Embed Size (px)
Citation preview

Review
10.1586/14760584.4.1.89 © 2005 Future Drugs Ltd. ISSN 1476-0584 89
CONTENTS
Aquaculture: agriculture of the oceans
Vaccines & disease control
Administration methods & factors affecting efficacy
Limitations in vaccine development
Bacterial fish vaccines
Viral fish vaccines
Fish vaccines against parasites
Expert opinion & five-year view
Key issues
References
Affiliations
www.future-drugs.com
Vaccines for fish in aquaculture
Ingunn Sommerset†, Bjørn Krossøy, Eirik Biering and Petter Frost
†Author for correspondenceIntervet Norbio AS, Thormøhlensgate 55,N-5008 Bergen, NorwayTel.: +47 55 543 958Fax: +47 55 960 [email protected]
KEYWORDS: aquaculture, bacteria, disease, fish, immunology, parasites, salmon, vaccination, vaccine, virus
Vaccination plays an important role in large-scale commercial fish farming and has been a key reason for the success of salmon cultivation. In addition to salmon and trout, commercial vaccines are available for channel catfish, European seabass and seabream, Japanese amberjack and yellowtail, tilapia and Atlantic cod. In general, empirically developed vaccines based on inactivated bacterial pathogens have proven to be very efficacious in fish. Fewer commercially available viral vaccines and no parasite vaccines exist. Substantial efficacy data are available for new fish vaccines and advanced technology has been implemented. However, before such vaccines can be successfully commercialized, several hurdles have to be overcome regarding the production of cheap but effective antigens and adjuvants, while bearing in mind environmental and associated regulatory concerns (e.g., those that limit the use of live vaccines). Pharmaceutical companies have performed a considerable amount of research on fish vaccines, however, limited information is available in scientific publications. In addition, salmonids dominate both the literature and commercial focus, despite their relatively small contribution to the total volume of farmed fish in the world. This review provides an overview of the fish vaccines that are currently commercially available and some viewpoints on how the field is likely to evolve in the near future.
Expert Rev. Vaccines 4(1), 89–101 (2005)
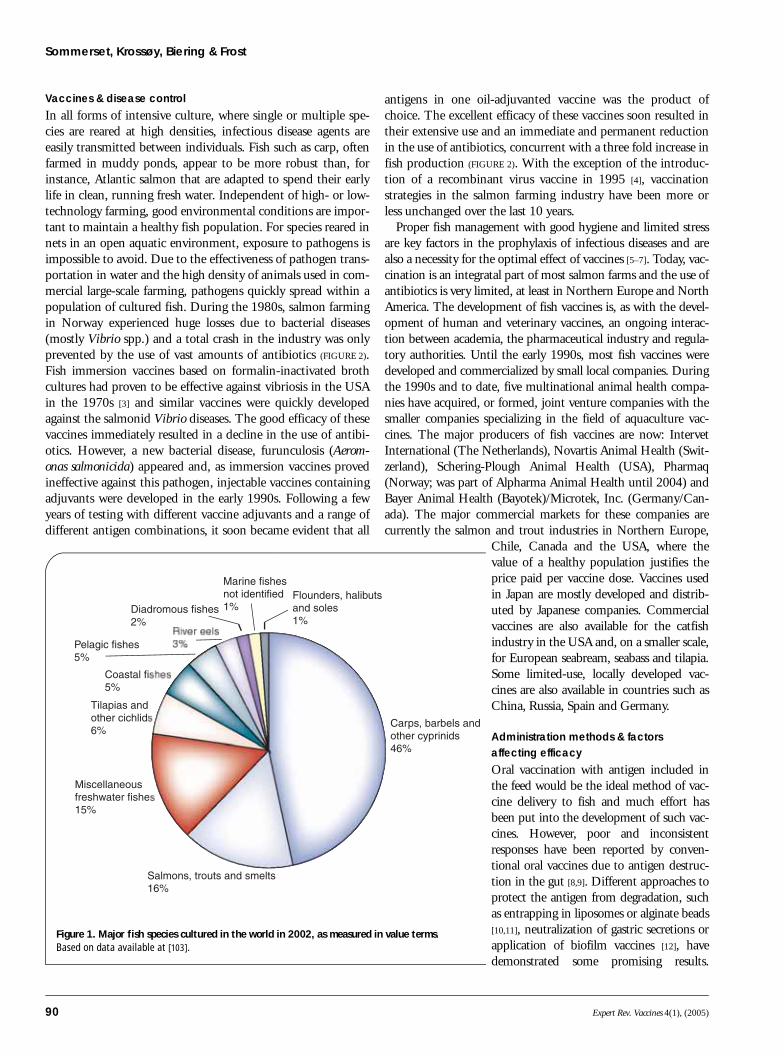
Aquaculture: agriculture of the oceansAquaculture is growing more rapidly than allother food animal-producing sectors; its contri-bution to global supplies of fish, crustaceansand molluscs increased from 3.9% of total pro-duction by weight in 1970 to 29% in2001 [101]. As the capture fishing industry hasdeclined and wild stocks diminished, the aqua-culture industry has become an importantsource of seafood. China is the largest fish pro-ducer, with 70% of the total volume and 53%of total value in 2002 [102,103]. Today, carps,barbels and other members of the cyprinidfamily dominate fish aquaculture (FIGURE 1).These fish species have a relatively low valuecompared with other types of farmed fish andare typically raised in simple pond systemswhere they are a food source for families indeveloping countries. The tendencies are, how-ever, that all fish farming will become moreindustrialized with the increasing investment
from multinational companies. Today, high-value species, such as salmon and trout,account for 7% of total volume and 16% oftotal value of cultivated fish worldwide. Atlan-tic salmon is one of the most intensivelyfarmed fish in the world, with Norway, Chile,the UK and Canada being the major produc-ers. Farming of high-value marine species suchas European seabass and seabream, halibut,flounder, cod, tuna, eel and amberjack/yellow-tail will probably increase as appropriate inten-sive aquaculture systems are developed.Although crustaceans (e.g., shrimp), molluscs(e.g., clams and oysters) and aquatic plants areimportant in global aquaculture, they do notpossess an adapted or specific immune system.Therefore, vaccines (at least by their traditionaldefinition) are not appropriate for inverte-brates and plants, and prophylactics in thefarming of these aquatic organisms will not bereviewed in this article.
