic acid inthe dark. Duplicate electrophoresis runs wereperformed for each sample.
Analysis of Electrophoregrams
Each electrophoresis run was analysed separately.Comparisons of the protein banding patternswere made between accessions within each gel.The protein bands were examined and assessedvisually and identified according to their number,position, size, staining intensity and the presenceor absence of the bands in the profile. Bandswhich were very clear, thick and densely stainedwere considered as major bands whereas thosethat were very thin and weakly stained were
considered as minor bands. An identificationletter was given to each of the major bands.
RESULTS
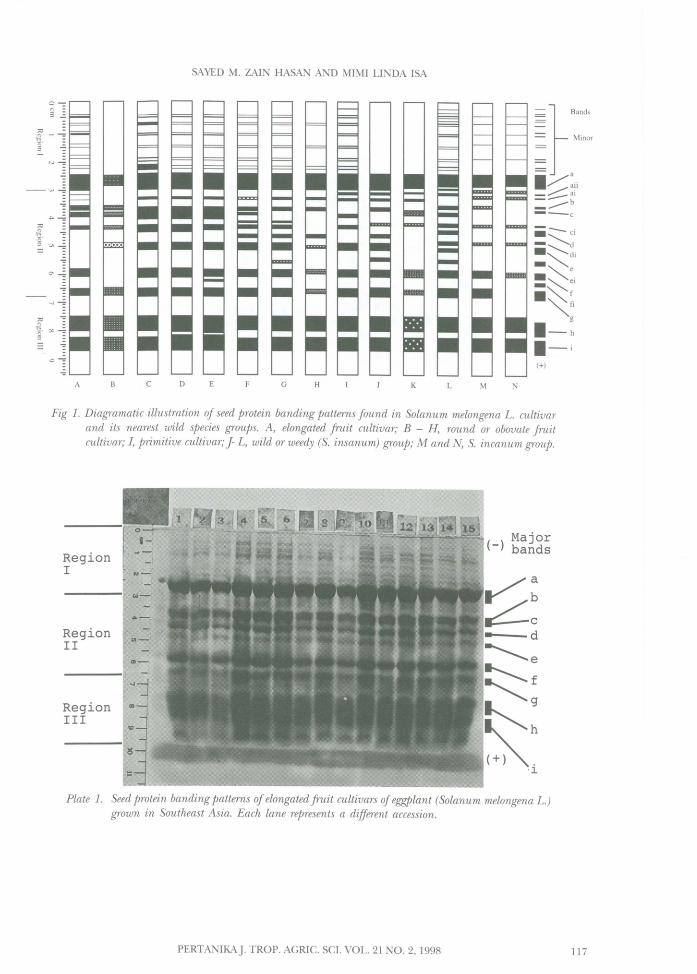
The electrophoretic banding pattern of SDSPAGE of seed proteins of eggplant (S. melongenaL.) and its nearest wild taxa representing diversemorphological groups and geographical originsare depicted in Plates 1-4 and Fig. 1. The bandingpatterns revealed variations in the numbers,positions, width, staining intensities and thepresence or absence of bands among differentaccessions and groups such that each accessionor group exhibited a unique banding pattern.The zymograms showed that eggplants havefive common major protein bands (a, f, g, hand i). The banding patterns can be dividedinto 3 regions: region I with one broad stronglystaining protein band (band a) moving at thesame rate in all accessions; region II with 6 - 11protein bands (bands b, c, d, e, f, g, and ai, ci,di, ei and fi) of which two bands (f and g) arefast moving at the anodal end and region IIIconsisting of two very broad strongly stainedbands (h and i) present in all accessions.Differences in the protein profiles are describedbelow.
116 PERTANIKAJ. TRap. AGRIe. SCI. VOL. 21 NO.2, 1998
SAYED M. ZAIN HAS AND MIMI LI DA ISA
=}BandS,M;""'
.~aii=:---~l-------c
(+)
A B c D E F G H K L M N
Fig 1. Diagramatic illustration of seed protein banding patterns found in Solanum melongena L. cultivarand its nearest wild species groups. A, elongated fruit cultivar; B - H, round or obovate fruitcultivar; I, primitive cultivar; j- L, wild or weedy (s. insanum) group; M and N, S. incanum group.
RegionI
RegionII
RegionIII
( ) Major- bands
Plate 1. Seed protein banding patterns ofelongated fruit cultivars ofeggplant (Solanum melongena L.)grown in Southeast Asia. Each lane represents a different accession.