For reprint orders, please contact [email protected]
Sommerset, Krossøy, Biering & Frost
90 Expert Rev. Vaccines 4(1), (2005)
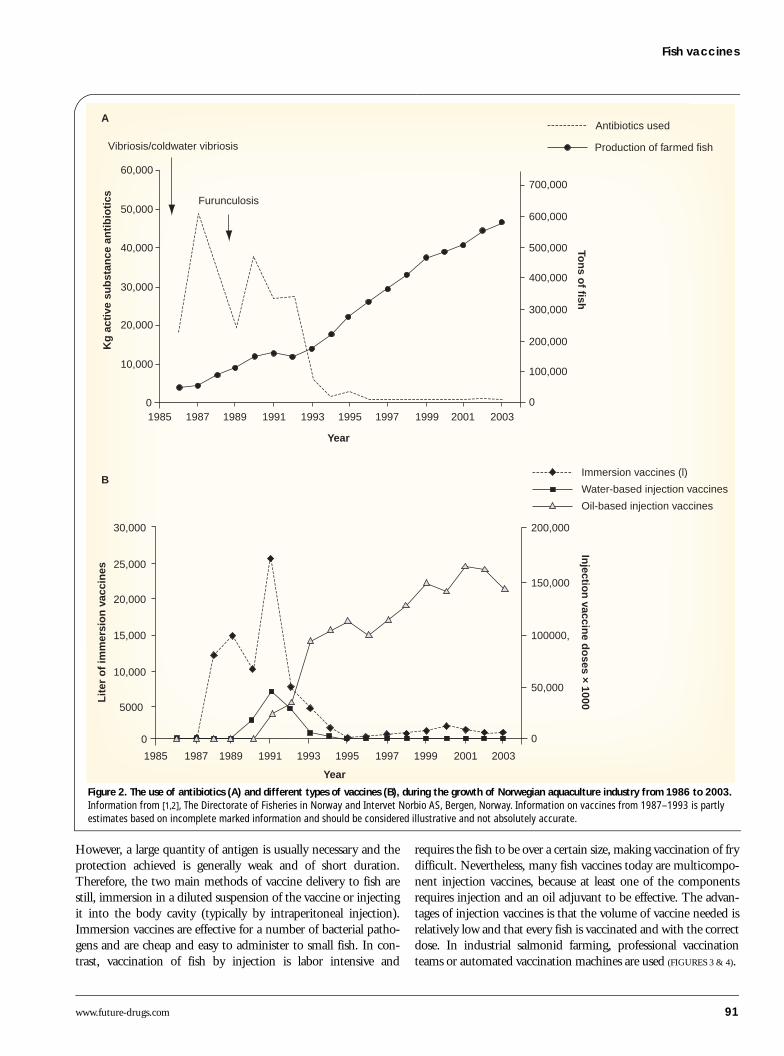
Vaccines & disease controlIn all forms of intensive culture, where single or multiple spe-cies are reared at high densities, infectious disease agents areeasily transmitted between individuals. Fish such as carp, oftenfarmed in muddy ponds, appear to be more robust than, forinstance, Atlantic salmon that are adapted to spend their earlylife in clean, running fresh water. Independent of high- or low-technology farming, good environmental conditions are impor-tant to maintain a healthy fish population. For species reared innets in an open aquatic environment, exposure to pathogens isimpossible to avoid. Due to the effectiveness of pathogen trans-portation in water and the high density of animals used in com-mercial large-scale farming, pathogens quickly spread within apopulation of cultured fish. During the 1980s, salmon farmingin Norway experienced huge losses due to bacterial diseases(mostly Vibrio spp.) and a total crash in the industry was onlyprevented by the use of vast amounts of antibiotics (FIGURE 2).Fish immersion vaccines based on formalin-inactivated brothcultures had proven to be effective against vibriosis in the USAin the 1970s [3] and similar vaccines were quickly developedagainst the salmonid Vibrio diseases. The good efficacy of thesevaccines immediately resulted in a decline in the use of antibi-otics. However, a new bacterial disease, furunculosis (Aerom-onas salmonicida) appeared and, as immersion vaccines provedineffective against this pathogen, injectable vaccines containingadjuvants were developed in the early 1990s. Following a fewyears of testing with different vaccine adjuvants and a range ofdifferent antigen combinations, it soon became evident that all
antigens in one oil-adjuvanted vaccine was the product ofchoice. The excellent efficacy of these vaccines soon resulted intheir extensive use and an immediate and permanent reductionin the use of antibiotics, concurrent with a three fold increase infish production (FIGURE 2). With the exception of the introduc-tion of a recombinant virus vaccine in 1995 [4], vaccinationstrategies in the salmon farming industry have been more orless unchanged over the last 10 years.
Proper fish management with good hygiene and limited stressare key factors in the prophylaxis of infectious diseases and arealso a necessity for the optimal effect of vaccines [5–7]. Today, vac-cination is an integratal part of most salmon farms and the use ofantibiotics is very limited, at least in Northern Europe and NorthAmerica. The development of fish vaccines is, as with the devel-opment of human and veterinary vaccines, an ongoing interac-tion between academia, the pharmaceutical industry and regula-tory authorities. Until the early 1990s, most fish vaccines weredeveloped and commercialized by small local companies. Duringthe 1990s and to date, five multinational animal health compa-nies have acquired, or formed, joint venture companies with thesmaller companies specializing in the field of aquaculture vac-cines. The major producers of fish vaccines are now: IntervetInternational (The Netherlands), Novartis Animal Health (Swit-zerland), Schering-Plough Animal Health (USA), Pharmaq(Norway; was part of Alpharma Animal Health until 2004) andBayer Animal Health (Bayotek)/Microtek, Inc. (Germany/Can-ada). The major commercial markets for these companies arecurrently the salmon and trout industries in Northern Europe,
Chile, Canada and the USA, where thevalue of a healthy population justifies theprice paid per vaccine dose. Vaccines usedin Japan are mostly developed and distrib-uted by Japanese companies. Commercialvaccines are also available for the catfishindustry in the USA and, on a smaller scale,for European seabream, seabass and tilapia.Some limited-use, locally developed vac-cines are also available in countries such asChina, Russia, Spain and Germany.
Administration methods & factors affecting efficacyOral vaccination with antigen included inthe feed would be the ideal method of vac-cine delivery to fish and much effort hasbeen put into the development of such vac-cines. However, poor and inconsistentresponses have been reported by conven-tional oral vaccines due to antigen destruc-tion in the gut [8,9]. Different approaches toprotect the antigen from degradation, suchas entrapping in liposomes or alginate beads[10,11], neutralization of gastric secretions orapplication of biofilm vaccines [12], havedemonstrated some promising results.
Carps, barbels andother cyprinids46%
Salmons, trouts and smelts16%
Miscellaneousfreshwater fishe15%
Tilapias andother cichlid6%
Coastal fi5%
Pelagic fishes5%
Diadromous fishes2%
Marine fishesnot identified1%
Flounders, halibutsand soles1%
Figure 1. Major fish species cultured in the world in 2002, as measured in value terms. Based on data available at [103].
Fish vaccines
www.future-drugs.com 91
However, a large quantity of antigen is usually necessary and theprotection achieved is generally weak and of short duration.Therefore, the two main methods of vaccine delivery to fish arestill, immersion in a diluted suspension of the vaccine or injectingit into the body cavity (typically by intraperitoneal injection).Immersion vaccines are effective for a number of bacterial patho-gens and are cheap and easy to administer to small fish. In con-trast, vaccination of fish by injection is labor intensive and
requires the fish to be over a certain size, making vaccination of frydifficult. Nevertheless, many fish vaccines today are multicompo-nent injection vaccines, because at least one of the componentsrequires injection and an oil adjuvant to be effective. The advan-tages of injection vaccines is that the volume of vaccine needed isrelatively low and that every fish is vaccinated and with the correctdose. In industrial salmonid farming, professional vaccinationteams or automated vaccination machines are used (FIGURES 3 & 4).
60,000
50,000
40,000
30,000
20,000
10,000
01985 1987 1989 1991 1993 1995 1997 1999 2001 2003
0
Kg
act
ive
sub
stan
ce a
nti
bio
tics
700,000
600,000
500,000
400,000
300,000
200,000
100,000
Antibiotics used
Production of farmed fish
A
Year
Vibriosis/coldwater vibriosis
FurunculosisTo
ns o
f fish
30,000
25,000
20,000
15,000
10,000
5000
0
1985 1987 1989 1991 1993 1995 1997 1999 2001 2003
0
200,000
150,000
100000,
50,000
Year
Lit
er o
f im
mer
sio
n v
acci
nes
Injectio
n vaccin
e do
ses × 1000
Immersion vaccines (l)
Water-based injection vaccines
Oil-based injection vaccines
B
Figure 2. The use of antibiotics (A) and different types of vaccines (B), during the growth of Norwegian aquaculture industry from 1986 to 2003. Information from [1,2], The Directorate of Fisheries in Norway and Intervet Norbio AS, Bergen, Norway. Information on vaccines from 1987–1993 is partly estimates based on incomplete marked information and should be considered illustrative and not absolutely accurate.

Sommerset, Krossøy, Biering & Frost
92 Expert Rev. Vaccines 4(1), (2005)
Fish are cold-blooded animals with a body temperature thatequals their surroundings. Dependent upon fish species and tem-perature, vaccination must be performed within a certain mini-mum period of time before the risk of their exposure topathogens [6]. The onset of an immune response is faster in warm-water species than in cold-water species. At an optimal tempera-ture for Atlantic salmon (10–12°C), antibodies are typically notdetected until at least 4–6 weeks after vaccination, whereas awarm-water species such as seabass, with an optimal temperatureof 22°C, can have detectable antibodies 1 week after immuniza-tion [13]. In addition to temperature, stress caused by environmen-tal or man-made factors such as photoperiod, seasonal changes,salinity, heavy metals, crowding, handling and transport, caninduce immune suppression and be a limiting factor for vaccineefficacy [14]. Unlike wild fish exposed to similar stressors, farmedfish cannot escape from these factors, and both acute and chronicstress can render fish susceptible to different opportunistic as wellas pathogenic microorganisms.
Limitations in vaccine developmentThe major goal of vaccination is to induce aspecific long-term protection against a cer-tain disease. It has been debated whether theeffective long-term protection of oil-adju-vant injection vaccines is due to immuno-logic memory in the fish or constant stimu-lation from the antigen depot. As theexisting empirically developed vaccines caninduce protection after a single administra-tion and until the fish are harvested, lesseffort has been put into the investigation ofthe actual mechanisms behind the protec-tion. In evolutionary terms, fish are the firstgroup of animals with the basic aspects ofthe immune system of higher vertebrates.Although fish immune systems are primi-tive compared with mammals, there seem tobe more similarities than differences [15,16].In the limited number of fish species stud-ied, the major antibody type is an immu-noglobulin (Ig)M-like isotype that typically
exists as a tetramer in its secreted form [17]. No isotypes corre-sponding to mammalian IgG, IgA or IgE have been identified infish and the secondary humoral immune response in fish is, ifpresent at all, less prominent than in mammals. Due to ease ofmeasurement, the presence and titer of specific antibodies is a fre-quently used immunologic assay. Assays to measure specific T-cellresponses are still in their infancy, however, cell lines from clonedfish lines (matching major histocompatibility complex [MHC]molecules) are now available for carp and rainbow trout [18]. Evenamong the relatively few fish species that are commercially farmedtoday, it is important to keep in mind that fish immunology can-not be considered a homogenous system. The number of fish spe-cies is much greater than the number of mammalian species andthe evolutionary distance between different teleost (bony fish)families are considerably larger than between mammals. Further-more, there are more than 25,000 species of fish living in habitatsranging from polar regions to the tropics, from muddy fresh
Figure 3. Large-scale vaccination of juvenile Atlantic salmon. Fish are transported in pipes from the rearing tanks to an anesthetic bath and anaesthetized fish are injected by the vaccination team.
Figure 4. Manual vaccination of Atlantic salmon. Two metallic bows are attached to the injection unit to aid a correct intraperitoneal injection of the vaccine dose.

Fish vaccines
www.future-drugs.com 93
water to the ocean, from dark high-pressure depths to bright low-pressure surface areas. It is therefore likely that different specieshave major differences in how they combat pathogens and also intheir response to vaccines. Lack of detailed knowledge of theimmune systems in different fish species limits the possibilities tostudy both pathogen and vaccine-induced immunity.
As with all veterinary vaccines, cost effectiveness in the fieldis an essential limitation to commercial vaccine development.For example, the calculations would be very different for apopulation of sturgeon used to produce Russian caviar versus apopulation of carp in a Chinese village pond. Fish generallyneed a large antigen dose compared with terrestrial animalsand cost-effective inactivated viral vaccines have proven diffi-cult to develop. In some species, all types of injection vaccines(or even immersion vaccines) are simply too expensive. Otherspecies are too vulnerable to handle the stress induced duringthe vaccination or may develop severe side effects post vaccina-tion. Yet, in other species, the major disease problems mayappear in the larval or fry stages, before the animal is largeenough to be vaccinated or have even developed a functionalimmune system. The apparent lack of maternal immunity infish also limits the possibilities to protect offspring by parentalvaccination [19,20].
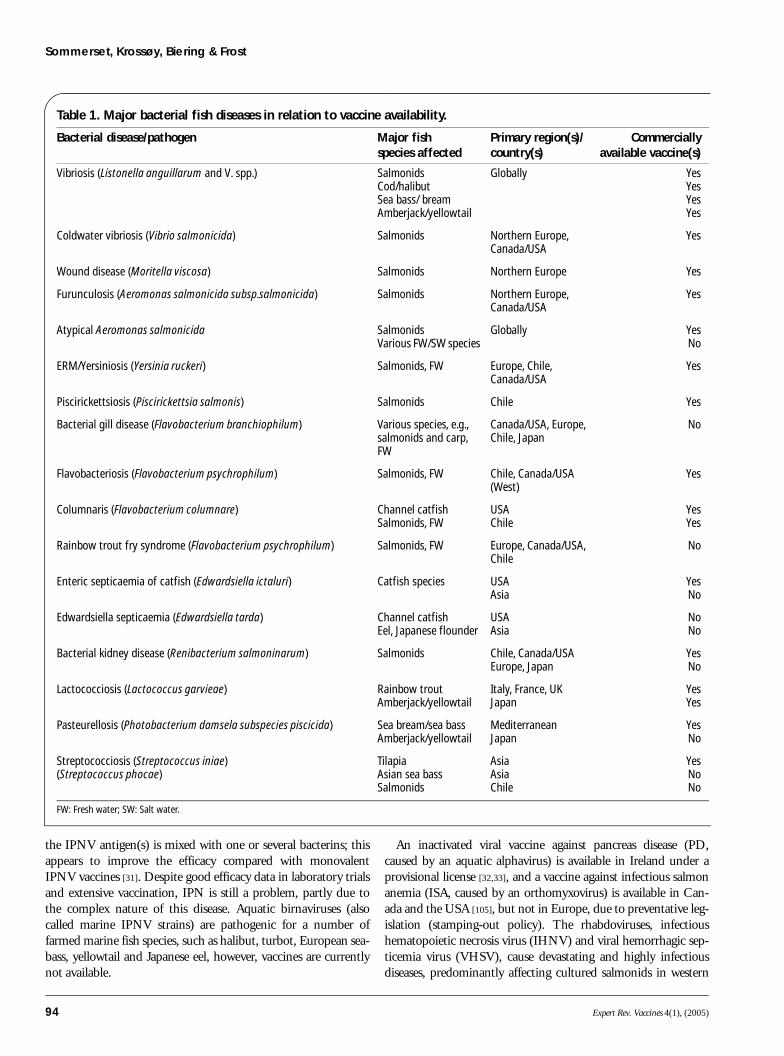
Bacterial fish vaccinesAt present, vaccines are available against many of the seriousbacterial diseases causing problems for the fish farming indus-tries (TABLE 1) [21]. The first commercially available bacterial vac-cines were against enteric redmouth disease (ERM, yersiniosis)and vibriosis, introduced in the USA in the late 1970s [3].These vaccines were based on inactivated whole-cell formula-tions and were administered by immersion. Such vaccines haveproven efficacious in preventing many of the major bacterialdiseases and the fact that these vaccines can be produced at lowcost makes them ideal for use in the aquaculture industry.Vibriosis has a global distribution in a wide range of fish spe-cies. The bacterial species causing classical vibriosis isListonella anguillarum (previous nomenclature, Vibrio anguil-larum), with more than ten serotypes characterized so far. Forthe salmonid and cold-water marine fish species such as codand halibut, different serotypes might be involved in diseaseoutbreaks; thus, vaccines designed for salmonids cannot auto-matically be used for the marine species. In addition to usingthe right serotypes, the main challenges for vaccination ofmarine species are that vaccination often has to be performedon very small juveniles/larvae and that outbreaks occur duringthe entire lifecycle. This probably means that in some speciesrevaccination will be necessary if the fish are to be protectedduring the entire production period.
Vibriosis is an example of a disease for which the simple inac-tivated bacterin vaccines works well but other bacteria haveproven more difficult to control by means of vaccination. In thechannel catfish industry, Edwardsiella ictaluri and Flavobacte-rium columnare are two of the most important pathogens, withlosses of up to US$60–80 million annually in the USA [22].
Vaccines containing E. ictaluri and F. columnare bacterins havelow efficacy and the use of live attenuated isolates is a new andpromising approach [23,24]. A live attenuated E. ictaluri vaccine,licensed in the USA a few years ago, has proven efficacious byimmersion of fish as early as 7–10 days post hatching [25]. Alive vaccine has also been licensed for use in salmonids inNorth America and Chile. This vaccine is based on the cross-protective property of Arthrobacter spp. against bacterial kidneydisease (BKD) caused by the intracellular bacterium Renibacte-rium salmoninarum [26]. Another intracellular bacterium forwhich development of vaccines has proven difficult is Piscirick-ettsia salmonis. This is the causative agent of salmonid rickett-sial septicemia (SRS, piscirickettsiosis), which is the most seri-ous disease problem in the Chilean aquaculture industry.Inactivated P. salmonis bacterins are available, however, due tolow efficacy, development of new vaccines based on recom-binant proteins is ongoing [27]. These new approaches mightoffer a solution for diseases where inactivated bacterins are inef-ficient, although the long-term performance of the vaccinesremains to be documented.
Viral fish vaccinesToday, most available virus vaccines for aquaculture are based oninactivated virus or recombinant subunit proteins. Inacti-vated/killed viral vaccines are generally not efficacious unless deliv-ered by injection and, as relatively high doses are needed to achieveprotection, cost-effective inactivated viral vaccines are difficult todevelop. Live viral vaccines have been tested with good results infish [28–30] and should be the optimal regarding protection, admin-istration and price. However, as discussed below, ecosafety aspectsof live virus vaccines are considered major drawbacks and currentlyhinder their use as commercial vaccines.