PERTANIKAJ. TRap. ACRIC. SCI. VOL. 21 0.2,1998 117
RegionI
RegionII
RegionIII
VARIABILITY I EGGPlANT (SOLANUM MELONGENA L.)
Majorbands
~1:~d1-
e
e1
ff1
gh
i
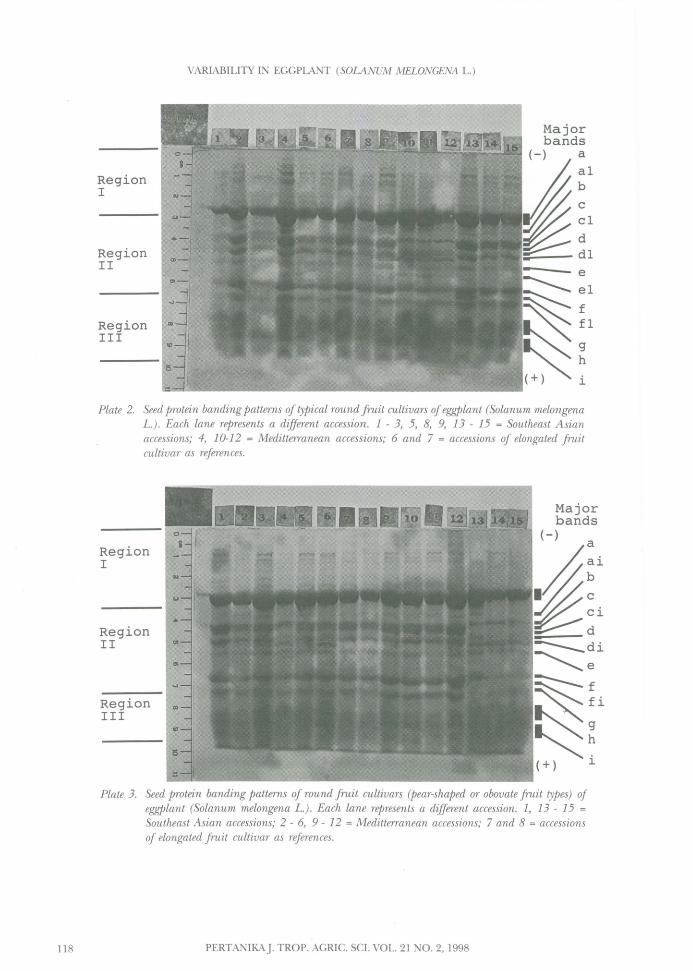
Plate 2. Seed protein banding patterns of typical round fruit cultivars of eggplant (Solanum melongenaL.). Each lane represents a different accession. 1 - 3, 5, 8, 9, 13 - 15 = Southeast Asianaccessions; 4, 10-12 = Meditterranean accessions; 6 and 7 = accessions of elongated fruitcultivar as references.
RegionI
RegionII
RegionIII
118
Plate 3. Seed protein banding patterns of round fruit cultivars (pear-shaped or obovate fruit types) ofeggplant (Solanum melongena L.). Each lane represents a different accession. 1, 13 - 15 =Southeast Asian accessions; 2 - 6, 9 - 12 = Medittermnean accessions; 7 and 8 = accessionsof elongated fruit cultivar as references.