The first viral vaccine for fish was produced by a Czechoslova-kian company (Bioveta) in 1982. The vaccine was against a carprhabdovirus, causing spring viremia of carp (SVC) and was basedon two inactivated strains of SVC virus emulsified in oil andadministered by injection. TABLE 2 summarizes the viral fish vac-cines commercially available today. Carp production is huge inAsia, both for food (common carp) and ornamental (koi carp)purposes. Although some severe viral diseases exist (e.g., causedby koi herpes virus, grass carp hemorrhage disease virus and SVCvirus), few viral vaccine trials have been published. To theauthors’ knowledge, the only commercial carp vaccine in Asia isan inactivated grass carp hemorrhage disease virus (a reovirus)vaccine, which has been widely used in China. A koi herpesvirusvaccine based on an attenuated strain of carp interstitial nephritisand gill necrosis virus is available in Israel [30,104].
In salmonid farming, commercial vaccines exist for some of themost common viral diseases. However, few, if any, of the viralvaccines are as efficacious as the bacterial vaccines. Disease causedby the birnavirus, infectious pancreatic necrosis virus (IPNV), isa major problem in the salmonid industry [31]. Commerciallyavailable IPNV vaccines are based on either inactivated cell cul-ture-propagated virus or recombinant structural proteins. MostIPNV vaccines exist as polyvalent oil-adjuvanted vaccines where
Sommerset, Krossøy, Biering & Frost
94 Expert Rev. Vaccines 4(1), (2005)
the IPNV antigen(s) is mixed with one or several bacterins; thisappears to improve the efficacy compared with monovalentIPNV vaccines [31]. Despite good efficacy data in laboratory trialsand extensive vaccination, IPN is still a problem, partly due tothe complex nature of this disease. Aquatic birnaviruses (alsocalled marine IPNV strains) are pathogenic for a number offarmed marine fish species, such as halibut, turbot, European sea-bass, yellowtail and Japanese eel, however, vaccines are currentlynot available.
An inactivated viral vaccine against pancreas disease (PD,caused by an aquatic alphavirus) is available in Ireland under aprovisional license [32,33], and a vaccine against infectious salmonanemia (ISA, caused by an orthomyxovirus) is available in Can-ada and the USA [105], but not in Europe, due to preventative leg-islation (stamping-out policy). The rhabdoviruses, infectioushematopoietic necrosis virus (IHNV) and viral hemorrhagic sep-ticemia virus (VHSV), cause devastating and highly infectiousdiseases, predominantly affecting cultured salmonids in western
Table 1. Major bacterial fish diseases in relation to vaccine availability.
Bacterial disease/pathogen Major fishspecies affected
Primary region(s)/ country(s)
Commerciallyavailable vaccine(s)
Vibriosis (Listonella anguillarum and V. spp.) Salmonids Cod/halibutSea bass/ breamAmberjack/yellowtail
Globally YesYesYesYes
Coldwater vibriosis (Vibrio salmonicida) Salmonids Northern Europe, Canada/USA
Yes
Wound disease (Moritella viscosa) Salmonids Northern Europe Yes
Furunculosis (Aeromonas salmonicida subsp.salmonicida) Salmonids Northern Europe, Canada/USA
Yes
Atypical Aeromonas salmonicida SalmonidsVarious FW/SW species
Globally YesNo
ERM/Yersiniosis (Yersinia ruckeri) Salmonids, FW Europe, Chile, Canada/USA
Yes
Piscirickettsiosis (Piscirickettsia salmonis) Salmonids Chile Yes
Bacterial gill disease (Flavobacterium branchiophilum) Various species, e.g., salmonids and carp, FW
Canada/USA, Europe, Chile, Japan
No
Flavobacteriosis (Flavobacterium psychrophilum) Salmonids, FW Chile, Canada/USA (West)
Yes
Columnaris (Flavobacterium columnare) Channel catfishSalmonids, FW
USAChile
YesYes
Rainbow trout fry syndrome (Flavobacterium psychrophilum) Salmonids, FW Europe, Canada/USA, Chile
No
Enteric septicaemia of catfish (Edwardsiella ictaluri) Catfish species USA Asia
YesNo
Edwardsiella septicaemia (Edwardsiella tarda) Channel catfishEel, Japanese flounder
USA Asia
NoNo
Bacterial kidney disease (Renibacterium salmoninarum) Salmonids Chile, Canada/USA Europe, Japan
YesNo
Lactococciosis (Lactococcus garvieae) Rainbow troutAmberjack/yellowtail
Italy, France, UK Japan
YesYes
Pasteurellosis (Photobacterium damsela subspecies piscicida) Sea bream/sea bassAmberjack/yellowtail
Mediterranean Japan
YesNo
Streptococciosis (Streptococcus iniae)(Streptococcus phocae)
Tilapia Asian sea bassSalmonids
Asia AsiaChile
YesNoNo
FW: Fresh water; SW: Salt water.

Fish vaccines
www.future-drugs.com 95
North America (IHNV), France (VHSV and IHNV) and Den-mark (VHSV). For more than 30 years, several attempts havebeen made to develop efficacious vaccines using the conventionalapproaches of killed or live virus vaccines. Despite their ability toinduce good protection in laboratory studies, the live vaccineswere demonstrated to be unsafe for field use and inactivated vac-cines required high doses [34,31]. Different recombinant subunitvaccines based on the IHNV and VHSV membrane glycoproteinhave been less successful [35–38]. However, DNA vaccines encod-ing the same viral glycoproteins are remarkably efficacious [39–41].Indeed, these DNA vaccines are protective when used at smalldoses (as little as 10 ng in trout fry) and efficacious as early as4–8 days and for up to 2 years post vaccination [42–46].
Viral nervous necrosis, caused by betanodaviruses, is a majorproblem in the farming of several marine fish species (e.g., Euro-pean seabass, Atlantic halibut, barramundi and several groupers)[47], yet commercial vaccines are unavailable. A few publicationshave demonstrated the effect of recombinant subunit formulations[48–50]. However, in most marine species, disease caused by betano-daviruses strike early in the lifecycle of the fish (larval or fry stages),before injection of these formulations are protective or applicable.
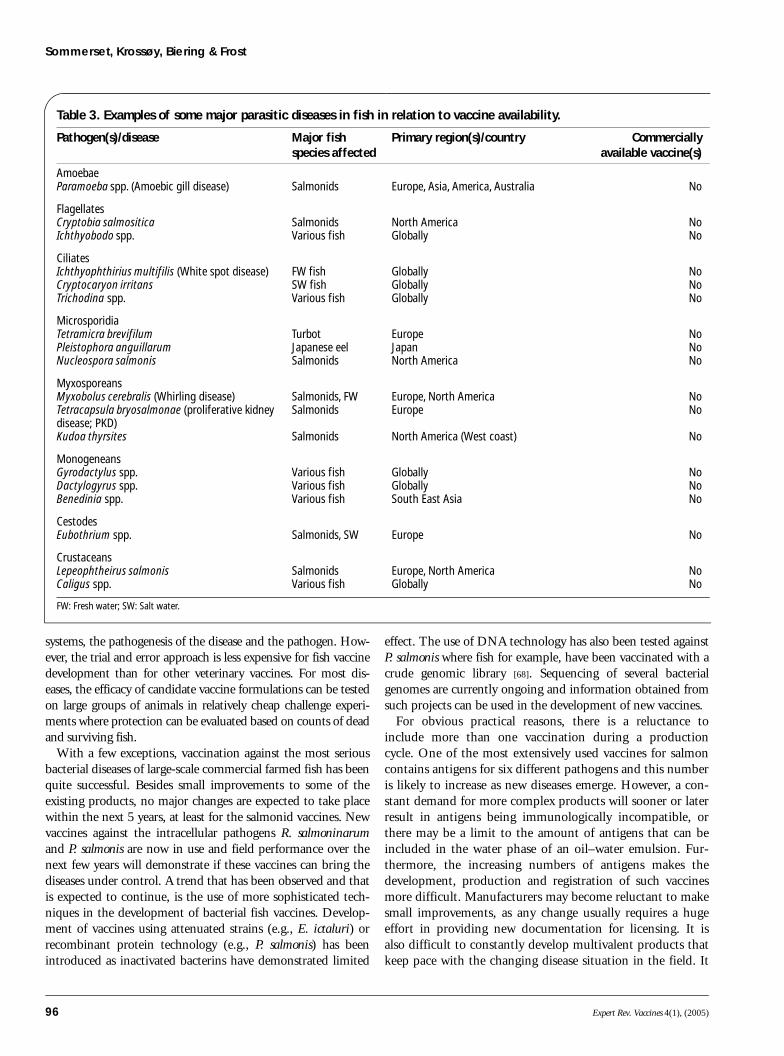
Fish vaccines against parasitesThere is a wide range of parasites in both wild and cultured fishstocks. Although parasitic diseases such as amoebic gill disease,white spot disease, whirling disease, proliferative kidney disease(PKD) and salmon lice infestation create severe problems in fishfarming, no parasite vaccines are commercially available (TABLE 3).
Instead, large amounts of antiparasitic pharmaceuticals are usedwith potentially negative environmental effects. In general, fishpossess both humoral and cell-mediated defense mechanismsagainst many parasites [51–53] and there are many reports onimmunity/increased resistance among fish surviving natural par-asite infections [54–58]. Therefore, there are no principal biologiclimitations hindering vaccine development against at least someparasitic diseases in fish. Experimental parasite vaccines based onwhole pathogens have been reported and live vaccines appear tobe superior to killed vaccines [59–64].
Cultivation of parasites for potential killed or live vaccines iseven more expensive than virus cultivation, as a host populationrather than cell cultures are usually required. In addition to thehigh costs, the use of natural hosts for cultivation of the parasitewould create major problems with respect to safety documenta-tion. Therefore, identification and production of protectiveantigens is probably the most feasible strategy towards commer-cial parasite vaccines, at least for low-cost vaccines. The success-fully developed vaccine against cattle tick (Boophilus microplus)[65–67], a terrestrial ectoparasite of cattle, demonstrates that thisstrategy is indeed feasible, albeit very difficult.
Expert opinion & five-year viewTo date, trial and error has been the main strategy for the devel-opment of fish vaccines and this empirical approach will probablycontinue in the short term. Development of vaccines for newlycultivated species (or new diseases in already domesticated fish) isusually restricted by a limited knowledge of the fish immune
Table 2. Major viral diseases in fish in relation to vaccine availability.
Viral disease/pathogen Major fish species affected Primary region(s)/country(s)
Commerciallyavailable vaccine(s)
Infectious pancreatic necrosis/IPNV, other aquatic birnaviruses
SalmonidsVarious marine species
Globally YesNo
Pancreas disease/PDV Salmon UK, Ireland, Norway Yes
Infectious salmon anemia/ISAV Salmonids Canada/USA (East), Norway, UK
Yes
Infectious hematopoietic necrosis/IHNV Salmonids Canada/USA (West) Yes
Viral hemorrhagic septicemia/VHSV Rainbow and brown trout, turbot, Japanese flounder
Europe, Asia No
Viral nervous necrosis/SJNNV and several other betanodavirus
Several marine fish species, e.g., sea bass, groupers, barramundi, halibut
Globally No
Iridoviral disease/RSIV Red sea bream, amberjack/yellowtail Asia YesYes
Channel catfish virus disease/CCV Channel catfish USA No
Spring viremia of carp: /SVCV Mostly carp species Europe No*
Grass carp hemorrhage disease/GCHDV Grass carp China Yes‡
*Previously available inactivated virus vaccine but no longer commercially available; ‡Previously available but may not be in use today.CCV: Channel catfish virus; GCHDV: Grass carp hemorrhage disease virus; IHNV: Infectious hematopoietic necrosis; IPNV: Infectious pancreatic necrosis virus; ISAV: Infectious salmon anemia; PDV: Pancreas disease virus; RSIV: Red sea bream iridovirus: SJNNV: Striped jack nervous necrosis virus: SVCV: Spring viremia of carp virus; VHSV: Viral hemorrhagic septicemia virus.
Sommerset, Krossøy, Biering & Frost
96 Expert Rev. Vaccines 4(1), (2005)
systems, the pathogenesis of the disease and the pathogen. How-ever, the trial and error approach is less expensive for fish vaccinedevelopment than for other veterinary vaccines. For most dis-eases, the efficacy of candidate vaccine formulations can be testedon large groups of animals in relatively cheap challenge experi-ments where protection can be evaluated based on counts of deadand surviving fish.
With a few exceptions, vaccination against the most seriousbacterial diseases of large-scale commercial farmed fish has beenquite successful. Besides small improvements to some of theexisting products, no major changes are expected to take placewithin the next 5 years, at least for the salmonid vaccines. Newvaccines against the intracellular pathogens R. salmoninarumand P. salmonis are now in use and field performance over thenext few years will demonstrate if these vaccines can bring thediseases under control. A trend that has been observed and thatis expected to continue, is the use of more sophisticated tech-niques in the development of bacterial fish vaccines. Develop-ment of vaccines using attenuated strains (e.g., E. ictaluri) orrecombinant protein technology (e.g., P. salmonis) has beenintroduced as inactivated bacterins have demonstrated limited
effect. The use of DNA technology has also been tested againstP. salmonis where fish for example, have been vaccinated with acrude genomic library [68]. Sequencing of several bacterialgenomes are currently ongoing and information obtained fromsuch projects can be used in the development of new vaccines.
For obvious practical reasons, there is a reluctance toinclude more than one vaccination during a productioncycle. One of the most extensively used vaccines for salmoncontains antigens for six different pathogens and this numberis likely to increase as new diseases emerge. However, a con-stant demand for more complex products will sooner or laterresult in antigens being immunologically incompatible, orthere may be a limit to the amount of antigens that can beincluded in the water phase of an oil–water emulsion. Fur-thermore, the increasing numbers of antigens makes thedevelopment, production and registration of such vaccinesmore difficult. Manufacturers may become reluctant to makesmall improvements, as any change usually requires a hugeeffort in providing new documentation for licensing. It isalso difficult to constantly develop multivalent products thatkeep pace with the changing disease situation in the field. It
Table 3. Examples of some major parasitic diseases in fish in relation to vaccine availability.
Pathogen(s)/disease Major fish species affected
Primary region(s)/country Commerciallyavailable vaccine(s)
Amoebae Paramoeba spp. (Amoebic gill disease) Salmonids Europe, Asia, America, Australia No
FlagellatesCryptobia salmositicaIchthyobodo spp.
SalmonidsVarious fish
North AmericaGlobally
NoNo
CiliatesIchthyophthirius multifilis (White spot disease)Cryptocaryon irritansTrichodina spp.
FW fishSW fishVarious fish
GloballyGloballyGlobally
NoNoNo
MicrosporidiaTetramicra brevifilumPleistophora anguillarumNucleospora salmonis
TurbotJapanese eelSalmonids
EuropeJapanNorth America
NoNoNo
MyxosporeansMyxobolus cerebralis (Whirling disease)Tetracapsula bryosalmonae (proliferative kidney disease; PKD)Kudoa thyrsites
Salmonids, FWSalmonids
Salmonids
Europe, North AmericaEurope
North America (West coast)
NoNo
No
MonogeneansGyrodactylus spp. Dactylogyrus spp.Benedinia spp.
Various fishVarious fishVarious fish
GloballyGloballySouth East Asia
NoNoNo
Cestodes Eubothrium spp. Salmonids, SW Europe No
CrustaceansLepeophtheirus salmonis Caligus spp.
SalmonidsVarious fish
Europe, North AmericaGlobally
NoNo
FW: Fresh water; SW: Salt water.

Fish vaccines
www.future-drugs.com 97
is likely that this development will sooner or later make itnecessary to apply more than one vaccination, thus enablingthe development of vaccines with fewer antigens.
Adverse effects following vaccination with adjuvant vac-cines are common problems and fish vaccines are no excep-tion to this rule [69]. As injection of nonadjuvant vaccines oradjuvant without antigens results in little or no side effects, itis clear that it is the combination of adjuvant and antigens,particularly crude bacterial broth antigens, which createsproblems. Adverse effects have been taken very seroiusly byvaccine manufacturers in recent years. Products with a greaterbenefit to risk ratio are now on the market and the quest fornew adjuvants with satisfactory efficacy and minimal sideeffects will continue. As bacterial antigens contribute signifi-cantly to the side effects, another solution to this problemwould be to identify and separate the protective antigensfrom those that induce side effects. For already effective bac-terin vaccines, this is not likely to happen in the near future,as the process of identifying and separating these componentsis expensive and laborious.
Injection vaccines developed for some of the new marine spe-cies in aquaculture may require adjuvants other than oil. In manyspecies, both the liver and the eggs are valuable food sources (theliver is also an important source of fish oil) and remnants of oiladjuvants in these products will be undesirable. Delivery systemsfor effective oral or immersion administration would be a majorimprovement to fish vaccination in the future. Recently, a Cana-dian company has made interesting progress regarding oraladministration of antigens [106], but it is unlikely that thisapproach will result in effective commercially available vaccineswithin a 5-year view. Even in humans, only the live polio vaccinehas been widely used with oral administration [70].