PERT IKA]. TROP. AGRIC. SCI. VOL. 21 0.2,1998
SAYED M. ZAIN HASAN AND MIMI LINDA ISA
RegionI
RegionII
RegionIII
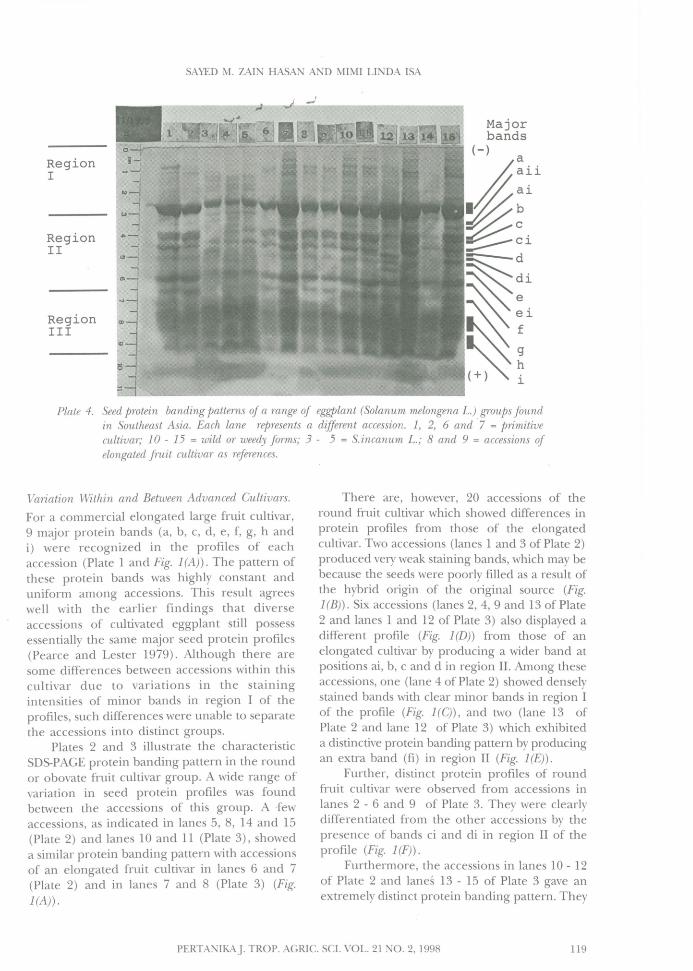
Plate 4. Seed protein banding patterns of a range of eggplant (Solanum melongena L.) groups foundin Southeast Asia. Each lane represents a different accession. 1, 2, 6 and ·7 = primitivecultivar; 10 - 15 = wild or weedy forms; 3 - 5 = S.incanum L.; 8 and 9 = accessions ofelongated fruit cultivar as references.
Variation Within and Between Advanced Cultivars.
For a commercial elongated large fruit cultivar,9 major protein bands (a, b, c, d, e, f, g, handi) were recognized in the profiles of eachaccession (Plate 1 and Fig. 1(A)). The pattern ofthese protein bands was highly constant anduniform among accessions. This result agreeswell with the earlier findings that diverseaccessions of cultivated eggplant still possessessentially the same major seed protein profiles(Pearce and Lester 1979). Although there aresome differences between accessions within thiscultivar due to variations in the stainingintensities of minor bands in region I of theprofiles, such differences were unable to separatethe accessions into distinct groups.
Plates 2 and 3 illustrate the characteristicSDS-PAGE protein banding pattern in the roundor obovate fruit cultivar group. A wide range ofvariation in seed protein profiles was foundbetween the accessions of this group. A .fewaccessions, as indicated in lanes 5, 8, 14 and 15(Plate 2) and lanes 10 and 11 (Plate 3), showeda similar protein banding pattern with accessionsof an elongated fruit cultivar in lanes 6 and 7(Plate 2) and in lanes 7 and 8 (Plate 3) (Fig.
1(A)).
There are, however, 20 accessions of theround fruit cultivar which showed differences inprotein profiles from those of the elongatedcultivar. Two accessions (lanes 1 and 3 of Plate 2)produced very weak staining bands, which may bebecause the seeds were poorly filled as a result ofthe hybrid origin of the original source (Fig.1(B)). Six accessions (lanes 2, 4, 9 and 13 of Plate2 and lanes 1 and 12 of Plate 3) also displayed adifferent profile (Fig. 1(D)) from those of anelongated cultivar by producing a wider band atpositions ai, b, c and d in region II. Among theseaccessions, one (lane 4 of Plate 2) showed denselystained bands with clear minor bands in region Iof the profile (Fig. 1(C)), and two (lane 13 ofPlate 2 and lane 12 of Plate 3) which exhibiteda distinctive protein banding pattern by producingan extra band (fi) in region II (Fig. 1(E)).
Further, distinct protein profiles of roundfruit cultivar were observed from accessions inlanes 2 - 6 and 9 of Plate 3. They were clearlydifferentiated from the other accessions by thepresence of bands ci and di in region II of theprofile (Fig. 1(F)).
Furthermore, the accessions in lanes 10 - 12of Plate 2 and land 13 - 15 of Plate 3 gave anextremely distinct protein banding pattern. They
PERTANIKAJ. TROP. AGRIe. SCI. VOL. 21 NO.2, 1998 119

VARlABILITY IN EGGPLANT (SOLANUM MELONGENA L.)
were easily distinguished from the otheraccessions by the absence of band c in theprofile (Fig. 1(G)). However, accessions in lanes14 and 15 of Plate 3 could be furtherdistinguished from the other accessions by theabsence of bands ci, d and ei in the profile (Fig.1 (H)).