The lack of effective viral vaccines is one of the main prob-lems facing fish vaccinology. Within the next 5 years, new and(or) improved virus vaccines will probably be developed forISA, PD, VHS and IHN. Experimental IHN and VHS DNAvaccines are highly effective and the DNA vaccine against IHNis currently being tested in controlled field trials in Canada. It islikely that an IHN vaccine will be the first licensed DNA vac-cine for fish. For aquaculture applications, DNA vaccines aresafer than attenuated live vaccines, but varying governmentalattitudes towards both DNA and genetically modified organ-ism vaccines (North America being more liberal than Europe)may limit the use of both these vaccine types.
Live attenuated virus vaccines would be the optimal fishvaccines if cost, efficacy and ease of administration were theonly concerns. However, commercial fish vaccine develop-ment is limited by safety concerns to the consumers, and tothe environment. Most cultured fish live in ponds or netswith no physical barriers to wild stocks. This limits the use oflive vaccines, whether they are attenuated by classical meansor by use of molecular biology. Although nonpathogenic tothe target animal, a live vaccine may have to be proven non-pathogenic to all other relevant species in the aquatic environ-ment. Such safety data, if possible to generate at all, will
demand an enormous cost and effort to collect. However, forhigh-value species reared in closed indoor tanks with sterilizedeffluent water, attenuated virus vaccines may become accepta-ble. In addition, if live virus vaccines are to become a realisticalternative also for species reared in nets in the ocean, theyneed to have a high level of safety, for example, by develop-ment of a replication-deficient virus (infective but unable toproduce new infective virus). These viruses or other modifiedmicroorganisms might be used as vehicles for other protectiveantigens. Based on reverse genetics methodology, a new liverecombinant IHNV vaccine strain has been developed by aFrench group [71,72]. As the small nonvirion protein Nvproved to be nonessential for the recombinant IHNV, the Nvgene may be used as a site of insertion for foreign gene expres-sion and potentially serve as a vector for expressing additionalantigens in the host.
An interesting aspect of the fish rhabdovirus glycoproteinDNA vaccines is their stimulation of strong innate antiviralresponses and the ability to induce early protection against het-erologous viruses [41,44,45,73]. In fish, the innate immune systemsprobably play a very important role in the protection againstinfectious diseases [74]. More attention should be paid to aspectsof the innate immune responses in the search for specificimmune stimulants and adjuvants. Recently, Jørgensen andcoworkers demonstrated that short DNA fragments (CpGDNA) protect salmon, to a certain extent, from IPNV chal-lenge, suggesting that CpG DNA may be an effective adjuvantin some viral vaccines [75].
With the use of modern genomics, strategies for fish para-site vaccine development should be feasible. The commercialBoophilus vaccine in cattle is an example of how a single pro-tein expressed in the tick gut, a protein that is not exposed tothe host during a natural infection (i.e., a concealed antigen),can be used as a vaccine antigen, at least in parasites that feedon host tissue that contains components of the immune sys-tem. To study a parasite in such detail that new essential bio-logic processes are identified, is a major task, both financiallyand scientifically. This will probably limit this kind of vac-cine development to high-value fish species where a singleparasite causes huge financial or ecologic problems. In ouropinion, the ectoparasitic copepods L. salmonis (salmonlouse) and Caligus spp., are currently the only fish parasitesthat fit this description. On salmon ongrowing farms thesecopepods are the major parasite problem and in the northernhemisphere salmon lice alone cause €50–100 million annuallosses through mortality, growth reduction, quality reductionand pharmaceutical costs [76]. Caligus spp. currently causessevere problems for the huge salmon industry in Chile.Unlike the salmon lice, Caligus spp., are not salmonid spe-cific and are found globally, and most cultured marine fishspecies, present and future, are likely to experience Caligusproblems, as production is intensified. In addition, the phar-maceuticals currently used against ectoparasites may sooneror later induce drug resistance. Therefore, efforts in basicresearch and commercial vaccine development against these

Sommerset, Krossøy, Biering & Frost
98 Expert Rev. Vaccines 4(1), (2005)
ectoparasites is likely to be intensified. As long as the marketpotential is large enough and vaccine development feasible,an effort towards product development is likely to take place.
AcknowledgementsInformation regarding commercial fish vaccines worldwidewould have been difficult to obtain without the invaluable
assistance from colleagues in Intervet. In particular WilliamEnright, Oscar Parra, Luc Grisez, Marian McLoughlin, RoyOlav Hovlid and Kari Thorsø are thanked for their effort.Dag H Knappskog, Frank Nilsen and Stephane Villoing areacknowledged for valuable comments during the prepara-tion of this manuscript and Irene Nygård for making theFIGURE 3 collage.
Key issues
• Most of today’s fish vaccines have been developed and commercialized for the salmon and trout farming industry, but vaccines are also available for other high-value fresh water and marine species.
• Currently, most vaccines are based on simple empirically developed inactivated pathogens. A few recombinant subunit vaccines are also available.
• Vaccines against bacterial diseases are, with a few exceptions, highly effective while more variable efficacy is apparent for viral vaccines. No vaccines against fish parasites exist.
• Injectable multivalent oil–emulsion vaccines for salmonids dominate the commercial fish vaccine market.• A relatively high antigenic mass is needed in most fish vaccines compared with similar vaccines used in higher vertebrates.• Environmental safety concerns currently hinder the development and use of live virus vaccines in fish.• Species diversity and limited knowledge of immune systems in fish limit the development of vaccines or new vaccine delivery
systems based on nonempirical strategies.
ReferencesPapers of special note have been highlighted as:
• of interest
•• of considerable interest
1 Grave K, Engelstad M, Sørli NE, Håstein T. Utilization of antibacterial drugs in salmonid farming in Norway during 1980–1988. Aquaculture 83, 347–358 (1990).
2 Lillehaug A, Lunestad BT, Grave K. Epidemiology of bacterial diseases in Norwegian aquaculture – a description based on antibiotic prescription data for the 10 year period 1991–2000. Dis. Aquat. Org. 53, 115–125 (2003).
3 Evelyn TPT. A historical review of fish vaccinology. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Eds.). Karger, Basel, Switzerland, 90, 3–12 (1997).
4 Frost P, Ness A. Vaccination of Atlantic salmon with recombinant VP2 of infectious pancreatic necrosis virus (IPNV), added to a multivalent vaccine, suppresses viral replication following IPNV challenge. Fish Shellfish Immunol. 7, 609–619 (1997).
• Demonstrates suppression of virus replication in Atlantic salmon vaccinated with what became the first commercially available recombinant subunit vaccine for fish.
5 Press CM, Lillehaug A. Vaccination in European salmonid aquaculture: a review of practices and prospects. Br. Vet. J. 151, 45–69 (1995).
6 Lillehaug A. Vaccination strategies in seawater cage cultures of salmonids. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Ed.). Karger, Basel, Switzerland, 90, 401–408 (1997).
7 Larsen JL, Pedersen K. Vaccination strategies in freshwater salmonid aquaculture. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Ed.). Karger, Basel, Switzerland, 90, 391–400 (1997).
8 Hart S, Wrathmell AB, Harris JE, Grayson TH. Gut immunology in fish: a review. Dev. Comp. Immunol.12, 453–480 (1988). Erratum in: Dev. Comp. Immunol. 13, 93–100 (1989).
9 Quentel C, Vigneulle M. Antigen uptake and immune responses after immersion vaccination. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Ed.). Karger, Basel, Switzerland, 90, 69–78 (1997).
10 Ire T, Watarai S, Iwasaki T, Kodama H. Protection against experimental Aeromonas salmonicida infection in carp by oral immunisation with bacterial antigen entrapped in liposomes. Fish Shellfish Immunol. 18, 235–242 (2005).
11 Maurice S, Nussinovitch A, Jaffe N, Shoseyov O, Gertler A. Oral immunization of Carassius auratus with modified recombinant A-layer proteins entrapped in alginated beads. Vaccine 23, 450–459 (2004).
12 Azad IS, Shankar KM, Mohan CV, Kalita B. Uptake and processing of biofilm and free-cell vaccines of Aeromonas hydrophila in indian major carps and common carp following oral vaccination-antigen localization by a monoclonal antibody. Dis. Aquat. Organ. 43, 103–108 (2000).
13 Hrubec TC.Ward D, Smith SA, Robertsen JL. Age related changes in humoral immune response of hybrid striped bass (Morone chrysops x Morone saxatilis). Vet. Immunol. Immunopathol. 101, 103–108 (2004).
14 Bly JE, Quiniou SM-A, Clem LW. Environmental effects on fish immune mechanisms. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Eds). Karger, Basel, Switzerland, 90, 33–43 (1997).
15 Manning MJ, Nakanishi T. The specific immune system: cellular defenses. In: The Fish Immune System. Iwama G, Nakanishi T (Eds), Academic Press, London, UK, 160–195 (1997).
16 Secombes CJ. Adaptive immunity in teleost II (cellular immunity). 3rd International Symposium on Fish Vaccinology. Abstract book, 45. Bergen, Norway, 9–11 April (2003).
17 Pilstrom L, Bengten E. Immunoglobulin in fish: genes, expression and structure. Fish Shellfish Immunol. 6, 243–262 (1996).

Fish vaccines
www.future-drugs.com 99
18 Nakanishi T, Fischer U, Dijkstra JM et al. Cytotoxic T-cell function in fish. Dev. Comp. Immunol. 26, 131–139 (2002).
• The authors give a good overview of cell-mediated immunity in fish and present their recent work on cytotoxic T-cell function employing isogeneic fish and cell lines.
19 Lillehaug A, Svatdal S, Endal T. Passive transfer of specific maternal immunity does not protect Atlantic salmon (Salmo salar L.) fry against yersiniosis. Fish Shellfish Immunol. 6, 521–535 (1996).
20 Tanaka T, Furukawa K, Suzuki Y, Aida K. Transfer of maternal antibody from mother to egg may have no protective meaning for larvae of Red sea bream Pagrus major, a marine teleost. Fish Sci. 65, 240–243 (1999).
21 Håstein T, Gudding R, Evensen Ø. Bacterial vaccines for fish – an update of the current situation worldwide In: Fish Vaccinology, Developments in Biologicals. Midtlyng PJ (Ed.), Karger, Basel, Switzerland, 121, 55–74 (2005).
•• Comprehensive and updated review regarding bacterial fish vaccines.
22 Shoemaker CA, Klesius PH, Evans JJ, Arias CR. Modified live vaccines against Edwadsiella ictaluri and Flavobacterium culumnare. Proceedings of the 28th Annual Eastern Fish Health Workshop, Gettysburg, PA, USA (2003).
23 Klesius PH, Shoemaker, CA. Development and use of modified live Edwardsiella ictaluri vaccine against enteric septicemia of catfish. In: Advances in Veterinary Medicin. Schultz R (Ed.), Academic Press, NY, USA, 523–537 (1999).
24 Arias CR, Shoemaker C, Evans JJ, Klesius PH. A comparative study of Edwardsiella ictaluri parent (EILO) and E. ictaluri rifampicin-mutant (RE-33) isolates using lipopolysaccharides, outer membrane proteins, fatty acids, Biolog, API 20E and genomic analyses. J. Fish Dis. 26, 415–421 (2003).
25 Shoemaker CA, Klesius PH, Bricker JM. Efficacy of a modified live Edwardsiella ictaluri vaccine in channel catfish as young as 7 days post hatch. Aquaculture 176, 189–193 (1999).
26 Griffiths SG, Melville KJ, Salonius K. Reduction of Renibacterium salmoninarum culture activity in Atlantic salmon following vaccination with avirulent strains. Fish Shellfish Immunol. 8, 607–619 (1998).
27 Kuzyk MA, Burian J, Machander D et al. An efficacious recombinant subunit vaccine against the salmonid rickettsial pathogen Piscirickettsia salmonis. Vaccine 19, 2337–2344 (2001).
• Demonstrates the immunostimulatory effect of mammalian T-cell epitopes in fish when fused with a recombinant Piscirickettsia salmonis protein.
28 Benmansour A, de Kinkelin P. Live fish vaccines: history and perspectives. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Eds). Karger, Basel, Switzerland, 90, 279–289 (1997).
29 Lopez-Doriga MV, Smail DA, Smith RJ et al. Isolation of salmon pancreas disease virus (SPDV) in cell culture and its ability to protect against infection by the wild-type agent. Fish Shellfish Immunol. 11, 505–522 (2001).
30 Ronen A, Perelberg A, Abramowitz J et al. Efficient vaccine against the virus causing a lethal disease in cultured Cyprinus carpio. Vaccine 21, 4677–4684 (2003).
31 Biering E, Villoing S, Sommerset I, Christie KE. Update on viral vaccines for fish. In: Fish Vaccinology, Developments in Biologicals. Midtlyng PJ (Ed.). Karger, Basel, Switzerland, 121, 97–113 (2005).
32 Christie KE, Mockett K, Fyrand K, Goovaerts D, Rødseth OM. Vaccination of Atlantic salmon Salmo salar L. against pancreas disease. 4th International Symposium on viruses of Lower Vertebrates, Weymouth, UK (1998).
33 McLoughlin, Christie KE, Knappskog DH et al. Field trial experiences with an inactivated monovalent pancreas disease virus vaccine. 3rd International Symposium on Fish Vaccinology, 9–11 April, Bergen, Norway (2003).
34 Winton JR. Immunization with viral antigens: Infectious haematopoietic necrosis. In: Fish Vaccinology, Developments in Biological Standardization. Gudding R, Lillehaug A, Midtlyng PJ, Brown F (Ed.) Karger, Basel, Switzerland, 90, 211–220 (1997).
35 Lorenzen N, Olesen NJ, Jorgensen PE et al. Molecular cloning and expression in Escherichia coli of the glycoprotein gene of VHS virus, and immunization of rainbow trout with the recombinant protein. J. Gen. Virol. 74, 623–630 (1993).
36 Lecocq-Xhonneux F, Thiry M, Dheur I et al. A recombinant viral hemorrhagic septicemia virus glycoprotein expressed in insect cells induces protective immunity in rainbow-trout. J. Gen. Virol. 75, 1579–1587 (1994).
37 Cain KD, LaPatra SE, Shewmaker B et al. Immunogenicity of a recombinant infectious hematopoietic necrosis virus glycoprotein produced in insect cells. Dis. Aquat. Org. 36, 67–72 (1999).
38 Simon B, Nomellini J, Chiou P et al. Recombinant vaccines against infectious hematopoietic necrosis virus: production by the Caulobacter crescentus S-layer protein secretion system and evaluation in laboratory trials. Dis. Aquat. Org. 44, 17–27 (2001).
39 Anderson ED, Mourich DV, Fahrenkrug SC et al. Genetic immunization of rainbow trout (Oncorhynchus mykiss) against infectious hematopoietic necrosis virus. Mol. Mar. Biol. Biotechnol. 5, 114–122 (1996).
• First study to demonstrate the high efficacy of fish rhabdoviral DNA vaccines.
40 Lorenzen N, Lorenzen E, Einer-Jensen K et al. Protective immunity to VHS in rainbow trout (Oncorhynchus mykiss, walbaum) following DNA vaccination. Fish Shellfish Immunol. 8, 261–270 (1998).
41 Kim C, Johnson M, Drennan J et al. DNA vaccines encoding viral glycoproteins induce nonspecific immunity and Mx protein synthesis in fish. J. Virol. 74, 7048–7054 (2000).
42 Corbeil S, LaPatra SE, Anderson E, Kurath G. Nanogram quantities of a DNA vaccine protect rainbow trout fry against heterologous strains of infectious hematopoietic necrosis virus. Vaccine 18, 2817–2824 (2000).
•• Demonstrates that a single dose of the DNA vaccine containing the glycoprotein gene of infectious hematopoietic necrosis induces protection in fish at a much lower dose than is usually reported in mammalian DNA vaccine studies.
43 Lorenzen E, Einer-Jensen K, Martinussen T, LaPatra SE, Lorenzen N. DNA vaccination of rainbow trout against viral hemorrhagic septicemia virus: A dose-response and time-course study. J. Aquat. Animal Health 12, 167–180 (2000).
44 LaPatra SE, Corbeil S, Jones GR et al. Protection of rainbow trout against infectious hematopoietic necrosis virus 4 days after specific or semispecific DNA vaccination. Vaccine 19, 4011–4019 (2001).
45 Sommerset I, Lorenzen E, Lorenzen N, Bleie H, Nerland AH. A DNA vaccine directed against a rainbow trout rhabdovirus induces early protection against a nodavirus challenge in turbot. Vaccine 21, 4661–4667 (2003).

Sommerset, Krossøy, Biering & Frost
100 Expert Rev. Vaccines 4(1), (2005)
• The authors demonstrate that a DNA vaccine containing the glycoprotein gene of viral hemorrhagic septicemia virus can induce a strong nonspecific protection against a completely heterologous virus in a nonsalmonid host.