Variation Within and Between Primitive Cultivarsand Wild Groups.
The photoelectrophoregram of seed proteinprofiles for different accessions of primitivecultivar, weedy and wild groups of eggplant isshown in Plate 4. For comparison, control linesof elongated cultivar were in lanes 8 and 9.Three accessions (lanes 1, 2 and 6) of primitivecultivar showed only slight differences in seedprotein profile from those of the elongatedcultivar. One accession (lane 7) of primitivecultivar and three accessions (lanes 10, 11 and13) of the weedy group shared a similar proteinbanding pattern (Fig. 1(1)) but showed somedifferences from those of the elongated cultivarby producing densely stained bands and thepresence of bands ai and aii in the profiles.
One accession (lane 14) of weedy group(Plate 4)) showed a clear difference in proteinprofile from those of the elongated cultivar bythe absence of bands c, d and e in the profile(Fig. 1(K)). One accession in lane 12 of the samegroup also showed the differences by the absenceof bands c and d and the presence of bands ci,di and ei in the profile (Fig. 1(j)). Anotheraccession of the weedy group (lane 15) alsoexhibited a specific protein banding patterndistinct from the other groups by producingbands in all position with thick and denselystained bands, particularly in region I of theprofile (Fig. 1(L)).
The truly wild S. incanum (lanes 3 - 5) wasclearly differentiated from the other groups bythe absence of bands c, d and e and the presenceof bands aii, ai, ci and di in the profile (Fig.1(M)). The accession in lane 3 is even moredistinct from the other accessions by the absence of bands band g in the profile (Fig. 1(N)).
DISCUSSION
The overall differential banding patterns of seedproteins revealed both qualitative andquantitative variations between and within thegroups of eggplant. There are 14 protein bandingpatterns (Fig. 1) characterizing the different types
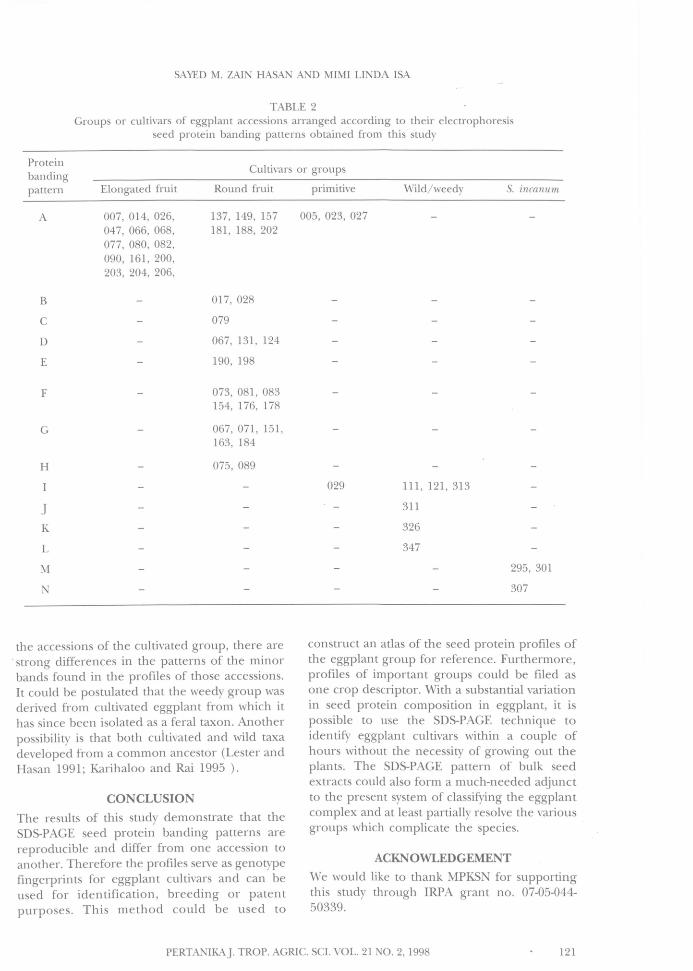
of eggplant used in this investigation. Majorbands which contributed to the variability ofprotein banding patterns were mainly found inregion II of the profile. These made it possibleto divide genotypes of eggplant into differentcultivars or groups (Table 2). These differencesappear reproducible, as similar protein profileswere obtained when further samples of the sameaccessions were analysed in different gels.
It is evident that the elongated fruit cultivarshad the most constant and uniform pattern ofm~or protein bands that gave characteristicgroup-specific seed protein banding patterns,thus enabling them to be distinguished easilyfrom those of the other groups (Fig. 1(A) andTable 2). Although elongated advanced cultivarshave many of the same bands, there are, however,small variations between accessions. Theelongated fruit type is the most common eggplantcultivar grown for commercial purposes. Thus,they may have a common parent in theirpedigrees which could explain the similarity oftheir electrophoregrams. It is common for theagricultu~alcultivar seed protein banding patternto be very stable during commercial seedproduction, but the pattern may alter slightlywhen an accession is multiplied, particularly if itis regenerated from a very small sample size(Gardiner and Forde 1987).
The other groups of eggplant did not showa strong common protein banding pattern, butdisplayed a great deal of variation at the groupslevel (Fig. 1(B-N)). Great variability in seed proteinprofiles was detected in the round fruit cultivarindicating that the cultivar actually consists ofdifferent genotypes. This high variability is notsurprising because round fruit cultivar is anoutbred heterogeneous population whoseaccessions were mostly obtained from smallisolated populations in diverse agro-ecosystems.
Primitive cultivar also showed variable seedprotein banding patterns between differentaccessions, and the protein banding pattern ofthe weedy group, including the truly wild S.incanum, was even more variable. The substantialdifferences in the protein profiles within thesepopulations are mainly due to the outbreedingnature of these taxa (Karihaloo and Rai 1995).
The majority of the accessions used in thisinvestigation could be readily distinguished bytheir soluble seed protein banding patterns.Although in a few cases the weedy accessionsshowed similar major protein compositions to
120 PERT lKA]. TROP. AGRIe. SCI. VOL. 21 0.2,1998
SAYED M. ZAIN HASAN AND MIMI LINDA ISA
TABLE 2Groups or cultivars of eggplant accessions arranged according to their electrophoresis
seed protein banding patterns obtained from this study
Cultivars or groups
Round fruit primitive
Proteinbandingpattern Elongated fruit
A 007, 014, 026,047, 066, 068,077, 080, 082,090, 161, 200,203, 204, 206,
B
C
D
E
F
G
H
JK
L
M
N
137, 149, 157181, 188, 202
017, 028
079
067, 131, 124
190, 198
073, 081, 083154, 176, 178
067, 071, 151,163, 184
075, 089
005, 023, 027
029
Wild/weedy
111, 121, 313
311
326
347
S. incanum
295, 301
307
the accessions of the cultivated group, there are.strong differences in the patterns of the minorbands found in the profiles of those accessions.It could be postulated that the weedy group wasderived from cultivated eggplant from which ithas since been isolated as a feral taxon. Anotherpossibility is that both cuitivated and wild taxadeveloped from a common ancestor (Lester andHasan 1991; Karihaloo and Rai 1995 ).
CONCLUSIONThe results of this study demonstrate that theSDS-PAGE seed protein banding patterns arereproducible and differ from one accession toanother. Therefore the profiles serve as genotypefingerprints for eggplant cultivars and can beused for identification, breeding or patentpurposes. This method could be used to
construct an atlas of the seed protein profiles ofthe eggplant group for reference. Furthermore,profiles of important groups could be filed asone crop descriptor. With a substantial variationin seed protein composition in eggplant, it ispossible to use the SDS-PAGE technique toidentify eggplant cultivars within a couple ofhours without the necessity of growing out theplants. The SDS-PAGE pattern of bulk seedextracts could also form a much-needed adjunctto the present system of classifying the eggplantcomplex and at least partially resolve the variousgroups which complicate the species.
ACKNOWLEDGEMENTWe would like to thank MPKSN for supportingthis study through IRPA grant no. 07-05-04450339.
PERTANIKAJ. TROP. AGRlC. SCI. VOL. 21 TO. 2, 1998 121

VARIABILITY IN EGGPLANT (SOLANUM MELONGENA L.)
REFERENCES
BREZH:\,EY, D.D. 1958. Vegetable plants fam.Solanaceae. In Flora of Cultivated Plants XX, ed.P.M. Zhukovsky, p. 302-324. Leningrad: StateAgri .. [Russian].