46 Kurath G. Overview of recent DNA vaccination development for fish. 3rd International Symposium on Fish Vaccinology, 9–11 April, Bergen, Norway (2003).
47 Munday BL, Kwang J, Moody N. Betanodavirus infections of teleost fish: a review. J. Fish Dis. 25, 127–142 (2002).
48 Husgard S, Grotmol S, Hjeltnes BK, Rodseth OM, Biering E. Immune response to a recombinant capsid protein of striped jack nervous necrosis virus (SJNNV) in turbot Scophthalmus maximus and Atlantic halibut Hippoglossus hippoglossus, and evaluation of a vaccine against SJNNV. Dis. Aquat. Org. 45, 33–44 (2001).
49 Yuasa K, Koesharyani I, Roza D et al. Immune response of humpback grouper, Cromileptes altivelis (Valenciennes) injected with the recombinant coat protein of betanodavirus. J. Fish Dis. 25, 53–56 (2002).
50 Sommerset I, Skern R, Biering E et al. Protection against Atlantic halibut nodavirus in turbot is induced by recombinant capsid protein vaccination but not following DNA vaccination. Fish Shellfish Immunol. 18, 13–29 (2005).
51 Secombes CJ, Chappell LH. Fish immune responses to experimental and natural infection with helminth parasites. Ann. Rev. Fish Dis. 6, 167–177 (1996).
52 Buchmann K, Lindenstrøm T, Bresciani J. Defence mechanisms against parasites in fish and the prospect for vaccines. Acta Parasitologica 46, 71–81 (2001).
53 Jones SRM. The occurrence and mechanisms of innate immunity against parasites in fish. Dev. Comp. Immunol. 25, 841–852 (2001).
54 McCallum HI. Acquired resistance of black mollies Poecilia latipinna to infection by Ichthyophthirius multifiliis. Parasitology 93, 251–261 (1986).
55 Sin YM, Ling KH, Lam TJ. Protection against velvet disease in goldfish recovered from Ichthyopthiriasis. Aquaculture 102, 187–191 (1992).
56 Burgess PJ, Matthews RA. Cryptocaryon irritans (Ciliophora): aquired immunity in the thick-lipped mullet, Chelon labrosus. Fish Shellfish Immunol. 5, 459–468 (1995).
57 Xu D-H, Klesius PH. Protective effect of cutaneous antibody produced by channel catfish, Ictalarus punctatus (Rafinesque), immune to Ichthyophthirius multifiliis Fouquet on cohabitated nonimmune catfish. J. Fish Dis. 26, 287–291 (2003).
58 Rubio-Godoy M, Tinsley RC. Immunity in rainbow trout, Oncorhynchus mykiss, against the monogean Discococtyle sagittata following primary infection. Parasitol. Res. 92, 367–374 (2004).
59 Dickerson HW, Brown J, Dawe DL, Gratzek JB. Terahymena pyriformis as the protective antigen against Ichthyophthirius multifiliis infection: comparison between isolates and ciliary preparations. J. Fish Biol. 24, 523–528 (1984).
60 Burkart MA, Clark TG. Immunization of channel catfish, Ictalarus punctatus Rafinesque against Ichthyophthirius multifiliis (Fouquet): killed versus live vaccines. J. Fish Biol. 13, 401–410 (1990).
61 Ling KH, Sin YM, Lam TJ. Protection of goldfish against some common ectoparasites protozoans using Ichthyophthirius multifiliis and Terahymena for vaccination. Aquaculture 116, 303–314 (1993).
62 Li S, Woo PTK. Efficacy of a live Cryptobia salmositica vaccine, and the mechanism of protection in vaccinated Onchorhyncus mykiss (Walbaum) against cryptobiosis. Vet. Immunol. Immunopathol. 48, 343–353 (1995).
63 Kim HK, Hwang YJ, Cho JB, Park SI. Immunization of cultured juvenile rockfish Sebastes schlegeli against Microcotyle sebatis (Monogenea). Dis. Aquat. Org. 40, 29–32 (2000).
64 Rubio-Godoy M, Sigh J, Buchmann K, Tinsley RC. Immunization of rainbow trout Oncorhynchus mykiss against Discococtyle sagittata (Monogenea). Dis. Aquat. Org. 55, 23–30 (2003).
65 Willadsen P, McKenna RV, Riding GA. Isolation from the cattle tick, Boophilus microplus, of antigenic material capable of eliciting a protective immunological response in the bovine host. Int. J. Parasitol. 18, 183–189 (1988).
•• Demonstrates the feasibility of vaccinating against an ectoparasite using a single concealed antigen.
66 Rand KN, Moore T, Sriskantha A et al. Cloning and expression of a protective antigen from the cattle tick B. microplus. Proc. Natl Acad. Sci. USA 86, 9657–9661 (1989).
67 Fuente J, Rodriguez M, Redondo M et al. Field studies and cost-effectiveness analysis of vaccination with Gavac™ against the cattle tick Boophilus microplus. Vaccine 16, 366–376 (1998).
68 Miquel A, Müller I, Ferrer P, Valenzuela PDT, Burzio LO. Immunoresponse of Coho salmon immunized with a gene expression library from Piscirickettsia salmonis. Biol. Res. 36, 313–323 (2003).
69 Midtlyng PJ, Lillehaug A. Growth of Atlantic salmon Salmo salar after intraperitoneal administration of vaccines containing adjuvants. Dis. Aquat. Org. 32, 91–97 (1998).
70 Clements CJ, Larsen G, Jodar L. Technologies that make administration of vaccines safer. Vaccine 22, 2054–2058 (2004).
71 Biacchesi S, Yu YX, Bearzotti M, Tafalla C, Fernandez-Alonso M, Bremont M. Rescue of synthetic salmonid rhabdovirus minigenomes. J. Gen. Virol. 81, 1941–1945 (2000).
72 Biacchesi S, Thoulouze MI, Bearzotti M, Yu YX, Bremont M. Recovery of NV knockout infectious hematopoietic necrosis virus expressing foreign genes. J. Virol. 74, 11247–11253 (2000).
73 Lorenzen N, Lorenzen E, Einer-Jensen K, LaPatra SE. Immunity induced shortly after DNA vaccination of rainbow trout against rhabdoviruses protects against heterologous virus but not against bacterial pathogens. Dev. Comp. Immunol. 26, 173–179 (2002).
74 Ellis AE. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 25, 827–839 (2001).
75 Jorgensen JB, Johansen LH, Steiro K, Johansen A. CpG DNA induces protective antiviral immune responses in atlantic salmon (Salmo salar L.). J. Virol. 77, 11471–11479 (2003).
76 Pike AW, Wadsworth SL. Sealice on salmonids: their biology and control. In: Advances in Parasitology. Baker JR, Muller R, Rollinson D (Eds). Academic Press, London, UK, 233–237 (1999).
Websites
101 FAO newsroom 2003 www.fao.org/english/newsroom/news/2003/14203-en.html(Accessed January, 2005)
102 The State of Food and Agriculture 2003–2004 www.fao.org/documents/show_cdr.asp?url_file = /docrep/006/y5160e/y5160e00.htm(Accessed January, 2005)
103 FishstatPlus 2.3 and databases for global aquaculture production in tons and values www.fao.org/fi/statist/fisoft/FISHPLUS.asp(Accessed January, 2005)
104 International koi herpesvirus workshop report www.frmltd.com/Workshop_KHV.htm (Accessed January, 2005)

Fish vaccines
www.future-drugs.com 101
105 Recombinant ISA vaccine informationwww.microtek-intl.com/research/Temp/IHNV&ISAVNEW/ISAV/rrISAV%20vaccine.htm(Accessed January, 2005)
106 Oral vaccine information www.perosbio.com (Accessed January, 2005)
Affiliations• Ingunn Sommerset, PhD
Intervet Norbio AS, Thormøhlensgate 55,N-5008 Bergen, NorwayTel.: +47 555 439 58Fax: +47 55 960 [email protected]
• Bjørn Krossøy, PhD
Intervet Norbio AS, Thormøhlensgate 55,N-5008 Bergen, NorwayTel.: +47 5554 3769Fax: +47 5596 [email protected]
• Eirik Biering, PhD
Institute of Marine Research, PO Box 1870, Nordnes,N-5817 Bergen, NorwayTel.: +47 5523 6949Fax: +47 5523 [email protected]
• Petter Frost, PhD
Institute of Marine Research, PO Box 1870, Nordnes,N-5817 Bergen, NorwayTel.: +47 5523 6362Fax: +47 5523 [email protected]





