COOKE, RJ. 1984. The characterisation and identification of crop cultivars by electrophoresis.Electrophoresis 5: 59-72.
ED~10:\'DS,J.M. and S.M. GLIDEV\'ELL. 1977. Acrylamidegel electrophoresis of seed protein from someSolanum (section Solanum) species. Plant Syst.Evo!. 127: 277-291.
G.-\RDI:\'ER, S.E. and M.B. FORDE. 1988. Identificationof cultivars and species of pasture legumes bysodium dodecylsulphate polyacrylamideelectrophoresis of seed proteins. Plant Varietiesand Seeds 1: 13-26.
GnTA, S.K and G. ROBBELE:\'. 1986. Identificationof rapeseed (Brassica napus) cultivars byelectrophoresis. Z. Pjlanzenzuchtg 96: 363-370
H-\..\IES, B.D. and D. RICKwOOD. 1986. Gel Electrophoresis of Protein. Oxford: IRL Press.
HCA~L-\:\', Z. and H. STEGHL\:\':\'. 1989. Use of electrophoretic analyses to verify morphologicallyidentical clones in a potato collection. PlantVarieties and Seeds 2: 155-161.
K-\RIHALOO, J.L. and M. R-\I. 1995. Significance ofmorphological variability in Solanum insanumL. (sensu lato). Plant Genetic Res. News 103: 24-26.
LESTER, R.X and S.M.Z. H-\.SA:\'. 1990. The distinction between Solanum melongena L. and Solanum incanum L. (Solanaceae). TAXON 39(3):521-523.
LESTER, R.N. and S.M.Z. HAs~'\. 1991. Origin anddomestication of the brinjal egg-plant, Solanummelongena, from S. incanum in Mrica and Asia.
In Solanaceae III. Taxonomy, Chemistry, Evolution, ed. J.G. Hawkes, R. . Lester and A.D.Skelding. p. 369-387. London: Academic Press.
PEARCE, KG. and R.N. LESTER. 1979. Chemotaxonomy of the cultivated eggplant - a new look atthe taxonomic relationships of Solanummelongena L. In The Biology and Taxonomy of theSolanaceae. eds. J.G. Hawkes, R. . Lester andA.D. Skelding. p. 615-628. London: AcademicPress.
RAo, T.N., Y.S. NERKAR and Y.D. PATIL. 1990. Identification of cultivars of cotton by sodiumdodecylsulphate polyacrylamide gelelectrophoresis (SDS-PAGE) of soluble seedproteins. Plant Varieties and Seeds 5: 83-91.
SATHAIAH, Y. and T.P. REDDY. 1985. Seed proteinprofiles of castor (Ricinus communis L.) andsome Jatropha species. Genet. Agr. 39: 35-43.
SMITH, P.M. 1976. The Chemotaxonomy ofPlants. London: Edward Arnold.
STEGHL-\:\':\', H., A.A. SHAH, E. KROGERRECKLE:\'FORTand M.M. H-\..\IZA. 1992. Sweet potato (Ipomoeabatatas L.): genotype identification byelectrophoretic methods and properties of theirproteins. Plant Varieties and Seeds 5: 83-91.
vV.-\..'\G, C., K B1-\..'\, H.X. ZH.-\..'\G, Z.M. ZHOC and J.A.W~'\G. 1994. Polyacrylamide gel electrophoresis of salt soluble protein for maize varietyidentification and genetic purity assessment.Journal Seed Science and Technology 22: 51-57.
Yns.-\..,\Is, T., M. MocsTAKAs and S. KARAKOLI. 1992.Seed protein electrophoresis for varietalidentification in rice (Oy)'za sativa L.). Journalof Agronomy and Crop Science 168(2): 95-99.
(Received 13 August 1996)(Accepted 24 March 1999)
122 PERTANIKAJ. TRap. AGRIC. SCI. VOL. 21 0,T0. 2, 1998






















![Effect of agricultural inputs and essential oils on the ... · spp. [Solanaceae]) y la berenjena (Solanum melongena L. [Solanaceae]) hacen una especial contribución a la seguridad](https://img.pdfslide.us/doc/110x75/5e88e70fc04b8a293e2a7345/effect-of-agricultural-inputs-and-essential-oils-on-the-spp-solanaceae-y.jpg)






